
Characterization of the White-rot Fungus, Phanerochaete carnosa,
through Proteomic Methods and Compositional Analysis of Decayed Wood Fibre
By
Sonam Mahajan
A thesis submitted in conformity with the requirements for the degree of Doctorate of Philosophy
Department of Chemical Engineering and Applied Chemistry University of Toronto
© Copyright by Sonam Mahajan 2011

ii
Characterization of the white-rot fungus, Phanerochaete carnosa,
through proteomic methods and compositional analysis of decayed wood fibre
Sonam Mahajan
Doctorate of Philosophy
Department of Chemical Engineering and Applied Chemistry
University of Toronto
2011
Abstract
Biocatalysts are important tools for harnessing the potential of wood fibres since they can
perform specific reactions with low environmental impact. Challenges to bioconversion
technologies as applied to wood fibres include low accessibility of plant cell wall polymers and
the heterogeneity of plant cell walls, which makes it difficult to predict conversion efficiencies.
White-rot fungi are among the most efficient degraders of plant fibre (lignocellulose), capable of
degrading cellulose, hemicellulose and lignin. Phanerochaete carnosa is a white-rot fungus that,
in contrast to many white-rot fungi that have been studied to date, was isolated almost
exclusively from fallen coniferous trees (softwood). While several studies describe the
lignocellulolytic activity of the hardwood-degrading, model white-rot fungus Phanerochaete
chrysosporium, the lignocellulolytic activity of P. carnosa has not been investigated.

iii
An underlying hypothesis of this thesis is that P. carnosa encodes enzymes that are particularly
well suited for processing softwood fibre, which is an especially recalcitrant feedstock, though a
major resource for Canada. Moreover, given the phylogenetic similarity of P. carnosa and P.
chrysosporium, it is anticipated that the identification of pertinent enzymes for softwood
degradation can be more easily conducted. In particular, this project describes the
characterization of P. carnosa in terms of the growth conditions that support lignocellulolytic
activity, the effect of enzymes secreted by P. carnosa on the chemistry of softwood feedstocks,
and the characterization of the corresponding secretome using proteomic techniques. Through
this study, cultivation methods for P. carnosa were established and biochemical assays for
protein activity and quantification were developed. Analytical methods, including FTIR and
ToF-SIMS were used to characterize wood samples at advancing stages of decay, and revealed
preferential degradation of lignin in the early stages of growth on all softwoods analyzed.
Finally, an in depth proteomic analysis of the proteins secreted by P. carnosa on spruce and
cellulose established that similar sets of enzyme activities are elicited by P. carnosa grown on
different lignocellulosic substrates, albeit to different expression levels.

iv
Acknowledgements
I would like to express my deepest gratitude to Dr. Emma Master for accepting me as her PhD
student 5 some years ago, in spite of the sufficient lack of background that I brought with me in
this demanding field. Over the years, I have acquired technical skills and developed the ability to
learn newer information better - it would certainly not have been possible without her kind
patience throughout my learning stages. I am very grateful for her constant guidance,
intellectually stimulating discussions, understanding of my personal and technical challenges,
and encouragement during the trying times.
I am also very grateful to Dr. Dragica Jeremic for all her technical assistance, and more
importantly, for her friendly guidance throughout the later years of my PhD. It would have been
very difficult for me to see the end of my PhD if it were not for her un-ending support.
I am thankful to the members of my committee, Dr. Elizabeth Edwards and Dr. Krishna
Mahadevan, for the feedback and direction they provided for this project.
I am also grateful to Dr. Robyn Goacher for her technical assistance and contribution to the fibre
characterization studies, and to Peter Brodersen for his help in the early phases of fibre
characterization experiments with ToF-SIMS. I would also like to express my thanks to Dr. Eric
Yang from Sunnybrooke for his collaboration through the Proteomic Studies and to Dr. Tony
Ung for his assistance in sugar analysis and kind offering of all technical resources whenever
required.
I will always remember Jacqueline, for our friendly PhD pep-talks and for the technical and
moral support in designing the mammoth fibre characterization experiments. Finally, my huge
appreciation for all the members of the Master Lab and BioZone for their cooperation, and for
accepting me as a relatively inert member of the group, especially during the final stages of my
PhD.

v
I am very grateful to my parents, Dr. Ravi Mahajan and Dr. Kalpana Mahajan, who inspired me
to begin this marathon journey in life. Over the last 5 years, I’ve learnt a significant amount and
in spite of the challenges, I would have never taken it up, if it were not for them. My loving
thanks to my sister, Samridhi for adding a fresh breathe of non-academic humor to my
sometimes humdrum life.
I am indebted to my husband, Ateet, for his never-ending patience, gentle encouragement, kind
technical assistance and acceptance of all sloppy standards at home. And to all my dearest
friends, whose warmth created for me a home away from home, whose company made dull
moments bright, and whose smiles, encouraging emails, texts and generous home visits with
food, kept me alive through the most trying times…we did it!
Finally, my heartfelt gratitude to all my spiritual teachers and mentors, who have carved on my
heart determination and faith in the absolute will, the ability to discern the temporary from the
eternal, and the desire to serve with perfection…
Things that are very difficult to do become easy to execute if one somehow or other simply
remembers Lord Caitanya Mahaprabhu. But if one does not remember Him, even easy things
become very difficult. To this Lord Caitanya Mahaprabhu I offer my respectful obeisances.
Caitanya Caritamrita Adi Lila 14.1

vi
Table of Contents
Table of Contents ......................................................................................................................... vi
List of Figures .................................................................................................................................x
List of Abbreviations ................................................................................................................... xi
Chapter 1 : Overview.....................................................................................................................1
Chapter 2 : Literature Review ......................................................................................................7
2.1 Lignocellulose: A Valuable Resource ...............................................................................7
2.2 Composition and Structure of Plant Cell Walls ..............................................................8
2.3 Bioconversion of Lignocellulose........................................................................................9
2.4 Lignocellulose-degrading Bacteria ...................................................................................9
2.5 Lignocellulose-degrading Fungi .....................................................................................10
2.5.1 White-Rot Fungi...................................................................................................11
2.5.2 Brown-Rot Fungi .................................................................................................11
2.6 Lignocellulose Active Enzymes .......................................................................................12
2.6.1 Carbohydrate Active Enzymes ...........................................................................12
2.6.1.1 Cellulases ..............................................................................................................13
2.6.1.2 Hemicellulases ......................................................................................................13
2.6.1.3 Fungal Oxidative Lignin Enzymes .....................................................................14
2.7 Investigative Approaches to Improve Lignocellulose Bioconversion ..........................14
2.7.1 Analytical Characterization of Wood Fibre ......................................................15
2.7.2 Biochemical Characterization of Wood-degrading Fungi ...............................18
Chapter 3 : Effect of Cultivation Conditions on the Expression of Cellulolytic Activity by Phanerochaete species P. chrysosporium and P. carnosa ................................................30
3.1 Abstract .............................................................................................................................30
3.2 Introduction ......................................................................................................................30

vii
3.3 Review of P. chrysosporium Cultivation and Lignocellulolytic Activity .....................32
3.4 Materials and Methods ....................................................................................................41
3.4.1 Microorganism and Materials ............................................................................41
3.4.2 Cultivation Conditions.........................................................................................41
3.4.3 Growth Measurements ........................................................................................42
3.4.4 Biochemical Assays ..............................................................................................43
3.5 Results ...............................................................................................................................45
3.5.1 Effect of Cultivation Condition on the Extent of P. carnosa Growth on Microcrystalline Cellulose. ..............................................................................................45
3.5.2 Effect of Nitrogen Concentration and Agitation on Lignocellulolytic Expression by P. carnosa. ................................................................................................48
3.6 Discussion..........................................................................................................................51
Chapter 4 : Mode of Coniferous Wood Decay by the White Rot Fungus Phanerochaete carnosa as Confirmed by FT-IR and ToF-SIMS ..................................................................56
4.1 Abstract .............................................................................................................................56
4.2 Introduction ......................................................................................................................56
4.3 Materials and Methods ....................................................................................................59
4.3.1 Fungal Strain and Cultivation Conditions ........................................................59
4.3.2 Fourier Transform Infrared Spectroscopy .......................................................59
4.3.3 Transmission Electron Microscropy - Energy-Dispersive X-ray Analysis (TEM-EDXA) ...................................................................................................................60
4.3.4 Time of Flight Secondary Ion Mass Spectroscopy ............................................60
4.3.5 Statistical Analysis ...............................................................................................61
4.4 Results ...............................................................................................................................61
4.4.1 FTIR Spectroscopy ..............................................................................................62
4.4.2 TEM-EDXA ..........................................................................................................68
4.4.3 ToF-SIMS .............................................................................................................68

viii
4.5 Discussion..........................................................................................................................71
Chapter 5 : Proteomic Characterization of Lignocellulose-degrading Enzymes Secreted by Phanerochaete carnosa Grown on Spruce and Microcrystalline Cellulose ..................79
5.1 Abstract .............................................................................................................................79
5.2 Introduction ......................................................................................................................79
5.3 Materials and Methods ....................................................................................................81
5.3.1 Cultivation Conditions.........................................................................................81
5.3.2 Protein Extraction ................................................................................................81
5.3.3 Protein Preparation and Analysis by Mass Spectrometry ...............................82
5.3.4 Peptide Sequence Annotation .............................................................................82
5.4 Results ...............................................................................................................................83
5.4.1 Preparation and Analysis of Peptide Samples...................................................83
5.4.2 Cellulases and Hemicellulases in Cellulose and Spruce Cultivations .............84
5.4.3 Oxidoreductases in Cellulose and Spruce Cultivations ....................................91
5.4.4 Carbohydrate Esterases, Proteases and Other Glycoside Hydrolases ............95
5.4.5 Peptides Annotated to Hypothetical Proteins Identified in Proteomic Analyses of P. chrysosporium ........................................................................................100
5.5 Discussion........................................................................................................................101
Chapter 6: Synthesis and Conclusions .....................................................................................107
Chapter 7: Engineering Relevance ...........................................................................................110
Appendix 1 : B3 Medium ..........................................................................................................113
Appendix 2: Supplemental Information for Chapter 4 ..........................................................115
Appendix 3: Supplemental Information for Chapter 5 ..........................................................119
Appendix 4: Substrate recognition and hydrolysis by a fungal xyloglucan-specific family 12 hydrolase ...............................................................................................................126
Appendix 6: Additional Information Collected During ResearchError! Bookmark not defined.

ix
List of Tables
Table 3.1 Summary of carbohydrate active enzyme production by Phanerochaete chrysosporium
Table 3.2 Expression of lignocellulolytic activity by Phanerochaete chrysosporium
Table 3.3 Cultivation of P. carnosa in B3 medium at 27˚C with 1 % microcrystalline cellulose, variable nitrogen, pH 4.5 Table 3.4 Protein concentration in P. carnosa culture supernatant during growth on microcrystalline cellulose
Table 3.5 Chitin content of P. carnosa cultivations after 24 days
Table 3.6 Endoglucanase activity in P. carnosa culture supernatant during growth on microcrystalline cellulose
Table 3.7 β-glucosidase activity in P. carnosa culture supernatant during growth on microcrystalline cellulose Table 3.8 Total cellulolytic activity in P. carnosa culture supernatant during growth on microcrystalline cellulose Table 3.9 MnP activity in P. carnosa culture supernatant during growth on microcrystalline cellulose Table 4.1 Assignment of FTIR peaks that distinguish control and decayed wood samples Table 5.1 Analysis of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce that correspond to predicted cellulases and hemicellulases Table 5.2 Analysis of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce that correspond to extracellular oxidases and proteins involved in lignin degradation Table 5.3 Analysis of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce that correspond to extracellular carbohydrate-active enzymes Table 5.4 Peptides from extracellular filtrates of P. carnosa grown on cellulose that were annotated to proteins from P. chrysosporium with unknown function

x
List of Figures Figure 3.1 SDS-PAGE of P.carnosa culture supernatants Figure 4.1 Spruce decay described by FTIR analysis Figure 4.2 Fir decay described by FTIR analysis Figure 4.3 Pine decay described by FTIR analysis Figure 4.4 PCA of normalized and mean-centered ToF-SIMS spectra: A) Fir, B) Pine, and C) Spruce Figure 4.5 PCA of mean-centered ToF-SIMS image of decayed pine (time point 5) Figure 5.1 Distribution of peptide annotations from proteins produced by P. carnosa grown on (A) crystalline cellulose, and (B) spruce wood chips Figure 6.1 Synthesis of information flow between the three key objectives of this thesis

xi
List of Abbreviations FTIR - Fourier Transform InfraRed ToF-SIMS - Time-of-Flight Secondary Ion Mass Spectrometry TEM-EDXA - Transmission Electron Microscope with Energy Dispersive X-ray Analysis SEM - scanning electron microscopy CAZymes - Carbohydrate Active Enzymes FOLymes - Fungal Oxidative Lignin enzymes GH - Glycoside Hydrolase CE - Carbohydrate esterase LiP - Lignin peroxidase MnP - Manganese peroxidase ABTS - 2,2'-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid) EGase - Endoglucanase CBH - Cellobiohydrolase CBD - Cellobiose Dehydrogenase BGL - beta-glucosidase FPU - Filter Paper Units XYN - Xylanase MAN - Mannanase PCR - Polymerase chain reaction Ribosomal ITS sequences – ribosomal internal transcribed spacer sequences BSA - Bovine Serum Albumin LN - Low nitrogen, stationary cultivations LNS - Low nitrogen cultivations with shaking HN - Low nitrogen, stationary cultivations PCA - Principal Component Analysis PLS Toolbox - Partial Least Squares Toolbox PDMS - Polydimethylsiloxane MWCO – Molecular weight cut off

1
Chapter 1 : Overview
Terrestrial plants synthesize nearly 200 billion tons of lignocellulosic biomass per year (Zhang
2008). Lignocellulose comprises the structural component of plant cell walls and is composed of
cellulose, hemicellulose, and lignin, as well as relatively low amounts of aliphatic and alicyclic
compounds, phenolic compounds, and proteins. The “lignocellulose biorefinery” aims to produce
renewable fuels, chemicals and new polymers from all the constituents of lignocellulose. As the
non-consumable portion of plant biomass, bioproducts from lignocellulose do not directly
compete with food resources. Moreover, lignocellulose can be supplied as a residual of
agricultural and forest industries.
Despite the advantages of lignocellulose, the complex and variable composition of this feedstock
creates significant technical barriers that can be broadly grouped into 1) the production of
fermentable sugars and technical polymers, and 2) the conversion of mixed sugars to fuels and
chemicals. Current research towards efficient production of fermentable sugars and polymeric
materials is aimed at improving the integration of pretreatment technologies and downstream
enzyme applications, and minimizing the production of compounds that inhibit fermentation
processes (Kabel et al. 2006). Still, the yield of fermentable sugars from enzyme treatments is
variable and incomplete, and the effect of commercially available enzymes often depends on the
nature of the substrate, rather than advertised enzyme activity (Lynd et al. 2008).
White-rot fungi are the most efficient degraders of lignocellulose as they can degrade cellulose
and hemicellulose, as well as lignin (Schmidt 2006). Many white-rot fungi have been isolated
from hardwoods. By contrast, Phanerochaete carnosa is a white-rot fungus that has been
isolated predominantly from softwood (Burdsall 1985). While several studies describe the
lignocellulolytic behaviour of the hardwood-degrading, model white-rot fungus P.
chrysosporium (Sato et al. 2007; Vanden Wymelenberg et al. 2005a; Vanden Wymelenberg et al.
2005b), the lignocellulolytic activity of P. carnosa has not been investigated. As a softwood
degrader, P. carnosa might encode enzymes that are particularly well suited for processing
softwood fibre, which consists of different lignin and hemicellulose composition, compared to
hardwood. Given the abundance and comparatively high recalcitrance of softwood fibre (Mosier

2
et al. 2005), enzymes that effectively process this material would benefit forest sectors in
northern countries, including Canada, where boreal forests occupy 77% of the forest land
(Natural Resources Canada 2011).
Accordingly, the main aim of this PhD thesis is to characterize the secretome of P. carnosa
during growth on lignocellulosic substrates, and to evaluate the pattern of wood decay by this
fungus over time. In addition to increasing our fundamental understanding of bioprocesses that
have evolved to transform recalcitrant biomass, the analyses described herein assesses the
potential of P. carnosa to be used for biopulping and as a supply of industrially relevant
enzymes.
In particular, the following specific research hypotheses were tested:
1. Cultivation conditions that promote the expression of lignocellulolytic enzymes in P.
chrysosporium will also induce the expression of lignocellulolytic enzymes in P. carnosa.
2. P. carnosa will exhibit selective degradation of lignin at early stages of softwood decay, and
having been isolated from softwood species, P. carnosa will efficiently degrade guaiacyl
lignin, which dominates in softwood.
3. The profile of proteins secreted by P. carnosa during growth on lignocellulose will depend
on the source of the substrate, and could be used to predict enzyme formulations that
optimally transform a particular biomass feedstock.
As summarized below, the research hypotheses listed above were tested as described in chapters
three, four and five, respectively.
Chapter 3: Effect of cultivation conditions on the expression of cellulolytic activity by
Phanerochaete species, P. chrysosporium and P. carnosa
This study describes the first reported effort to cultivate and biochemically characterize
Phanerochaete carnosa. Accordingly, the primary aim of this study was to establish growth
conditions that induce the expression of lignocellulolytic enzymes by this organism, and to
investigate the effect of nitrogen concentration, temperature and agitation on the production of
cellulolytic and lignin-degrading activities. Protein concentration in culture supernatants, fungal

3
growth, and biochemical assays for cellulases, lignin peroxidases and manganese peroxidases,
were measured. By comparing the effect of cultivation condition on the production of
lignocellulolytic activity by P. carnosa and P. chrysosporium, differences in enzyme activities
and their regulation were also evaluated.
Chapter 4: Mode of coniferous wood decay by the white-rot fungus Phanerochaete carnosa
as elucidated by FTIR and ToF-SIMS (a version of this chapter has been accepted for
publication)
The objective of this study was to investigate the effect of conifer species and stage of
degradation on the mode of softwood decay by P. carnosa (i.e., selective vs. simultaneous
decay). The three wood species investigated were white spruce, lodgepole pine and balsam fir,
all of which are abundant in the North American boreal forests, and comprise common mixed
feedstocks for pulp and paper industries. FTIR and ToF-SIMS were used to monitor changes in
lignocellulose composition resulting from fungal activity. Additionally, TEM and SIMS imaging
were used to evaluate the selectivity of the degradation across cell walls by determining the
localization of lignin prior and post-decay.
Chapter 5: Proteomic analysis of the secretome of P. carnosa grown on microcrystalline
cellulose and spruce wood chips (a version of this chapter was published in Applied and
Microbial Biotechnology)
The objective of this research was to describe the secretome of P. carnosa while growing on
cellulose and spruce. A comparative study of the extracellular proteins expressed in both
cultivation conditions was performed to determine whether a model cellulose substrate elicited a
similar profile of lignocellulose-active enzymes as a complex lignocellulosic substrate. Since the
genome sequence of P. carnosa was not available, the proteomic protocols were modified to
accommodate de novo sequencing and manual validation of peptides along with sequence-
similarity based annotations. In this way, the regulation of lignocellulose-active enzyme
expression in P. carnosa was investigated, as was the possibility to define enzyme formulations
that are optimal for particular feedstocks. Finally, the proteomic data resulting from this study
were analyzed to predict enzyme activities that may have evolved in P. carnosa to promote
softwood degradation.

4
As summarized in Chapters 6 and 7, the results of the research described herein elucidate the
growth conditions that support lignocellulolytic activity of P. carnosa, the differential effects of
P. carnosa on the chemistry of softwood feedstocks, and the expression profile of enzymes
secreted by P. carnosa during growth on cellulose and a lignocellulosic feedstocks. As a result,
the softwood-degrading activity of P. carnosa was confirmed and the applied potential of P.
carnosa and encoded enzymes was explored.
Summary of Scholarly Contributions: A. Peer-reviewed Publications: MASc 1. S. Mahajan, S.K. Konar and D.G.B. Boocock. Standard Biodiesel from Soybean Oil by a Single Chemical Reaction. Journal of American Oil Chemists’ Society. 2006. 84: 641-644. 2. S. Mahajan, S.K. Konar and D.G.B. Boocock. Determining acid number for biodiesel. Journal of American Oil Chemists’ Society. 2006. 83: 567-570. 3. S. Mahajan, S.K. Konar and D.G.B. Boocock. Variables Affecting the Production of Standard Biodiesel. Journal of American Oil Chemists’ Society. 2007. 84: 189-195. PhD 4. J. Powlowski, S. Mahajan, M. Schapira and E.R. Master. Substrate Recognition and Hydrolysis by a Fungal Xyloglucan-Specific Family 12 Hydrolase. Carbohydrate research. 2009. 344(10):1175-9. (see Appendix 4). (Contribution: Performed enzymatic digestion for
xyloglucans and detected products through HPLC) 5. S. Mahajan and E.R. Master. Proteomic Characterization of Lignocellulose-degrading Enzymes Secreted by Phanerochaete carnosa Grown on Spruce and Microcrystalline Cellulose. Applied Microbiology and Biotechnology. 2010. 86(6):1903-14. (see Chapter 5) 6. S. Mahajan, D. Jeremic, R. E. Goacher and E. R. Master. Mode of coniferous wood decay by the white-rot fungus Phanerochaete carnosa as elucidated by FTIR and ToF-SIMS. 2011 (accepted with revisions; see Chapter 4). (Contribution: established growth conditions for P.
carnosa on softwood species; harvested samples and performed FTIR analysis; prepared

5
samples for ToF-SIMS and performed data analyses in collaboration with D. Jeremic and R.E.
Goacher.) B. Conference Presentations (Presenter is underlined): MASc 1. S. Mahajan, S.K. Konar and D.G.B. Boocock. ‘Production of Standard Biodiesel from Soybean Oil: A one-step process using NaOH’ (speaker + poster). 55th Canadian Chemical Engineering Conference, Toronto, ON, October 2005 PhD 2. S. Mahajan and E.R. Master. ‘Proteomic Analysis of Softwood-degrading Fungi’ (speaker). 56th Canadian Chemical Engineering Conference, Sherbrooke, QC, October 2006 3. S. Mahajan and E.R. Master. ‘Proteomic Analysis of Softwood-degrading Fungi’ (poster). 56th Canadian Chemical Engineering Conference, Sherbrooke, QC, October 2006 4. S. Mahajan. ‘Unit Operations Laboratory: Problem based learning’ (speaker). 56th Canadian Chemical Engineering Conference, Sherbrooke, QC, October 2006 5. S. Mahajan and E.R. Master. ‘Proteomic Analysis of Softwood-degrading Fungi towards Biomimetic Enzyme Applications’ (poster). The World Congress on Industrial Biotechnology and Bioprocessing, Toronto, ON, July 2006 6. S. Mahajan and E.R. Master. ‘Proteomic Analysis of Softwood-degrading Fungi’ (poster). 107th General Meeting of the American Society for Microbiology.Toronto, ON May 2007 7. S. Mahajan and E.R. Master. ‘Refining models of Lignocellulose Biotransformation towards Biomimetic Enzyme Applications’ (poster). 10th International Congress on Biotechnology in the Pulp and Paper Industry, Madison WI, June 2007 8. J. MacDonald, S. Mahajan and E. R. Master ‘Transcription Profiles and Proteomic Analysis of P. carnosa Grown on Softwood Feedstocks for Tailored Applications of Hydrolytic Enzymes.’ MIE BioForum Mie, Japan. Sep 2008 9. S. Mahajan and E.R. Master. ‘Proteomic Analysis of the Secretome of Softwood-Degrading Fungi’ (poster). 4th annual conference US HUPO North Bethesda, MD, March 2008 10. S Mahajan, E Yang and E.R. Master. ‘The secretome of the softwood-degrading Basidiomycete, Phanerochaete carnosa, grown on cellulose and spruce.’

6
Gordon Research Conference (GRC): Cellulosomes, Cellulases & Other Carbohydrate Modifying Enzymes. Proctor Academy, NH, USA. July 26-31, 2009. 11. J. MacDonald, S. Mahajan and E. R. Master. ‘Proteomic and Transcriptomic Analysis of Lignocellulose-degrading enzymes in softwood-degrading fungus, Phanerochaete carnosa’ Lignobiotech One Symposium, Reims, France, March 2010 Chapter 1 References Burdsall HHJ (1985) A contribution to the taxonomy of the genus Phanerochaete (Corticiaceae,
Aphyllophorales). Mycological Memoir No. 10. Braunschweig. Kabel MA, van der Maarel MJEC, Klip G, Voragen AGJ, Schols HA (2006) Standard assays do
not predict the efficiency of commercial cellulase preparations towards plant materials. Biotechnology and Bioengineering 93 (1):56-63.
Lynd LR, Laser MS, Bransby D, Dale BE, Davison B, Hamilton R, Himmel M, Keller M,
McMillan JD, Sheehan J, Wyman CE (2008) How biotech can transform biofuels. Nature Biotechnology 26 (2):169-172
Mosier N, Wyman C, Dale B, Elander R, Lee YY, Holtzapple M, Ladisch M (2005) Features of
promising technologies for pretreatment of lignocellulosic biomass. Bioresource Technology 96 (6):673-686
Sato S, Liu F, Hasan K, Tien M (2007) Expression analysis of extracellular proteins from
Phanerochaete chrysosporium grown on different liquid and solid substrates. Microbiology 153:3023-3033
Schmidt O (2006) Wood and Tree Fungi. Biology, Damage, Protection and Use. Springer-
Verlag, New York Vanden Wymelenberg A, Sabat G, Martinez D, Rajangam AS, Teeri TT, Gaskell J, K KPJ,
Cullen D (2005a) The Phanerochaete chrysosporium secretome: Database predictions and initial mass spectrometry peptide identifications in cellulose-grown medium Journal of Biotechnology 118 (1):17-34
Vanden Wymelenberg A, Sabat G, Martinez D, Rajangam AS, Teeri TT, Gaskell J, Kersten PJ,
Cullen D (2005b) The Phanerochaete chrysosporium secretome: database predictions and initial mass spectrometry peptide identifications in cellulose-grown medium. Journal of Biotechnology 118:17-34
Zhang YH (2008) Reviving the carbohydrate economy via multi-product lignocellulose
biorefineries. Journal of Industrial Microbiology Biotechnology 35 (5):367-375.

7
Chapter 2 : Literature Review
2.1 Lignocellulose: A Valuable Resource
Lignocellulose is the major structural component of woody plants. It is a network of lignin,
cellulose and hemicellulose that is chemically bonded through non-covalent forces and covalent
cross-linkages (Perez et al. 2002).
Terrestrial plants synthesize nearly 200 billion tons of lignocellulosic biomass per year (Zhang
2008). Forests, which are the primary source of lignocellulosic biomass, cover approximately
30% of the total land area, and comprise a significant source of energy, timber, and pulp (Global
Forest Resources Assessment 2005). Protective functions of forests include soil and water
conservation, biological diversity and mitigation of climate change. While conventional research
in wood fibre structure and composition focused on properties that improve timber, pulp and
paper qualities, present research has expanded to include bioconversion of wood fibre to
platform sugars that can be fermented to fuels and chemicals, and bioprocessing techniques that
increase the sustainability of processes used to convert wood fibre to high-quality pulp and other
value-added materials.
Large amounts of lignocellulosic residues (6 to 3849 million t and from 64 to 561 million t, for
US and Canada) are generated through forestry and agricultural practices, with the most common
disposal being combustion (Magdalena 2008). This is not only an environmental concern but
also represents an under-utilization of a renewable resource that can be converted to energy and
biochemicals. For instance, over 75% of organic chemicals are produced from five primary base
chemicals: ethylene, propylene, benzene, toluene and xylene. The aromatic compounds could be
produced from lignin, while the low molecular weight aliphatic compounds could be obtained
from alcohols that result from fermentation of sugars generated from bioconversion of
hemicellulose and cellulose (Howard et al. 2003). Moreover, the alcohols produced could be
utilized as a biofuel. And chemicals like vanillin, xylitol, and furfural from lignocellulosic wastes
can be used in industrial products including herbicides, pharmaceuticals, and household products
(Howard et al. 2003; Ragauskas et al. 2006).

8
2.2 Composition and Structure of Plant Cell Walls
The main components of plant cell walls are cellulose, lignin, and hemicellulose. Cellulose and
hemicellulose are polysaccharides, whereas lignin is an aromatic polymer synthesized from
phenylpropanoid precursors (Sjostrom 1983). Wood from different tree species is typically
composed of 40–50% α-cellulose, 20–35% hemicellulose and 15–35% lignin (Perez et al. 2002).
With the exception of cellulose, these polymers are synthesized inside the cell and then
organized outside the cell membrane. The primary cell wall is deposited first and is characterized
by relatively amorphous cellulose structure. During cell differentiation, the primary cell wall
expands and elongates, then secondary cell walls are synthesized, which are characterized by
cellulose microfibrils with higher crystallinity and altered hemicellulose content (Ding and
Himmel, 2006). Differentiated xylem cells with a secondary cell wall are called secondary xylem
or wood.
Cellulose is the most abundant organic polymer on earth, and is composed of repeating
cellobiose subunits that consist of β-1,4-linked glucose. The degree of polymerization (DP) of
wood-derived cellulose is typically greater than 10,000 and adjacent cellulose polymers interact
through hydrogen bonds, forming highly stable structures that contain both amorphous and
crystalline regions (Lerouxel et al. 2006; McCann and Carpita 2008). Cellulose is synthesized
by a cellulose synthase complex that is located within the cytoplasmic membrane of plant cells.
In plant cells, the cellulose synthase is comprised of many enzymes that include 36 cellulose
synthase enzymes assembled as rosette structure (Taylor et al. 2000). Accordingly, 36 cellulose
molecules are thought to emerge from each rosette structure.
Like cellulose, hemicellulose backbones are composed of β-1,4-linked sugars. However, the
particular sugar composition of hemicellulose depends on the source of the polysaccharide. For
instance, xyloglucan is the main hemicellulose of primary cell walls and consists of a β-1,4-
linked glucose backbone that is decorated with xylose, galactose and sometimes fucose
branching sugars (Hayashi and Kaida 2011). By contrast, the main hemicellulose in secondary
cells walls of hardwoods and softwoods is xylan and galactoglucomannan, respectively. Xylan is
composed of β-1,4-linked xylose that can be substituted by arabinose and glucuronic acid; xylan

9
can also be acetylated. By contrast, wood-derived mannans are composed of β-1,4-linked
mannose and glucose that can be substituted by galactose (Stenius and Vuorinen 1999). These
polymers have a lower DP than cellulose, averaging between 100 and 200 and have a lower
crystallinity (Rowell 2005a). While cellulose is synthesized by cellulose synthases located within
the cytoplasmic membrane, hemicelluloses are generated in the Golgi complex, and then secreted
to the plant cell wall (Keegstra 2010).
Lignin is a complex polymer that is composed of phenylpropane units linked together by carbon-
carbon (C-C) and ether (C-O-C) linkages. Lignin provides structural support to plant fibres, as
well as resistance against microbial attack and improved water transport in xylem and phloem
tissues. Precursors of lignin biosynthesis are p-coumaryl alcohol, coniferyl alcohol, and sinapyl
alcohol, and while p-coumaryl alcohol predominates in grasses to form H-lignin, coniferyl
alcohol is the main monolignol in softwood G-lignin, and hardwood G/S-lignin contains both
sinapyl and coniferyl monolignols (Humphreys and Chapple 2002).
Besides carbohydrates and lignin, the cell wall contains hydroxyproline-rich glycoproteins,
arabinogalactan proteins, glycine-rich proteins, and proline-rich proteins (Showalter 2001). Other
extraneous compounds that do not contribute to the structure of cell walls are grouped as
extractives and ash (Pettersen 1984).
2.3 Bioconversion of Lignocellulose
Physical and chemical processes have been developed to pretreat wood fibres to separate
cellulose, hemicellulose, and lignin; however, these processes can decrease the quality of the
polymers and create by-products that inhibit the fermentation of resulting sugars. Alternatively,
biocatalysts (enzymes) can perform specific reactions under comparatively mild reaction
conditions, and could be used to improve the pretreatment process (Lynd et al. 2008).
2.4 Lignocellulose-degrading Bacteria
Evidence of bacterial degradation of lignin is sparse. Streptomycete species have been shown to
degrade low levels of lignin (Crawford, 1978; Watanabe et al. 2003), and multicopper oxidases

10
with laccase activity have been isolated from bacteria, although these enzymes are expected to
mainly participate in sporulation (Claus 2004; Malherbe and Cloete 2002).
Accordingly, bacteria are generally considered secondary lignocellulose degraders, and can
degrade cellulose and hemicellulose both aerobically and anaerobically (Walker and Wilson
1991). Examples of aerobic (hemi)cellulose degraders include Thermobifida fusca and
Cellulomonas composti, as well as several other bacteria (Béguin and Aubert 1994). In these
cases, degradation is initiated by the concerted activity of cell-associated and free extracellular
cellulases and hemicellulases (Walker and Wilson 1991). By contrast, anaerobic cellulose-
degrading bacteria typically anchor relevant hydrolase activities to the cell through a cellulosome
complex. Cellulosomes are multienzyme complexes that bind to the bacterial cell wall and
promote the uptake of solubilized sugars by the hydrolytic organism (Bayer et al. 2008).
Clostridium thermocellum and C. cellulolyticum are among the best-studied anaerobic, cellulose-
degrading bacteria (Bayer et al. 2008). Notably, these organisms have been extensively
evaluated for their potential to promote simultaneous saccharification and fermentation (SSF) or
consolidated bioprocessing (CBP) (Lynd et al. 2002).
2.5 Lignocellulose-degrading Fungi
Fungi are the primary degraders of lignocellulose (Rabinovich et al. 2002; Sanchez 2009). In
addition to secreting enzymes that are critical to lignocellulose decomposition, fungal growth on
lignocellulose is promoted by the formation of mycelia that allow filamentous fungi to transport
nutrients, including nitrogen and iron, to the carbon-rich lignocellulosic substrate (Hammel
1997). Many fungi are also more resistant to wood-derived biocides that limit bacterial growth.
These compounds include tannins and various phenolic compounds (terpenes, stilbenes,
flavonoids and tropolones) that are particularly abundant in the heartwood of fallen trees.
The majority of wood-degrading fungi that have been characterized to date are members of the
phylum Basidiomycota and are characterized by either brown-rot or white-rot decay.

