Download - ESTUARINE ScienceDirect

&
A vailable online a t w w w .sciencedirect.comi föS* ESTUARIN E
ScienceDirect c o a s t a l
ELSEVIER E stu a rin e , C oasta l and S helf S cience 71 (2007) 580—592
AND
S H E L F SC IEN C Ew w w .elsev ier.com /locate /ecss
Long-term changes in summer phytoplankton com m unities of the open northern Baltic Sea
Sanna Suikkanen*, Maria Laamanen, Maija H u t tunen
F innish In stitu te o f M arine Research , P .O . B ox 2, FI-00561 H elsink i, F in la n d
R eceived 5 June 2006; accepted 6 S ep tem ber 2006 A vailable online 17 O ctober 2006
A bstract
Changes in the b iom ass and species com position o f phytoplankton m ay reflect m ajor sh ifts in environm ental cond itions. W e investigated relationships betw een the late sum m er b iom ass o f different phytoplankton taxa and environm ental facto rs , and th e ir long -te rm (1979—2003) trends in tw o areas o f the B altic Sea, the northern Baltic proper (NBP) and the G ulf o f Finland (G F ), w ith s ta tis tica l analyses. An increasing trend was found in late sum m er tem peratu re and chlorophyll a o f the surface w ater layer (0—10 m ) in both areas. T h e re w as also a significant decrease in sum m er salinity and an increase in w inter dissolved inorganic nitrogen to phosphorus (D IN :D IP) ratio in the N BP, as well as in creases in w in ter DIN concen tra tions and D IN ;S i0 4 ratio in the GF. Simultaneously, the b iom ass o f chrysophytes and chlorophytes increased in both areas. In the NBP, also the biom ass o f dinophytes increased and that of euglenophytes decreased , w hereas in th e GF, cyanobacteria increased and cryptophytes decreased . R edundancy analysis (RDA) indicated that sum m er tem pera tu re and w in ter D IN concentration were the most im portant factors with respect to changes in the phytoplankton com m unity structure. Thus, the phy top lank ton com m u n ities seem to reflect both hydrographic changes and the ongoing eu troph ication process in the northern Baltic Sea.© 2006 E lsevier Ltd. All rights reserved.
K eyw ords; p h y to p lan k to n ; B altic S ea; long -term c h a n g es; eu troph ica tion ; m ultivariate analysis
1. Introduction
P hy to p lan k to n fo rm th e e n erg e tic b a s is o f open sea food w ebs, and b ecause o f th e ir re la tiv e ly h igh grow th ra tes and d irect d ep en d en cy on bo th h y d ro g rap h ic fa c to rs and d issolved n u trien ts, p h y to p lan k to n sp ec ie s are im p o rtan t ind ica to rs of w a te r quality in p e lag ic a reas. H ow ever, due to both natural and m eth o d o lo g ical reaso n s , p h y to p la n k to n sp ec ie s co m position and ab u n d an ce d a ta a re h ig h ly v a riab le , and consequently th e ir use as w a te r q u a lity in d ic a to rs is u su a lly re stric ted and only ch lo ro p h y ll a c o n ce n tra tio n s are m o n ito red . A lso, due to dev elo p m en t in tax o n o m ica l c lass ifica tio n and m ethodology, su ffic ien tly long c o m p a ra b le tim e se ries o f phy top lank ton are rare.
* C o rrespond ing author.E -m a il a ddress; s an na.su ikkanen@ fim r.fi (S. S u ik k an en ),
0272 -7714 /$ - see fro n t m a tte r © 2 0 0 6 E lsev ie r L td . A ll righ ts reserved, doi; 10 .1016 /j.ecss.2006 .09 .004
D ue to co ncern o ver the p o llu tio n o f th e B a ltic Sea, the surrounding sta tes s ig n ed th e C o n v en tio n on the P ro tection o f the M arine E n v iro n m en t o f th e B altic S e a A re a in 1974. To im prove the k n o w led g e on th e lo n g -te rm c h a n g e s in the factors re la ted to eu tro p h ica tio n and p o llu tio n o f th e sea , the Baltic M arine E nv ironm en t P ro tec tio n C o m m iss io n (H elsinki C om m ission , H E L C O M ) has been c o o rd in a tin g a jo in t m onitoring p rogram m e o f the B a ltic S ea (B M P, la te r C O M B IN E ) since 1979. D ata on p h y sica l, ch em ica l and b io lo g ic a l param eters o f the open sea h ave now been c o lle c ted fo r o v e r 25 years. A c co rd ing to the la test a sse ssm en t o f th e s ta te o f the Baltic Sea, covering years 1 9 9 4 -1 9 9 8 (H E L C O M , 2 0 0 2 ) , eu trophication is still a m ajo r p rob lem in m o st su b -a rea s , d e sp ite the successful reductions in p o in t-so u rce n u tr ie n t lo ad in g .
T h e typ ica l annual p h y to p la n k to n su c c ess io n in the open northern B altic Sea is ch a ra c te rised by a w in te r biom ass m in im um stage, a v e rn al b loom o f d ia to m s a n d d inofiagellates that d ep le tes th e in o rg an ic n u trien ts in th e e u p h o tic layer and leads
s u b ito e .V . l ic e n s e d c u s to m e r c o p y s u p p lie d a n d p r in te d fo r F la n d e r s M a rin e in s ti tu te L ib rary (S L I0 5 X 0 0 2 2 5 E )

5. Su ikkanen et a l. ! E s tu a r in e . C o asta l a n d S h e l f Sc ience 71 (201)71 5 8 0 —5 9 2 581
to a su m m er m in im um stag e, a la te su m m er m ax im u m , m ain ly f o rm e d by cy an o b acte ria , and a la te au tu m n b io m a ss decline (N ie m i, 1973). T h is p ap er fo c u ses on the n u tr ie n t-p o o r late s u m m e r (July—A ugust) p e rio d , w h en h igh w ater tem p era tu res a n d stab le w eather co n d itio n s fa v o u r th e d ev elopm en t o f n itro g e n -fix in g cy an o b acte ria , an d m ass-o ccu rren ces fo rm ed by A p h a n izo m e n o n flo s -a q u a e , N o d u la r ia spum igena an d A n a b a e n a spp. occu r frequen tly . In a d d itio n to th e filam entous a n d co lon ia l cy an o b acte ria , th e la te su m m er co m m u n ity also ty p ic a lly con tains several sp ec ies o f c ry p to p h y tes , d ino flagel- la te s , chrysophytes, e u g len o p h y tes an d ch lo ro p h y tes (K ono- n e n , 1988).
T h e aim of o u r study w as to u se th e H E L C O M m o n ito rin g d a ta to determ ine if long-te rm c h an g e s in hydro g rap h y and n u t r ie n t conditions are re flected in th e la te sum m er p h y to p la n k to n com m unity . W e evalu a ted th e 197 9 —2003 d a ta se t fo r the la te sum m er p e rio d , c o n sistin g o f q u an tita tiv e p h y to p lan k to n o b se rv a tio n s , a s w ell as severa l en v iro n m en ta l fac to rs in tw o s u b -a re a s o f th e northern B altic S ea ; th e northern B altic p ro p er (N B P ) and the G u lf o f F in lan d (G F ). In a recen t an a ly sis , W as- m u n d and U hlig (2003) used th e B M P data in an in v estiga tion o f p h y to p lan k to n trends in th e so u th e rn and c en tra l B a ltic Sea. In con trast, lo n g -te rm p h y to p la n k to n co m m unity analyses h a v e no t been m ad e s in ce the en d o f th e 1980s fo r the northern B a lt ic Sea (K ononen and N iem i, 1984; K ononen, 1988).
2. M aterials and m ethods
2 .1 . S tu d y area
T h e Baltic S ea is a sem i-e n c lo se d , b rack ish w a te r se a (ca. 4 2 2 0 0 0 km 2, m ean dep th 55 m ), w ith a very re stric ted co n n ec t io n to the N orth S ea v ia the D a n ish stra its. It is ch arac te rized b y s tro n g seasonal tem p era tu re v a ria tio n , as w ell as a north - s o u th surface sa lin ity g rad ien t, w ith su rface w a ter sa lin ities d e c re a s in g from ca. 20 in th e K a tteg a t to 1—2 in th e n o rth ern a n d easte rn areas. T he n o rth e rn B a ltic p ro p er c o m p rise s an a r e a o f 2 9 1 0 0 km 2 (Fig. 1). C o n n e c ted to th e N B P w ithou t a s i l l , the G u lf o f F in lan d (2 9 5 0 0 k m 2) is s itu a te d b e tw een F in la n d and E ston ia . It has a su rfa c e w a ter sa lin ity g rad ien t ra n g in g from ab o u t 7 in the w e st to fresh w a te r co n d itio n s in th e N eva Bay, R ussia . G F h as a large c a tch m e n t area (4 2 1 00 0 km 2) and is h e av ily in flu en ced by n u trien t load ing
from the surrounding ag ricu ltu ra l and urban areas, especia lly from the c ity o f St. P e te rsb u rg by th e ru n -o ff from the R iver N eva. The nutrient re se rv e s o f th e G F a re also affec ted by the phosphorus-rich B altic Sea d eep w aters flow ing free ly into the g u lf due to the s ill-free connection to th e B altic proper. T he G F rece ives tw o to th ree tim es the average B altic Sea external nutrient in p u ts re la tiv e to its surface area (H E L C O M , 2002), and it is thus considered as o n e o f the m ost eu trophicated su b -b asin s o f the B altic Sea.
T h e study area is stra tified , w ith a deep, perm anen t ha lo - cline (at a depth o f ca. 60 —80 m ) and a sum m er th erm ocline ( 1 0 - 2 0 m). O ur data o rig in a ted from five H E L C O M m onito ring sta tions (Fig. 1): th ree in the northern B altic p roper; H3 (58° 35 ' 00" N , 18° 14' 0 0 " E; bo ttom depth 450 m ), H2 (59° 0 2 ' 00", 21° 0 5 ' 00"; 166 m ) an d H I (59° 29 ' 0 0", 22° 54' 00"; 82 m ), and tw o in the G u lf o f F in land ; F3 (59° 51 ' 06", 24° 50 ' 03"; 77 m ) a n d F I (60° 04 ' 01", 26° 2 1 ' 00"; 66 m ).
2.2. D ata
T h e orig inal da ta c o lle c ted by th e B M P since 1979 consist o f observations o f b io lo g ica l and env ironm en ta l param eters m ade in d ifferen t tim es o f the year. T h e m onitoring p ro g ram m e focuses on q u an tifica tion o f sum m er phy top lank ton , w hich is why th e cu rren t p a p e r deals w ith th e nu trien t defic ien t late sum m er period, b e tw een the en d o f Ju ly and the beg inn ing o f Septem ber, 1979—2003. D ata fro m years 1982 (G F) and 1998—1999 (both sub -reg ions) a re lack ing , as the sam ples w ere taken outside th e tim e period o f in terest during those years.
T h e phytop lankton data co n sis ted o f 90 quantitative observa tions (50 from the N B P and 40 fro m the G F), re su ltin g in up to th ree observations per y e a r at e ach o f the sub-regions. In 1979—1997, the sam pling d ay s w ere d istribu ted be tw een 29 July and 3 Septem ber, w hereas in 2 0 0 0 —2003, sam ples w ere taken betw een 30 Ju ly and 20 A u g u st each year, due to a ch an g e in the sam pling schedu le . In tegra ted sam ples o f the u p p e r w ater layer (0—10 m ) w ere p reserv ed w ith acid L ugoi so lu tion , and analysed using an inverted m icroscope (U te- rm öhl, 1958). D uring m ost o f the s tudy period (1 9 8 2 —2003), only one person (M aija H uttunen) w as responsib le fo r species iden tification and counting , w h ich en su red the com parab ility
FinlandR ussia
• F I G u lf o f F in landS w ed en
N o rth e rn * B a ltic p ro p e r H I
Estonia
100 km
Fig. 1. L o ca tio n o f th e m o n ito r in g sta tions in the B a ltic Sea. T h e dashed line rep resen ts th e borders o f the tw o sub -a reas studied.
s u b i to e .V . l ic e n s e d c u s to m e r c o p y s u p p lie d a n d p r in te d fo r F la n d e r s M a rin e In s ti tu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )

582 S. S u ikkanen e t al. / E stuarine . C oasta l a n d S h e l f Sc ience 7 / (2 0 0 7 ) 5 8 0 - 5 9 2
o f the da ta . A lto g e th e r ca. 2 0 0 taxa, m o stly iden tified to sp e cies leve], w ere fo u n d in th e sam ples. T h e sp ec ies n o m en c la tu re is a cc o rd in g to H ä llfo rs (2 0 0 4 ). T h e b iom ass ca lcu la tions w ere m ad e a cc o rd in g to th e M an u a l fo r M arine M o n ito rin g in the C O M B IN E P ro g ra m m e o f H E L C O M (A nnex C -6; h ttp :// sea .h e lco m .fi/M o n as /C o m b in eM an u a l2 /co n ten ts ,h tm l). C h lo rophyll a c o n ce n tra tio n in sam p le s tak en from d iscrete dep ths w as m ea su re d sp e c tro p h o to m e tric a lly acco rd ing to H E L C O M (1988), an d av erag ed o v e r 0 —10 m .
