
Evaluación de la actividad polinizadora de Apis mellifera L. y otros
insectos asociados a la floración de tres cultivares de almendro (Prunus
dulcis [Mill] Webb) y tres cultivares de peral asiático (Pyrus pyrifolia
Nakai), en la localidad de Nogales, V región.
Profesor guía: Sergio De La Cuadra I. Profesor corrector: Eduardo Gratacós N. Alumna: Mónica Martínez Domínguez.
PONTIFICIA UNIVERSIDAD CATOLICA DE VALPARAISO FACULTAD DE AGRONOMIA TALLER DE LICENCIATURA
ÁREA FRUTICULTURA
Quillota, Octubre de 2003

2
ÍNDICE DE MATERIAS
1. INTRODUCCIÓN 1
2. REVISIÓN BIBLIOGRÁFICA 4
2.1. Situación actual del cultivo del almendro 4
2.2. Situación actual del cultivo del peral asiático 4
2.3. Antecedentes de la especie 5
2.3.1. Almendro 5
2.3.2. Peral asiático 5
2.3.3. Variedades de almendro 6
2.3.4. Variedades de peral asiático 7
2.4. Floración 8
2.4.1. Comportamiento floral del almendro 8
2.4.2. Comportamiento floral del peral asiático 9
2.4.3. Traslape de floración 10
2.5. Importancia de la polinización 10
2.6. Tipos de polinización 11
2.6.1. Autopolinización 11
2.6.2. Polinización cruzada 11
2.7. Compatibilidad 12
2.8. Período efectivo de polinización 13
2.9. Insectos polinizantes 13
2.10. La abeja melífera como polinizador 16
2.11. Condiciones climáticas que afectan la polinización 18
3. MATERIALES Y MÉTODO 20
3.1. Ubicación y época del estudio 20
3.2. Característica climatológica de la zona de estudio 20
3.3. Material vegetal utilizado 20

3
3.4. Determinación de insectos que visitan las flores de almendro
y peral asiático 21
3.5. Determinación del comportamiento polinizador de la abeja
melífera en las flores 22
3.6. Identificación de insectos asociados a la floración 22
3.7. Determinación de la evolución de la floración 23
3.8. Registro de temperatura 23
3.9. Aplicación del modelo de evaluación de los insectos como
polinizadores 23
3.9.1. Número de individuos / m2 /tiempo 24
3.9.2 Presencia en las flores 24
3.9.3 Tipo de recolección de los insectos 24
3.9.4 Frecuencia de visita a las flores 24
3.10. Porcentaje de cuaja en los cultivares de ambas especies 25
3.11. Análisis estadístico 25
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS 26
4.1. Identificación de insectos visitantes de las flores de almendro
y peral asiático 26
4.2. Comportamiento de los insectos que visitan las flores de
almendro y peral asiático 30
4.2.1. Comportamiento polinizador de Apis mellifera L en almendro 30
4.2.2. Comportamiento polinizador de Apis mellifera L en peral asiático 33
4.2.3. Comportamiento de Drosophila melanogaster Meigen
como potencial polinizador 36
4.3. Curvas de floración 42
4.3.1. Curvas de floración de almendro 42
4.3.2. Curvas de floración de peral asiático 44
4.4. Relación entre el nivel de floración y la visita de Apis mellifera L. 46
4.4.1. Relación entre el número de flores abiertas y la visita
de Apis mellifera L. en almendro 46

4
4.4.2. Relación entre el número de flores abiertas y la visita
de Apis mellifera L. en peral asiático 50
4.5. Relación de la temperatura y la visita de Apis mellifera L. 54
4.5.1. Relación de la temperatura y la visita de Apis mellifera L.
en almendro 54
4.5.2. Relación entre la temperatura y la visita de Apis mellifera L.
en peral asiático 55
4.6. Porcentaje de frutos cuajados en almendro y peral asiático 55
4.7. Aplicación del modelo para evaluar a los insectos como
polinizadores 57
5. CONCLUSIONES 60
6. RESUMEN 61
7. LITERATURA CITADA 62
ANEXOS

5
1. INTRODUCCIÓN
El cultivo de frutales de nuez, como es el caso del almendro, se ha constituido en una
alternativa interesante de exportación para Chile, debido a que sus márgenes de
rentabilidad se han acercado a los de frutas frescas. A nivel mundial se está
reconociendo nutricionalmente a los frutos de nuez por su aporte para la salud. La
incorporación de variedades y el mejoramiento tecnológico, han contribuido a
aumentar la productividad y rentabilidad de los productos de naturaleza seca.
En el caso del peral asiático, a pesar de que el mercado es limitado y hoy está
estabilizado entre oferta y demanda, existen nichos en el Hemisferio Norte, donde se
encuentran comunidades de asiáticos residentes, y en Oriente, que demandan frutos
de grandes calibres y de excelente calidad, generando buenos retornos.
La porción comestible del fruto del almendro, corresponde a la semilla o almendra,
por lo que la polinización de la flor y su posterior fecundación, son de fundamental
importancia para posibilitar que el máximo número de flores logren dar semillas,
considerando además que se trata de una especie autoincompatible, la presencia de
frutos partenocárpicos es improbable. De esta forma, para obtener cosechas
adecuadas, todas las flores deben ser polinizadas en forma cruzada, ya que el
porcentaje de cuaja será determinante en su productividad.
Por otro lado, en peral asiático, la presencia de semillas es importante para la
obtención de frutos de gran calibre y bien formados, características de calidad
exigidas por el mercado, y además, como la mayoría de los cultivares de peral
asiático son autoestériles, se necesita una adecuada polinización cruzada, que
asegurará una cuaja adecuada.

6
Debido a lo anterior, es claro que la polinización libre y cruzada, en la que juegan un
importante papel las abejas, es la mejor forma de asegurar la calidad óptima de la
fruta, se hace necesario evaluar la actividad de la abeja melífera Apis mellifera L. y
otros insectos que estarían asociados a la floración del almendro y del peral asiático.
Esta investigación, plantea que Apis mellifera L. es la especie de mayor importancia
como agente polinizador durante la floración del almendro (Prunus dulcis [Mill]
Webb) y del peral asiático (Pyrus pyrifolia Nakai), y se esperaría la presencia de otros
insectos asociados a la floración de ambas especies, en la localidad de Nogales, V
región.
Por esta razón, se han planteado los siguientes objetivos:
OBJETIVO GENERAL:
Evaluar la actividad polinizadora de Apis mellifera L. y de otros insectos como
polinizadores del almendro (Prunus dulcis [Mill] Webb) en los cultivares Non Pareil,
Price y Carmel, y del peral asiático (Pyrus pyrifolia Nakai) en los cultivares Hosui,
Shinseiki y Nijisseiki.
OBJETIVOS ESPECÍFICOS:
Identificar los insectos asociados a la floración del almendro (Prunus dulcis [Mill]
Webb) en los cultivares Non Pareil, Price y Carmel, y del peral asiático (Pyrus
pyrifolia Nakai) en los cultivares Hosui, Shinseiki y Nijisseiki.
Asociar la floración del almendro (Prunus dulcis [Mill] Webb) en los cultivares Non
Pareil, Price y Carmel, y del peral asiático (Pyrus pyrifolia Nakai) en los cultivares

7
Hosui, Shinseiki y Nijisseiki, con el comportamiento polinizador de Apis mellifera y
otros insectos.
Relacionar la visita de los insectos a las flores de almendro (Prunus dulcis [Mill]
Webb) en los cultivares Non Pareil, Price y Carmel, y del peral asiático (Pyrus
pyrifolia Nakai) en los cultivares Hosui, Shinseiki y Nijisseiki, con la temperatura del
lugar de observación.
Aplicar un modelo para evaluar a los insectos como polinizadores, de acuerdo a su
comportamiento en las flores de almendro (Prunus dulcis [Mill] Webb) en los
cultivares Non Pareil, Price y Carmel, y del peral asiático (Pyrus pyrifolia Nakai) en
los cultivares Hosui, Shinseiki y Nijisseiki.

8
2. REVISIÓN BIBLIOGRÁFICA
2.1. Situación actual del cultivo del almendro:
Tras un período de cuestionamiento a nivel mundial por el alto contenido calórico de
este producto, en la actualidad se está reconociendo a los frutos de nuez como un
importante aporte para la salud, debido a su perfil químico rico en antioxidantes
(vitamina E), flavonoides y compuestos fenólicos (ODEPA, 2001).
En almendras Estados Unidos es el mayor productor (alrededor del 40% del total
mundial) y oferente, ya que exporta entre el 70-80% de los volúmenes transados, que
superan las 200.000 toneladas, en su mayor parte sin cáscara (ODEPA, 2001).
Las exportaciones de Chile de volúmenes cercanos a las 2.500 toneladas, se envían
principalmente hacia países de América del Sur como Argentina, Brasil, Uruguay,
Perú, Colombia, Venezuela y Ecuador (ODEPA, 2001).
Según el VI Censo Agropecuario de 1997, se calcula que la superficie de huertos
comerciales de almendros en 1998, bordeó las 5.750 ha. De éstas, alrededor de 33%
se encontraría en etapa de formación y gran parte de las restantes en creciente y plena
producción. Las plantaciones en Chile están concentradas en la Región Metropolitana
presentando un 48,7% y en la VI región con un 33,2% (ODEPA, 2001).
2.2. Situación actual del cultivo de peral asiático:
Aunque el escenario actual presenta una sobreoferta en los mercados externos para el
caso de las pomáceas, generando retornos estrechos debido a menores precios, los
perales asiáticos constituyen una alternativa de producción, orientados hacia
mercados específicos en el Hemisferio Norte, para satisfacer la demanda de

9
comunidades asiáticas residentes en países como Estados Unidos, basado en una
mayor planificación y un alto nivel tecnológico de las plantaciones, generando
retornos adecuados a los volúmenes producidos. Las plantaciones de peral asiático
representan el 5,3% del total de hectáreas destinadas a la fruticultura nacional, lo que
corresponde a un total de 1.075 ha, concentrándose en la VI-VII regiones (ODEPA,
2001).
2.3. Antecedentes de la especie:
2.3.1. Almendro
El almendro (Prunus dulcis [Mill] Webb), es una especie de hoja caduca,
perteneciente a la familia de las Rosáceas (GIL, 2000). Es originario de Asia (sudeste
de Rusia, Afganistán e Irán) de allí se extendió a la región del Mar Mediterráneo, en
cuyos países es de gran importancia con variedades propias. En América,
posteriormente, cobró importancia en Estados Unidos (GIL, 1997).
Se incluye en el grupo de árboles que producen nueces o frutas secas, en que la parte
comestible corresponde a la semilla. Este tipo de producto se caracteriza por ser más
bien un alimento que una fruta, por su alto contenido de ácidos grasos y proteínas
(ROJAS, 1993).
2.3.2. Peral asiático
Los perales orientales o asiáticos cultivados se han clasificado como Pyrus pyrifolia
Nakai corresponde a una especie de hoja caduca, perteneciente a la familia de las
Rosáceas (GIL, 2000). Tiene su origen en el centro y sur de China y Japón (GIL,
1997). Actualmente su cultivo está difundido por China, Japón, Corea del Sur y

10
Estados Unidos (principalmente en California) en el hemisferio Norte y por Nueva
Zelanda y Australia en el Hemisferio Sur (IGLESIAS, 1993).
Las variedades japonesas son selecciones de Pyrus pyrifolia Nakai, mientras que las
Chinas son híbridos complejos entre Pyrus pyrifolia, Pyrus ussuriensis y Pyrus
bretschneideri (KRAUSE, LOMBARD y STEBBINS, 1987).
Las peras asiáticas son también llamadas “salad pears” (peras de ensaladas), Nashi
(pera en japonés), Li (pera en chino), peras Orientales, peras Chinas, peras Japonesas
o también se les denomina “peras-manzanas”, debido a su textura crocante, y jugosa
o por su forma parecida a la manzana, pero su sabor difiere al ser algo más áspero,
con un sabor dulce, pero sub-ácido (KRAUSE, LOMBARD y STEBBINS, 1987).
2.3.3. Variedades de almendro
Las variedades usadas en Chile son todas de origen californiano. Prácticamente, se
han usado las mismas en los mismos períodos. En California se usan variedades
encontradas a fines del siglo pasado o comienzos de este siglo, como Non Pareil, que
continúa siendo importante, debido a su producción constante y alta demanda
(ROJAS, 1993). En California Non Pareil representa el 43% de las 186.159 hectáreas
de almendros plantados, seguido por Carmel con una participación del 18%
(CONNELL, 2000).
Non Pareil: originada en Suisun, California en 1879. El árbol es de tamaño medio,
parcialmente abierto. Produce tanto en dardos como en brotes largos. De floración
moderadamente temprana, resiste medianamente las heladas en los primeros estados
de yema. La nuez tiene cáscara papel con un alto rendimiento de semilla (65-70%),
sin embargo, es pobremente sellada lo que la hace susceptible al daño de insectos (en
California) y a los pájaros. La semilla es suave, uniforme, atractiva y de color claro.

11
Con un largo medio de 23 mm y un peso medio entre 1.14-1.29 g/semilla (ROJAS,
1993).
Carmel: descubierta en Le Grand, California. El árbol presenta vigor regular, más
cerrado que Non Pareil, redondeado y de tamaño medio. Produce en brotes largos
como en dardos. De floración moderadamente temprana, es un buen polinizante para
Non Pareil. La nuez globosa altamente sellada, con un rendimiento de semilla del
60%. La semilla es relativamente angosta, con un largo medio de 24 mm, y color
moderadamente claro (ROJAS, 1993).
Price: corresponde a una selección de árbol de semilla descubierta en Durham,
California. El árbol tiene buen vigor, un poco abierto, pero más cerrado que Non
Pareil. Puede alternar floración y producción. Produce bien en dardos y algo en brotes
largos, es notoria la producción en racimos, tanto en dardos como en el ápice de
ramas largas, lo que aparentemente reduce la densidad de flores al año siguiente. De
floración conjunta o un día antes que Non Pareil, que lo hace un buen polinizante
para Non Pareil. La semilla tiene un largo que varía entre 20-24 mm, de color
moderadamente clara. Tiene tendencia a producir almendras dobles (10-20%) en un
porcentaje relativamente alto (ROJAS, 1993).
2.3.4. Variedades de peral asiático
Las variedades se caracterizan en función del color de la epidermis, pudiendo
dividirse en dos grupos: variedades de piel verde, amarilla o verde amarillenta,
generalmente exentas de russeting y variedades de piel bronceada, con más o menos
russeting (IGLESIAS, 1993).
Hosui (abundancia): variedad japonesa, obtenida por cruzamiento de Kikosui x
Yakumo x Yacumo. Es, actualmente, una de las mejores variedades, por calibre,

12
aspecto visual (bronceado) y calidad gustativa del fruto. A pesar de esto, su manejo
en plantaciones es delicado por su fuerte vigor y la acrotonía marcada del árbol;
sensible a la caída de flores o pequeños frutos, especialmente los primeros años hasta
que el árbol no está equilibrado. Su entrada en producción es más lenta que en otras
variedades. Presenta numerosos botones florales laterales en ramas del año. Se
poliniza con Nijisseiki, Shinseiki, Shinko, Shinsui (IGLESIAS, 1993).
Shinseiki (Nuevo Siglo): variedad japonesa, obtenida por cruzamiento de Nijisseiki x
Chojuro. Es la variedad de epidermis amarilla más interesante. Fruto de buen calibre
y agradable al consumo, epidermis sensible a los roces y al russeting. Árbol de vigor
medio, muy productivo y de fácil conducción. Fructifica en madera de un año
(especialmente sobre brindilla) hasta tres años. Utilizado habitualmente como
polinizador de numerosas variedades. De floración posterior a Hosui. Se poliniza con
Hosui, Nijisseiki, Kosui, Kikusui, Shinsui, Chojuro (IGLESIAS, 1993).
Nijisseiki (Siglo XX): variedad japonesa, procedente de semillero, de origen
desconocido. Ampliamente cultivada en Japón y California. Fruto de epidermis
verde-amarillenta, con lenticelas evidentes de color oscuro, más sensible a las
rozaduras y al russeting que Shinseiki; calibre medio. Árbol de vigor medio, erecto y
denso, con entrenudos cortos (tipo spur), muy ramificado. Variedad muy fértil y fácil
de conducir, pero necesita un raleo muy importante, y un portainjerto vigoroso para
obtener buen calibre. Se poliniza con Hosui, Shinseiki, Chojuro (IGLESIAS, 1993).
2.4. Floración:
2.4.1. Comportamiento floral del almendro
Las flores del almendro son simples, dispuestas lateralmente en dardos y ramillas
(GIL, 2000).

13
La flor del almendro es hermafrodita con 5 sépalos verdes, 5 pétalos blancos, de base
rosada, con 20-40 estambres en cuya base existen nectarios, 1 pistilo con estigma,
estilo y ovario ínfero monocarpelar con 2 óvulos, cada uno con una ovocélula
(WESTWOOD, 1982).
Aunque las flores son hermafroditas y funcionales, hay que destacar que esta especie
frutal diploide (2n=16) presenta en sus órganos florales una pronunciada
autoincompatibilidad de tipo gametofítico, es decir, la germinación suele producirse
normalmente, pero el crecimiento del tubo es inhibido en el estilo y cubierto de
callosa. Así se impide la autofertilización y se impone la fecundación cruzada entre
cultivares (WESTWOOD, 1982).
2.4.2. Comportamiento floral del peral asiático
Las flores del peral asiático corresponden a un corimbo, donde las flores basales son
las primeras en abrir y son más fuertes que las terminales, y las provenientes de
yemas terminales en dardos son más fuertes que las de yemas laterales de ramillas.
Todas las variedades asiáticas presentan autoincompatibilidad de tipo gametofítica
(GIL, 2000).
La inflorescencia del peral contiene siete u ocho flores, de color blanco, los pétalos
tiene “forma de uña” (suborbiculares a oblongos anchos), los sépalos reflexos o
extendidos, estambres de 20-30, anteras rojas o púrpuras, estilos 2 a 5 libres,
estrechamente unidos en la base por el disco, tiene 2 óvulos por lóculo, y el cáliz a
diferencia de Pyrus communis L es caduco (WESTWOOD, 1982).

