
Anatomy of the cochlea
Lecture 7

2
Note in particular◦ Insertion of stereocilia into cuticular plate◦ Basal body (rudimentary kinocilium)◦ OHCs contain extensive subsurface cisternae and
intracellular structures concentrated around the edges of the cell(In contrast with IHCs)
◦ Afferent and efferent nerve endings around base of cells
Structure of OHCs & IHCs (see figs)

Kinocilia are found on the apical surface of hair cells and are involved in both the morphogenesis of the hair bundle and mechanotransduction. Vibrations (either by movement or sound waves) cause displacement of the hair bundle, resulting in depolarization or hyperpolarization of the hair cell. The depolarization of the hair cells in both instances causes signal transduction via neurotransmitter release.
Kinocilia

4
OHC

5
(This could be an error – efferents are thought to terminate on afferent endings rather than directly on IHC)
IHC

6
Perilymph◦ Similar in composition to other extra-cellular fluids
& CSF◦ Cochlear aqueduct connects ST (near base) to
subarachnoid space
Are some differences in composition
◦ Very high in Na+, very low in K+ ions◦ Source – probably blood vessels within cochlea
Cochlear fluids

7
Endolymph◦ Similar in composition to intra-cellular fluids
(unusual)◦ High in K+, low in Na+ ions◦ Source unclear – perhaps derived from perilymph
Stria vascularis plays important role in maintaining ionic composition

8
Note that reticular lamina (not BM) forms the boundary between endolymph & perilymph in ST
◦Hence the distinction between scala media (bounded by BM) and endolymphatic space
◦While hair bundles are exposed to endolymph, lateral walls & bases are bathed in perilymph

9
Stapes vibrations set up initial pressure waves in SV ◦ Correspond to normal propagation of sound in fluid
Very fast, longitudinal waves◦ Wavelength much greater than dimensions of
scala vestibuli => instantaneous fluid pressure is more or less the same throughout SV
◦ Vibrations in SV do not pass through helicotrema (except for very low frequencies)
Cochlear physiology

10
Reissner’s membrane no impedance barrier – ‘acoustically transparent’
Result is a pressure differential across BM/organ of Corti complex, causing it to vibrate between points of attachment

11
Note – Reissner’s membrane probably does not vibrate with same pattern as (parallel to) BM as suggested in the figure
Fluid pressure eventually released at (flexible) round window

12
Although the initial fluid pressure at any instant is the same throughout SV,
... the BM does not simply vibrate as one unit in response to pressure differential across it
Rather, interaction between fluid pressures and BM mechanics results in a wave of displacement of BM◦ Wave appears to travel (relatively slowly) from
base to apex of cochlea
The travelling wave
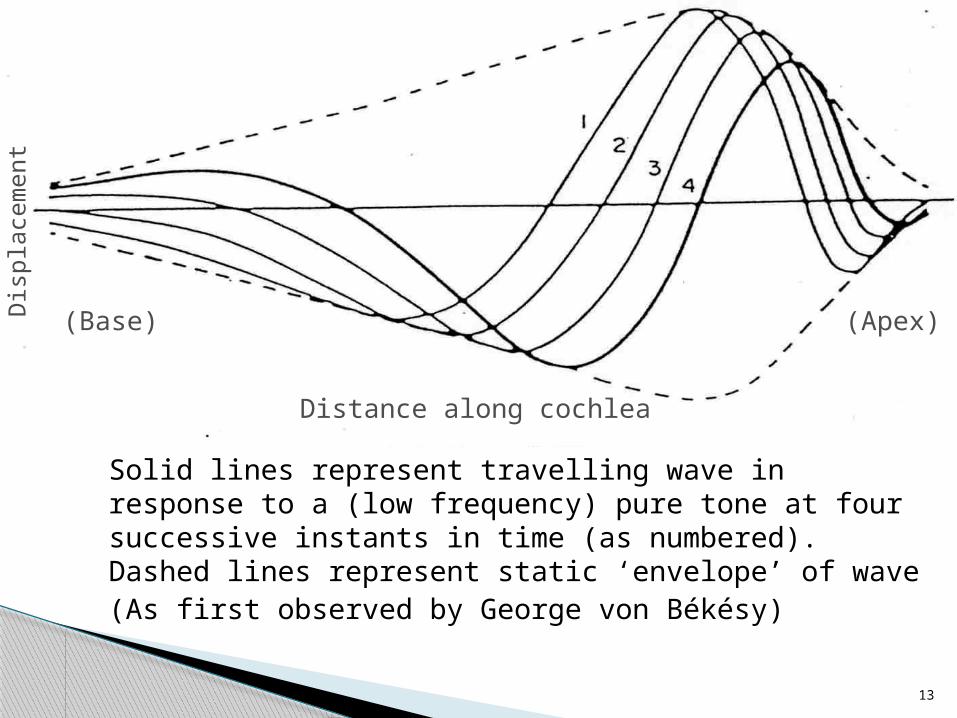
13
Solid lines represent travelling wave in response to a (low frequency) pure tone at four successive instants in time (as numbered). Dashed lines represent static ‘envelope’ of wave(As first observed by George von Békésy)
Distance along cochlea
(Base) (Apex)Dis
plac
emen
t