T he en v ironm en ta l d a ta con sisted o f sim ultaneous m easu rem ents o f tem p era tu re and salin ity , as w ell as concentra tions o f ino rganic nu trien ts (phosp h a te-p h o sp h o ru s , n itrate, nitrite and am m o nium -n itrogen and silica te -silicon ) m easured at the sam e stations du rin g the p reced in g w in ter (N ovem ber—January). As there w ere often several n u trien t m easurem ents per station and winter, these w ere averag ed fo r the analysis. Long-term trends in surface w a ter n u trien t con cen tra tio n s are m ost reliably detected in w in ter sam ples because in w in te r season the least am ount o f nutrients are bou n d to b io lo g ica l m ateria l. In sum m er, concen trations o f inorgan ic n u trien ts, especia lly that of n itrogen, are often below the d e tection lim its . H ow ever, variability m ay be g reat due to e.g . o rg an ic aggrega tes and sporadic events such as upw elling . W e also ex am in ed early sum m er n u trien t concen trations in sam p les taken b e fo re th e phytop lankton sam pling , but the large variab ility o f these da ta , due to the reasons m entioned above, p reven ted th e ir in c lu sio n in th e analysis. T h e rationale fo r com paring w in tertim e n u trien t concen tra tions w ith sum m er phytop lankton d a ta w as th at w in te r n u trien t trends can b e expected to be linked to su m m er co m m u n itie s in a long tim e scale. T he w a ter chem istry p a ram e te rs w ere de te rm ined according to K oro leff (1979), an d e ac h v a lu e represen ts the m ean fo r the 0 —10 m w ater layer. T h e lim it o f de tec tio n in the nutrient m easurem ent w as 0.01 [iM . S a lin ity w as m easu red using the Practical Salinity Scale.
2.3. S ta tistica l a n a lyses
B oth th e p h y to p la n k to n an d the en v iro n m en ta l da ta w ere analysed fo r th e p re sen c e o f m o n o to n ie increasing or d ec reasing trends by th e n o n -p a ra m e tr ic M an n —K endall te s t using the M A K E SE N S 1.0 a p p lic a tio n (S a lm i et a l., 2002). A n o n -p ara m etric tren d te s t w as u sed , b e c a u se all d a ta w ere no t norm ally d istribu ted . In the M a n n —K en d all p ro ced u re , the d a ta need not confo rm to any p a r tic u la r d is trib u tio n an d m issin g values are allow ed (G ilb ert, 1987).
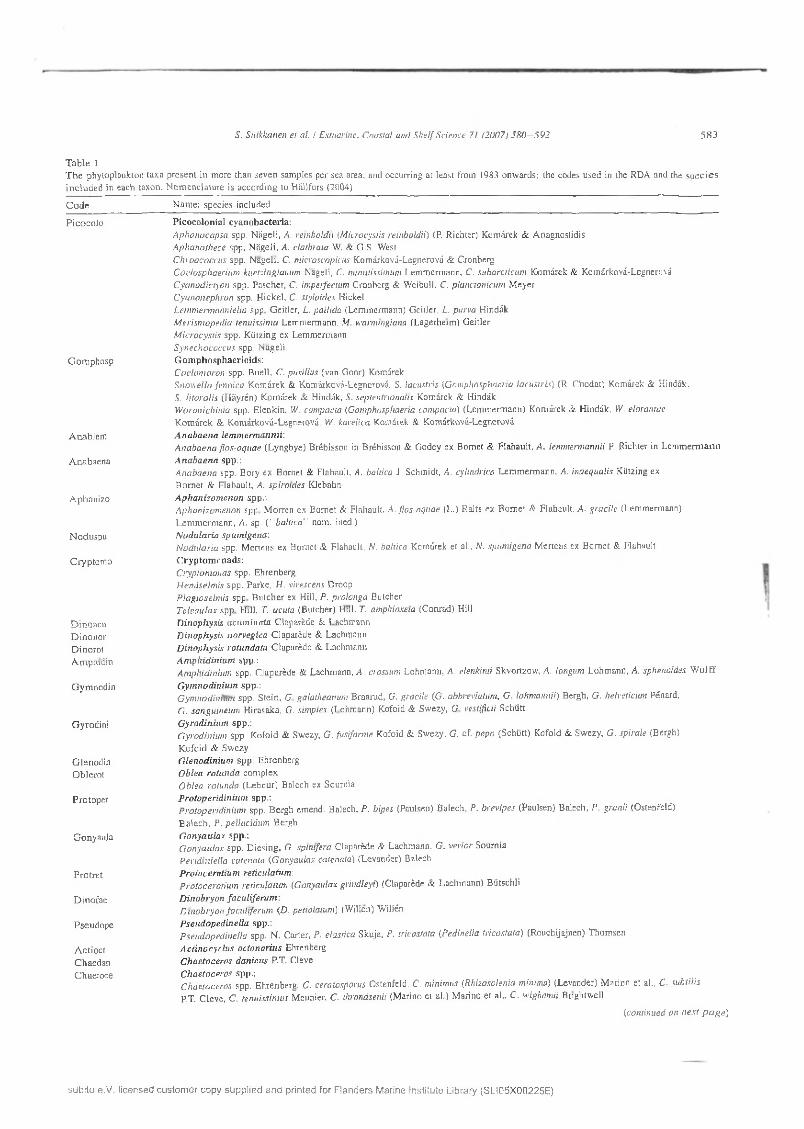
R e la tio n sh ip s b e tw ee n th e p h y to p lan k to n com position and the en v iro n m en ta l fa c to rs w e re a sse ssed using redundancy analysis (R D A ), w h ich is a l in e a r m eth o d o f d irec t o rd ination (te r B raak , 1994). F o r the a n a ly s is , w e se lec ted phy top lank ton species o r w id e r tax o n o m ic g ro u p s w h ich w ere iden tified co n sisten tly o ver m o st o f the s tu d y p e rio d (a t least fro m 1983 on w ards), an d w h ich o c cu rre d in m ore th an seven sam ples in each sea a rea (T able 1). T h e final n u m b er o f taxa in the analysis w as 32 in th e N B P an d 28 in th e GF. A ll b iom ass da ta w ere log(.v-F l) - tra n s fo rm e d to s ta b iliz e variance and reduce the in fluence o f d o m in a n t ta x a on th e o rd ination . R D A was p e rfo rm ed u sin g C A N O C O fo r W in d o w s 4 .0 ( te r B raak and
S m ilauer, 1998), w ith fo rw ard se lec tio n to identify the env ironm ental v a ria b le s th a t b est e x p la in ed the phytoplankton species co m position . T h e s ign ificance o f th e variables and the first o rd in atio n axis w a s d e te rm in ed using M o n te C arlo perm uta tion testing (9999 p e rm u ta tio n s), im p le m e n ted in C AN OCO.
3. Results
3.1. L ong-term trends in env iro n m en ta l fa c to r s and phytop lankton
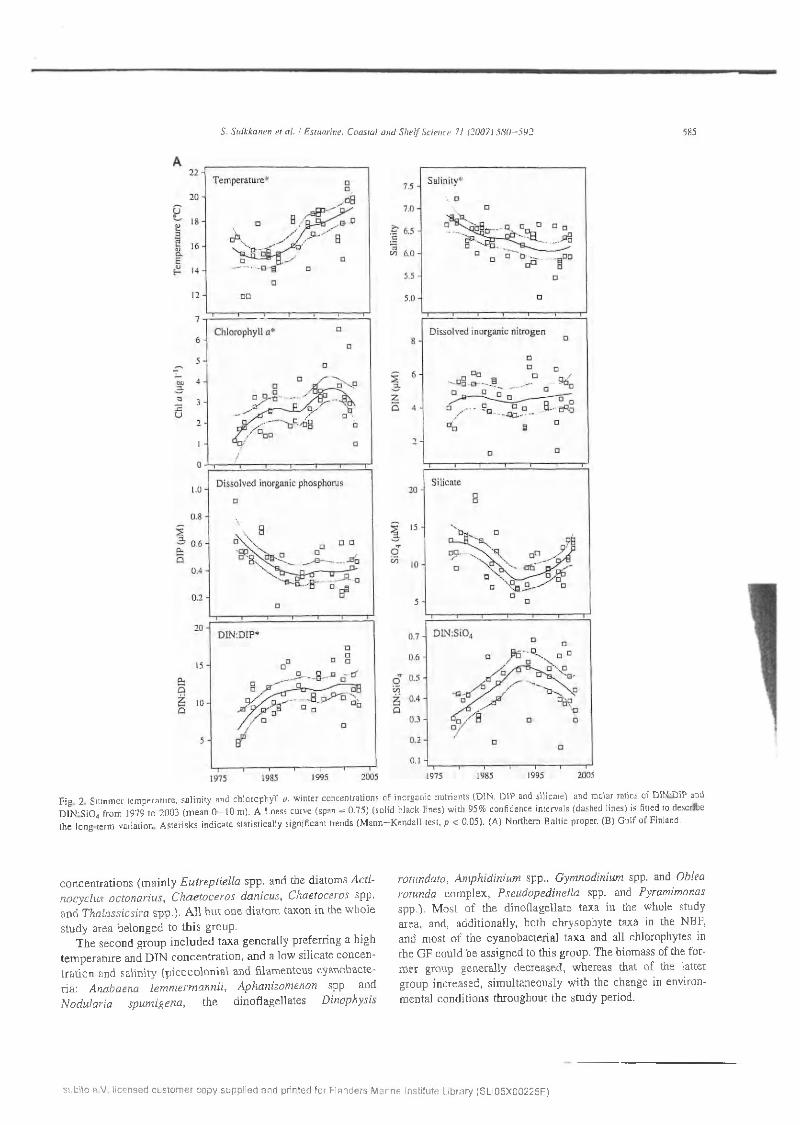
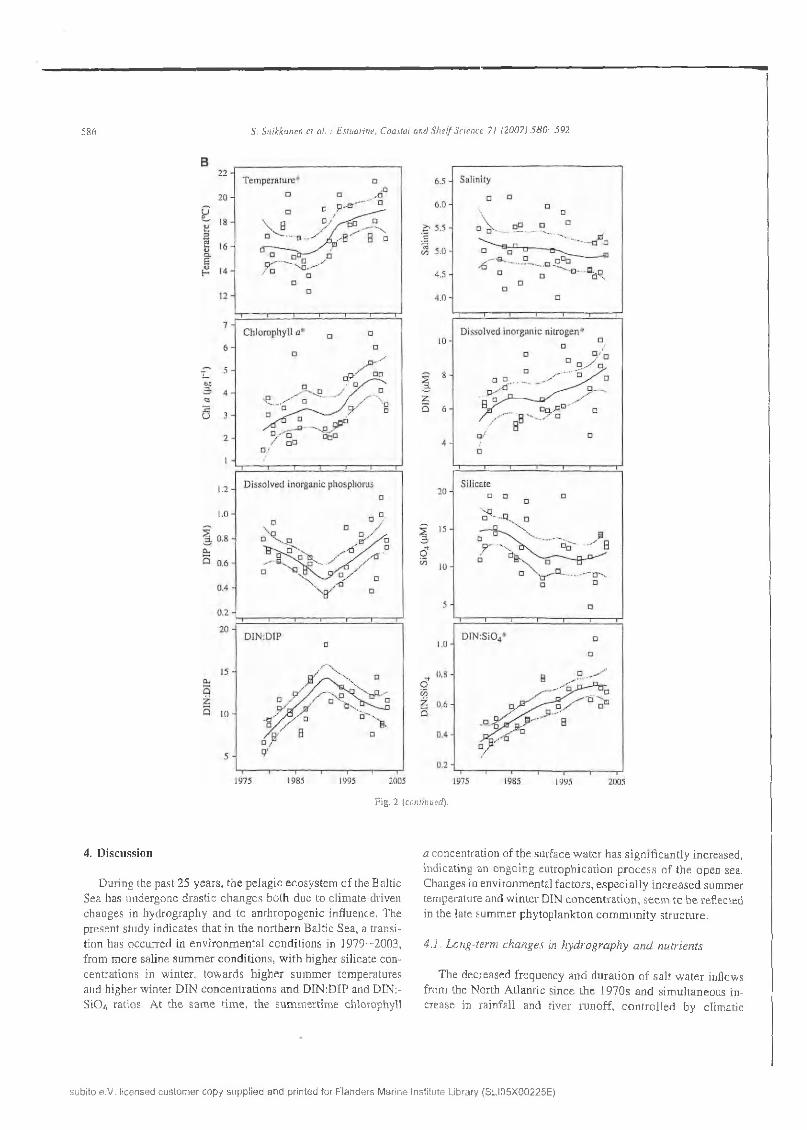
In the northern B a ltic proper, s ig n ifican t (M ann—Kendall test, p < 0 .05) in creas in g tren d s w ere o b se rv ed for sum m er tem pera tu re and ch lo ro p h y ll a co n ce n tra tio n , and w inter d isso lv ed in o rgan ic n itro g e n to p h o sp h o ru s (D IN :D IP) ratio , w hereas sum m er sa lin ity show ed a d e c re a s in g trend (Table 2, Fig. 2A ). In the G u lf o f F in lan d , in c re as in g trends w ere found fo r sum m er tem p e ra tu re and ch lo ro p h y ll a, and w inter D IN co n cen tra tion an d D IN :S i0 4 ra tio (F ig . 2B). In the an a lysis o f early su m m er n u trie n t c o n ce n tra tio n s (data not show n), no sign ifican t trends w ere o b se rv ed d u e to a large variance (M ann—K endall test,/? > 0 .05 ), H ow ever, th e re was a decreasing tendency in D IN co n ce n tra tio n s in b o th sub-areas, but e specially in th e GF.