11
2.5.1 White-Rot Fungi
Fungi causing white-rot decay secrete enzymes that degrade lignin, hemicellulose and cellulose,
leaving the residual wood fibre with a bleached appearance. Two main patterns of white-rot
decay have been distinguished by microscopic and ultra structural investigations (Liese 1970).
Simultaneous white-rot (“corrosion rot”) is exemplified by combined degradation of
carbohydrates and lignin at early and late stages of decay. Examples of fungi that elicit
simultaneous white-rot include Fomes fomentarius, Phellinus robustus, and Trametes versicolor
(Blanchette 1984, Blanchette 1994). By contrast, selective (sequential) white-rot is exemplified
by early degradation of lignin and hemicelluloses followed by cellulose degradation.
Ceriporiopsis subvermispora and Phlebia radiata are perhaps the best studied fungi to elicit
selective white-rot decay (Ander and Eriksson, 1977; Blanchette, 1991; Fackler et al., 2007).
Often, the sequential white-rot fungi “selectively” degrade lignin and hemicellulose in small
elongated cavities within a wood tissue such that decayed regions are surrounded by tissue that
appears sound (Blanchette 1984). With advancing decay and delignification of the middle
lamella and primary cell wall, the wood sample typically acquires a fibrous texture (Schmidt
2006). Importantly, elicitation of simultaneous or sequential white-rot decay can depend on the
wood that is being degraded, the stage of wood degradation, and the particular fungal strain used
in the study (Messner and Srebonik, 1994). For instance, certain strains of Phanerochaete
chrysosporium (e.g. BKM-F-1767), cause selective decay of deciduous wood samples, while
many other strains cause simultaneous wood decay (Blanchette 1992).
2.5.2 Brown-Rot Fungi
Brown-rot decay is characterized by rapid degradation of cellulose and hemicellulose, and
retention of a modified lignin residue (Yelle et al. 2008). To date, brown-rot fungi were mainly
isolated from coniferous (softwood) trees and represent approximately 7 % of isolated wood-
rotting basidiomycetes (Martínez et al. 2005). Brown-rot decay can cause rapid loss in the
structural strength of construction wood (Green III and Highley 1997), and decayed woods are
typically characterized by reddish brown color and dry, crumbly and brittle consistency
(Martínez et al. 2005). Examples of brown-rot fungi include Fomitopsis lilacino-gilva,
Laetiporus portentosus, Postia placenta, Gloeophyllum trabeum and Serpula lacrymans. In
contrast to the numerous enzymes secreted by white-rot fungi, brown-rot fungi appear to initiate

12
the degradation of wood polysaccharides using Fenton chemistry, whereby diffusible hydroxyl
radicals penetrate the wood surface and depolymerize cellulose and hemicelluloses while leaving
lignin intact (Jensen Jr. et al. 2001).
Studies on the phylogeny and substrate preference of wood decaying fungi suggest that the
brown-rot fungi have evolved multiple times from white-rot fungi. Hibbet et al. (2001) predict
that white-rot, tetrapolar mating systems, and ability to degrade conifers and hardwoods is a
primitive characteristic of wood-degrading fungi, whereas brown-rot, bipolar mating systems,
and exclusive decay of conifers, evolved subsequently from these ancestral characteristics
(Hibbett and Donoghue 2001).
2.6 Lignocellulose Active Enzymes
Given the polymeric and complex composition of lignocellulose, it is not surprising that the
initial attack of this substrate is achieved through the concerted activity of several extracellular
microbial enzymes. Lignocellulose-active enzymes that are produced by white-rot fungi are
particularly valuable for biomass conversion, since they can be used to selectively transform both
lignin and polysaccharides (Kirk and Cullen 1998). The enzymes that contribute to this activity
can be broadly classified as Carbohydrate-Active enzymes (CAZymes) and Fungal Oxidative
Lignin enzymes (FOLymes) (Cantarel et al. 2009; Levasseur et al. 2008). The following is a
summary of the main enzymes involved in these degradation processes.
2.6.1 Carbohydrate Active Enzymes
A sequence-based classification scheme for carbohydrate-active enzymes was developed in
1991, called the CAZy database (CArbohydrate enZYme database) (Cantarel et al. 2009;
Henrissat 1991). At present, this database is comprised of 125 glycoside hydrolase families, 92
glycoside transfer families, 22 polysaccharide lyase families and 16 carbohydrate esterase
families. Glycoside hydrolases hydrolyze the glycosidic bonds between α-linked or β-linked
sugars, using a retaining or inverting mechanism (Davies and Henrissat 1995). Polysaccharide
lyases cleave polysaccharide chains via a β-elimination mechanism resulting in the formation of
a double bond at the newly formed non-reducing end, whereas carbohydrate esterases catalyze
the deacetylation and demethylation of substituted polysaccharides. Efficient degradation of

13
polysaccharides requires cooperation or synergistic interactions between enzymes responsible for
cleaving the different linkages. Significant research has been done to demonstrate and
understand synergy between various isolated enzymes for degradation of microcrystalline
cellulose (Avicel) and commercial xylans (de Vries and Visser 2001). For instance, hydrolysis
of xylan by an Aspergillus xylanase was increased in the presence of accessory enzymes that
catalyze the hydrolysis of xylan side chains (Paszczynski et al. 1988).
2.6.1.1 Cellulases
Hydrolysis of cellulose to glucose requires the activity of endoglucanases (endo-cellulases),
cellobiohydrolases and β-glucosidases. While endo-cellulases hydrolyze glycosidic linkages at
internal positions within cellulose molecules, cellobiohydrolases release cellobiose from the
reducing or non-reducing end of cellulose and β-glucosidases hydrolyze cellobiose to glucose.
These enzymes work synergistically to degrade both amorphous and crystalline cellulose, and
endo-cellulases and cellobiohydrolases are often associated with cellulose-binding modules to
promote their activity on polymeric substrates (Kirk and Cullen 1998). Of the 125 GH families,
fungal cellulases belong to GH families 5, 6, 7, 9, 12, 44, 45, 48, 61 and 74 (Dashtban et al.
2009). In addition to the hydrolytic enzymes, oxidative enzymes also participate in cellulose
degradation (Kirk and Cullen 1998). For example, quinone oxidoreductase (cellobiose
dehydrogenase) reduces quinones and phenoxy radicals in the presence of cellobiose, which is
oxidized to cellobiono-δ-lactone. And cellobiose oxidase uses molecular oxygen to oxidize
cellobiose and longer cello-oligomers to corresponding acids.
2.6.1.2 Hemicellulases
Wood hemicelluloses include xylan, (galacto)glucomannan, and xyloglucan. These
polysaccharides contain a β-1,4-linked sugar backbone that can be acetylated or substituted by
sugar branches (Scheller and Ulvskov 2010). Given the diversity of hemicelluloses, many
glycoside hydrolases, as well as carbohydrate esterases participate in their degradation. For
instance, xylan degradation requires the activity of xylanases, xylosidases, arabinofuranosidases,
galactosidases, deacetylases, glucuronidases, glucuronyl esterases, and feruloyl esterases. Like
cellulases, many of these activities function synergistically and are associated with carbohydrate-
binding modules that promote enzyme activity on polymeric substrates (de Vries et al. 2000;

14
Hervé et al. 2010). So far, fungal hemicellulases were identified in nineteen GH families: 1, 2, 3,
5, 10, 11, 26, 27, 36, 39, 43, 51, 53, 54, 62, 67, 74,115, and 116, and nine CE families: 1, 2, 3, 4,
5, 6, 12, 15 and 16.
2.6.1.3 Fungal Oxidative Lignin Enzymes
Similar to carbohydrate-active enzymes, enzymes involved in lignin catabolism have also been
grouped into sequence-based families and integrated in a database, named the Fungal Oxidative
Lignin enzymes (FOLy) (Levasseur et al. 2008).
Laccases and peroxidases are extracellular, lignolytic enzymes that are key to enzymatic lignin
degradation (Perez et al. 2002; ten Have and Teunissen, 2001). The peroxidases include lignin
peroxidase (LiPs) and manganese-dependent peroxidase (MnP). Both LiP and MnP oxidize
corresponding substrates through two consecutive one-electron oxidation steps with
intermediate cation radical formation (Sanchez 2009). LiP degrades non-phenolic lignin units (up
to 90% of the polymer) whereas MnP generates Mn3+, which acts as a diffusible oxidant of
phenolic or non-phenolic lignin units likely mediated through lipid peroxidation reactions
(Cullen and Kersten 2004; Moen and Hammel, 1994).
Laccases are blue copper oxidases that catalyze the one-electron oxidation of phenolics, aromatic
amines, and other electron-rich substrates with the concomitant reduction of O2 to H2O (d'Souza
et al. 1999). Like Mn(III) chelates, laccases oxidize the phenolic units in lignin to phenoxy
radicals, which can lead to aryl-C cleavage (Kawai et al. 1988). Laccase can also oxidize non-
phenolic substrates in the presence of certain auxiliary substrates such as 2,2´-azino-bis-3-
ethylthiazoline-6- sulfonate (Call and Muncke 1997).
2.7 Investigative Approaches to Improve Lignocellulose Bioconversion
Challenges associated with bioconversion of lignocellulose can be attributed to the distinct
composition and structure of different plant cell walls. For instance, the accessibility of enzymes
to fibre polymers can vary, and the enzymes required to depolymerize cell wall components
depends on the source of fibre. Moreover, since plant cell walls are a composite of different

15
polymers, multiple enzymes are necessary to process fibres, and the enzyme requirement is
likely to change as the fibre is treated.
Methods for increasing the efficiency of biocatalysts include screening for organisms with novel
enzymes, improvement of existing industrial strains, enzyme engineering and development of
enzyme cocktails that are tailored to particular lignocellulosic feedstocks. Considerable progress
has been made concerning the biochemistry of purified, fungal enzymes that participate in wood
degradation. However, the effect of enzyme activities on natural lignocellulosic feedstocks is
limiting. It is also unclear how the profile of enzymes secreted by wood-degrading fungi
depends on the composition of the lignocellulosic feedstock.
It is anticipated that by characterizing the effect of lignocellulose composition on the expression
of lignocellulolytic enzymes by fungi, and enhanced analytical tools to characterize the impact of
these enzymes on the lignocellulose substrate, the development of more efficient bioprocesses
can be developed. The following is a summary of some of the analytical and molecular biology
tools that have recently been used to increase our understanding of lignocellulose bioconversion
by fungi.
2.7.1 Analytical Characterization of Wood Fibre
Degradation patterns associated with advanced stages of white-rot and brown-rot decay can be
identified macroscopically and microscopically. However, a precise analysis of the effect of
degradation requires chemical assessment of the residual fibre. Lignin in wood is traditionally
measured by the Klason method, which is based on total acid hydrolysis of polysaccharides and
gravimetric estimation of the precipitated lignin. This, however, is a time consuming method and
not particularly suited to characterizing partially decayed or modified wood (Martínez et al.
2005). Alternatively, the direct characterization of fibre chemistry facilitates the analysis of
fungal decay, since these methods require minimal sample preparation and quantity (Stenius and
Vuorinen 1999). These methods typically use dry fibre samples and enable either surface
analysis or bulk analysis of fibre chemistry. Examples of analytical techniques for fibre surface
analysis include X-ray photoelectron spectroscopy (XPS) and secondary ion mass spectroscopy
(SIMS), as well as atomic force microscopy (AFM) and attenuated total reflectance (ATR).

16
Common analytical techniques for bulk analysis of fibre chemistry include nuclear magnetic
resonance (NMR), Fourier transform infrared (FTIR), Raman and ultraviolet/visible (UV/Vis)
spectroscopy, as well as pyrolysis gas chromatography (Py-GC) (Stenius and Vuorinen 1999).
Additionally, microscopic methods, including transmission electron microscopy (TEM), have
been used to detect morphological changes in wood cell walls that occur during wood decay. In
the present thesis, FTIR, ToF-SIMS and TEM were used to analyze residual fibre composition
and anatomy, and so these methods will be further described below.
ToF-SIMS is a powerful technique that provides chemical information about a solid surface by
ionizing the sample surface with a primary ion beam and determining the mass-to-charge ratio of
the secondary ion fragments (Benninghoven 1994). ToF-SIMS has been used to characterize
pulp fibre (Rowell 2005), as well as spatially resolve different components on the surface of
wood, providing compositional information across cell walls (Jung et al. 2010). Further, Kangas
et al., (2004) compared different types of fines separated from thermomechanical pulp based on
the chemical composition as determined by ToF-SIMS (Kangas and Kleen 2004). In another
study, Saito et al., (2008) used ToF-SIMS to investigate the distribution of trace elements and
lignin in contiguous rings to highlight differences between sapwood and heartwood of
Chamaecyparis obtusa. ToF-SIMS imaging was also used to conclude that the ray parenchyma
cells play a role in defining the state of elements during the transition from sapwood to
heartwood (Saito et al. 2008). Another application of this technology has been to determine the
surface distribution of lignin, carbohydrates and metal for aspen and spruce wood tissue-sections
(Tokareva et al. 2007). Additionally, effects of processing techniques like refining and pulping
on fibre surfaces have also been determined using ToF-SIMS (Fardim and Duran 2003).
Importantly, in a recent publication, researchers from our own group conducted a detailed
characterization of wood samples to improve ToF-SIMs analysis of lignocellulosic materials by
substantially increasing the annotation of corresponding secondary ions (Goacher et al. 2011).
FTIR has been used to distinguish angiosperms and gymnosperms (Schultz et al. 1985),
determine the lignin, glucose and xylose content of different woods (McDonald 1976), and
characterize the variation in fibre chemistry of natural and transgenic plants (Mouille et al.
2006). FTIR has also been used to detect compositional changes in cell wall polymers as a result

17
of chemical extraction (McCann et al. 1992). FTIR is especially suited to rapid chemical
characterization of small quantities of wood with minimum sample preparation and has been
used to analyze changes in wood fibre chemistry after degradation by different brown and white-
rot fungi. The recently published investigation of spruce degradation by the brown-rot fungi G.
trabeum and P. placenta, describes for the first time, the application of FTIR imaging
microscopy coupled with multivariate analysis (Fackler et al. 2010). In their study, Fackler et al
(2010) concluded that the brown-rot decay was most significant in the outer cell wall regions in
the early stages of decay. This was accompanied by loss in glycosidic bonds and pectic
substances.
Standard wavelengths that describe chemical bonds characteristic of lignin and cellulose have
been reported (Faix 1992; Faix et al. 1991; Kacurakova et al. 2000; Kotilainena et al. 2000;
Labbe et al. 2005; Pandey and Nagveni 2007; Pandey and Pitman 2003; Schwanninger et al.
2004). Commonly, visual comparison of spectra and lignin to carbohydrate peak area ratios have
been used to characterize the activity of brown-rot and white-rot fungi on wood fibre (Pandey
and Nagveni 2007). However, the utilization of chemometric tools would allow more efficient,
complete and un-biased assessment of large sets of FTIR data.
Typically, delignification is not uniformly distributed throughout wood samples, and so bulk
chemical methods to characterize the mode of fungal decay (i.e., selective or simultaneous) can
be misleading. Conventional SEM and TEM have been used as effective tools to confirm the
ability of fungi to degrade wood cell walls, and specific morphological features observed during
selective and simultaneous wood decay have been established. Simultaneous white-rot results in
thinning of wood cell walls along the circumference of the lumen, while selective white-rot leads
to gradual delignification of middle lamella and cell walls without any observable changes in cell
morphology (Blanchette and Reid 1986; Blanchette et al. 1985; Ruel et al. 1981; Ruel et al.
1986). Visualization of lignin by staining wood samples with either Bromine or KMnO4 has
greatly enhanced the capability of TEM to decipher the type and extent of wood decay caused by
microbes (Fromm et al. 2003; Saka et al. 1978). Notably, an extensive review on the utility of
microscopic techniques to characterize wood biodegradation has been previously published
(Daniel 1994; Daniel 2003).

18
2.7.2 Biochemical Characterization of Wood-degrading Fungi
2.7.2.1. Techniques to Measure Fungal Growth.
Monitoring fungal growth is often used to assess degradation potential. Because fungal
mycelium is difficult to separate from solid substrates like wood, rapid methods for measuring
fungal growth are by necessity indirect, and measure either constituents of fungal cells or factors
relating to their metabolic processes (Gottlieb and van Etten 1964). These indirect methods
however, have their own limitations. While visual inspection is the simplest method to assess
fungal growth, poor visibility of hyphae limits its application for detecting small differences.
Additionally, hyphae can be obscured by substrate and living mycelium can not be differentiated
from the dead (Boyle and Kropp 1992). In their study to develop methods to measure growth of
filamentous fungi on wood, Boyle et al., (1992) compared visual inspection, substrate dry weight
loss, rate of fluroescein diacetate hydrolysis, extractable protein content and chitin content of the
colonized substrate. The authors concluded that each assay measured a different aspect of the
growth and chitin gave the best measure of biomass.
There are several methods available for determination of chitin content (Ekblad and Nisholm
1996; Zamani et al. 2008). An assay of glucosamine based on acidic hydrolysis is considered
sensitive and to have low susceptibility to interfering compounds. Hydrolysis of chitin in 6 M
HCl for 16 hours at 80˚C is approximately 90 % complete and yields acetyl and glucosamine
residues. Alternative methods include freeze-drying and evaporation under reduced pressure,
which are comparatively arduous. Further, alkaline hydrolysis typically releases only 50 % of
glucosamine residues and requires numerous washings to remove excess base. However as a
cautionary note, the slight release of other sugars by HCl used in this assay can create minor
interferences during the color reaction, leading to slight over estimations of chitin concentration.
This disadvantage could be avoided by using alkaline hydrolysis, but it appears to be largely
compensated by simplicity of this method (Plassard et al. 1982).

19
2.7.2.2. Techniques to Characterize the Secreted Enzymes that Participate in Lignocellulose
Degradation.
Traditionally, biochemical assays have been used to characterize lignocellulose degrading
enzymes. These include assays that use model and often chromogenic substrates to determine
specific cellulolytic, hemicellulolytic and ligninolytic activities (Himmel et al. 1997).
Biochemical characterization of enzymes through activity assays requires previous knowledge of
expected activities, and is often limited by the availability of commercial substrates. As a result,
biochemical characterization of culture supernatant through conventional enzyme activity studies
limits the possibility to discover new enzymes and proteins, as well as novel protein
combinations that promote lignocellulose degradation.
Phanerochaete chrysosporium was the first lignocellulose degrading fungus to have its genome
sequenced (Martinez et al. 2004). Since then, the genomes of over 25 basidiomycetes have been
sequenced (http://genome.jgi-psf.org/). With the sequencing of genomes from lignocellulose
degrading microorganisms, advances in instrumentation for protein separation, and protein
identification by mass-spectrometry, new proteins that contribute to lignocellulose
transformation can now be identified (Vanden Wymelenberg et al. 2005b). For example,
proteomic analyses of Gloeophyllum trabeum grown on spruce sawdust identified secreted
enzymes and proteins that were not distinguished by previous biochemical assays of culture
supernatant (Abbas et al. 2005; Varela et al. 2003). Notably, less than 15 % of the proteins
secreted by G. trabeum had been previously identified by biochemical or sequence data. This
potential of genomic and proteomic approaches to identify new enzymes is further highlighted
by the systematic analysis of the P. placenta genome, transcriptome and proteome (Martinez et
al. 2009). While oxidases predicted to generate hemicellulases and extracellular Fe2+ and H2O2
were identified, perhaps most striking was the relatively low number of cellulolytic glycoside
hydrolases (GH) encoded by the P. placenta genome, and apparent lack of GHs containing
family 1 carbohydrate-binding modules.
Genomic and proteomic analyses of the white-rot basidiomycete Phanerochaete chrysosporium
have revealed many glycoside hydrolases and other carbohydrate-active proteins that were not
differentiated by previous biochemical assays of culture supernatant (Vanden Wymelenberg et

20
al. 2006; Vanden Wymelenberg et al. 2005a). Notably, more than 65% of the proteins secreted
by P. chrysoporium grown on crystalline cellulose corresponded to previously uncharacterized
glycoside hydrolases, or proteins with unknown function. Genomic analyses indicate that P.
chrysosporium is distinguished from other eukaryotic genomes by encoding more than twice as
many carbohydrate hydrolyzing enzymes than synthesis enzymes. Since less than 20 % of
predicted glycoside hydrolases encoded by P. chrysosporium were characterized before the
genome sequence was published, this example clearly illustrates the potential for genome
sequencing to increase the arsenal of industrially relevant enzymes (Martinez et al. 2004; Vanden
Wymelenberg et al. 2006).
As might be predicted from biomass composition, the characterization of extracellular proteins
from P. chrysosporium grown on cellulose and wood substrates consistently detect the
expression of GH3 β-glycosidases, GH6 and GH7 cellobiohydrolases, GH10 xylanases, and
GH12 endoglucanases; polygalacturonases, α-galactosidase, aspartic acid proteases, and GH88
d-4,5-unsaturated glucuronyl hydrolases are also frequently detected (Abbas et al. 2005;
Ravalason et al. 2008; Sato et al. 2007; Vanden Wymelenberg et al. 2009; Vanden Wymelenberg
et al. 2005a). Notably, lignin peroxidases and manganese peroxidases were often missed in
proteomic analyses of P. chrysosporium grown on cellulose and wood substrates, although
detectable by RT-PCR (Sato et al. 2009; Vanden Wymelenberg et al. 2009). Proteomic analyses
of P. chrysosporium have also revealed substrate dependent expression profiles of hypothetical
proteins, identified secreted proteins not predicted from initial bioinformatics analyses, and have
improved gene models for this organism (Vanden Wymelenberg et al. 2006). More recently,
pyrosequencing technology was used to sequence an expression library from P. chrysosporium
grown on oak (Sato et al. 2009). These data were consistent with previously reported proteomic
analyses, and underscore the significance of gene expression levels in microbial responses to
substrate composition.
For several years, a main focus in lignocellulose degradation research has been to isolate and
identify organisms that are hyper-producers of stable lignocellulolytic enzymes. Now, the
application of genomic and proteomic techniques can be used to reveal new enzymes from un-
cultured organisms, or enzymes that are difficult to detect using conventional biochemical

21
assays. The challenge, however, is to couple these advanced molecular techniques with
analytical methods that characterize the effects of new enzymes and enzyme mixtures on
complex, industrially relevant substrates and predict which proteins directly participate in
lignocellulose utilization.
Chapter 2 References
Abbas A, Koc H, Liu F, Tien M (2005) Fungal degradation of wood: initial proteomic analysis of extracellular proteins of Phanerochaete chrysosporium grown on oak substrate. Current Genetics 47 (1):49-56.
Ander, P. and Eriksson, K.E. (1977) Selective degradation of wood components by white-rot fungi. Physiol. Plant. 41:239-248.
Bayer EA, Lamed R, White BA, Flint HJ.(2008) From cellulosomes to cellulosomics. The Chemical Record. (6):364-77.
Béguin P, Aubert J-P (1994) The biological degradation of cellulose. FEMS Microbiology Reviews 13 (1):25-58
Benninghoven A (1994) Chemical analysis of inorganic and organic surfaces and thin films by static Time-of-Flight Secondary Ion Mass Spectrometry (TOF-SIMS). Angewandte Chemie International Edition in English 33 (10):1023-1043.
Blanchette RA (1984) Screening wood decayed by white rot fungi for preferential lignin degradation. Applied and Environmental Microbiology. 48 (3):647-653
Blanchette RA, Otjen L, Effland MJ, Eslyn WE (1985) Changes in structural and chemical components of wood delignified by fungi. Wood Science and Technology 19:35-46
Blanchette R, Reid ID (1986) Ultrastructural aspects of wood delignification by Phlebia
(Merulius) tremellosus. Appl Environ Microbiol 52:239-245 Blanchette RA (1991) Delignification by wood-decay fungi. Annual Reviews of Phytopathology
29:381-398 Blanchette, R.A., Burnes, T.A., Eerdmans, M.M. and Akhtar, M. (1992) Evaluating isolates of
Phanerochaete chrysosporium and Cerzporiopsis subz:errnispora for use in biological pulping processes. Holzforschung 46, 109-115. 645-662.
Blanchette, R.A, Obst JR, Timell, TE (1994) Biodegradation of comparession wood and tension
wood by white and brown-rot fungi. Holzforschung 48: 34-42.

22
Boyle DC, Kropp BR (1992) Development and comparison of methods for measuring growth of filamentous fungi on wood. Canadian Journal of Microbiology 38:1053-1060
Call HP, Muncke I (1997) History, overview and applications of mediated lignolytic systems, especially laccase-mediator systems (lignozyme(R)-process). Journal of Biotechnology 53:163-202
Cantarel BL, Coutinho PM, Rancurel C, Bernard T, Lombard V, Henrissat B (2009) The Carbohydrate-Active EnZymes database (CAZy): an expert resource for Glycogenomics. Nucleic Acids Research 37 (suppl 1):D233-D238.
Claus H (2004) Laccases: structure, reactions, distribution. Micron 35 (1-2):93-96
Crawford DL.(1978) Lignocellulose decomposition by selected Streptomyces strains.Appl Environ Microbiol. 35(6):1041-5.
Cullen D, Kersten PJ (2004) Enzymology and molecular biology of lignin degradation. In: Brambl R, Marzluf GA (eds) The Mycota III Biochemistry and Molecular Biology. Springer-Verlag, Berlin-Heidelberg,
d'Souza TM, Merritt CS, Reddy AC (1999) Lignin modifying enzymes of the white rot basidiomycete Ganoderma lucidum. Applied and Environmental Microbiology 65 (12):5307-5313
Daniel G (1994) Use of electron microscopy for aiding our understanding of wood biodegradation. FEMS Microbiology Reviews 13:199-233
Daniel, G. (2003) Microview of wood under degradation by bacteria and fungi.In: Wood
Deterioration and Degradation. Advances in Our Changing World. B. Goodell, D. D. Nicholas and T. P. Schultz, ACS Symposium Series. 845:34 - 72
Dashtban M, Schraft H, Qin W (2009) Fungal bioconversion of lignocellulosic residues - opportunities and perspectives. International Journal of Biological Sciences 5:578-595
Davies G, Henrissat B (1995) Structures and mechanisms of glycosyl hydrolases. Structure 3:853-859
de Vries RP, Kester HCM, Poulsen CH, Benen JAE, Visser J (2000) Synergy between enzymes from Aspergillus involved in the degradation of plant cell wall polysaccharides. Carbohydrate Research 327 (4):401-410
de Vries RP, Visser J (2001) Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiology and Molecular Biology Reviews 65 (4):497-522
Ding SY, Himmel ME. (2006) The maize primary cell wall microfibril: a new model derived from direct visualization. Journal of Agriculture and Food Chemistry.54(3):597-606.

23
Ekblad A, Nisholm T (1996) Determination of chitin in fungi and mycorrhizal roots by an improved HPLC analysis of glucosamine. Plant and Soil 178:29-35
Fackler K, Stevanic J, Ters T, Hinterstoisser B, Schwanninger M, Salmén L (2010) Localisation and characterisation of incipient brown-rot decay within spruce wood cell walls using FTIR imaging microscopy. Enzyme and Microbial Technology 47 (6):257-267
Faix O (ed) (1992) Fourier Transform Infrared Spectroscopy. Methods in lignin chemistry. Springer, Berlin-Heidelberg
Faix O, Bremer J, Schmidit O, Stevanovic T (1991) Monitoring of chemical changes in white-rot degraded beech wood by pyrolysis-gas chromatography and Fourier transform infrared spectroscopy. Journal of Analytical and Applied Pyrolysis 21:147-162
Fardim P, Duran N (2003) Modification of fibre surfaces during pulping and refining as analysed by SEM, XPS, and TOF-SIMs. Colloids and Surfaces A; Physiochemical Engineering Aspects 223:263-276
34r cc J, Rockel B, Lautner S, Windeisen E, Wanner G (2003) Lignin distribution in wood cell walls determined by TEM and backscattered SEM techniques. Journal of Structural Biology 143 (1):77-84
Global Forest Resources Assessment (2005). Food and Agriculture Organization of the United Nations, Rome
Goacher R, Jeremic D, Master ER (2011) Expanding the library of secondary ions that distinguish lignin and polysaccharides in ToF-SIMS analysis of wood. Analytical Chemistry. 83(3):804-12
Gottlieb D, Van Etten JL (1964) Biochemical changes during the growth of fungi. Journal of Bacteriology 88 (1):114-121
Green III F, Highley TL (1997) Mechanism of brown-rot decay: Paradigm or Paradox. International Biodeterioration and Biodegradation 39 (2-3):113-124
Hammel KE (1997) Fungal Degradation of Lignin. In: Cadisch G, Gillier KE (eds) Driven by Nature: Plant litter quality and decomposition. CAB International, pp 33-45
Hayashi T, Kaida R (2011) Hemicelluloses as recalcitrant components for saccharification in wood. Routes to Cellulosic Ethanol 2:45-52
Henrissat B (1991) A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem J 280 (2):309-316
Hervé C, Rogowski A, Blake A, Marcus S, Gilbert H, Knox J (2010) Carbohydrate-binding modules promote the enzymatic deconstruction of intact plant cell walls by targeting and proximity effects. Proc Natl Acad Sci U S A 107 (34):15293-15298

24
Hibbett DS, Donoghue MJ (2001) Analysis of character correlations among wood decay mechanisms, mating systems, and substrate ranges in homobasidiomycetes. Systems Biology 50:215-242
Himmel ME, Adney William S, Baker John O, Elander R, McMillan James D, Nieves Rafael A, Sheehan John J, Thomas Steven R, Vinzant Todd B, Zhang M (1997) Advanced Bioethanol Production Technologies: A Perspective. In: Fuels and Chemicals from Biomass, vol 666. ACS Symposium Series. American Chemical Society, pp 2-45.
Howard RL, Abotsi E, Jansen van Rensburg EL, Howard S (2003) Lignocellulose biotechnology: issue of bioconversion and enzyme production. African Journal of Biotechnology 2 (12):602-619
Humphreys JM, Chapple C (2002) Rewriting the lignin roadmap. Current Opinion in Plant Biology 5 (3):224-229
Jensen Jr. KA, Houtman CJ, Ryan ZC, Hammel KE (2001) Pathways for extracellular Fenton chemistry in the brown rot basidiomycete Gloeophyllum trabeum. Applied and Environmental Microbiology 67 (6):2705-2711
Jung S, Foston M, Sullards CM, Ragauskas AJ (2010) Surface characterization of dilute acid pretreated Populus deltoides by TOF-SIMS. Energy Fuels 24:1347-1357
Kacurakova M, Capek P, Sasinkova V, Wellner N, Ebringerova A (2000) FTIR study of plan cell wall model compounds: pectic polysacchardes and hemicelluloses. Carbohydrate polymers 43:195-203
Kangas H, Kleen M (2004) Surface chemical and morphological properties of mechanical pulp fines. Nordic Pulp and Paper Research Journal 19 (2):191-199
Kawai S, Umezawa T, Higuchi T (1988) Degradation mechanisms of phenolic P-1 lignin substructure model compounds by laccase of Coriolus versicolor. Archives of Biochemistry and Biophysics 262 (1):99-110
Keegstra K (2010) Plant Cell Walls. Plant Physiol 154 (2):483-486. doi:10.1104/pp.110.161240
Kirk TK, Cullen D (1998) Enzymology and molecular genetics of wood degradation by white-rot fungi. In: Young RA, Akhtar M (eds) Environmentally Friendly Technologies for the Pulp and Paper lndustry. John Wiley & Sons, Inc,
Kotilainena RA, Toivanena T-J, Aléna RJ (2000) FTIR monitoring of chemical changes in softwood during heating. Journal of Wood Chemistry and Technology 20 (3):307-320
Labbe N, Rials TG, Kelley SS, Cheng Z-M, Kim J-Y, Li Y (2005) FTIR imaging and pyrolysis-molecular beam mass spectrometry:new tools to investigate wood tissues. Wood Science and Technology 39:61-77

25
Lerouxel O, Cavalier DM, Liepman AH, Keegstra K (2006) Biosynthesis of plant cell wall polysaccharides - a complex process. Current Opinion in Plant Biology 9 (6):621-630
Levasseur A, Piumi F, Coutinho P, Rancurel C, Asther M, Delattre M, Henrissat B, Pontarotti P, Asther M, Record E (2008) FOLy: an integrated database for the classification and functional annotation of fungal oxidoreductases potentially involved in the degradation of lignin and related aromatic compounds. Fungal Genetic Biology 45 (5):638-645
Liese W (1970) Ultrastructural aspects of woody tissue disintegration as reported in wood and tree fungi by Schmidt Olaf; 2006. Ann Rev Phytopath 8:231-258
Lynd LR, Laser MS, Bransby D, Dale BE, Davison B, Hamilton R, Himmel M, Keller M, McMillan JD, Sheehan J, Wyman CE (2008) How biotech can transform biofuels. Nature Biotechnology 26 (2):169-172
Lynd LR, Weimer PJ, van Zyl WH, Pretorius IS (2002) Microbial cellulose utilization: fundamentals and biotechnology. Microbiol Mol Biol Rev 66 (3):506-577.
Lynd LR, Wyman CE, Gerngross T (1999) Biocommodity Engineering. Biotechnology Progress 15 (5):777-793
Magdalena G (2008) An evaluation of U.S. and Canadian lignocellulosic biomass supply and ethanol production costs. University of Toronto (dissertation)
Malherbe S, Cloete TE (2002) Lignocellulose biodegradation: Fundamentals and applications. Reviews in Environmental Science and Biotechnology 1 (2):105-114.
Martínez ÁT, Speranza M, Ruiz-Dueñas FJ, Ferreira P, Camarero S, Guillén F, Martínez MJ, Gutiérrez A, del Río JC (2005) Biodegradation of lignocellulosics: microbial, chemical, and enzymatic aspects of the fungal attack of lignin. International Microbiology 8:195-204
Martinez D, Challacombe J, Morgenstern I, Hibbett D, Schmoll M, Kubicek CP, Ferreira P, Ruiz-Duenas FJ, Martinez AT, Kersten P, Hammel KE, Vanden Wymelenberg A, Gaskell J, Lindquist E, Sabat G, Splinter BonDurant S, Larrondo LF, Canessa P, Vicuna R, Yadav J, Doddapaneni H, Subramanian V, Pisabarro AG, LavÃn JL, Oguiza JA, Master E, Henrissat B, Coutinho PM, Harris P, Magnuson JK, Baker SE, Bruno K, Kenealy W, Hoegger PJ, KÃes U, Ramaiya P, Lucas S, Salamov A, Shapiro H, Tu H, Chee CL, Misra M, Xie G, Teter S, Yaver D, James T, Mokrejs M, Pospisek M, Grigoriev IV, Brettin T, Rokhsar D, Berka R, Cullen D (2009) Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proceedings of the National Academy of Sciences 106 (6):1954-1959
Martinez D, Larrondo LF, Putnam N, Sollewijn Gelpke MD, Huang K, Chapman J, Helfenbein KG, Ramaiya P, Detter JC, Larimer F, Coutinho PM, Henrissat B, Berka R, Cullen D, Rokhsar D (2004) Genome sequence of the lignocellulose degrading fungus Phanerochaete chrysosporium strain RP78. Nature Biotechnology 22:695-700

26
McCann MC, Carpita NC (2008) Designing the deconstruction of plant cell walls. Current Opinion in Plant Biology 11 (3):314-320
McCann MC, Hammouri M, Wilson R, Belton P, Roberts K (1992) Fourier Transform Infrared Microspectroscopy is a new way to look at plant cell walls. Plant Physiol 100 (4):1940-1947.
McDonald RS (1976) Infrared spectrometry. Analytical Chemistry 48 (5):196-216
Moen MA, Hammel KE (1994) Lipid peroxidation by the manganese peroxidase of Phanerochaete chrysosporium is the basis for phenanthrene oxidation by the inact fungus. Applied and Environmental Microbiology. 60: 1956-61.
Mouille G, Witucka-Wall H, Bruyant M-P, Loudet O, Pelletier S, Rihouey C, Lerouxel O, Lerouge P, Hofte H, Pauly M (2006) Quantitative trait loci analysis of primary cell wall composition in Arabidopsis. Plant Physiol 141 (3):1035-1044.
Pandey KK, Nagveni HC (2007) Rapid characterisation of brown and white rot degraded chir pine and rubberwood by FTIR spectroscopy European Journal of Wood and Wood Products 65 (6):477-481
Pandey KK, Pitman AJ (2003) FTIR studies of the changes in wood chemistry following decay by brown-rot and white-rot fungi. International Biodeterioration and Biodegradation 52:151-160
Paszczynski A, Crawford RL, Blanchette RA (1988) Delignification of wood chips and pulps by using natural and synthetic porphyrins: models of fungal Decay. Applied and Environmental Microbiology 54 (1):62-68
Perez J, Munoz-Durado J, de la Rubia T, Martinez J (2002) Biodegradation and biological treatment of cellulose, hemicellulose and lignin : an overview. International Microbiology 5:53-63
Pettersen RC (1984) The Chemical Composition of Wood. Advances in Chemistry Series 207:57-126
Plassard CS, Mousain DG, Salsaca LE (1982) Estimation of mycelial growth of basidiomycetes by means of chitin determination. Phytochemistry 21 (2):345-348
Rabinovich ML, Melnik MS, Bolobova AV (2002) Microbial Cellulases- A Review. Applied Biochemical Microbiology 38:305-321
Ragauskas AJ, Williams CK, Davison BH, Britovsek G, Cairney J, Eckert CA, Frederick WJ Jr, Hallett JP, Leak DJ, Liotta CL, Mielenz JR, Murphy R, Templer R, Tschaplinski T. (2006) The path forward for biofuels and biomaterials. Science. 311(5760):484-9.
Ravalason H, Jan G, Mollé D, Pasco M, Coutinho P, Lapierre C, Pollet B, Bertaud F, Petit-Conil M, Grisel S, Sigoillot J-C, Asther M, Herpoël-Gimbert I (2008) Secretome analysis of