14
2.4.3. Traslape de floración
Se refiere a la coincidencia de floración entre los cultivares del huerto. Una cuaja
máxima resulta cuando los cultivares presentan el mismo estado o fase de floración
aproximadamente al mismo tiempo (CONNELL, 2000; SOCÍAS I COMPANY,
FELIPE y ESPADA, 1995). Para evitar un pobre traslape es importante la selección
de polinizantes dentro de un mismo grupo de floración de cultivares, incluyendo más
de dos cultivares en una plantación, de manera que florezcan ligeramente antes y
después que el cultivar comercial (SOTOMAYOR, 1998; KESTER, GRADZIEL y
MICKE, 1994).
2.5. Importancia de la polinización:
La transferencia de granos de polen de una antera a un estigma es denominada
polinización. La polinización cruzada es la transferencia de polen entre flores de dos
cultivares diferentes, de la misma especie o relacionadas; la autopolinización es la
transferencia de polen entre flores de un mismo cultivar (GIL-ALBERT, 1996).
La reproducción sexual y el desarrollo de la semilla dependen de la polinización. La
fecundación tiene lugar al unirse el núcleo masculino del tubo polínico con la oósfera
en el saco embrionario (WESTWOOD, 1982).
La polinización es un requisito previo al cuajado en la mayoría de los frutos, los
durazneros, ciruelos, cerezos, almendros y damascos deben tener semillas para que se
desarrolle el fruto. Normalmente, las manzanas y peras deben tener semillas, aunque
se pueden desarrollar frutos sin semillas en algunos cultivares, normalmente se
producen frutos semillados por polinización cruzada (WESTWOOD, 1982).

15
Particularmente, en almendro se deben polinizar el mayor número de flores posible,
para obtener altos niveles de cuaja, logrando producciones rentables, ya que un
elevado número de frutos no afecta el calibre (DICENTA et al., 2000;
SOTOMAYOR, 1997).
2.6. Tipos de polinización:
2.6.1. Autopolinización
Según ROOT (1976), corresponde a la transferencia de polen al estigma de la misma
flor o de otras flores presentes en el mismo pie de la planta o bien a flores en pies
distintos pero del mismo genotipo.
Cuando la autopolinización ocurre en una misma flor abierta, se denomina
polinización cercana o autogamia (GIL, 2000).
2.6.2. Polinización cruzada
Corresponde a la transferencia de polen desde la flor de un cultivar a otra planta de un
cultivar distinto, por lo que existe un intercambio genético (GIL, 2000; RALLO,
1976).
Para que ocurra esto, las plantas deben disponer de medio atractivos como colores,
olores, polen y néctar, para que sean visitadas por insectos polinizadores (Mc
GREGOR, 1976).

16
2.7. Compatibilidad:
La autoincompatibilidad es el mecanismo desarrollado en algunos vegetales
superiores para evitar que plantas hermafroditas fértiles, produzcan semillas después
de autopolinizarse (BATLLE, ROMERO y VARGAS, 1999).
La capacidad de producir fruta con semilla viable corresponde a la fertilidad,
condición que depende de la viabilidad del polen y del óvulo, además de la
compatibilidad o capacidad del polen de germinar en el estigma, crecer por el estilo y
fecundar el óvulo sin crear rechazo. La incapacidad de producir semillas es la
infertilidad o esterilidad cuyas causas pueden ser morfológicas (diferencias de tamaño
entre tubo polínico y estilo), citológicas (gametos inviables y defectuosos) y
fisiológicas, tanto gametofítica como esporofítica (polen incapaz de germinar o crecer
con la rapidez suficiente para efectuar la fecundación por una reacción de rechazo)
(GIL, 2000).
La incompatibilidad entre polen y estilo, es uno de los factores de mayores
consecuencias económicas en la producción de especies frutales pertenecientes a la
familia de las Rosáceas, que incluye almendro y peral (BATLLE, ROMERO y
VARGAS, 1999).
En las Rosáceas la incompatibilidad está gobernada por un gen multialélico, S, y es
de tipo gametofítica, ya que se produce un rechazo del polen si el alelo S coincide con
uno de los alelos del estilo. Esta inhibición ocurre en el estilo después de un
crecimiento inicial del tubo polínico (SOTOMAYOR, 2001; BATLLE, ROMERO y
VARGAS, 1999).

17
2.8. Período efectivo de polinización:
El período de polinización efectiva (PPE) término acuñado por Williams (1965), es la
diferencia entre el número de días que el polen requiere para germinar y efectuar la
fecundación de la ovocélula y el número de días que la ovocélula permanece viable y
receptiva después de la antesis (WESTWOOD, 1982).
En almendro la flor es receptiva a la polinización el día que abre y su receptividad va
decreciendo en los próximos tres a cuatro días (Mc GREGOR, 1976).
En peral, el período efectivo de polinización es muy dependiente de la temperatura, el
tubo polínico llega al óvulo y lo fecunda en un día a 20° C, en 3-4 días bajo 10° C
(GIL, 2000).
2.9. Insectos polinizantes:
Según ROOT (1976), señala que en la apreciación de los insectos polinizantes, es
necesario considerar dos grupos:
-Insectos polinizantes silvestres, sobre los cuales el hombre no tiene influencia.
-Insectos polinizantes domesticados, sobre los cuales se tiene un control y están
representados principalmente por la abeja melífera, (Apis mellifera L.) y algunos
Bómbidos.
Todas las plantas con flores llamativas son polinizadas por insectos, los cerezos,
ciruelos, manzanos, perales, durazneros y nectarines, principalmente por abejas. Las
avispas silvestres y ocasionalmente otros insectos poseen también una participación
en la polinización de plantas frutales, sin embargo, el éxito de la operación requiere la

18
instalación temporal de colmenas (DE LA CUADRA, 1992; LÓPEZ y
SOTOMAYOR, 1992; WESTWOOD, 1982).
En la gran mayoría de las especies, el polen es transferido por medio de las abejas,
aunque a veces también existen otros insectos polinizadores como abejorros, moscas
y coleópteros (PEÑA, 1987). En la mayoría de los frutales el polen es pesado y
pegajoso, características que lo hacen poco móvil a través del viento, pero sí
adherente al cuerpo y patas de los insectos (RAZETO, 1999).
Según BOSH, BLAS y LACASA (1992); de todos los insectos que visitan las flores
de las plantas fanerógamas para recolectar néctar y/o polen son, sin duda, los
llamados apoideos o abejas los más eficientes como polinizadores, comprendiendo a
Apis mellifera L. y más de 20000 especies de abejas repartidas por todo el mundo.
En estudios realizados en Australia, en flores de ciruelo japonés (Prunus salicina L.),
LANGRIDGE y GOODMAN (1985), encontraron un 88,5% de visitas de Apis
mellifera L., correspondiendo el resto a sírfidos, dípteros y lepidópteras. Señalan que
las flores de longanberry (Rubus loganobaccus L.)fueron visitadas por un 98,6% de
abejas melíferas, siendo el resto dípteros o coleópteros.
CARBÓ (1991) señala que la polinización del manzano se atribuye en 60-95% a Apis
mellifera L., y en estudios realizados en España obtuvo un 38% más de producción
en árboles de manzano polinizados libremente por las abejas.
Según CALZONI y SPERANZA (1998) los abejorros o bombus (Bombus terrestris
L.) exhiben una actividad diaria a campo abierto, durante la tarde y temprano en la
mañana, mientras que la abeja melífera presenta actividad durante la parte central del
día.

19
En estudios realizados en peral (Pyrus communis L.), MAYER y LUNDEN (1997)
describen que los bombus (Bombus occidentalis L.) presentan un menor tamaño en
relación a las abejas melíferas y transportan cargas de polen significativamente más
pequeñas, ya que acarrean cargas cercanas al 12% de su peso corporal, mientras que
las abejas trasladan cargas del 19% de su peso. Además, señalan que Apis mellifera L.
trabaja más rápido visitando un mayor número de flores por minuto; por lo tanto, los
bombus son agentes complementarios en la polinización del peral.
SILVESTER (1997) en un estudio en flores de murta (Ugni molinae Turez)no
encontró presencia de Apis mellifera L. a pesar de existir colmenas en el lugar,
detectando solamente abejorros. Por otra parte, LOBOS (1995), en flores de
frambueso (Rubus idaeus L.) detectó la presencia de Apis mellifera L. en un 93%
siendo el resto abejorros y dípteros.
En Chile, LÓPEZ y ROJAS (1992), en flores de chirimoyo (Annona cherimola Mill.)
detectaron la presencia de insectos polinizadores como Carpophilus hemipterus y
Colopterus sp. coleópteros pertenecientes a la familia Nitidulidae, insectos plaga
como Drosophila melanogaster díptero perteneciente a la familia Drosophilidae, que
según BRNCIC (1992) corresponde a una especie doméstica y generalista en cuanto a
los sitios de ovoposición y requerimientos nutricionales, distribuida alrededor de todo
el mundo.
Según PEÑA (1987), otros apoideos de interés agrícola pertenecen a la familia
Megachilidae, conocidas como abejas cortadoras de hojas, incluye un grupo
numeroso de especies, que son capaces de transportar polen en la parte ventral del
abdomen.
En estudios realizados en España por BOSH, BLAS y LACASA (1992) describen a
la especie del orden Hymenóptera Osmia cornuta como un excelente polinizador de

20
las flores de almendro, porque muestra una clara predilección por el polen de las
rosáceas, por lo que tiende a visitar estas flores aún cuando exista otras plantas en
flor. Además, tanto los machos como las hembras aterrizan siempre sobre los órganos
reproductores de las flores al momento de recolectar polen y/o néctar, presenta
actividad a partir de los 12°C, es poco agresiva ya que nunca ataca a personas ni
animales, y las hembras muestran un alto grado de gregarismo, por lo que tienden a
nidificar cerca unas de otras.
Las condiciones climáticas a principios de primavera limitan la actividad pecoreadora
de la abeja melífera, por lo que se hace necesario evaluar si existen otros insectos
polinizadores asociados a la floración de especies de floración temprana (CALZONI
y SPERANZA, 1998).
2.10. La abeja melífera (Apis mellifera L.) como polinizador:
Las abejas son los principales agentes polinizadores, imprescindibles para las
variedades autoincompatibles y beneficiosas para las autocompatibles (GIL, 2000).
MONTENEGRO y ÁVILA (1995), señalan que factores como el tamaño del grano de
polen, morfología de los órganos florales, simetría floral, color de la corola, cantidad
y calidad del néctar influyen sobre el atractivo que las especies de flores ejercen sobre
las abejas.
En el hábito de trabajo de las abejas, una porción de ellas es colectora de néctar y el
resto de polen, lo que depende de las características florales y de las necesidades de la
colmena. Entre las colectoras de néctar hay abejas que trabajan por encima de la flor
y otras por el lado, siendo las primeras las más apropiadas para la polinización. Éste
es un hábito que se adquiere si las flores del huerto son de tipo cerrado, lo que las
obliga a trabajar por encima (GIL, 2000).

21
La alimentación de la abeja melífera consiste, principalmente, en néctar (miel) y
polen. El néctar provee la mayor parte de los mono y oligosacáridos que la abeja
adulta necesita y el polen es la fuente más importante de proteínas, lípidos, minerales,
tanto para la abeja adulta como para la larva (ROUGIER et al., 1994; VARELA,
SCHUCK y MONTENEGRO, 1991).
Las recolectoras de polen son más eficientes que las recolectoras de néctar como
agentes polinizadores, ya que entran en contacto real con las partes reproductivas de
la flor, adhiriéndose los granos de polen en el pelo de su cuerpo, por lo que tiene una
mayor posibilidad de transferencia del polen (CONNELL, 2000; LOPER, THORP y
BERDEL, 1985).
Mc GREGOR (1976) señala que las pecoreadoras de néctar son activas a lo largo del
día. Cuando las flores pierden sus pétalos debido a la ocurrencia de precipitaciones y
vientos fuertes, pueden permanecer aún atractivas para las pecoreadoras de néctar
(CONNELL, 2000). Además, CASTAÑEDA (2000) señala que prefieren realizar sus
colectas en las partes del árbol que se encuentran más expuestas a la luz solar.
Según SOTOMAYOR (1997), las abejas no sólo trasladan polen de una flor a otra,
sino también transfieren polen a otras abejas dentro de la colmena, las que a su vez
visitan otras flores, prolongándose algunas horas la viabilidad del polen por la
temperatura y humedad favorables de sus cuerpos.
Las abejas deben sentirse a gusto desde el primer momento que salen a volar en un
huerto, por ello deben ser ubicadas en el lugar cuando hay suficiente cantidad de
flores abiertas, que oscila entre 10-30%, para acostumbrarse a ellas, pero no deben
existir flores extrañas distractoras como las que presentan las malezas (GIL, 2000).

22
Existe un número de abejas que se dedica a pecorear superficies relativamente
pequeñas, lo que implica que las variedades polinizantes deben estar lo
suficientemente cercanas para ofrecer a la abeja la posibilidad de desplazarse de una
variedad a otra (RALLO, 1986).
NEIRA et al. (1996) señalan que el tiempo empleado por las abejas en sus visitas a
las flores varía según la cantidad de polen y néctar que la flor ofrezca, además del
tipo de flor, estado fenológico en relación a la edad de la flor, el número de insectos
competidores presentes y las condiciones climáticas.
GARY, WITHERELL y MARSTON (1978) mencionan que las abejas tienden a
aprovechar los recursos alimenticios en territorios cercanos a su colonia, y la
exclusión de la mayoría de las pecoreadoras desde colonias distantes puede ser
explicada por disuasivos de competencia y distancia.
La abeja permanece fuera de la colmena pecoreando hasta que completa su carga de
polen o néctar, y el tiempo que demora depende de la distancia a la que se encuentra
la fuente de alimento y del volumen de néctar que exista (NEIRA et al., 1997).
2.11. Condiciones climáticas que afectan la polinización:
Entre los factores más importantes se encuentra la temperatura que afecta la
germinación del polen y posterior fecundación del óvulo, ya que con temperaturas
inferiores a 12° C el grano de polen tarda demasiado en germinar y el tubo polínico
tiene un desarrollo muy lento como para alcanzar el óvulo en estado aún receptivo.
Por otro lado, con temperaturas superiores a 25° C el óvulo y el grano de polen
maduran y senescen demasiado rápido(RAZETO, 1999). La actividad de la abeja es
óptima con temperaturas entre 18-30° C, es muy baja a 15° C y prácticamente nula a
10° C (GIL, 2000).

23
Vientos superiores a 24 km/h durante la floración impiden el vuelo de la abeja y
deshidrata rápidamente los estigmas, pero es necesario para sacudir flores, liberar el
polen y trasladarlo en el caso de especies anemófilas. Las precipitaciones influyen
negativamente en el vuelo de la abeja melífera, evitan la polinización y pueden
arrastrar el polen, y el agua libre que se deposita sobre las anteras, puede hacer
germinar al polen, inutilizándolo, ya que provoca su maduración en condiciones
adversas (GIL, 2000).
SOTOMAYOR (1998) señala que los períodos fríos y nubosos durante la floración
afectan la cuaja, ya que para una adecuada fecundación las flores deben ser
polinizadas desde su apertura con temperaturas ambientales cálidas.

24
3. MATERIALES Y MÉTODO
3.1. Ubicación y época del estudio:
El estudio se realizó en la sociedad agrícola “El Carmen de Pucalán”, ubicado en la
comuna de Nogales, situado en 32° 43’ latitud sur y 71° 14’ longitud oeste.
Perteneciente a la provincia de Quillota, V región, Chile. Las observaciones se
realizaron en el período de floración de ambas especies, durante los meses de agosto
y septiembre del año 2002.
3.2. Característica climatológica de la zona de estudio:
La comuna de Nogales posee un clima mediterráneo subtropical semiárido, inserto en
el agroclima de La Ligua, el cual se extiende entre los paralelos 31° y 32° 50’ latitud
sur. El sector recibe una atenuada influencia marítima.
El régimen térmico de esta zona se caracteriza por una temperatura media anual de
14,4° C, con una máxima media del mes más cálido (febrero) de 26,5° C, donde la
acumulación térmica de primavera-verano es de 1.600-3.400 grados día (base 10° C),
y una mínima media del mes más frío (julio) de 4,5° C, donde la acumulación de
horas frío puede alcanzar a 754 horas bajo 7° C (NOVOA et al., 1989).
3.3. Material vegetal utilizado:
El predio “El Carmen de Pucalán” posee 43,6 ha de almendros y 10 ha de peral
asiático, entre otros cultivos. Para el caso de los almendros, el estudio se realizó en un
sector de 4,4 ha, donde las variedades comerciales y polinizantes se distribuyen en
grupos de dos hileras del cv. Non Pareil, una hilera del cv. Price y dos hileras del cv.
Carmel, lo que corresponde a una disposición varietal de 40%, 20% y 40%,

25
respectivamente, orientadas en sentido norte-sur, plantadas en el año 1996, sobre
camellones, a una distancia de 6 x 4 m (Anexo 1).
En el caso del peral asiático, el estudio se realizó en un sector de 6,4 ha, separado por
una cortina natural de cassuarina (Casuarina equisetifolia) del resto de las especies,
donde las variedades comerciales y polinizantes se distribuyen en grupos de dos
hileras del cv. Hosui, una hilera del cv. Shinseiki y dos hileras de cv. Nijisseiki, lo
que corresponde a una disposición varietal de 40%, 20% y 40% respectivamente, con
orientación oeste-este, plantadas en el año 1988, sobre camellones, a una distancia de
5 x 3 m (Anexo 2).
Considerando que cada variedad se encuentra en una hilera completa, además de la
distribución de las tres variedades en el sector, se seleccionaron tres hileras de manera
que una variedad se encuentre inmediatamente al lado de la otra. Las observaciones
se realizaron en 12 árboles eligiendo cuatro árboles por variedad, al azar, a una
distancia equidistante entre ellos.
En almendro se colocaron 15 colmenas/ha el día 4 de agosto de 2002 y en peral
asiático se distribuyeron 10 colmenas/ha el día 1 de septiembre de 2002.
3.4. Determinación de los insectos que visitan las flores de almendro y peral:
Durante todo el período de floración de ambas especies, se realizaron observaciones
tres veces por semana, en dos bloques horario, desde las 10:00 hr hasta las 13:00 hr y
desde las 13:30 hr hasta las 16:30 hr, con un tiempo de duración de tres minutos en
cada una de las dos caras del árbol.
Para la observación se eligió una superficie de 1 m2 tanto en la cara oeste como en la
cara este en almendro y en la cara norte y cara sur en peral asiático, a la altura de la

26
vista del observador, la que fue marcada con una cinta de polietileno de color
amarillo. En una tablilla de anotaciones, se registraron todas las visitas de insectos, y
el tipo de recolección que realizaron (estableciendo que cuando recolectan polen
muerden las anteras de la flor, cuando recolectan néctar se posan sobre la flor,
extendiendo su lengua para comenzar a libar y cuando recolectan polen y néctar
simultáneamente presentan ambos comportamientos) determinado visualmente,
durante los tres minutos de observación, en cada una de las dos caras del árbol, en los
dos bloques horario señalados, para lo cual se utilizó un cronómetro.
3.5.Determinación del comportamiento polinizador de los insectos en las flores:
Para la realización de este estudio se utilizó la superficie de 1m2 elegida
anteriormente sobre las caras de los árboles, y se determinó el número de flores
visitadas durante un minuto y la permanencia de los insectos sobre la flor visitada,
según recolectaba polen o néctar, medido en segundos, lo que se determinó
cronometrando este tiempo. Esta medición se realizó en el cultivar Non Pareil en
almendro y en el cultivar Hosui en peral asiático, sobre un árbol de cada cultivar
elegido al azar, una vez al día entre 13:00 y 13:30 horas.
3.6. Identificación de insectos asociados a la floración:
Para la captura de los insectos asociados a la floración de almendro y peral asiático,
se utilizó una red entomológica y un frasco entomológico. Los insectos capturados se
colocaron en otro frasco correctamente etiquetado con la fecha de captura, para su
posterior identificación, que fue realizada en el laboratorio de Entomología
perteneciente al Instituto de Biología de la Pontificia Universidad Católica de
Valparaíso.