In the p h y to p lan k to n b iom ass d a ta (F ig . 3), significant increasing trends w ere fo u n d fo r d in o p h y te s , chrysophytes and ch lo ro p h y tes in the n o rth ern B altic p roper (Table 2, F ig . 3A ), and fo r cy an o b acte ria , ch ry so p h y tes and ch lo ro phy tes in th e G u lf o f F in lan d (F ig . 3 B ). N egative trends w ere obse rv ed fo r e u g len o p h y tes in the n o rth e rn B altic p roper and fo r c ry p to p h y tes in th e G u lf o f F in lan d .
3 .2 . R e la tio n sh ip s be tw een p h y to p la n k to n a n d environm en ta l fa c to rs
T h e p h y to p lan k to n tax a se lec ted fo r th e R D A and the sp e cies in c lu d ed in each co m b in a tio n tax o n a re listed in Table 1. T h e 32 co m b in a tio n tax a u sed in the R D A consisted o f 11 d inophytes, 6 cy an o b acte ria , 6 d ia to m s, 4 chlorophytes, 2 ch ry sophy tes and 1 tax o n o f c ry p to p h y tes , euglenophytes and zooflagella tes, i.e. E bria tr ipartita (T ab le 3). M ost o f the included sp ec ies are ty p ica l o f w arm b rack ish water, but so m e o f the taxa a lso con ta in lacu strin e o r co ld -w ater species. S om e o f the sp ec ies u se m ixo- o r h e te ro tro p h ic nu trition , although they a re trad itio n a lly re g a rd e d as phytoplankton sp ec ies (T ab le 3). T h e m o st co m m o n ta x a in th e m aterial, o c cu rrin g in all sam ples, w ere A p h a n izo m e n o n spp. and P yram im o n a s spp. C ryp to m o n ad s w ere a lso fo u n d in all sam ples in the GF. A p h a n izo m en o n spp. and c ry p to m o n ad s had the h ig h est m ean b io m ass in both sea areas (8 7 .9 an d 68.1 pg T 1 in the NBP, and 103.7 an d 99.3 p g l -1 in the G F, respectively). T he th ird h ig h est b iom asses w ere fo rm ed by p ico co lo n ia l cy an o bac teria in th e N B P and th e to x ic c y an o b ac te riu m N odu laria spum igena in th e G F (T able 3). G o n y a u la x spp ., D inobryon fa cu life ru m , C ylindro theca c lo s teriu m a n d N itzsch ia spp. w ere enco u n tered freq u en tly en o u g h in th e N B P only, and thus they w ere excluded fro m the a n a ly sis o f th e G F data.
s u b ito e .V . l ic e n s e d c u s to m e r c o p y s u p p l ie d a n d p r in te d fo r F la n d e r s M a r in e In s titu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )
S. Su ikkanen el aí. / Estuarine . C oasta l a n d S lie lf Science 71 (2007) 5 8 0 —5 9 2 5 8 3
Table 1T h e phy top lank ton tax a p re sen t in m ore than seven sam ples p e r sea area, and occurring at leas t from 1983 onw ard s; the codes u sed in the RDA and the s p e c ie s in c lu d e d in each taxon. N o m en c la tu re is acco rd in g to H ällfors (2004)
Code N am e: sp ec ies included
P ic o c o lo
G o m p h o sp
Anablem
A n ab aen a
A p h a n iz o
N o d u s p u
C ry p to m o
D in o a c u
D in o n o rD in o ro tA m p h id in
G y m n o d in
G yro d in i
G len o d inO b le ro t
P ro to p e r
G o n y au la
P ro tre t
D in o fac
P se u d o p e
A c tio c tC h ae d anC h ae to ce
P ic o c o lo n ia l c y a n o b a c te r ia :A p h a n o c a p sa spp. N iigeli, A. reinhold ii (M icrocystis reinhold ii) (P. R ichter) K om árek & A nagnostid is A p h a n o th ece spp, N iigeli, A. cla thra ta W, & G .S . WestC h ro o co cc u s spp. N ag eb , C. m icroscopicus K om árková-L egnerová & CronbergC oelo sp h a eriu m kue tzing ianum N ageb , C. m inutissim um L em m erm ann , C. subarc ticum K om árek & K om árková-L egnerová C yanod ic tyon spp. P ascher, C . im perfectum C ronberg & W eibull. C. p lanc ton icum M eyer C ya n o n ep h ro n spp. H icke l, C. stylo ides H ickelL em m erm a n n ie lla spp. G eitler, L. pallida (Lem m erm ann) G eitler, L. pa rva H indák M erism o p ed ia tenu issim a L em m erm ann, M. warm ingiana (L agerheim ) G eitler M ic ro c ystis spp. K ü tz in g ex Lem m erm ann S yn ech o co cc u s spp. N ageb G o m p h o s p h a e r io id s :C oelo m o ro n spp. B uell, C. pusillus (van G oor) KomárekS n o w e lla fe n n ic a K om árek & K om árková-L egnerová, S. lacustris (G om phosphaeria la cu stris ) (R. C hodat) K om árek & H indák.S. lito ra lis (H áy rén ) K om árek & H indák, S. sep tentrionalis K om árek & H indákW o ron ich in ia spp. E lenk in , W. com pacta (G om phosphaeria com pacta) (L em m erm ann) K om árek & H indák , W. elorantae K o m árek & K om árková-L egnerová, W . karelica K om árek & K om árková-L egnerová A n a b a e n a lem m erm a n n ii'.A n a b a e n a flo s-a q u a e (L yngbye) B rébisson in B rébisson & G odey ex B om et & F lah au lt, A . lem m erm annii P. R ichter in L e m m e rm a n n A n a b a e n a spp .:A n a b a e n a spp. B ory ex B o m et & F lahau lt, A. baltica J. S chm idt, A. cylindrica L em m erm ann , A. inaequa lis K ützing ex B o m e t & F lahau lt, A. sp iro ides K lebahn A p h a n iz o m e n o n spp .:A p h a n izo m en o n spp. M orren ex B om et & F lahault. A. flo s-aquae (L .) R alis ex B o m e t & F lahau lt, A . g racile (Lem m erm ann) L e m m e rm an n , A . sp . ( " b a ltic a " nom . ined.)N o d u la r ia spu m ig en a :N o d u la r ia spp. M ertens ex B om et & F lahault, N . baltica K om árek e t al., N . spu m ig en a M ertens ex B o m e t & Flahault C ry p to m o n a d s :C ryp to m o n a s spp . E hrenbergH e m ise lm is spp . P arke, H . virescens D roopP la g io se lm is spp. B u tc h e r ex H ill, P ■ pro longa B utcherT e le a u la x spp. H ill, T. acuta (B utcher) H ill. T. am phioxeia (C onrad) HillD in o p h y s is a c u m in a ta C laparède & Lachm annD in o p h y s is norveg ica C laparède & Lachm annD in o p h y s is ro tu n d a ta C laparède & L achm annA m p h id in iu m spp .:A m p h id in iu m spp. C laparède & L achm ann, A . crassum Lohm ann, A. e lenkin ii Skvortzov/, A . longum L ohm ann , A. sphenoides W u l f f
G y m n o d in iu m spp .:G ym n o d in iu m spp. S te in , G. gala theanum B raan id , G. gracile (G. abbrevia tum , G . lohm annii) Bergh, G . helveticum Pénard,G . sa n g u in eu m H irasak a , G. sim plex (Lohm ann) K ofoid & S w ezy , G . vestificii S chü tt
G y ro d in iu m spp .:G yro d in iu m spp. K ofo id & Swezy, G . fu s ifo rm e Kofoid & S w ezy, G . cf. p ep o (S chü tt) K ofoid & S w ezy, G. sp ira le (Bergh)
K o fo id & S w ezyG le n o d in iu m s p p . E hrenbergO blea r o tu n d a co m p lexO blea ro tunda (L ebou r) B alech ex SourniaP r o to p e r id in iu m sp p .:P ro to p e rid in iu m spp. B ergh em end. B alech , P . bipes (Paulsen) B alech , P. brevipes (Paulsen) B alech , P. g ran ii (O stenfeld)
B a lech , P . p e lluc idum B ergh G o n y a u la x sp p .:G o n y a u la x spp. D ies in g , G . sp in ifera C laparède & Lachm ann, G . ve rio r Sournia P e rid in ie lla carenata (G onya u la x catenata) (L evanderi B alech P r o to c e ra tiu m re tic u la tu m :P ro to cera tiu m re ticu la tum (G onyau lax grindleyi) (C laparède & L achm ann) B ütschli D in o b r y o n fa c u life r u m :D in o b ryo n fa c u life ru m (D . petio la tum ) (W illen) W illén P s e u d o p e d in e lla spp .:P seu d o p e d in e lla spp. N \ Carter, P. elastica S ku ja, P. tricostata (P edinella tr icosta ta) (Rouchijajnen) T hom sen
A c tin o c y c lu s o c to n a riu s Ehrenberg C h a e to cero s d a n ic u s P.T. C leve C h a e to cero s sp p .:C haetoceros spp. E hrenberg , C . ceratosporus O stenfeld, C. m in im us (R h izo so len ia m inim a) (L evanderi M arino e t al„ C. sub tilis P.T. C leve , C . tenu issim us M eunier, C . throndsenii (M arino el al.) M arino e t a l„ C . w igham ii B rightw ell
(con tinued on n e x t p a g e )
s u b i to e .V . l i c e n s e d c u s to m e r c o p y s u p p l ie d a n a p r in te d fo r F la n d e r s M a rin e In s titu te L ib ra ry (S L 1 0 5 X 0 0 2 2 5 E )
584 S. S u ikkanen er a l. / E stuarine , C oasta l a n d S h e lf Sc ience 71 (2007) 5 8 0 —5 9 2
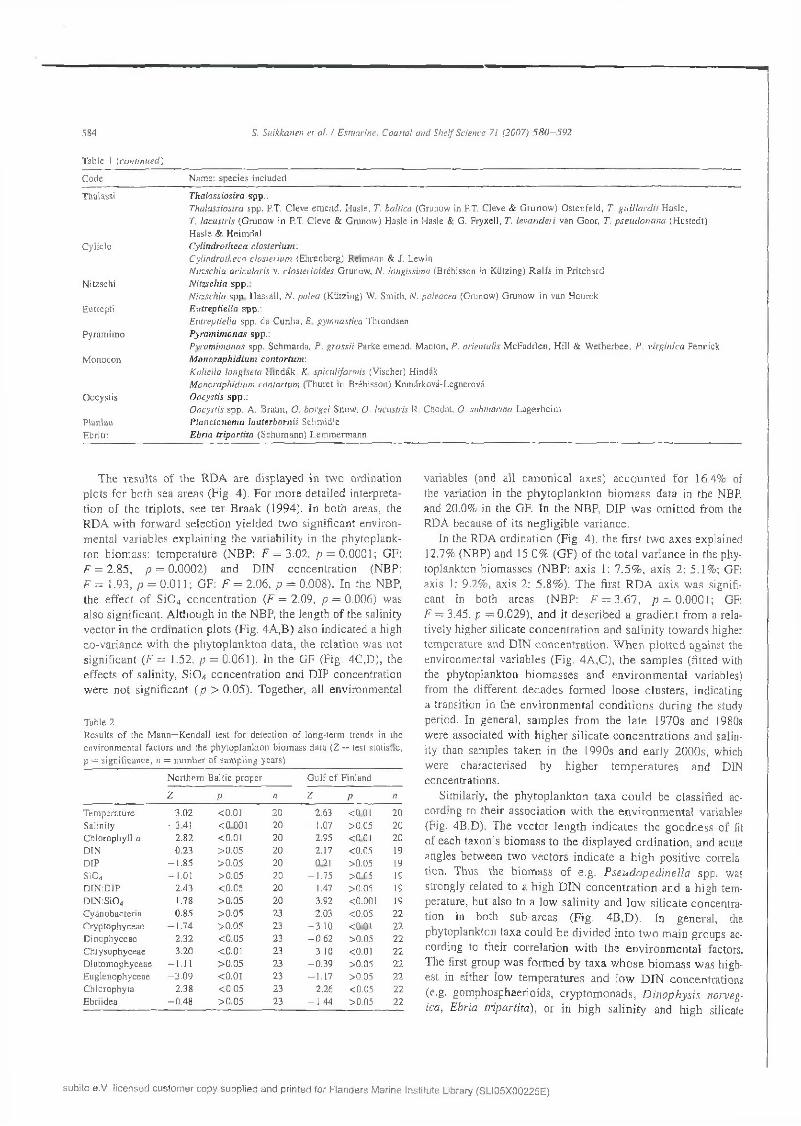
T able 1 (c o n tin u ed )
C ode N am e: sp ec ies inc luded
T halassi
C yliclo
N itzsch i
E utrepti
P y ram iin o
M onocon
O ocystis
P lan lauE britri
T h a la ss io s ira spp .:T h a lassio sira spp . P.T. C leve em en d . H asle , T. baltica (G runow in P.T. C leve & G ru n o w ) O sten fe ld , T. g u i l la r d i i H asie ,T. lacustris (G runow in P.T. C leve & G n inow ) H asle in H asle & G . F ryxell, T . le v a n d e r i van G oor, T. p se u d o n a n a (H usted t) H as le & H eim dal C ylin d ro th ec a closterium -.C ylindro theca c lo ster iu m (E hrenberg) R eim ann & J. Lew inN itzsch ia acicu la ris v. clo ster io ides G runow , N. long issim a (B rébisson in K iitzing) R a lfs in P ritch ard N itzsc h ia sp p .:N itzsch ia spp . H assa ll, N . p a le a (K iitzing) W. S m ith , /V. pa leacea (G ninow ) G runow in van H eu rck E u tre p tie lla sp p .:E utrep tie lla spp. d a C u n h a , E. gym nastica T hrondsen P y ra m im o n a s spp .:P yram im onas spp . S chm arda , P . g ro ss ii P arke em end . M antón, P . orien ta lis M cF ad d en , H ill & W e th erb ee , P . virg in ica Penniek M o n o r a p h id iu m c o n to r tu m :K oliella long ise ta H in d ák , K. sp icu lifo rm is (V ischer) H indák M o n oraph id ium con to rtum (T h u re t in B réb isson) K om árková-L egnerová O ocystis s p p .:O ocystis spp . A . B raun , O. borgei S now , 0 . lacustris R . C hodat, 0 . subm arina L a g e rh e im P la n c to n e m a la u te r b o rn ii Schm id le E b ria tr ip a r tita (S ch u m an n ) L em m erm ann