27
Phanerochaete chrysosporium strain CIRM-BRFM41 grown on softwood. Applied Microbiology and Biotechnology 80 (4):719-733.
Rowell RM (2005) Handbook of wood chemistry and wood composites. In. CRC Press,
Ruel K, Barnoud F, Eriksson K-E (1981) Micromorphological and ultrastructural aspects of spruce wood degradation by wild-type Sporotrichum pulverulentum and its cellulase-less mutant Cel 44. Holzforschung 25:157-171
Ruel K, Joseleau J-P, Johnsrud SC, Eriksson K-E (1986) Ultrastructural aspects of birch degradation by Phanerochaete chrysosporium and two of its cellulase deficient mutants. Holzforschung 40:5-9
Saito K, Mitsutani T, Imai T, Matsushita Y, Yamamoto A, Fukushima K (2008) Chemical differences between sapwood and heartwood of Chamaecyparis obtusa detected by ToF-SIMS. Applied Surface Science 255 (4):1088-1091
Saka S, Thomas RJ, Gratzl JS (1978) Lignin distribution: Determination by energy-dispersive analysis of X-rays. Tappi 61 (1):73-76
Sanchez C (2009) Lignocellulosic Residues - Biodegradation and bioconversion by fungi. Biotechnology Advances 27:185-194
Sato S, Liu F, Hasan K, Tien M (2007) Expression analysis of extracellular proteins from Phanerochaete chrysosporium grown on different liquid and solid substrates. Microbiology 153:3023-3033
Sato S, Feltus F, Iyer P, Tien M (2009) The first genome-level transcriptome of the wood-degrading fungus Phanerochaete chrysosporium grown on red oak. Current Genetics 55 (3):273-286.
Scheller HV, Ulvskov P (2010) Hemicelluloses. Annual Review of Plant Biology 61 (1):263-289.
Schmidit O (2006) Wood and Tree Fungi. Biology, Damage, Protection and Use. Springer-Verlag, New York
Schultz TP, Templeton MC, McGinnis GD (1985) Rapid determination of lignocellulose by diffuse reflectance Fourier transform infrared spectrometry. Analytical Chemistry 57 (14):2867-2869.
Schwanninger, M., Rodrigues, J., Pereira, H., Hinterstoisser, B. (2004) Effects of short-time vibratory ball milling on the shape of FTIR spectra of wood and cellulose. Vibrational Spectroscopy 36:23-40
Showalter AM (2001) Introduction: plant cell wall proteins. Cellular and molecular life sciences 58 (10):1361-1362

28
Sjostrom E (1983) Wood Chemistry, Second Edition: Fundamentals and Applications. Academic Press,
Stenius P, Vuorinen T (1999) Direct characterization of chemical properties of fibres. In: Sjostrom E, Alen R (eds) Analytical Methods in Wood Chemistry, Pulping, and Papermaking, vol 2. Springer -Verlag,
Taylor NG, Laurie S, Turner SR (2000) Multiple cellulose synthase catalytic subunits are required for cellulose synthesis in Arabidopsis. Plant Cell 12 (12):2529-2540.
ten Have R, Teunissen P (2001) Oxidative mechanisms involved in lignin degradation by white-rot fungi. Chemical Reviews 11:3397-3414
Tokareva EN, Fardim P, Pranovich aV, Fargerholm H-P, Daniel G, Holmbom B (2007) Imaging of wood tissue by TOF-SIMS: Critical evaluation and development of sample preparation techniques. Applied Surface Science 253:7569-7577
Vanden Wymelenberg VA, Sabat G, Martinez D, Rajangam AS, Teeri TT, Gaskell J, K KPJ, Cullen D (2005a) The Phanerochaete chrysosporium secretome: Database predictions and initial mass spectrometry peptide identifications in cellulose-grown medium Journal of biotechnology 118 (1):17-34
Vanden Wymelenberg VA, Sabat G, Martinez D, Rajangam AS, Teeri TT, Gaskell J, Kersten PJ, Cullen D (2005b) The Phanerochaete chrysosporium secretome: database predictions and initial mass spectrometry peptide identifications in cellulose-grown medium. Journal of Biotechnology 118:17-34
Vanden Wymelenberg AV, Minges P, Sabat G, Martinez D, Aerts A, Salamov A, Grigoriev I, Shapiro H, Putnam N, Belinky P, Dosoretz C, Gaskell J, Kersten P, Cullen D (2006) Computational analysis of the Phanerochaete chrysosporium v2.0 genome database and mass spectrometry identification of peptides in ligninolytic cultures reveal complex mixtures of secreted proteins. Fungal Genetics and Biology 43 (5):343-356
Vanden Wymelenberg VA, Gaskell J, Mozuch M, Kersten P, Sabat G, Martinez D, Cullen D (2009) Transcriptome and secretome analysis of Phanerochaete chrysosporium reveal complex patterns of gene expression. Appl Environ Microbiol. 75(12): 4058-68.
Varela E, Mester T, Tien M (2003) Culture conditions affecting biodegradation components of the brown-rot fungus Gloeophyllum trabeum. Archives of Microbiology 180 (4):251-256.
Walker LP, Wilson DB (1991) Enzymatic hydrolysis of cellulose: An overview. Bioresource Technology 36 (1):3-14
Watanabe Y, Shinzato N, Fukatsu T. (2003) Isolation of actinomycetes from termites' guts. Biosci Biotechnol Biochem. 67(8):1797-801.
Yelle, D. J., J. Ralph, F. C. Lu, and K. E. Hammel (2008) Evidence for cleavage of lignin by a

29
brown rot basidiomycete. Environmental Microbiology. 10:1844-1849.
Zamani A, Jeihanipour AJ, Edebo L, Niklasson C, Taherzadeh MJ (2008) Determination of glucosamine and N-acetyl glucosamine in fungal cell walls. Journal of Agricultural and Food Chemistry 56 (18):8314-8318
Zhang YH (2008) Reviving the carbohydrate economy via multi-product lignocellulose biorefineries. Journal of Industrial Microbiology Biotechnology 35 (5):367-375.

30
Chapter 3 : Effect of Cultivation Conditions on the Expression of Cellulolytic Activity by Phanerochaete species
P. chrysosporium and P. carnosa
3.1 Abstract
This chapter describes the first biochemical and growth study of the white-rot fungus
Phanerochaete carnosa. A range of simple compounds (glucose, glycerol, cellulose) and
lignocellulosic substrates (coniferous softwoods: pine, fir, spruce; and deciduous hardwoods:
maple and poplar) were validated as potential substrates for P. carnosa. In-depth characterization
of P. carnosa cultivation was pursued using microcrystalline cellulose. Culture conditions and
activity studies indicate that P. carnosa grows optimally at temperatures between 27ºC and 30ºC,
and confirmed that lignocellulolytic activity is present in extracellular filtrates of cultures grown
on cellulose. Low nitrogen (2.5 mM), stationary cultures led to highest growth measurements
(as determined by chitin content) and final protein concentration in culture supernatant.
However, while high nitrogen condition (25 mM) was consistently correlated to low specific
cellulase and MnP activity, shaking at 150 rpm did not appear to significantly effect the secretion
of lignocellulose-degrading enzyme in P. carnosa.
3.2 Introduction
Efficient biotransformation of wood components is critical for the production of renewable
energy and chemicals from this abundantly available resource. Fungi are the major decomposers
of higher plants, and the largest group of wood-degrading fungi is the Basidiomycetes. The terms
brown-rot and white-rot have been used to classify two types of decay by these fungi. White-rot
fungi enzymatically degrade all major components of wood (i.e., hemicellulose, cellulose and
lignin). By contrast, brown-rot fungi typically initiate rapid degradation of cellulose and
hemicellulose through Fenton chemistry, and modify the residual lignin without substantial
depolymerization. A different type of decay that is similar in macroscopic appearance to brown-
rotted wood is caused by soft-rot fungi. This form of wood decay is usually important in
extremely wet environments, or environments that undergo frequent wet-dry cycles, where more
aggressive wood degrading fungi cannot survive. Of all these, the white-rot fungi are known to
cause most wide-spread and significant decay (Eriksson 1990; Otjen et al. 1987).

31
Within the white-rot fungi, there are selective and simultaneous degraders, that is, fungi that
preferentially degrade lignin before attacking cellulose and hemicellulose (e.g. Ceriporiopsis
subvermispora) and those that degrade all components at similar rates (e.g. Trametes versicolor),
respectively. The fungi that can selectively remove lignin without extensive cellulose
degradation are of prime interest specifically for the pulping process as well as bioremediation
due to their established ability to degrade the recalcitrant lignin.
Phanerochaete chrysosporium is commonly used to study the production and regulation of
lignocellulolytic enzymes by white-rot fungi. This model white-rot fungus belongs to the wood
inhabiting Agaricomycete class of the Basidiomycota phylum. There are over 90 species in this
genus that are currently described all over the world (Wu et al. 2010). P. chrysosporium was
first isolated from wood chip piles and was given the name Chrysosporium lignorum (Bergman
and Nilsson 1966). This name was later changed to Sporotichum pulverulentum and finally the
present name was assigned to it based on its perfect state (the stage at which spores are formed
after nuclear fusion). The fungus is characterized as thermotolerant and has an optimum growth
rate at around 38-39˚C (Eriksson 1978).
Phanerochaete carnosa is a comparatively poorly characterized member of the family
Phanerochaete sensu strictum, and was previously isolated from balsam fir (Abies balsamea) as
well as other softwoods (de Koker et al. 2003). The phylogenetic analysis reported by de Koker
et al. (2003) groups P. carnosa with P. arizonica, P. burtii and P. sanguinea, and reveals that the
ribosomal ITS sequences of P. carnosa and P. chrysosporium share 89% identity. However, the
isolation of P. carnosa being primarily from softwoods is different from other well-studied
white-rot fungi like P. chrysosporium, which were primarily isolated from hardwoods.
Accordingly, it is possible that P. carnosa has evolved to express unique lignocellulolytic
activities that effectively transform softwoods, which would constitute a valuable source of
enzymes, given the typical recalcitrance and abundance of this biomass feedstock.
Since the cultivation and biochemical characterization of P. carnosa has not been described, the
primary aim of this first study was to establish growth conditions that induce the expression of
lignocellulolytic enzyme by this organism. This information will provide a basis for further
investigation of lignocellulose conversion by P. carnosa to utilize and enhance its potentially

32
unique lignocellulolytic activity. Moreover, by comparing the effect of cultivation conditions on
the production of lignocellulolytic activity by P. carnosa and P. chrysosporium, differences in
enzyme activities and their regulation can be evaluated.
Given the phylogenetic similarity of P. carnosa and P. chrysosporium, growth conditions that
promote lignocellulolytic activity in P. chrysosporium were predicted to also promote
lignocellulolytic activity by P. carnosa. Therefore, in order to establish the growth conditions
and investigate the lignocellulolytic enzyme profiles of P. carnosa in the present study,
published literature on P. chrysosporium was used as a basis for defining initial experimental
conditions.
3.3 Review of P. chrysosporium Cultivation and Lignocellulolytic Activity
General maintenance and sporulation of P. chrysosporium has been achieved using potato
dextrose agar (PDA). Tien and Kirk (1988) reported a medium containing 1% glucose, 1% malt
extract, 0.2% peptone, 0.2% yeast extract, 0.1%, aspargine, 0.2% KH2PO4, 0.1% MgSO4.7H2O,
0.0001% thiamine, and 2% agar for maintenance and sporulation. In P. chrysosporium, cellulose
degradation occurs during primary metabolism, while lignin mineralization has been
characterized as a secondary metabolic event that is triggered by carbon, nitrogen or sulfur
starvation (Jeffries et al 1981; Kirk and Farrell 1987).
The P. chrysosporium genome sequence reveals that this organism encodes at least 10 LiPs, 5
MnPs and 6 copper radical oxidases, in addition to several other gene products that might
contribute to lignin degradation, including glyoxal oxidases, (Martinez et al. 2004). Growth
conditions that trigger the production of lignin degrading enzymes by P. chrysosporium were
recently reviewed (Singh and Chen 2008). To summarize, the expression of lignin-degrading
enzymes by submerged liquid cultures of P. chrysosporium is promoted by limiting carbon,
nitrogen and/or sulfur availability, as well as increasing organic acid (α-hydroxy acids) and trace
element concentration. Increased agitation, temperature and pH appeared to reduce lignin
degrading activity. The effect of agitation was more pronounced in large flasks with high
shaking, which was predicted to generate shear forces that can denature secreted enzymes. It has
also been proposed that enzyme inactivation may occur due to higher level of dissolved oxygen

33
in agitated cultures than in stationary cultures. This has been attributed to the higher level of
active oxygen content in the culture which can oxidize, fragment, and cross-link proteins (Wolff
et al. 1986). Further, high oxygen concentrations can inhibit fungal growth at low glucose
concentrations (Leisola et al. 1984).
Decades worth of studies now mean that P. chrysosporium is typically cultivated between pH
4.2-6.2 (optimal below 6.0) at 25-39˚C (optimal 37˚C) in either stationary or shaking cultivations
(up to 300 rpm; optimal below 200 rpm). The growth medium is also amended with 0.02%
ammonium tartarate and 1% glucose or glycerol, as well as trace metals including calcium,
manganese, iron, zinc, copper and magnesium. Notably, adding detergents such as Tween 80 or
non-reactive foam-stabilizing agents such as polyethylene glycol (PEG) to the culture medium
can also increase lignolytic activity in culture supernatants (Asther et al. 1987; Venkatadri and
Irvine 1993). Moreover, the highest LiP production of 3,800 U/ml was obtained by
immobilizing fungal mycelia in polyurethane foam, whereas the highest MnP production
observed was 1,375 U/ml with solid-state fermentation using steam-exploded straw as substrate.
Some effective repressors of lignin-degrading enzyme production are glutamic acid, glutamine
and ammonium ion.
While the effect of cultivation conditions on the expression of lignolytic activity by P.
chrysosporium was recently reviewed, corresponding effects on the expression of cellulolytic
activities are more dispersed in the published literature. Similar to lignolytic enzymes,
carbohydrate active enzyme expression is highly dependent on the nitrogen and carbon source
supplied to the culture medium, pH, temperature, and presence of inhibitors (Table 3.1 and Table
3.2). For instance, early studies of white-rot fungi indicated that cellulases are induced in the
presence of cellulosic substrates, and that cellulase expression is regulated through catabolite
repression (Eriksson 1978; Jensen 1971; Johansson 1966; Smith and Gold 1979). These studies
employed viscometric measurements to determine endoglucanase activity on
carboxymethylcellulose, and colorimetric assays to measure glucosidase activity on p-
nitrophenyl-D-glycopyranosides. In general, early biochemical analyses of P. chrysosporium
confirmed that cellulose is a good substrate to investigate cellulolytic and hemicellulolytic
activity by this fungus (Bao et al. 1994; Eriksson and Rzedowski 1969). Early studies also

34
determined that asparagine is an effective nutrient source and that aeration with oxygen enhances
the activity of the cellulase systems (Ander and Eriksson 1977; Levonen-Munoz and Bone
1984). No clear impact of agitation on cellulolytic activity has been reported, and the pH
optimum for cellulolytic activity in basidiomycetes is typically between 3.5 to 5.5 and the
temperature optima are between 35-50˚C (Baldrian and Valaskova 2008; Eriksson 1978; Smith
and Gold 1979; Eriksson and Hamp 1978).
More recently, the effect of substrate, inducing compounds, and cultivation conditions on
lignocellulolytic expression by P. chrysosporium has been evaluated using molecular techniques,
including quantitative PCR and proteomic methods. PCR is particularly useful for monitoring
microbial activity when corresponding enzymes are not easily accessible or distinguished by
activity alone. One of the early studies to use PCR to investigate P. chrysosporium activity grew
the fungus on four different carbon sources: ball-milled straw, microcrystalline cellulose
(Avicel), high glucose concentration (2 %), and low glucose concentration (0.2 %); all under
nitrogen limiting conditions (0.23 g/L) (Broda et al. 1995). This research revealed differential
expression of cellulase gene families, depending on the substrate and stage of cultivation.
Specifically, the expression of genes that encode lignin-degrading and cellulose-degrading
enzymes was observed on straw and microcrystalline cellulose, suggesting lignocellulose-
dependent expression of these enzymes (Broda et al. 1995).
RT-PCR was also used to investigate the regulation of LiPs and MnPs by P. chrysosporium,
(Janse et al. 1998), and several more recent studies have used RT-PCR to identify inducers and
repressors of lignocellulolytic genes. For example, Suzuki et al., (2008) used real-time
quantitative RT-PCR to investigate carbon catabolite repression in P. chrysosporium. They
concluded that cellobiohydrolase genes (cel6A, cel7d and cdh) were drastically influenced by
increased levels of glucose concentration, whereas no significant changes were observed in β-
glucosidase gene (bgl3A) (Suzuki et al. 2008). Recent advances in RT-PCR techniques that
improve the quantification of gene transcripts produced at low levels, were also used to quantify
transcript levels of glycoside hydrolases genes (cel6A and cel7A to cell 7F/G) in cultures
containing glucose, cellulose and cellooligosaccharides (Suzuki et al. 2009). Cellulose,
cellotriose and cellotetraose demonstrated clear upregulation of transcript levels as compared to
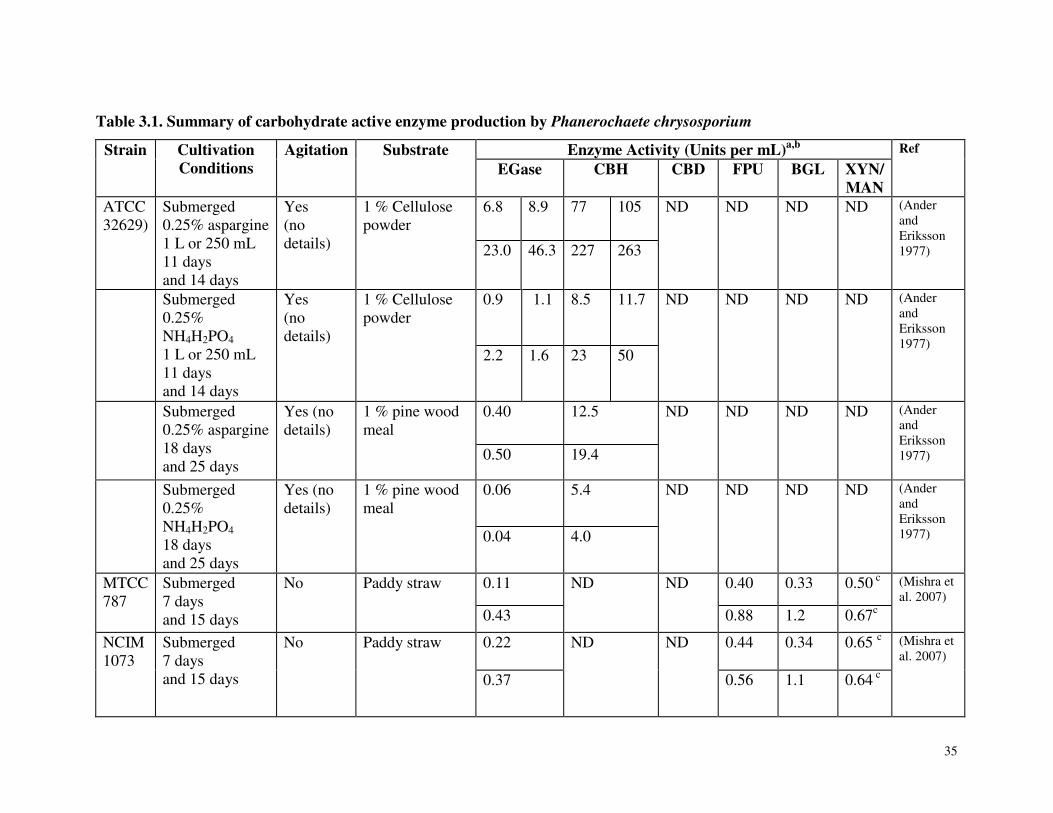
35
Table 3.1. Summary of carbohydrate active enzyme production by Phanerochaete chrysosporium
Strain Cultivation Conditions
Agitation Substrate Enzyme Activity (Units per mL)a,b Ref
EGase CBH CBD FPU BGL XYN/ MAN
ATCC 32629)
Submerged 0.25% aspargine 1 L or 250 mL 11 days and 14 days
Yes (no details)
1 % Cellulose powder
6.8 8.9 77 105 ND ND ND ND (Ander and Eriksson 1977)
23.0 46.3 227 263
Submerged 0.25% NH4H2PO4 1 L or 250 mL 11 days and 14 days
Yes (no details)
1 % Cellulose powder
0.9 1.1 8.5 11.7 ND ND ND ND (Ander and Eriksson 1977) 2.2 1.6 23 50
Submerged 0.25% aspargine 18 days and 25 days
Yes (no details)
1 % pine wood meal
0.40
12.5 ND ND ND ND (Ander and Eriksson 1977)
0.50 19.4
Submerged 0.25% NH4H2PO4 18 days and 25 days
Yes (no details)
1 % pine wood meal
0.06
5.4 ND ND ND ND (Ander and Eriksson 1977)
0.04 4.0
MTCC 787
Submerged 7 days and 15 days
No Paddy straw 0.11 ND ND 0.40 0.33 0.50 c (Mishra et al. 2007) 0.43 0.88 1.2 0.67c
NCIM 1073
Submerged 7 days and 15 days
No Paddy straw 0.22
ND ND 0.44 0.34 0.65 c (Mishra et al. 2007)
0.37 0.56 1.1 0.64 c
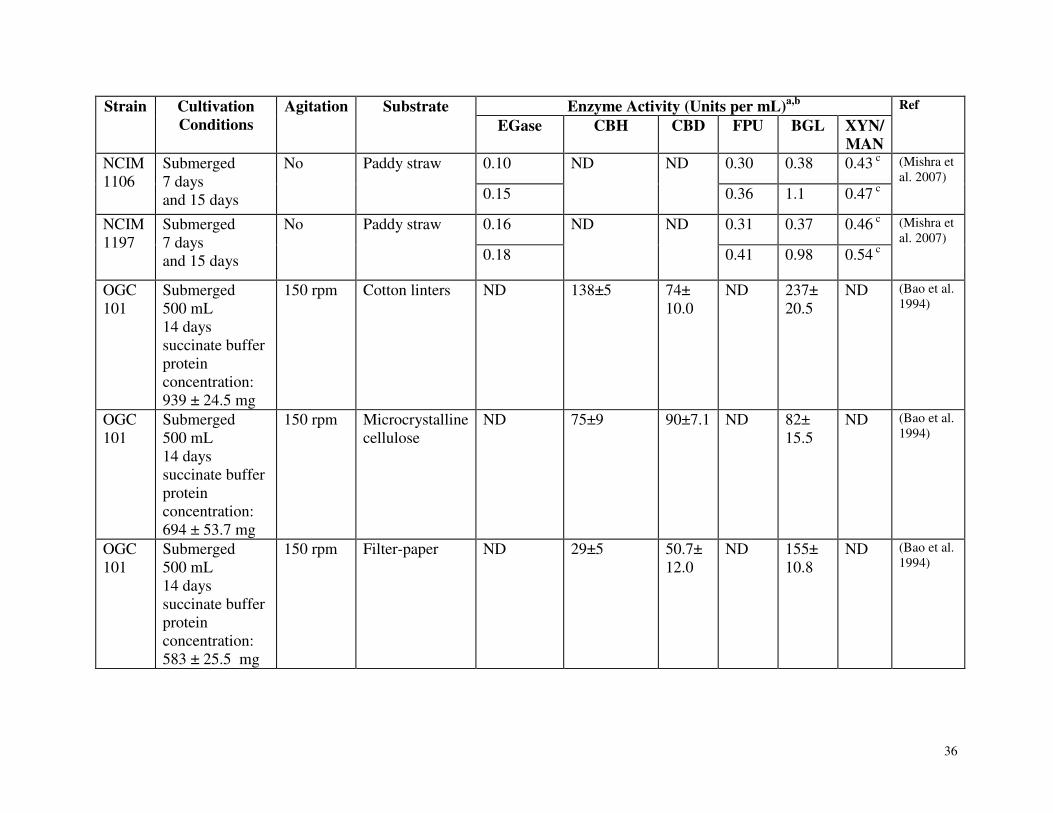
36
Strain Cultivation Conditions
Agitation Substrate Enzyme Activity (Units per mL)a,b Ref
EGase CBH CBD FPU BGL XYN/ MAN
NCIM 1106
Submerged 7 days and 15 days
No Paddy straw 0.10 ND ND 0.30 0.38 0.43 c (Mishra et al. 2007)
0.15 0.36 1.1 0.47 c
NCIM 1197
Submerged 7 days and 15 days
No Paddy straw 0.16 ND ND 0.31 0.37 0.46 c (Mishra et al. 2007)
0.18 0.41 0.98 0.54 c
OGC 101
Submerged 500 mL 14 days succinate buffer protein concentration: 939 ± 24.5 mg
150 rpm Cotton linters ND 138±5 74± 10.0
ND 237± 20.5
ND (Bao et al. 1994)
OGC 101
Submerged 500 mL 14 days succinate buffer protein concentration: 694 ± 53.7 mg
150 rpm Microcrystalline cellulose
ND 75±9 90±7.1 ND 82± 15.5
ND (Bao et al. 1994)
OGC 101
Submerged 500 mL 14 days succinate buffer protein concentration: 583 ± 25.5 mg
150 rpm Filter-paper ND 29±5 50.7± 12.0
ND 155± 10.8
ND (Bao et al. 1994)
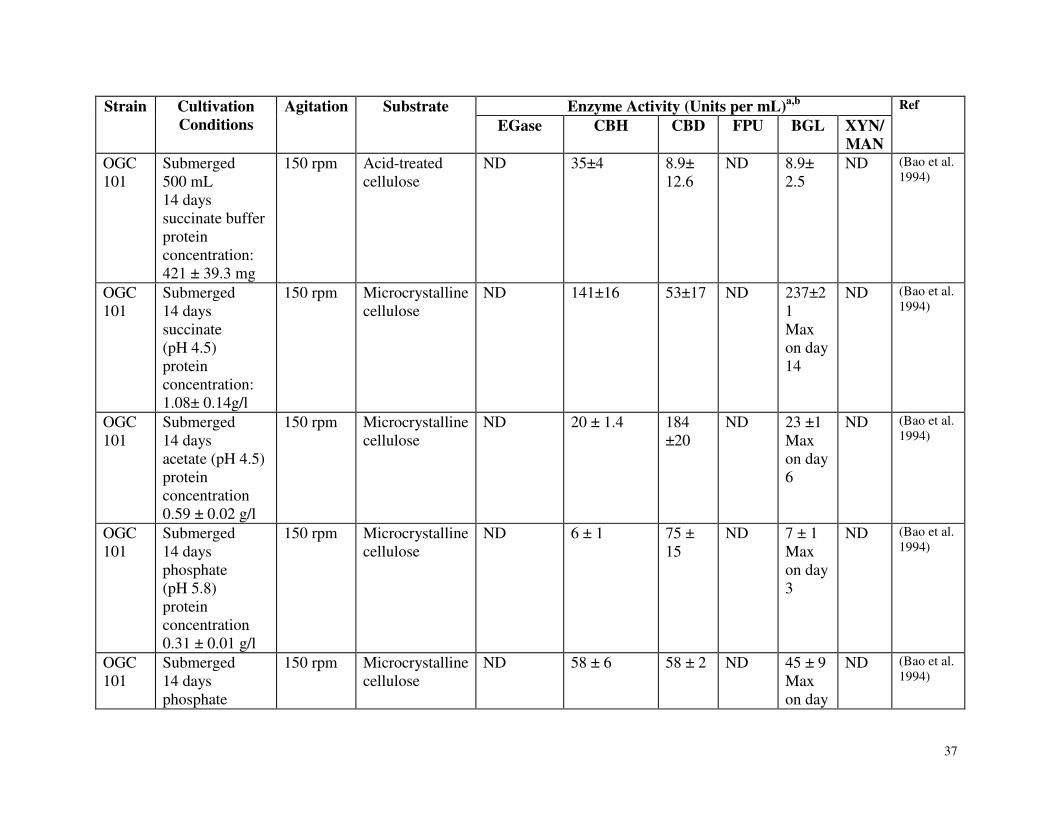
37
Strain Cultivation Conditions
Agitation Substrate Enzyme Activity (Units per mL)a,b Ref
EGase CBH CBD FPU BGL XYN/ MAN
OGC 101
Submerged 500 mL 14 days succinate buffer protein concentration: 421 ± 39.3 mg
150 rpm Acid-treated cellulose
ND 35±4 8.9± 12.6
ND 8.9± 2.5
ND (Bao et al. 1994)
OGC 101
Submerged 14 days succinate (pH 4.5) protein concentration: 1.08± 0.14g/l
150 rpm Microcrystalline cellulose
ND 141±16 53±17 ND 237±21 Max on day 14
ND (Bao et al. 1994)
OGC 101
Submerged 14 days acetate (pH 4.5) protein concentration 0.59 ± 0.02 g/l
150 rpm Microcrystalline cellulose
ND 20 ± 1.4 184 ±20
ND 23 ±1 Max on day 6
ND (Bao et al. 1994)
OGC 101
Submerged 14 days phosphate (pH 5.8) protein concentration 0.31 ± 0.01 g/l
150 rpm Microcrystalline cellulose
ND 6 ± 1 75 ± 15
ND 7 ± 1 Max on day 3
ND (Bao et al. 1994)
OGC 101
Submerged 14 days phosphate
150 rpm Microcrystalline cellulose
ND 58 ± 6 58 ± 2 ND 45 ± 9 Max on day
ND (Bao et al. 1994)
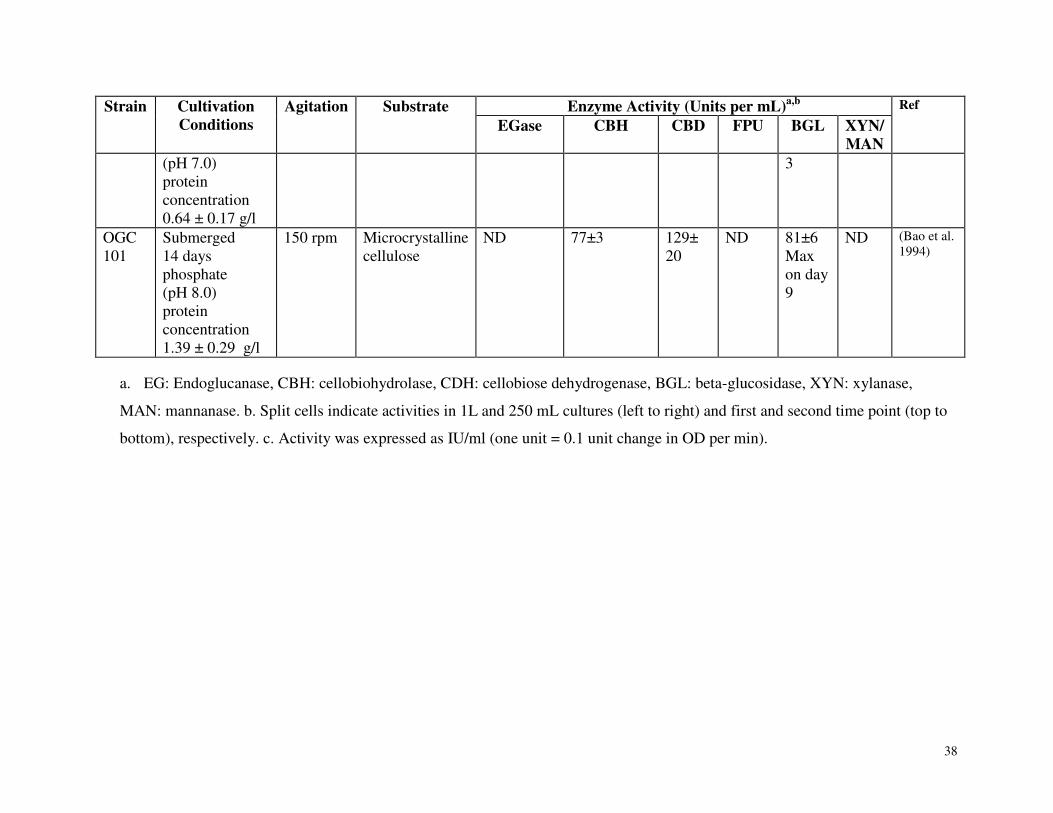
38
Strain Cultivation Conditions
Agitation Substrate Enzyme Activity (Units per mL)a,b Ref
EGase CBH CBD FPU BGL XYN/ MAN
(pH 7.0) protein concentration 0.64 ± 0.17 g/l
3
OGC 101
Submerged 14 days phosphate (pH 8.0) protein concentration 1.39 ± 0.29 g/l
150 rpm Microcrystalline cellulose
ND 77±3 129± 20
ND 81±6 Max on day 9
ND (Bao et al. 1994)
a. EG: Endoglucanase, CBH: cellobiohydrolase, CDH: cellobiose dehydrogenase, BGL: beta-glucosidase, XYN: xylanase,
MAN: mannanase. b. Split cells indicate activities in 1L and 250 mL cultures (left to right) and first and second time point (top to
bottom), respectively. c. Activity was expressed as IU/ml (one unit = 0.1 unit change in OD per min).
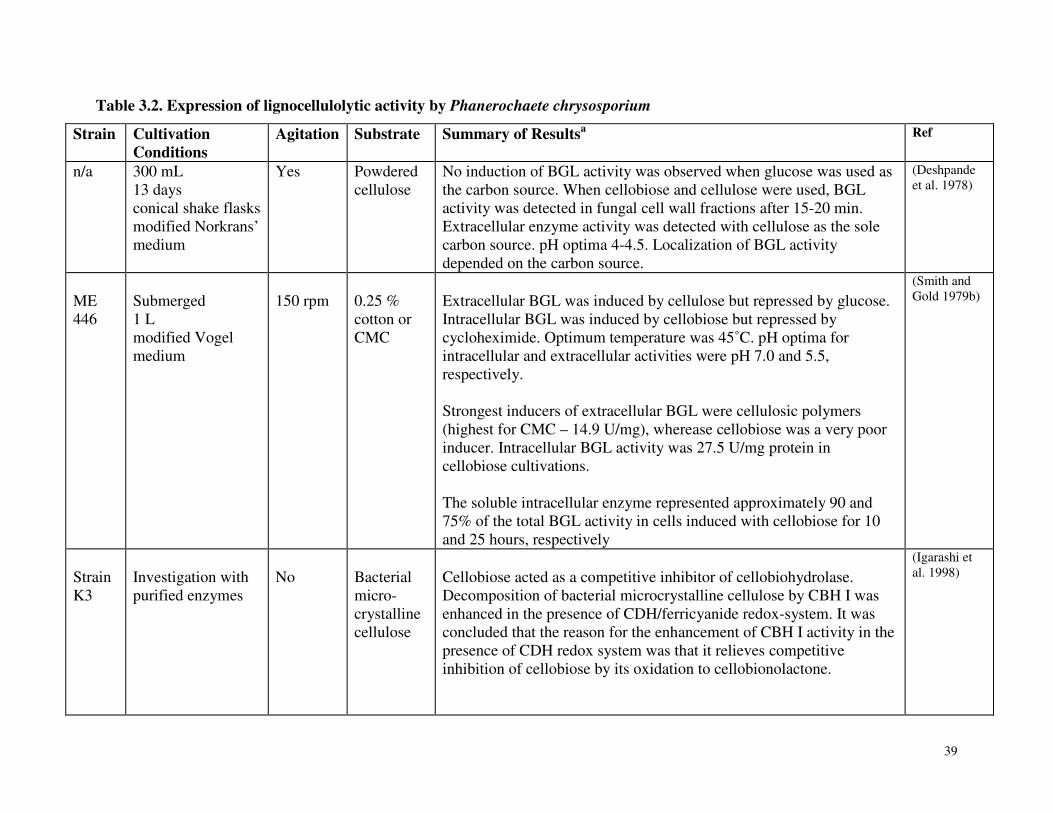
39
Table 3.2. Expression of lignocellulolytic activity by Phanerochaete chrysosporium
Strain Cultivation Conditions
Agitation Substrate Summary of Resultsa Ref
n/a 300 mL 13 days conical shake flasks modified Norkrans’ medium
Yes Powdered cellulose
No induction of BGL activity was observed when glucose was used as the carbon source. When cellobiose and cellulose were used, BGL activity was detected in fungal cell wall fractions after 15-20 min. Extracellular enzyme activity was detected with cellulose as the sole carbon source. pH optima 4-4.5. Localization of BGL activity depended on the carbon source.
(Deshpande et al. 1978)
ME 446
Submerged 1 L modified Vogel medium
150 rpm
0.25 % cotton or CMC
Extracellular BGL was induced by cellulose but repressed by glucose. Intracellular BGL was induced by cellobiose but repressed by cycloheximide. Optimum temperature was 45˚C. pH optima for intracellular and extracellular activities were pH 7.0 and 5.5, respectively. Strongest inducers of extracellular BGL were cellulosic polymers (highest for CMC – 14.9 U/mg), wherease cellobiose was a very poor inducer. Intracellular BGL activity was 27.5 U/mg protein in cellobiose cultivations. The soluble intracellular enzyme represented approximately 90 and 75% of the total BGL activity in cells induced with cellobiose for 10 and 25 hours, respectively
(Smith and Gold 1979b)
Strain K3
Investigation with purified enzymes
No
Bacterial micro-crystalline cellulose
Cellobiose acted as a competitive inhibitor of cellobiohydrolase. Decomposition of bacterial microcrystalline cellulose by CBH I was enhanced in the presence of CDH/ferricyanide redox-system. It was concluded that the reason for the enhancement of CBH I activity in the presence of CDH redox system was that it relieves competitive inhibition of cellobiose by its oxidation to cellobionolactone.
(Igarashi et al. 1998)
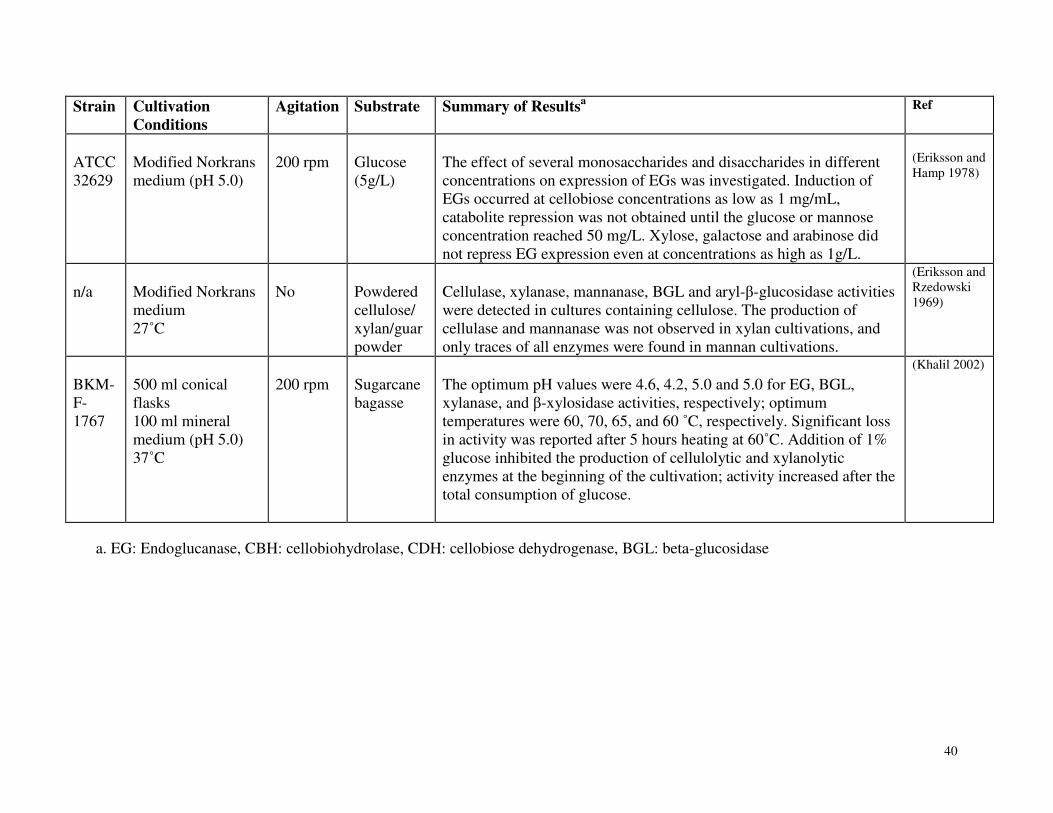
40
Strain Cultivation Conditions
Agitation Substrate Summary of Resultsa Ref
ATCC 32629
Modified Norkrans medium (pH 5.0)
200 rpm
Glucose (5g/L)
The effect of several monosaccharides and disaccharides in different concentrations on expression of EGs was investigated. Induction of EGs occurred at cellobiose concentrations as low as 1 mg/mL, catabolite repression was not obtained until the glucose or mannose concentration reached 50 mg/L. Xylose, galactose and arabinose did not repress EG expression even at concentrations as high as 1g/L.
(Eriksson and Hamp 1978)
n/a
Modified Norkrans medium 27˚C
No
Powdered cellulose/ xylan/guar powder
Cellulase, xylanase, mannanase, BGL and aryl-β-glucosidase activities were detected in cultures containing cellulose. The production of cellulase and mannanase was not observed in xylan cultivations, and only traces of all enzymes were found in mannan cultivations.
(Eriksson and Rzedowski 1969)
BKM-F-1767
500 ml conical flasks 100 ml mineral medium (pH 5.0) 37˚C
200 rpm
Sugarcane bagasse
The optimum pH values were 4.6, 4.2, 5.0 and 5.0 for EG, BGL, xylanase, and β-xylosidase activities, respectively; optimum temperatures were 60, 70, 65, and 60 ˚C, respectively. Significant loss in activity was reported after 5 hours heating at 60˚C. Addition of 1% glucose inhibited the production of cellulolytic and xylanolytic enzymes at the beginning of the cultivation; activity increased after the total consumption of glucose.
(Khalil 2002)
a. EG: Endoglucanase, CBH: cellobiohydrolase, CDH: cellobiose dehydrogenase, BGL: beta-glucosidase