27
3.7. Determinación de la evolución de la floración:
Se realizó un seguimiento de la floración, para lo cual se eligió una rama de similar
vigor (diámetro basal) en cada una de las dos caras del árbol, en los 12 árboles de
cada especie, y fue registrado el número de yemas reproductivas totales y se
contabilizó en cada medición el número de flores abiertas. Esto, para relacionar el
número de flores abiertas en el tiempo con la visita de los insectos.
3.8. Registro de temperatura:
La temperatura se registró cada una hora, utilizando termómetros ubicados en el
predio. Estos datos se recuperaron en una planilla Excel para la elaboración de
resultados. Además, se registró la condición climática (despejado, parcial, nublado y
precipitaciones) al momento de la observación.
3.9. Aplicación del modelo de evaluación de los insectos como polinizadores:
Se determinó el valor como polinizador de los insectos detectados en las flores de
ambos cultivos. Se utilizó el modelo descrito por VALDÉS (2002) usado en palto,
que incluye las siguientes variables:
VP=A x B x C x D
Donde:
VP = valor como polinizador
A = Número de individuos por m2/tiempo
B = Presencia en las flores
C = Tipo de recolección del insecto (néctar, polen, néctar + polen)
D = Frecuencia de visita a las flores

28
3.9.1. Número de individuos por m2/tiempo:
Se registraron las visitas de los insectos en la superficie de 1m2 marcada en cada cara
del árbol, durante los tres minutos de observación por cara, en dos bloques horario
diferentes, durante todo el período de floración de ambas especies. El valor utilizado
para esta variable correspondió al número promedio de insectos que visitaron la
superficie de observación.
3.9.2. Presencia en las flores:
Se asignaron dos valores que representaron la presencia o no presencia de insectos en
la flor de almendro y peral asiático, ya que la visita real sobre la flor determina la
transferencia efectiva de polen. Se asignó valor 0 si no visitaba las flores y valor 1 si
la visitaba.
3.9.3. Tipo de recolección de los insectos:
Se asignó valor 1 a los insectos que en su recolección tuvieron contacto con las partes
reproductivas de la flor en almendro y peral asiático, ya sea cuando recolectaron
polen o bien néctar. Se asignó un valor 0 a los insectos que no presentaron un
contacto real con los órganos sexuales de la flor, para el caso de ambos cultivos, ya
que no realizarían la polinización.
3.9.4. Frecuencia de visita a las flores (D)
Se utilizó el valor promedio de flores visitadas durante un minuto, dato obtenido
anteriormente en el estudio del comportamiento polinizador de los insectos en las
flores de ambos cultivos.

29
3.10. Porcentaje de cuaja en los cultivares de ambas especies:
Se contabilizó el número de frutos cuajados en las ramas de los 12 árboles en cada
especie, seleccionadas anteriormente para el estudio de la evolución floral. En
almendro el recuento de frutos fue realizado el 13 de septiembre de 2002, y en el caso
de peral asiático se realizó el 11 de octubre de 2002 (antes de raleo).
El porcentaje de cuaja se obtuvo a través de la siguiente relación:
% = (N° de frutos cuajados / N° de flores totales) x 100
3.11. Análisis estadístico:
Se realizó un análisis de correlación para determinar si existió relación entre el
número de abejas con la temperatura, y con el número de flores abiertas. En el
análisis estadístico se utilizó un nivel de significación de 5%.

30
4. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
4.1. Identificación de insectos visitantes de las flores de almendro y peral asiático:
Se presentó una baja diversidad de especies de insectos que visitaron las flores de
almendro (Prunus dulcis [Mill] Webb) y peral asiático (Pyrus pyrifolia Nakai). En el
Cuadro 1 se presenta el listado de insectos identificados durante el período de
floración de ambas especies cultivadas. Se encontraron solamente dos especies que se
incluyen en los órdenes Hymenóptera y Díptera. Dentro del orden Hymenóptera se
identificó la especie Apis mellifera L. perteneciente a la familia Apidae y del orden
Díptera se identificó la especie Drosophila melanogaster Meigen perteneciente a la
familia Drosophilidae.
CUADRO 1. Listado de insectos identificados durante el período de floración de almendro y peral asiático. Nogales, 2002.
Orden Familia Especie Cultivo HYMENÓPTERA
DÍPTERA
Apidae
Drosophilidae
Apis mellifera L.
Drosophila melanogaster Meigen
Almendro y Peral asiático
Almendro y Peral asiático
Los resultados obtenidos concuerdan con lo expuesto por Mc GREGOR (1976),
quien señala que Apis mellifera L. prácticamente es el único insecto polinizador de
importancia económica en almendro y en peral (Pyrus communis L.). Además, este
autor señala que las flores de peral son visitadas por insectos del orden Hymenóptera,
Díptera y Coleóptera, siendo Apis mellifera L. el visitante predominante, en
comparación a otras especies de dípteros.

31
La presencia de Drosophila melanogaster Meigen se explicaría por su
comportamiento generalista en cuanto a sus sitios de ovoposición y dietas
alimenticias, por lo que pueden ser encontradas en sitios cerrados, chacras, huertos,
jardines y lugares muy alejados de las actividades humanas donde comparten recursos
con especies endémicas (BRNCIC, 1992). Además señala que en Chile se encuentran
alrededor de siete especies cosmopolitas, las que pueden alimentarse de exudados de
savia, néctar y polen de las flores de varias especies.
En los resultados se observa una baja diversidad de especies de insectos, debido a las
condiciones climáticas que se presentaron durante el período de floración de
almendro y peral asiático. RALLO (1986), señala que la polinización por abejas
silvestres y otros insectos procedentes de fuera de la explotación, se limita si se tienen
condiciones meteorológicas adversas como baja temperatura, escasa luminosidad,
además de limitar su actividad a pocos metros de su colmena o lugar de nidificación.
Esta baja diversidad de insectos también se podría explicar por la escasa presencia de
flora nativa en las inmediaciones, ya que el huerto presenta un alto grado de prácticas
culturales, como control de malezas y vegetación natural, junto a la aplicación de
agroquímicos. Según BOSH, BLAS y LACASA (1992), el aumento de la superficie
dedicada a cultivos en zonas rurales, supone la destrucción de lugares de nidificación
de poblaciones de insectos polinizadores. Al respecto, CALZONI y SPERANZA
(1998) señalan que la aplicación de productos químicos para la protección de plagas y
malezas, ha llevado a una disminución drástica de abejas nativas y otros insectos
útiles en la agricultura.
Los Cuadros 2 y 3 presentan el número y porcentaje de insectos totales detectados en
las flores de los cultivares de almendro y peral asiático, respectivamente. Se observa
que Apis mellifera L. es la especie de mayor participación porcentual en almendro y
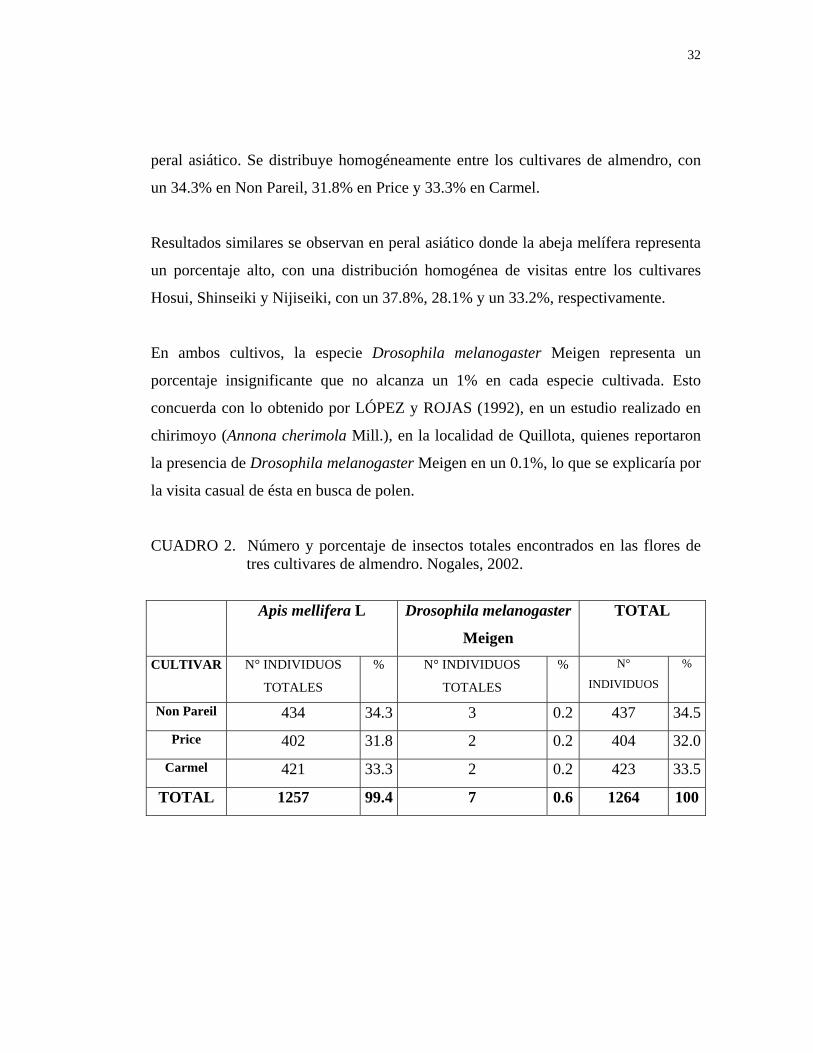
32
peral asiático. Se distribuye homogéneamente entre los cultivares de almendro, con
un 34.3% en Non Pareil, 31.8% en Price y 33.3% en Carmel.
Resultados similares se observan en peral asiático donde la abeja melífera representa
un porcentaje alto, con una distribución homogénea de visitas entre los cultivares
Hosui, Shinseiki y Nijiseiki, con un 37.8%, 28.1% y un 33.2%, respectivamente.
En ambos cultivos, la especie Drosophila melanogaster Meigen representa un
porcentaje insignificante que no alcanza un 1% en cada especie cultivada. Esto
concuerda con lo obtenido por LÓPEZ y ROJAS (1992), en un estudio realizado en
chirimoyo (Annona cherimola Mill.), en la localidad de Quillota, quienes reportaron
la presencia de Drosophila melanogaster Meigen en un 0.1%, lo que se explicaría por
la visita casual de ésta en busca de polen.
CUADRO 2. Número y porcentaje de insectos totales encontrados en las flores de tres cultivares de almendro. Nogales, 2002.
Apis mellifera L Drosophila melanogaster
Meigen
TOTAL
CULTIVAR N° INDIVIDUOS
TOTALES
% N° INDIVIDUOS
TOTALES
% N°
INDIVIDUOS
%
Non Pareil 434 34.3 3 0.2 437 34.5Price 402 31.8 2 0.2 404 32.0
Carmel 421 33.3 2 0.2 423 33.5
TOTAL 1257 99.4 7 0.6 1264 100

33
CUADRO 3. Número y porcentaje de insectos totales encontrados en las flores de tres cultivares de peral asiático. Nogales, 2002.
Apis mellifera L Drosophila melanogaster
Meigen
TOTAL
CULTIVAR N° INDIVIDUOS
TOTALES
% N° INDIVIDUOS
TOTALES
% N°
INDIVIDUOS
%
Hosui 435 37.8 5 0.4 440 38.2Shinseiki 324 28.1 3 0.3 327 28.4Nijiseiki 383 33.2 2 0.2 385 33.4
TOTAL 1142 99.1 10 0.9 1152 100
El alto porcentaje de abejas melíferas encontradas en almendro y peral asiático, se
explica por la colocación y distribución de colmenas en el huerto, manejo
fundamental para que ocurra la polinización cruzada y fructificación en almendro
(DICENTA et al., 2000; SOTOMAYOR, 1997; SOCÍAS I COMPANY, FELIPE y
ESPADA, 1995; DE LA CUADRA, 1992).
MAYER y LUNDEN (1997), IGLESIAS (1993) señalan que al considerar la
floración precoz que presenta el peral asiático, que ocurre cuando las condiciones
climáticas pueden ser adversas, se hace necesario la colocación de colmenas para
asegurar una adecuada polinización, la cual es imprescindible para la obtención de
buenas producciones y frutos de forma regular.
Además, Mc GREGOR (1976) asevera que no hay ninguna duda en colocar colmenas
de abejas melíferas en almendro y peral, siendo las colonias fuertes las más eficientes
en comparación a las colonias débiles, particularmente, cuando se presentan bajas
temperaturas al inicio de primavera. Las colmenas fuertes con crías activas realizan
un mayor pecoreo y polinización cruzada (CONNELL, 2000).

34
Junto a lo anterior DE LA CUADRA (1992) señala que en los cultivos que florecen
temprano en primavera, cuando se presentan días nublados, con bajas temperaturas y
precipitaciones, se hace necesario disponer de una alta población de abejas, que
compense la inactividad de las abejas en estos días.
4.2.Comportamiento de los insectos que visitan las flores de almendro y peral
asiático:
4.2.1. Comportamiento polinizador de Apis mellifera L. en almendro.
Las abejas melíferas visitaron las flores de los cultivares de almendro durante todo el
período de observaciones, debido a que las colmenas se encontraban instaladas al
momento de iniciar este estudio. Además en este período no se presentó floración de
otras especies que significaran fuentes de alimento, descartando la competencia con
la oferta que proporcionan las flores de almendro.
En el Cuadro 4 se presentan el número y porcentaje total de abejas observadas en
flores de los tres cultivares de almendro, durante el período de observación.

35
CUADRO 4. Número y porcentaje total de abejas (Apis mellifera L.) encontradas en ambos períodos de observación (mañana y tarde), durante todo el periodo de floración en los tres cultivares de almendro. Nogales, 2002.
NÚMERO DE ABEJAS
HORARIO Mañana
10:00-13:00 hr
Tarde
13:30-16:30 hr
TOTAL
Non Pareil 203 231 434
Porcentaje 46,7% 53.2% 100%
Price 153 249 402
Porcentaje 38,0% 61,9% 100%
Carmel 209 212 421
Porcentaje 49,6% 50,3% 100%
Como se observa en el Cuadro 4, las abejas visitaron las flores de los tres cultivares
de almendro en ambos períodos de observación, presentando una mayor actividad
durante la tarde (Anexos 7,8 y 9).
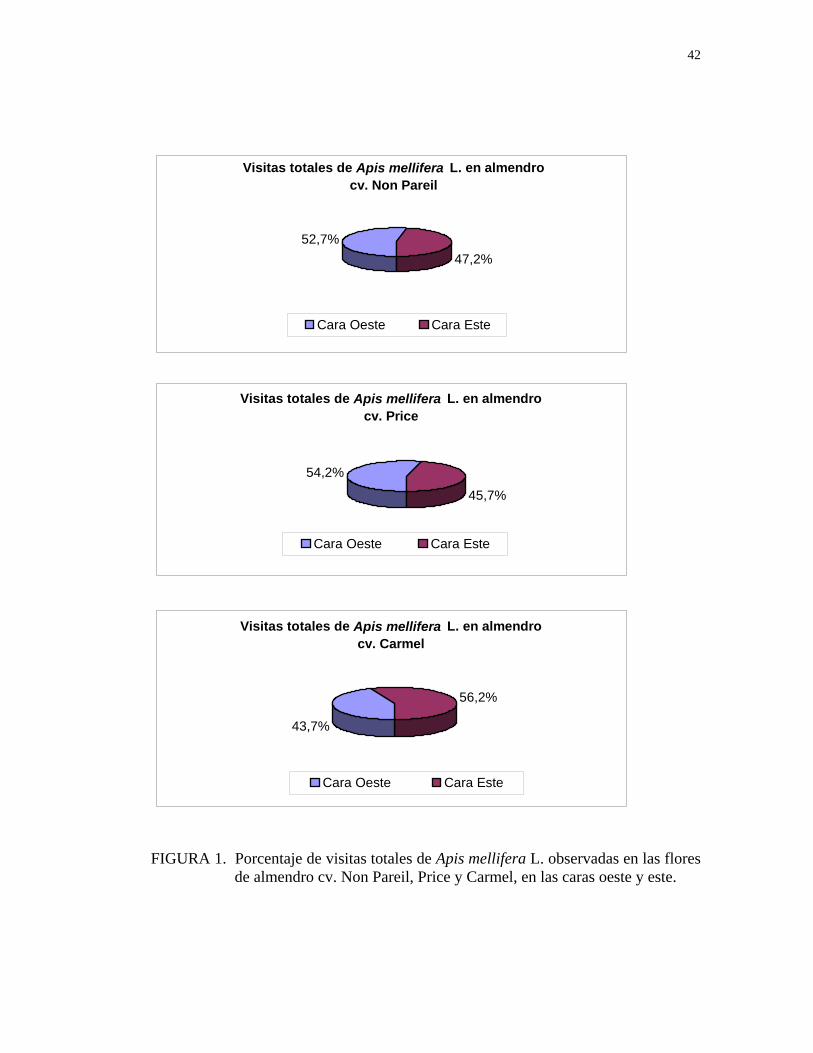
De acuerdo con los resultados obtenidos, la mayoría de las abejas visitaron las flores
de la cara oeste de los cultivares Non Pareil y Price, lo que representa un 52,7% y
54,2% respectivamente, del total de visitas. En cambio la cara este del cultivar
Carmel fue la más visitada, correspondiendo a un 56,2% del total de visitas (Figura
1).
Este comportamiento concuerda con lo descrito por CALZONI y SPERANZA (1998)
quienes señalan que la actividad pecoreadora aumenta con la temperatura y la luz
solar, lo que ocurre cuando las hileras están dispuestas en sentido norte-sur quedando
la cara oeste más expuesta a la luz solar durante la tarde. En cambio los resultados
obtenidos en el cultivar Carmel no coinciden con lo expuesto por estos autores, lo que
se explicaría por un mayor porcentaje de floración en la cara este (Figura 3); al

36
respecto VARELA, SCHUCK y MONTENEGRO (1991) señalan que las especies
con mayor oferta floral son visitadas por un elevado número de abejas.
Se observó en almendro que la mayoría de las abejas recolectaron polen (Cuadro 5),
lo que se explicaría porque las abejas tienden a utilizar especies que tienen polen de
alto contenido de nitrógeno y el almendro posee un alto contenido proteico en su
polen (ROUGIER et al., 1994). Además el polen es la fuente más importante de
proteínas, lípidos, vitaminas y minerales para la abeja adulta y también para la
nutrición de larvas (ROUGIER et al., 1994; VARELA, SCHUCK y
MONTENEGRO, 1991).
CUADRO 5. Tipo de recolección de la abeja melífera en las flores de almendro. Nogales, 2002.
ALMENDRO NECTAR POLEN NÉCTAR + POLEN TOTAL
Porcentaje 37,1% 58,1% 4,8% 100%
Como se observa en el Cuadro 5 el mayor porcentaje corresponde a la recolección de
polen con un 58,1%, lo que concuerda con GARY, WITHERELL y MARSTON
(1978) quienes reportan que entre un 30-50% aproximadamente de las abejas son
recolectoras de polen. Las pecoreadoras de polen son más eficientes como agentes
polinizadores que las pecoreadoras de néctar, ya que al morder las anteras
desempolvan el polen que tienen en su cuerpo sobre el estigma de la flor,
transfiriendo así efectivamente el polen (CONNELL, 2000; LOPER, THORP y
BERDEL, 1985).
El Cuadro 6 presenta el número de flores visitadas por Apis mellifera L. en almendro
durante un minuto y el tiempo de permanencia sobre las flores medido en segundos,
según el tipo de recolección realizada.

