
AGGRESSIVE BEHAVIOR Volume 9, pages 133-155
Effects of Midbrain Central Gray Lesions on Spontaneous and Electrically Induced Aggression in the Rat J. MOS, J.H.C.M. Lammers, A.M. van der Poel, B. Bermond, W. Meelis, and M.R. Kruk
University Medical Centre, Department of Pharmacology, Sylvius Laboratories, Wassenaarseweg 72, 2333 AL Leiden, The Netherlands
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Large electrolytic lesions were placed in the midbrain central gray of male rats. Their effects on hypothalamically induced aggression, switch-off behaviour, and locomotion were investigated. A number of these animals were also tested for territorial intermale aggression in order to compare electrically induced and spontaneous aggression. Large lesions resulted in an increase of the current threshold to induce aggression by hypo- thalamic stimulation. Smaller, but still quite large, lesions decreased the threshold current for hypothalamic aggression. After the operation a decrease in the threshold for switch-off was present, both in the experimental and the control group. Current thresholds for locomotion were decreased after the lesions only in the experimental group.
Spontaneous aggression was temporarily decreased after the lesion. No indication was found that other behavioural elements of the animal were distorted by the lesion.
The parallel between the effects on spontaneous and electrically induced aggression makes it attractive to ascribe a role to the neural circuit of hypothalamus and central gray in territorial aggression. However, even with large lesions the animals were still capable of fighting, hence the central gray is not indispensable. An attempt was made to explain the differential effects that differently sized central gray lesions have on hypothalamic aggression.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Key words: rat, midbrain central gray, lesion, ESB, intraspecific aggression, hypothalamus
INTRODUCTION
In a previous paper we postulated that the central gray and the hypothalamus are part of a neural circuit subserving intraspecific aggression in rats [Mos et al, 19821. It was argued that attacks induced by stimulation of this circuit have many character- istic elements in common with offensive aggression as the Blanchards [1977a,b] and Adams [ 19791 have defined it. Aggression induced by hypothalamic stimulation is
Received for publication August 2, 1982; accepted March 8, 1983.
Address correspondence to J. Mos, Sylvius Laboratories, Department of Pharmacology, Wassenaar- seweg 72,2333 AL Leiden, The Netherlands.
0 1983 Alan R. Liss, Inc.

I34 Mos et a1
accompanied with teeth-chattering and piloerection by the attacking animal. This animal shows no signs of flight, but actively approaches the opponent, sometimes displaying sideways threat. During fighting this animal is always “on top” and never shows the characteristic defensive pattern “on the back” [Blanchard and Blanchard, 1977a,b]. An indication that defensive components may also be evoked stems from the data on the bite patterns. Stimulation-induced attacks result in bite patterns with most lesions on the back and on the head. Since defensive animals almost exclusively bite the head and offensive animals predominantly bite the Back, this pattern may be interpreted as a mixture of offensive and defensive tendencies. Hitherto, this bite pattern has not been settled definitely and it is now investigated in more depth [Olivier et al, 19821. In our view hypothalamically induced attacks must be seen as mainly offensively motivated.
Regarding this hypothalamic-central gray circuit subserving attacks in rats a number of questions remained unanswered. The first question concerns the organization of this neural circuitry. In cats a similar circuit has been described for rage reactions [Hunsperger, 1956; Fernandez De Molina and Hunsperger, 1962; Skultety, 19631. Fernandez De Molina and Hunsperger [ 19621 hypothesized a circuit descending from the amygdala through the hypothalamus to the central gray. In their opinion, the central gray is the most important part of this circuit. Lesioning of the midbrain central gray resulted in a complete blockade of hypothalamically induced reactions when more than 80% of the central gray had been destroyed. Since stimulation of the central gray still produced rage reactions even after lesioning of the hypothalamus, this hypothesis seems plausible.
Recently however, Bandler et a1 [ 19801 have considered the possible importance of ascending projections. They induced predatory attacks by electrical stimulation in the hypothalamus of cats. Knife cuts in the medial forebrain bundle at the level of the anterior hypothalamus/posterior preoptic area ipsilaterally blocked the terminal biting component of the predatory attack in cats. This means that the behaviour during stimulation included approach and seizing the rat, but the bite at the head and neck region which usually comes as the final element in this sequence failed to be displayed. Such a finding may fit in a “descending” model for aggression but leaves room for speculations about the role of ascending projections. Especially the visual aspect of the attacks is now being studied [Bandler et al, 19801.
As we have demonstrated [Mos et al, 19821 both ascending and descending projec- tions arise from hypothalamic attack sites in rats. In cats this has been demonstrated by others [Chi and fly^, 1971a,b] for the quiet bite attack as well as for the “affective” attack or defense induced by hypothalamic stimulation. Since there are several output pathways, it is not clear why central gray lesions should completely block all aggressive acts, offensive or defensive, the more so if ascending projections indeed play an important role as Bandler suggests.
Another question pertains to the importance of the neural circuit for territorial aggression. It is known from studies by Adams that midbrain central gray units are active particularly during defense in cats and rats [Adams, 1968; Pond et al, 19771. Large lesions of the central gray in rats abolish defensive reflexive fighting resulting from footshock [Edwards and Adams, 19741. In cats, defense is diminished following large lesions of the central gray and the adjacent tegmentum [Glusman et al, 1961; Hunsperger, 1956; Fernandez De Molina and Hunsperger, 1962; Skultety, 19631. However, data about the effect of lesions on electrically induced behaviour as well as on spontaneously occurring aggression are scarce [Berntson, 19721. Such an approach

Central Gray Lesions and Aggression in Rats 135
may not only answer the above-mentioned questions but could be particularly useful in assessing the value of electrical brain stimulation studies. Therefore we implanted stimulation electrodes in the hypothalamus and housed a number of these rats in large cages where they could establish a territory. After repeatedly testing the basal level of performance of these animals, lesions were placed in the central gray and the subsequent effects were studied for four weeks.
MATERIAL AND METHODS Animals
Brown-eyed, beige-coloured male Wistar rats (CPBWE-zob), derived from the Central Institute of Laboratory Animals (CPB-TNO, Zeist , The Netherlands) were used as experimental animals. Before the operation, the rats were housed in groups of six to ten in large macrolon cages, in low-noise rooms with a relative humidity of 75 % and a temperature of 22 "C. An inverse day and night cycle was maintained( 14L/ lOD), with night starting at 8 AM. All experiments were performed during the rats' dark period. Food and water were given freely. Prior to operation the animals were handled once daily for a week. After the first operation the rats were housed individually in small macrolon cages or in large home cages where they were allowed to establish their territory.
Male albino Wistar rats (CPBWU-random) weighting between 160 and 200 gm served as partners in both types of aggresstion tests.
Electrodes Bipolar Pt 10 % Ir electrodes-external diameter 150 pm-were used for stimulation
in the hypothalamus. These were made by twisting two Teflon-insulated wires (Med- wire PtIr 3T) over a length of 15 mm. Only the cross section at the tip was bare. Indifferent electrodes were made of 70-pm-diameter diamel-insulated Pt wire (John- son Ltd., England) which were bared for about 2 mm at the tips.
The electrodes used for lesioning the central gray were monopolar epoxyresin- coated PtlO%Ir wires with a diameter of 0.25 mm (Plastic Products). The tips were bared ranging from 0.5 to 1 mm in order to vary the size of the lesion. All electrodes were carefully checked for leakage before implantation.
Surgery and Lesioning Thirty-nine male CPBWE-zob rats weighing between 300 and 500 gm were oper-
ated by essentially the same procedure as described by Kruk et a1 [ 19791. Hypotha- lamic electrodes were implanted at the coordinates 7.2 mm A.P., 4.5 mm M.L., and - 18.25 mm D.V. under an angle of 10" to the midsagittal plane of the rat brain. This roughly corresponds with the 5.5 mm A.P., 1.5 mm M.L., and -2.5 mm D.V. coordinates in the stereotaxic atlas of Konig and Klippel [ 19631.
An indifferent electrode was implanted to the right of the bregma, under the skull, just touching the cerebral fluid. The electrodes were soldered to a miniature Am- phenol plug [see Kruk et al, 19781, which was fixed to the skull by screws and dental cement. Unlike in previous descriptions, a piece of india rubber was embedded in the dental cement just underneath of the connector on the skull; its purpose is described later.
After the initial threshold determinations for hypothalamically induced behaviours and tests for territorial aggression (eight weeks), the rats were operated on for a