41
glucose for most genes, at all time points, while cellobiose was a weak inducer of cellulase gene
transcription by comparison (Suzuki et al. 2010).
The specific objective of the current study was to establish conditions and methods to cultivate
P. carnosa, as well as monitor the growth and the profile of proteins secreted by this fungus. In
addition to enabling subsequent investigations and growth studies on wood, comparison of
optimum cultivation conditions to those defined for other wood-degrading Basidiomycetes, will
help elucidate evolutionary adaptations among the white-rot fungi.
3.4 Materials and Methods
3.4.1 Microorganism and Materials
P. carnosa strains (FP-135508-Sp, HHB-10118-Sp, RLG-7412-Sp) were obtained from the US
Department of Agriculture (USDA) Forest Products Laboratory, Madison, WI. All strains were
maintained on YMPG agar media; yeast (2 % w/v), maltose (10 % w/v), peptone (2 % w/v),
glucose (1 % w/v) at 27°C. Strain HHB-10118 was selected for detailed cultivation studies,
given its consistent growth rate and phenotype. The internal transcribed spacer (ITS) region was
sequenced to confirm the identity of the fungus (de Koker et al. 2003). Avicel (Fluka 11365),
which is a commercial preparation of wood-derived microcrystalline cellulose, and contains
approximately 30 % of amorphous structure, was used as the carbon source in liquid cultivations,
and ammonium tartarate (Sigma A4767) was used as the nitrogen source.
3.4.2 Cultivation Conditions
Liquid cultures were prepared in 200 mL B3 medium (Appendix 1) (Hon, D.N-S 1984; Jai, M.,
et. al., 1996). Liquid media were then inoculated with 0.1 mg/mL blended mycelia from 10-day
liquid YMPG cultures. Cultivation temperatures were 22°C, 27°C, 30°C, and 37°C, and cultures
were incubated for up to 18 days under stationary conditions or with shaking (150 rpm). The
impact of nitrogen concentration on the secretion of lignocellulolytic activity was tested in
stationary cultivations grown for 27 days at 27 °C with 1 % Avicel and 2.5 or 25 mM ammonium
tartrate (Table 3.2).

42
3.4.3 Growth Measurements
Chitin measurement was used to compare the extent of growth in all cultivation conditions
(Plassard et al. 1982). Briefly, three replicate cultivations were harvested after five time points
(Table 3.3), and the mycelia were separated from the culture supernatant using Mira Cloth. The
mycelia were then oven dried overnight and the final dry weight of the mycelia was recorded.
Precisely weighed amounts (~0.2 g) of each mycelial sample were transferred to a glass tube
with a screw-cap lid, incubated in 5 mL of 6 M HCI at room temperature for 3 h with the lid
open, capped and then hydrolyzed for 16 h at 80°C. Aliquots of 1 mL hydrolysate were
transferred to 5 mL of 1.25 M NaOAc to bring the pH of the solution to approximately pH 3, and
then further processed at room temperature. Specifically, 1 mL of the solution at pH 3 was mixed
with 1 mL of 5 % KHSO4 and 1 mL of 5 % NaNO2. After shaking for 5 min, the mixture was
allowed to stand for 15 min to allow for deamination, and then 1 mL of 12.5 % ammonium
sulfonate was added to release excess nitric acid (note: This step is accompanied by vigorous
mixing to prevent over-flow of the solution from reaction vial due to effervescence.) After
shaking for 5 min, 1 mL of 0.5 % MBTH (Methylbenzthiazolinone-2-hydrazone) was added. The
reaction was allowed to proceed for 1 h without any shaking. For colorimetric detection of the
anhydromannose-MBTH-FeCl3 complex at 653 nm; 1 mL of 0.5 % FeCl3 was added and the
solution was left to develop for 30 min. The absorbance was stable up to 20 h.
For protein concentration measurements, culture supernatants from the three biological replicates
prepared for each cultivation condition were pooled to obtain adequate amounts of protein for all
biochemical assays. Prior to pooling, the biological reproducibility of protein profiles was
determined using SDS-page gels. The pooled samples were then centrifuged for 20 min at 4500
rpm and concentrated at 4 °C using Pall’s Jumbosep (FD010K65) centrifugal units at a cutoff of
10 kDa. Protein concentration was measured using the Bio-rad Protein Assay according to the
manufacturer’s instructions. The absorption measurements were performed at 595 nm. A BSA
stock solution of 2 mg/ml was used to generate a standard curve from 0.05-0.5 mg/mL of
protein.

43
3.4.4 Biochemical Assays
The standard endoglucanase assay contained 50 mM sodium acetate (pH 4.5) and 0.3 wt % of
carboxymethyl cellulose (CMC). The final reaction volume was 100 µL and reactions were
incubated at 27°C for up to 1 h. The reaction products were developed using the BCA assay for
reducing ends (Doner et al. 1992), and measured at 565 nm. Glucose was used to generate a
standard curve.
β-glucosidase activity was determined by measuring the amount of p-nitrophenol released from
p-nitrophenol-β-D-glucoside (pNPG) using 0.6 % pNPG by weight in 50 mM sodium acetate
(pH 4.5) (Bowers et al. 1980). The reaction products were developed using 100 µL of 1M
Na2CO3 and measured at 410 nm; pNP (0-0.05µmol) was used to generate a standard curve.
Filter paper units (FPUs) were measured using Whatman No.1 filter paper. An office hole-
puncher was used to produce equal sized filter paper samples (~ 1 mm2), which were submerged
in 50 mM sodium acetate (pH 4.5). The total reaction volume was 200 µL and reactions were
incubated at 27°C for up to 1 hour. The reaction products were again developed using the BCA
assay, and glucose was used to generate a standard curve (Ghose 1987).
Lignin peroxidase activity was measured by monitoring oxidation of 3,5-dimethoxybenzyl
alcohol to veratraldehyde at 310 nm (Tien and Kirk 1984). An extinction coefficient of 9.3
mmol-1 cm-1 was used to determine product formation (Kapich et al. 2004). Manganese
peroxidase (MnP) activity was measured by monitoring the oxidation of ABTS at 415 nm
(Katagiri et al. 1995). The reaction mixture contained 7.8 mM ABTS, 0.1 mM MnSO4, 50 mM
sodium lactate, 0.1 mM H2O2, and 20 mM sodium succinate buffer (pH 4.5). An extinction
coefficient of 3.6 mmol-1 cm-1 was used to determine product formation. The assay was
performed in the presence and absence of MnSO4 to determine Mn-dependent and Mn-
independent activity.
All assays were performed in triplicate at the temperature and pH of the cultivation. Enzyme
activities are expressed as units (U) per ml of culture supernatant, where one unit of enzyme
activity is defined as the amount of enzyme generating 1 µmol of product per minute.
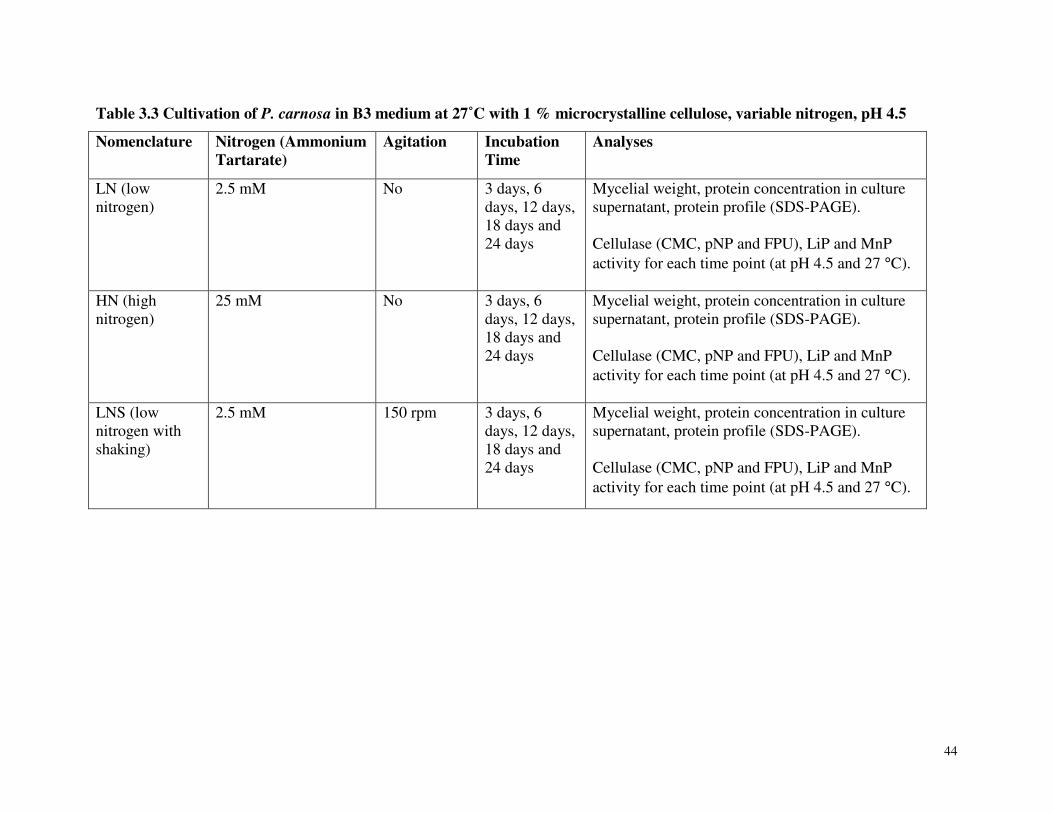
44
Table 3.3 Cultivation of P. carnosa in B3 medium at 27˚C with 1 % microcrystalline cellulose, variable nitrogen, pH 4.5
Nomenclature Nitrogen (Ammonium Tartarate)
Agitation Incubation Time
Analyses
LN (low nitrogen)
2.5 mM No 3 days, 6 days, 12 days, 18 days and 24 days
Mycelial weight, protein concentration in culture supernatant, protein profile (SDS-PAGE). Cellulase (CMC, pNP and FPU), LiP and MnP activity for each time point (at pH 4.5 and 27 °C).
HN (high nitrogen)
25 mM No 3 days, 6 days, 12 days, 18 days and 24 days
Mycelial weight, protein concentration in culture supernatant, protein profile (SDS-PAGE). Cellulase (CMC, pNP and FPU), LiP and MnP activity for each time point (at pH 4.5 and 27 °C).
LNS (low nitrogen with shaking)
2.5 mM 150 rpm 3 days, 6 days, 12 days, 18 days and 24 days
Mycelial weight, protein concentration in culture supernatant, protein profile (SDS-PAGE). Cellulase (CMC, pNP and FPU), LiP and MnP activity for each time point (at pH 4.5 and 27 °C).

45
3.5 Results
Initial growth studies performed to evaluate the range of compounds that could be used to
cultivate P. carnosa confirmed that P. carnosa grew on commercial substrates including
glycerol, glucose and microcrystalline cellulose (Avicel), as well as chips of lodgepole pine,
white spruce, balsam fir, maple and poplar, submerged in B3 medium. To characterize the
growth of P. carnosa in more detail, cultivations were prepared using Avicel as the carbon
source.
3.5.1 Effect of Cultivation Condition on the Extent of P. carnosa Growth on
Microcrystalline Cellulose.
Preliminary cultivations of P. carnosa were grown on micro-crystalline cellulose (Avicel) at
22˚C, 27°C, 30°C and 37°C. From visual inspection of cultivations and weight of collected
mycelia, it was concluded that cultures incubated at 27°C led to the highest fungal growth. The
air-dried weight of mycelia collected after 5 weeks at 22˚C, 27˚C, 30˚C and 37˚C were 0.99g,
0.95g, 0.70 and 0.74g, respectively. In contrast to P. chrysosporium, which grows optimally at
37°C, growth of P. carnosa was not visible at 37°C,
As reviewed above, nitrogen concentration and agitation affects the expression of
lignocellulolytic enzymes by P. chrysosporium. Therefore, the impact of these two parameters
on the secretion of lignocellulolytic activity by P. carnosa grown at 27°C on Avicel was
investigated (Table 3.3). P. carnosa grew well in low and high nitrogen conditions as well as in
cultivations agitated at 150 rpm. At each of the time points indicated in Table 3.3, the extent of
fungal growth was determined by protein concentrations in culture supernatants, and by
measuring chitin content (Plassard et al. 1982).
The final protein concentrations for high nitrogen (HN) and low nitrogen with shaking (LNS)
cultivations were similar, while the low nitrogen, stationary (LN) cultivations had the highest
protein concentration at the last time point (Table 3.4). After 12 days of cultivation, protein
concentrations in both HN and LNS cultivations appeared to decrease, suggesting proteolysis. By
constrast, the protein concentration in LN cultivations increased steadily over 24 days.
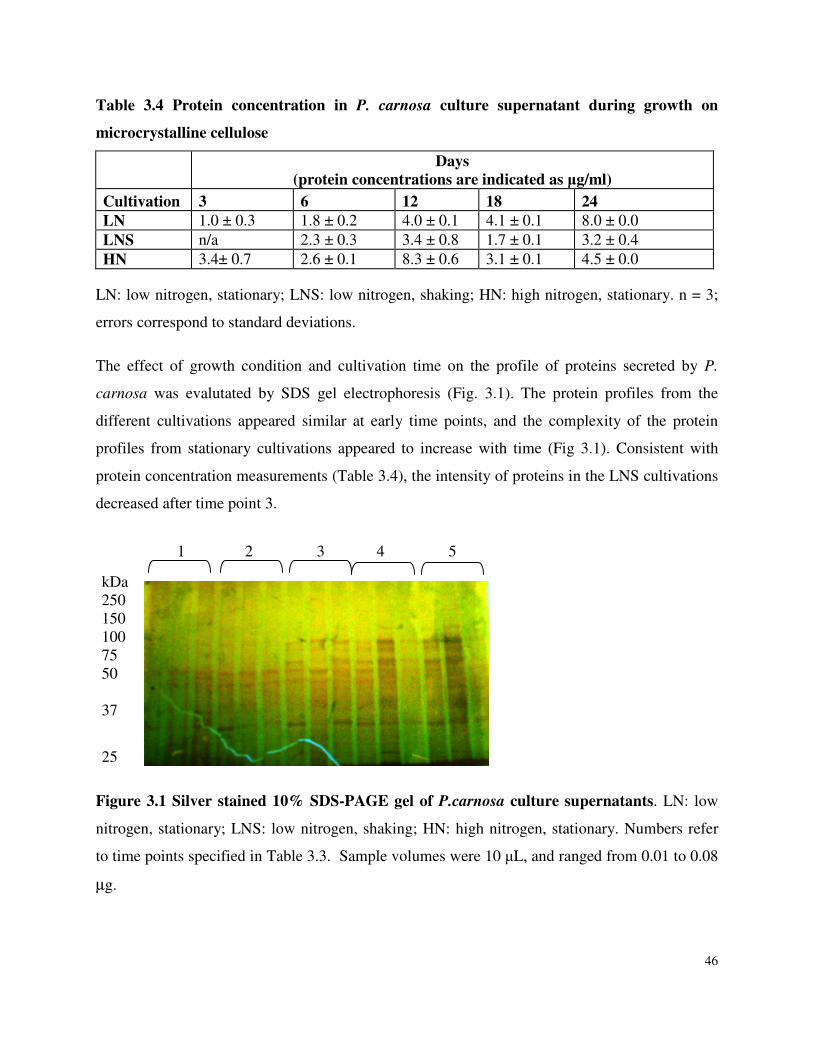
46
Table 3.4 Protein concentration in P. carnosa culture supernatant during growth on
microcrystalline cellulose
Days
(protein concentrations are indicated as µg/ml)
Cultivation 3 6 12 18 24
LN 1.0 ± 0.3 1.8 ± 0.2 4.0 ± 0.1 4.1 ± 0.1 8.0 ± 0.0 LNS n/a 2.3 ± 0.3 3.4 ± 0.8 1.7 ± 0.1 3.2 ± 0.4 HN 3.4± 0.7 2.6 ± 0.1 8.3 ± 0.6 3.1 ± 0.1 4.5 ± 0.0
LN: low nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary. n = 3;
errors correspond to standard deviations.
The effect of growth condition and cultivation time on the profile of proteins secreted by P.
carnosa was evalutated by SDS gel electrophoresis (Fig. 3.1). The protein profiles from the
different cultivations appeared similar at early time points, and the complexity of the protein
profiles from stationary cultivations appeared to increase with time (Fig 3.1). Consistent with
protein concentration measurements (Table 3.4), the intensity of proteins in the LNS cultivations
decreased after time point 3.
Figure 3.1 Silver stained 10% SDS-PAGE gel of P.carnosa culture supernatants. LN: low
nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary. Numbers refer
to time points specified in Table 3.3. Sample volumes were 10 µL, and ranged from 0.01 to 0.08
µg.
kDa 250 150 100 75 50 37
25
1 2 3 4 5

47
Indirect methods including visual inspection, substrate dry weight loss, rate of diacetate
hydrolysis, extractable protein content and chitin content have previously been used to quantify
fungal growth on substrates like wood chips, where separation between mycelia and substrate is
challenging. In a comparative study of these methods, Boyle and Kropp (1992) concluded that
chitin (a polymer of N-acetyl-p-glucosamine, the major cell wall component in fungi) is the most
suitable means of measuring fungal growth. Ergosterol, which is a fungus specific membrane
component, is another means to quantify fungal growth. While ergosterol measurements can give
an estimate of the living fungal biomass at time of harvest, chitin content is a measure of both
living and dead fungal biomass, and so can be used to estimate total growth (Ekblad and
Nisholm 1996). Literature cites several methods to determine chitin content (Ekblad and
Nisholm 1996; Francois 2006; Zamani et al. 2008). The method developed by Plassard et al.
(1982) has been used to determine chitin content in similar cultivations, and so was used in the
current study (Papinutti and Lechner 2008; Levin et al. 2008). As anticipated, low nitrogen
stationary cultivations resulted in the highest fungal growth and protein concentration. FTIR of
the mycelial samples was also investigated as a means to obtain relative mycelial growth.
However, due to the low mycelial density, the FTIR spectra of the test samples were very similar
to the controls containing Avicel alone.
While chitin was initially measured from 0.3-30 mg (dry weight) of mycelia recovered from
cellulose cultivations, the mycelial sample had to be increased to approximately 200 mg before
reliable absorption measurements could be obtained. As a result, chitin was measured in oven
dried mycelial samples collected from 24-day cultures, which corresponded to comparatively
high visible growth. Microcrystalline cellulose that remained in the 24-day P. carnosa
cultivations was aggretated with fungal mycelia, and so could not be removed prior to chitin
measurements. Notably, up to 0.8 mg of cellulose was found to contribute to the calorimetric
reading. However, since cellulose in exceess of 0.8 mg did not increase background readings,
the absorption of 0.8 mg of microcrystalline cellulose was used as a reference in the assay. Chitin
measurements were then used to estimate the growth of P. carnosa in the different cultures; this
analysis was consistent with protein concentrations measured in the culture media, where the
highest chitin content was observed for LN cultures (Table 3.5).
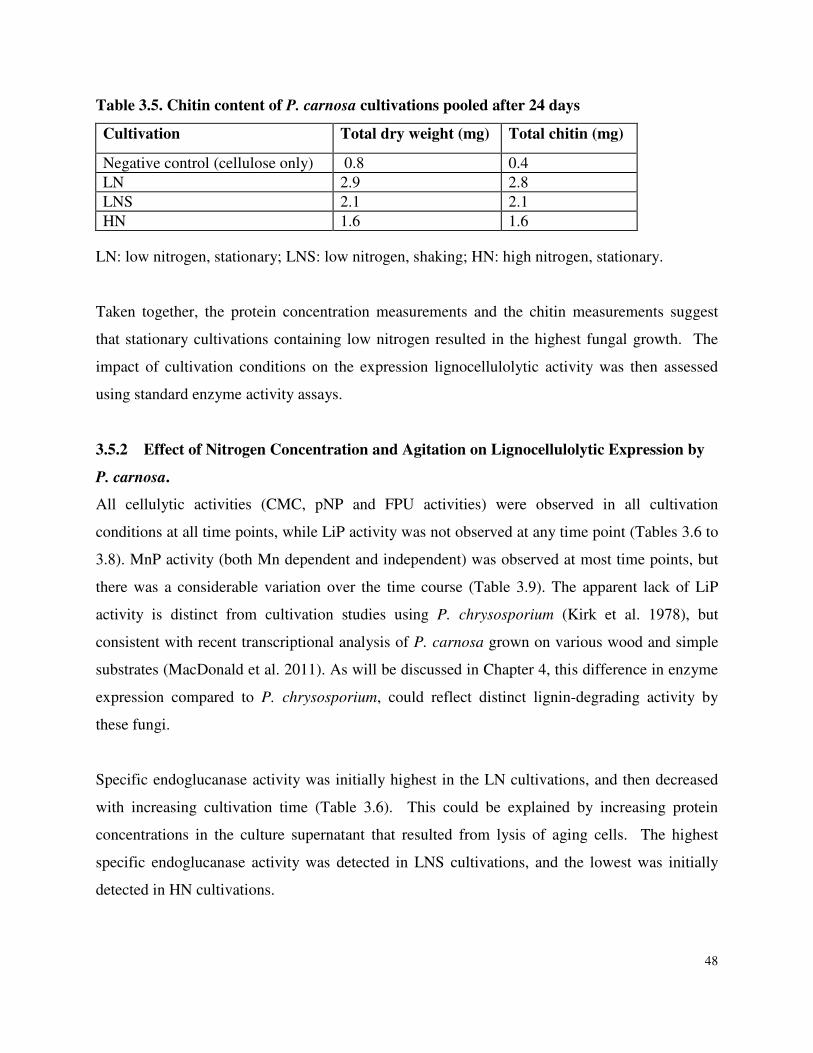
48
Table 3.5. Chitin content of P. carnosa cultivations pooled after 24 days
Cultivation Total dry weight (mg) Total chitin (mg)
Negative control (cellulose only) 0.8 0.4 LN 2.9 2.8 LNS 2.1 2.1 HN 1.6 1.6
LN: low nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary.
Taken together, the protein concentration measurements and the chitin measurements suggest
that stationary cultivations containing low nitrogen resulted in the highest fungal growth. The
impact of cultivation conditions on the expression lignocellulolytic activity was then assessed
using standard enzyme activity assays.
3.5.2 Effect of Nitrogen Concentration and Agitation on Lignocellulolytic Expression by
P. carnosa.
All cellulytic activities (CMC, pNP and FPU activities) were observed in all cultivation
conditions at all time points, while LiP activity was not observed at any time point (Tables 3.6 to
3.8). MnP activity (both Mn dependent and independent) was observed at most time points, but
there was a considerable variation over the time course (Table 3.9). The apparent lack of LiP
activity is distinct from cultivation studies using P. chrysosporium (Kirk et al. 1978), but
consistent with recent transcriptional analysis of P. carnosa grown on various wood and simple
substrates (MacDonald et al. 2011). As will be discussed in Chapter 4, this difference in enzyme
expression compared to P. chrysosporium, could reflect distinct lignin-degrading activity by
these fungi.
Specific endoglucanase activity was initially highest in the LN cultivations, and then decreased
with increasing cultivation time (Table 3.6). This could be explained by increasing protein
concentrations in the culture supernatant that resulted from lysis of aging cells. The highest
specific endoglucanase activity was detected in LNS cultivations, and the lowest was initially
detected in HN cultivations.
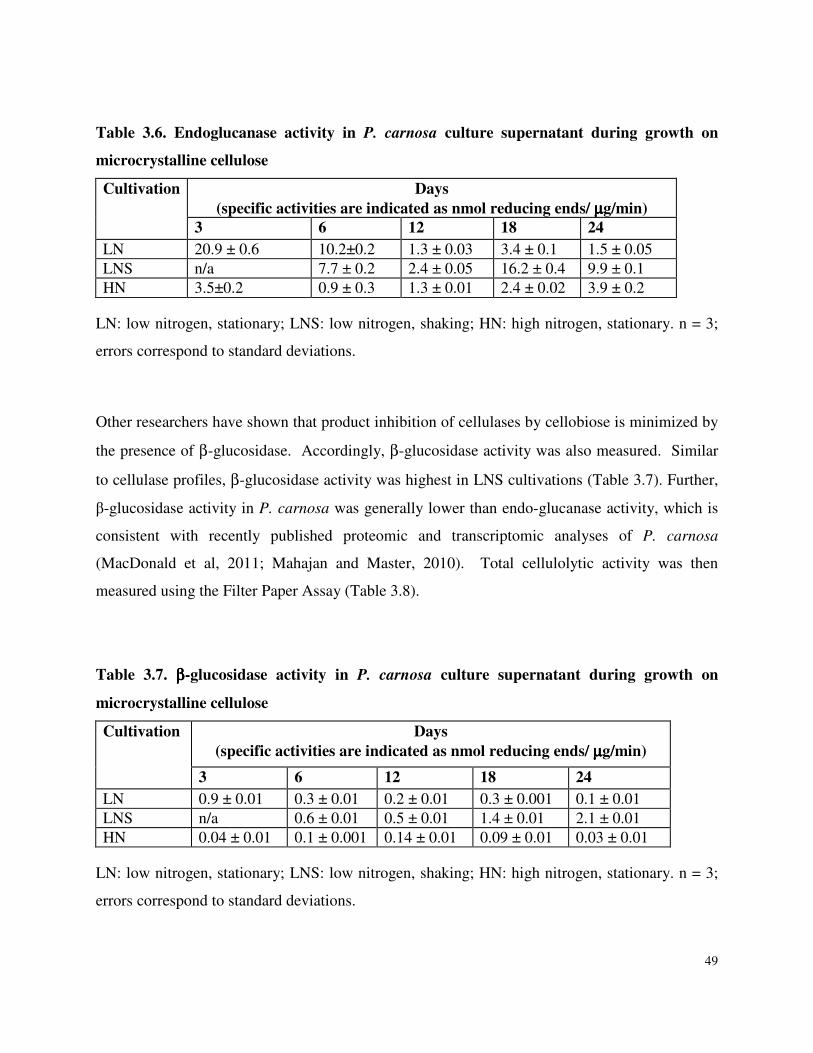
49
Table 3.6. Endoglucanase activity in P. carnosa culture supernatant during growth on
microcrystalline cellulose
Cultivation Days
(specific activities are indicated as nmol reducing ends/ µµµµg/min) 3 6 12 18 24
LN 20.9 ± 0.6 10.2±0.2 1.3 ± 0.03 3.4 ± 0.1 1.5 ± 0.05 LNS n/a 7.7 ± 0.2 2.4 ± 0.05 16.2 ± 0.4 9.9 ± 0.1 HN 3.5±0.2 0.9 ± 0.3 1.3 ± 0.01 2.4 ± 0.02 3.9 ± 0.2
LN: low nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary. n = 3;
errors correspond to standard deviations.
Other researchers have shown that product inhibition of cellulases by cellobiose is minimized by
the presence of β-glucosidase. Accordingly, β-glucosidase activity was also measured. Similar
to cellulase profiles, β-glucosidase activity was highest in LNS cultivations (Table 3.7). Further,
β-glucosidase activity in P. carnosa was generally lower than endo-glucanase activity, which is
consistent with recently published proteomic and transcriptomic analyses of P. carnosa
(MacDonald et al, 2011; Mahajan and Master, 2010). Total cellulolytic activity was then
measured using the Filter Paper Assay (Table 3.8).
Table 3.7. ββββ-glucosidase activity in P. carnosa culture supernatant during growth on
microcrystalline cellulose
Cultivation Days
(specific activities are indicated as nmol reducing ends/ µµµµg/min)
3 6 12 18 24
LN 0.9 ± 0.01 0.3 ± 0.01 0.2 ± 0.01 0.3 ± 0.001 0.1 ± 0.01 LNS n/a 0.6 ± 0.01 0.5 ± 0.01 1.4 ± 0.01 2.1 ± 0.01 HN 0.04 ± 0.01 0.1 ± 0.001 0.14 ± 0.01 0.09 ± 0.01 0.03 ± 0.01
LN: low nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary. n = 3;
errors correspond to standard deviations.
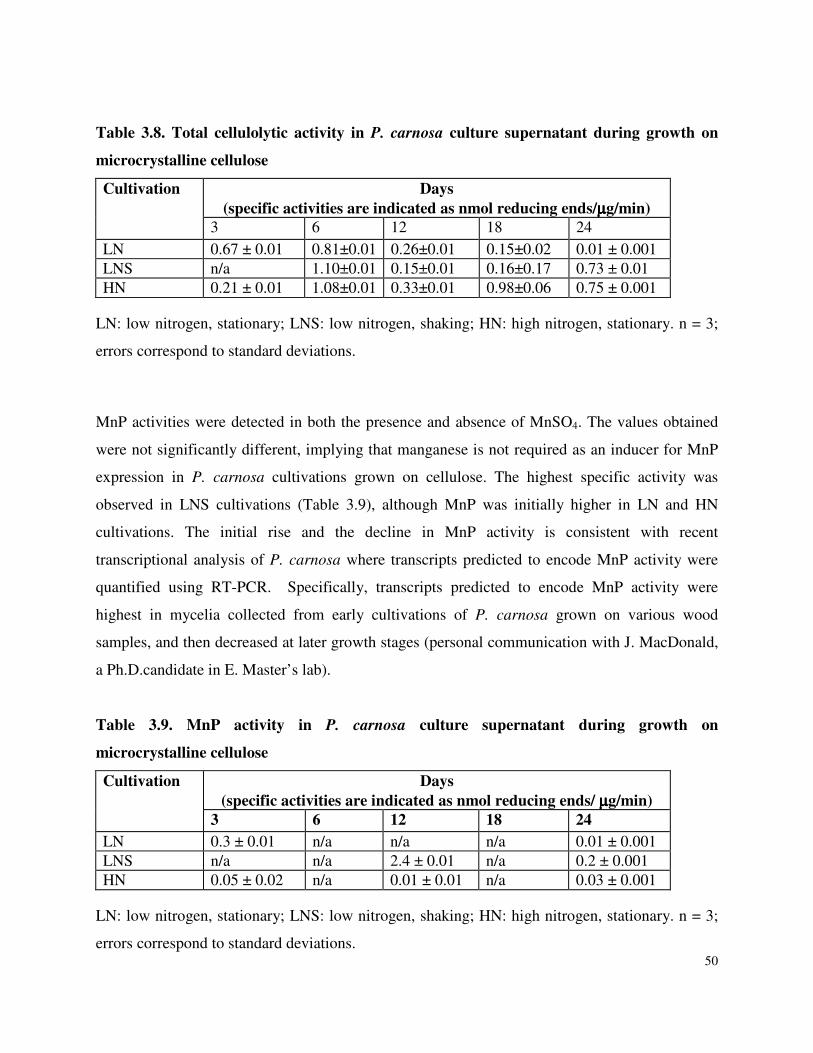
50
Table 3.8. Total cellulolytic activity in P. carnosa culture supernatant during growth on
microcrystalline cellulose
Cultivation Days
(specific activities are indicated as nmol reducing ends/µµµµg/min) 3 6 12 18 24
LN 0.67 ± 0.01 0.81±0.01 0.26±0.01 0.15±0.02 0.01 ± 0.001 LNS n/a 1.10±0.01 0.15±0.01 0.16±0.17 0.73 ± 0.01 HN 0.21 ± 0.01 1.08±0.01 0.33±0.01 0.98±0.06 0.75 ± 0.001
LN: low nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary. n = 3;
errors correspond to standard deviations.
MnP activities were detected in both the presence and absence of MnSO4. The values obtained
were not significantly different, implying that manganese is not required as an inducer for MnP
expression in P. carnosa cultivations grown on cellulose. The highest specific activity was
observed in LNS cultivations (Table 3.9), although MnP was initially higher in LN and HN
cultivations. The initial rise and the decline in MnP activity is consistent with recent
transcriptional analysis of P. carnosa where transcripts predicted to encode MnP activity were
quantified using RT-PCR. Specifically, transcripts predicted to encode MnP activity were
highest in mycelia collected from early cultivations of P. carnosa grown on various wood
samples, and then decreased at later growth stages (personal communication with J. MacDonald,
a Ph.D.candidate in E. Master’s lab).
Table 3.9. MnP activity in P. carnosa culture supernatant during growth on
microcrystalline cellulose
Cultivation Days
(specific activities are indicated as nmol reducing ends/ µµµµg/min)
3 6 12 18 24
LN 0.3 ± 0.01 n/a n/a n/a 0.01 ± 0.001 LNS n/a n/a 2.4 ± 0.01 n/a 0.2 ± 0.001 HN 0.05 ± 0.02 n/a 0.01 ± 0.01 n/a 0.03 ± 0.001
LN: low nitrogen, stationary; LNS: low nitrogen, shaking; HN: high nitrogen, stationary. n = 3;
errors correspond to standard deviations.