T h e re su lts o f th e R D A are d isp lay ed in tw o o rd ination p lo ts fo r bo th sea a rea s (Fig. 4 ). F o r m o re d e ta iled in te rp re ta tion o f th e trip lo ts , see te r B ra ak (1994). In bo th areas, the R D A w ith fo rw ard se lec tio n y ie ld ed tw o sig n ifican t env iron m enta l va riab les ex p la in in g th e v a riab ility in the p h y to p lan k ton b io m ass: tem p e ra tu re (N B P: F = 3 .02 , p = 0 .0001; GF: F = 2 .85 , p = 0 .0 0 0 2 ) and D IN c o n cen tra tio n (NBP; F = 1.93, p = 0.011 ; GF: F = 2 .06 , p = 0 .008). In th e NBP, th e e ffec t o f S i 0 4 co n ce n tra tio n (F = 2 .09 , p = 0 .006) was a lso s ig n ifican t. A lth o u g h in th e NBP, th e length o f th e sa lin ity v ec to r in the o rd in a tio n p lo ts (F ig . 4 A ,B ) also in d ica ted a high c o -v arian ce w ith th e p h y to p lan k to n d a ta , the re la tio n w as not s ig n ifican t (F = 1.52, p = 0 .0 6 1 ). In th e G F (Fig. 4C ,D ), the e ffec ts o f sa lin ity , S i 0 4 c o n cen tra tio n an d D IP co n cen tra tion w ere n o t sig n ifican t ( p > 0 ,0 5 ). T o gether, all en v ironm en ta l
T able 2R esu lts o f the M an n —K endall te s t fo r d e tec tio n o f long -term trends in the env ironm en ta l fac to rs and the p h y to p lan k to n b io m ass d a ta (Z = test s tatistic , p = s ign ificance , n = n u m b e r o f sam p lin g years)
N o rth e rn B altic p ro p e r G u lf o f F in land
Z P n Z P n
T em p era tu re 3 .02 < 0.01 20 2.63 < 0.01 20S alin ity - 3 .4 1 < 0.001 20 - 1 .0 7 > 0 .0 5 20C h lo ro p h y ll a 2 .8 2 < 0.01 20 2.95 < 0.01 20DIN 0.23 > 0 .0 5 20 2.17 < 0 .05 19D IP - 1 .8 5 > 0 .0 5 20 0.21 > 0 .05 19SÍO 4 - 1 .0 1 > 0 .0 5 20 - 1 .7 5 > 0 .0 5 19DIN:DTP 2.43 < 0 .0 5 20 1.47 > 0 .0 5 19D L N :S i04 1.78 > 0 .0 5 20 3.92 < 0.001 19C yanobac te ria 0 .85 > 0 .0 5 23 2.03 < 0 .0 5 22C ryp to p h y ceae - 1 .7 4 > 0 .0 5 23 - 3 .1 0 < 0.01 22D in o p h y ceae 2 .3 2 < 0 .0 5 23 - 0 .6 2 > 0 .0 5 22C hrysophyceae 3.20 < 0.01 23 3,10 < 0.01 22D ia to m o p h y ceae - 1.11 > 0 .0 5 23 - 0 .3 9 > 0 .0 5 22E ug len o p h y ceae - 3 .0 9 < 0.01 23 - 1 .1 7 > 0 .0 5 22C h lo ro p h y ta 2 .38 < 0 .0 5 23 2.26 < 0 .0 5 22E b riid ea - 0 .4 8 > 0 .0 5 23 - 1 .4 4 > 0 .0 5 2 2
variables (and a ll c an o n ica l ax es) a cc o u n te d fo r 16.4% of the variation in th e p h y to p la n k to n b io m a ss d a ta in th e NBP, and 20,0% in th e GF. In the NBP, D IP w a s o m itted from the RD A because o f its n e g lig ib le va rian ce .
In the R D A o rd in a tio n (Fig. 4), th e f irs t tw o ax es explained 12.7% (N B P) and 15.0% (G F ) o f th e to ta l v a ria n ce in the phytoplankton b iom asses (N B P: ax is 1: 7 .5 % , ax is 2: 5 .1% ; GF: axis 1: 9 .2% , ax is 2: 5 .8 % ). T h e first R D A ax is w as significan t in both a reas (N B P : F = 3 .6 7 , p = 0 .0 0 0 1 ; GF: F = 3.45, p = 0 .029), an d it d e sc rib e d a g ra d ie n t from a relatively h ig h er s ilica te co n ce n tra tio n and sa lin ity to w ard s higher tem perature and D IN c o n ce n tra tio n . W h en p lo tte d against the env ironm ental v a riab le s (F ig . 4 A ,C ), th e sa m p le s (fitted with the p h y to p lank ton b io m a sses and e n v iro n m en ta l variables) from the d iffe ren t d e ca d es fo rm e d lo o se c lu s te rs , indicating a transition in the e n v iro n m en ta l c o n d itio n s d u rin g th e study period. In g enera l, sa m p le s fro m th e la te 1970s an d 1980s w ere assoc ia ted w ith h ig h e r s i l ic a te c o n c e n tra tio n s an d salinity than sam ples taken in the 1990s a n d e a r ly 2 0 0 0 s , which w ere ch arac te rised by h ig h er te m p e ra tu re s an d DIN concen tra tions.
S im ilarly , the p h y to p la n k to n ta x a c o u ld be c lass ified accord ing to th e ir a sso c ia tio n w ith th e e n v iro n m e n ta l variables (Fig. 4B ,D ). T h e v ec to r len g th in d ic a te s th e g o o d n e ss o f fit o f each tax o n ’s b io m a ss to th e d isp la y e d o rd in a tio n , and acute angles b e tw een tw o v e c to rs in d ic a te a h ig h p o sitiv e correlation. T hus, the b io m ass o f e .g . P se u d o p e d in e lla spp . was strongly re la ted to a h ig h D IN c o n c e n tra tio n an d a h igh tem perature, but a lso to a low sa lin ity and lo w s il ic a te co n cen tra tion in bo th su b -a reas (F ig . 4 B ,D ). In g e n era l, the phy top lank ton ta x a co u ld b e d iv id e d in to tw o m ain g ro u p s accord ing to th e ir co rre la tio n w ith the e n v iro n m e n ta l factors. T he first group w as fo rm e d b y ta x a w h o se b io m a ss w as highest in e ith e r low tem p era tu res an d lo w D IN co n cen tra tions (e.g. g om phosphaerio ids, c ry p to m o n ad s , D in o p h y s is norvegica, E bria tripartita ), or in h ig h sa lin ity and h ig h silicate
s u b ito e .V . l i c e n s e d c u s to m e r c o p y s u p p lie d a n d p r in te d fo r F la n d e r s M a rin e In s titu te L ib rary (S L I0 5 X 0 0 2 2 5 E )
S . Su ikkanen e t a l. I Estuarine. C oasta l a n d S lw l f Science 71 (2007) 5 8 0 - 5 0 2 585
22 -T em pera tu re
2 0 -
UCU3S<uD.E
t -
□□
C h lo rophy ll a'
.....■CU
D isso lved inorgan ic phosphorus
o.Q
0 .4 -
0 .2 -
20 -D IN :D IP *
5ZQ
2005199519851975
7.0 -
6.5 - o..
co 6,0 -
5 .0 -
D isso lved inorganic n itro g en
S3.
z5
Silicate20 -
O* 1 0 -
D IN :SiO ,0.7
0.6
-Q-a'zo
0.3 ' □ /
0.2
1995 200519851975
F ig 2 S um m er tem pera tu re , s a lin ity and ch lo rophy ll a , w in te r concentrations o f inorganic nu trien ts (D IN , D IP and s ilica te), and m olar ratios o f D IN:DIP and D IN -SiO * from 1979 to 20 0 3 (m e a n 0 - 1 0 m ). A Loess curve (span = 0 .75) (so lid b lack lines) w ith 95% confidence in tervals (dashed lines) is fitted to describe th e long-term variation A ste risk s ind ica te s ta tis tic a lly significant trends (M a n n -K e n d a ll test, p < 0 .05). (A) N orthern B altic proper, (B) G u lf o f Finland.
co n cen tra tio n s (m ain ly E u trep tie lla spp . and the d iatom s A ctinocyc lu s o c to n a r iu s , C h a e to cero s d a n icu s , C haetoceros spp. and T halassiosira sp p .). A l l b u t one d ia to m taxon in the w ho le s tudy area b e lo n g ed to th is group.
T h e second g ro u p in c lu d e d tax a genera lly p referring a h igh tem p e ra tu re and D IN co n cen tra tio n , an d a low silica te co n cen tra tio n and sa lin ity (p ico c o lo n ia l and filam entous cyanobacteria : A nabaena le m m erm a n n ii, A phan izom enon spp. and N odularia, sp um igena , th e d inoflagellates D inophysis
rotundata , A m p h id in iu m spp., G ym nodinium spp. and Oblea ro tunda co m plex , P seudopedinella spp. and Pyram im onas spp .). M o st o f th e d ino flagella te taxa in the w hole study area, and, add itionally , both chrysophyte taxa in the NBP, and m ost o f the cyanobacteria l tax a and a ll chlorophytes in the G F could be assigned to th is group. T he biom ass o f the form er group gen era lly decreased , w hereas that o f the latter g roup increased , sim ultaneously w ith the change in environm enta l co nd itions th roughout th e study period.
s u b ito e .V . l i c e n s e d c u s to m e r c o p y s u p p l ie d a n d p r in te d fo r F la n d e r s M arin e In s titu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )
586 S . S u ikkanen c t a l. I E stuarine , C oastal a n d S h e l f Science 71 (2007) 5 8 0 - 5 9 2
T e m p era tu re
?ya2
C h lo ro p h y ll a*
.P -□□
aÖ
D isso lv ed inorgan ic phosphorus
Q
0.2
20 -D IN :D IP
19851975 2005
S alin ity6.5
6 .0 -
□Dc
3 5.0
4.5
4.0
D isso lved in o rg a n ic n itro g en1 0 -
2Q
□ /
S ilica te2 0 -
- Q . ,
10 -
D IN :SiO ,
O<75SQ n . /
0 .4 -
0 .2 -
1975 1985 2005
Fig . 2 (continued).
4. Discussion
D u rin g the p a s t 25 y e a rs , th e p e la g ic eco sy stem o f the B altic Sea has und erg o n e d ra s tic c h a n g e s b o th due to c lim ate-d riven ch an g es in h y d ro g rap h y an d to a n th ro p o g e n ic in fluence. The p re sen t study in d ica tes th a t in th e n o rth e rn B altic S ea , a transitio n h as o ccu rred in e n v iro n m en ta l co n d itio n s in 1979—2003, fro m m ore sa line su m m e r c o n d itio n s , w ith h igher silica te conc en tra tio n s in w in ter, to w ard s h ig h e r su m m er tem pera tu res and h igher w in ter D IN c o n ce n tra tio n s an d D IN iD IP and D IN :- S i 0 4 ra tio s. A t th e sa m e tim e , th e su m m ertim e ch lorophyll
a co n cen tra tion o f th e su rface w a te r has s ig n if ic a n tly increased , ind ica ting an o n g o in g eu tro p h ica tio n p ro c e ss o f th e open sea. C hanges in en v iro n m en ta l fa c to rs , e sp e c ia lly in c re a se d sum m er tem pera tu re an d w in te r DIN c o n c e n tra tio n , se e m to b e reflected in th e late su m m er p h y to p la n k to n c o m m u n ity s tru c tu re .