136 Mos et a1
second time. The skull was fixed in a David Kopf stereotaxic apparatus and a thin layer of cement just behind the connector was removed, exposing the india rubber. Since this is elastic, it could be easily whipped out of the surrounding dental cement, thereby exposing the skull. Holes could then be drilled, after which the lesion electrodes were implanted bilaterally under an angle of 10" to the midsagittal plane. The coordinates of the lesion electrodes ranged from 3.90 to 4.50 mm A.P., 3.05 mm M.L., and -15.55 mm D.V. This roughly corresponds with the 2.5-2.8 mm A.P., 0.4 mm M.L., and -0.8 mm D.V. coordinates in the atlas of Konig and Klippel [1963]. The electrodes were fixed by a cyanolit glue to the skull and the cavity was filled with dental cement. After the cement had hardened, the electrodes were connected to a Grass LM4 RF lesion maker. We took advantage of the fact that the central gray is a medially located structure. Lesions were made by passing a current between the two electrodes and not as is usually done, between one electrode and an indifferent. The most intense heat production will therefore be generated in the region between the two electrodes. The lesion electrodes were placed at the lateral borders of the central gray. This procedure resulted in very localized, smooth-edged lesions. The parameters of the lesion procedure were as follows: 100 kHz frequency, 20 seconds duration, 40-80 V, with a current of approximately 20 mamp. During the 20 seconds the voltage was increased until a sudden drop in the current appeared. This is probably due to the hydrolysis of water. Then the voltage was adjusted to return to the original current flow of about 20 mamp.
Control animals underwent the same surgical treatment, including implantation of electrodes, but without the actual lesioning by the RF lesion generator. Some animals were given an intragastric injection of 10 ml glucose after the operation because they failed to drink. Both after the first and the second operation the rats were allowed to recover for a week.
Test Cages For the stimulation-induced aggression tests we used a cage with 60-cm-high walls
on three sides, a 50 X 100 cm floor area covered with sawdust, and a 60 X 100 cm glass observation window. This cage was also used for locomotion tests but in that case had a feltlike carpet on the bottom instead of sawdust. By means of black lines drawn on this carpet, the cage was divided into eight squares. Switch-off tests were performed in a smaller circular Plexiglass cage of 40-cm diameter, which was placed in the larger test cage. In the wall of this 35-cm-high cage a lever was fixed at 3.5 cm above the floor. This lever was connected to a switching unit on the top of the large cage.
Animals used in the territorial aggression experiments were permanently housed in large cages of 100 X 50 X 75 cm. These cages were sound-insulated and furnished with constant ventilation. An inverted day and night scheme as described above was established, while food and water were given freely. A glass window in the front of the cage permitted observation. A movable partition was used to introduce an intruder at the beginning of the test sessions.
Stimulation Techniques
In all stimulation experiments the rats were stimulated with 40-Hz biphasic square- wave pulses with a phase duration of 0.2 msec and a phase interval of 12.5 msec. Pulses were delivered by two Grass PSIU6 isolated constant current sources con- nected to a Grass S88 stimulator or a modular Ortec stimulation equipment. The

Central Gray Lesions and Aggression in Rats 137
current, measured as the voltage drop across a 1,OOO-Q resistor in series with the electrode, and the voltage were displayed on a Tektronix 5103N oscilloscope. Dura- tion and current intensity were always carefully equalized to minimize net charge flow. By adjustable timers the current was switched to a dummy resistor or to the rat. During the initial testing trials the pulse trains lasted two minutes alternated by an intertrain interval of one minute. During the routine current threshold determinations a 10-second pulse train was alternated with a 50-second intertrain interval. A slight modification was used for switch-off behaviour (see description Behavioural tests).
Behavioural Tests Stimulation-induced aggression. Experimental animals were placed in the test
cage with a naive partner. Testing usually started at current intensities of 50 to 100 pamp. Current was raised in steps of 50-100 pamp, depending upon the observed behaviour, until a clear aggressive response was obtained. At this current intensity the animal was stimulated several times before an attempt was made to determine a current threshold. Testing was discontinued when an upper limit of 500 pamp was reached or when motor responses precluded further testing.
Current thresholds were determined according to the up-and-down method of Dixon and Mood [1948] as modified by Wetherill [1966]. When a trial yielded an attack within ten seconds, the current was decreased with fixed steps, until no attack occurred in the ten-second stimulation period (=one response change). Then the current was raised with the same fixed step size until the animal attacked again (=second response change). The current was varied in this manner until six response changes had been obtained. From these six response changes a mean current intensity was estimated, which will theoretically induce attacks in 50% of the trials [see Kruk et al, 1979; Mos et al, 19821. This current intensity will henceforth be referred to as the aggression threshold. The criterion in these experiments is at least one attack in ten seconds. The intensity of the attacks was scored as follows: a, attack jump; b, bite and jump; c, severe bite; d, gentle bite; e, skin pulling; f, clinch flight. For a more elaborate description of the behaviour see Kruk et a1 [ 19791.
Territorial aggression. Animals with two positive electrodes for aggression in the hypothalamus were selected and housed in special cages. Before the actual aggression tests, the cage was divided into two parts by a movable partition. A naive albino Wistar rat weighing between 160 and 200 gm was placed in the empty compartment. After five minutes adaptation, the partition was raised under remote control to prevent disturbance by the investigator. For 15 minutes the behaviour of both the resident and the intruder were recorded on Sony V62 high-density videotape. A Sony DXC2000A videocamera, which was particularly sensitive to red light and a Sony AV-3670 ACE videorecorder were used. Two 15-W red darkroom bulbs provided the required light.
The videotapes were later analyzed and a protocol of the behaviour was scored on punchtape which was fed into the computer. With the aid of especially developed computer programs the data were reduced to frequencies, duration, mean bout length, and latencies of the observed behavioural elements. The aggression in the first three tests was only observed but not recorded on videotape, because during the first few tests the aggressive behaviour of the resident male is usually lower than during subsequent tests. Prior to the lesion two tests were recorded with a week’s interval. On these days no other tests were performed. One, two, and four weeks after the lesion the aggression tests were recorded again.

138 Mos et a1
Locomotion. A number of the electrodes that supported aggression were also tested for stimulation-induced locomotion. The rats were placed in the test cage with the feltlike carpet five minutes before the actual threshold determination. The loco- motor activity due to exploration, which rats normally show in new or seminew environments, is then greatly reduced (unpublished observations in this laboratory). Stimulation for the first threshold started at the current thresholds for aggression. The number of squares traversed was counted. The rat had to cross a line between two squares with all four paws before a count was scored. The criterion for threshold determinations was six squares crossed in ten seconds. Previous unpublished investi- gations had shown that this criterion was never reached during spontaneous locomotor activity. Subsequent threshold determinations were started at the current value of the previous threshold. The threshold procedure was otherwise the same as employed for aggression. The mean prelesion threshold for locomotion was approximately 33 % higher than the mean value for aggression. The threshold values for locomotion and aggression correlated significantly (P < .02).
Switch-off. Many aggressive electrodes also support switch-on (unpublished data) and/or switch-off behaviour after a short period of training. In the present study switch-off was tested in two ways. Using a leverpress test it was determined whether switch-off could be obtained at current threshold values for aggression. This proce- dure was as follows: Animals were placed in the circular cage and stimulated continuously for five minutes at the current thresholds for aggression, with otherwise identical stimulation parameters. Pressing the lever interrupted the stimulation for five seconds. Pressing the lever again in this period had no effect. The animal received five to ten trials of five minutes at this current intensity. If after some training the rat responded with approximately 30 presses in the five-minute period, it was considered positive for switch-off. For such electrodes current thresholds for switch-off were determined as a second procedure. In these threshold determinations a ten-second stimulation train was alternated with a 50-second period without stimulation. During the stimulation pressing the lever interrupted the current for one second. The thresh- old procedure was otherwise identical to that for aggression. The criterion for switch- off was at least one press during the stimulation period. This threshold was always determined directly after the five-minute presses test. Thus, the switch-off test consisted of a five-minute leverpress test, followed by a threshold determination. Mean switch-off thresholds were 60% lower than the aggression thresholds. The threshold values for aggression and switch-off showed significant correlation (P < 0.002).
Histological Procedures Upon completion of the experiments the rats yere deeply anaesthetized with ether
and perfused intracardially with saline followed by a 4 % formaldehyde/saline solu- tion. After storage in formaldehyde for a week, the brains were removed from the skull and again stored in formaldehyde for another week. Then the brain was frozen and cut on a freezing microtome at 20 pm. All sections with the lesion in the central gray and the sections with the electrode track in the hypothalamus were stained according to the Kliiwer-Barrera method.
The anterior-posterior coordinates of the electrodes in the hypothalamus were assessed by comparing the relevant sections with the standard sections of our own unpublished atlas. In contrast to commercially available atlases, sections in this atlas