51
3.6 Discussion
Similar to P. chrysosporium, the cellulolytic enzyme activity expressed by P. carnosa was
lowest in HN (high nitrogen) stationary cultivations (Barclay et al. 1993). While β-glucosidase
activity in P. chrysosporium was typically higher than endoglucanase activity (Tables 3.1 and
3.2), the opposite trend was observed in P. carnosa cultivations (Tables 3.6 and 3.7). As
described in Chapter 5, comparatively low β-glucosidase activity in P. carnosa cultures is
consistent with the comparatively few peptides corresponding to GH3 enzymes detected in P.
carnosa culture supernatants.
The highest lignin degrading activity was measured in LNS cultivations using the MnP assay.
Low-nitrogen concentration also induces the expression of lignolytic activity by P.
chrysosporium, where nutrient limiting conditions are thought to induce secondary metabolism
(Kirk and Farrell 1987). Decreased MnP activity in later time points sampled from P. carnosa
cultivations was consistent with recent transcription analyses performed by J. MacDonald (a PhD
candidate in E. Master’s lab), where it was observed that MnP expression was highest during
early growth on wood samples, and then declined (personal communication with J. MacDonald).
Shaking at 150 rpm did not have a detrimental effect on lignocellulolytic activity, and in fact
increased the activities measured in this study. Notably, early studies with P. chrysosporium
revealed that agitation (rpm = 125, 150) of submerged cultures can suppress lignin degrading
activity (Kirk et al. 1978), perhaps as a result of pellet formation (Shimada 1981). However,
other studies report increased lignin degrading activity in P. chrysosporium cultivations shaking
at 150 rpm, due to increased diffusion of oxygen in the culture medium (Leisola et al. 1984).
Providing oxygen while reducing shear stress might be key (Venkatadri and Irvine 1993).
It is evident that final lignocellulolytic activity measurements were low, despite the complex
extracellular enzyme profiles observed by SDS-PAGE (Fig 3.1). This can be attributed to several
factors, including the presence of proteases or enzyme inhibitors. Based on the review of P.
chrysosporium, it is possible that changes to cultivation conditions including pH, nitrogen
source, carbon source, or addition of inducers including oligosaccharides and cellobiose oxidase,
could increase the production of lignocellulolytic enzymes by P. carnosa. Further, the addition

52
of surfactants like Tween, or organic acids is likely to impact production of lignin degrading
peroxidases. Still, it is important to remember that biochemical assays of enzyme activity are
limited to the availability of appropriate substrates, and so can underestimate the range of
lignincellulolytic activities present in culture supernatants. Accordingly, as described in
Chapters 4 and 5, further characterization of lignocellulose conversion by P. carnosa applied
analytical tools that could retrieve more detailed descriptions of enzymes secreted by P. carnosa
during growth on lignocellulosic substrates, and the corresponding impact on wood fibre
composition and structure.
Chapter 3 References
Ander P, Eriksson KE (1977) Selective degradation of wood components by white-rot fungi. Physiology Plant 41:239-248
Asther M, Corrieu G, Drapron R, Odier E (1987) Effect of Tween 80 and oleic acid on ligninase production by Phanerochaete chrysosporium INA-12. Enzyme and Microbial Technology 9 (4):245-249
Baldrian P, Valaskova V (2008) Degradation of cellulose by basidiomycetous fungi. FEMS Microbiology Reviews 32:501-521
Bao W, Lymar E, Renganathan V (1994) Optimization of cellobiose dehydrogenase and β−glucosidase production by cellulose-degrading cultures of Phanerochaete chrysosporium. Applied and Microbial Biotechnology 42:642-646
Barclay CD, Legge RL, Farquhar GF (1993) Modelling the growth kinetics of Phanerochaete
chrysosporium in submerged static culture. Appl Environ Microbiol 59 (6):1887-1892
Bergman O, Nilsson T (1966) Research Notes. As published by Erikkson et. al., 1978.
Bowers G, N. Jr., McComb RB, Christensen RC, Schaffer R (1980) High-purity 4-nitrophenol: purification, characterization and specifications for use as a spectrophotometric reference material. Clinical chemistry 26 (6):724-729
Boyle DC, Kropp BR (1992) Development and comparison of methods for measuring growth of filamentous fungi on wood. Canadian Journal of Microbiology 38 (10):1053-1060
Broda P, Birch PR, Brooks PR, Sims PF (1995) PCR-mediated analysis of lignocellulolytic gene transcription by Phanerochaete chrysosporium: substrate-dependent differential expression within gene families. Appl Environ Microbiol 61 (6):2358-2364

53
De Koker TH, Nakasone KK, Haarhof J, Burdsall jr HH, Janse BJH (2003) Phylogenetic relationships of the genus Phanerochaete inferred from the internal transcribed spacer region. Mycological Research 107 (9):1032-1040
Deshpande V, Eriksson K-EL, Pettersson B (1978) Production, purification and partial characterization of 1,4-β-glucosidase enzymes from Sporotrichum pulverulentum. European Journal of Biochemistry 90:191-198
Ekblad A, Nisholm T (1996) Determination of chitin in fungi and mycorrhizal roots by an improved HPLC analysis of glucosamine. Plant and Soil 178:29-35
Eriksson KE, Rzedowski W (1969) Extracellular enzyme systems utilized by the fungus Chrysosporium lignorum for the breakdown of cellulose. Archives of Biochemistry and Biophysics 129:683-688
Eriksson KE (1978) Enzyme mechanisms involved in cellulose hydrolysis by the rot fungus Sporotrichum pulverulentum. Biotechnology and Bioengineering 20:317-322
Eriksson KEL, Hamp SG (1978) Regulation of endo-1,4-β-glucanase production in Sporotrichum pulverulentum. European Journal of Biochemistry 90:183-190
Eriksson KEL, Blanchette RA, Ander P. (1990) Microbial and Enzymatic Degradation of Wood and Wood Components. Springer-Verlag. Berlin, Germany.
Francois JM (2006) A simple method for quantitative determination of polysaccharides in fungal cell walls. Nature Protocols 1 (6): 2995-3000
Ghose TK (1987) Measurement of cellulase activities. Pure and Applied Chemistry 59:257-268
Hibbett D (2007) After the gold rush, or before the flood? Evolutionary morphology of mushroom-forming fungi (Agaricomycetes) in the early 21st century. Mycology Research 111:1001-1018
Igarashi K, Sampy JD, Eriksson K-EL (1998) Cellobiose dehydrogenase enhances Phanerochaete chrysosporium cellobiohydrolase I activity by reliveing product inhibition. European Journal of Biochemistry 253:101-106
Janse BJH, Gaskell J, Akhtar M, Cullen D (1998) Expression of Phanerochaete chrysosporium genes encoding lignin peroxidases, manganese peroxidases, and glyoxal oxidase in wood. Appl Environ Microbiol 64 (9):3536-3538
Jeffries TW, Choi S, Kirk TK (1981) Nutritional regulation of lignin degradation by Phanerochaete chrysosporium. Appl Environ Microbiol.42(2):290-6.
Jensen KF (1971) Cellulolytic enzymes of Stereum gausapatum. Phytopathology 61:134-138
Johansson M (1966) A comparison between the cellulolytic activity of white and brown rot fungi. I. The activity on insoluble cellulose. Physiology Plant 19:709-722

54
Khalil AI (2002) Production and characterization of cellulolytic and xylanolytic enzymes from the ligninolytic white-rot fungus Phanerochaete chrysosporium grown on sugarcane bagasse. World Journal of Microbiology and Biotechnology 18:753-759
Kirk TK, Schlutz E, Connors WJ, Lorenz LF, Zeikus JG (1978) Influence of culture parameters on lignin metabolism by Phanerochaete chrysosporium. Archives of Microbiology 117:277-285
Kirk TK, Farrell RL (1987) Enzymatic 'combustion': The microbial degradation of lignin. Annual Reviews Microbiology 41:465-505
Leisola MSA, Ulmer DC, Fiechter A (1984) Factors affecting lignin degradation in lignocellulose by Phanerochaete chrysosporium. Archives of Microbiology 137 (2):171-175.
Levin L, Herrmann C, Papinutti VL (2008) Optimization of lignocellulolytic enzyme production by the white-rot fungus Trametes trogii in solid-state fermentation using response surface methodology. Biochemical Engineering Journal 39 (1):207-214
Levonen-Munoz E, Bone DH (1984) Effect of different gas environments on bench-scale solid state fermentation of oat straw by white-rot fungi. Biotechnology and Bioengineering 27:382-387
MacDonald J, Doering M, Canam T, Gong Y, Guttman D, Campbell M, Master ER (2011) Transcriptomic analysis of the softwood-degrading white-rot fungus Phanerochaete
carnosa reveals concerted gene expression responses to growth on coniferous and deciduous wood. Applied and Environmental Microbiology. [Epub ahead of print]
Martinez D, Larrondo LF, Putnam N, Sollewijn Gelpke MD, Huang K, Chapman J, Helfenbein KG, Ramaiya P, Detter JC, Larimer F, Coutinho PM, Henrissat B, Berka R, Cullen D, Rokhsar D (2004) Genome sequence of the lignocellulose degrading fungus Phanerochaete chrysosporium strain RP78. Nature Biotechnology 22:695-700
Mishra BK, Pandey AK, Lata (2007) Lignocellulolytic enzyme production from submerged fermentation of paddy straw. Indian Journal of Microbiology 47:176-179
Otjen L, Blanchette R, Effland M, Leathem G (1987) Assessment of 30 white rot basidiomycetes for selective lignin degradation. Holzforschung. 41 (6):343-349
Papinutti L, Lechner B (2008) Influence of the carbon source on the growth and lignocellulolytic enzyme production by Morchella esculenta strains. Journal of Industrial Microbiology and Biotechnology 35 (12):1715-1721.
Plassard CS, Mousain DG, Salsaca LE (1982) Estimation of mycelial growth of basidiomycetes by means of chitin determination Phytochemistry 21 (2):345-348

55
Shimada M, Nahatsubo F, Kirk TK, Higuchi T (1981) Biosynthesis of the secondary metabolite veratryl alcohol in relation to lignin degradation in Phanerochaete chrysosproium. Archives of Microbiology. 129: 321-324
Singh D, Chen S (2008) The white-rot fungus Phanerochaete chrysosporium: conditions for the production of lignin-degrading enzymes. Applied Microbiology and Biotechnology 81:399-417
Smith MH, Gold MH (1979) Phanerochaete chrysosporium ß-glucosidases: Induction, cellular localization, and physical characterization. Applied and Environmental Microbiology 37 (5):938-942
Suzuki H, Igarashi K, Samejima M (2008) Real-time quantitative analysis of carbon catabolite derepression of cellulolytic genes expressed in the basidiomycete Phanerochaete
chrysosporium. Applied Microbiology and Biotechnology 80 (1):99-106
Suzuki H, Igarashi K, Samejima M (2009) Quantitative transcriptional analysis of the genes encoding glycoside hydrolase family 7 cellulase isozymes in the basidiomycete Phanerochaete chrysosporium. FEMS Microbiology Letters 299 (2):159-165.
Suzuki H, Igarashi K, Samejima M (2010) Cellotriose and cellotetraose as inducers of the genes encoding cellobiohydrolases in the Basidiomycete Phanerochaete chrysosporium. Appl Environ Microbiol 76 (18):6164-6170.
Vanden Wymelenberg VA, Sabat G, Martinez D, Rajangam AS, Teeri TT, Gaskell J, K KPJ, Cullen D (2005) The Phanerochaete chrysosporium secretome: Database predictions and initial mass spectrometry peptide identifications in cellulose-grown medium Journal of Biotechnology 118 (1):17-34
Venkatadri R, Irvine RL (1993) Cultivation of Phanerochaete chrysosporium and production of lignin peroxidase in novel biofilm reactor systems: Hollow fiber reactor and silicone membrane reactor. Water Research 27 (4):591-596
Wolff SP, Garner A, Dean RT (1986) Free radicals, lipids and protein degradation. Trends in Biochemical Sciences 11 (1):27-31
Zamani A, Jeihanipour AJ, Edebo L, Niklasson C, Taherzadeh MJ (2008) Determination of glucosamine and N-acetyl glucosamine in fungal cell walls. Journal of Agricultural and Food Chemistry 56 (18):8314-8318

56
Chapter 4 : Mode of Coniferous Wood Decay by the White Rot Fungus Phanerochaete carnosa as Confirmed by FT-IR
and ToF-SIMS
4.1 Abstract
The softwood degrading white-rot fungus, Phanerochaete carnosa was investigated for its ability
to degrade three coniferous woods: balsam fir, white spruce and lodgepole pine. P. carnosa
grew similarly on these three wood species, and like the hardwood-degrading white-rot fungi
Ceriporiopsis subvermispora and Phanerochaete chrysosporium BKM-F-1767, P. carnosa
demonstrated selective degradation of lignin, as observed by FTIR and ToF-SIMS analyses.
Lignin degradation across-the-wall was determined by the ratio of lignin concentration between
distinct cell wall regions as measured by TEM and by ToF-SIMS, and was shown to be uniform.
This study evaluates the ability of a white-rot fungus to utilize softwood lignin and reveals the
industrial potential of the lignocellulolytic activity elicited by this fungus.
4.2 Introduction
Basidiomycetes are among the most important degraders of lignocellulosic biomass (Lundell et
al. 2010). The two most common modes of wood degradation by basidiomycetes are brown-rot
and white-rot decay. Brown-rot decay is characterized by extensive degradation of cellulose and
hemicellulose with only limited modification of lignin. Examples of well-studied brown-rot
basidiomycetes include Gloeophyllum trabeum, Serpula lacrymans, and Postia placenta
(Hogberg et al. 2006; Jensen et al. 2001; Martinez et al. 2009). These fungi appear to initiate the
depolymerization of wood polysaccharides using Fenton chemistry, whereby diffusible hydroxyl
radicals penetrate the wood surface, leaving the lignin as a modified residue (Jensen et al. 2001;
Yelle et al. 2008). By contrast, white-rot fungi secrete numerous glycoside hydrolases,
carbohydrate esterases and lignin-degrading peroxidases that target all of the main components
of wood, including cellulose, hemicellulose and lignin, leaving residual wood with a bleached
appearance (Hatakka 1994; Kersten and Cullen 2007; Tuor et al. 1995).

57
Certain white-rot fungi, such as Trametes versicolor, appear to simultaneously degrade lignin
and polysaccharides, while other white-rot fungi, including Ceriporiopsis subvermispora, appear
to selectively degrade lignin and then only slowly hydrolyze cellulose (Blanchette 1991;
Blanchette et al. 1997; Fackler et al. 2006; Pandey and Pitman 2003). The ability of fungi to
selectively degrade lignin has been used to pretreat fibre and defibrillate wood in the production
of mechanical pulp (Fackler et al. 2006; Scott et al. 2002).
While many white-rot fungi characterized to date have been isolated from hardwoods (Hibbett
and Donoghue 2001), Phanerochaete carnosa, studied here, was isolated almost exclusively
from softwood species (Burdsall 1985). As boreal forests comprise more than 45% of the North
America’s forested area and softwoods are comparatively recalcitrant sources of lignocellulosic
biomass (Birdsey et al. 2006), the glycoside hydrolases and lignin-degrading peroxidases
produced by P. carnosa constitute an important source of industrially relevant enzymes. The
secretome and transcriptome of the P. carnosa grown on lignocellulosic substrates were recently
described, and show that similar sets of enzymes are elicited by P. carnosa grown on different
lignocellulosic substrates although to different expression levels, suggesting that adaptations to
degrade lignocellulose might result from fine-tuning the expression of similar enzyme families
(Mahajan and Master 2010; MacDonald et al., 2011). Although the production of
lignocellulolytic activities by P. carnosa has been studied, the pattern of wood decay by this
fungus over time is not known. By characterizing the progression of chemical and morphological
changes of softwood decayed by P. carnosa, the aim of the current study was to predict whether
the expression of secreted hemicellulases and cellulases likely follows initial lignin degradation
by this organism. That is, to determine if P. carnosa can be classified as a selective or
simultaneous degrader.
Previous reports describe several analytical methods that can be used to investigate anatomical
and chemical changes introduced in wood fibres upon microbial decay. Electron microscopy
techniques, including scanning electron microscopy (SEM) and transmission electron
microscopy (TEM) have been used to determine micro-morphological degradation patterns in
plant cell walls (Blanchette 1991). Lignin-specific dyes such as phloroglucinol and safranin are
used for the visualization of lignin by light microscopy (Hakala et al. 2004). Changes in fibre

58
chemistry have been monitored directly using Fourier transform infrared spectroscopy (FTIR),
nuclear magnetic resonance spectroscopy (NMR), pyrolysis mass spectrometry (Pyr-MS), ion
chromatography, and thermochemolysis (Blanchette 1995; Davis et al. 1994; Faix et al. 1991;
Irbe et al. 2006; Kelley et al. 2002). FTIR is especially suited to rapid chemical characterization
of small quantities of wood with minimum sample preparation and has been used to analyze
changes in wood fibre chemistry after degradation by different brown and white-rot fungi (Faix
et al. 1991; Pandey and Nagveni 2007; Pandey and Pitman 2003). For example, Pandey and
Pitman (2003) use FTIR to analyze the chemical composition of pine (softwood) and beech
(hardwood) following treatment with the brown-rot Coniophora puteana, the non-selective
white-rot T. versicolor and the selective white-rot P. chrysosporium. In their study, brown-rot
decay was indicated by a relative increase in the lignin-to-carbohydrate peak area ratio, selective
white-rot decay was demonstrated by a relative decrease in lignin and xylan content, and the
simultaneous decay led to similar reduction in carbohydrate and lignin peaks.
Time-of-flight secondary ion mass spectrometry (ToF-SIMS) is another versatile technique that
has previously been applied to characterize surfaces of pulp, fibres and wood samples (Rowell
2005). In addition to parallel analysis of organic and inorganic constituents of wood fibre, ToF-
SIMS provides chemical images with a mass spectrum at each pixel. As a result, ToF-SIMS is
capable of spatially resolving different components on the surface of wood, providing
information about the composition across cell walls (Jung et al. 2010). ToF-SIMS analysis of
lignin and carbohydrate polymers in wood had been limited to a few wood peaks assigned in
literature until the recent identification of approximately 40 additional confirmed lignocellulose-
descriptive peaks through ToF-SIMS analysis of pine species (Goacher et al. 2011).
The current study investigates the effect of conifer species and stage of degradation on the mode
of softwood decay by P. carnosa. Specifically, P. carnosa was grown on wood chips prepared
from white spruce, lodgepole pine and balsam fir. These three wood species are not only of
prime commercial value in the North American boreal forests, but they also comprise common
mixed feedstocks for pulp and paper industries. Specifically, FTIR was employed to monitor
temporal changes in lignocellulose composition resulting from fungal activity. To evaluate the
mode of degradation across cell walls and regions of lignin degradation (i.e. cell wall or middle

59
lamella), TEM was then used to determine the localization of lignin prior and post-decay.
Finally, ToF-SIMS analysis was performed to simultaneously describe the chemical composition
and spatial resolution of lignocellulose components and to validate results obtained using FTIR
and TEM.
4.3 Materials and Methods
4.3.1 Fungal Strain and Cultivation Conditions
Phanerochaete carnosa strain HHB-10118-sp was obtained from the US Department of
Agriculture (USDA) Forest Products Laboratory, Madison, WI, and was maintained on YMPG
medium containing 2 g yeast extract, 10 g malt extract, 2 g peptone, 10 g glucose, 2 g KH2PO4, 1
g MgSO4·7H2O, 1 g asparagine per 1 L in H2O. Wood cultivations were prepared by using a
Waring blender to grind balsam fir (Abies balsamea), lodgepole pine (Pinus contorta), white
spruce (Picea glauca), and then sifting air-dried samples through US standard mesh size 6 (3.35
mm2) and 14 (1.5 mm2) sieves. Wood particles that were retained on mesh size 14 were
recovered, and 4 g samples were transferred to a total of sixty (3 species x 5 time points x 3 test
replicates and one control) 500 mL beakers containing 10 mL of B3 buffer (Appendix 1; Kirk et
al. 1978). Thiamine-HCl, ammonium tartrate, and 2,2- dimethylsuccinic acid were added as
filter-sterilized solutions, while all other media were steam sterilized for 20 to 30 min.
Each culture medium was inoculated with an 11 mm circular agar plug taken from the growing
edge of P. carnosa cultivated on solid YMPG. Cultivations were incubated under stationary
conditions at 27 °C until the diameter of the surface mycelial mat was 1 cm, 4 cm, 6 cm and 8
cm; the final sample was taken after 12 weeks of cultivation. At each growth point, a metal cutter
with 2.8 cm diameter was used to isolate residual wood particles from the centre of each
cultivation. Wood samples were rinsed with milli-Q water to remove mycelia and were air dried
before analysis.
4.3.2 Fourier Transform Infrared Spectroscopy
A minimum of three wood slivers from each time point for each wood species were randomly
chosen and powdered using a bench-top ball mill (WIG-L-BUG model, Dentsply Rinn). A small
amount of the milled sample (~ 1 mg) was mixed with KBr (200 mg) and the mixture pelletized

60
using a die (1.3 cm diameter) and a hydraulic press. A Bruker Tensor 27 FTIR was used to
record the absorbance between 4000 and 400 cm-1 with a resolution of 4 cm-1. The spectra
representing the average of 32 scans were corrected for atmospheric vapor compensation;
baseline was corrected using the rubber band method (Opus software, v. 5.0). Spectra were
normalized for unit-vector and mean-centred prior to the principal component analysis (PCA) in
Matlab v. 7.8.0 software.
The following compounds were run as standards to assist lignocellulose FTIR assignments for
unreported peaks: microcrystalline cellulose (Avicel PH-101), tamarind xyloglucan, birchwood
xylan, arabinogalactan, guar (galactomannan), pectin, and mannose were obtained from Sigma-
Aldrich; konjac glucomannan was obtained from Megazyme, and Klason lignin was prepared by
Dr. D. Jeremic (PDF in E. Master’s lab) from white spruce according to TAPPI standards (T222
om-83). The FTIR spectra were obtained and analyzed as described above.
4.3.3 Transmission Electron Microscropy - Energy-Dispersive X-ray Analysis (TEM-
EDXA)
Lignin is usually visualized with TEM-EDXA by labeling lignin with KMnO4 or bromine
(Sjöström 1983; Stein et al. 1992). In this study, six late decay wood samples (time point 5) and
six early decay wood samples (time point 1) from spruce cultivations were brominated following
the protocol of Saka et al (1978). The brominated samples were then embedded in low viscosity
embedding medium (Modification E) (Spurr 1968). Thin sections (100 nm) of the embedded
samples were obtained by using a Leica EM UC 6 microtome, and Diatome Ultra 35˚ knife (MC
12734), and analyzed using an HD-2000 transmission electron microscope (Hitachi High
Technologies America, Inc) equipped with an Oxford Instruments Inca X-sight detector.
4.3.4 Time of Flight Secondary Ion Mass Spectroscopy
Since samples observed under TEM-EDXA showed very similar patterns of lignin distribution
across the cell walls, cross-section surfaces of a fibre sample from each wood species
representing growth points one and five were prepared using a microtome (Leica EM UC 6).
ToF-SIMS spectra of these cross-sections were collected by Dr. R. Goacher (PDF in C. Mims
and E. Edwards’ lab) using a ToF-SIMS IV (IonTof, Muenster, Germany). 25 keV Bi3+ primary

61
ions were utilized to generate high mass resolution spectra in the bunched mode (0.2 pA pulsed
current) and high spatial resolution images in burst alignment mode. Charge compensation was
applied with a low energy electron beam. Base vacuum was 5 × 10-9 mbar. Spectra were
calibrated to the CH3+, H3O
+, C2H3+ and C3H5
+ ions using IonSpec v.4.1 software. At m/z 91, the
mass resolution (M/∆M) was ~5,000 in bunched mode and ~250-300 in burst alignment mode;
depending on sample roughness. To determine local variations in degradation, one spot on each
sample (100×100 to 200×200 µm2) was analyzed and split into three to four regions of interest to
generate replicates. Prior to PCA, peak intensities were Poisson-corrected to compensate for
minor detector saturation (Stephan et al. 1994). A peak list consisting of 36 definitive
lignocellulosic m/z ratios was employed for the analyses in both spectral and spatial models.
These peaks comprised groups 1 and 2 of Goacher et al. (2011), minus peaks at m/z 19 and 31
due to suspicion of moisture effects and at m/z 45, 59, 73, 131, and 147 because of PDMS
(polydimethylsiloxane) interferences. ToF-SIMS high mass resolution spectra were normalized
to unit area and mean-centred. ToF-SIMS images were mean-centred without normalization.
4.3.5 Statistical Analysis
PLS Toolbox v. 5.8.2 (Eigenvector Research, Inc.) was used with MATLAB v. 7.8.0 (The
MathWorks, Inc.) to perform PCA of spectra obtained from FTIR and ToF-SIMS. MIA Toolbox
(Eigenvector Research, Inc.) was used for PCA analysis of ToF-SIMS nominal mass resolution
ion images. PCA loadings were analyzed to determine the specific lignocellulose components
that changed significantly over time.
4.4 Results
Radial extension of fungal mycelia has been used to measure growth and compare growth
profiles on solid substrates. Previous reports also describe that visual estimates of fungal growth
on wood chips are consistent with indirect methods including substrate dry weight loss, rate of
diacetate hydrolysis, or extractable protein content and chitin content, where separation of
mycelia and substrate is challenging (Boyle and Kropp 1992; Marin et al. 2005). To monitor
growth in the present study, radial extension of P. carnosa from the agar plug was measured, and
wood samples from the centre of each cultivation were harvested at pre-defined radial
measurements. The time required to reach each radial extension was similar between cultivation

62
replicates and across wood species, indicating a similar growth progression on all the three
species selected (Appendix 2, Fig. S4.1). Gradual darkening of wood chips was observed in all
cultivations. This darkening has been correlated to chemical changes due to lignin decay (Fackler
et al. 2007; Fackler et al. 2003). These decay products and compositional changes induced in the
material were investigated using FTIR and ToF-SIMS spectral analysis and the abundance of
lignin in different cell wall layers was monitored by TEM-EDXA and ToF-SIMS imaging.
4.4.1 FTIR Spectroscopy
The PCA of wood FTIR spectra in the fingerprint region (800-1800 cm-1) clearly separated the
advanced decayed samples from the controls (Fig. 4.1 to 4.3). Considering the natural variability
of wood samples, Hotelling’s T-square statistics were used to initially identify samples from the
same time point and species that generated scans with high leverage as well as residual variance
(i.e., dangerous outliers). Despite the removal of the obvious outliers, the initial principal
component (PC1) of the analysis of spruce and fir identified variability among the data due to
sample origin.
PC1 of pine and subsequent PCs of fir and spruce (PC2 and PC3, respectively) revealed
progressive lignin decay by P. carnosa (Fig. 4.1 to 4.3). All control samples exhibited relatively
high peak intensity at ~1512 and ~1262 cm-1 as determined by PCA (Table 4.1). The 1512 cm-1
peak corresponds to aromatic skeletal vibrations of lignin in softwoods (Faix and Beinhoff 1988;
Pandey and Pitman 2003). The PCA loading at 1262 cm-1 does not correspond to an actual peak
in the spectra, but rather the difference in the slope of the peak with 1268 cm-1 maxima (guaiacyl
ring breathing). It has been previously shown that the 1263 cm-1 peak appears in a first derivative
IR spectra of lignin, confirming its suggested relation to 1268 cm-1 peak (Faix 1992). Therefore,
for all of the examined species, the significant aromatic assignments for lignin were
distinguished by PCA as higher in control samples than the decayed samples.
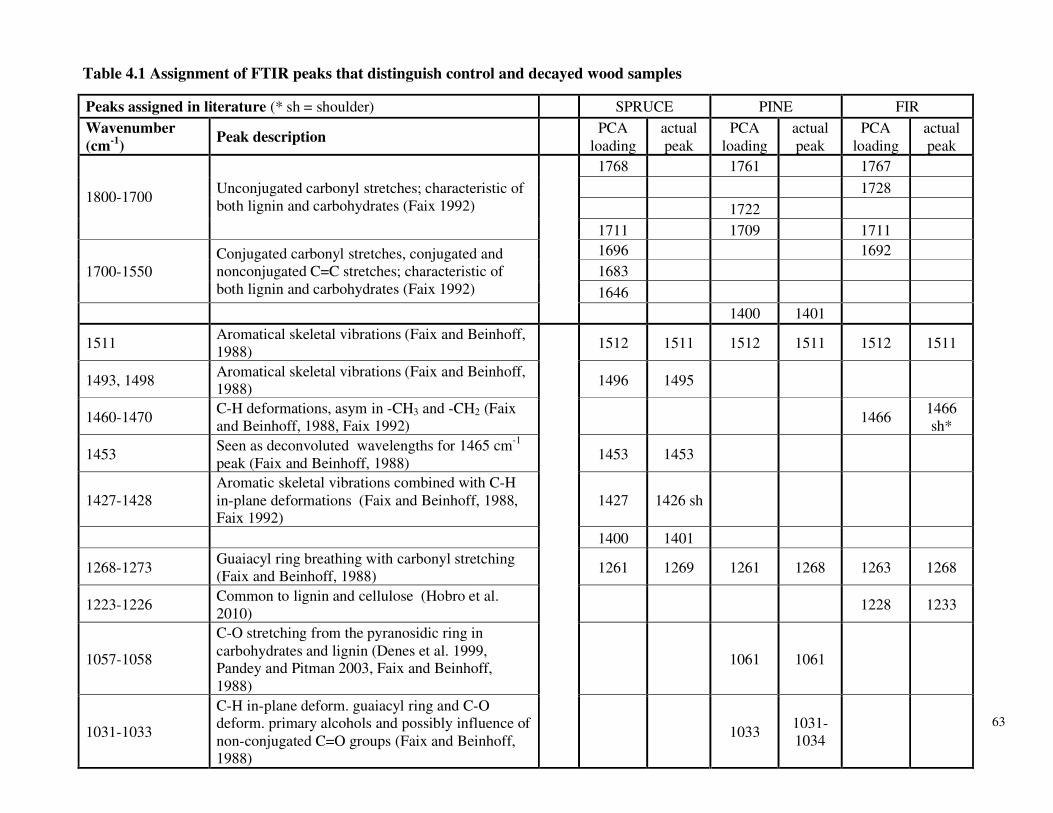
63
Peaks assigned in literature (* sh = shoulder) SPRUCE PINE FIR
Wavenumber (cm-1)
Peak description PCA
loading actual peak
PCA loading
actual peak
PCA loading
actual peak
1768 1761 1767
1728
1722 1800-1700
Unconjugated carbonyl stretches; characteristic of both lignin and carbohydrates (Faix 1992)
1711 1709 1711
1696 1692
1683 1700-1550 Conjugated carbonyl stretches, conjugated and nonconjugated C=C stretches; characteristic of both lignin and carbohydrates (Faix 1992) 1646
1400 1401
1511 Aromatical skeletal vibrations (Faix and Beinhoff, 1988)
1512 1511 1512 1511 1512 1511
1493, 1498 Aromatical skeletal vibrations (Faix and Beinhoff, 1988)
1496 1495
1460-1470 C-H deformations, asym in -CH3 and -CH2 (Faix and Beinhoff, 1988, Faix 1992)
1466 1466 sh*
1453 Seen as deconvoluted wavelengths for 1465 cm-1 peak (Faix and Beinhoff, 1988)
1453 1453
1427-1428 Aromatic skeletal vibrations combined with C-H in-plane deformations (Faix and Beinhoff, 1988, Faix 1992)
1427 1426 sh
1400 1401
1268-1273 Guaiacyl ring breathing with carbonyl stretching (Faix and Beinhoff, 1988)
1261 1269 1261 1268 1263 1268
1223-1226 Common to lignin and cellulose (Hobro et al. 2010)
1228 1233
1057-1058
C-O stretching from the pyranosidic ring in carbohydrates and lignin (Denes et al. 1999, Pandey and Pitman 2003, Faix and Beinhoff, 1988)
1061 1061
1031-1033
C-H in-plane deform. guaiacyl ring and C-O deform. primary alcohols and possibly influence of non-conjugated C=O groups (Faix and Beinhoff, 1988)
1033 1031-1034
Table 4.1 Assignment of FTIR peaks that distinguish control and decayed wood samples

64
Fig. 4.1 Spruce decay described by
FTIR analysis. A) Grouping on PCs 2
and 3 for normalized and mean-
centered FTIR data. Closed symbols
represent test data; open symbols
represent control data. Symbol shapes
represent time progression: time point
1 (O), 2 (), 3 (�), 4 ( ), 5 (◊). B)
Loadings for PC 3, separating more
and less decayed samples. Horizontal
dotted lines at magnitude |0.05|
represent thresholds for loading
significance. Peaks with highest
loadings for the control samples
denotes their loss in the decayed
samples. C) Mean baseline corrected
FTIR data normalized to total intensity
between 800 and 1800 cm-1: control
time point 1 (thick black line), test time
point 2 (thin black line) and test time
point 5 (thin grey line). Grey vertical
dashed lines are visual guides to
compare significant loadings in (B)
with FTIR spectra in (C).
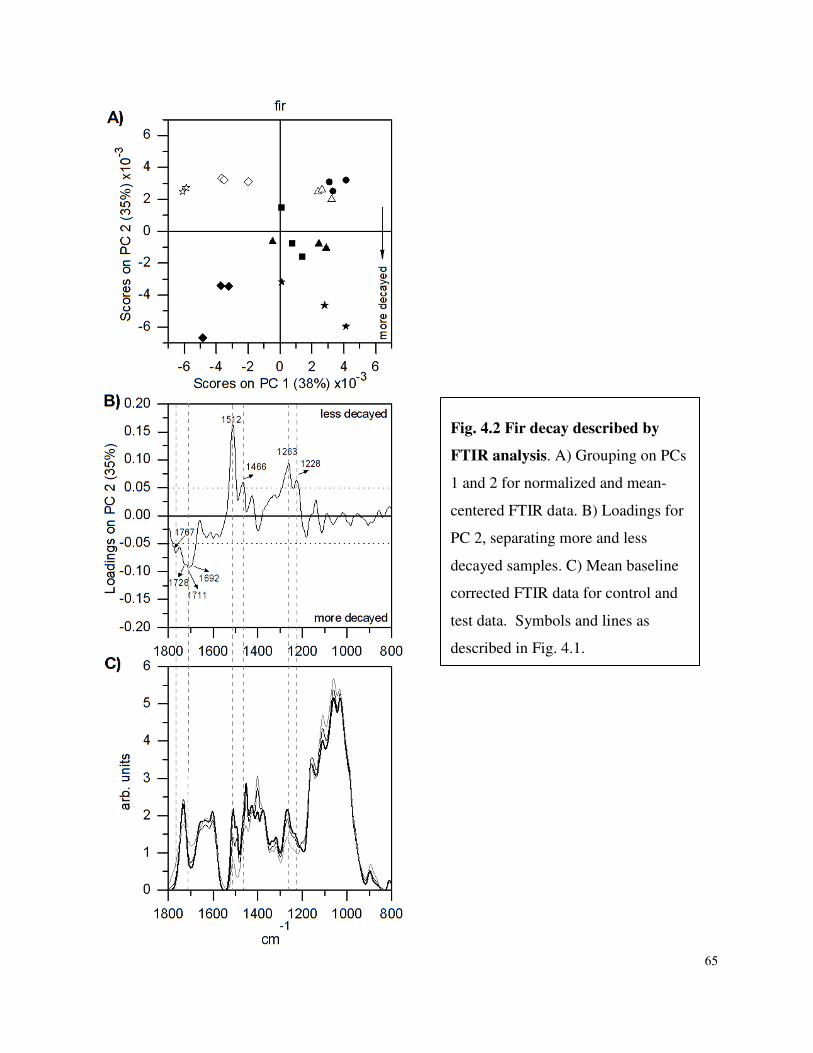
65
Fig. 4.2 Fir decay described by
FTIR analysis. A) Grouping on PCs
1 and 2 for normalized and mean-
centered FTIR data. B) Loadings for
PC 2, separating more and less
decayed samples. C) Mean baseline
corrected FTIR data for control and
test data. Symbols and lines as
described in Fig. 4.1.
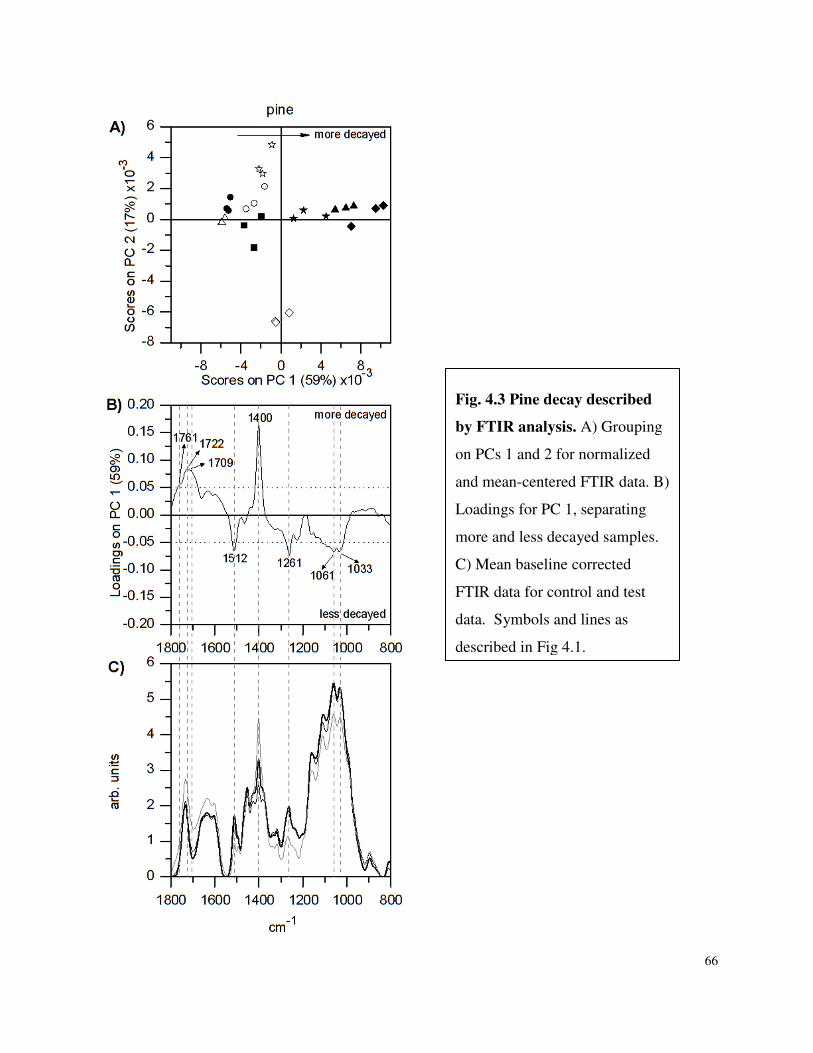
66
Fig. 4.3 Pine decay described
by FTIR analysis. A) Grouping
on PCs 1 and 2 for normalized
and mean-centered FTIR data. B)
Loadings for PC 1, separating
more and less decayed samples.
C) Mean baseline corrected
FTIR data for control and test
data. Symbols and lines as
described in Fig 4.1.