4.1. L ong-term ch a n g es in h y d ro g ra p h y a n d n u tr ien ts
T h e d ecreased frequency a n d d u ra tio n o f sa lt w a te r inflows from the N orth A tlan tic s in ce th e 1970s an d sim u ltan eo u s in crease in ra in fa ll an d riv er ru n o ff , c o n tro lle d b y c lim atic
s u b ito e .V . l ic e n s e d c u s to m e r c o p y s u p p lie d a n d p r in te d fo r F la n d e r s M a rin e In s titu te L ib ra ry (S L 1 0 5 X 0 0 2 2 5 E )
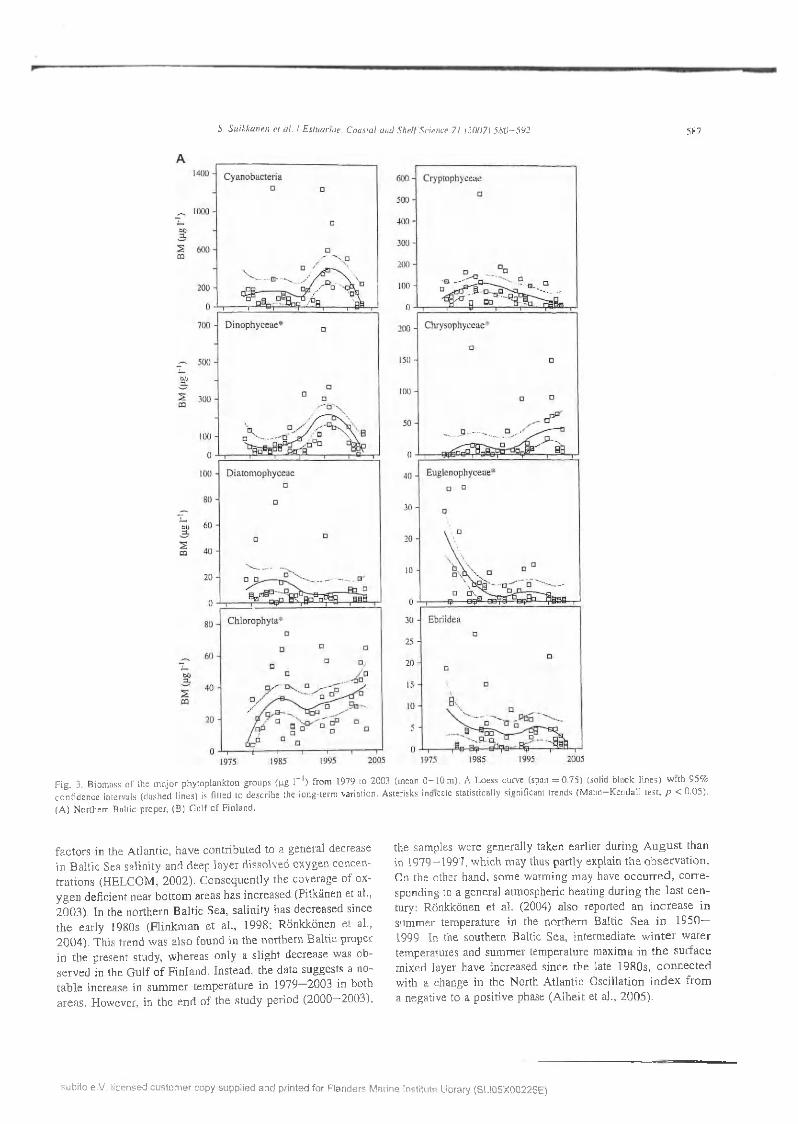
S. S u ikkanen e t a l. / E stuarine . C oastal and S h e l f Science 71 (20071 5 S U -5 9 2 587
1400 - C yanobacteria Cryptophyceae600 -
500 -1000 -
400 -
300-
2 00 -
1 0 0 -2 0 0 -
B O
D inophyceae ' 2(jo . C hrysophyceae700 -
150 -500 -
100 -
CQ
5 0 -100
Euglenophyceae'100 - D ia tom ophyceae 40 -
SO -30 -
60 -so=L2 0 -
I 40 -
2 0 -
Ebriideago . C h lo rophy ta 30 -
60 -20 -
oof «■CQ
tx.
10 -
200519852005 1975199519851975
F ig . 3. B iom ass o f Ute m a jo r p h y top lank ton g roups (p g I " 1) from 1979 to 2003 (m ean 0 - 1 0 m ). A L oess curve (span = 0 .7 5 ) (solid b la c k lin e s ) w ith 9 5 % c o n fid en ce intervals (d ash ed lin es ) is fitted to desc rib e the long -term variation . A sterisks indicate sta tis tica lly significant trends (M a n n -K e n d a ll te st, p < 0 .0 5 ).
(A ) N orthern B altic p roper, (B ) G u lf o f F in land .
fa c to rs in th e A tlan tic , h ave co n trib u ted to a general decrease in B altic S ea sa lin ity and deep lay e r d isso lv ed oxygen concentra tio n s (H E L C O M , 2 0 0 2 ). C o n seq u en tly the coverage o f ox y g e n defic ien t near b o tto m areas h as in creased (P itkänen et al.,2 0 0 3 ). In the n o rth ern B altic Sea, sa lin ity has d ecreased since th e early 1980s (F lin k m an e t al., 1998; R önkkönen e t al.,2 0 0 4 ). T his trend w as a lso found in th e n o rth ern B a ltic p roper in th e p resen t study, w h ereas on ly a s lig h t decrease w as ob se rv ed in th e G u lf o f F in land . In stead , th e d a ta suggests a no ta b le increase in su m m er tem p era tu re in 1979—2003 in both a rea s . H ow ever, in th e e n d o f the s tu d y period (2 0 0 0 -2 0 0 3 ) ,
the sam ples w ere generally taken earlie r d u rin g A u g u s t th a n in 1979—1997, w h ich m ay thus p a rtly explain th e o b se rv a tio n . On th e o ther h and , so m e w arm ing m ay have o c c u r re d , c o rre sponding to a general atm ospheric heating d u rin g th e la s t c e n tury: R önkkönen e t a l. (2004) a lso reported an in c re a s e in sum m er tem perature in the n o rth ern B altic S e a i n 1950— 1999. In the sou thern B altic Sea, in te rm ed iate w in te r w a te r tem peratures and su m m er tem pera tu re m ax im a in th e su rfa c e m ixed layer h ave in creased s in ce th e late 1980s, c o n n e c te d w ith a change in the N orth A tlan tic O scilla tio n in d e x f ro m a negative to a p o sitiv e phase (A lh e it et al., 2 0 0 5 ).
s u b i to e .V . l i c e n s e d c u s to m e r c o p y s u p p lie d a n d p r in te d fo r F la n d e r s M a r in e In s titu te L ib rary (S L I0 5 X 0 0 2 2 5 E )
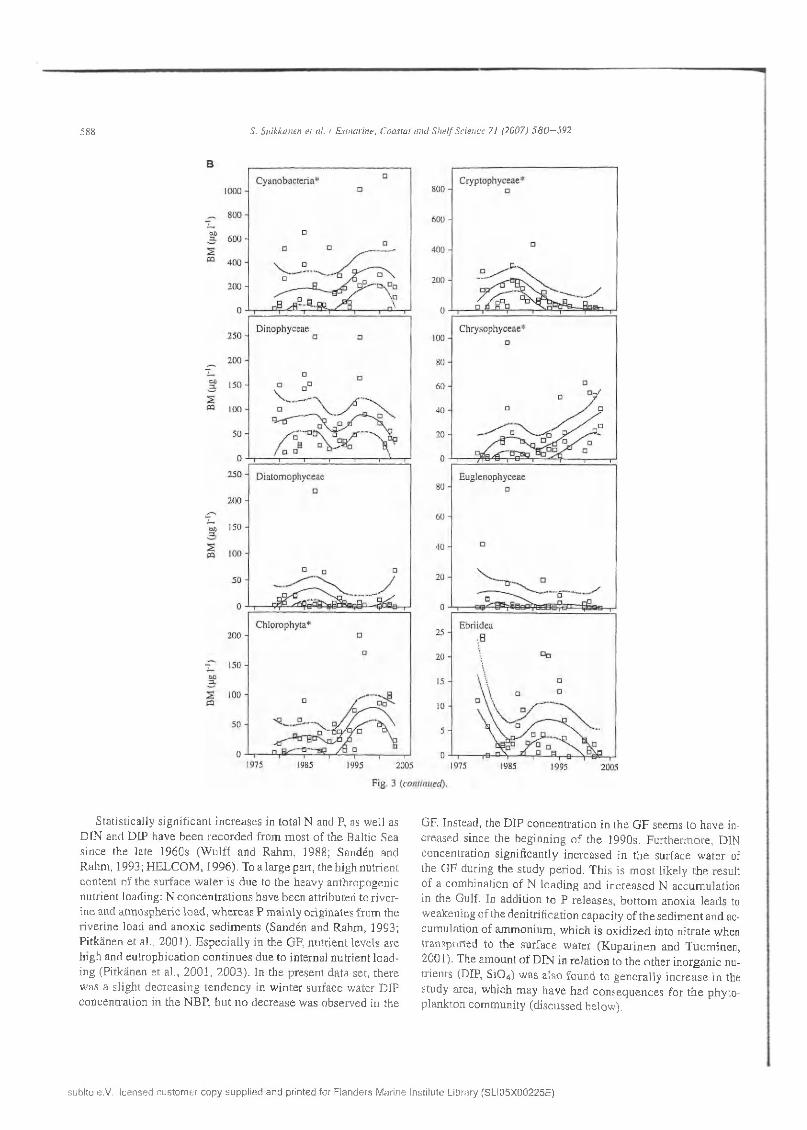
588 S. Su ikkanen e t al. / E stuarine , C oasta l a n d S h e l f Sc ience 71 (2007) 5 8 0 —592
B
C ryp to p h y ceae1C y an o b a c te ria ’800 -1000 -
800 - 600 -
3 600 - Sm 400 -
400 -
2 0 0 -200 -
D inophyceae C h ry so p h y ceae’250 - 1 0 0 -
2 0 0 - 80 -
B 'SOISm 1 0 0 -
60 -
40 -
50 - 20 -
250 - D ia tom ophyceae E uglenophyceae80 -
200 -
60 -
40 -
2 0 -50 -
C h lo ro p h y ta ’ E briidea25 -200 -
2 0 -■ ff 150 -M32 mo -cû 7-0 ao '■ 10 -
50 -
975 1985 2005
F ig . 3 (con tinued ).
1975 1985 1995
S ta tistica lly s ig n ifican t in c re a se s in to ta l N and P, as w ell as D IN and D IP have b e e n re c o rd e d fro m m o st o f th e B altic Sea since th e la te 1960s (W u lff an d R ah m , 1988; Sandén and R ahm , 1993; H E L C O M , 1996). T o a larg e part, the h ig h nu trien t co n ten t o f th e su rfa ce w a te r is d u e to th e h eavy an th ro p o g en ic nu trien t load ing : N co n c e n tra tio n s h ave b een a ttrib u ted to riv e rin e and a tm o sp h e ric lo ad , w h e re a s P m ain ly o rig in a tes fro m the riv e rin e load an d a n o x ic se d im e n ts (S andén and R ah m , 1993; P itk än en e t al., 2 0 0 1 ). E sp e c ia lly in th e GF, n u tr ie n t leve ls are h ig h and e u tro p h ica tio n co n tin u e s d u e to in te rnal n u trie n t lo a d in g (P itkänen e t al., 2 0 0 1 , 2 0 0 3 ). In th e p re sen t d a ta set, th e re w as a s lig h t d e c re as in g te n d e n c y in w in te r su rface w a ter D IP c o n cen tra tio n in th e N B P , b u t n o d e c re a se w as o b se rv ed in the
GF. Instead , the D IP c o n ce n tra tio n in th e G F seem s to h av e inc rea sed s in ce th e b e g in n in g o f th e 1990s. F u rth e rm o re , DIN co n cen tra tio n s ig n ifican tly in c reased in th e su rface w a te r of th e G F during th e s tu d y pe rio d . T h is is m o st like ly th e resu lt o f a co m b in a tio n o f N lo a d in g an d in c re a sed N accu m u la tio n in th e G ulf. In ad d itio n to P re leases , b o tto m an o x ia lead s to w eaken ing o f the d en itrifica tio n cap ac ity o f th e se d im e n t an d acc u m u la tio n o f am m o n iu m , w h ich is o x id iz e d into n itra te w hen transported to th e su rface w a ter (K u p a rin e n an d T u o m in en , 2 001). T he am o u n t o f D IN in re la tio n to th e o th e r in o rg an ic nutrien ts (DIP, S i0 4) w as a lso fo u n d to g e n e ra lly in c re a se in the study area, w h ich m ay h av e h ad c o n se q u en c es fo r th e p h y to p lan k to n co m m u n ity (d iscu ssed below ).