Central Gray Lesions and Aggression in Rats 139
were made at 75-pm intervals, which allowed a more accurate localization of the electrode tips.
The lesions in the central gray were indicated on drawings of sections of our own atlas. We used planes spaced by 375-pm intervals. The lesions on these drawings were measured by means of a Tektronix X-Y tablet, connected to a PDP 11/60 computer. The drawings were laid on this tablet and the circumference of the lesion was followed with a metal pencil. Computer programs calculated the surface of the lesion. Since on these drawings the central gray was indicated, a distinction could be made between the part of the lesion inside the central gray and that part of the lesion which was outside the central gray (henceforth referred to as “external lesion”). Within the central gray we had previously [Mos et al, 19821 delimited an area where degenerating axons were found following destruction of hypothalamic attack sites. The overlap of the lesion with this degeneration area-the “degeneration lesion”- could therefore be measured too. Hence the effects of the lesions could be correlated to four measures of the lesion-namely, the total lesion, the external lesion, the central gray lesion, and the degeneration lesion.
Statistical Procedures
Analysis of variance for randomized block designs was performed with the thresh- old values for aggression, locomotion, and switch-off tests as well as with the frequency of switch-off leverpresses. The same analysis was performed with the data of spontaneous aggression, social interactions, defense, and self-grooming . Both the experimental and the control group were treated in this way. When such an analysis revealed significance, a Student’s t-test or a column contrast test according to Scheffk [1959] were performed to test for significance between the columns. Spearman rank correlations between the effect of the lesion and its size were calculated. The proce- dure for the Spearman rank test was as described in Siege1 [1956]. A chi square test was performed to test for differences in intensity of attack before and after the lesion. The procedure of the adjusted residuals as described by Everitt [1977] was used to see which cells differed from a random distribution.
EXPERIMENT 1 Introduction
From a degeneration study [Mos et al., 19821 it is known that the central gray is a major projection area for neurons in hypothalamic attack sites. A lesion in the central gray therefore destroys an important output pathway from hypothalamic sites. Ac- cording to Hunsperger [ 19561 and Skultety [ 19631, large lesions should abolish hypothalamic aggression. In some pilot experiments we placed small unilateral le- sions, which appeared to be ineffective in reducing hypothalamic aggression.
The aim of this study was to place large bilateral lesions in the central gray and to study its effect on aggression. Lesion studies often deal with the problem of how to be certain that the observed effect is a specific one. Many mechanisms can be proposed as to how a lesion could exert its effect. To evaluate whether lesioning specifically affects aggression, the effects on hypothalamically induced switch-off behaviour and locomotion were also studied. Stimulation by electrodes in the hypo- thalamus that induce aggression very often results in increased locomotor activity. Neurophysiological studies reveal that locomotion and aggression probably have a

140 Mos eta1
different anatomical substrate, because the excitability differs [Kruk and van der Poel, 19801. It thus seems that different neural substrates mingle at the site of the electrode. It has been demonstrated for different forms of aggression in the cat by Berntson [ 19721 that at more caudal levels neural pathways may be separate.
If the projection to the central gray is indeed specific for aggression, then locomo- tion and switch-off should remain unaffected following central gray lesions. Or at least the performance of all three behaviours should not deteriorate to the same extent after the lesion.
Another problem with lesion studies is that most often the lesion is not confined to the brain structure in target. Therefore one should be able to differentiate between the lesion of the intended structure and the lesion outside this structure. This can be done by measuring the size of the lesion and correlating it with the effect such a lesion produces. If a certain region is functionally homogeneous, then the more tissue is destroyed, the larger the effect should be. If a region has nothing to do with the behaviour in study, no correlation should be found.
In the following experiment the effect of central gray lesions on aggression as well as on other hypothalamically induced behaviours was studied. Moreover the lesion was subdivided into several regions and the extent of destruction to that region was correlated with the effect of the lesion.
RESULTS Histology. A survey of the localization of the hypothalamic electrodes is given in
Figure 1. The anterior-posterior, the dorsoventral, and the mediolateral coordinates were each correlated with the change in aggression threshold. No significant correla- tion was observed with any of these parameters. Hence it can be concluded that it is impossible to delimit hypothalamic areas that are particularly sensitive to lesions of the central gray. It seems that the population of hypothalamic attack sites is homoge- neous in this respect.
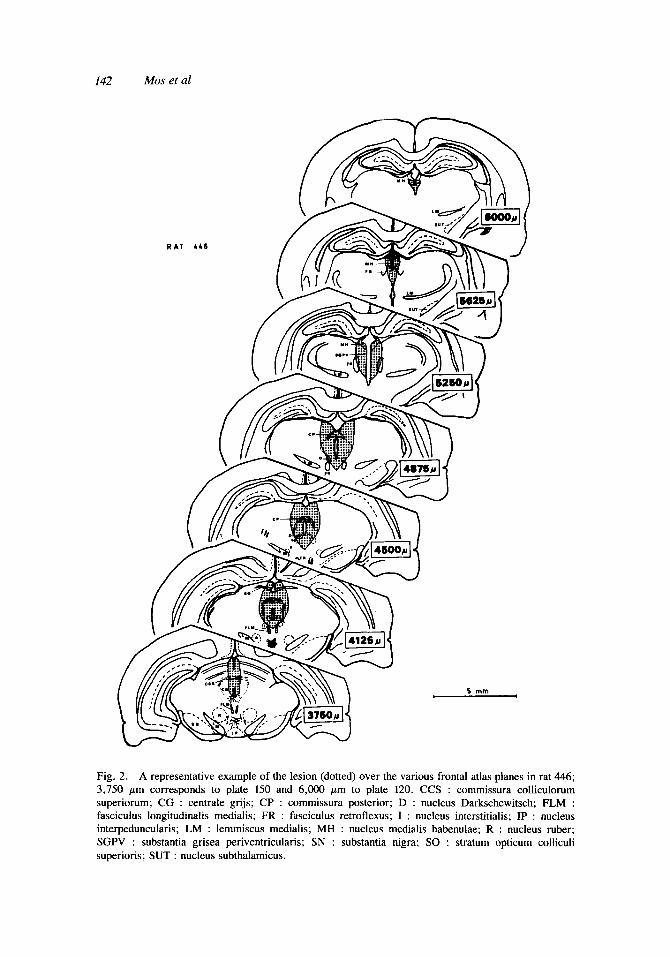
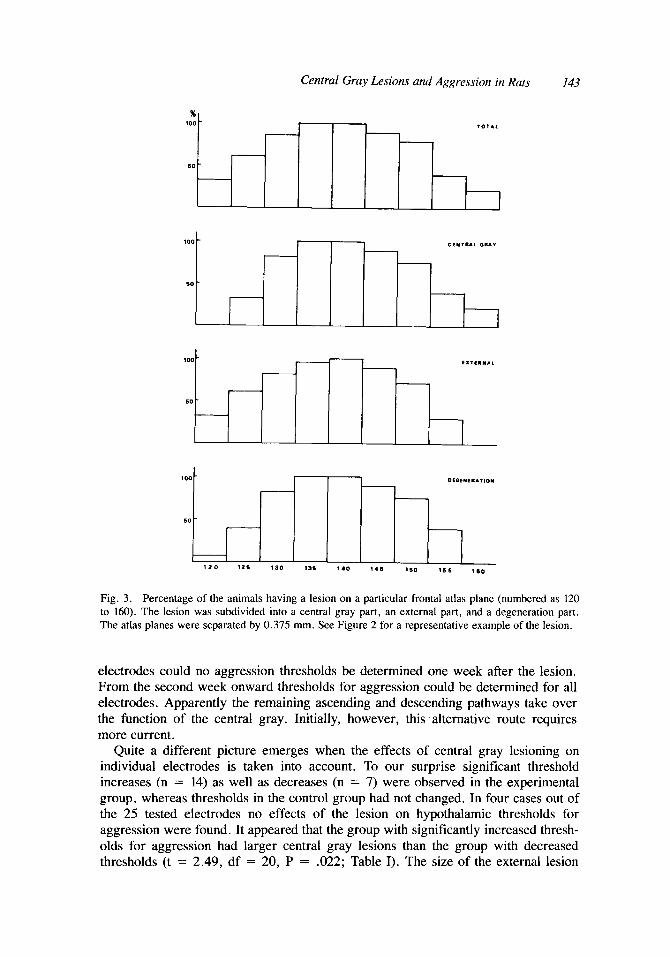
An example of a representative central gray lesion is seen in Figure 2. The size of the lesions varied from 5.3 to 18.7 mm3. The distribution of the lesions over the various planes is given in Figure 3. The lesions all comprised planes 135 and 140, though the degree of damage could differ considerably. This also holds for the more specific classification of the lesion into its various parts. The most rostra1 lesion ran from plane AP 3.7 to 6.5 mm, the most caudal one from AP 2.0 to 4.3 mm.
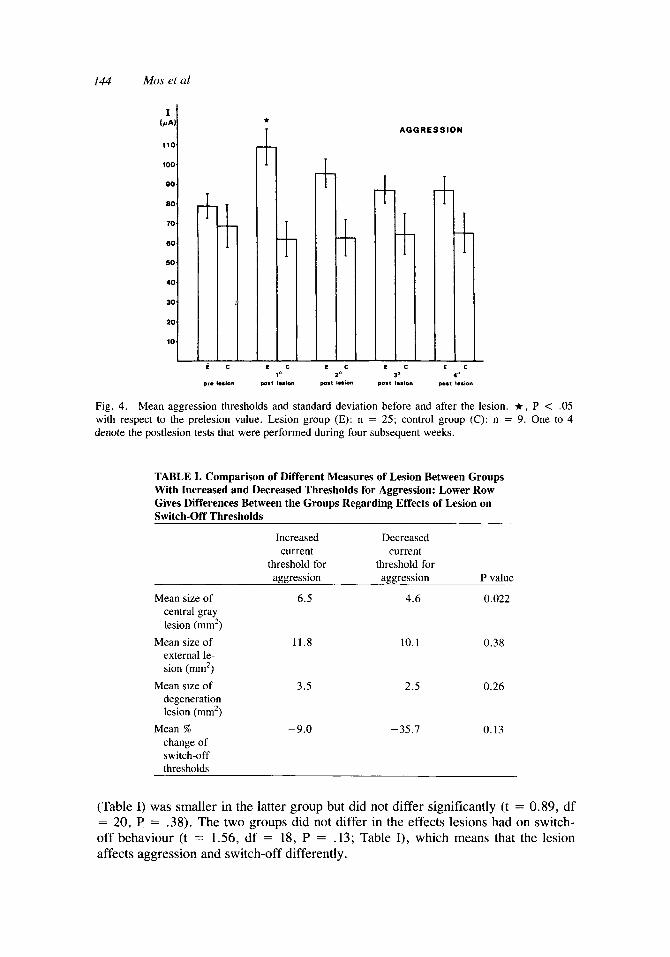
Aggression. In 39 rats electrodes were implanted bilaterally into the hypothala- mus. Ultimately 34 electrodes inducing aggression in 24 animals remained. Two rats died during the experiment and a number of electrodes were discarded because of the gradual appearance of convulsions during the threshold determinations. Twenty-five electrodes in 18 animals belonged to the experimental group; nine electrodes in six animals served as control group. When two aggressive electrodes were in the same animal, they were both independently tested during the entire experiment. The lesions to the central gray were always divided into a left part and a right part. Prior to the lesion at least six current thresholds for aggression were determined. The mean value of the last three thresholds served as prelesion value. Starting one week after the lesion, thresholds were determined during four weeks, one threshold each week. Analysis of variance was performed separately on the results of both the control group and the experimental group. This was not significant for the control group (P = .085) but closely approximated the 5 % level. As can be seen in Figure 4, the