67
Nearly all of the peaks that corresponded to other significant loadings (>|0.05|) in control
samples were previously reported in milled-wood lignin (MWL) and/or G and GS lignin
dehydrogenation polymers by Faix and Beinhoff (1988). Exceptions include certain peaks in the
fir control samples that are not exclusive to lignin (Table 4. 1). For example, the peak at 1228
can be attributed to both lignin and hemicelluloses (Hobro et al. 2010). Still, the only peak not
associated with lignin that showed high peak loading in a control sample, was at 1400 cm-1 for
spruce. This peak was also highest in the 1480-1300 cm-1 region of non-cellulosic carbohydrates,
including arabinogalactan, guar galactomannan, mannose, pectin, tamarind xyloglucan and
birchwood xylan (Appendix 2, Table S4.1).
The PC loadings that describe the decayed samples indicate either bonds that were not attacked
by the fungus or else were generated through biological modification of the substrate. With the
exception of the loading at 1400 cm-1 that described decayed pine, PCA of all samples showed
high loadings in the 1800-1700 cm-1 region and the 1700-1550 cm-1 region. The PCA loading at
~1767 cm-1 and ~1711 cm-1 described decayed spruce, pine and fir, but were not detected in raw
spectra of the samples or standards and instead were correlated to shoulders (Fig. 4.1 to 4.3;
Appendix 2, Fig. S4.2). Similarly, the loading at 1722 cm-1 in decayed pine samples and loading
at 1728 cm-1 in decayed fir corresponded to peaks that were not detected in the raw spectra. All
of these loadings describe slopes of peaks in the 1800-1700 cm-1 region of the corresponding
spectra, where the 1734-1732 cm-1 region defines the peak maximum. This region describes
unconjugated carbonyl stretches that can be formed either in lignin or hemicelluloses (Faix
1992). For instance, three peaks in the 1737-1710 cm-1 region (1737, 1720 and 1710 cm-1) are
detected in lignin spectra after deconvolution (Faix and Beinhoff 1988) and like the peak at 1228
cm-1, a peak maximum at 1734 cm-1 has also been attributed to both lignin and hemicelluloses
(Hobro et al. 2010; Pandey and Pitman 2003)
The decayed samples were also characterized by significant loadings in the 1700-1550 cm-1
region, which were identified as slopes of peaks that are characteristic of conjugated carbonyl
stretches (Table 4.1). For instance, aromatic rings of milled wood lignin absorb at ~1595 cm-1
(Colom et al. 2003; Faix and Beinhoff 1988; Pandey and Pitman 2003), while Klason lignin
displays carbon stretching at 1682 cm-1 and aromatic skeletal vibration breathing with C=O

68
stretching at 1603 cm-1 (Kubo and Kadla 2005). Although outside of the range of important
loadings that describe the data reported here, conjugated carbonyl stretch in degraded cellulose
has been reported at 1620 cm-1(Lojewska et al. 2005).
In summary, the PCA of FTIR spectra described the decayed samples by peaks characteristic of
lignin and carbohydrates, while the same analysis distinguished the control samples primarily by
lignin-specific peaks that were lost upon degradation. Accordingly, the PCA of FTIR spectra
indicated that progressive decay by P. carnosa was mainly described by change in lignin
composition. TEM and ToF-SIMS were performed next to evaluate the localization of lignin
decay and to characterize the biochemical transformations in more detail.
4.4.2 TEM-EDXA
Lignin concentration in different regions of the cell wall differs considerably, with the highest
reported concentration being in cell corners and middle lamellae (Fromm et al. 2003; Saka et al.
1982). Here, the spatial degradation of lignin was investigated to describe microscopic
progression of P. carnosa attack on the softwood species.
Spruce samples from growth point one and five were selected for TEM-EDXA image analyses
given the clear compositional transformations detected in these samples as characterized by
FTIR. In this study, line scans of brominated samples across the cell wall and cell corners
demonstrated that the ratio of lignin in the cell walls to lignin in the middle lamella did not
change significantly with decay (Appendix 2, Table S4.1). No apparent change in lignin
concentration between cell walls suggests that P. carnosa simultaneously degraded lignin present
in the cell walls and middle lamella. Further, wood decay by P. carnosa did not appear to cause
morphological changes in the decayed samples.
4.4.3 ToF-SIMS
ToF-SIMS analysis was used to validate the FTIR results of lignin degradation and also the
lignin concentration pattern across fibre cell walls as observed by TEM-EDXA. Chemical
changes due to decay were assessed using an extended list of peak annotations for lignocellulosic
materials (Goacher et al. 2011).

69
A clear progression of lignin degradation in each of the three wood species tested was revealed
when comparing ToF-SIMS spectra of samples from early and late P. carnosa cultivations (Fig
4.4). The lignin peaks described by Goacher et al. (2011) characterized the samples at early
growth points (except m/z 67 for spruce and fir), while peaks characteristic of polysaccharides
were more prevalent in the later growth points.
However, the contributions of each lignin and polysaccharide peak depended on the wood
species. For example, degraded spruce samples exhibited lower enhancement of high-mass
polysaccharide peaks, suggesting that bioconversion activity had progressed further in these
samples. Also, the m/z 137 peak characteristic of guaiacyl lignin (Saito et al. 2005) was
particularly influential in separating degraded spruce and fir samples from corresponding
controls, but less influential for the pine samples. Finally, the prevalence of the lignin peak at
m/z 67 at the later time points of decayed spruce and fir samples may indicate a degradation
product that arose at this mass.
For all wood species, the PCA models of ToF-SIMS cross-sections of control and decayed
samples demonstrated higher concentrations of lignin in middle lamella and cell corners relative
to the intensities in the cell walls (Fig. 4.5, Appendix 2 Fig. S4.3 and S4.4), further supporting
the TEM-EDXA observations that P. carnosa simultaneously degrades lignin in the cell walls
and middle lamella.
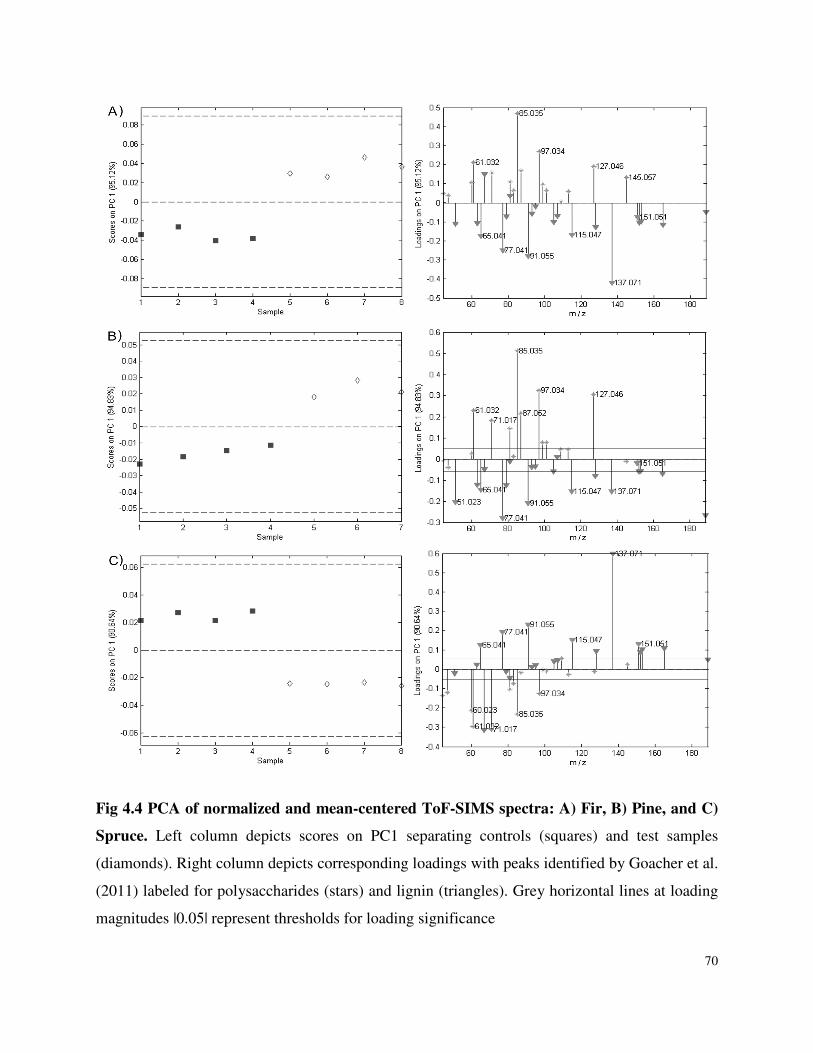
70
Fig 4.4 PCA of normalized and mean-centered ToF-SIMS spectra: A) Fir, B) Pine, and C)
Spruce. Left column depicts scores on PC1 separating controls (squares) and test samples
(diamonds). Right column depicts corresponding loadings with peaks identified by Goacher et al.
(2011) labeled for polysaccharides (stars) and lignin (triangles). Grey horizontal lines at loading
magnitudes |0.05| represent thresholds for loading significance
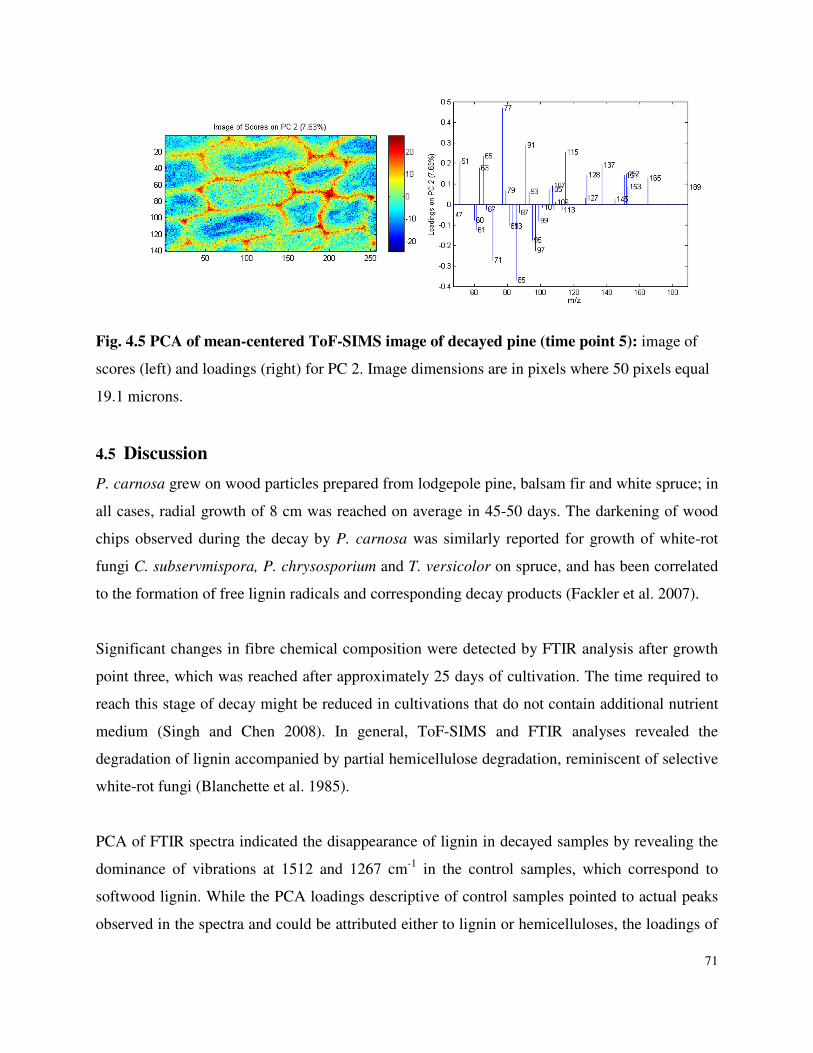
71
Fig. 4.5 PCA of mean-centered ToF-SIMS image of decayed pine (time point 5): image of
scores (left) and loadings (right) for PC 2. Image dimensions are in pixels where 50 pixels equal
19.1 microns.
4.5 Discussion
P. carnosa grew on wood particles prepared from lodgepole pine, balsam fir and white spruce; in
all cases, radial growth of 8 cm was reached on average in 45-50 days. The darkening of wood
chips observed during the decay by P. carnosa was similarly reported for growth of white-rot
fungi C. subservmispora, P. chrysosporium and T. versicolor on spruce, and has been correlated
to the formation of free lignin radicals and corresponding decay products (Fackler et al. 2007).
Significant changes in fibre chemical composition were detected by FTIR analysis after growth
point three, which was reached after approximately 25 days of cultivation. The time required to
reach this stage of decay might be reduced in cultivations that do not contain additional nutrient
medium (Singh and Chen 2008). In general, ToF-SIMS and FTIR analyses revealed the
degradation of lignin accompanied by partial hemicellulose degradation, reminiscent of selective
white-rot fungi (Blanchette et al. 1985).
PCA of FTIR spectra indicated the disappearance of lignin in decayed samples by revealing the
dominance of vibrations at 1512 and 1267 cm-1 in the control samples, which correspond to
softwood lignin. While the PCA loadings descriptive of control samples pointed to actual peaks
observed in the spectra and could be attributed either to lignin or hemicelluloses, the loadings of

72
decayed samples were not as clear and described changes in slopes in regions between 1800 and
1590 cm-1 overall suggesting changes in residual lignin and hemicelluloses. Notably, the PCA of
FTIR and ToF-SIMS also revealed that the progression of decay was most aggressive in spruce
samples, since similar levels of decay were achieved after the third growth point. Further, the
PCA of ToF-SIMS spectra composed of previously assigned peaks more clearly separated early
and late decay by lignin degradation. Given the clarity of the ToF-SIMS data and the capability
of this technique to give both spectral and spatial information of the samples, these analyses
reinforce the utility of ToF-SIMS for chemical analysis of complex biomass samples.
During delignification or selective decay, cell walls retain their morphology, but gradually
become un-reactive to lignin stains and increasingly reactive to cellulose stains as the lignin is
removed (Kirk and Cullen 1998). For instance, morphological alterations of cell walls were not
detected in pinewood chips degraded by the selective white-rot C. subvermispora, even though
staining with uranyl acetate indicated that the permeability of the secondary wall had changed
(Blanchette et al. 1997). Similarly, changes in the cell morphology and the ratio of lignin in cell
walls and middle lamellae were not detected when comparing early and late decayed samples by
TEM-EDXA and ToF-SIMS in the present study.
Most white-rot fungi that have been characterized to date typically degrade syringyl (S) lignin in
preference to guaiacyl (G) lignin (Faix et al. 1985). For instance, the secondary cell wall layer in
hardwoods is primarily composed of syringyl units and is typically more receptive to white-rot
attack than the cell corners, which are rich in guaiacyl units (Blanchette et al. 1987). By contrast,
the current results indicated that P. carnosa is proficient at attacking and modifying the guaiacyl
lignin component in softwood (as denoted by ToF-SIMS as m/z 137, and by FTIR as 1512 and
1268 cm-1). Though not detected in the samples characterized here, syringyl lignin is described
by a peak at 1128-1125 cm-1 in FTIR spectra and m/z 167 and 181 in ToF-SIMS spectra (Saito et
al. 2005)..
The degradation of guaiacyl lignin by P. carnosa is consistent with recent transcriptomic
analyses of P. carnosa grown on woody substrates (MacDonald et al. 2011). MacDonald et al.
(2011) revealed that the expression of transcripts encoding manganese peroxidases (MnP) in P.

73
carnosa were higher than those predicted to encode lignin peroxidases (LiP), and that the P.
carnosa genome encoded more genes that were predicted to encode MnPs than LiPs
(MacDonald et al. 2011). This is in contrast to P. chrysosporium, which encodes 10 LiPs and 5
MnPs (Martinez et al. 2004). Since the oxidants produced by MnP, including Mn3+, are smaller
than LiPs and associated oxidants, it is possible that products of MnP activity can diffuse more
easily through denser guaiacyl lignin structures present in softwood fibre, and promote softwood
decay by P. carnosa (Fackler et al. 2007; Hammel and Cullen 2008). As described in the next
Chapter, the similar pattern of decay observed for lodgepole pine, balsam fir and white spruce is
also consistent with the proteomic analysis of P. carnosa, which highlighted the secretion of
similar lignocellulolytic enzymes during growth on spruce and cellulose, implying the elicitation
of a similar decay strategy for different lignocellulosic substrates (Mahajan and Master 2010).
To summarize, P. carnosa can degrade guaiacyl lignin moieties and elicits selective white-rot
decay of lodgepole pine, balsam fir and white spruce. These results suggest that P. carnosa could
benefit bioprocess treatments used to reduce the recalcitrance of softwood species that
predominate in many boreal forests. Still, it is important to bear in mind that the pattern of white-
rot decay (i.e., selective or simultaneous decay) by wood-degrading fungi can vary depending on
cultivation conditions. For example, Ganoderma applanatum elicits simultaneous decay when
grown on Acer saccharum, Populus alba, Populus tremuloides, as does Ischnaderma resinosum
when grown on Populus deltoides (Blanchette 1984). However, both of these organisms were
correlated with selective decay of other tree species (Blanchette 1984). Accordingly, although P.
carnosa elicited selective degradation of lignin in lodgepole pine, balsam fir and white spruce, it
is plausible that the mode of decay may differ when grown on significantly different feedstocks,
including hardwood species.
Chapter 4 References
Birdsey RA, Jenkins JC, Johnston M, Huber-Sannwald E (2006) Chapter 11: North American Forests. In: Climate Change Science Program (CCSP) Product 2.2. USA.
Blanchette RA (1984) Screening wood decayed by white rot fungi for preferential lignin degradation Applied and Environmental Microbiology 48 (3):647-653

74
Blanchette RA, Otjen L, Effland MJ, Eslyn WE (1985) Changes in structural and chemical components of wood delignified by fungi. Wood Science and Technology 19:35-46
Blanchette RA, Otjen L, Carlson MC (1987) Lignin distribution in cell walls of birch wood decayed by white rot Basidiomycetes. Phytopathology 77 (5):684-690
Blanchette RA (1991) Delignification by wood-decay fungi. Annual Reviews of Phytopathology 29:381-398
Blanchette RA (1995) Degradation of the lignocellulosic complex in wood. Canadian Journal of Botany 73 (Supplemental 1):S999 - S1010
Blanchette RA, Krueger EW, Haight JE, Akhtar M, Akin DE (1997) Cell wall alterations in loblolly pine wood decayed by the white-rot fungus, Ceriporiopsis subvermispora
Journal of Biotechnology 53 (2-3):203-213
Boyle DC, Kropp BR (1992) Development and comparison of methods for measuring growth of filamentous fungi on wood. Canadian Journal of Microbiology 38:1053-1060
Burdsall HHJ (1985) A contribution to the taxonomy of the genus Phanerochaete (Corticiaceae,
Aphyllophorales). Mycological Memoir No. 10. J. Cramer, Braunschweig.
Colom X, Carrillo F, Nogués F, Garriga P (2003) Structural analysis of photodegraded wood by means of FTIR spectroscopy. Polymer Degradation and Stability 80 (3):543-549
Davis MF, Schroeder HA, Maciel GE (1994) Solid state 13C nuclear magnetic resonance studies of wood decay. II White rot decay of paper birch. Holzforschung 48:186-192
Fackler K, Schwanninger M, Hinterstoisser B, Messner K (2003) Bio-modification of spruce wood by Ceriporiopsis subvermispora: Comparison of the effects of three different strains. In: 12th International Symposium on Wood and Pulping Chemistry, Madison USA. pp 291-294
Fackler K, Gradinger C, Hinterstoisser B, Messner K, Schwanninger M (2006) Lignin degradation by white rot fungi on spruce wood shavings during short-time solid-state fermentations monitored by near infrared spectroscopy. Enzyme and Microbial Technology 39 (7):1476-1483
Fackler K, Gradinger C, Schmutzer C, Tavzes C, Burgert I, Schwanninger M, Hinterstoisser B, Watanabe T, Messner K (2007) Biotechnological wood modification with selective white-rot fungi and its molecular mechanisms. Food Technology and Biotechnology 45 (3):269-276
Faix O, Mozuch MD, Kirk TK (1985) Degradation of gymnosperm (guaiacyl) vs. angiosperm (syringyl/guaiacyl) lignins by Phanerochaete chrysosporium. Holzforschung - International Journal of the Biology, Chemistry, Physics and Technology of Wood 39 (4):203-208

75
Faix O, Beinhoff O (1988) FTIR spectra of milled wood lignins and lignin polymer models (DHP's) with enhanced resolution obtained by deconvolution. Journal of Wood Chemistry and Technology 8 (4):505-522
Faix O, Bremer J, Schmidt O, Stevanovic T (1991) Monitoring of chemical changes in white-rot degraded beech wood by pyrolysis-gas chromatography and Fourier transform infrared spectroscopy. Journal of Analytical and Applied Pyrolysis 21:147-162
Fromm J, Rockel B, Lautner S, Windeisen E, Wanner G (2003) Lignin distribution in wood cell walls determined by TEM and backscattered SEM techniques. Journal of Structural Biology 143 (1):77-84
Goacher R, Jeremic D, Master ER (2011) Expanding the library of secondary ions that distinguish lignin and polysaccharides in ToF-SIMS analysis of wood. Analytical Chemistry. 83:804-812.
Hakala TK, Maijal P, Konn J, Hatakka A (2004) Evaluation of novel wood-rotting polypores and corticioid fungi for the decay and biopulping of Norway spruce (Picea abies) wood Enzyme and Microbial Technology 34 (3-4):255-263
Hammel KE, Cullen D (2008) Role of fungal peroxidases in biological ligninolysis. Current Opinion in Plant Biology 11 (3):349-355
Hatakka A (1994) Lignin-modifying enzymes from selected white-rot fungi: production and role from in lignin degradation. FEMS Microbiology Reviews 13 (2-3):125-135
Hibbett DS, Donoghue MJ (2001) Analysis of character correlations among wood decay mechanisms, mating systems, and substrate ranges in homobasidiomycetes. Systems Biology 50:215-242
Hobro A, Kuligowski J, Döll M, Lendl B (2010) Differentiation of walnut wood species and steam treatment using ATR-FTIR and partial least squares discriminant analysis (PLS-DA). Analytical and Bioanalytical Chemistry 398 (6):2713-2722
Högberg N, Svegården IB, Kauserud H (2006) Isolation and characterization of 15 polymorphic microsatellite markers for the devastating dry rot fungus, Serpula lacrymans. Molecular Ecology Notes 6 (4):1022-1024
Irbe I, Andersons B, Chirkova J, Kallavus U, Andersone I, Faix O (2006) On the changes of pinewood (Pinus sylvestris L.). Chemical composition and ultrastructure during the attack by brown-rot fungi Postia placenta and Coniophora puteana International Biodeterioration and Biodegradation 57 (2):99-106
Jensen KA, Jr, Houtman CJ, Ryan ZC, Hammel KE (2001) Pathways for extracellular Fenton chemistry in the brown rot basidiomycete Gloeophyllum trabeum. Applied Environmental Microbiology 67:2705-2711

76
Jung S, Foston M, Sullards CM, Ragauskas AJ (2010) Surface characterization of dilute acid pretreated Populus deltoides by TOF-SIMS. Energy Fuels 24:1347-1357
Kelley SS, Jellison J, Goodell B (2002) Use of NIR and pyrolysis-MSMS coupled with multivariate analysis for detecting the chemical changes associated with brown-rot biodegradation of spruce wood. FEMS Microbiology Letters 209 (1):107-111
Kersten P, Cullen D (2007) Extracellular oxidative systems of the lignin-degrading Basidiomycete Phanerochaete chrysosporium. Fungal Genetics and Biology 44 (2):77-87
Kirk TK, Schultz E, Connors WJ, Lorenz LF, Zeikus JG (1978) Influence of culture parameters on lignin metabolism by Phanerochaete chrysosporium. Archives of Microbiology 117:277-285
Kirk TK, Cullen D (1998) Enzymology and molecular genetics of wood degradation by white-rot fungi. In: Young RA, Akhtar M (eds) Environmentally Friendly Technologies for the Pulp and Paper lndustry. John Wiley & Sons, Inc,
Kubo S, Kadla JF (2005) Hydrogen bonding in lignin: A Fourier Transform infrared model compound study. Biomacromolecules 6 (5):2815-2821
Lojewska J, Miskowiec P, Lojewski T, Proniewicz LM (2005) Cellulose oxidative and hydrolytic degradation: In situ FTIR approach. Polymer Degradation and Stability 88 (3):512-520
Lundell TK, Mäkelä MR, Hildén K (2010) Lignin-modifying enzymes in filamentous basidiomycetes – ecological, functional and phylogenetic review. Journal of Basic Microbiology 50 (1):5-20.
MacDonald J, Doering M, Canam T, Gong Y, Guttman D, Campbell M, Master ER (2011) Transcriptomic analysis of the softwood-degrading white-rot fungus Phanerochaete
carnosa reveals concerted gene expression responses to growth on coniferous and deciduous wood. Applied and Environmental Microbiology. [Epub ahead of print]
Mahajan S, Master ER (2010) Proteomic characterization of lignocellulose-degrading enzymes secreted by Phanerochaete carnosa grown on spruce and microcrystalline cellulose. Applied Microbiology and Biotechnology 86 (6):1903-1914
Marin S, Ramos AJ, Sanchis V. (2005) Comparison of methods for the assessment of growth of food spoilage moulds in solid substrates. International Journal of Food Microbiology 99:329-341
Martinez D, Larrondo LF, Putnam N, Sollewijn Gelpke MD, Huang K, Chapman J, Helfenbein KG, Ramaiya P, Detter JC, Larimer F, Coutinho PM, Henrissat B, Berka R, Cullen D, Rokhsar D (2004) Genome sequence of the lignocellulose degrading fungus Phanerochaete chrysosporium strain RP78. Nature Biotechnology 22:695-700

77
Martinez D, Challacombe J, Morgenstern I, Hibbett D, Schmoll M, Kubicek CP, Ferreira P, Ruiz-Duenas FJ, Martinez AT, Kersten P, Hammel KE, Vanden Wymelenberg A, Gaskell J, Lindquist E, Sabat G, Splinter BonDurant S, Larrondo LF, Canessa P, Vicuna R, Yadav J, Doddapaneni H, Subramanian V, Pisabarro AG, Lavin JL, Oguiza JA, Master E, Henrissat B, Coutinho PM, Harris P, Magnuson JK, Baker SE, Bruno K, Kenealy W, Hoegger PJ, Kües U, Ramaiya P, Lucas S, Salamov A, Shapiro H, Tu H, Chee CL, Misra M, Xie G, Teter S, Yaver D, James T, Mokrejs M, Pospisek M, Grigoriev IV, Brettin T, Rokhsar D, Berka R, Cullen D (2009) Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proceedings of the National Academy of Sciences 106 (6):1954-1959
Pandey KK, Pitman AJ (2003) FTIR studies of the changes in wood chemistry following decay by brown-rot and white-rot fungi. International Biodeterioration and Biodegradation 52:151-160
Pandey KK, Nagveni HC (2007) Rapid characterisation of brown and white rot degraded chir pine and rubberwood by FTIR spectroscopy. European Journal of Wood and Wood Products 65 (6):477-481
Rowell RM (2005) Handbook of Wood Chemistry and Wood Composites. CRC Press,
Saito K, Kato T, Tsuji Y, Fukushima K (2005) Identifying the characteristic secondary ions of lignin polymer using TOF-SIMS. Biomacromolecules 6:678-683
Saka S, Whiting P, Fukazawa K, Goring DAI (1982) Comparative studies on lignin distribution by UV microscopy and bromination combined with EDXA. Wood Science and Technology 16:269-277
Scott GM, Akhtar M, R.E S, Houtman CJ (2002) Recent developments in biopulping technology In: R L, L V (eds) Biotechnology in the pulp and paper industry, Madison WI, Elsevier Science
Singh D, Chen S (2008) The white-rot fungus Phanerochaete chrysosporium: conditions for the production of lignin-degrading enzymes. Applied Microbiology and Biotechnology 81:399-417
Sjöström E (1983) Wood Chemistry, Second Edition: Fundamentals and Applications. Academic Press,
Spurr AR (1968) A low-viscosity epoxy resin embedding medium for electron microscopy Journal of Ultrastructure Research 26 (1-2):31-43
Stein BD, Klomparens KL, Hammerschmidt R (1992) Comparison of bromine and permanganate as ultrastructural stains for lignin in plants infected by the fungus Colletotrichurn lagenariurn. Microscopy research and technique 23:201-206

78
Stephan T, Zehnpfennig J, Benninghoven A (1994) Correction of dead time effects in time-of-flight mass spectrometry. Journal of Vacuum Science and Technology A 12:405-410
Tuor U, Winterhalter K, Fiechter A (1995) Enzymes of white-rot fungi involved in lignin degradation and ecological determinants for wood decay. Journal of Biotechnology 41 (1):1-17

79
Chapter 5 : Proteomic Characterization of Lignocellulose-degrading Enzymes Secreted by Phanerochaete carnosa
Grown on Spruce and Microcrystalline Cellulose
5.1 Abstract
Proteins secreted by the white-rot, softwood degrading fungus Phanerochaete carnosa during
growth on cellulose and spruce were analyzed using tandem mass spectrometry and de novo
sequencing. Homology driven proteomics was applied to compare P. carnosa peptide sequences
to proteins in P. chrysosporium using MS BLAST and non-gapped alignment. In this way, 665
and 365 peptides from cellulose and spruce cultivations, respectively, were annotated. Predicted
activities included endoglucanases from glycoside hydrolase (GH) families 5, 16, and 61,
cellobiohydrolases from GH6 and GH7, GH3 β-glucosidases, xylanases from GH10 and GH11,
GH2 β-mannosidases, and de-branching hemicellulases from GH43 and CE15. Peptides
corresponding to glyoxal oxidases, peroxidases, and glycopeptides that could participate in lignin
degradation were also detected. Overall, predicted activities detected in extracellular filtrates of
spruce cultures represented a subset of those identified in cellulose cultures, suggesting that the
adaptation of P. carnosa to growth on lignocellulose might result from fine-tuning the expression
of similar enzyme families.
5.2 Introduction
Genomic and proteomic analyses of basidiomycetes are providing more complete descriptions of
enzyme combinations that play a critical role in carbon-cycling and can be applied in the
production of energy and chemicals from lignocellulosic biomass. Progress in this area is
exemplified by recent genomic and proteomic analyses of the white-rot fungus Phanerochaete
chrysosporium and the brown-rot fungus Postia placenta (Martinez et al. 2004; Vanden
Wymelenberg et al. 2009; Martinez et al. 2009). For instance, less than 20 % of predicted
glycoside hydrolases encoded by P. chrysosporium were characterized before the genome
sequence was published. And while proteomic analysis of P. chrysosporium grown on cellulose
and wood substrates consistently detect the expression of GH3 β-glycosidases, GH6 and GH7

80
cellobiohydrolases, GH10 xylanases, and GH12 endoglucanases, more than 65 % of the proteins
secreted by P. chrysoporium grown on crystalline cellulose corresponded to previously
uncharacterized glycoside hydrolases, esterases and proteases, as well as proteins with unknown
function (Abbas et al. 2005; Ravalason et al. 2008; Sato et al. 2007; Wymelenberg et al. 2005;
Vanden Wymelenberg et al. 2006; Vanden Wymelenberg et al. 2009). By contrast, genomic and
proteomic analysis of P. placenta identified oxidases predicted to generate extracellular Fe2+ and
H2O2 and relatively few cellulolytic glycoside hydrolases (GH) (Martinez et al. 2009). The
apparent lack of GHs encoded by P. placenta that contain carbohydrate-binding modules was
particularly striking.
As described in previous chapters, Phanerochaete carnosa is a white-rot fungus isolated from
balsam fir (Abies balsamea) as well as other softwoods, and shown to belong to the family
Phanerochaete sensu strictum (de Koker et al. 2003). Given the abundance of softwood and the
unique challenges associated with its hydrolysis, enzymes that have evolved to efficiently
transform softwood feedstocks are particularly relevant both industrially and for fundamental
investigation. To ascertain the diversity and regulation of carbohydrate-active and lignolytic
enzymes secreted by P. carnosa, a proteomic approach was applied to compare the secretome of
this fungus while growing on microcrystalline cellulose and spruce wood chips. This study
represents the first detailed characterization of the carbohydrate-active and lignin-degrading
enzymes produced by P. carnosa. Further, comparative proteomics was performed to reveal
enzymes and proteins secreted by P. carnosa that can accelerate the bioconversion of softwood
feedstocks to fermentable sugars. Since the P. carnosa genome sequence was not available
when this study was performed, the genome sequence of P. chrysosporium was used to facilitate
the annotation of peptide sequences retrieved. In this way, this study describes a proteomic
approach to investigate the enzymatic basis for feedstock preference, while also contributing to
the still limited number of investigations that have applied homology driven proteomics to
annotate peptides from organisms that lack genome sequence information (Junqueira et al. 2008).