37
CUADRO 6. Número de flores visitadas por la abeja en un minuto y tiempo de permanencia en segundos sobre flores de almendro según el tipo de recolección. Nogales, 2002.
Tiempo permanencia (seg/flor)
ALMENDRO N° flores
visitadas/min Polen Néctar
Promedio 5,4 6,5 9,3
Se observa en el Cuadro 6 que la abeja permanece durante un mayor período de
tiempo sobre la flor al succionar néctar, demorando en promedio 9,3 segundos. El
tiempo de permanencia que la abeja emplea sobre la flor, varía de acuerdo al volumen
de polen y néctar, además de la concentración de azúcares en el néctar (NEIRA et al.,
1997). Al respecto RALLO (1986) señala que la flor de almendro secreta diariamente
entre 1,1 y 4,5 mg de néctar con un contenido de 29 a 35% de azúcares y
aproximadamente 1,3 mg de polen; por lo que se considera una flor atractiva para las
abejas.
Según MONTENEGRO y ÁVILA (1995), la actividad de la abeja melífera implica
una intensa acción recolectora, efectuada a través de numerosas visitas en períodos de
tiempo cortos a flores circundantes a sus colmenas, favoreciendo a las plantas en su
polinización.
4.2.2. Comportamiento polinizador de Apis mellifera L. en peral asiático.
Las flores de peral asiático fueron visitadas por un gran número de abejas melíferas
desde el comienzo de la floración, debido a que las colmenas ya estaban instaladas en
esta etapa. Además, esta presencia podría ser explicada por el término de la floración
de almendro (especie que se encontraba a una distancia de 200 m) obligando a la
abeja a volar hacia otras fuentes de alimento, por lo que se produce un reemplazo de
especies en flor (VARELA, SCHUCK y MONTENEGRO, 1991).
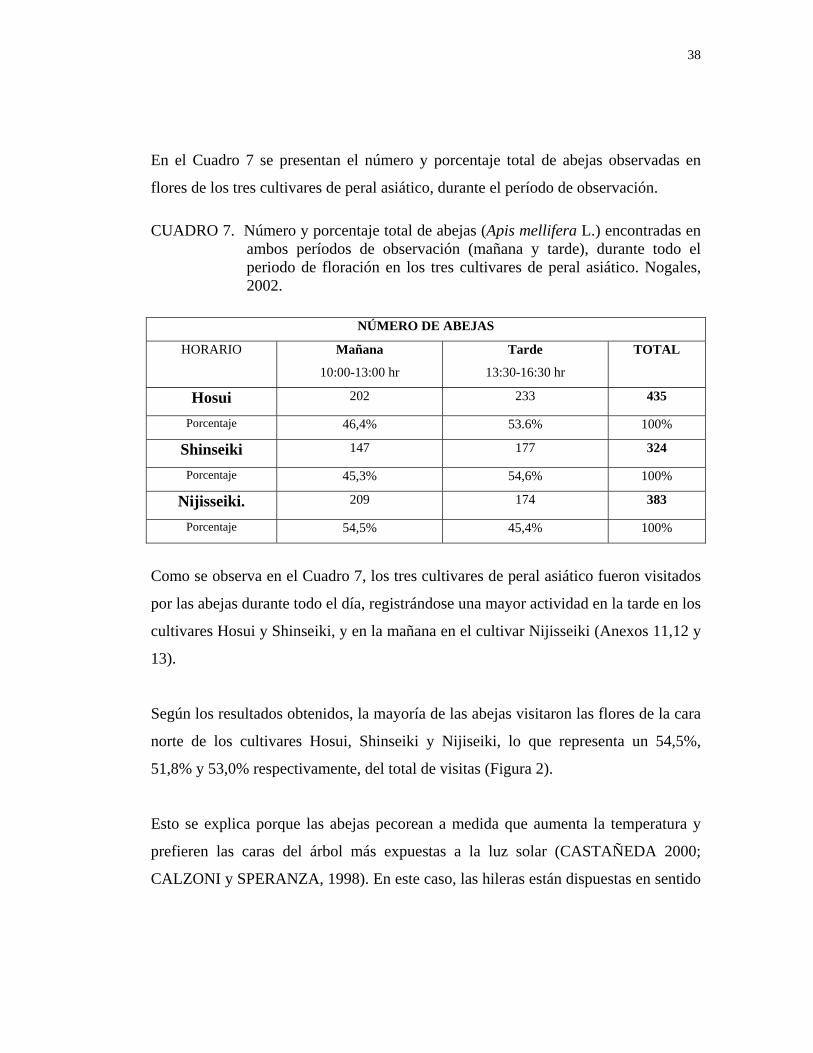
38
En el Cuadro 7 se presentan el número y porcentaje total de abejas observadas en
flores de los tres cultivares de peral asiático, durante el período de observación. CUADRO 7. Número y porcentaje total de abejas (Apis mellifera L.) encontradas en
ambos períodos de observación (mañana y tarde), durante todo el periodo de floración en los tres cultivares de peral asiático. Nogales, 2002.
NÚMERO DE ABEJAS
HORARIO Mañana
10:00-13:00 hr
Tarde
13:30-16:30 hr
TOTAL
Hosui 202 233 435
Porcentaje 46,4% 53.6% 100%
Shinseiki 147 177 324
Porcentaje 45,3% 54,6% 100%
Nijisseiki. 209 174 383
Porcentaje 54,5% 45,4% 100%
Como se observa en el Cuadro 7, los tres cultivares de peral asiático fueron visitados
por las abejas durante todo el día, registrándose una mayor actividad en la tarde en los
cultivares Hosui y Shinseiki, y en la mañana en el cultivar Nijisseiki (Anexos 11,12 y
13).
Según los resultados obtenidos, la mayoría de las abejas visitaron las flores de la cara
norte de los cultivares Hosui, Shinseiki y Nijiseiki, lo que representa un 54,5%,
51,8% y 53,0% respectivamente, del total de visitas (Figura 2).
Esto se explica porque las abejas pecorean a medida que aumenta la temperatura y
prefieren las caras del árbol más expuestas a la luz solar (CASTAÑEDA 2000;
CALZONI y SPERANZA, 1998). En este caso, las hileras están dispuestas en sentido

39
oeste-este quedando la cara norte expuesta a la luz solar durante la mayor parte del
día, que también influye en un mayor porcentaje de floración (Figura 4).
Según NEIRA et al., (1997) la variación del patrón de actividad de las abejas durante
el día depende del atractivo que ejerce la variedad, de las variaciones que sufriría el
volumen y la concentración de azúcar del néctar por efecto de la temperatura y
humedad ambiental, influyendo en la frecuencia de visitas de Apis mellifera L., lo que
podría explicar los resultados obtenidos en el cultivar Nijisseiki.
En el Cuadro 8 se muestran el tipo de recolección realizada en mayor porcentaje por
las abejas en su visita a las flores de peral asiático. Al respecto, RALLO (1986)
señala que no existen abejas recolectoras estrictas de polen ni tampoco de néctar,
aunque una u otra tendencia sea dominante, ya que cuando una abeja recolectora de
polen llega a la flor procede simultáneamente a recoger cierta cantidad de néctar y
polen, esta inevitable doble aptitud le es absolutamente necesaria, pues requieren el
néctar para humedecer los granos de polen, con el fin de aglutinarlos en los cestillos
de las patas traseras.
CUADRO 8. Tipo de recolección de la abeja melífera en las flores de peral asiático.
Nogales, 2002.
PERAL ASIÁTICO NECTAR POLEN NÉCTAR + POLEN TOTAL
Porcentaje 43,2% 51,6% 5,1% 100%
Del Cuadro 8 se desprende que las abejas recolectan polen en mayor porcentaje en las
flores de peral asiático presentando un 51,6%. Este resultado concuerda con lo
reportado por Mc GREGOR (1976), quien señala que las abejas frecuentemente
visitan las flores del peral en búsqueda de su polen, porque es muy abundante y
atractivo, visitando flores de otras especies en búsqueda de néctar. El néctar de la flor

40
de peral tiene una baja concentración de azúcares (entre 2 a 20%), con poco aroma y
un tanto acíbar (RALLO, 1986).
El Cuadro 9 presenta el número de flores visitadas por Apis mellifera L. en peral
asiático durante un minuto y el tiempo de permanencia sobre las flores cronometrado
en segundos según el tipo de recolección realizada.
CUADRO 9. Número de flores visitadas por la abeja en un minuto y tiempo de
permanencia en segundos sobre flores de peral asiático según el tipo de recolección. Nogales, 2002.
Tiempo permanencia (seg/flor) PERAL
ASIÁTICO N° flores
visitadas/min Polen Néctar
Promedio 5,1 6,4 6,2
Se observa en el Cuadro 9 que la abeja visita en promedio 5,1 flores de peral asiático
en un minuto, resultados que se contraponen a lo señalado por MAYER y LUNDEN
(1997) quienes reportaron en peral, que la abeja visitaba en promedio 11,9 flores en
un minuto.
De acuerdo a los resultados obtenidos, la abeja permanece durante períodos de tiempo
similares al recolectar tanto polen como néctar sobre las flores de peral asiático.
Según NEIRA et al., (1996) la cantidad de polen y néctar que ofrece una flor influye
en el tiempo empleado por la abeja en sus visitas a las flores. RALLO (1986) señala
que la flor de peral secreta diariamente entre 0,8 a 2mg de néctar con un contenido de
2 a 20% de azúcares y aproximadamente 0,6 a 1,8mg de polen, que es abundante y
parece presentar un estímulo para las abejas pecoreadoras de polen.
4.2.3. Comportamiento de Drosophila melanogaster Meigen como potencial
polinizador.

41
El número de drosophilas encontradas en las flores de almendro y peral asiático es
insignificante (Cuadros 2 y 3), además se observó que recolectaban néctar, el cual
liban por medio de su boca especializada para esto (PEÑA, 1987), sin necesidad de
tener contacto con las partes reproductoras de la flor (Anexos10 y 14).
Además, esta especie posee hábitos de alimentación generalistas y muy variables
(BRNCIC, 1992; PEÑA, 1987), por lo tanto, no presentaría las características que
posee un potencial polinizador.
42
FIGURA 1. Porcentaje de visitas totales de Apis mellifera L. observadas en las flores
de almendro cv. Non Pareil, Price y Carmel, en las caras oeste y este.
Visitas totales de Apis mellifera L. en almendro cv. Non Pareil
52,7%47,2%
Cara Oeste Cara Este
Visitas totales de Apis mellifera L. en almendro cv. Price
54,2%
45,7%
Cara Oeste Cara Este
Visitas totales de Apis mellifera L. en almendro cv. Carmel
43,7%
56,2%
Cara Oeste Cara Este

43
FIGURA 2. Porcentaje de visitas totales de Apis mellifera L. observadas en las flores
de peral asiático cv. Hosui, Shinseiki y Nijisseiki, en las caras norte y sur.
Visitas totales de Apis mellifera L. en peral asiático cv. Hosui
54,5%
45,5%
Cara Norte Cara Sur
Visitas totales de Apis mellifera L. en peral asiático cv. Shinseiki
51,8%48,1%
Cara Norte Cara Sur
Visitas totales de Apis mellifera L. en peral asiático cv. Nijiseiki
53,0%46,9%
Cara Norte Cara Sur

44
FIGURA 3. Porcentaje de floración de almendro cv. Non Pareil, Price y Carmel, en
las caras oeste y este.
Floración en almendro cv. Non Pareil
59%
41%
Cara Oeste Cara Este
Floración en almendro cv. Price
54%46%
Cara Oeste Cara Este
Floración en almendro cv. Carmel
49%51%
Cara Oeste Cara Este

45
FIGURA 4. Porcentaje de floración de peral asiático cv. Hosui, Shinseiki y
Nijisseiki, en las caras norte y sur.
Floración en peral asiático cv. Hosui
55%
45%
Cara Norte Cara Sur
Floraciónen peral asiático cv. Shinseiki
57%
43%
Cara Norte Cara Sur
Floración en peral asiático cv. Nijiseiki
49%51%
Cara Norte Cara Sur

46
4.3.Curvas de floración:
Se determinó el número de flores abiertas en almendro y peral asiático durante todo el
período de observación, y se confeccionaron curvas de floración para cada cultivar
(Anexos 5 y 6).
4.3.1. Curva de floración de almendro.
En la Figura 5 se observa el traslape de floración de los tres cultivares de almendro,
en la localidad de Nogales durante agosto de 2002.
Al comenzar las mediciones se presentaron precipitaciones los días 5 y 7 de agosto,
por lo que observó un bajo número de flores abiertas. A partir del día 9 de agosto los
tres cultivares comenzaron a presentar una acelerada floración expresada en un alto
porcentaje de flores abiertas, alcanzando el “peak” el día 14 de agosto los tres
cultivares, por lo que al observar la tendencia de las curvas se advierte una
coincidencia de floración marcada entre los tres cultivares.
Además, se distingue que los cultivares polinizantes Carmel y Price mantienen un
mayor porcentaje de flores abiertas con respecto al cultivar comercial Non Pareil
durante todo el período de floración.
Los resultados obtenidos anteriormente concuerdan con lo reportado por KESTER,
GRADZIEL y MICKE (1994), quienes señalan que para asegurar una óptima
polinización cruzada en almendro, es necesario la existencia de variedades de
floración coincidente con no más de dos a tres días de diferencia en la época de
floración.

47
FIGURA 5. Traslape de floración entre tres cultivares de almendro expresado en
porcentaje de flores abiertas. Nogales, 2002. pp: precipitaciones
0
10
20
30
40
50
60
70
80
2-A
go
5-A
go
7-A
go
9-A
go
12-A
go
14-A
go
16-A
go
19-A
go
21-A
go
23-A
go
26-A
go
28-A
go
30-A
go
Fecha
Porc
enta
je d
e flo
res
abie
rtas
(%
)
Non pareil Price Carmel
pppp

48
4.3.2. Curva de floración de peral asiático.
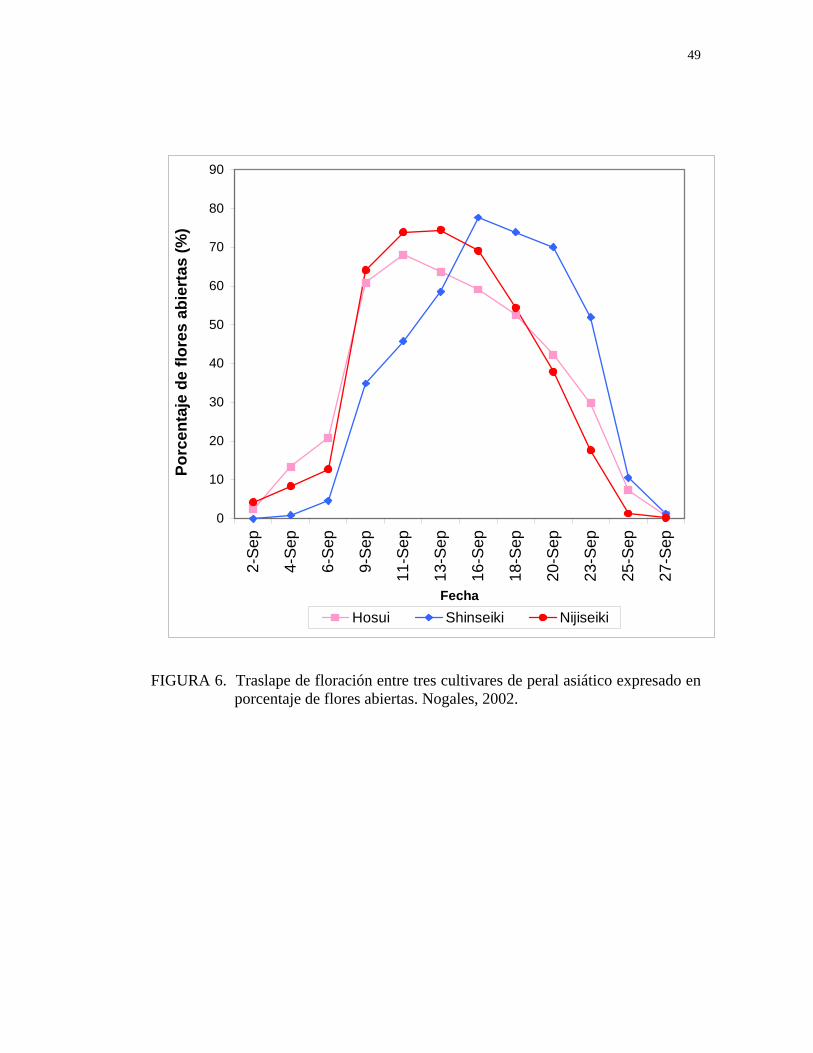
Las mediciones comenzaron el día 2 de septiembre donde se observó que los
cultivares Hosui y Nijisseiki se encontraban en inicio de floración, presentando una
tendencia creciente de flores abiertas, aumentando notablemente a partir del día 6 de
septiembre, marcando una diferencia con el cultivar Shinseiki que comenzó su
floración dos días después que estos cultivares, aumentando las flores abiertas en un
menor porcentaje hasta el día 9 de septiembre, a partir del cual presenta un
incremento en la floración, alcanzando un “peak” de flores abiertas el día 16 de
septiembre a pesar de la diferencia inicial en el porcentaje con respecto a los otros
cultivares, que alcanzaron un menor “peak” de floración cinco días antes (Figura 6).
Estos resultados son similares a los obtenidos en un estudio realizado por
CONTARDO (1993) quien señala que el cultivar Nijisseiki cubre mejor el período de
plena flor que el cultivar Hosui, en comparación con el cultivar Shinseiki, y que el
cultivar Hosui presenta un período de plena flor más extendido y de menor intensidad
en relación a los otros dos cultivares.
IGLESIAS (1993) señala que las épocas de floración en peral asiático fluctúan según
las variedades y pueden ser modificadas más o menos por las temperaturas
primaverales. Además, reporta que Hosui es una de las variedades de floración más
precoz, seguida por Nijisseiki. Por otra parte, señala que la floración de Shinseiki es
ligeramente posterior a Hosui.
Se observa que las curvas de floración coinciden logrando el traslape entre los tres
cultivares, durante septiembre de 2002 en la localidad de Nogales. La concordancia
de floración entre variedades es fundamental para asegurar una buena polinización,
para la obtención de producciones adecuadas (IGLESIAS, 1993).
49
FIGURA 6. Traslape de floración entre tres cultivares de peral asiático expresado en porcentaje de flores abiertas. Nogales, 2002.
0
10
20
30
40
50
60
70
80
90
2-S
ep
4-S
ep
6-S
ep
9-S
ep
11-S
ep
13-S
ep
16-S
ep
18-S
ep
20-S
ep
23-S
ep
25-S
ep
27-S
ep
Fecha
Porc
enta
je d
e flo
res
abie
rtas
(%)
Hosui Shinseiki Nijiseiki