Central Gray Lesions and Aggression in Ruts 141
8 3 2 5 ~ 8 1 0 0 ~
7 9 5 0 ~
( 7800p
7575p 7 3 5 0 , ~
7125p 6900p
Fig. I . Survey of the localization of the hypothalamic electrode placements on various planes of our own atlas. Anterior-posterior coordinates are given in pm. t , aggressive; 0 , nonaggressive; a, aggressive, but no thresholds could be determined.
thresholds tend to be slightly lower after the sham-lesion, but in any case there is no increase in thresholds. The analysis of variance for the experimental group revealed significance (P < .OOl). The Scheffk test for column contrasts showed that only the first postlesion threshold was significantly increased with respect to the prelesion value. The second, third, and fourth thresholds were still increased but no longer significant (P = .07, .69, .69, respectively). Thus the original expectation, derived from the literature, that large lesions abolish aggression [Skultety, 1963; Hunsperger, 19561, could not be confirmed.
At least at one level of the rostrocaudal extent of the central gray, the degeneration bundle, had been completely destroyed in all but three rats, thus presumably blocking a major output pathway of hypothalamic attack sites. Nevertheless, only for two
142 Mos eta1
R A T L L 6
5 m m
Fig. 2. A representative example of the lesion (dotted) over the various frontal atlas planes in rat 446; 3,750 pm corresponds to plate 150 and 6,000 pn to plate 120. CCS : commissura colliculorum superiorum; CG : centrale grijs; CP : commissura posterior; D : nucleus Darkschewitsch; FLM : fasciculus longitudinalis medialis; FR : fasciculus retroflexus; I : nucleus interstitialis; IP : nucleus interpeduncularis; LM : lemmiscus medialis; MH : nucleus medialis habenulae; R : nucleus ruber; SGPV : substantia grisea periventricularis; SN : substantia nigra; SO : stratum opticum colliculi superioris; SUT : nucleus subthalamicus.
Central Gray Lesions and Aggression in Rats 143
x 100
50
l 0 T l l -
-
I
100
50
Fig. 3 . Percentage of the animals having a lesion on a particular frontal atlas plane (numbered as 120 to 160). The lesion was subdivided into a central gray part, an external part, and a degeneration part. The atlas planes were separated by 0.375 mm. See Figure 2 for a representative example of the lesion.
- DEOENf lA110N -
- - - -
electrodes could no aggression thresholds be determined one week after the lesion. From the second week onward thresholds for aggression could be determined for all electrodes. Apparently the remaining ascending and descending pathways take over the function of the central gray. Initially, however, this alternative route requires more current.
Quite a different picture emerges when the effects of central gray lesioning on individual electrodes is taken into account. To our surprise significant threshold increases (n = 14) as well as decreases (n = 7) were observed in the experimental group, whereas thresholds in the control group had not changed. In four cases out of the 25 tested electrodes no effects of the lesion on hypothalamic thresholds for aggression were found. It appeared that the group with significantly increased thresh- olds for aggression had larger central gray lesions than the group with decreased thresholds (t = 2.49, df = 20, P = .022; Table I). The size of the external lesion
144 Mos eta1
110
50
40
30
20
10 k E C i E C
AGGRESSION
E C E C E C 1 0 30 b'
pr. I d o n p0.1 I.*(M ~0.t Id- ~a.1 I.sion p0.1 I . S ~
Fig. 4. Mean aggression thresholds and standard deviation before and after the lesion. It, P < .05 with respect to the prelesion value. Lesion group (E): n = 25; control group (C): n = 9. One to 4 denote the postlesion tests that were performed during four subsequent weeks.
TABLE I. Comparison of Different Measures of Lesion Between Groups With Increased and Decreased Thresholds for Aggression: Lower Row Gives Differences Between the Groups Regarding Effects of Lesion on Switch-Off Thresholds
Increased Decreased current current
threshold for threshold for aggression aggression P value
Mean size of 6.5 4.6 0.022 central gray lesion (rnrn')
external le- sion (rnm2)
degeneration lesion (rnm2)
change of switch-off thresholds
Mean size of 11.8 10.1 0.38
Mean size of 3.5 2 .5 0.26
Mean % -9.0 -35.7 0.13
(Table I) was smaller in the latter group but did not differ significantly (t = 0.89, df = 20, P = .38). The two groups did not differ in the effects lesions had on switch- off behaviour (t = 1.56, df = 18, P = .13; Table I), which means that the lesion affects aggression and switch-off differently.