14
◦ Entire BM/organ of Corti complex (also TeM) vibrates
◦ Can be regarded as a transverse wave, similar to waves (ripples) on surface of water
◦ Direction of travel results from decrease in BM stiffness (that accompanies increase in its width) from base to apex Wave appears to travel from base to apex regardless
of whether the stimulus is applied at the base of cochlea or elsewhere

15
Wave slows and builds in amplitude as it travels, then peaks and decays almost immediately
For a particular signal frequency, waves are contained within ‘static’ envelope (dotted lines in figure)
Position of peak along BM depends uniformly on signal frequency – place-frequency map
See also animations at www reference!
Key features of the travelling wave

16
For high frequency signals, maximum BM response (peak of travelling wave envelope) is near the base of the cochlea
Lower frequencies peak towards apex Also due primarily to BM stiffness decrease
from base to apex
The place-frequency map

17

18
Response (vibration) at a particular site is also extremely sharply ‘tuned’ to this best (‘characteristic’) frequency◦ (If you change frequency slightly, the response
drops dramatically) Sharply tuned place-frequency map is a
primary basis of frequency selectivity in the auditory system◦ (Perhaps not at low frequencies)
Response for a particular frequency spreads much more into adjacent ‘higher-frequency’ (basal) regions than ‘lower-frequency’ (apical) regions

19
‘Vertical’ displacements of BM/organ of Corti, TeM give relative shear between reticular lamina & TeM◦ Points at which BM, TeM ‘hinged’ are important
… which causes HC stereocilia (hair bundles) to be deflected
Stimulation of the hair cells

20

21
Classical view –◦ Hair bundles defelected away from modiolus for
BM upward associated with excitation◦ Towards modiolus for BM downward compresses
the tip links and is associated with inhibition◦ Relationship between BM displacement & HC
bundle deflection is evidently highly complex

22
Achieved by IHCs◦ Are ‘true receptor cells’◦ (Vast majority of NVIII afferents synapse with
them) Endolymph is at a steady potential (voltage)
of ~ +80 mV (relative to perilymph)◦ The endocochlear potential (EP)◦ Maintained by metabolic ion-pumping mechanism
of stria vascularis
Stereociliary deflections to neural discharges (see figure)

23

24
HCs (like most cells) also maintain a steady internal potential◦ The intracellular (resting) potential◦ Maintained by cellular ion transfer processes◦ ~ –45 mV in IHCs◦ ~ –70 mV in OHCs

25
Combination of potentials acts as a ‘battery’◦ Recall that stereocilia contains ion channels
Certain percentage are always open◦ ‘Battery’ drives a steady (‘DC’) ionic current
(mainly K+) from endolymph, through ion channels, into HC (and out through basolateral wall)
However, it’s clear that deflection of stereocilia is the mechanical input to the HCs◦ OHC bundles – directly coupled to TeM◦ IHC bundles – probably via intervening fluid only◦ Recall rigidity of stereocilia – ‘pivot’ at point of
attachment to HC, rather than ‘bend’ (as often described)

26
In quiet, the system is in equilibrium – essentially steady (DC) potentials, currents
Deflections of hair bundles thought to open or close ion channels◦ Cross-links at tips of stereocilia physically them
pull open?!◦ Cause variations in ion currents flowing
through cell
Mechano-electrical transduction

27
Deflection of hair bundle away from modiolus◦ Opens ion channels◦ Increases current into HC◦ Depolarises cell
(Resting potential heads back towards zero)
HC resting potential (negatively polarised)
Influx of K+ ions depolarises cell
0 V
+
–

28
Depolarisation of IHCs causes release of neurotransmitter at base of cell◦ Neurotransmitter probably glutamate◦ Ca2+ ions probably mediate release◦ Initiates (increases likelihood of) firing of
primary afferent neurons And conversely, deflection of bundle
towards modiolus … reduces likelihood of firing of primary afferents

29
Can be steady (DC) or fluctuating (AC) over time
‘Resting’ potentials (DC)◦ Endocochlear potential (+80 mV)◦ HC intracellular resting potentials
HC receptor potentials◦ See figure – responses to 50-ms tone bursts (at
various frequencies as marked)◦ Receptor potential is the intracellular voltage
change in response to sound
Cochlear potentials

30
Hair cell receptor potentials

31
Have both a fluctuating (AC) and a steady (DC) component◦ AC component broadly follows waveform of
stimulus (see upper traces of figure)◦ But the average amplitude of the AC waveform is
not the same as when the stimulus is off (at the end of the burst)
◦ The slight positive shift in the average AC response is equivalent to a DC (steady) component superimposed on it
Receptor potentials

32
◦ Note that DC component is depolarising (positive-going)(Recall that in IHCs, depolarisation causes neurotransmitter release)
◦ Also note that the AC component of the receptor potential gets smaller with increasing stimulus frequency & the DC relatively larger

33
Massed effects of electrical activity of individual cells. Can be recorded at various sites in & around cochlea. (Potentially clinically useful)◦ Cochlear microphonic – voltage recorded as
extracellular correlate of AC component of receptor currents/potentials. Predominantly due to activity of OHCs
◦ Summating potential – extracellular correlate of DC component of receptor currents. Due to both IHCs & OHCs(?)
Extracellular evoked potentials (see figure)

34

35
◦ Compound action potential – summed activity of a number primary afferent neurons firing together at the onset of a stimulus Typically recorded in response to clicks or tone-bursts Distinct negative-going peak within ~ 1 ms of
transient stimulus, denoted N1 (secondary N2 peak sometimes seen)