s u b ito e .V . l i c e n s e d c u s to m e r c o p y s u p p l ie d a n d p r in te d fo r F la n d e r s M a r in e In s titu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )
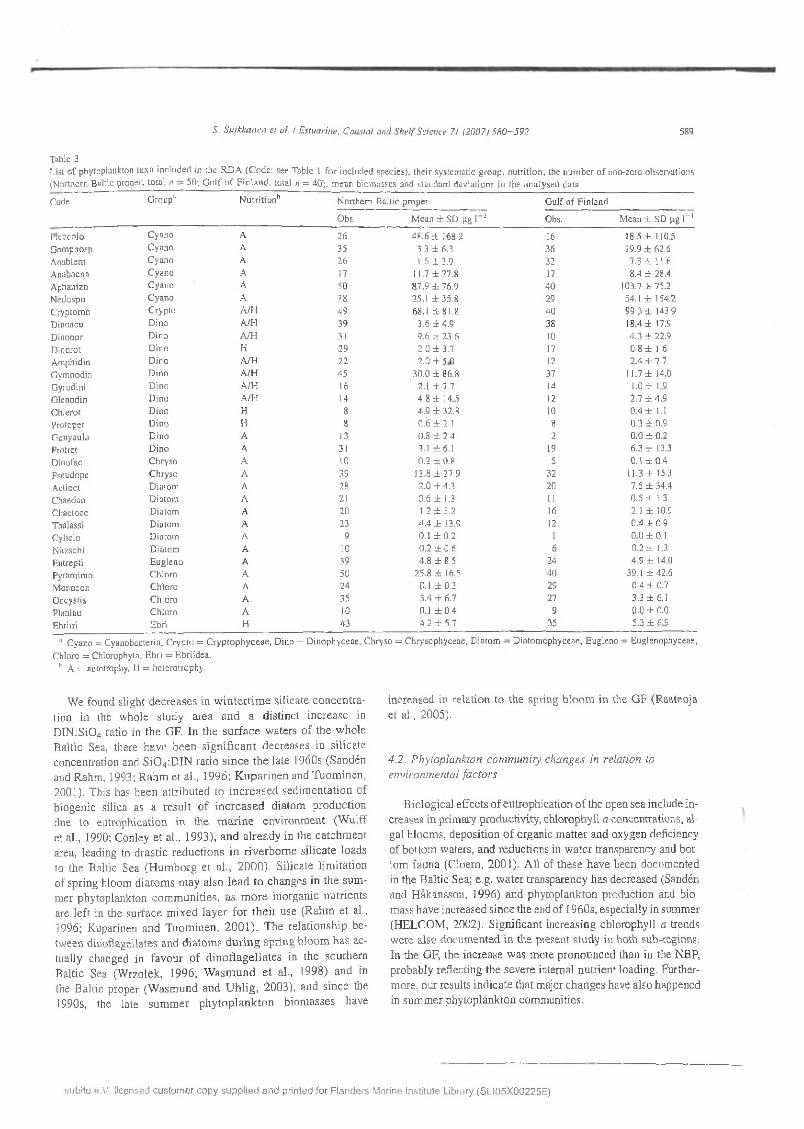
S. S u ik k a n e n et a i / E stuarine , C oasta l anti S h e l f Science 71 (2007) 5 8 0 - 5 9 2 589
Table 3List o f phytoplankton taxa in c luded in the R D A (C ode : see T able 1 fo r inc luded species), th e ir system atic g roup , n u tritio n , the n um ber o f non-zero observations (N orthern Baltic proper, to tal ;i = 50; G u lf o f F in la n d , to tal n = 40 ). m ean b iom asses and standard deviations in the an a ly sed da ta
Code G roup11 N u tritio n 11 N orthern B altic proper G ulf o f F in land
Obs. M ean ± SD pg 1 1 Obs. M ean ± SD pg I“ 1
Picocolo C yano A 26 48 .6 ± 168.2 16 18.5 ± 110.5G om phosp C yano A 35 3.3 ± 6.3 36 19.9 ± 6 2 .6Anablem C yano A 26 1.6 ± 3.9 32 7.3 ± 11.6A nabaena C yano A 17 11.7 ± 77.8 17 8 .4 ± 28.4A phanizo C yano A 50 87.9 ± 76.9 40 103.7 ± 75.2Noduspu C yano A 38 25.1 ± 35.8 29 54.1 ± 154.2Cryptom o C ryp to A h i 49 68.1 ± 81.8 40 99 .3 ± 143.9D inoacu D ino A/I-I 39 3.6 ± 4.9 38 18.4 ± 17.9
D inonor D ino A /H 31 9.6 ± 23.6 10 4 .3 ± 22.9D inorot D ino H 29 2 .0 ± 3 .7 17 0.8 ± 1.6A m phidin D ino A/M 22 2 .0 ± 5 .0 12 2 .4 ± 7.7G ym nodin D ino A /H 45 30 .0 ± 86.8 37 11.7 ± 14.0
G yrodini D ino A /H 16 2.1 ± 7 .7 14 1.0 ± 1.9
G lenodin D ino A /H 14 4,8 ± 14.5 12 2.7 ± 4.9O blerot D ino H 8 4.9 ± 32.8 10 0 .4 ± 1.1P rotoper D ino H 8 0 .6 ± 2 .1 8 0.3 ± 0.9
G onyaula D ino A 13 0.8 ± 2.4 2 0 .0 ± 0.2
Protret D ino A 31 3.1 ± 6 .1 19 6.3 ± 13.3
D inofac C hryso A 10 0.2 ± 0.8 5 0.1 ± 0.4
Pseudope C hryso A 39 13.8 ± 2 7 .9 32 11.3 ± 15.3Actioct D iatom A 28 2 .0 ± 4 .3 20 7.5 ± 34.4
Chaedan D iatom A 21 0 .6 ± 1.3 11 0.5 ± 1.3
C haetoce D iatom A 20 1.2 ± 3 .2 16 2.1 ± 10.9
Thalassi D iatom A 23 4 .4 ± 13.9 12 0 .4 ± 0,9
Cyliclo D iatom A 9 0.1 ± 0 .2 1 0.0 ± 0 .1
N itzschi D iatom A 10 0 .2 ± 0 .6 6 0.2 ± 1.3
Eutrepti E ug leno A 39 4.8 ± 8.5 24 4.9 ± 14.0
Pyram im o C hlora A 50 25 .8 ± 16.5 40 39.1 ± 4 2 .6
M onocon C hlora A 24 0.1 ± 0 .3 29 0 .4 ± 0.7
O ocystis C hlo ra A 35 3.4 ± 6.7 27 3.3 ± 6 .1
Planlau C hlora A 10 0.1 ± 0 .4 9 0.0 ± 0 ,0
Ebri tri Ebri H 43 4.2 ± 5.7 35 5.3 ± 6 .9
;l C yano = C yanobacteria. C ryp to = C ry p to p h y c e a e , D ino = D inophyceae , C hryso = C hrysophyceae , D ia tom = D iatom ophyceae, E ugleno = E uglenophyceae,
C hlora = C hlorophyta, E bri = E briidea . h A = autotrophy, H = hetero trophy.
W e found slight d ec reases in w in te r tim e silica te co n cen tra tion in the w hole study area a n d a d istinc t in crease in D IN :S i0 4 ratio in th e GF. In the su rfa c e w aters o f th e w hole B altic Sea, there have been s ig n ific a n t decreases in silicate concentration and S i0 4:D IN ratio s in c e th e late 1960s (Sandén and Rahm , 1993; R ahm et al., 1996; K u p a rin en and T uom inen , 2001). This has been a ttribu ted to in c re a se d sed im en tatio n o f b iogenic silica as a resu lt o f in c re a se d d iatom p roduction due to eutrophication in th e m a r in e en v ironm en t (W ulff et al., 1990; Conley e t al., 1993), a n d a lread y in the catchm en t area, leading to d rastic red u ctio n s in r iv e rb o m e silica te loads to th e Baltic Sea (H u m b o rg e t a l., 2 0 0 0 ). S ilica te lim ita tion o f spring bloom diatom s m ay also le a d to changes in th e sum m er phytoplankton co m m u n itie s, a s m o re inorganic nu trien ts are le ft in th e su rface m ix ed la y e r fo r th e ir use (R ahm e t al., 1996; Kuparinen and T u om inen , 2 0 0 1 ) . T h e re la tio n sh ip b e tw een d inoflagellates and d ia tom s d u r in g sp rin g b lo o m has actually changed in fav o u r o f d in o fla g e lla te s in th e sou thern B altic Sea (W rzoiek , 1996; W asm u n d e t al., 1998) an d in the B altic proper (W asm u n d and U h lig , 2 003), and since the 1990s, the late su m m er p h y to p la n k to n b iom asses have
in creased in re la tion to th e spring b lo o m in the G F (R aateoja et al., 2005).
4.2. P hytop lankton co m m unity ch anges in relation to environm enta l fa c to rs
B iolog ical effects o f eutrophication o f the open sea include in creases in prim ary productiv ity , chlorophyll a concentrations, algal b loom s, deposition o f organic m atte r and oxygen deficiency o f bo ttom w aters, and reductions in w a te r transparency and bottom fau n a (C loem , 2001). A ll o f th ese h ave been docum ented in the B altic Sea; e .g . w a ter transparency has decreased (Sandén and H âkansson, 1996) and phytoplankton production and biom ass have increased since the end o f 1960s, especially in sum m er (H ELC O M , 2002). S ign ifican t increasing chlorophyll a trends w ere also docum ented in the present study in both sub-regions. In th e GF, the increase was m ore p ronounced than in the NBP, p robably reflecting the severe internal nutrient loading. Furtherm ore, our results ind ica te that major changes have also happened in sum m er phytop lank ton com m unities.
s u b ito e .V . l ic e n s e d c u s to m e r c o p y s u p p l ie d a n d p r in te d fo r F la n d e r s M arin e In s ti tu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )
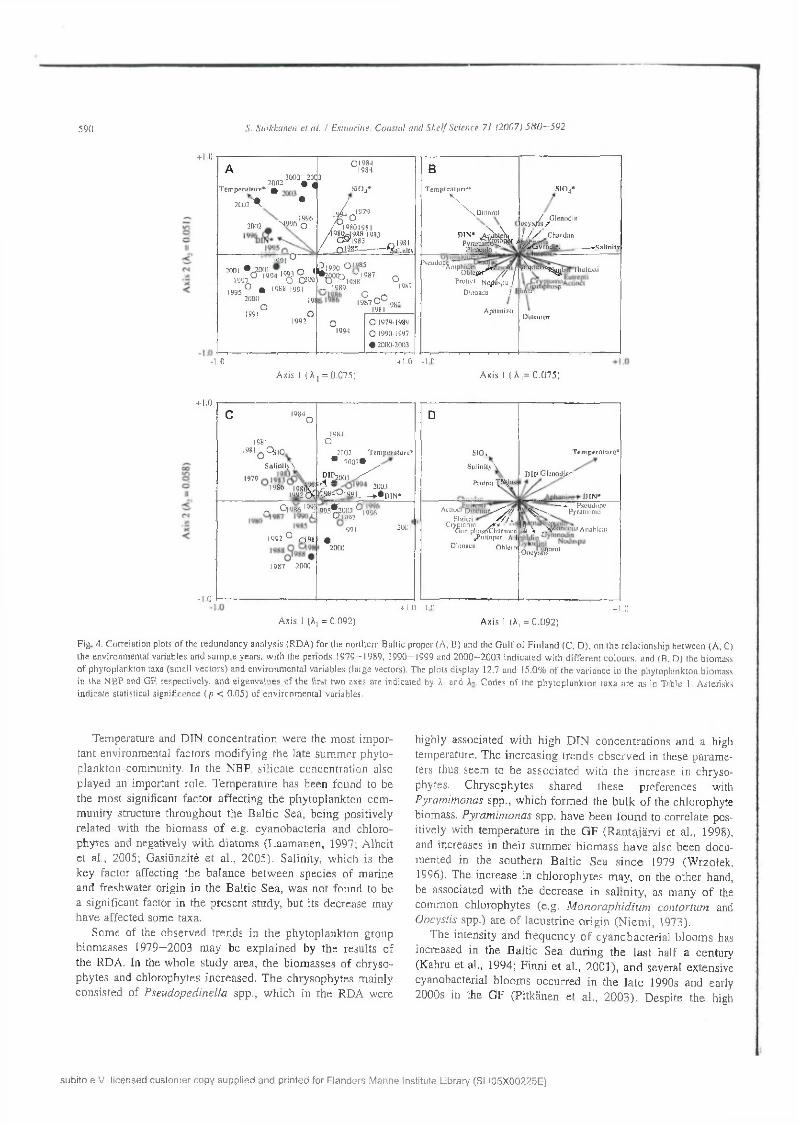
590 S . S u ikkanen c l at. / Estuarine . C oasta l and S h e l f Sc ience 71 12 0 0 7 ) 5 8 0 —592
+ 1.0
A2 0 0 2 200 V J
T e m p e r a tu r e * ©
2 0 0 2 ^ . * 1096
2002 N W f t o
0 1 9 8 41984
3
S I O /
' / o '979/ 1 9 8 0 1 9 8 1
/ 9 8 0 -1 9 8 8 1983 / 0 9 ) 9 8 3 | 9 8 | / _JDÍ2S¿—- — f a l t a n *
BT e m p e ra tu re *
* ^ S\ vD ¡nom i
D IN * yX XP y ra im rn rS fR g t ^
P leocola
S i O /
/ G lenod in o c y s f í s /
/ / / C h aed an
U O f C i y r o d in i ---------» S a l in i ty
1991 °
2001 #d°Vù)4 « » a O «1997 U o a o o
l 9 9 5 ° « IOS» 19912000 i9(
o1991 O
1992
3 , 9 9 0 0 1 9 8 5
V w * 7 O„ 1989 I9 »’’
, â o o |W ,1981
>seuJ° P A ,Í íh íd i* O b le s s ^ / / ¡Prom et NojKÍspu /
D inoacu /
A p han izo
yN o c í5Rp^ iuu T h a la ss i
bit tr i r
D in o n n rO
1994O 1979-1989
O 1990-1997
• 2000-2003
- 1.0 + 1 .0 - 1 .0
A xis 1 (A , = 0 .075) A xis 1 (A ,= 0 .0 7 5 )
+ 1.0
- 1 .0
C '9s40
1981W q Q s i o .