Central Gray Lesions and Aggression in Rats 145
%
lo0
60
1
soL 125 1%
C G LES,O*
i m -
IS 140 14s 1W 156 100
T K
100
SO I 110 12s 13E i 36 140
L I 85 160 1ss
Fig. 5 . Percentage of animals having a lesion on the planes 120 to 160. The upper bar diagram is the distribution for the animals with increased thresholds for aggression, the lower diagram for the animals with decreased thresholds for aggression. A chi square analysis revealed no difference between the groups in the distribution over the various atlas planes.
The distribution of the lesions over the frontal planes was also compared (see Fig. 5). No systematic change was found in the distribution, as was demonstrated by a chi square test (central gray lesion: x2 = 12.3, df = 8, P = .14; external lesion: x2 = 13.9, df = 8, P = .08). It thus seems that the amount of destruction to the central gray rather than the place of the lesions is the determining factor.
The groups with increased and decreased thresholds were also compared for the effects of the lesions on the intensity of the attack (Table 11). No change in intensity of attacks was found in the group with decreased thresholds after the lesion (x2 = 4.7, df = 3, P = .19). In the group of electrodes with increased thresholds the attacks were less intense after the lesion, ie, fewer attack jumps and more gentle bites were observed (x2 = 25.8, df = 4, P < .OOOl). The same was found in the group without change in the thresholds for aggression (x2 = 16.4, df = 4, P C .005).
In the group with increased thresholds a further subdivision could be made. The increase was temporary in four electrodes, whereas for ten electrodes the lesion effects lasted. The former group had smaller external lesions, but probably due to the
146 Mos eta1
TABLE 11. Intensity of the Attacks Before and After the Lesion*
a b C d e
Increased current Prelesion 5 ( + ) 40( + 107(-) 29 0 threshold after Postlesion 2(-) 37(-) 218(+) 99 4
No change current Prelesion 3 13(+) 74(-) 18 0 threshold after Postlesion 14 22(-) 106( +) 39 0 lesion
Decreased current Prelesion 0 19 25 7 0 threshold after Postlesion 4 9 60 11 1 lesion
lesion
*Chi square table of the pre- and postlesion intensities for aggression. The total group of 25 electrodes was divided into three subgroups (see text). The intensity of the attacks decreases from a to e. The marks (+) or (-) denote in which cell of the matrix the frequency is higher (+) or lower (-) than expected.
SWITCH-OFF : THRESHOLDS
SWITCH-OFF : PRESSES
50 "1 a
20
10
E C E C L C E C E C 1' 20 ,a *o
P,. I.., o" pa., I..,O" m0.t I..IO" PO.! I..IO* PO., I..IO"
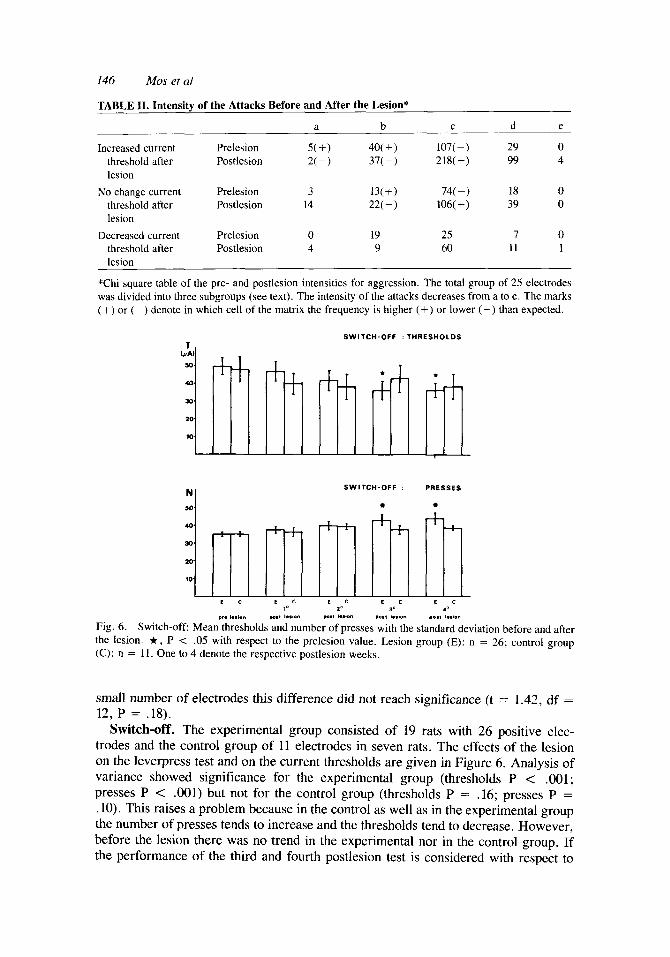
Fig. 6. Switch-off Mean thresholds and number of presses with the standard deviation before and after the lesion. t , P < .05 with respect to the prelesion value. Lesion group (E): n = 26; control group (C): n = 11. One to 4 denote the respective postlesion weeks.
small number of electrodes this difference did not reach significance (t = 1.42, df = 12, P = .18).
Switch-off. The experimental group consisted of 19 rats with 26 positive elec- trodes and the control group of 11 electrodes in seven rats. The effects of the lesion on the leverpress test and on the current thresholds are given in Figure 6. Analysis of variance showed significance for the experimental group (thresholds P < .001; presses P < .OOl) but not for the control group (thresholds P = .16; presses P = .lo). This raises a problem because in the control as well as in the experimental group the number of presses tends to increase and the thresholds tend to decrease, However, before the lesion there was no trend in the experimental nor in the control group. If the performance of the third and fourth postlesion test is considered with respect to
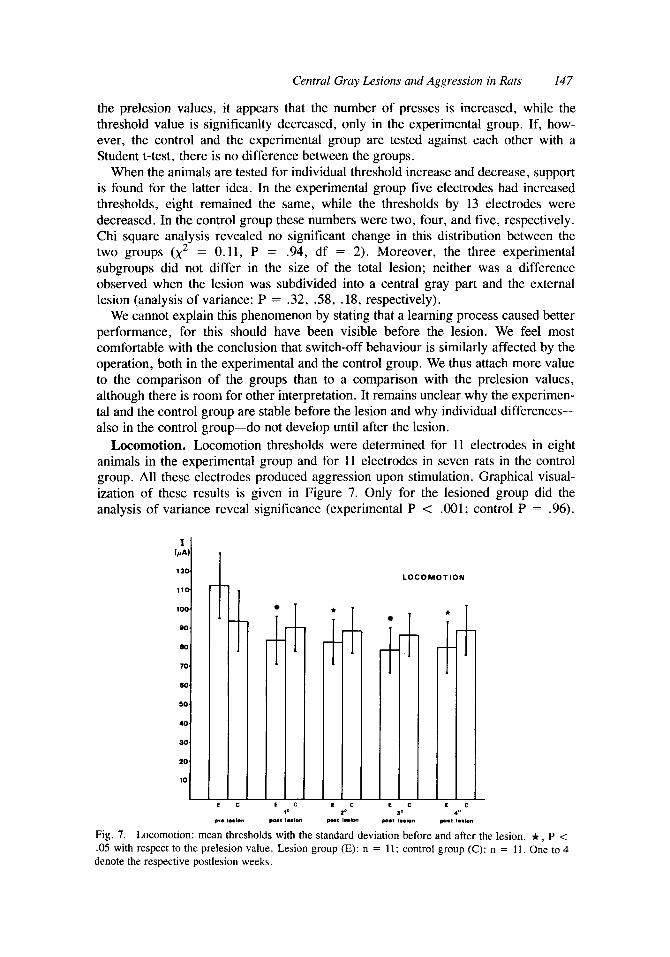
Central Gray Lesions and Aggression in Rats 147
the prelesion values, it appears that the number of presses is increased, while the threshold value is significanlty decreased, only in the experimental group. If, how- ever, the control and the experimental group are tested against each other with a Student t-test, there is no difference between the groups.
When the animals are tested for individual threshold increase and decrease, support is found for the latter idea. In the experimental group five electrodes had increased thresholds, eight remained the same, while the thresholds by 13 electrodes were decreased. In the control group these numbers were two, four, and five, respectively. Chi square analysis revealed no significant change in this distribution between the two groups (x2 = 0.11, P = .94, df = 2). Moreover, the three experimental subgroups did not differ in the size of the total lesion; neither was a difference observed when the lesion was subdivided into a central gray part and the external lesion (analysis of variance: P = .32, .58, .18, respectively).
We cannot explain this phenomenon by stating that a learning process caused better performance, for this should have been visible before the lesion. We feel most comfortable with the conclusion that switch-off behaviour is similarly affected by the operation, both in the experimental and the control group. We thus attach more value to the comparison of the groups than to a comparison with the prelesion values, although there is room for other interpretation. It remains unclear why the experimen- tal and the control group are stable before the lesion and why individual differences- also in the control group-do not develop until after the lesion.
Locomotion. Locomotion thresholds were determined for 11 electrodes in eight animals in the experimental group and for 11 electrodes in seven rats in the control group. All these electrodes produced aggression upon stimulation. Graphical visual- ization of these results is given in Figure 7. Only for the lesioned group did the analysis of variance reveal significance (experimental P < .001; control P = .96).
60
50
40
20
10 i i - E
10 E C
2'
LOCOMOTION
1 p,. I..iO" PO., 1..,on Po., I"mn pO.1 *.LO" pO.1 1..,on
Fig. 7. Locomotion: mean thresholds with the standard deviation before and after the lesion. * , P < .05 with respect to the prelesion value. Lesion group (E): n = 11; control group (C): n = 11, One to 4 denote the respective postlesion weeks.