50
4.4. Relación entre el nivel de floración y la visita de Apis mellifera L.:
4.4.1. Relación entre el número de flores abiertas y la visita de la abeja melífera en almendro.
Se realizó un recuento del número de flores abiertas en los tres cultivares de almendro
y se correlacionó con el número total de abejas que visitaron a estos cultivares, para
determinar posibles relaciones entre ellos.
La correlación entre ambos parámetros fue alta para los tres cultivares, es decir, a
medida que aumentó la floración, aumentaron también las visitas de abejas melíferas
(Anexo 15).
Como se observa en las Figuras 7, 8 y 9 el número de visitas de abejas aumenta al
existir un número mayor de flores abiertas (Anexo 5), esto se acentúa en días
despejados (Anexo 3), disminuyendo en días nublados o parciales, llegando a cero
visitas cuando ocurren precipitaciones. Se observa que las visitas se reanudan al
existir días despejados, pero en menor medida, ya que existe un bajo número de flores
abiertas.
Según CONNELL (2000), los factores abióticos del medio ambiente regulan el vuelo
de las abejas, de esta forma las abejas pecorean cuando las temperaturas son
superiores a 12,8° C y no lo hacen cuando hay lluvia; la nubosidad también reduce el
vuelo de la abeja especialmente cuando ocurre con temperaturas cercanas al umbral
(12,8° C).

51
FIGURA 7. Relación entre el número de visitas de Apis mellifera L. y el número de flores abiertas en el cultivar Non Pareil.
pp: precipitaciones
0
10
20
30
40
50
60
70
80
90
100
2-A
go
5-A
go
7-A
go
9-A
go
12-A
go
14-A
go
16-A
go
19-A
go
21-A
go
23-A
go
26-A
go
28-A
go
30-A
goFecha
N° t
otal
de
visi
tas
de A
pis
mel
lifer
a L
.
0
50
100
150
200
250
300
350
400
N° d
e flo
res
abie
rtas
N° total de visitas de Apis mellifera N° flores abiertas
pppp

52
FIGURA 8. Relación entre el número de visitas de Apis mellifera L. y el número de flores abiertas en el cultivar Price.
pp: precipitaciones
0
10
20
30
40
50
60
70
80
90
2-A
go
5-A
go
7-A
go
9-A
go
12-A
go
14-A
go
16-A
go
19-A
go
21-A
go
23-A
go
26-A
go
28-A
go
30-A
goFecha
N° t
otal
de
visi
tas
de A
pis
mel
lifer
a L
.
0
50
100
150
200
250
300
350
400
450
N° d
e flo
res
abie
rtas
N° total de visitas de Apis mellifera N° flores abiertas
pppp

53
FIGURA 9. Relación entre el número de visitas de Apis mellifera L. y el número de flores abiertas en el cultivar Carmel.
pp: precipitaciones
0
10
20
30
40
50
60
70
80
90
100
2-A
go
5-A
go
7-A
go
9-A
go
12-A
go
14-A
go
16-A
go
19-A
go
21-A
go
23-A
go
26-A
go
28-A
go
30-A
goFecha
N° t
otal
de
visi
tas
de A
pis
mel
lifer
a L
.
0
100
200
300
400
500
600
700
800
N° d
e flo
res
abie
rtas
N° total de visitas de Apis mellifera N° flores abiertas
pppp

54
4.4.2. Relación entre el número de flores abiertas y la visita de la abeja melífera en peral asiático.
Se contabilizó el número de flores abiertas en los tres cultivares de peral asiático y se
correlacionó con el número total de abejas que visitaron a estos cultivares.
La correlación entre ambos parámetros fue medianamente alta para los tres cultivares,
es decir, a medida que aumentó la floración, aumentaron también las visitas de abejas
melíferas (Anexo 16).
En las Figuras 10, 11 y 12 se observa que el número de abejas visitantes aumenta a
mayor número de flores abiertas (Anexo 6), esto se acentúa en días despejados
(Anexo 4), disminuyendo en días nublados o parciales. El día 9 de septiembre se
observa un bajo número de abejas melíferas en el cultivar Hosui lo que se explicaría
por la oferta más atractiva de alimento por parte del cultivar Nijisseiki con un mayor
número de flores abiertas, donde se registró un mayor número de insectos. El cultivar
Shinseiki en esta fecha presentaba un menor número de flores abiertas que el cultivar
Nijiseiki por lo que los resultados fueron similares. Las visitas se reanudan al existir
días despejados, pero en menor número, ya que existe un descenso de flores abiertas
hacia el término de la floración.

55
FIGURA 10. Relación entre el número de visitas de Apis mellifera L. y el número de flores abiertas en el cultivar Hosui.
0
20
40
60
80
100
120
140
2-S
ep
4-S
ep
6-S
ep
9-S
ep
11-S
ep
13-S
ep
16-S
ep
18-S
ep
20-S
ep
23-S
ep
25-S
ep
27-S
epFecha
N° t
otal
de
visi
tas
de A
pis
mel
lifer
a L
.
0
50
100
150
200
250
300
350
N° d
e flo
res
abie
rtas
N° total de visitas de Apis mellifera N° flores abiertas

56
FIGURA 11. Relación entre el número de visitas de Apis mellifera L. y el número de flores abiertas en el cultivar Shinseiki.
0
10
20
30
40
50
60
70
80
90
2-S
ep
4-S
ep
6-S
ep
9-S
ep
11-S
ep
13-S
ep
16-S
ep
18-S
ep
20-S
ep
23-S
ep
25-S
ep
27-S
epFecha
N° t
otal
de
visi
tas
de A
pis
mel
lifer
a L
.
0
50
100
150
200
250
300
350
N° d
e flo
res
abie
rtas
N° total de visitas de Apis mellifera N° flores abiertas
57
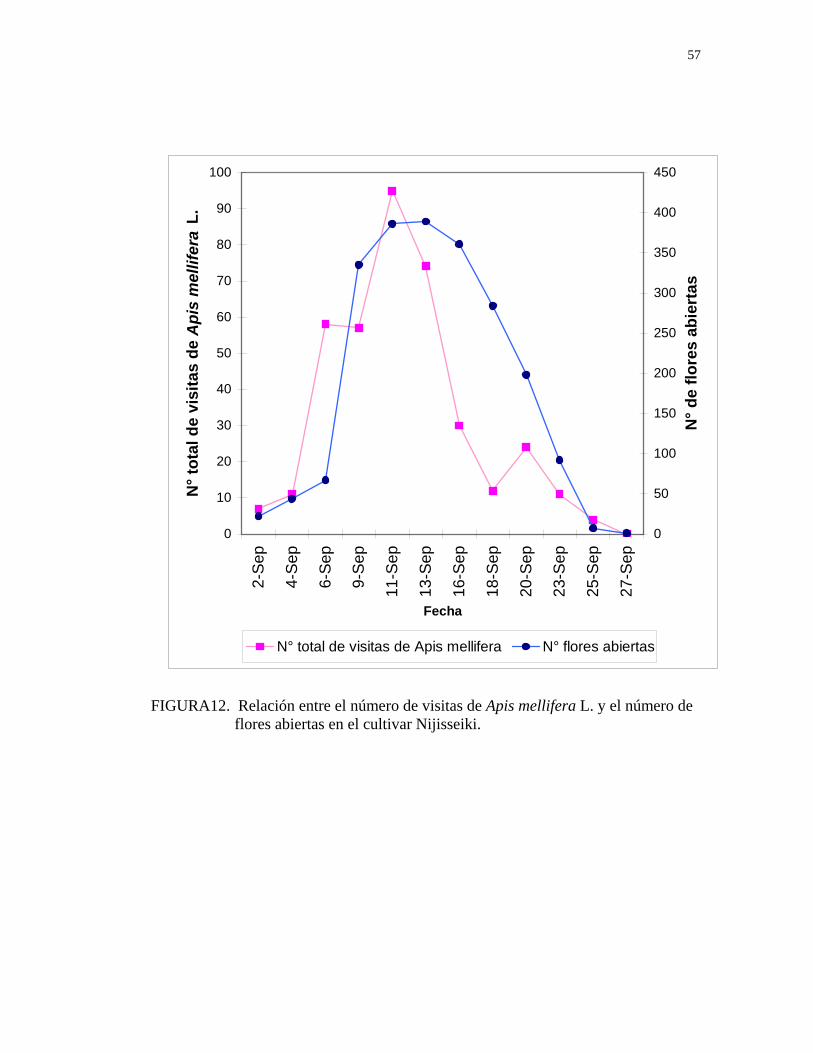
FIGURA12. Relación entre el número de visitas de Apis mellifera L. y el número de flores abiertas en el cultivar Nijisseiki.
0
10
20
30
40
50
60
70
80
90
100
2-S
ep
4-S
ep
6-S
ep
9-S
ep
11-S
ep
13-S
ep
16-S
ep
18-S
ep
20-S
ep
23-S
ep
25-S
ep
27-S
epFecha
N° t
otal
de
visi
tas
de A
pis
mel
lifer
a L
.
0
50
100
150
200
250
300
350
400
450
N° d
e flo
res
abie
rtas
N° total de visitas de Apis mellifera N° flores abiertas

58
4.5. Relación de la temperatura y la visita de Apis mellifera L.:
4.5.1. Relación entre la temperatura y la visita de Apis mellifera L. en almendro
El número de abejas observadas en los tres cultivares, tanto en la mañana como en la
tarde está correlacionada positivamente con la temperatura (Anexo 15), lo que indica
que el número de visitas aumenta junto con la temperatura.
Por otro lado, el número de abejas que visitan la cara este en los tres cultivares
durante la mañana se correlaciona positivamente con la temperatura (Anexo 15), lo
que se explicaría por una mayor exposición de la cara este a la luz solar en la mañana
debido a la disposición de las hileras en sentido norte-sur. El número de visitas
observadas en la cara este, durante la tarde, se correlaciona positivamente con la
temperatura en los cultivares Price y Carmel, es decir, a medida que aumenta la
temperatura se incrementa la actividad de las abejas, y medianamente en el cultivar
Non Pareil (Anexo 15), debido a un menor porcentaje de floración y visitas de abejas
en la cara este del cultivar Non Pareil (Figuras 1 y 3).
Las visitas de las abejas en la cara oeste durante la mañana se correlacionan
positivamente con la temperatura en los cultivares Non Pareil y Carmel, y
medianamente con el cultivar Price, y en la tarde se correlacionan positivamente en
los cultivares Non Pareil y Price (Anexo 15).
Se observó presencia de abejas a partir de los 12,9° C, con una mayor actividad
durante la tarde, ya que se presentaron temperaturas más altas que en la mañana, lo
que concuerda con lo señalado por RALLO (1986), quien señala que las abejas
desarrollan mayor actividad con temperaturas comprendidas entre los 15 y 26° C, y
decrece al disminuir la temperatura, llegando a ser nula por debajo de los 10° C
(Anexo 17).

59
4.5.2. Relación entre la temperatura y la visita de Apis mellifera L. en peral asiático
El número de abejas observadas en los tres cultivares, tanto en la cara norte como en
la cara sur, durante la mañana y la tarde no se correlaciona con la temperatura, a
excepción de las visitas de abejas en la cara norte del cultivar Shinseiki durante la
mañana que se correlaciona levemente con la temperatura (Anexo 16).
Se registró visitas de abejas con temperaturas de 12,7° C, presentando una mayor
actividad durante la tarde donde se presentaron temperaturas mayores en
comparación a las temperaturas de la mañana. El día 18 de septiembre se observó una
disminución importante del número de abejas encontradas en las flores, y se presentó
la temperatura promedio más baja del período de observación en este cultivo, que
junto a una condición climática de día nublado, pudo haber influido en la escasez de
visitas de abejas en relación al resto del período, donde se observó un predominio de
días despejados, en los cuales hubo una mayor actividad de abejas (Anexo 18).
En general, en los tres cultivares se observó una disminución del número de visitas de
abejas a contar del día 11 de septiembre, aun cuando existían temperaturas favorables
para su actividad y además se presentaba un número considerable de flores abiertas,
lo que se explicaría por la baja atracción que posee la flor del peral asiático como
fuente de alimento para la abeja.
4.6. Porcentaje de frutos cuajados en almendro y en peral asiático:
En los Cuadros 10 y 11 se presenta el porcentaje de frutos cuajados en ambas
especies.

60
CUADRO 10. Porcentaje de frutos cuajados según exposición en los tres cultivares de almendro.
Exposición Non Pareil Price Carmel
Oeste 13,2% 33,4% 38,7%
Este 34,8% 34,2% 24,0%
Se observa en el Cuadro 10 un bajo porcentaje de frutos cuajados en la cara oeste del
cultivar Non Pareil y en la cara este del cultivar Carmel, a pesar que ambas caras
presentaron un alto porcentaje de floración (Figura 3), lo que se explicaría por la
distribución varietal, ya que estas caras no quedan expuestas a un cultivar polinizante
(Anexo 1). De los resultados se puede inferir la importancia que posee la distribución
de los cultivares en el huerto y la distancia de los cultivares comerciales con respecto
a los polinizantes, para favorecer una adecuada polinización cruzada, ya que se
observa un mayor porcentaje de frutos cuajados en las caras que se encuentran frente
al cultivar donador de polen.
CUADRO 11. Porcentaje de frutos cuajados según exposición en los tres cultivares
de peral asiático.
Exposición Hosui Shinseiki Nijisseiki
Norte 54,9 42,2 56,6
Sur 49,0 43,5 46,5
En el Cuadro 11 se observa un mayor porcentaje de frutos cuajados en la cara norte
de los cultivares Hosui y Nijisseiki, lo que podría explicarse por el alto porcentaje de
floración en esta cara, atribuible a la orientación de la hilera en sentido oeste-este que
permite una mayor exposición de la cara norte a la luz solar (Figura 4) (Anexo 2).

61
4.7. Aplicación del modelo para evaluar a los insectos como polinizadores:
El modelo descrito por VALDÉS (2002) permite asignar a los insectos que visitan las
flores de las especies un valor como polinizador, que incluye todas las características
del comportamiento que presentan estos insectos, como el número de individuos
observados, presencia en las flores, tipo de recolección y frecuencia de visita a las
flores.
De acuerdo al comportamiento de Drosophila melanogaster Meigen en almendro y
peral asiático, esta especie no presentó características como potencial polinizador, por
lo tanto, se calculó el valor como polinizador (VP) de Apis mellifera L. especie más
abundante en almendro y peral asiático a través del siguiente modelo:
VP = A x B x C x D
En los Cuadros 12 y 13 se presenta el valor como polinizador de Apis mellifera L., y
su porcentaje de participación en relación a otros insectos presentes en almendro y
peral asiático.
CUADRO 12. Valor como polinizador (VP) de Apis mellifera L., y porcentaje de
participación en almendro. Nogales, 2002.
Especie A D VP %
N° individuos (promedio/m2/t)
Frecuencia de visitas (flores/min)
Valor polinizador
ALMENDRO 8,1 5,4 43,7 100 t= 3 minutos/cara árbol

62
CUADRO 13. Valor como polinizador (VP) de Apis mellifera L., y porcentaje de participación en peral asiático. Nogales, 2002.
Especie A D VP %
N° individuos (promedio/m2/t)
Frecuencia de visitas (flores/min)
Valor polinizador
PERAL ASIÁTICO
7,9 5,1 40,3 100
t=3 minutos/cara árbol
En esta evaluación se utilizó el número promedio de abejas melíferas visitantes por
m2, valor que se obtuvo con la información de los Cuadros 2 y 3. Las variables
presencia en las flores (B) y tipo de recolección (C) no se incluyeron , porque se les
asignó un valor 1, ya que la abeja siempre visitó la flor y tuvo contacto con los
órganos reproductivos de la flor en ambos cultivos. LOPER, THORP y BERDEL
(1985) señalan que las abejas colectoras de polen pueden transferir el polen que
llevan adherido en el pelo del cuerpo a las colectoras de néctar al tener contacto con
ellas cuando regresan al interior de la colmena. La variable frecuencia de visita a las
flores es un buen indicador de la eficiencia de la abeja como agente polinizador, ya
que al visitar un mayor número de flores por minuto transfiere una mayor cantidad de
polen, valor que se encuentra en los Cuadros 6 y 9.
Para el caso de Drosophila melanogaster Meigen, la variable tipo de recolección (C)
tiene un valor 0, ya que al visitar la flor en busca de néctar no tuvo contacto real con
los órganos sexuales, por lo que el valor como polinizador (VP) es 0.
De los Cuadros 12 y 13 se desprende que la participación de Apis mellifera L. como
insecto polinizador es de un 100% para el caso de ambas especies, lo que concuerda
con lo señalado por la mayoría de los autores quienes señalan que esta especie es el
principal agente polinizador en almendro y en peral asiático. En estos resultados

63
influye la presencia de colmenas de abejas durante el período de observación, manejo
comercial realizado para la obtención de adecuadas producciones en ambas especies.
En estudios realizados por LANGRIDGE y GOODMAN (1985b) reportan que la
gran proporción de abejas encontradas entre los insectos visitantes de las flores de
especies de prunus indican su importancia como polinizador, debido al movimiento
que realizan de flor en flor y de planta en planta, lo que asegura una adecuada
polinización cruzada.
Por otra parte, señalan que el bajo número de insectos nativos que visitan estas flores
indican la insuficiencia total de estas especies para los propósitos de polinización.
La contribución de las abejas melíferas en la reproducción de las especies vegetales y
en la polinización de cultivos hace que esta especie sea reconocida a nivel mundial
(RALLO, 1986).