81
5.3 Materials and Methods
5.3.1 Cultivation Conditions
As previously described, Phanerochaete carnosa strain HHB-10118 was obtained from the US
Department of Agriculture (USDA) Forest Products Laboratory, Madison, WI. The strain was
grown on liquid or solid medium under stationary conditions at 27 °C, and was maintained on
YMPG agar slants (yeast extract (2 % w/v), maltose (10 % w/v), peptone (2 % w/v), glucose (1
% w/v) and agar (2 %w/v)) at 4 °C.
Cellulose medium was prepared in 1 L flasks and contained 1 % wood-derived microcrystalline
cellulose (Avicel PH-101, Fluka) submerged in 200 mL B3 medium (low nitrogen, 0.1 mg L-1
thiamine HCl, 0.2 mg L-1 ammonium tartrate, 10 mM dimethyl succinate, pH 4.5) (Kirk et al.
1978). Wood cultivations were prepared in autoclavable 22 X 28 cm polypropylene bags
containing a gas exchange filter (Unicorn Imp & Mfg Corp TX USA) (Sato et al. 2007).
Briefly, chips of white spruce (Picea glauca) measuring approximately 0.1 cm X 0.2 cm X 1.2
cm were washed with milliQ water, steam sterilized for 30 min, and 225 g of the chips were
submerged in 350 mL of B3 medium. The inoculum was obtained from P. carnosa cultivations
grown for 7 days at 27 °C in liquid YMPG medium. The mycelium was filtered through sterile
Miracloth, rinsed with B3 buffer (pH 4.5), and then blended three times for 10 s using a sterile
Waring blender. The mycelium suspension was transferred to each culture to obtain an initial
mycelium concentration of 40 mg (fresh weight) 100 mL-1. Cellulose and spruce cultivations
were grown at 27 °C under stationary conditions for 4 and 12 weeks, respectively. Triplicate
cultivations were prepared for each growth condition; cellulose and spruce growth medium
without fungal inoculation served as abiotic controls. Extents of fungal growth on cellulose was
determined by measuring final dry weight and total chitin in each cultivation (Plassard et al.
1982).
5.3.2 Protein Extraction
Following vacuum filtration of cultivation supernatants through Miracloth, mycelium was rinsed
with B3 buffer and the filtrate was recovered then pooled with the corresponding culture
supernatant. Liquid fractions were centrifuged at 5300 x g for 20 min at 4 °C, and then filtered
using 0.45 µm syringe filters. The filtrate was concentrated approximately 20 times using

82
Amicon Centrifugal devices (5000 MWCO polyethersulfone membrane; Centricon-20 cat:
UFC2BCC24). Protein concentration was measured by the Bio-Rad Bradford assay, and
concentrated samples were stored at -80 ˚C for proteomic analysis.
5.3.3 Protein Preparation and Analysis by Mass Spectrometry
Protein preparations (20 µg each) were fractionated by 1-D SDS-PAGE and each lane was
excised into 10 fractions using a clean razor blade. In-gel trypsin digestion was then performed
by treating each gel segment with 40 µL of 50 mM NH4HCO3 and 12.5 ng µL-1 of modified
trypsin (Promega, sequencing grade) (Shevchenko et al. 1996). Peptides were extracted from the
gel by shaking for 20 min at room temperature with 50 µL of 5 % formic acid twice, and then
with 50 µL of 5 % formic acid in 30 % acetonitrile. The peptide extracts were concentrated to
roughly 20 µL by vacuum centrifuge. Concentrated peptide samples were loaded onto a
HPLC_Chip (160 nL high capacity sample enrichment column and 75-µm x 150 mm SB-C18
separation column, Agilent Technologies, Santa Clara, CA, USA) and separated by flow rate at
300 nL min-1, with solvent A (0.2 (v/v) % formic acid in water) and solvent B (100 %
acetonitrile) and the following gradients: 3 %, 35 %, 80 %, 100 % of solvent B at 0, 50, 54 and
56 min after injection, respectively. The LC-MS/MS analysis was carried out by an Agilent 1100
HPLC-chip and 6340 ion trap system with MS scan range from 300 to 1,300 m/z. Three
precursor ions were selected from each MS scan for back-to-back collision induced dissociation
and electron transfer dissociation (CID/ETD). A 30 s dynamic exclusion period was applied to
the precursor previously selected twice for MS/MS, i.e., two rounds, each with CID/ETD.
(SunnyBrook Health Science Centre, Proteomics Core Facility).
5.3.4 Peptide Sequence Annotation
Since the P. carnosa genome sequence was not available at the time of analysis, it was necessary
to identify peptide spectra through de novo sequencing. The SHERENGA de novo sequencing
tool (version A.03.03.084, Agilent Technologies) was used to explicitly read peptide sequences
from high-quality fragment ion spectra (Dancik et al. 1999). Peptide sequences that received a
SHERENGA score greater than 100 were matched to all predicted proteins from Phanerochaete
chrysosporium (v.2.1) using MS BLAST ((Shevchenko et al. 2001),
http://genetics.bwh.harvard.edu/msblast/index.html). To increase the confidence of protein

83
matches based on a homology-driven approach, only alignments receiving an MS BLAST score
of 55 or above were considered, which is consistent with other scoring schemes based on MS
BLAST analyses (Shevchenko et al. 2001, Junqueira et al. 2008). Peptide sequences were also
compared to predicted proteins from Phanerochaete chrysosporium using BLASTp and the
following parameter settings: no gaps, word length 5, and gap extension cost 100. The PAM30
substitution matrix was selected given the short peptide sequences that were queried in the
BLAST search. Initial analyses were automated using freely available BioPython source code
(Cock et al. 2009), and peptide matches were manually inspected. Percent identity was computed
as the number of contiguous amino acid matches divided by the total peptide length. To avoid
over prediction of protein hits, protein sequences receiving only one peptide match were ignored
if the corresponding peptide aligned to another protein with additional peptide matches. The
MS/MS spectra of peptides that were annotated were then manually interpreted to verify the
validity of de novo sequences predicted using SHERENGA. Non-gapped direct alignments using
BLASTp was also performed to compare peptide sequences from extracellular filtrates of
cellulose and spruce cultivations.
5.4 Results
5.4.1 Preparation and Analysis of Peptide Samples
Cellulose and spruce cultivations prepared for proteomic analyses of P. carnosa extracellular
filtrates were grown at 27 °C. While cellulose cultivations were harvested after four weeks of
incubation, spruce cultivations were grown for an additional eight weeks given the comparative
recalcitrance of the substrate.
Protein secreted from Phanerochaete carnosa grown on cellulose and spruce was fractionated by
SDS-PAGE to confirm that concentrated proteins from triplicate cellulose and spruce
cultivations generated reproducible protein profiles. Approximately 5.4 mg and 9.8 mg of protein
were recovered from 200 mL of cellulose-grown and 350 mL of spruce-grown culture
supernatant, respectively. Proteins were fractionated by one-dimensional gel electrophoresis,
each lane was cut into at least ten segments, and each section was processed by in-gel trypsin
digestion. Peptides eluted from each gel segment were then analyzed by LC-MS/MS. This
procedure enabled the characterization of proteins that might not have been detected by two-

84
dimensional gel electrophoresis. This approach also avoided reported difficulties associated with
proteins extracted from wood-grown cultivations, which produce streaks in electrophoresis gels
that mask protein spots (Sato et al. 2007).
De novo sequencing was performed to explicitly read peptide sequences. In total, 3222 and 1763
peptide sequences were recovered from cellulose and spruce cultivations of P. carnosa,
respectively. Of these, 665 and 365 could be annotated based on sharing at least 70 % sequence
identity to predicted proteins from the white-rot fungus Phanerochaete chrysosporium. Notably,
direct non-gapped alignments also indicated that only 16 % (284) of the peptides obtained from
extracellular filtrates of spruce cultures shared greater than 70 % identity to peptides extracted
from cellulose cultivations.
Peptide sequences with best matches to predicted glycoside hydrolases, esterases,
oxidoreductases, monooxygenases, and proteins with unknown function were manually
inspected; all hits reported here correspond to proteins with predicted secretory signal sequences
and that lack predicted transmembrane sequences (Fig. 5.1). In most cases, non-gapped direct
alignments were able to validate the MS BLAST results, since many of the same peptide matches
were predicted in both cases (Tables 5.1 to 5.3). However, the percent identity of peptide and
protein sequences from P. carnosa and P. chrysosporium, respectively, did not correlate well to
MS BLAST score. Therefore, matches to enzyme families identified through both MS BLAST
and non-gapped alignment are reported in Tables 5.1 and 5.2, while enzyme families that were
predicted using non-gapped alignment only are summarized in the supplemental material
(Appendix 3, Table S5.1). Since P. carnosa peptide sequences were annotated using predicted
protein sequences from a different, albeit related, organism, P. carnosa peptide annotations are
discussed below in terms of predicted enzyme activity rather than expression of a specific
protein.
5.4.2 Cellulases and Hemicellulases in Cellulose and Spruce Cultivations
Cellulose degradation by white-rot fungi is mediated by the concerted activity of three
cellulolytic enzymes: endoglucanases, cellobiohydrolases and β-glucosidases (Baldrian and
Valaskova 2008). Evidence of these cellulolytic activities was found in the secretome
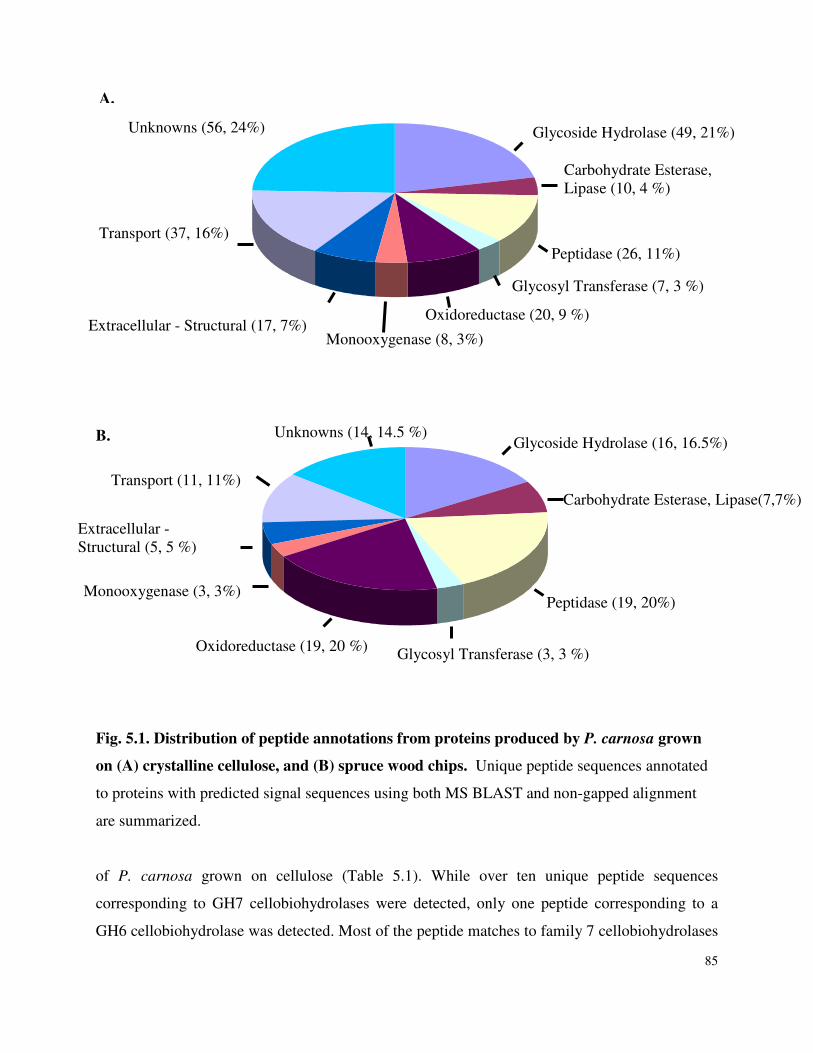
85
B.
Fig. 5.1. Distribution of peptide annotations from proteins produced by P. carnosa grown
on (A) crystalline cellulose, and (B) spruce wood chips. Unique peptide sequences annotated
to proteins with predicted signal sequences using both MS BLAST and non-gapped alignment
are summarized.
of P. carnosa grown on cellulose (Table 5.1). While over ten unique peptide sequences
corresponding to GH7 cellobiohydrolases were detected, only one peptide corresponding to a
GH6 cellobiohydrolase was detected. Most of the peptide matches to family 7 cellobiohydrolases
Glycoside Hydrolase (16, 16.5%)
Carbohydrate Esterase, Lipase(7,7%)
Peptidase (19, 20%)
Oxidoreductase (19, 20 %) Glycosyl Transferase (3, 3 %)
Monooxygenase (3, 3%)
Extracellular - Structural (5, 5 %)
Transport (11, 11%)
Unknowns (14, 14.5 %)
Glycoside Hydrolase (49, 21%)
Carbohydrate Esterase, Lipase (10, 4 %)
Peptidase (26, 11%)
Oxidoreductase (20, 9 %)
Glycosyl Transferase (7, 3 %)
Monooxygenase (8, 3%) Extracellular - Structural (17, 7%)
Transport (37, 16%)
Unknowns (56, 24%)
A.

86
were distributed over conserved regions of this GH family. This result confounds the ability to
predict the number of unique GH7 enzymes secreted by P.carnosa. However, since only those
protein hits containing at least one unique peptide match are listed in Table 5.1, the higher
number of peptides corresponding to a GH7 cellobiohydrolase likely results from the secretion of
multiple GH7 isozymes. MS BLAST analyses identified three unique peptide sequences
corresponding to GH3 glycoside hydrolases in the culture of P. carnosa grown on cellulose. GH
family 3 enzymes exhibit a broad range of substrate specificity, including β-glucosidases that
hydrolyze cellobiose to glucose.
Comparatively few peptide sequences corresponding to GH families with uniquely endo-
cellulolytic activities were identified in extracellular filtrates of P. carnosa grown on cellulose or
spruce. Non-gapped alignments identified peptide sequences that were matched to GH5 and
GH74 enzymes in P. chrysosporium; so it is conceivable that GH5 and GH74 enzymes secreted
by P. carnosa catalyze additional cellulolytic activity (Appendix 3 Table S5.1). Five peptide
sequences corresponding to GH61 endoglucanases were also identified in cellulose cultivations
(two by both MS BLAST and BLASTp). The Cel61A enzyme from Aspergillus kawachii has
endoglucanase activity, and so it is tempting to conclude that GH61 enzymes could play a role in
cellulose degradation by P. carnosa. However, the biological function of GH61 enzymes
remains unclear. Recent structural analyses of a GH61 enzyme could not unambiguously assign
amino acid residues involved in catalysis and substrate binding (Karkehabadi et al. 2008).
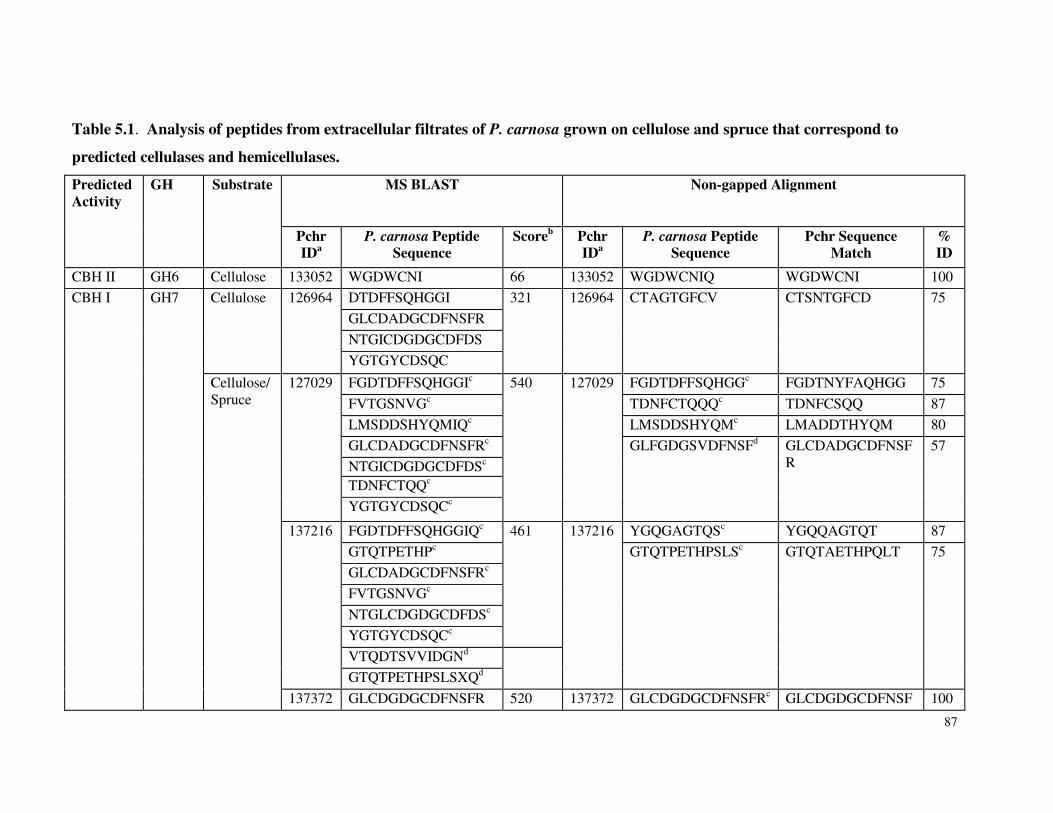
87
Table 5.1. Analysis of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce that correspond to
predicted cellulases and hemicellulases.
Predicted Activity
GH
Substrate
MS BLAST
Non-gapped Alignment
Pchr IDa
P. carnosa Peptide Sequence
Scoreb Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
CBH II GH6 Cellulose 133052 WGDWCNI 66 133052 WGDWCNIQ WGDWCNI 100
CBH I GH7 Cellulose
126964
DTDFFSQHGGI 321
126964
CTAGTGFCV
CTSNTGFCD
75
GLCDADGCDFNSFR
NTGICDGDGCDFDS
YGTGYCDSQC
Cellulose/ Spruce
127029
FGDTDFFSQHGGIc 540
127029
FGDTDFFSQHGGc FGDTNYFAQHGG 75
FVTGSNVGc TDNFCTQQQc TDNFCSQQ 87
LMSDDSHYQMIQc LMSDDSHYQMc LMADDTHYQM 80
GLCDADGCDFNSFRc GLFGDGSVDFNSFd
GLCDADGCDFNSFR
57
NTGICDGDGCDFDSc TDNFCTQQc
YGTGYCDSQCc
137216
FGDTDFFSQHGGIQc 461
137216
YGQGAGTQSc YGQQAGTQT 87
GTQTPETHPc GTQTPETHPSLSc
GTQTAETHPQLT
75
GLCDADGCDFNSFRc
FVTGSNVGc
NTGLCDGDGCDFDSc
YGTGYCDSQCc
VTQDTSVVIDGNd
GTQTPETHPSLSXQd
137372 GLCDGDGCDFNSFR 520 137372 GLCDGDGCDFNSFRc GLCDGDGCDFNSF 100
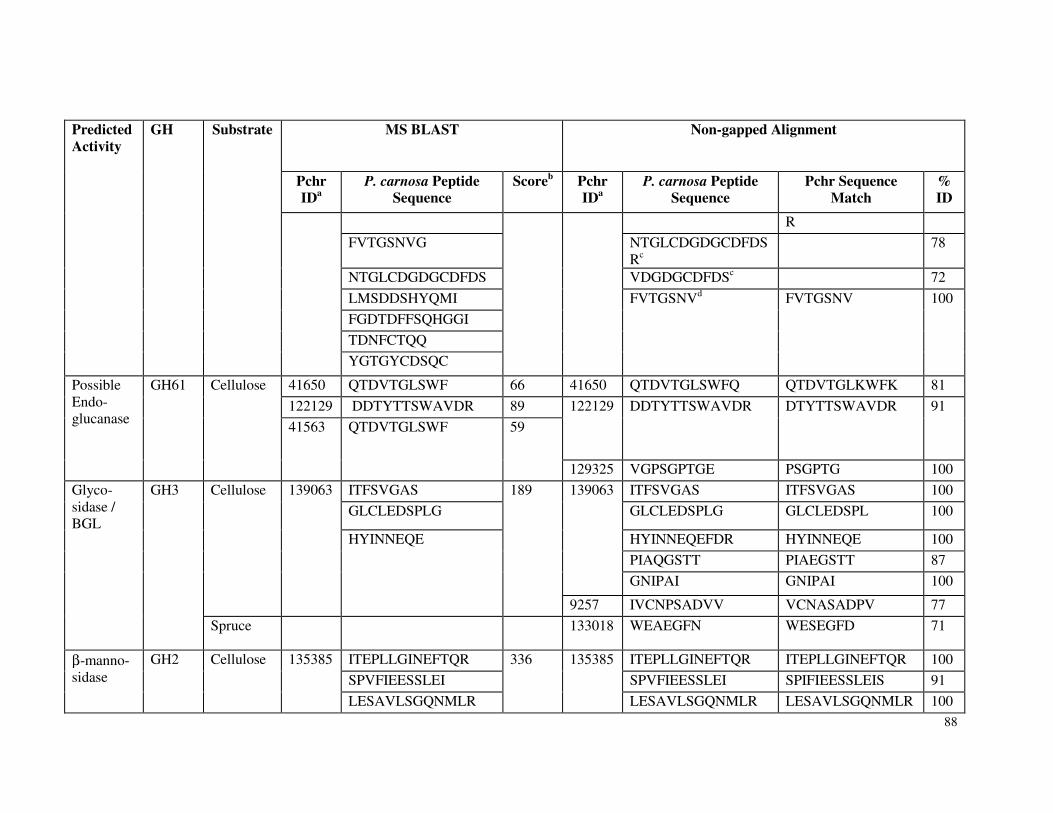
88
Predicted Activity
GH
Substrate
MS BLAST
Non-gapped Alignment
Pchr IDa
P. carnosa Peptide Sequence
Scoreb Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
R
FVTGSNVG NTGLCDGDGCDFDSRc
78
NTGLCDGDGCDFDS VDGDGCDFDSc 72
LMSDDSHYQMI FVTGSNVd FVTGSNV 100
FGDTDFFSQHGGI
TDNFCTQQ
YGTGYCDSQC
Possible Endo-glucanase
GH61
Cellulose
41650 QTDVTGLSWF 66 41650 QTDVTGLSWFQ QTDVTGLKWFK 81
122129 DDTYTTSWAVDR 89 122129 DDTYTTSWAVDR DTYTTSWAVDR 91
41563
QTDVTGLSWF
59
129325 VGPSGPTGE PSGPTG 100
Glyco-sidase / BGL
GH3
Cellulose
139063
ITFSVGAS 189
139063
ITFSVGAS ITFSVGAS 100
GLCLEDSPLG GLCLEDSPLG GLCLEDSPL 100
HYINNEQE
HYINNEQEFDR HYINNEQE 100
PIAQGSTT PIAEGSTT 87
GNIPAI GNIPAI 100
9257 IVCNPSADVV VCNASADPV 77
Spruce 133018 WEAEGFN WESEGFD 71
β-manno-sidase
GH2
Cellulose
135385
ITEPLLGINEFTQR 336
135385
ITEPLLGINEFTQR ITEPLLGINEFTQR 100
SPVFIEESSLEI SPVFIEESSLEI SPIFIEESSLEIS 91
LESAVLSGQNMLR LESAVLSGQNMLR LESAVLSGQNMLR 100
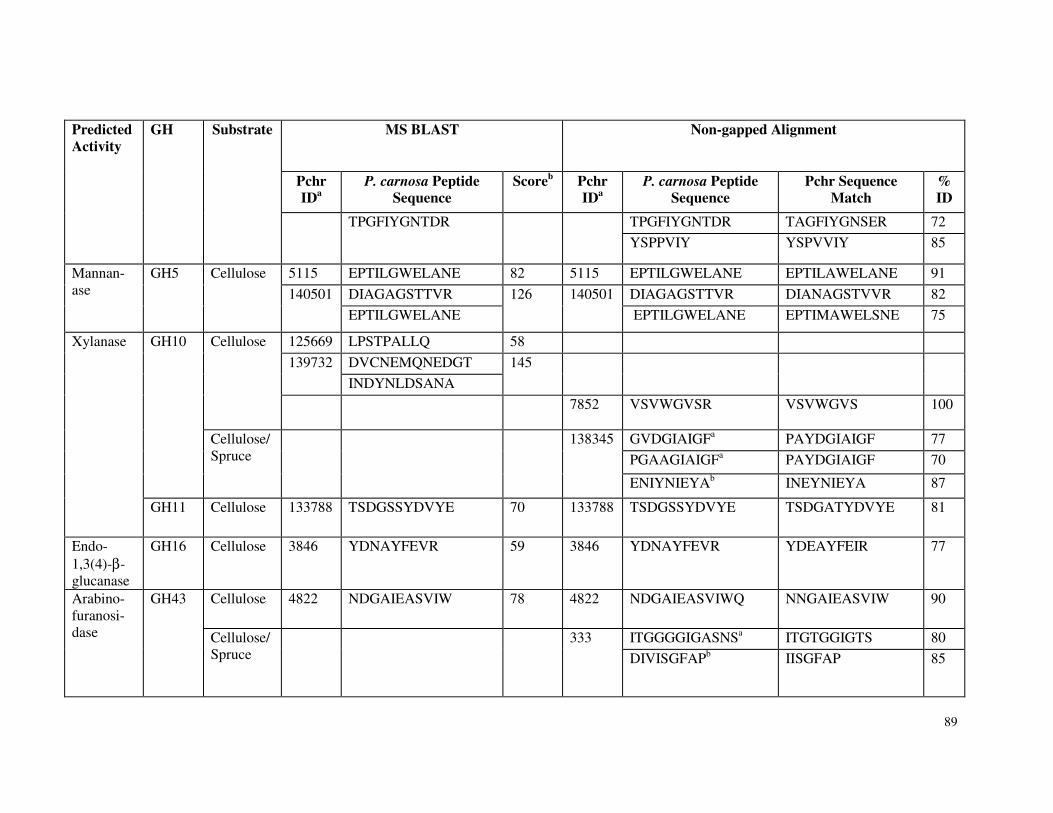
89
Predicted Activity
GH
Substrate
MS BLAST
Non-gapped Alignment
Pchr IDa
P. carnosa Peptide Sequence
Scoreb Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
TPGFIYGNTDR
TPGFIYGNTDR TAGFIYGNSER 72
YSPPVIY YSPVVIY 85
Mannan-ase
GH5
Cellulose
5115 EPTILGWELANE 82 5115 EPTILGWELANE EPTILAWELANE 91
140501
DIAGAGSTTVR 126
140501
DIAGAGSTTVR DIANAGSTVVR 82
EPTILGWELANE EPTILGWELANE EPTIMAWELSNE 75
Xylanase
GH10
Cellulose 125669 LPSTPALLQ 58
139732
DVCNEMQNEDGT 145
INDYNLDSANA
7852 VSVWGVSR VSVWGVS 100
Cellulose/Spruce
138345
GVDGIAIGFa PAYDGIAIGF 77
PGAAGIAIGFa PAYDGIAIGF 70
ENIYNIEYAb INEYNIEYA 87
GH11
Cellulose
133788
TSDGSSYDVYE
70
133788
TSDGSSYDVYE
TSDGATYDVYE
81
Endo-1,3(4)-β-glucanase
GH16 Cellulose 3846 YDNAYFEVR 59 3846 YDNAYFEVR YDEAYFEIR 77
Arabino-furanosi-dase
GH43
Cellulose
4822
NDGAIEASVIW
78
4822
NDGAIEASVIWQ
NNGAIEASVIW
90
Cellulose/Spruce
333
ITGGGGIGASNSa ITGTGGIGTS 80
DIVISGFAPb IISGFAP 85
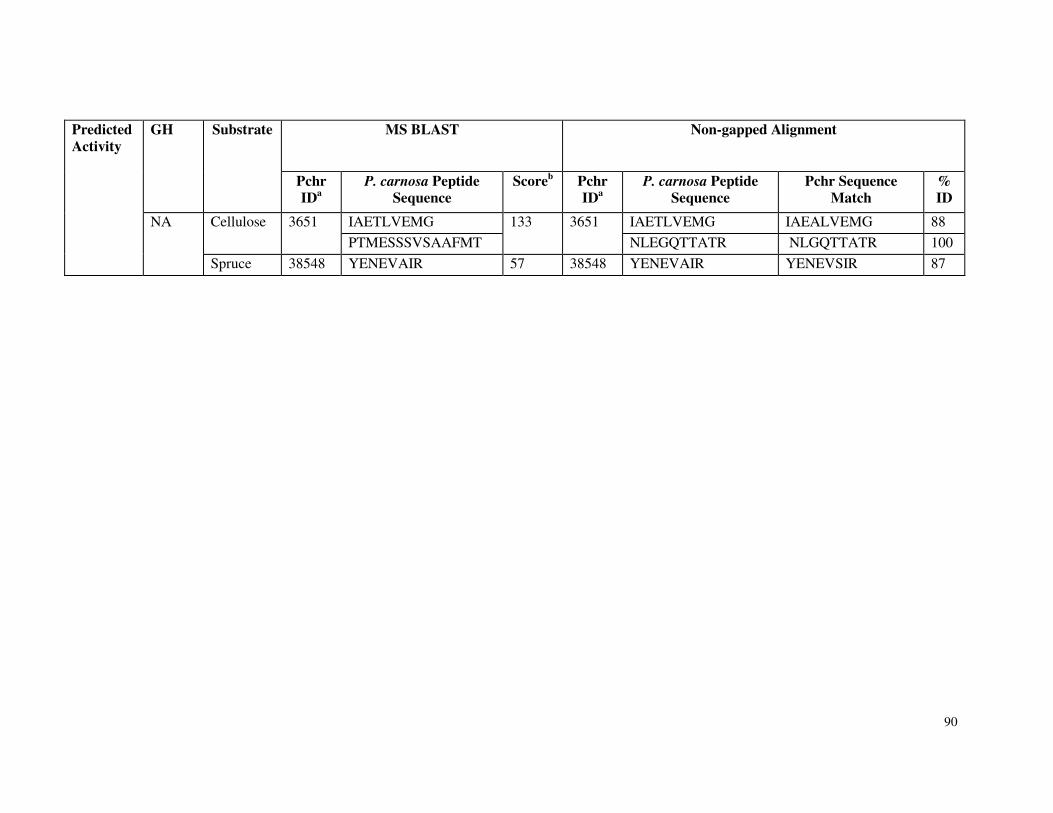
90
Predicted Activity
GH
Substrate
MS BLAST
Non-gapped Alignment
Pchr IDa
P. carnosa Peptide Sequence
Scoreb Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
NA
Cellulose
3651
IAETLVEMG 133
3651
IAETLVEMG IAEALVEMG 88
PTMESSSVSAAFMT NLEGQTTATR NLGQTTATR 100
Spruce 38548 YENEVAIR 57 38548 YENEVAIR YENEVSIR 87

91
a Additional information for protein from P. chrysosporium can be accessed by ending the
following URL with the model number, e.g. http://genome.jgi-psf.org/cgi-
bin/dispGeneModel?db=Phchr1&id=6482; bAnnotation score from MS BLAST; c Detected in
extracellular filtrates of cellulose cultivations; d Detected in extracellular filtrates of spruce
cultivations. Figure S5.1 summarizes Table 5.1 as pie graphs.
Both MS BLAST and non-gapped alignments matched peptide sequences from P. carnosa to
Pchr5115 and Pchr140501, which are predicted to encode mannanase activity and were detected
in P. chrysosporium cultivations grown on Wood’s medium and spruce (Ravalason et al. 2008;
Vanden Wymelenberg et al. 2009). Both methods also identified a GH2 mannosidase in P.
carnosa cultures grown on cellulose. While a GH2 mannosidase was detected in extracellular
filtrates of P. chrysosporium maintained on replete medium (Vanden Wymelenberg et al. 2009),
GH2 enzymes have not been recovered by proteomic analyses of P. chrysosporium grown on
cellulose or woody substrates.
Seven peptides sequences matching GH10 xylanases were detected in both cellulose and spruce
cultivations. By contrast, only one peptide sequence from cellulose cultures was matched to a
GH11 xylanase. Both MS BLAST and non-gapped alignments matched a similar number of
peptide sequences to α-arabinofuranosidases, supporting other studies that emphasize the
importance of debranching enzymes in the hydrolysis of lignocellulosic substrates (de Vries et al.
2000).
5.4.3 Oxidoreductases in Cellulose and Spruce Cultivations
Peptides recovered from both cellulose and spruce cultivations were matched to Pchr4636 (Table
5.2), a manganese peroxidase (MnP) that was previously detected in P. chrysosporium
cultivations grown on spruce (Ravalason et al. 2008). Non-gapped alignment also identified
peptides from spruce cultivations that matched Pchr131707, a lignin peroxidase (LiP) that is
encoded by a gene located on scaffold 19 of the P. chrysosporium genome (Appendix 3 Table 1).
While the expression cellobiose dehydrogenease activity (CDH) was not detected, peptides from
extracellular filtrates of P. carnosa grown on spruce were matched to a putative multicopper