148 Mosetal
Scheffe column contrasts were significant for all four postlesion thresholds as com- pared to the prelesion values in the experimental group. When individual threshold changes are considered, the conclusion remains the same. In the experimental group all but one electrode exhibited a decrease in thresholds after the lesion. In the control group four electrodes had decreased thresholds, four remained the same, whereas in three electrodes a threshold increase was observed.
Locomotion thresholds are decreased in lesioned animals, meaning that it is easier to activate the animal and/or that the spontaneous locomotion is increased. This could mean that an inhibition mechanism in the central gray has been destroyed. If so, most lesions will have destroyed it, because there is no correlation between the size of the various parts of the lesions and the change in locomotion (P = .63). Neither is the effect mediated by decrease of overall performance, because if the animal is inactive after the lesion, this would result in higher locomotion thresholds. The contrary appears to be true.
Discussion Despite the heterogeneous response following central gray lesions, it is clear that
complete destruction of the hypothalamic projection to the central gray does not completely inhibit hypothalamic aggression permanently. All the same, large lesions of the central gray result in an increase of the current required to induce aggression by hypothalamic stimulation. The lesion to structures outside the central gray is probably not responsible for the effects on aggression. There is no difference between the groups with increased and decreased thresholds, regarding the extent of this lesion, nor in its distribution over the various frontal planes.
Taking the line that we induce aggression of an offensive type, it is difficult to assess the role of the central gray. In any case it is not the main descending relay station for offense, because in most animals-in at least one frontal plane-the projection from the hypothalamus to the central gray had been effectively destroyed. The remaining output pathways from hypothalamic attack sites are apparently suffi- cient to propagate nervous activity needed for attacks. In contrast defensive aggression is completely and permanently abolished following extensive lesions of the central gray and adjoining tissue [Edwards and Adams, 19741. This suggests that the role of the central gray for offensive attacks is much less important than for defensive attacks. Although the central gray is not required for offense, its proper functioning definitely affects tne current thresholds for hypothalamic attacks.
The most puzzling thing to explain is why some lesions facilitate and others inhibit aggression. A similar phenomenon has also been reported by Schmitt et a1 [1979], who observed differential effects of central gray lesions upon hypothalamically in- duced escape reactions. The central gray is known to have reciprocal connections to the hypothalamus. Perhaps the smaller lesions result in hyperactivity of the remaining part of this feedback system, whereas the larger lesions may destroy feedback from the central gray completely. Following an idea of Adams [1979] regarding different neural substrates for offense, defense, and submission, it is possible that the lesions destroy different neural pathways. However, this idea cannot be verified by the present material.
Finally it should be mentioned that Edwards and Adams [I9741 also observed that small central gray lesions resulted in a decrease of the current threshold in electro- shock-induced fighting. They stressed that only large lesions were effective in block-

Central Gray Lesions and Aggression in Rats 149
ing the defensive response but did not suggest a mechanism which might explain the effects of smaller lesions.
Some of the thresholds were only temporarily increased after the lesion. Edwards and Adams [1974] suggested that the adjacent tegmentum might take over functions formerly performed by the central gray. The partial recovery is then dependent upon the sparing of the tegmentum. There is some evidence in their data that supports this view. Since large lesions of the central gray are likely to invade a substantial part of the tegmentum, the finding of Skultety that large central gray lesions result in complete blockade of hypothalamic rage could be attributed to the fact that his large lesions damaged the tegmentum considerably. We find a significant correlation (P = .005) between the amount of destruction of the central gray and of the surrounding tegmentum and superior colliculus. We compared the electrodes having lasting lesion effects with those where the lesion effects were transient. In the latter group (n = 4) the external lesions were smaller, but unfortunately this difference did not reach significance.
The effects of the lesion seem to be specific in the sense that they do not impair every behavioural test. If there is a lesion effect on switch-off at all (see discussion in the paragraph Switch-Off of the Results section), the time course of these changes differs from the effects of the lesion on aggression. The effects on locomotion demonstrate that the lesions do not result in a general deterioration of the animal’s condition. In that case a threshold increase should be expected, but this does not happen.
The lack of any correlation between the lesion size and the effect on locomotion cannot be explained by us. Edwards and Adams [1974] have found that there is only a small region of common overlap between the lesions in animals having increased locomotion [Edwards and Adams, 1974: Fig. 5D]. There was one frontal atlas plane in which all animals had a lesion. This indicates that the central gray is not homoge- neous with respect to locomotion. Homogeneous means that each part of the tissue is of equal importance for the behaviour under study. The more tissue is destroyed, the larger the effect will be. Homogeneity is a necessary condition for meaningful interpretation of the correlations between the size of the brain lesions and the behavioural effect.
Furthermore it deserves attention that superior colliculus lesions may result in an increased locomotion, too [Kallman and Isaac, 19801. In our animals very often parts of the superior colliculus are also damaged. The results of Edwards and Adams [ 19741 and Kallmann and Isaac [1980] deal with spontaneous locomotion. This may also be relevant here. Our threshold determinations most likely are sensitive for increases in spontaneous locomotion. The criterion in the threshold determinations is chosen in such a way that the animal never reaches it due to spontaneous locomotion in unlesioned animals.
EXPERIMENT 2 Introduction
Electrical stimulation of the central gray may induce aggressive behaviour [Mos et al, 19821. No doubt electrical brain stimulation studies can be criticized by drawing attention to the highly unnatural intervention that is produced. Electrical stimulation will activate many cell bodies and axons of passage that lie under the electrode tip,