64
5. CONCLUSIONES
Existe una baja diversidad de especies de insectos asociados a la floración de
almendro y peral asiático, encontrándose solamente dos especies de los órdenes
Hymenóptera y Díptera.
Hymenóptera se presenta como el orden de mayor relevancia, siendo Apis mellifera
L. la especie predominante en las flores de los tres cultivares de almendro y peral
asiático, con un 99,4% y 99,1% de visitas, respectivamente.
Existió un alto grado de correlación entre el número de flores abiertas y el número de
visitas de Apis mellifera L. en almendro y un grado medianamente alto en peral
asiático.
El grado de correlación entre las visitas de la abeja a las flores de almendro y la
temperatura del lugar de observación fue positivo. En peral asiático no existió
correlación entre ambos parámetros.
Apis mellifera L. presenta una participación del 100% en la polinización de almendro
y peral asiático, no existiendo otros insectos que participen en la polinización de estos
cultivos, en la localidad de Nogales, V región.

65
6. RESUMEN
Entre el 2 de agosto y el 27 de septiembre de 2002 se realizó un estudio para evaluar
la actividad polinizadora de Apis mellifera L. y otros insectos asociados a la floración
de almendro y peral asiático. Para esto se seleccionaron doce árboles en cada especie
estudiada, en la localidad de Nogales, V región.
Para la determinación de los insectos asociados a la floración de almendro y peral
asiático se realizaron observaciones tres veces por semana en dos bloques horario
diferentes, desde 10:00 hr a 13:00 hr y desde 13:30 hr a 16:30 hr, sobre una superficie
de 1m2 marcada en ambas caras de los árboles seleccionados.
Para la identificación de los insectos detectados durante el período de observación, se
procedió a la captura de un ejemplar de la especie registrada, utilizando una red
entomológica.
Se observó que existe una baja diversidad de especies de insectos, siendo Apis
mellifera L. la especie predominante en ambos cultivos estudiados. Por su parte, la
presencia de Drosophila melanogaster Meigen representó valores menores a 1%.
Apis mellifera L. fue la única especie que presentó atributos de potencial agente
polinizador en almendro y peral asiático, por lo que se determinó un modelo de
evaluación que incluye número de individuos en 1m2, tipo de recolección, frecuencia
de visitas a las flores. Se determinó que Apis mellifera L. participa en un 100% en la
polinización de almendro y peral asiático, en la localidad de Nogales, V región.

66
7. LITERATURA CITADA
BATLLE, I., ROMERO, M.A., y VARGAS, F.J. 1999. Aplicación de las ribonucleasas (RNasas) estilares al cultivo y mejora genética del almendro. Fruticultura Profesional 104: 54-58.
BOSCH, J., BLAS, M., y LACASA, A. 1992. Osmia cornuta (Hymenoptera;
megachilidae), un nuevo polinizador para los almendros. Fruticultura Profesional 44: 65-71.
BRNCIC, D. 1992. Abundancia, distribución y estacionalidad de adultos de las
especies cosmopolitas del género Drosophila (díptera, drosophilidae) en Chile. Acta Entomológica Chilena 17: 11-19.
CALZONI, G.L., and SPERANZA, A. 1998. Insect controlled pollination in
Japanese plum (Prunus salicina Lindl.). Scientia Horticulturae 72: 227-237.
CARBÓ, J. 1991. Mejora de la polinización de los manzanos con la introducción de
abejas durante la floración. Fruticultura Profesional 38: 65-68. CASTAÑEDA, A. 2000. Identificación y eficiencia de insectos polinizadores del
aguacatero en los estados de México y Michoacán, México. Tesis de Postgrado. Montecillo, Texcoco, México. Colegio de Post graduados, Institución de Enseñanza e Investigación en Ciencia Agrícola. 87p.
CONNELL, JH. 2000. Pollination of almonds: practices and problems.
HortTechnology 10(1): 116-119. CONTARDO, C. 1993. Determinación de la fenología floral y curva de crecimiento
del fruto en tres variedades de peral asiático (Pyrus pyrifolia Nakai, Pyrus serotina R., Pyrus bretscneideri), para dos localidades de la V región. Tesis Ing. Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 144p.
DE LA CUADRA, S. 1992. La importancia de las abejas en la polinización de los
frutales. La Palma 5: 27-34. DICENTA, F., ORTEGA, E., CANOVAS, J.A., y EGEA, J. 2000. Hacia la
plantación monovarietal de almendro con variedades autocompatibles. Fruticultura Profesional 11: 11-18.

67
GARY, N., WITHERELL, P., and MARSTON, J. 1978. Distribution and foraging activities of honeybees during almond pollination. Journal of Apicultural Research 17(4): 188-194.
GIL-ALBERT, F. 1996. Tratado de arboricultura frutal. Madrid, Mundi-Prensa. vol
2. 248p. GIL, G. 1997. Fruticultura: el potencial productivo. Santiago, Ediciones
Universidad Católica de Chile. 342p _________. 2000. Fruticultura: la producción de fruta. Primera edición. Santiago,
Ediciones Universidad Católica de Chile. 583p. IGLESIAS, I. 1993. El nashi. Fruticultura Profesional 54: 15-34. KESTER, D.E., GRADZIEL, T.M., and MICKE, W.C. 1994. Identifying pollen
incompatibility groups in California almond cultivars. Journal of the American Society for Horticultural Science 119(1): 106-109.
KRAUSE, W., LOMBARD, P., and STEBBINS, R. 1987. Cultivo de Peras
Asiáticas. Universidad de Chile, Facultad de Ciencias Agrarias y Forestales. Cultivo de peras rojas y asiáticas. Santiago, 29-30 de abril 1987. pp39-56
LANGRIDGE, D.F., and GOODMAN, R.D. 1985a. Honeybee pollination of
loganberries (Rubus loganobaccus L.H. Bailey). Australian journal of Experimental Agriculture 25(1): 224-226.
_________., and _________. 1985b. Honeybee pollination of Japanese plums
(Prunus salicina Lindl. Cv. Satsuma) in the Goulburn valley, Victoria. Australian journal of Experimental Agriculture 25(1): 227-230.
LOBOS, N. 1995. Entomofauna asociada al frambueso (Rubus idaeus L.) y
evaluación del atrayente feromoral Beescent en la actividad polinizadora de Apis mellifera L. Tesis Ing. Agr. Valdivia, Universidad Austral de Chile, Facultad de Ciencias Agrarias. 72p.
LOPER, G.M., THORP, R.W., and BERDEL, R. 1985. Improving honey bee
pollination efficiency in almonds. California Agriculture 39(11-12): 19-20.

68
LÓPEZ, E., y ROJAS, R. 1992. Artrópodos asociados a la floración del chirimoyo (Annona cherimola Mill.) en la localidad de Quillota, quinta región, Chile. Acta Entomológica Chilena 17: 101-106.
_________., y SOTOMAYOR, C. 1992. Las abejas como polinizantes en frutales.
Chile Agrícola 17(180): 270-272. MAYER, D.F., and LUNDEN, JD. 1997. A comparison of commercially managed
bumblebees and honey bees (Hymenoptera: apidae) for pollination of pears. Acta Horticulturae 437: 283-288.
Mc GREGOR, S. 1976. Insect pollinator of cultivated crop plant. Washington.
Departament of Agriculture United States. 441p. MONTENEGRO, G., y ÁVILA, G. 1995. Continua actividad de Apis mellifera en
Lo Blanco, V región de Chile. Ciencia e Investigación Agraria 22(1-2): 44-48.
NEIRA, M., NALLAR, J., PESSOT, R., y CARRILLO, R. 1996. Actividad
polinizadora de Apis mellifera L., en peral, bajo la influencia de dos atrayentes. Agro Sur 24(2): 167-176.
_________., PALACIOS, J., CARRILLO, R., PESSOT, R., y MUNDACA, N. 1997.
Conducta polinizadora de Apis mellifera L. y Bombus ruderatus F. (Hymenoptera: apidae) sobre dos cultivares de Cranberry, Vaccinium macrocarpon Ait. (Ericaceae). Agro-Ciencia 13(3): 337-344.
NOVOA, R., VILLASECA, R., DEL CANTO, P., ROVANET, J., SIERRA, C., DEL
POZO, A. 1989. Mapa Agroclimático de Chile. Santiago, INIA. 221p. OFICINA DE ESTUDIOS Y POLÍTICAS AGRARIAS. 2001. Mercado
agropecuario. N° 102. OFICINA DE ESTUDIOS Y POLÍTICAS AGRARIAS. 2001. Mercado
agropecuario. N° 112. PEÑA, L. 1987. Introducción a los insectos de Chile. Santiago, Editorial
Universitaria. 253p. RALLO, J. 1986. Frutales y abejas. Madrid, Publicaciones de Extensión Agraria.
231p.

69
RAZETO, B. 1999. Para entender la fruticultura. 3a ed. Santiago, Universidad de Chile. 373p.
ROJAS, C. 1993. Cultivo del almendro. Universidad Católica de Valparaíso ,
Facultad de Agronomía. Curso, reciclaje para ingenieros agrónomos, frutales de hoja caduca. Santiago, vol 2. pp1-35.
ROOT, A. 1976. ABC y XYZ de la Apicultura. Enciclopedia de la cría científica y
practica de las abejas. 15a. Ed. Buenos Aires. 670p ROUGIER, D., TIMMERMANN, B., FUENTES, E.,YATES, L., BAS, F., y
MONTENEGRO, G. 1994. Relación entre la selectividad de la abeja melífera (Apis mellifera) y el contenido de proteína cruda del grano de polen. Diagnóstico en la flora nativa de Chile. Ciencia e Investigación Agraria 21(1-2): 47-52.
SILVESTER, R. 1997. Entomofauna asociada a las flores de murta (Ugni molinae
Turez) y su evaluación como polinizadores. Tesis Ing. Agr. Valdivia, Universidad Austral de Chile, Facultad de Ciencias Agrarias. 63p.
SOCIAS I COMPANY, R., FELIPE, A.J., y ESPADA, J.L. 1995. Diseño de
plantación y cuajado en el almendro. Fruticultura Profesional 72: 11-13. SOTOMAYOR, C. 1997. Evaluación de diferentes modalidades de polinización en
almendro. Ciencia e Investigación Agraria 24(1): 7-11. _________. 1998. Polinización y cuaja frutal del almendro. Universidad Católica de
Chile. Seminario situación actual y perspectivas tecnológicas del almendro. Santiago, 30 y 31 de julio1998. pp. 36-48
_________. 2001. Polinización y fructificación del almendro. Chile Agrícola
26(253): 258-261. VALDÉS, C. 2002. Evaluación de la actividad de Apis mellifera L. y otros insectos
asociados a la floración del palto (Persea americana Mill.) cv. Hass en dos localidades de la V región (Quillota y La Ligua). Taller de Licenciatura. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 76p.
VARELA, D., SCHUCK, M., y MONTENEGRO, G. 1991. Selectividad de Apis
mellifera L., en su recolección de polen en la vegetación de Chile central (región metropolitana). Ciencia e Investigación Agraria 18(1-2): 73-78.

70
WESTWOOD, M. 1982. Fruticultura de Zonas Templadas. Madrid, Mundi-Prensa. 461p.

71
ANEXOS

72
ANEXO 1. Esquema de la distribución varietal en huerto de almendro.
:Non Pareil :Price :Carmel

73
ANEXO 2. Esquema de la distribución varietal en huerto de peral asiático.
:Hosui :Shinseiki :Nijisseiki

74
ANEXO 3. Registro de temperaturas durante el periodo de mediciones para almendro, Nogales V región.
TEMPERATURA (°C) PRECIPITACIONES CONDICIÓN Promedio Mañana Tarde Promedio 02-08-02 14,8 16,7 15,8 0,0 Despejado 05-08-02 10,9 11,4 11,2 6,4 Precipitaciones 07-08-02 15,7 13,9 14,8 9,4 Precipitaciones 09-08-02 20,2 25,5 22,9 0,0 Despejado 12-08-02 22,7 28,5 25,6 0,0 Despejado 14-08-02 19,2 21,9 21 0,0 Despejado 16-08-02 13,7 17,2 15,5 0,0 Nublado 19-08-02 13,9 15,7 14,8 0,0 Parcial 21-08-02 18 22,7 20,4 0,0 Despejado 23-08-02 12,6 13,5 13,1 0,6 Precipitaciones 26-08-02 14 15,5 14,8 9,4 Precipitaciones 28-08-02 12,9 15,5 14,2 0,0 Despejado 30-08-02 18,4 19,9 19,2 0,0 Despejado
Fuente: Estación meteorológica, de la Sociedad Agrícola “El Carmen de Pucalán”

75
ANEXO 4. Registro de temperaturas durante el periodo de mediciones para peral asiático, Nogales V región.
TEMPERATURA (°C) CONDICIÓN Fecha Mañana Tarde Promedio
02-09-02 22,5 25,5 24 Despejado 04-09-02 15,2 13,8 14,5 Nublado 06-09-02 18,5 21 19,8 Parcial 09-09-02 22,2 18,4 20,3 Parcial 11-09-02 21,1 22 21,6 Despejado 13-09-02 21,9 19,9 20,9 Despejado 16-09-02 18,8 12,7 15,8 Parcial 18-09-02 13,7 15,1 14,4 Nublado 20-09-02 22 25,2 23,6 Despejado 23-09-02 24,9 28,5 26,7 Despejado 25-09-02 25,5 26,4 26 Despejado 27-09-02 15,4 18,5 17 Parcial
Fuente: Estación meteorológica, de la Sociedad Agrícola “El Carmen de Pucalán
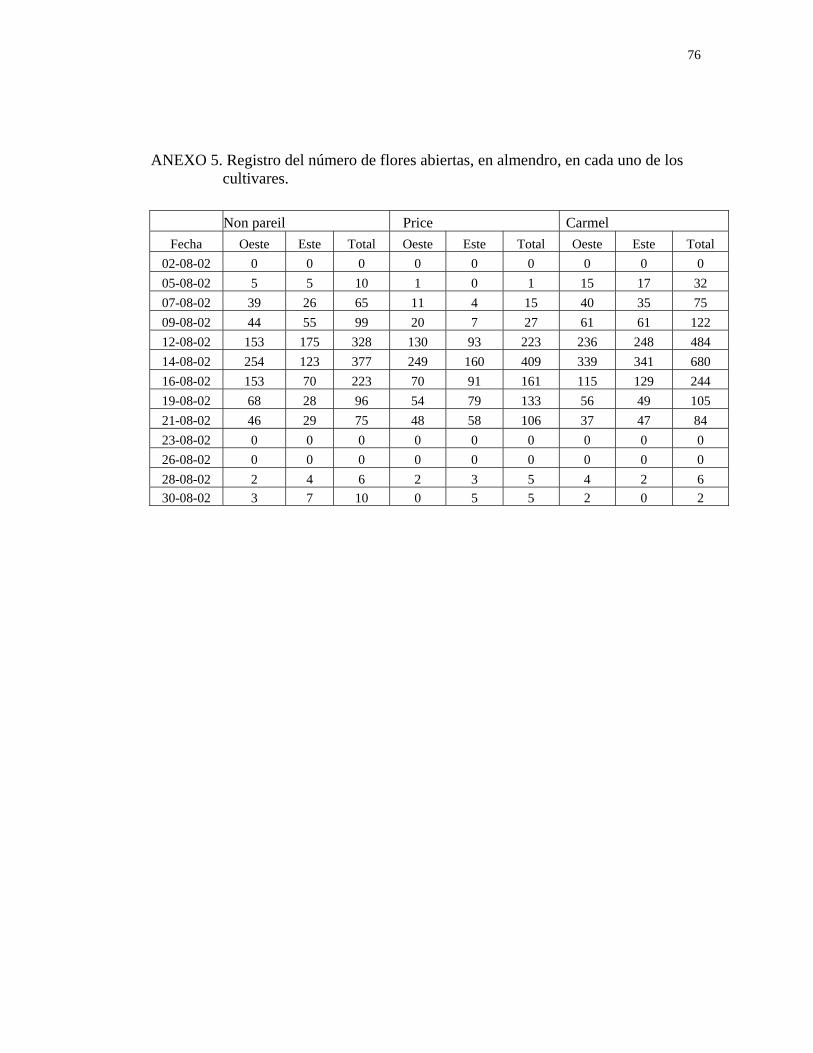
76
ANEXO 5. Registro del número de flores abiertas, en almendro, en cada uno de los cultivares.
Non pareil Price Carmel Fecha Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 05-08-02 5 5 10 1 0 1 15 17 32 07-08-02 39 26 65 11 4 15 40 35 75 09-08-02 44 55 99 20 7 27 61 61 122 12-08-02 153 175 328 130 93 223 236 248 484 14-08-02 254 123 377 249 160 409 339 341 680 16-08-02 153 70 223 70 91 161 115 129 244 19-08-02 68 28 96 54 79 133 56 49 105 21-08-02 46 29 75 48 58 106 37 47 84 23-08-02 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 28-08-02 2 4 6 2 3 5 4 2 6 30-08-02 3 7 10 0 5 5 2 0 2