S a lin ily X
1979 1786 I98K L 1992 ÔÇ
1984O
2002 T e m p e r a tu r e '' • 2002®
j » > Xi d 200.1 fc » ¿ O i9 9 j_____ ► • d i n *
D
S io ,
S a lin U y V
P ru tre t W h S!
T e m p e r a tu r e *
D I P G le n o d in ^
D IN *
- , O |086 m i
1992 ° r )9 8
1987 2000
™ f e 7° .9 9 6
1991 20011 ©
2000
A ctluct D \ n o n a ¡ ^ ^ ¿ V Ebrllrl v y y ,
C ry p to m o / * ‘ G o m p litw iC h a e m e d
»Pro toper A li D inoacu Ohlercn
— ■» Pscu tlupe V ^ s . P y ram im u
1 \ /^ « » n u c o n A n a h lc m
O o c y s (ß ü ral
+ 1 .0 - 1 .0 + 1.0
A xis 1 (A, = 0 .092) A xis 1 (A, = 0 .0 9 2 )
F ig . 4 . C orre la tion plots o f the red u n d an c y an a ly sis (R D A ) fo r the northern B altic proper (A, B) and the G u lf o f F in la n d (C . D ), on th e re la tio n sh ip betw een (A , C) the environm ental variables and s am p le years , w ith the periods 1 9 7 9 - 1989. 1 9 9 0 -1 9 9 9 and 2 0 0 0 -2 0 0 3 in d ica ted w ith d iffe re n t co lo u rs , and (B , D ) the biom ass o f phy top lank ton taxa (sm all vec to rs) and en v iro n m en ta l va riab les (large vectors). T he plots d isp lay 12.7 and 15.0% o f th e v arian ce in th e phy top lank ton biom ass in the N B P and GF, respectively , a n d e igenva lues o f th e first tw o axes are indicated by A| and A - C odes o f the p h y to p lan k to n lax a a re a s in T able 1. A sterisks in d ica te statistical s ign ificance ( / ; < 0 .0 5 ) o f en v iro n m en ta l variables.
T em perature and D IN c o n ce n tra tio n w ere the m ost im p o rtan t environm ental fac to rs m o d ify in g th e late su m m er p h y to p lank ton com m unity. In th e NBP, s ilic a te co n cen tra tio n also p lay ed an im portan t ro le. T em p era tu re has been found to be th e m ost significant fa c to r a ffec tin g th e p h y to p lan k to n co m m u n ity structure th ro u g h o u t th e B a ltic Sea, b e in g positively re la ted w ith the b io m ass o f e.g . cy an o b a c te ria and ch lo ro p h y tes and negatively w ith d ia to m s (L aam an en , 1997; A lh e it e t al., 2005; G asiüna itè e t a l., 2 0 0 5 ). S alin ity , w h ich is the k ey fac to r affecting th e b a lan ce b e tw een sp ec ies o f m arine an d freshw ater orig in in th e B a ltic S ea , w as no t found to be a s ig n ifican t factor in th e p re sen t study , bu t its decrease m ay h av e affec ted som e taxa.
S om e o f the obse rv ed tren d s in th e p h y to p lan k to n group b iom asses 1979—200 3 m ay b e ex p la in ed by th e resu lts o f th e R D A . In th e w h o le s tu d y area , th e b io m asses o f ch ryso p h y tes and ch lo rophy tes in creased . T h e ch ry so p h y tes m ain ly co n sis ted o f P seu d o p ed in e lla spp ., w h ich in th e R D A w ere
highly assoc ia ted w ith h ig h D IN co n ce n tra tio n s an d a high tem peratu re. T h e in c re a s in g tre n d s o b se rv e d in these p a ram eters thus seem to b e a sso c ia ted w ith th e in crease in ch ry so phytes. C h ry so p h y tes sh a red th ese p refe ren ces with P yram im onas spp ., w h ich fo rm e d the b u lk o f the ch lo rophy te b iom ass. P yra m im o n a s spp . h av e been fo u n d to co rre la te positively w ith tem p era tu re in th e G F (R a n ta jä rv i et a l., 1998), and increases in th e ir su m m e r b io m a ss h a v e also b een d o cu m ented in the so u th e rn B a ltic S ea s in c e 1979 (W rzo lek , 1996), T h e in crease in c h lo ro p h y te s m ay, on the o th e r hand, be assoc ia ted w ith th e d e c re a se in sa lin ity , as m any o f the com m on ch lo ro p h y tes (e .g . M o n o ra p h id iu m co n to r tu m and O ocystis spp .) are o f lacu s tr in e o rig in (N iem i, 1973).
T h e in tensity and freq u en cy o f c y ano bac teria l b lo o m s has increased in th e B a ltic S e a d u rin g th e la s t h a lf a cen tury (K ahru e t al., 1994; F in n i e t a l., 2 0 0 1 ), a n d several extensive cyanobacteria! b lo o m s o ccu rred in th e la te 1990s and early 2000s in th e G F (P itk än en e t a l., 2 0 0 3 ). D esp ite th e high
s u b ito e .V . l i c e n s e d c u s to m e r c o p y s u p p l ie d a n d p r in te d fo r F la n d e r s M a rin e In s titu te L ib ra ry (S L 1 0 5 X 0 0 2 2 5 E )

S. S u ik ka n en e t a!. / E stuarine , C o asta l a n d S h e l f Science 71 (201)7) 5 8 0 - 5 9 2 591
v a ria b ility in the b iom ass da ta , w e o b se rv ed a s ig n ifican t in c re a se in cyanobacteria ! b io m a ss , m a in ly fo rm ed by A p h a n izo m e n o n flo s-a q u a e an d N o d u la r ia sp u m ig en a , in the G F during 1 9 7 9 —2003. U sing the B M P da ta , W asm u n d and U h lig (2003) a lso fo u n d an au tu m n in c rease in c y an o b a c te ria in th e B altic p ro p e r in 1979—1999, b u t in th e sa m p le s from 2 0 0 0 —2003, th is trend seem ed to h av e lev e lled o ff. T h e in crease in the G F w as possib ly asso c ia ted w ith th e in c rease in tem pera tu re a n d in o rgan ic nu trien ts: in th e R D A , gom pho sp h aerio id s w e re the only cy an o b ac te ria l ta x o n n o t a sso c ia ted w ith high tem p e ra tu re s and h igh D IN co n ce n tra tio n . In th e GF, g o m p h o sp h a e rio id s are m ost a b u n d a n t d u rin g the cool, w e ll-m ix ed s p r in g and au tum n p e rio d s (L aa m a n e n , 1997). T em pera tu re ( > 1 6 °C ) is the m ain fa c to r d e te rm in in g the o n se t and in ten s ity o f b loom s d o m in a ted by N . sp u m ig e n a in the B altic S ea (W asm u n d , 1997; K an o sh in a e t a h , 2 0 0 3 ), w h ereas the b io m a s s o f A ph a n izo m en o n sp p . m ay be s tro n g ly re la ted to D IN c o n ce n tra tio n (Jaan u s and P e llik k a , 2 0 0 3 ). A lso the s lig h t dec re a s e in salin ity , co m b in e d w ith the in creased tem p era tu re , m a y have favoured e .g . th e p ico c o lo n ia l cy an o b acte ria . A n a b a e n a spp. and A p h a n izo m en o n spp . (L aam an en , 1997; H äll- fo rs , 2004).
O n the o ther h and , w e fo u n d no s tro n g asso c ia tio n betw een w in te r D IP co n cen tra tio n an d th e n itro g en -fix in g c y an o b ac te r ia l b io m ass , a lth o u g h in c reased c y an o b a c te ria l b loom s in the G F have often been a ttrib u ted to in c reas in g D IP c o n cen tra t io n s and d ecreas in g D IN .'D IP ra tio s , o rig in a tin g from the se d im e n t efflux (P itk än en e t al., 2 0 0 3 ). In stead , the b io m ass o f b o th A ph a n izo m en o n spp. and N o d u la r ia sp u m ig en a co rre la te d positively w ith w in te r D IN co n ce n tra tio n . I t h as been s ta te d , how ever, th at c y an o b a c te ria l b lo o m s in the G F have an a sso c ia tio n w ith the a m o u n t o f e x ce ss D IP le ft o v e r from th e spring b lo o m (K iirik k i et a l., 2 0 0 1 ), and no t so m uch w ith the n u trien t c o n ce n tra tio n s d u rin g m id -w in ter, w hen th e sam p les fo r th is s tudy w ere tak e n . A s d iscu ssed above, th e v e rn al b iom ass has d e c re a se d (R a a teo ja e t al., 2 005), p oss ib ly leav ing a g re a te r a m o u n t o f D IP fo r the use o f sum m er c o m m u n itie s , in c lu d in g th e c y an o b a c te ria , w h ich h av e in c re a se d in b io m ass . In c re a se d n itro g e n -fix in g cy an o b acte ria m a y , on the o th er hand , h ave c o n tr ib u te d to th e in creased D I N level in the w in te r su rfa ce w a te r (K o n o n en and N iem i, 1 9 8 4 ). M oreover, in b o th su b -a rea s , th e re w as a d ecreas ing ten d e n cy in early su m m e r D IN co n ce n tra tio n s , w h ich m ay h a v e favoured th e c y a n o b a c te r ia o v e r o th er sp ec ies unab le to f ix a tm ospheric N.
T h e b iom ass o f c ry p to p h y te s d e c re ased in the GF, w h ich m a y be co nnec ted w ith th e in c re a se in tem p era tu re : in the R D A , the c ry p to p h y tes w e re a sso c ia te d w ith low tem p era tu res . In te re s tin g ly , the decrease in c ry p to p h y te s co in c id ed w ith the in c re a s e in c y an o b ac te ria in th e GF. E x tra c ts o f the m o st c o m m o n B altic c y an o b ac te ria (A p h a n izo m e n o n flo s -a q u a e , N o d u la r ia spum igena an d A n a b a e n a sp p .) h ave recen tly been fo u n d to inh ib it th e g ro w th o f c ry p to p h y te s in lab o ra to ry exp e rim e n ts e m p lo y in g a n a tu ra l B a ltic S e a p lan k to n co m m unity (S u ik k a n e n e t ah , 2005).
In th e NBP, m an y o f th e d in o p h y te s (e .g . D in o p h ysis ro tu n d a ta , G ym nod in ium spp ., O b lea ro tu n d a co m p lex ) m ay have
been favoured by the in c re a s in g w a ter tem p era tu re and /or DIN concentra tion (cf. N iem i, 1973; K ononen , 1988; H ällfors,2004). A lso, the general in c re a se o f sm all-celled species (chrysophytes, ch lorophy tes), a g g reg a te s o f ageing cyan o b acte ria w ith associated b ac teria a n d excre ted D O M m ay have benefited the dinoflagellates, m a n y o f w h ich are m ixo- or het- ero trophs. W asm und and U h lig (2003) a lso reported an increase in sp rin g and sum m er d in o p h y te b iom ass in the N B P in 1979— 1999, coincid ing w ith a decrease in d ia tom s. In c reasing dinoflag ella tes m ay be a cause o f c o n c e n t, as so m e B altic dinoflagellata species are also po ten tially to x ic (H E L C O M , 1996).
In sp ite o f the ra th e r low sa m p lin g frequency and the high variab ility in som e variables, w e b e lieve that the B M P data gives a reliable p ic tu re o f th e genera l changes in th e pelagic ecosystem o f the B altic Sea. T h e ra th e r low percen tag e o f variance in the phytop lankton d a ta exp la in ed by the env iro n m en tal variab les in the R D A is quite com m on in com plex eco lo g ica l m onito ring data ( te r B raak and S m ilauer, 1998), and is partly due to o th er fac to rs , not con sid ered in th is study, in fluencing phytop lankton p o p u la tio n s (e.g. grazing , sed im entation , m icronutrien ts, in h ib ito ry substances, light conditions).
To conclude, the long-term d ev elo p m en t o f the late sum m er phytop lankton com m unities in th e northern B altic S ea seem s to reflect the com bination o f o n g o in g eu tro p h ica tio n and clim ate- driven changes in hydrography. T h e sign ifican t in crease o f ph y to p lank ton biom ass, ind icated as ch lorophyll a concen tra tion , is due to a considerab le increase in cyanobacteria , v a rio u s flagellates and chlorophytes. O ur resu lts ag ree w ith those o f Kononen (1988), w ho also no ticed an in crease in n an o p lan k to n ic flagellates, ch lorophytes, p ico co lo n ia l cy an o b acte ria and hetero tro- ph ic species in the G F in 1972—1985, suggesting an increase o f th e re la tive im portance o f reg en e ra ted p roduction an d the m ic rob ia l loop. A cy an o b acte ria -flag ella te dom inated com m unity is generally poor food fo r m an y g raze rs (O fficer an d Ryther, 1980), but the large-scale co n seq u en ces o f the changes in phyto p lank ton fo r h igher tro p h ic lev e ls and th e ir connections w ith observed long-term changes in e.g . Zooplankton, ben th ic and fish com m unities are ye t unknow n.