150 Mos et a1
and activation will not be confined to one type of fibre or neuron. Perhaps this lack of selectivity can also explain why the morphology of the induced behaviour does not always match naturally occurring behaviour [Bandler and Tork, 198 11. Consequently the relevance of the induced behaviour in terms of its significance in natural situations has been questioned. In studies of the relationship between brain and behaviour one cannot escape this problem, except perhaps as regards some types of chemostimula- tion, since technical facilities do not permit any more selective intervention of brain processes.
Ideally one should therefore apply many control procedures to establish the role of the electrically stimulated tissue in naturally occuring behaviour. One of these could be the recording of units at sites where upon stimulation a particular behaviour is induced. On the other hand, destruction of these sites should diminish the occurrence of this behaviour in situations where it occurs spontaneously.
In our experiment we could study the effects of central gray lesions on both territorial (exp. 2) and stimulation-induced aggression (exp. 1) in the same ruts. It should be expected that if the neural substrate in the central gray plays an active and selective role in naturally occuring aggression, lesions should reduce territorial aggression, leaving other behavioural parameters largely unaffected. Rats with two aggressive electrodes in the hypothalamus were selected and studied for territorial aggression and hypothalamic aggression. Thus we compared the change in territorial aggression, which is generally viewed as being offensive by nature, and the change in hypothalamic aggression after the lesions in the same animals. The behaviour of the partners was scored to assess the role of the partner in the changed behaviour of the experimental animals.
Results Before the lesion two tests lasting 15 minutes each were recorded on video, with an
interval of one week between the two tests. The mean scores of these two tests served as prelesion values. One week after the lesion, the first postoperative aggression test was recorded. The second and the fourth week after the lesion another test was recorded. The aggression test in the third postoperative week was not recorded on video and is not incorporated in the following treatment of the data. Ten animals belonged to the experimental group and four to the control group.
The observed behaviours were divided into four categories: aggression, defense, social behaviour, and self-grooming . Aggression consisted of hipthrow, pushing, lateral threat, frontal threat, quiet and severe bites, fighting, chasing and keeping down, or full aggressive posture [Bermond, 19781. Elements belonging to defensive behaviour were freezing and scanning. Social behaviour included social grooming, genital investigation, and mounting. Defensive behaviour in the experimental animals and aggression in the test partners hardly ever occurred and could therefore not be treated statistically.
The parameters we employed to describe the behaviour of the rats were frequency, duration, percentage of time spent on a particular behaviour, and mean bout length. Analysis of variance was performed separately with the data for frequency, duration, percentage of time spent, and mean bout length for the different groups. Since the prelesion values may differ between the experimental and the control group (see Fig. 8), tests for significant changes after the lesion were always performed within either the experimental or the control group. Thus the postlesion and postsham performance
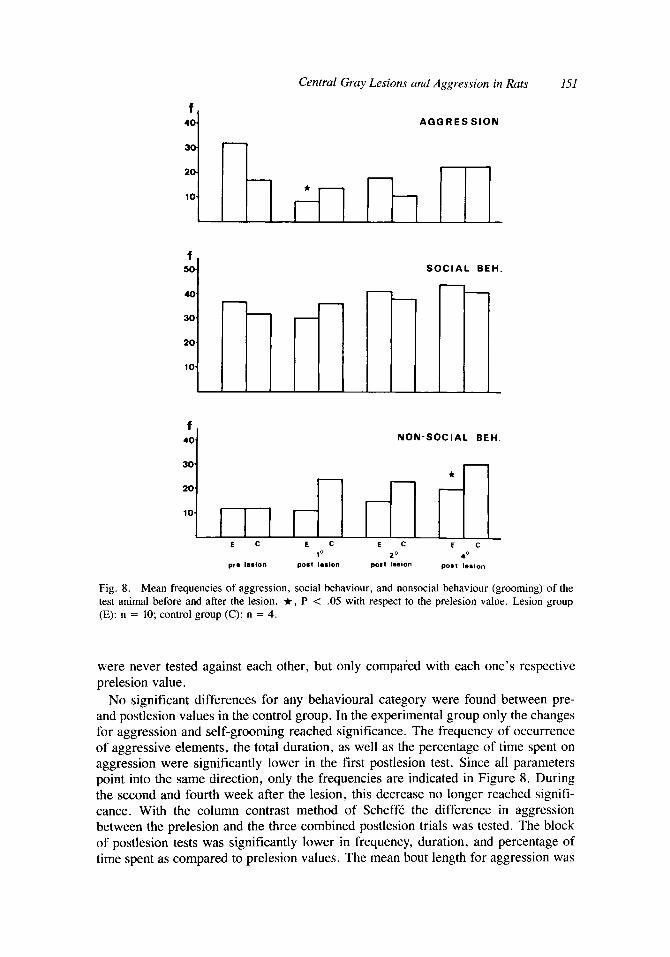
Central Gray Lesions and Aggression in Ruts 151
* - - - - 10.
A Q G R E S S lON
-
20
10 h f
4 0
3 0
20.
S O C I A L BEH.
N O N - S O C I A L BEH.
10 20 4 O
pro lesion post lesion post I e h n post lesion
Fig. 8. Mean frequencies of aggression, social behaviour, and nonsocial behaviour (grooming) of the test animal before and after the lesion. It, P < .05 with respect to the prelesion value. Lesion group (E): n = 10; control group (C): n = 4.
were never tested against each other, but only compared with each one’s respective prelesion value.
No significant differences for any behavioural category were found between pre- and postlesion values in the control group. In the experimental group only the changes for aggression and self-grooming reached significance. The frequency of occurrence of aggressive elements, the total duration, as well as the percentage of time spent on aggression were significantly lower in the first postlesion test. Since all parameters point into the same direction, only the frequencies are indicated in Figure 8. During the second and fourth week after the lesion, this decrease no longer reached signifi- cance. With the column contrast method of Scheff6 the difference in aggression between the prelesion and the three combined postlesion trials was tested. The block of postlesion tests was significantly lower in frequency, duration, and percentage of time spent as compared to prelesion values. The mean bout length for aggression was

1.52 Mos eta1
not significantly altered by the lesion. In parallel, the mean current threshold for hypothalamic aggression in these animals was increased by 35 % .
Self-grooming frequency was increased in the fourth postlesion test, whereas the mean bout length of self-grooming was significantly higher in the first postlesion test (not shown). The total postlesion block was not changed compared to prelesion values for any of the parameters we used to describe the behaviour. The changes in self- grooming caused by the lesion are therefore not so consistent as is the case for aggression.
Social behaviour was not impaired by the lesion (Fig. 8). Diminished aggression can therefore not be attributed to a loss of social contact. These social contacts usually precede the occurrence of aggressive actions [Lehman and Adams, 19771. It therefore seems that the animal is well capable of starting social interactions, but that the lesions selectively block the tendency of the territory owner to attack the intruder.
Histology. Correlation of the lesion size and the decrease in aggression showed that the extent of the lesion of the degeneration bundle significantly correlated with the mean change in duration and the percentage of time spent on aggression (P = .028, .04 respectively). The drop in frequency of aggressive elements correlated with the lesion of the degeneration bundle only in the first postlesion test (P = .03). No correlation was found between the lesion size and the change in self-grooming (P = .57).
The correlation with the destruction of the hypothalamic projection bundle to the central gray and not with the total central gray lesion is especially noteworthy. It makes it very attractive to attribute an important role to the neural pathway from the hypothalamus to the central gray in spontaneous aggression.
Discussion
Edwards and Adams [ 19741 found that large lesions of the central gray and adjacent tegmentum abolished defensive behaviour of foot-shocked rats. The change in boxing of rats with smaller lesions in the central gray was not significant. On the contrary, the mean threshold current used to induce defensive boxing is lower in these animals. The response to pain did not change in the lesioned animals, while there were deficits in escape performance and freezing. The latter changes did not require the lesion of the total central gray as is the case for boxing deficits. Edwards and Adams [1974] have suggested that the central gray is a nodal point in the neural organization of pain-induced defensive boxing. Similarly, Blanchard et a1 [ 19811 have recently de- scribed central gray lesions as impairing defensive reactions toward painful and nonpainful stimuli.
Our results show that the central gray only plays a minor role in territorial, offensive aggression. Temporary, but nevertheless significant changes in territorial aggression were observed after lesioning the central gray. Like in the experiment on stimulation-induced attacks, the central gray is not vital for offensive attacks, whereas it may be for defense. There is little reason-as in the study of Edwards and Adams- to suspect a general depressant effect of the lesion to be the cause of the observed behavioral changes. As can be seen in Figure 8, there is no drop in social activities and grooming.
In spontaneous territorial aggression the effects of lesions tend to decrease in time. Similar fading of lesion effects has been observed by Blanchard et a1 [ 1977~1. Lesions in the septa1 region decreased the offensive aggression of a-males in a colony toward

Central Gray Lesions and Aggression in Rats 153
intruders. These effects decreased in time, but, as in our experiments, no complete recovery was found. We could not find a correlation between the amount of recovery and the damage to the tegmentum. The phenomenon of recovery of function is still a difficult problem. It would, however, be of great scientific and clinical value to understand the underlying principles. Unfortunately our precise subdivision of the lesion into several parts does not provide an explanation either. Further studies are therefore needed to solve this important question.
A study of Skultety [1958] has revealed that extensive lesions of the central gray have no effect on vegetative functions like gastric motility, insuline tolerance, water metabolism, and temperature regulation. Some cats were hyperactive after the lesion, which is in contrast to the findings of Bailey and Davis [1942], who reported hypokinesia and akinesia in cats and monkeys. Later studies of Lindsley et a1 [1950] and French and Magoun [1952] no more confirmed the results of Bailey and Davis. No hypokinesia is found in our rats; on the contrary, the decreased postlesion locomotion thresholds rather support the idea of hyperactivity [Skultety, 19581. Edwards and Adams [1974] also reported increased ambulation scores in the open field, especially in rats with extensive central gray lesions. It therefore seems unlikely that the reduced spontaneous aggression should be an indirect effect of impaired locomotion.
GENERAL DISCUSSION
In the two experiments evidence was adduced for a-be it minor-role of the central gray in offensive attacks. In brief the evidence is as follows: Large central gray lesions increase thresholds for hypothalamically induced aggression. This cannot be explained by a general depressant effect of the lesion, as is shown in the locomotion and switch-off controls. The size of the external lesions is not correlated with the effects on aggression. Territorial aggression toward intruders is decreased by the lesions, but this effect gets weaker in time. The lesion affects no other behaviours studied. It needs to be stressed that despite the impairments, the animals are well capable of fighting, even with substantial central gray lesions. Hence the central gray is not indispensable.
The parallel which is found between the effects of the lesion on hypothalamic aggression and on territorial aggression is an indication that the neural circuit, formed by the hypothalamus and the central gray, may serve a function in spontaneous aggression. This is further supported by the correlation of the effects on spontaneous aggression with the damage to the place in the central gray where most degeneration is seen after lesioning hypothalamic attacks sites. As has been demonstrated in a previous article on stimulation-induced aggression in the central gray [Mos et al, 19821, this place is the primary target to induce aggression by electrical stimulation. Large lesions of this site also result in an increase of the thresholds for hypothalamic aggression. It therefore seems inviting to attribute a function of this degeneration region to both induced and spontaneous aggression. As far as we know such a simultaneous study of lesion effects on natural and induced aggression has only been accomplished by Berntson [1972]. We think that by the combined use of different methods in the same animal, our knowledge of the relation between brain and behaviour can be truly improved.