77
ANEXO 6. Registro del número de flores abiertas, en peral asiático, en cada uno de los cultivares
Hosui Shinseiki Nijiseiki Fecha Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 5 7 12 0 0 0 12 10 22 04-09-02 32 32 64 0 3 3 21 23 44 06-09-02 50 49 99 11 6 17 32 35 67 09-09-02 177 113 290 89 40 129 142 193 335 11-09-02 182 143 325 113 56 169 178 208 386 13-09-02 168 136 304 128 88 216 199 190 389 16-09-02 155 127 282 147 140 287 177 184 361 18-09-02 131 120 251 144 129 273 156 128 284 20-09-02 112 90 202 143 116 259 112 86 198 23-09-02 76 66 142 113 79 192 46 46 92 25-09-02 22 13 35 16 23 39 5 2 7 27-09-02 4 0 4 2 3 5 1 0 1
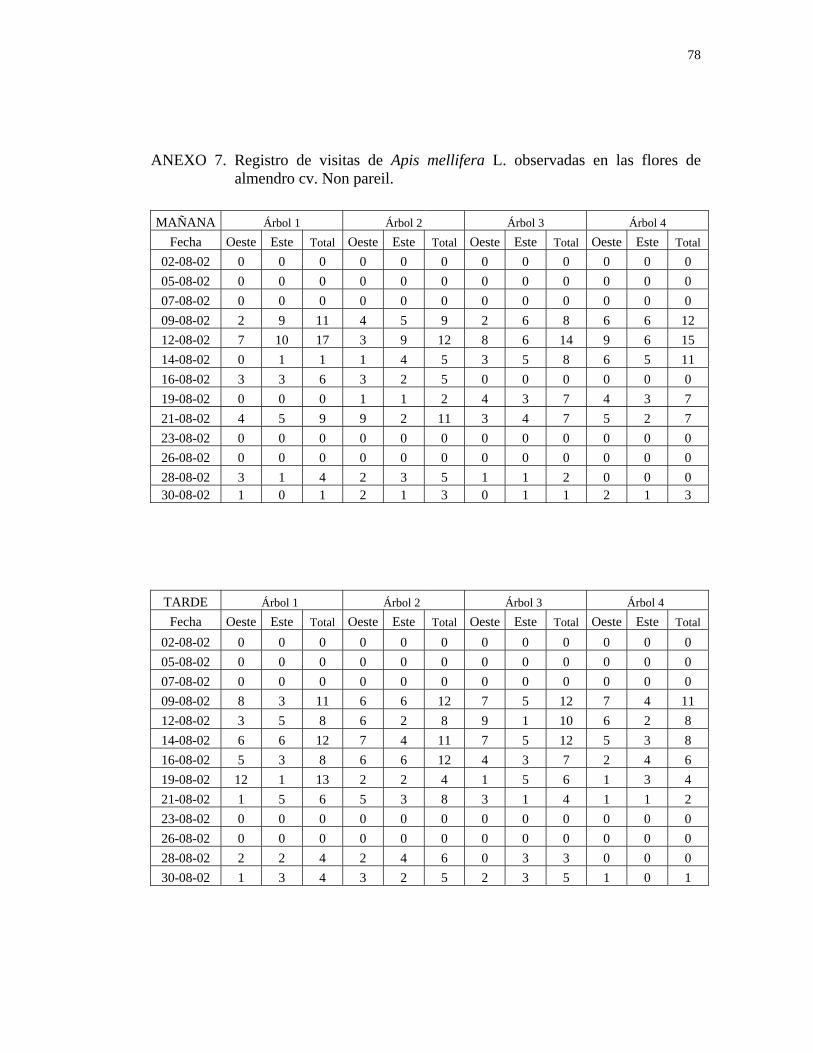
78
ANEXO 7. Registro de visitas de Apis mellifera L. observadas en las flores de almendro cv. Non pareil.
MAÑANA Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Oeste Este Total Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 0 0 0 09-08-02 2 9 11 4 5 9 2 6 8 6 6 12 12-08-02 7 10 17 3 9 12 8 6 14 9 6 15 14-08-02 0 1 1 1 4 5 3 5 8 6 5 11 16-08-02 3 3 6 3 2 5 0 0 0 0 0 0 19-08-02 0 0 0 1 1 2 4 3 7 4 3 7 21-08-02 4 5 9 9 2 11 3 4 7 5 2 7 23-08-02 0 0 0 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 0 0 0 28-08-02 3 1 4 2 3 5 1 1 2 0 0 0 30-08-02 1 0 1 2 1 3 0 1 1 2 1 3
TARDE Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Oeste Este Total Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 0 0 0 09-08-02 8 3 11 6 6 12 7 5 12 7 4 11 12-08-02 3 5 8 6 2 8 9 1 10 6 2 8 14-08-02 6 6 12 7 4 11 7 5 12 5 3 8 16-08-02 5 3 8 6 6 12 4 3 7 2 4 6 19-08-02 12 1 13 2 2 4 1 5 6 1 3 4 21-08-02 1 5 6 5 3 8 3 1 4 1 1 2 23-08-02 0 0 0 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 0 0 0 28-08-02 2 2 4 2 4 6 0 3 3 0 0 0 30-08-02 1 3 4 3 2 5 2 3 5 1 0 1
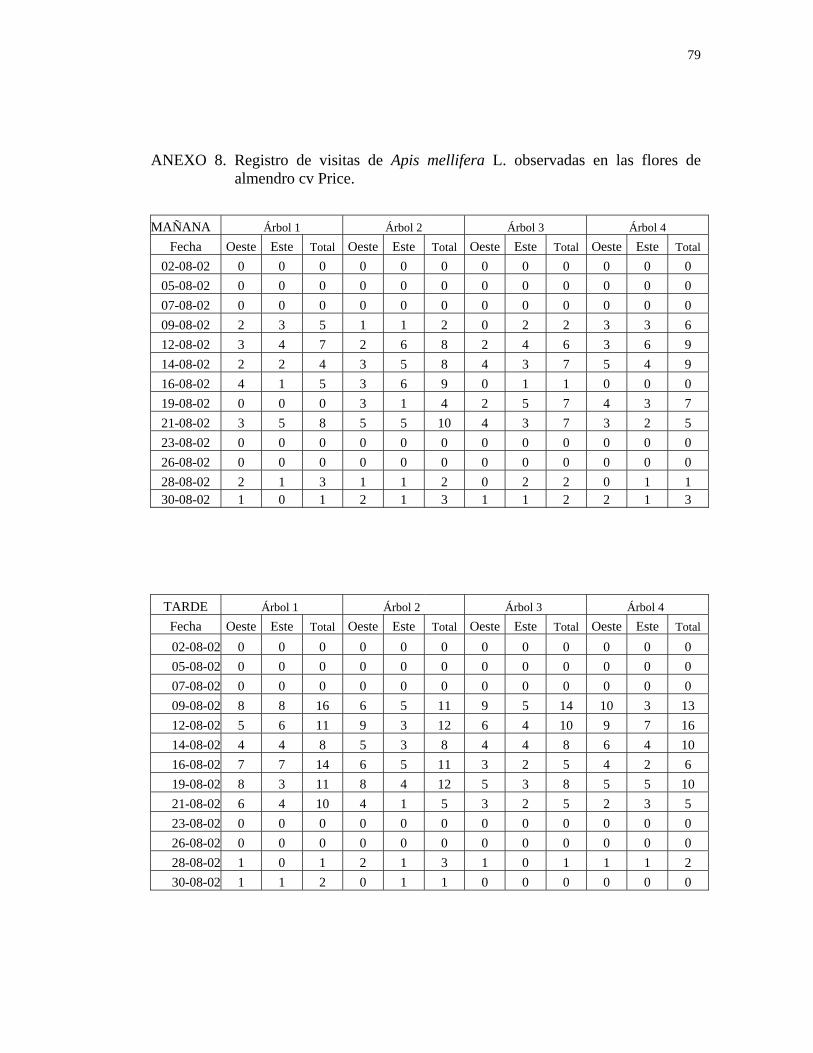
79
ANEXO 8. Registro de visitas de Apis mellifera L. observadas en las flores de almendro cv Price.
MAÑANA Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Oeste Este Total Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 0 0 0 09-08-02 2 3 5 1 1 2 0 2 2 3 3 6 12-08-02 3 4 7 2 6 8 2 4 6 3 6 9 14-08-02 2 2 4 3 5 8 4 3 7 5 4 9 16-08-02 4 1 5 3 6 9 0 1 1 0 0 0 19-08-02 0 0 0 3 1 4 2 5 7 4 3 7 21-08-02 3 5 8 5 5 10 4 3 7 3 2 5 23-08-02 0 0 0 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 0 0 0 28-08-02 2 1 3 1 1 2 0 2 2 0 1 1 30-08-02 1 0 1 2 1 3 1 1 2 2 1 3
TARDE Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Oeste Este Total Oeste Este Total Oeste Este Total Oeste Este Total 02-08-02 0 0 0 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 0 0 0 09-08-02 8 8 16 6 5 11 9 5 14 10 3 13 12-08-02 5 6 11 9 3 12 6 4 10 9 7 16 14-08-02 4 4 8 5 3 8 4 4 8 6 4 10 16-08-02 7 7 14 6 5 11 3 2 5 4 2 6 19-08-02 8 3 11 8 4 12 5 3 8 5 5 10 21-08-02 6 4 10 4 1 5 3 2 5 2 3 5 23-08-02 0 0 0 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 0 0 0 28-08-02 1 0 1 2 1 3 1 0 1 1 1 2 30-08-02 1 1 2 0 1 1 0 0 0 0 0 0
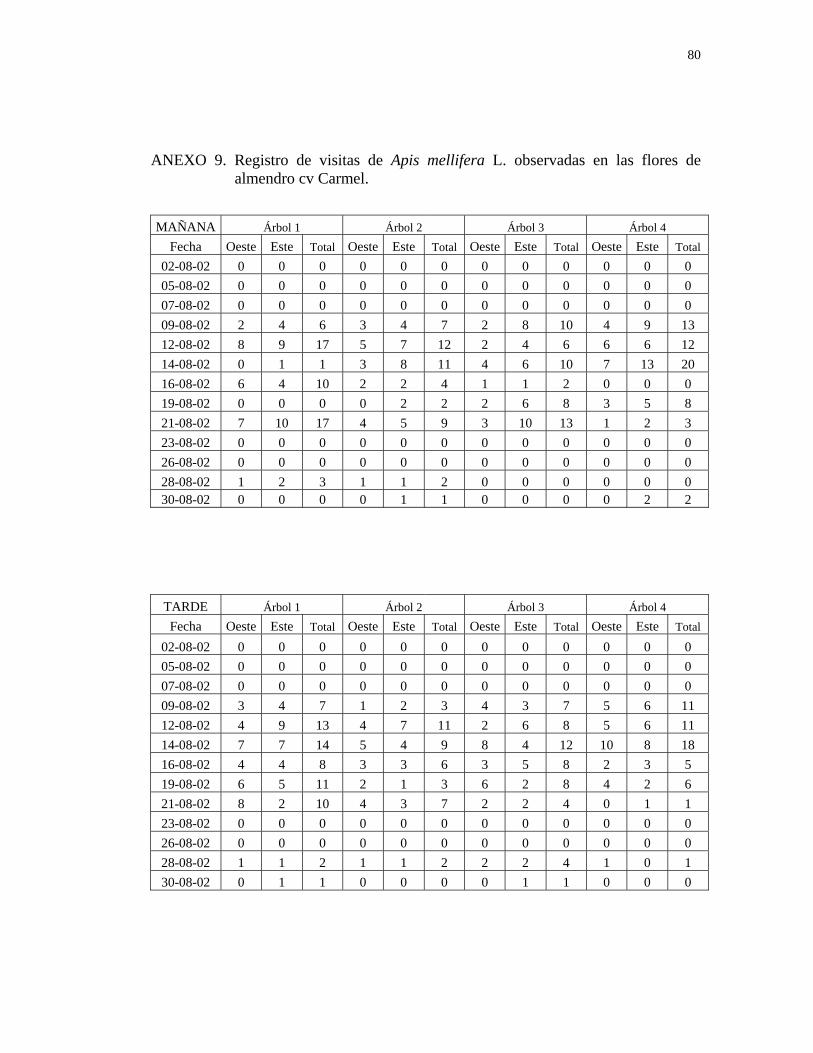
80
ANEXO 9. Registro de visitas de Apis mellifera L. observadas en las flores de almendro cv Carmel.
MAÑANA Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Oeste Este Total Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 0 0 0 09-08-02 2 4 6 3 4 7 2 8 10 4 9 13 12-08-02 8 9 17 5 7 12 2 4 6 6 6 12 14-08-02 0 1 1 3 8 11 4 6 10 7 13 20 16-08-02 6 4 10 2 2 4 1 1 2 0 0 0 19-08-02 0 0 0 0 2 2 2 6 8 3 5 8 21-08-02 7 10 17 4 5 9 3 10 13 1 2 3 23-08-02 0 0 0 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 0 0 0 28-08-02 1 2 3 1 1 2 0 0 0 0 0 0 30-08-02 0 0 0 0 1 1 0 0 0 0 2 2
TARDE Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Oeste Este Total Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 0 0 0 09-08-02 3 4 7 1 2 3 4 3 7 5 6 11 12-08-02 4 9 13 4 7 11 2 6 8 5 6 11 14-08-02 7 7 14 5 4 9 8 4 12 10 8 18 16-08-02 4 4 8 3 3 6 3 5 8 2 3 5 19-08-02 6 5 11 2 1 3 6 2 8 4 2 6 21-08-02 8 2 10 4 3 7 2 2 4 0 1 1 23-08-02 0 0 0 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 0 0 0 28-08-02 1 1 2 1 1 2 2 2 4 1 0 1 30-08-02 0 1 1 0 0 0 0 1 1 0 0 0
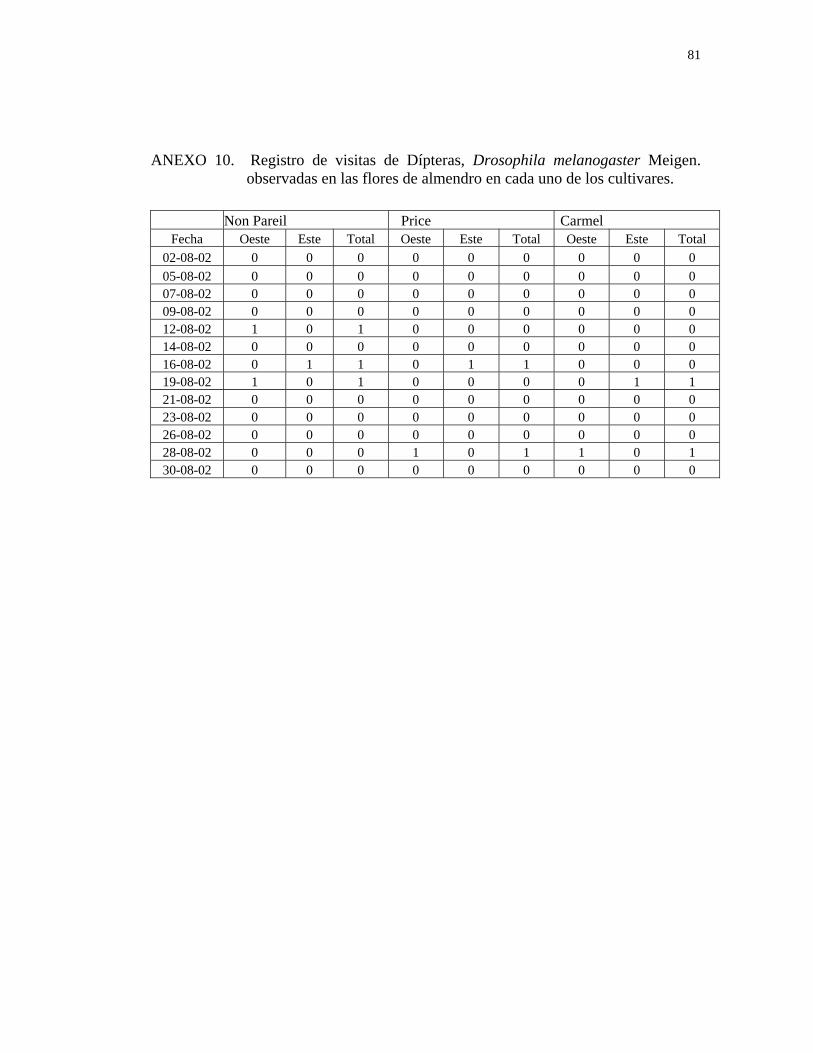
81
ANEXO 10. Registro de visitas de Dípteras, Drosophila melanogaster Meigen. observadas en las flores de almendro en cada uno de los cultivares.
Non Pareil Price Carmel Fecha Oeste Este Total Oeste Este Total Oeste Este Total
02-08-02 0 0 0 0 0 0 0 0 0 05-08-02 0 0 0 0 0 0 0 0 0 07-08-02 0 0 0 0 0 0 0 0 0 09-08-02 0 0 0 0 0 0 0 0 0 12-08-02 1 0 1 0 0 0 0 0 0 14-08-02 0 0 0 0 0 0 0 0 0 16-08-02 0 1 1 0 1 1 0 0 0 19-08-02 1 0 1 0 0 0 0 1 1 21-08-02 0 0 0 0 0 0 0 0 0 23-08-02 0 0 0 0 0 0 0 0 0 26-08-02 0 0 0 0 0 0 0 0 0 28-08-02 0 0 0 1 0 1 1 0 1 30-08-02 0 0 0 0 0 0 0 0 0
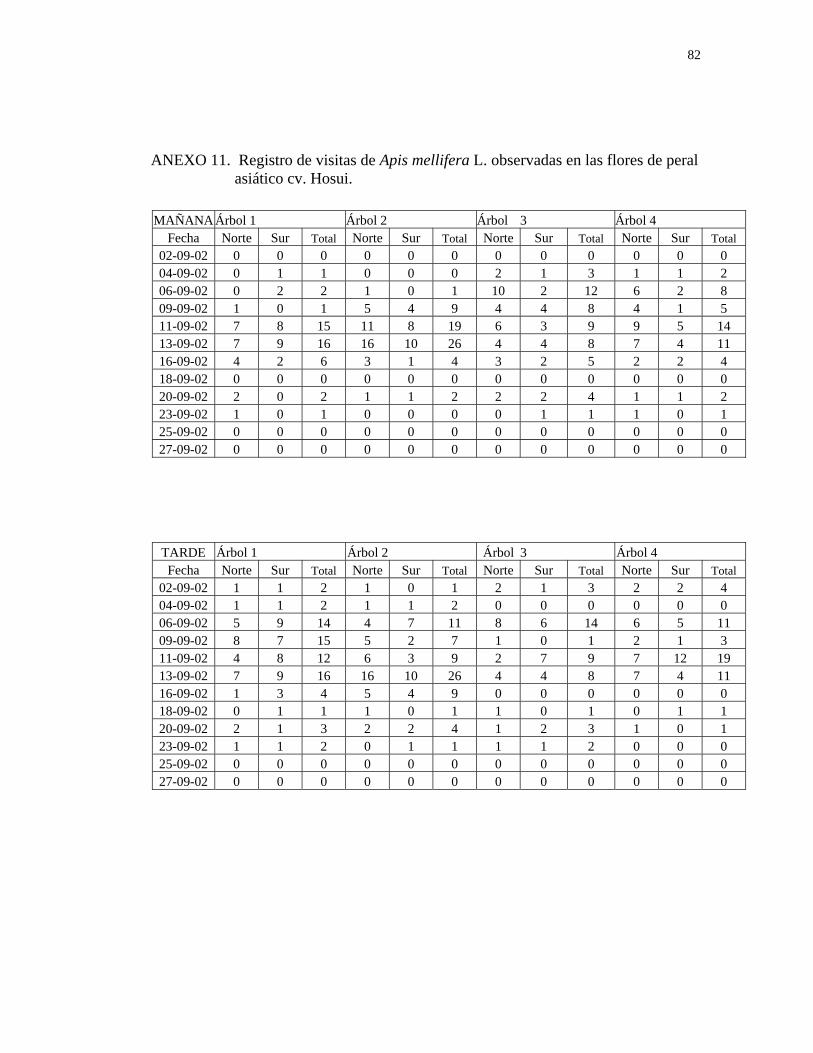
82
ANEXO 11. Registro de visitas de Apis mellifera L. observadas en las flores de peral asiático cv. Hosui.
MAÑANA Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Norte Sur Total Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 0 0 0 0 0 0 0 0 0 0 0 0 04-09-02 0 1 1 0 0 0 2 1 3 1 1 2 06-09-02 0 2 2 1 0 1 10 2 12 6 2 8 09-09-02 1 0 1 5 4 9 4 4 8 4 1 5 11-09-02 7 8 15 11 8 19 6 3 9 9 5 14 13-09-02 7 9 16 16 10 26 4 4 8 7 4 11 16-09-02 4 2 6 3 1 4 3 2 5 2 2 4 18-09-02 0 0 0 0 0 0 0 0 0 0 0 0 20-09-02 2 0 2 1 1 2 2 2 4 1 1 2 23-09-02 1 0 1 0 0 0 0 1 1 1 0 1 25-09-02 0 0 0 0 0 0 0 0 0 0 0 0 27-09-02 0 0 0 0 0 0 0 0 0 0 0 0
TARDE Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Norte Sur Total Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 1 1 2 1 0 1 2 1 3 2 2 4 04-09-02 1 1 2 1 1 2 0 0 0 0 0 0 06-09-02 5 9 14 4 7 11 8 6 14 6 5 11 09-09-02 8 7 15 5 2 7 1 0 1 2 1 3 11-09-02 4 8 12 6 3 9 2 7 9 7 12 19 13-09-02 7 9 16 16 10 26 4 4 8 7 4 11 16-09-02 1 3 4 5 4 9 0 0 0 0 0 0 18-09-02 0 1 1 1 0 1 1 0 1 0 1 1 20-09-02 2 1 3 2 2 4 1 2 3 1 0 1 23-09-02 1 1 2 0 1 1 1 1 2 0 0 0 25-09-02 0 0 0 0 0 0 0 0 0 0 0 0 27-09-02 0 0 0 0 0 0 0 0 0 0 0 0