Acknowledgements
W e are grateful to th e s ta ff o f the F in n ish In stitu te o f M arine R esearch fo r sam pling , h y d rograph ical m easurem ents and chem ica l analyses. J. E ng strö m -Ö st, M . K arjalainen and M. V iitasalo are acknow ledged for co n structive com m ents on th e m anuscrip t. F inan cia l su p p o rt to S .S. was p rov ided by th e M aj and Tor N essling F o u n d a tio n and the W alter and A n d rée de N ottbeck F oundation .
References
A lheit, J ., M öllm ann, C ., D u tz , J., K ornilovs, G ., L oew e, P., M ohrho lz , V.,W asm und, N ., 2005 . S ynchronous eco log ica l reg im e sh ifts in th e centralB altic Sea and the N orth Sea in the la te 1980s. IC E S Journal o f M arineS cience 62, 1 2 0 5 -1 2 1 5 .
te r B raak , C .J.F., 1994. C anonical com m unity o rd ina tion . P art 1: B as ic theoryand linear m ethods. E cosc ience 1, 1 2 7 -1 4 0 .
s u b ito e .V . l i c e n s e d c u s to m e r c o p y s u p p lie d a n d p r in te d fo r F la n d e r s M a rin e in s ti tu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )

592 S . S u ikkanen er al. / E stuarine , C o asta l a n d S h e l f Sc ien ce 71 (2 0 0 7 ) 5 8 0 —592
te r B raak. C.J.F., S m ilau er, P., 1998. C A N O C O R efe ren ce M an u a l and U ser's G u ide to C A N O C O fo r W indow s: S o ftw are fo r C ano n ica l C o m m un ity O rd in a tio n (version 4 ). M ic ro c o m p u te r P ow er, I thaca , NY, 351 pp.
C loem , J .E ., 2001. O u r e v o lv in g co n c ep tu a l m ode l o f th e coasta l eu troph ication p roblem . M arine E c o lo g y P ro g ress S eries 210 , 223—253.
C onley , D .J., S chelske , C .L ., S to erm er, E .F ., 1993. M odifica tion o f the b iogeo chem ica l cyc le o f s i lic a w ith eu tro p h ica tio n . M arin e E co logy Progress S eries 101, 1 7 9 -1 9 2 .
F inn i, T., K ononen, K ., O lsonen , R ., W alls tröm , K ., 2001 . T h e h isto ry o f cyanobacterial b loom s in th e B a ltic Sea. A m bio 30, 172— 178.
F linkm an , J., A ra , E ., V uorinen , I., V iitasa lo , M ., 1998. C h an g es in northern Baltic Zooplankton and h errin g n u tritio n fro m 1980s to 1990s: top-dow n and bottom -up p rocesses a t w ork . M arin e E co logy P rog ress S eries 1 6 5 ,127—136.
G asiflna ité , Z .R ., C ardoso , A .C ., H e isk an en , A .-S ., H enriksen , P., K auppila , P., O len ina , I., P ilk a ity tè , R ., P u rin a , I„ R az inkovas, A ., S agert, S., S chubert, H „ W asm und , N ., 2 005 . S easo n a lity o f co a sta l phytop lank ton in th e Baltic S ea: in flu en ce o f sa lin ity a n d eu troph ica tion . E stuarine, C oasta l and S h e lf S c ie n ce 65 , 2 3 9 —252.
G ilbert, R .O ., 1987. S ta tis tic a l M e th o d s fo r E n v iro n m en ta l P o llu tion M on ito ring. Van N ostrand R ein h o ld C o ., N ew Y ork, 320 pp.
H ällfors, G ., 2004. C h e c k lis t o f B a ltic S ea P h y to p lan k to n S pec ies (Including S om e H etero trophic P ro tis tan G ro u p s). B altic S ea E n v iro n m en t P roceedings 9 5 , 208 pp.
H E L C O M , 1988. G u id e lin es fo r th e B a ltic M o n ito rin g P rog ram m e fo r the T h ird S tage. P art D . B io lo g ica l D e te rm in an d s . B altic S e a E nv ironm ent P roceedings 27D , 161 pp.
H E L C O M , 1996. T h ird P e rio d ic A ssessm en t o f th e S ta te o f the M arine Enviro n m en t o f the B altic S ea 1989—1993. B ack g ro u n d D o cu m en t. B altic Sea E nvironm ent P ro ceed in g s 64B , 2 5 2 pp.
H E L C O M , 2002. E n v iro n m en t o f th e B a ltic S ea A rea 1994— 1998. B altic Sea E nvironm ent P roceed ings 82B , 2 1 5 pp .
H um borg , C., C onley, D .J., R ah m , L ., W ulff, F ., C ociasu , A ., Ittekko t, V.,2000 . S ilicon re ten tio n in riv e r b a s in s : fa r-reach in g effec ts on b iogeochem istry and aquatic food w ebs in co a sta l m a rin e env ironm en ts. A m bio 29, 4 5 - 5 0 .
Jaanus , A ., Pellikka, K „ 2 0 0 3 . D oes ex cess iv e p h o sp h o ru s necessa rily cause increasing b iom ass o f d ia zo tro p h ic c y a n o b ac te ria? P ro ceed in g s o f th e E stonian A cadem y o f S c ie n ces B io logy . E c o lo g y 52, 2 0 5 —217.
K ahru , M ., H orstm ann, U ., R ud , O ., 1994. S a te lli te de tec tio n o f inc reased cy anobacteria b loom s in th e B a ltic S ea : n a tu ra l fluc tuation o r ecosystem change? Am bio 23 , 4 6 9 —472 .
K anosh ina, I., Lips, U ., L e p p an en , J .-M ., 2003 . T h e in fluence o f w ea th er con ditions (tem perature a n d w in d ) o n cy an o b ac te ria l b lo o m deve lopm en t in the G u lf of F in land (B a ltic S ea). H a rm fu l A lg ae 2, 2 9 —41.
K iirikki, M ., Inkala, A ., K uosa , H ., P itk ä n en , H ., K uusisto , M ., S arkku la , J „2001. E valuating the e ffec ts o f n u tr ien t lo a d reduc tion o n the b iom ass of toxic n itrogen-fix ing cy a n o b a c te r ia in the G u lf o f F in la n d , B altic Sea. Boreal E nv ironm ent R esea rc h 6, 131—146.
lo nonen, K ., 1988. P h y to p la n k to n s u m m e r a s sem b lag es in re la tion to env ironm enta l factors a t the en tra n c e to th e G u lf o f F in land d u rin g 1972— 1985. K ie ler M eeresfo rschungen S o n d e rh e ft 6 , 2 8 1 —294.
K ononen, K „ N iem i, À ., 1984. L o n g -te rm v a ria tio n o f th e phy top lank ton co m position at the en tran ce to th e G u lf o f F in la n d . O p h e lia S upp l. 3 , 101—110.
K oroleff, F., 1979. M eriv e d en y le is im m ä t k e m ia llise t ana lyysim enete lm ät. M eri 7 , 1 -6 0 (in F inn ish ).
K uparinen , J„ T uom inen , L ., 2 0 0 1 . E u tro p h ica tio n and self-purification: counteractions fo rced by la rg e -sc a le cy c les a n d h y d ro d y n am ic processes. A m bio 30, 190—194.
L aam an en , M .J., 1997. E n v iro n m e n ta l facto rs a ffec ting the occu rren ce o f diffe ren t m o rp h o lo g ica l fo rm s o f cy a nop roka ryo tes in th e northern Baltic S ea . Jou rnal o f P lan k to n R e s e a rc h 19, 1385— 1403.
N iem i, Â ., 1973. E c o lo g y o f p h y to p la n k to n in th e T v ä rm in n e area , S W coast o f F in la n d . I. D y n am ics o f h y d ro g rap h y , nu trien ts , ch lo ro p h y ll a and phyto p lan k to n . A c ta B o ta n ica F e n n ic a 100, 1—68.
O fficer, C .B ., R y ther, J .H ., 1980 . T h e po ssib le im portance o f silicon in marine e u tro p h ica tio n . M arin e E c o lo g y P ro g ress S eries 3 , 83—91.
P itk än en , H ., L e h to ran ta , J . , R a ik e , A ., 2001 . In te rnal n u trien t fluxes counterac t d ec reases in ex te rn a l lo a d : th e case o f the estuaria! ea stern G u lf o f Finla n d , B a ltic S ea . A m b io 3 0 , 195—201.
P itk än en , H ., L e h to ran ta , J ., P e lto n e n , H ., L a ine , A ., K otta , J ., K otta , I„ M o sk a len k o , P., M äk in en , A ., K an g as , P., P erttilä , M ., K iirikk i, M ., 2003. B en th ic re lea se o f p h o s p h o ru s a n d its re la tio n to en v iro n m en ta l conditions in the estuaria! G u lf o f F in la n d , B altic S ea , in the ea rly 2000s. Proceedings o f th e E s to n ian A cad em y o f S c ie n ces B io logy . E co lo g y 52 , 173—192.
R aa teo ja , M ., S eppälä , J . , K u o sa , H ., M y rb erg . K ., 2005. R e c e n t ch an g es in trop h ic s ta te o f the B altic S ea a lo n g S W c o a st o f F in land . A m bio 3 4 ,1 8 8 — 191.
R ahm , L., C on ley , D ., S an d én , P., W ulff, F., S tâ lnacke , P., 1996. T im e series an a ly sis o f n u trien t in p u ts to the B altic Sea and ch a n g in g D S i:D lN ratios. M arin e E c o lo g y P rog ress S e r ie s 130, 2 2 1 -2 2 8 .
R an ta jä rv i, E ., G ran , V., H ä llfo rs , S ., O lsonen, R ., 1998. E ffec ts of en v iro n m en ta l fac to rs on th e p h y to p lan k to n c o m m u n ity in the G u lf of F in la n d — u n a tte n d e d h ig h f req u en c y m easu rem en ts and multivariate ana ly ses . H y d ro b io lo g ia 3 6 3 , 127—139.
R önkkönen , S ., O javeer, E ., R a id , T ., V iitasalo , M „ 2004 . L o n g -te rm changes in B a ltic h e rr in g (C lu p e a h a r e n g u s m em b ra s) g row th in the G u lf o f Finland . C a n a d ian J o u rn a l o f F is h e r ie s and A quatic S c ie n ces 61, 219—229.
S a lm i, T„ M äättä , A ., A n tti la , P., R uoho -A iro la , T ., A m nell, T., 2002, D e te c tin g T rends o f A n n u a l V alues o f A tm ospheric P o llu tan ts by the M a n n -K e n d a ll T est a n d S e n 's S lo p e E stim ates — th e E xcel Template A p p lic a tio n M A K E S E N S . F in n ish M e teo ro log ica l In s titu te , Helsinki, P u b lic a tio n s on A ir Q u a lity 31, 35 pp.
S an d én , P., H âk an sso n , B ., 1996. L o n g -te rm trends in S ecch i d ep th in the B a ltic S ea . L im n o lo g y and O c ean o g ra p h y 4 1 , 346—351.
S an d én , P., R ahm , L ., 1993. N u tr ie n t trends in th e B a ltic S ea , Environm etrics 4 , 7 5 - 1 0 3 .
S u ik k an en , S ., F is taro l, G .O ., G ra n é li , E ., 2005 . E ffec ts o f cy an o b ac te ria l al- le lo ch em ica ls on a n a tu ra l p la n k to n com m unity . M arin e E co lo g y Progress S erie s 2 8 7 , 1—9.
U term öh l, H ., 1958. Z u r V erv o llk o m m n u n g d e r qu an tita tiv en Phytoplankton- M eth o d ik . M itte ilu n g en . In te rn a tio n a le V erein igung fü r T h e o re tisch e und A n g ew an d te L im n o lo g ie 9 , 1—38 .
W asm und , N ., 1997. O ccu rren c e o f cy a n o b ac te ria l b loom s in the B a ltic Sea in re la tion to en v iro n m en ta l co n d itio n s . In te rna tiona le R evue d e r Gesamten H y d ro b io lo g ie 82 , 169—184.
W asm und , N ., U hlig , S ., 2003 . P h y to p la n k to n trends in the B altic Sea. ICES Jo u rn a l o f M arin e S c ie n c e 60 , 177—186.
W asm und , N ., N ausch , G ., M atth äu s , W., 1998. P h y to p lan k to n sp rin g bloom s in th e so u th e rn B a ltic S e a — sp a tio -tem p o ra l d ev e lo p m en t an d long-term trends. Jo u rn a l o f P lan k to n R e se a rc h 2 0 , 1 0 9 9 -1 1 1 7 .
W rzo fek , L „ 1996. P h y to p la n k to n in th e G dansk B a s in in 197 9 -1 9 9 3 . O cean o lo g ica l S tu d ie s 1-2, 8 7 - 1 0 0 .
W ulff, F., R ah m , L ., 1988 . L o n g -te rm , seaso n a l a n d sp a tia l v a r ia tio n s of n itrogen , p h o sp h o ru s and s ilic a te in the B altic : an overv iew . Marine E n v iro n m en ta l R esea rc h 26 , 19—37,
W ulff, F., S tig e b ran d t, A ., R ah m , L ., 1990. N utrien t d ynam ics in the Baltic S ea . A m b io 19, 1 2 6 -1 3 3 .
s u b ito e .V . l i c e n s e d c u s to m e r c o p y s u p p l ie d a n d p r in te d fo r F la n d e r s M arin e In s titu te L ib ra ry (S L I0 5 X 0 0 2 2 5 E )