154 Mos eta1
Furthermore, we substantiated the need for appropriate control experiments in lesion studies. It appeared that locomotion and switch-off were suited as such controls. Ideally controls should have no relation to the behaviour of particular interest. Switch- off may form an exception. The idea that aversive stimulation may result in aggression has been favoured by some authors [Ulrich, 1966; Moyer, 19681. In our experiments we find no relation between aggression and switch-off. Following the line of thought that switch-off is a measure for aversiveness, even an increase in aggression should be expected. There is a tendency for higher scores in the leverpress test. This should suggest increased aversiveness of the stimulation at threshold current for aggression. However, we found no increased aggression. This lesion experiment therefore gives no reason to endorse the aversiveness hypothesis to “explain” hypothalamically induced aggression.
ACKNOWLEDGMENTS
We want to thank Dr. M. Wijnans and Dr. J. Tijssen for their statistical advice, Prof. Dr. J.S. Ploem and K. van der Broek for the use of the X-Y tablet, and Dr. F. Putters for his computer programs for processing the behavioural data. We thank Prof. Dr. E.M. Cohen and Prof. Dr. E.L. Noach for their critical reading of the manuscript and Miss R.H.F. de Ru for correcting the manuscript.
This study was supported in part by a grant from the foundation for Biological Research (BION), which is subsidized by the Netherlands Organization for the Advancement of Pure Research (ZWO). Portions of this research were submitted by the first author in partial fulfillment of the requirements for a MD degree at the University of Leiden.
REFERENCES
Adams DB (1968): Cells related to fighting behav- iour recorded from midbrain central gray neu- ropil of cat. Science 159:894-896.
Adams DB (1979): Brain mechanisms for offense, defense, and submission. The Behavioral and Brain Sciences 2:201-241.
Bailey P, Davis EW (1942): Effects of lesions of the periaqueductal gray matter in the cat. Pro- ceedings of the Society for Experimental Biol- ogy and Medicine 51:305-306.
Bandler R, Halliday R, Abeyewardene S (1980): Centrally elicited aggressive behaviour in the cat: A changing view. In Girgis M, Kiloh LG (eds): “Limbic Epilepsy and the Dyscontrol Syndrome. ” Elseviers/North Holland Biomed- ical Press. Amsterdam.
Bandler R, Tork I (1981): Behavioural-anatomical techniques as a means of studying the neural substrate for aggression. In Benton D, Brain P (eds): “The Biology of Aggression. ” Alphen aan den Rijn, The Netherlands Sijthoff-Noor-
Bermond B (1978): “Neurohormonal Regulation of Aggressive and Sexual Behaviour in the Rat.” Thesis State University Amsterdam.
dhoff, pp 367-382.
Berntson GG (1972): Blockade and release of hy- pothalamically and naturally elicited aggres- sive behaviors in cats following midbrain lesions. Journal of Comparative Physiology and
Blanchard RJ, Blanchard DC (1977a): Aggressive behaviour in the rat. Behavioral Biology
Blanchard RJ, Blanchard DC, Takahashi T, Kelley MJ (1977b): Attack and defensive behaviour in the albino rat. Animal Behaviour 25:622-634.
Blanchard DC, Blanchard RJ, Takahashi LK, Tak- ahashi T (1977~): Septa1 lesions and aggressive behavior. Behavioral Biology 21: 157-161.
Blanchard DC, Williams G, Lee EMC, Blanchard RJ (1981): Taming of wild rattus norvegicus by lesions of the mesencephalic central gray. Physiological Psychology 9: 157-163.
Chi CC, Flynn JP (1971a): Neural pathways asso- ciated with hypothalamically elicited attack be- haviour in cats. Science 171:703-706.
Chi CC, Flynn JP (1971b): Neuroanatomic projec- tions related to biting attack elicited from hy- pothalamus in cats. Brain Research 35:49-66.
Dixon WJ, Mood AM (1948): A method for ob-
Psychology 81 :541-554.
21: 197-224.

Central Gray Lesions and Aggression in Rats 155
taining and analyzing sensitivity data. Journal of the American Statistical Association 43: 109- 126.
Edwards MA, Adam DB (1974): Role of mid- brain central gray in pain-induced defensive boxing of rats. Physiology and Behavior
Everitt BS (1977): “The Analysis of Contingency Tables.” New York: John Wiley and Sons Inc.
Fernandez De Molina A, Hunsperger RW (1962): Organization of the subcortical system govern- ing defence and flight reactions in the cat. Jour- nal of Physiology 160:200-213.
French JD, Magoun HW (1952): Effects of chronic lesions in the central cephalic brain stem of monkeys. Archives of Neurology and Psychia- try, 68591-604.
Glusman M, Won W, Burdock EI, Ransohoff J (1961): Effects of midbrain lesions on “sav- age” behavior induced by hypothalamic lesions in the cat. Transactions of the American Neu- rological Association 86:216-218.
Hunsperger RW (1956): Role of substantia grisea centralis mesencephali in electrically-induced rage reactions. Progress in Neurobiology
Kallman MD, and Isaac W (1980): Disruption of illumination dependent activity by superior col- liculus destruction. Physiology and Behavior
Konig JFR, and Klippel RA (1963): “The Rat Brain, a Stereo-Taxic Atlas of the Forebrain and Lower Parts of the Brain Stem. Baltimore: The Williams and Wilkins Company.
Kruk MR, Kuiper P, Meelis W (1978): An airpres- sure-operated commutator system for electrical brain stimulation in a fighting rat. Physiology and Behavior 21: 125-127.
Kruk MR, Van der Poel AM, De Vos-Frerichs TP (1979): The induction of aggressive behaviour by electrical stimulation in the hypothalamus of male rats. Behaviour 70:292-322.
Kruk MR, and Van der Poel AM (1980): Is there evidence for a neural correlate of an aggressive behavioural system in the hypothalamus of the rat. In McConnell, Boer, Romijn, van der Poll, Corner (eds): “Adaptive Capabilities of the Nervous System, Progress in Brain Research
13: 113- 121.
1:289-294.
25 145-49.
53:385-390.
Lehman MN, Adams DB (1977): A statistical and motivational analysis of the social behaviors of the male laboratory rat. Behaviour 61:238-275.
Lindsley DB, Schreiner LH, Knowles WB and Magoun, HW (1950): Behavioral and EEG changes following chronic brain stem lesions in the cat. EEG Clinical Neurophysiology
Mos J, Kruk MR, Van der Poel AM, Meelis W (1982) Aggression induced by electrical stimu- lation in the midbrain central gray of male rats. Aggressive Behavior 8: 26 1-284.
Moyer KE (1968): Kinds of aggression and their physiological basis. Communications in Be- havioral Biology 2:65-87.
Olivier B, Mos J, Van der Poel AM, Kruk MR (1982): Woundpatterns following aggressive interactions in rat: Strain independent, situa- tion dependent. Abstract at the Meeting of the International Society for Research on Aggres- sion at Mexico City, 1982.
Pond FJ, Sinnamon HM, A d a m DB (1977): Sin- gle unit recording in the midbrain of rats dur- ing shock-elicited fighting behavior. Brain Research 120:469-484.
Scheffe, H (1959): “The Analysis of Variance.” New York: John Wiley and Sons, Inc.
Schmitt P, Paunovic VR, Karli P (1979): Effects of mesencephalic central gray and raphe nuclei lesions on hypothalamically induced escape. Physiology and Behavior 23:85-95.
Siege1 S (1956): Non-Parametric Statistics for the Behavioural Sciences. New York: McGraw Hill Book Company.
Skultety FM (1958): The behavioral effects of de- structive lesions of the periaqueductal gray matter in adult cats. Journal of Comparative Neurology 110:337-366.
Skultety FM (1963): Stimulation of periaqueductal gray and hypothalamus. Archives of Neurol-
Ulrich R (1966): Pain as a cause of aggression. American Zoology 6:643-662.
Wetherill GB (1966): Sequential estimation of points on quanta1 response curves. In Barrie G, Wetherill G (eds): “Sequential Methods in Sta- tistics.” London: Methuen and Co, Ltd., pp
21483-498.
ogy 8~608-620.
162- 179.