83
ANEXO 12. Registro de visitas de Apis mellifera L. observadas en las flores de peral asiático cv. Shinseiki.
MAÑANA Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Norte Sur Total Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 0 0 0 0 0 0 0 0 0 0 0 0 04-09-02 0 0 0 0 0 0 0 0 0 1 0 1 06-09-02 0 0 0 1 0 1 0 0 0 1 2 3 09-09-02 1 0 1 1 2 3 2 1 3 1 1 2 11-09-02 4 3 7 7 3 10 4 4 8 5 4 9 13-09-02 2 2 4 3 2 5 1 1 2 3 3 6 16-09-02 4 2 6 2 2 4 3 4 7 6 8 14 18-09-02 0 1 1 0 0 0 0 1 1 0 0 0 20-09-02 6 4 10 4 7 11 3 6 9 1 2 3 23-09-02 2 2 4 2 1 3 1 0 1 2 1 3 25-09-02 2 0 2 0 0 0 0 0 0 0 0 0 27-09-02 0 1 1 0 1 1 0 0 0 0 1 1
TARDE Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Norte Sur Total Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 0 0 0 0 0 0 0 0 0 0 0 0 04-09-02 0 0 0 2 0 2 1 0 1 1 0 1 06-09-02 0 1 1 2 2 4 1 0 1 2 1 3 09-09-02 2 0 2 1 1 2 0 0 0 1 1 2 11-09-02 3 0 3 3 3 6 3 6 9 14 12 26 13-09-02 3 3 6 2 5 7 2 4 6 9 6 15 16-09-02 6 7 13 4 3 7 2 0 2 0 1 1 18-09-02 3 0 3 1 0 1 0 0 0 1 1 2 20-09-02 3 6 9 2 3 5 2 7 9 3 3 6 23-09-02 2 1 3 2 2 4 4 1 5 2 1 3 25-09-02 1 0 1 1 2 3 0 1 1 1 1 2 27-09-02 1 0 1 0 0 0 0 0 0 0 0 0

84
ANEXO 13. Registro de visitas de Apis mellifera L. observadas en las flores de peral asiático cv. Nijisseiki.
MAÑANA Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Norte Sur Total Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 0 1 1 0 0 0 0 0 0 1 0 1 04-09-02 1 2 3 0 0 0 0 1 1 2 2 4 06-09-02 0 2 2 0 1 1 0 2 2 9 5 14 09-09-02 2 4 6 2 2 4 2 1 3 7 15 22 11-09-02 6 8 14 8 8 16 9 8 17 13 8 21 13-09-02 7 3 10 5 5 10 6 4 10 4 4 8 16-09-02 4 1 5 3 2 5 3 3 6 2 1 3 18-09-02 0 1 1 1 0 1 0 0 0 0 1 1 20-09-02 1 1 2 0 3 3 1 1 2 1 2 3 23-09-02 1 1 2 0 1 1 1 1 2 0 0 0 25-09-02 0 0 0 2 0 2 0 0 0 0 0 0 27-09-02 0 0 0 0 0 0 0 0 0 0 0 0
TARDE Árbol 1 Árbol 2 Árbol 3 Árbol 4 Fecha Norte Sur Total Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 0 0 0 1 0 1 2 0 2 1 1 2 04-09-02 1 1 2 0 1 1 0 0 0 0 0 0 06-09-02 2 4 6 3 2 5 2 1 3 10 15 25 09-09-02 7 8 15 0 1 1 2 1 3 3 0 3 11-09-02 0 0 0 3 3 6 3 5 8 9 4 13 13-09-02 6 5 11 4 2 6 9 2 11 5 3 8 16-09-02 3 1 4 2 2 4 1 1 2 1 0 1 18-09-02 0 0 0 4 0 4 1 1 2 1 2 3 20-09-02 2 1 3 3 2 5 2 2 4 1 1 2 23-09-02 1 1 2 2 1 3 1 0 1 0 0 0 25-09-02 0 1 1 1 0 1 0 0 0 0 0 0 27-09-02 0 0 0 0 0 0 0 0 0 0 0 0
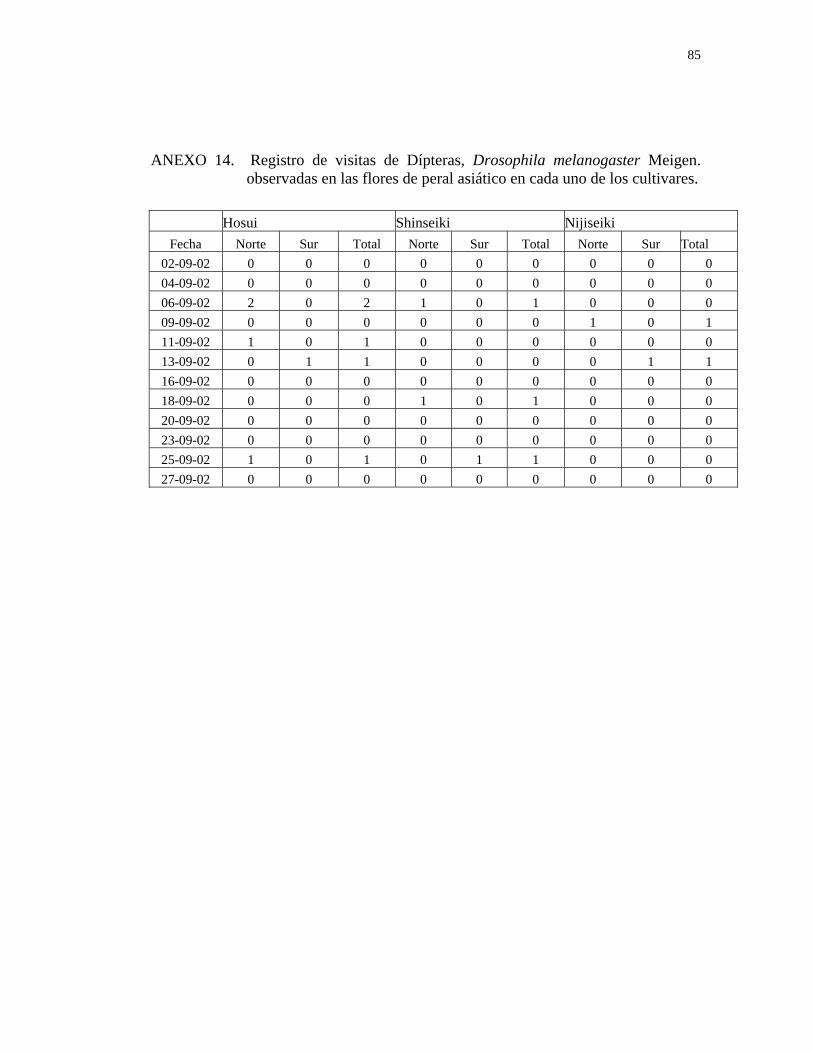
85
ANEXO 14. Registro de visitas de Dípteras, Drosophila melanogaster Meigen. observadas en las flores de peral asiático en cada uno de los cultivares.
Hosui Shinseiki Nijiseiki Fecha Norte Sur Total Norte Sur Total Norte Sur Total
02-09-02 0 0 0 0 0 0 0 0 0 04-09-02 0 0 0 0 0 0 0 0 0 06-09-02 2 0 2 1 0 1 0 0 0 09-09-02 0 0 0 0 0 0 1 0 1 11-09-02 1 0 1 0 0 0 0 0 0 13-09-02 0 1 1 0 0 0 0 1 1 16-09-02 0 0 0 0 0 0 0 0 0 18-09-02 0 0 0 1 0 1 0 0 0 20-09-02 0 0 0 0 0 0 0 0 0 23-09-02 0 0 0 0 0 0 0 0 0 25-09-02 1 0 1 0 1 1 0 0 0 27-09-02 0 0 0 0 0 0 0 0 0

86
ANEXO 15. Correlaciones entre número de abejas, temperatura y número de flores abiertas en los tres cultivares de almendro.
Variables Non Pareil Price Carmel N° de abejas totales/T° mañana 0,834 0,704 0,783 N° de abejas totales /T° tarde 0,759 0,748 0,718 N° de abejas totales/ n° de flores totales 0,735 0,715 0,838 N° de abejas cara oeste/T° mañana 0,778 0,628 0,757 N° abejas cara oeste/T° tarde 0,842 0,734 0,588 N° de abejas cara este/T° mañana 0,801 0,733 0,778 N° abejas cara este/T° tarde 0,656 0,762 0,793
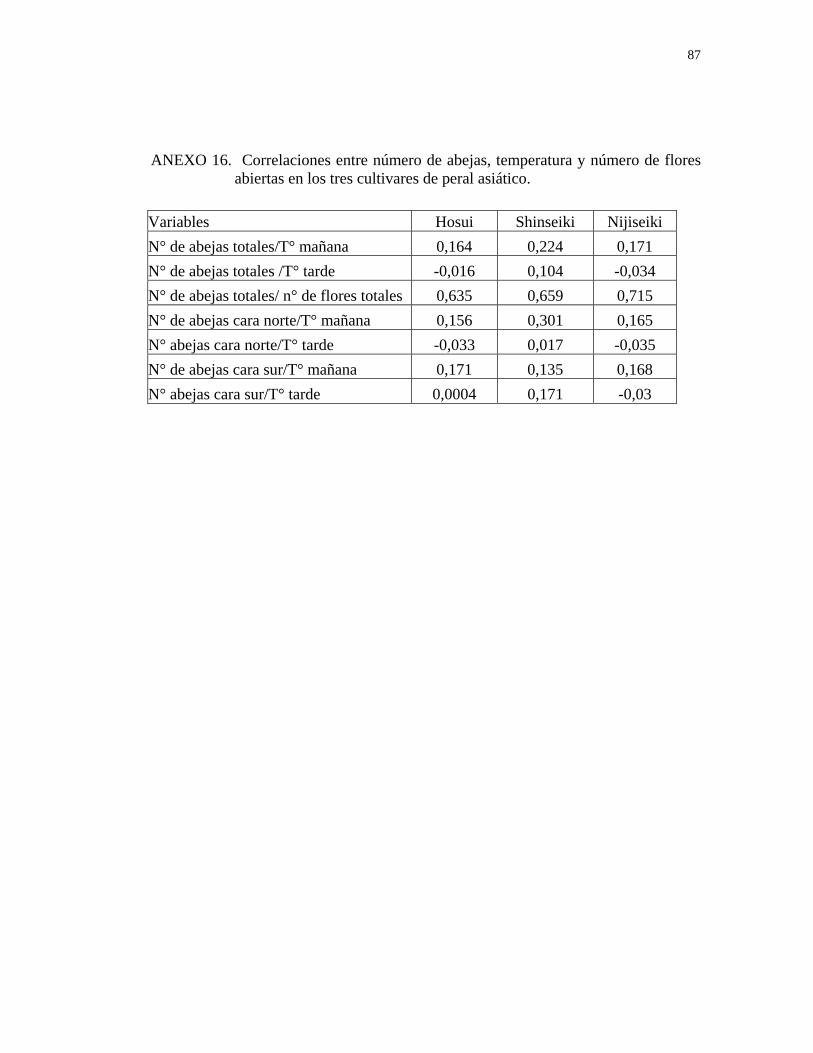
87
ANEXO 16. Correlaciones entre número de abejas, temperatura y número de flores abiertas en los tres cultivares de peral asiático.
Variables Hosui Shinseiki Nijiseiki N° de abejas totales/T° mañana 0,164 0,224 0,171 N° de abejas totales /T° tarde -0,016 0,104 -0,034 N° de abejas totales/ n° de flores totales 0,635 0,659 0,715 N° de abejas cara norte/T° mañana 0,156 0,301 0,165 N° abejas cara norte/T° tarde -0,033 0,017 -0,035 N° de abejas cara sur/T° mañana 0,171 0,135 0,168 N° abejas cara sur/T° tarde 0,0004 0,171 -0,03

88
ANEXO 17. Registro de temperatura y visitas durante, mañana y tarde en los tres cultivares de almendro.
Mañana Temperatura Non Pareil Price Carmel 02-08-02 14,8 0 0 0 05-08-02 10,9 0 0 0 07-08-02 15,7 0 0 0 09-08-02 20,2 40 15 36 12-08-02 22,7 58 30 47 14-08-02 19,2 25 28 42 16-08-02 13,7 11 15 16 19-08-02 13,9 16 18 18 21-08-02 18 34 30 42 23-08-02 12,6 0 0 0 26-08-02 14 0 0 0 28-08-02 12,9 11 8 5 30-08-02 18,4 8 9 3
Tarde Temperatura Non Pareil Price Carmel 02-08-02 16,7 0 0 0 05-08-02 11,4 0 0 0 07-08-02 13,9 0 0 0 09-08-02 25,5 46 54 28 12-08-02 28,5 34 49 43 14-08-02 21,9 43 34 53 16-08-02 17,2 33 36 27 19-08-02 15,7 27 41 28 21-08-02 22,7 20 25 22 23-08-02 13,5 0 0 0 26-08-02 15,5 0 0 0 28-08-02 15,5 13 7 9 30-08-02 19,9 15 3 2
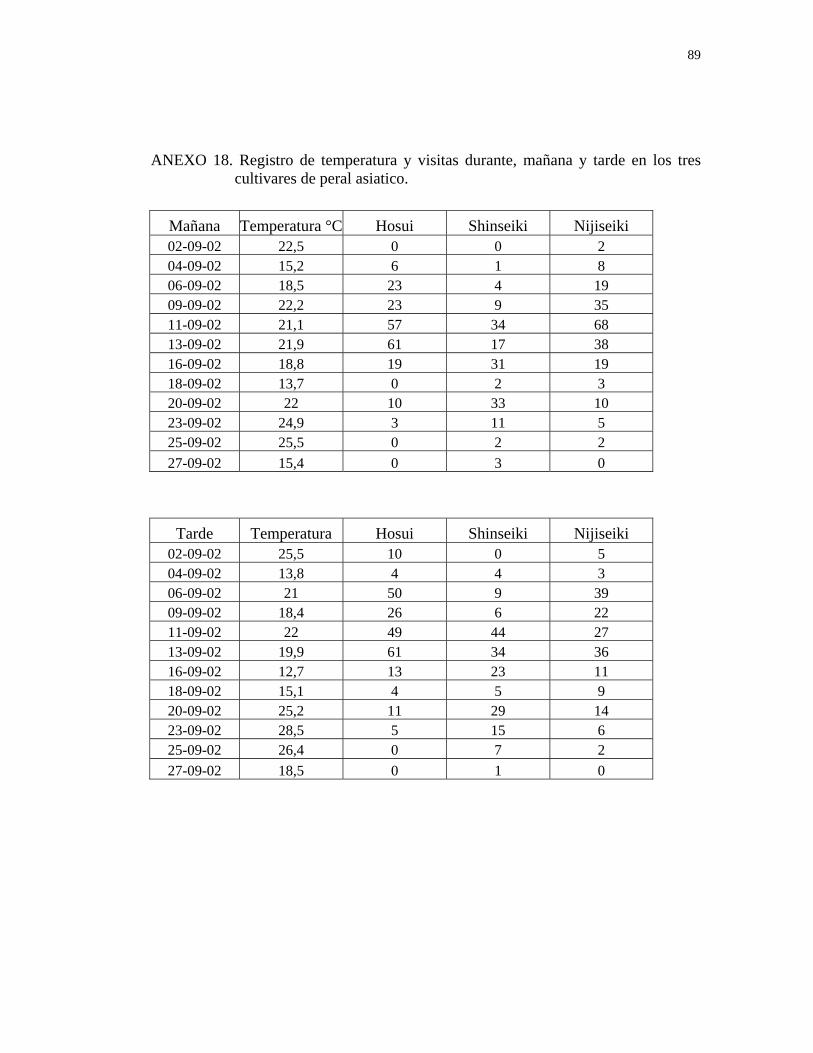
89
ANEXO 18. Registro de temperatura y visitas durante, mañana y tarde en los tres cultivares de peral asiatico.
Mañana Temperatura °C Hosui Shinseiki Nijiseiki 02-09-02 22,5 0 0 2 04-09-02 15,2 6 1 8 06-09-02 18,5 23 4 19 09-09-02 22,2 23 9 35 11-09-02 21,1 57 34 68 13-09-02 21,9 61 17 38 16-09-02 18,8 19 31 19 18-09-02 13,7 0 2 3 20-09-02 22 10 33 10 23-09-02 24,9 3 11 5 25-09-02 25,5 0 2 2 27-09-02 15,4 0 3 0
Tarde Temperatura Hosui Shinseiki Nijiseiki 02-09-02 25,5 10 0 5 04-09-02 13,8 4 4 3 06-09-02 21 50 9 39 09-09-02 18,4 26 6 22 11-09-02 22 49 44 27 13-09-02 19,9 61 34 36 16-09-02 12,7 13 23 11 18-09-02 15,1 4 5 9 20-09-02 25,2 11 29 14 23-09-02 28,5 5 15 6 25-09-02 26,4 0 7 2 27-09-02 18,5 0 1 0