92
oxidase and Cir1, a CBM-containing reductase that is related to CDH1 (Table 5.2, Appendix 3
Table 1). Multicopper oxidases are not implicated in lignin degradation by P. chrysosporium;
instead, they may function as extracellular ferroxidases with potential to modulate Fenton
reactions (Kersten and Cullen 2007; Larrondo et al. 2003). Notably, the majority of peptide
matches to oxidoreductases were to enzymes thought to be involved in extracellular peroxide
generation, including glyoxal oxidases and GMC oxidoreductases (Table 5.2). Further, a peptide
sequence matching Pchr1923 was detected in culture filtrates of P. carnosa grown on cellulose.
Pchr1923 shares over 70 % sequence identity to deduced amino acid sequence of glp2, which
encodes a glycopeptide implicated in the production of hydroxyl radicals involved in lignin and
carbohydrate degradation (Baldrian and Valaskova 2008; Tanaka et al. 2007).
Peptides corresponding to cytochrome P450s and monooxygenases were also detected in both
cellulose and spruce cultivations (Table 5.2, Appendix 3 Table 1). Notably, Pchr37971, Pchr663,
Pchr38849 and Pchr139445 are CYP2E-type enzymes, a class of P450 monooxygenases that
participate in the bioconversion of exogenous aromatic compounds. Although only a few
peptides corresponding to monooxygenase activity were detected in spruce cultivations, the
dependence of monooxygenase expression on biomass feedstock composition will be interesting
to pursue given the potential of this enzyme family to degrade oleoresins in softwood.
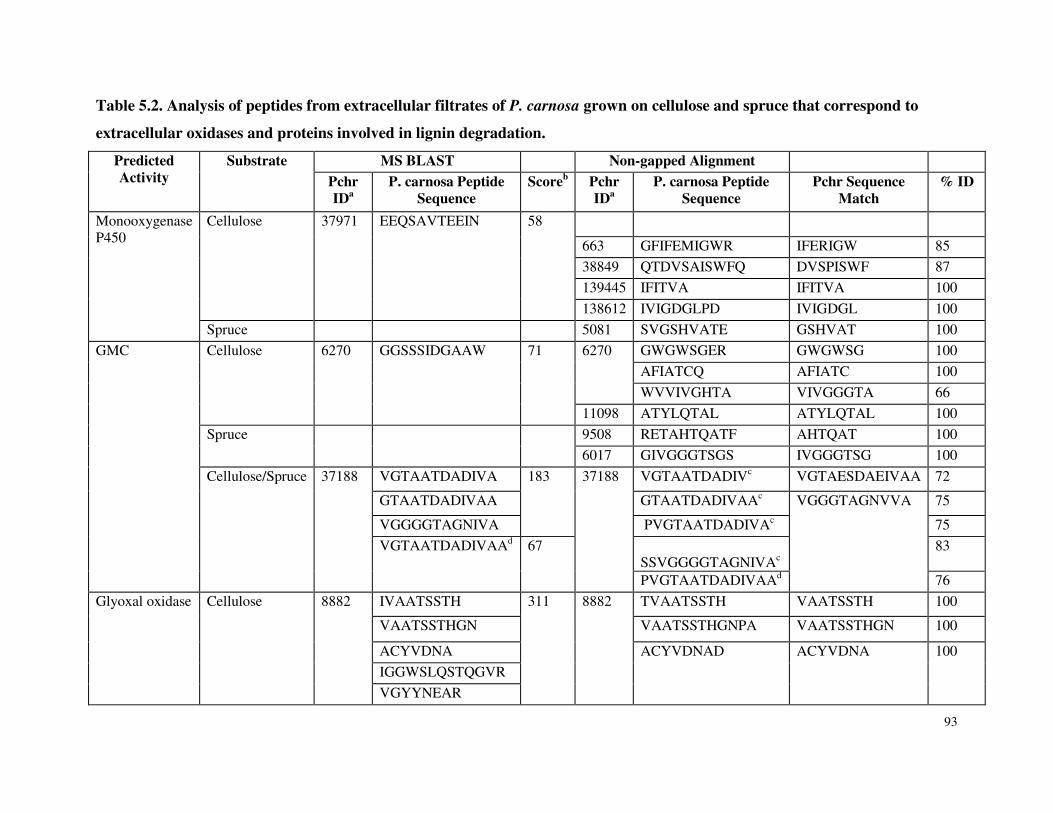
93
Table 5.2. Analysis of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce that correspond to
extracellular oxidases and proteins involved in lignin degradation.
Predicted Activity
Substrate
MS BLAST Non-gapped Alignment
Pchr IDa
P. carnosa Peptide Sequence
Scoreb Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
Monooxygenase P450
Cellulose
37971
EEQSAVTEEIN
58
663 GFIFEMIGWR IFERIGW 85
38849 QTDVSAISWFQ DVSPISWF 87
139445 IFITVA IFITVA 100
138612 IVIGDGLPD IVIGDGL 100
Spruce 5081 SVGSHVATE GSHVAT 100
GMC
Cellulose
6270
GGSSSIDGAAW
71
6270
GWGWSGER GWGWSG 100
AFIATCQ AFIATC 100
WVVIVGHTA VIVGGGTA 66
11098 ATYLQTAL ATYLQTAL 100
Spruce
9508 RETAHTQATF AHTQAT 100
6017 GIVGGGTSGS IVGGGTSG 100
Cellulose/Spruce
37188
VGTAATDADIVA 183
37188
VGTAATDADIVc VGTAESDAEIVAA 72
GTAATDADIVAA GTAATDADIVAAc VGGGTAGNVVA
75
VGGGGTAGNIVA PVGTAATDADIVAc 75
VGTAATDADIVAAd
67
SSVGGGGTAGNIVAc
83
PVGTAATDADIVAAd 76
Glyoxal oxidase
Cellulose
8882
IVAATSSTH 311
8882
TVAATSSTH VAATSSTH 100
VAATSSTHGN VAATSSTHGNPA VAATSSTHGN 100
ACYVDNA ACYVDNAD
ACYVDNA
100
IGGWSLQSTQGVR
VGYYNEAR
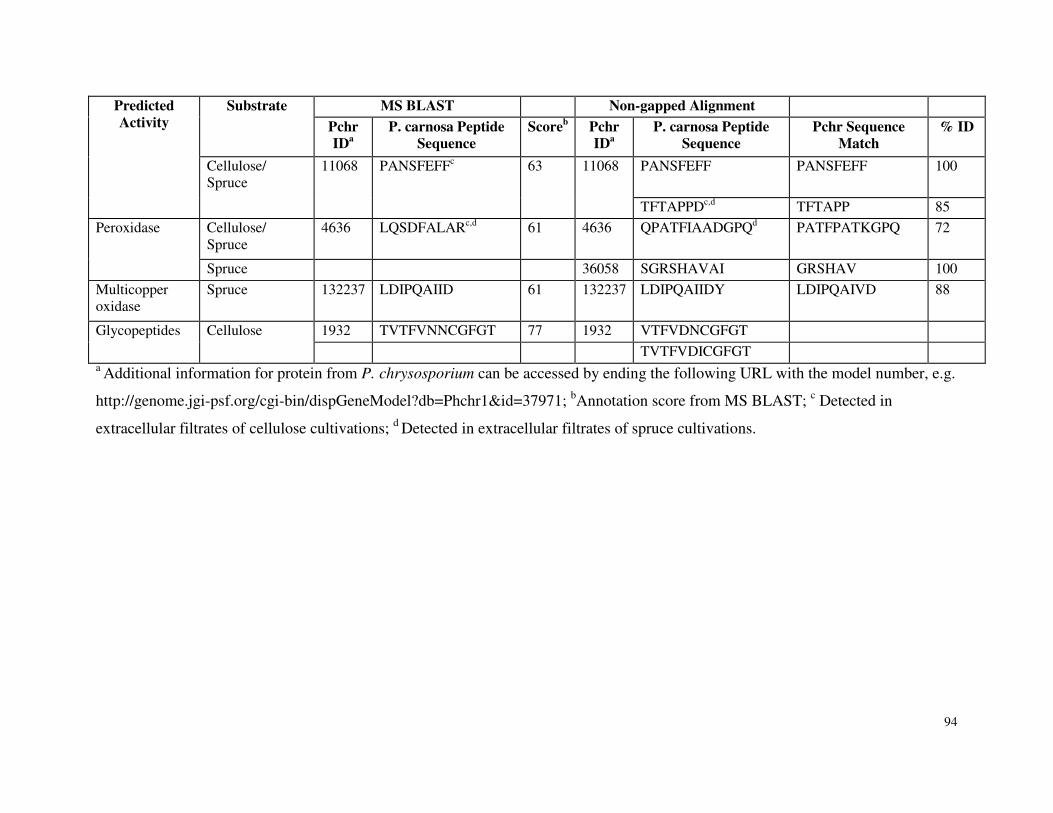
94
Predicted Activity
Substrate
MS BLAST Non-gapped Alignment
Pchr IDa
P. carnosa Peptide Sequence
Scoreb Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
Cellulose/ Spruce
11068
PANSFEFFc
63
11068
PANSFEFF PANSFEFF 100
TFTAPPDc,d TFTAPP 85
Peroxidase
Cellulose/ Spruce
4636 LQSDFALARc,d 61 4636 QPATFIAADGPQd PATFPATKGPQ 72
Spruce 36058 SGRSHAVAI GRSHAV 100
Multicopper oxidase
Spruce 132237 LDIPQAIID 61 132237 LDIPQAIIDY LDIPQAIVD 88
Glycopeptides
Cellulose
1932 TVTFVNNCGFGT 77 1932 VTFVDNCGFGT
TVTFVDICGFGT a Additional information for protein from P. chrysosporium can be accessed by ending the following URL with the model number, e.g.
http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db=Phchr1&id=37971; bAnnotation score from MS BLAST; c Detected in
extracellular filtrates of cellulose cultivations; d Detected in extracellular filtrates of spruce cultivations.

95
5.4.4 Carbohydrate Esterases, Proteases and Other Glycoside Hydrolases
Approximately 10 % of the peptide sequences identified from P. carnosa culture filtrates were
matched to predicted esterases in P. chrysosporium. Lipase and esterase enzymes typically
demonstrate broad substrate specificity, and have been implicated in the hydrolysis of lignin-
carbohydrate complexes. For instance, glucuronoyl esterases were recently identified and shown
to hydrolyze linkages between lignin and glucuonate branching sugars of xylan (Spanikova et al.
2007). Both MS BLAST and non-gapped alignment identified peptides from cellulose and
spruce cultivations that matched Pchr6482, a predicted glucuronoyl esterase from P.
chrysosporium. Non-gapped alignment identified three additional sequences from cellulose
cultivations with greater than 75 % identity to Pchr130517, a second putative glucuronoyl
esterase encoded by P. chrysosporium. Notably, in a separate study, a full-length cDNA
transcript isolated from P. carnosa was predicted to encode a protein that shares over 50 % and
80 % sequence identity to Pchr130517 and Pchr6482, respectively (data not shown).
Peptide sequences that were matched to proteolytic activities accounted for 25 % and 40 % of the
peptides analyzed from cellulose and spruce cultures, respectively (Fig. 5.1). Secreted proteases
could participate in nitrogen cycling and acquisition through hydrolysis of fungal proteins, as
well as plant proteins that are bound to lignocellulose. The relative susceptibility of
lignocellulose degrading enzymes to secreted fungal proteases remains to be elucidated.
In both cellulose and spruce cultivations, the majority of peptide sequences annotated to
extracellular glycoside hydrolases were matched to enzymes involved in cellulose and
hemicellulose degradation. Still, extracellular filtrates of P.carnosa grown on cellulose and
spruce also contained several peptides that were matched to amylases involved in starch
degradation (Table 5.3). MS BLAST and non-gapped alignment also identified peptides
sequences corresponding to enzymes involved in pectin degradation, namely a CE8 pectin
esterase and a GH28 pectinase, respectively (Table 5.3, Appendix 3, Table S5.1).
Similar to P. chrysosporium, P. carnosa appears to secrete a GH88 candidate d-4,5-unsaturated
β-glycosidase during growth on cellulose and spruce (Ravalason et al. 2008; Vanden
Wymelenberg et al. 2009) (Appendix 3, Table S5.1). GH88 d-4,5-unsaturated β-glycosidases

96
have been implicated in the degradation of unsaturated oligosaccharides, likely produced by
polysaccharide lyases (Nankai et al. 1999). Notably, peptides corresponding to a PL14 lyase
were also detected in cellulose cultivations (Table 5.3). Eight unique peptides from P.carnosa
culture filtrates were matched to GH92 α-1,2-mannosidases; only one peptide sequence was
matched to a GH47 α-mannosidase. While GH47 α-mannosidases are thought to participate in
post-translational modification of secreted proteins (Yoshida et al. 1993), most GH92 α-1,2-
mannosidases that have been identified and characterized to date are of bacterial origin.
Nevertheless, P. chrysosporium is predicted to encode four GH92 enzymes, and the Pchr1930
and Pchr133585 protein matches identified in my study correspond to genes that are upregulated
in P. chrysosporium during growth on cellulose (Vanden Wymelenberg et al. 2009).
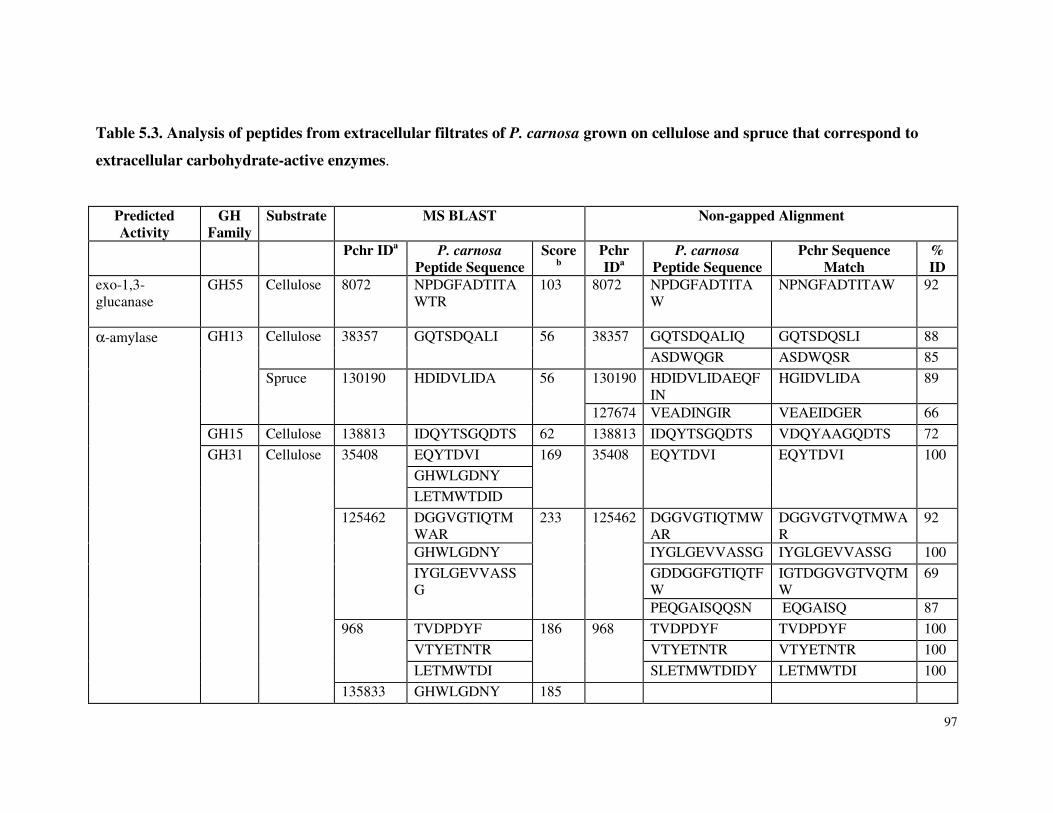
97
Table 5.3. Analysis of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce that correspond to
extracellular carbohydrate-active enzymes.
Predicted Activity
GH Family
Substrate MS BLAST Non-gapped Alignment
Pchr IDa P. carnosa Peptide Sequence
Scoreb
Pchr IDa
P. carnosa Peptide Sequence
Pchr Sequence Match
% ID
exo-1,3-glucanase
GH55
Cellulose
8072
NPDGFADTITAWTR
103
8072 NPDGFADTITAW
NPNGFADTITAW
92
α-amylase
GH13
Cellulose
38357
GQTSDQALI
56
38357
GQTSDQALIQ GQTSDQSLI 88
ASDWQGR ASDWQSR 85
Spruce
130190
HDIDVLIDA
56
130190 HDIDVLIDAEQFIN
HGIDVLIDA 89
127674 VEADINGIR VEAEIDGER 66
GH15 Cellulose 138813 IDQYTSGQDTS 62 138813 IDQYTSGQDTS VDQYAAGQDTS 72
GH31
Cellulose
35408
EQYTDVI 169
35408
EQYTDVI
EQYTDVI
100
GHWLGDNY
LETMWTDID
125462
DGGVGTIQTMWAR
233
125462
DGGVGTIQTMWAR
DGGVGTVQTMWAR
92
GHWLGDNY IYGLGEVVASSG IYGLGEVVASSG 100
IYGLGEVVASSG
GDDGGFGTIQTFW
IGTDGGVGTVQTMW
69
PEQGAISQQSN EQGAISQ 87
968
TVDPDYF 186
968
TVDPDYF TVDPDYF 100
VTYETNTR VTYETNTR VTYETNTR 100
LETMWTDI SLETMWTDIDY LETMWTDI 100
135833 GHWLGDNY 185
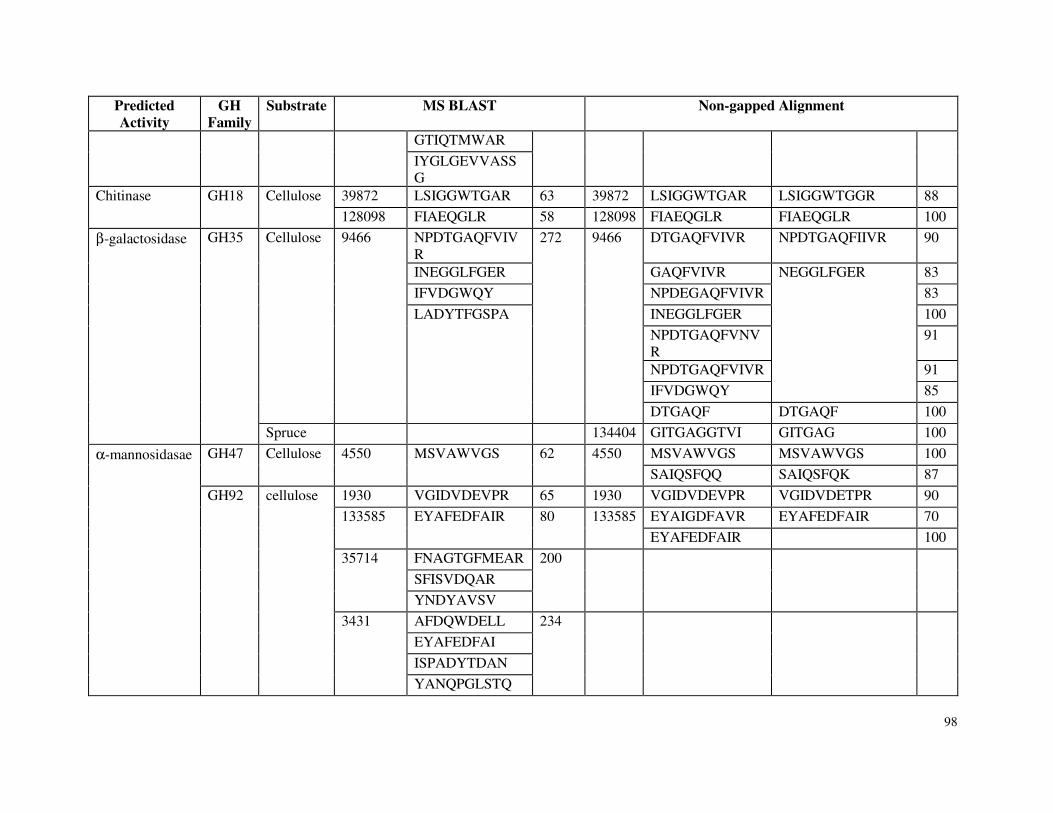
98
Predicted Activity
GH Family
Substrate MS BLAST Non-gapped Alignment
GTIQTMWAR
IYGLGEVVASS
G Chitinase
GH18
Cellulose
39872 LSIGGWTGAR 63 39872 LSIGGWTGAR LSIGGWTGGR 88
128098 FIAEQGLR 58 128098 FIAEQGLR FIAEQGLR 100
β-galactosidase
GH35
Cellulose
9466
NPDTGAQFVIVR
272
9466
DTGAQFVIVR NPDTGAQFIIVR 90
INEGGLFGER GAQFVIVR NEGGLFGER
83
IFVDGWQY NPDEGAQFVIVR 83
LADYTFGSPA
INEGGLFGER 100
NPDTGAQFVNVR
91
NPDTGAQFVIVR 91
IFVDGWQY 85
DTGAQF DTGAQF 100
Spruce 134404 GITGAGGTVI GITGAG 100
α-mannosidasae
GH47
Cellulose
4550
MSVAWVGS
62
4550
MSVAWVGS MSVAWVGS 100
SAIQSFQQ SAIQSFQK 87
GH92
cellulose
1930 VGIDVDEVPR 65 1930 VGIDVDEVPR VGIDVDETPR 90
133585
EYAFEDFAIR
80
133585
EYAIGDFAVR EYAFEDFAIR 70
EYAFEDFAIR 100
35714
FNAGTGFMEAR 200
SFISVDQAR
YNDYAVSV
3431
AFDQWDELL 234
EYAFEDFAI
ISPADYTDAN
YANQPGLSTQ
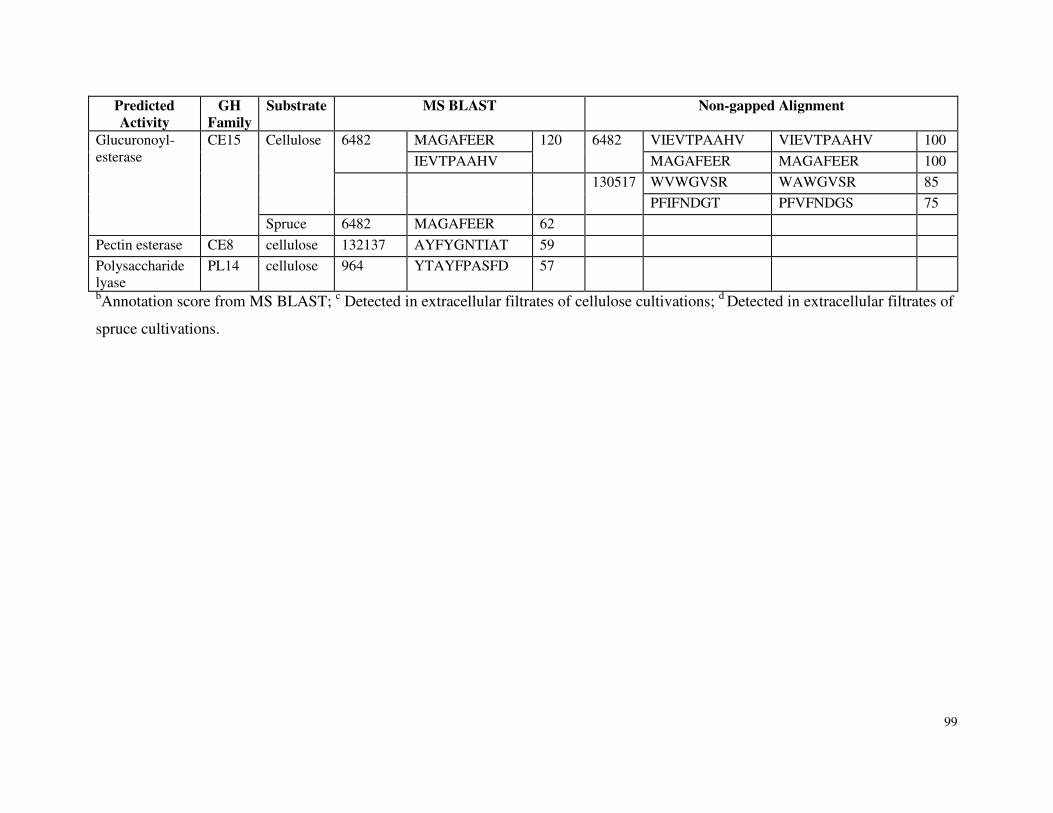
99
Predicted Activity
GH Family
Substrate MS BLAST Non-gapped Alignment
Glucuronoyl-esterase
CE15
Cellulose
6482
MAGAFEER 120
6482
VIEVTPAAHV VIEVTPAAHV 100
IEVTPAAHV MAGAFEER MAGAFEER 100
130517
WVWGVSR WAWGVSR 85
PFIFNDGT PFVFNDGS 75
Spruce 6482 MAGAFEER 62
Pectin esterase CE8 cellulose 132137 AYFYGNTIAT 59
Polysaccharide lyase
PL14 cellulose 964 YTAYFPASFD 57
bAnnotation score from MS BLAST; c Detected in extracellular filtrates of cellulose cultivations; d Detected in extracellular filtrates of
spruce cultivations.

100
5.4.5 Peptides Annotated to Proteins with Unknown Function, Identified in Proteomic
Analyses of P. chrysosporium
An important outcome of proteomic analysis is the detection of proteins with unknown function
whose expression can be correlated to specific cultivation conditions. Seven unique peptide
sequences from cellulose cultivations were assigned to six P. chrysosporium proteins that have
predicted signal sequences but unknown function (Table 5.4). Matches to Pchr3328 and
Pchr138739 were to proteins previously identified in culture supernatant of P. chrysosporium
grown on spruce and cellulose (Vanden Wymelenberg et al. 2006; Ravalason et al. 2008; Vanden
Wymelenberg et al. 2009). As noted by Vanden Wymelenberg et al. (2006), Pchr138739 shares
homology to proteins also found in filamentous Ascomycetes. Further, the carboxy-terminus of
Pchr138739 contains a series of proline residues and is rich in serine and threonine, reminiscent
of linker sequences that connect catalytic domains and carbohydrate binding modules.
Table 5.4. Peptides from extracellular filtrates of P. carnosa grown on cellulose that were
annotated to proteins from P. chrysosporium with unknown function.
Pchr IDa P. carnosa Peptide Sequence Scoreb
5038 SNIEDLTIR 57
4562 IPGAVQVYGIES 61
3328
ISDTDFSQR 126
QVIEWTDID
3053 TIIDGDDFT 56
1934 EDDVPSMSA 57
138739 NPADTSETDGIR 69
a Additional information for protein from P. chrysosporium can be accessed by ending the
following URL with the model number, e.g. http://genome.jgi-psf.org/cgi-
bin/dispGeneModel?db=Phchr1&id=5038; bAnnotation score from MS BLAST.

101
5.5 Discussion
Proteomic analyses of wood-degrading microorganisms reveal a rich repertoire of biomass-
degrading proteins, many of which are yet to be characterized. In addition to identifying new
families of carbohydrate-active enzymes, correlating substrate composition to profile of secreted
proteins can inform the design of enzyme formulations that are most suitable for processing
specific biomass feedstocks. Boreal forests represent one of the largest biomes on Earth, and
bioprocessing softwood remains a significant challenge. Accordingly, the aim of this study was
to characterize the secretome of P.carnosa grown on spruce and microcrystalline cellulose to i)
uncover potential adaptations that allow this white-rot fungus to efficiently grow on softwood
feedstocks, and ii) evaluate substrate-dependent protein secretion by this organism. Given the
phylogenetic similarity of P. carnosa and P. chrysosporium, the genome sequence of P.
chrysosporium could be used to annotate sequenced peptides from P.carnosa; comparative
proteomics was also enabled.
A comparison of peptide annotations from both cellulose and spruce cultivations of P. carnosa to
proteomic data for P. chrysosporium reveals their potential to secrete similar families of
cellulolytic and lignin-degrading enzymes. For example, cellobiohydrolases, endoglucanases,
and β-glycosidases were identified in P. carnosa secretomes, as were peptide sequences
corresponding oxidative enzymes involved in lignin degradation. The relative abundance of
GH7 peptide sequences compared to GH6 peptide sequences that were identified in the P.
carnosa secretome suggests that like P. chrysosporium, GH7 isozymes are more highly
represented in P. carnosa than GH6. Also similar to P. chrysosporium, xylanolytic enzymes,
including GH10 and GH11 xylanases, debranching hydrolases, and glucuronoyl esterases were
identified in extracellular filtrates of P.carnosa cultivations. Notably, while GH2 β-
mannosidases have not been detected in extracellular filtrates of P. chrysosporium grown on
cellulosic substrates, a GH2 β-mannosidase was predicted in extracellular filtrates of P. carnosa
grown on cellulose and spruce. Since mannan is the main hemicellulose in softwood fibre, this
result raises the question whether up-regulation of mannan-degrading enzymes could contribute
to preferential growth of P. carnosa on softwood.

102
P. carnosa peptide sequences were also matched to a multicopper oxidase, Cir1, and two distinct
glycopeptides. These proteins have been implicated in lignocellulose degradation, but have not
been identified in previous proteomic analyses of P. chrysosporium. Cir1 and certain membrane-
bound glycopeptides are likely involved in generating hydroxyl radicals that participate in
carbohydrate and lignin degradation. Given the lack of peptide sequences from P. carnosa
extracellular filtrates that were matched to exclusively endo-acting cellulases, an intriguing
possibility is that oxidative enzymes play a significant role in cellulose degradation by P.
carnosa.
The range of predicted glycoside hydrolases detected in the culture medium of P. carnosa grown
on spruce was represented in the culture medium of P. carnosa grown on cellulose. These
results are consistent with those reported by Sato et al. (2007) and Vanden Wymelenberg et al.
(2005) who detect similar profiles of cellulases and hemicellulases in culture supernatants of P.
chrysosporium grown on cellulose and wood substrates. Moreover, these results support the
possibility that like P. chrysosporium, P. carnosa optimizes its growth on different
lignocellulosic substrates by modifying the relative abundance of a similar pool of glycoside
hydrolases and oxidative enzymes. Transcriptional analyses are currently underway to test this
hypothesis. Quantitative transcription profiles will also complement proteomic analyses by
narrowing the set of proteins with unknown function that are most important to lignocellulose
degradation. Still, 1229 peptides were unique to spruce cultivations and could not be annotated
based on identity to predicted proteins from P. chrysopsorium. Many of these will correspond to
intracellular proteins that are detected in culture supernatants due to cell lysis. However, some
will likely correspond to secreted or intracellular proteins involved in the hydrolysis or
metabolism of softwood components that enable the growth of P. carnosa on this feedstock. For
comparison, MS BLAST was used to align peptide sequences from P. carnosa to P. placenta
protein models. This analysis did not lead to the identification of additional GH families or
lignin-degrading enzymes in extracellular filtrates of P. carnosa grown on cellulose or spruce.
Notably, peptides from cellulose cultivations were most frequently matched to predicted GH31
enzymes in P. placenta, while peptides from spruce cultivations were most frequently matched
to predicted proteases. While the number of peptide matches to a particular protein family
provides some indication of the relative abundance of that family in culture supernatants,

103
differences in digestibility by trypsin, and subsequent ionization and detection by LC/MS limits
quantificative comparisons of proteins families detected in the current analysis.
In summary, proteomic analysis of extracellular filtrates of P. carnosa grown on cellulose and
spruce revealed a similar range of glycoside hydrolases and oxidative activities. The distribution
of cellulases and xylanases predicted in extracellular filtrates of P. carnosa were also similar to
those identified in cultures of P. chrysosporium. However, the profile of secreted proteins
detected in cultures of P. carnosa could be distinguished by the prediction of mannan-degrading
enzymes and oxidative enzymes that were not previously identified in proteomic analysis of P.
chrysosporium.
Clearly, the similarity of P. carnosa and P. chrysoporium was sufficient to apply a homology
driven proteomic approach to identify a core set of carbohydrate-active and lignin-degrading
enzymes that are secreted by P. carnosa grown on cellulosic substrates. However, in December,
2010, the P. carnosa genome sequence was completed and annotated using automated
algorithms, and so the proteomic data collected for P. carnosa was re-analyzed to determine
whether additional carbohydrate-active or lignin-degrading enzymes could be identified.
Through this analysis, additional 116 and 11 peptides that were predicted to originate from
lignocellulose-active enzymes were annotated from cellulose and spruce samples, respectively
(Appendix 3, Tables S5.2 and S5.3). Interestingly, new assignments that corresponded to lignin-
degrading activities were comparatively high in peptide samples originating from spruce
cultures. Is is anticipated that proteins identified using the P.carnosa genome but not by
comparison to P. chrysosoporium, comprise a starting point for the discovery of novel enzymes
that could accelerate the performance of a core set of lignocellulose-degrading enzymes on
softwood feedstocks.
Chapter 5 References
Abbas A, Koc H, Liu F, Tien M (2005) Fungal degradation of wood: initial proteomic analysis of extracellular proteins of Phanerochaete chrysosporium grown on oak substrate. Curr Gen 47: 49-56

104
Baldrian P, Valaskova V (2008) Degradation of cellulose by basidiomycetous fungi. FEMS Microbiol Rev 32: 501-521
Cock PJ, Antao T, Chang JT, Chapman BA, Cox CJ, Dalke A, Friedberg I, Hamelryck T, Kauff
F, Wilczynski B, de Hoon MJ (2009) Biopython: freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 25: 1422-1423
Dancik V, Addona TA, Clauser KR, Vath JE, Pevzner PA (1999) De novo peptide sequencing
via tandem mass spectrometry. J Comput Biol 6: 327-342 de Koker TH, Nakasone KK, Haarhof J, Burdsall Jr HH, Janse BJ (2003) Phylogenetic
relationships of the genus Phanerochaete inferred from the internal transcribed spacer region. Mycol Res 107: 1032-1040
de Vries RP, Kester HCM, Poulsen CH, Benen JAE, Viser J (2000) Synergy between enzymes
from Aspergillus involved in the degradation of plant cell wall polysaccharides. Carbohydrate Res 327: 401-410
Hibbett DS, Donoghue MJ (2001) Analysis of character correlations among wood decay
mechanisms, mating systems, and substrate ranges in homobasidiomycetes. Sys Biol 50: 215-242
Ishida T, Yaoi K, Hiyoshi A, Igarashi K, Samejima M (2007) Substrate recognition by glycoside
hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete
chrysosporium. FEBS J 274: 5727-5736 Junqueira M, Spirin V, Balbuena TS, Thomas H, Adzhubei I, Sunyaev S, Shevchenko A (2008)
Protein identification pipeline for the homology-driven proteomics. J Proteomics 71: 346-356
Karkehabadi S, Hansson H, Kim S, Piens K, Mitchinson C, Sandgren, M (2008) The first
structure of a glycoside hydrolase family 61 member, Cel61B from Hypocrea jecorina, at 1.6 A resolution. J Mol Biol 383: 144-154
Kersten P, Cullen D (2007) Extracellular oxidative systems of the lignin-degrading
Basidiomycete Phanerochaete chrysosporium. Fungal Genet Biol 44: 77-87 Kirk TK, Schultz E, Connors WJ, Lorenz LF, Zeikus JG (1978) Influence of culture parameters
on lignin metabolism by Phanerochaete chrysosporium. Arch Microbiol 117: 277-285 Larrondo LF, Salas L, Melo F, Vicuna R, Cullen D (2003) A novel extracellular multicopper
oxidase from Phanerochaete chrysosporium with ferroxidase activity. Appl Environ Microbiol 69: 6257-6263

105
Martinez AT, Speranza M, Ruiz-Duenas FJ, Ferreira P, Camarero S, Guillen F, Martínez MJ, Gutiérrez A, del Río JC (2005) Biodegradation of lignocellulosics: microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int Microbiol 8: 195-204
Martinez D, Larrondo LF, Putnam N, Gelpke MD, Huang K, Chapman J, Helfenbein KG,
Ramaiya P, Detter JC, Larimer F, Coutinho PM, Henrissat B, Berka R, Cullen D, Rokhsar D (2004) Genome sequence of the lignocellulose degrading fungus Phanerochaete chrysosporium strain RP78. Nat Biotechnol 22: 695-700
Martinez D, Challacombe J, Morgenstern I, Hibbett D, Schmoll M, Kubicek CP, Ferreira P, Ruiz-Duenas FJ, Martinez AT, Kersten P, Hammel KE, Vanden Wymelenberg A, Gaskell J, Lindquist E, Sabat G, Bondurant SS, Larrondo LF, Canessa P, Vicuna R, Yadav J, Doddapaneni H, Subramanian V, Pisabarro AG, Lavín JL, Oguiza JA, Master E, Henrissat B, Coutinho PM, Harris P, Magnuson JK, Baker SE, Bruno K, Kenealy W, Hoegger PJ, Kües U, Ramaiya P, Lucas S, Salamov A, Shapiro H, Tu H, Chee CL, Misra M, Xie G, Teter S, Yaver D, James T, Mokrejs M, Pospisek M, Grigoriev IV, Brettin T, Rokhsar D, Berka R, Cullen D (2009) Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proc Natl Acad Sci USA 106: 1954-1959
Nankai H, Hashimoto W, Miki H, Kawai S, Murata K (1999) Microbial system for
polysaccharide depolymerization: enzymatic route for xanthan depolymerization by Bacillus sp. strain GL1. Appl Environ Microbiol 65: 2520-2526
Plassard CS, Mousain DG, Salsac LE (1982) Estimation of mycelial growth of basidiomycetes
by means of chitin determination. Phytochem 21: 345-348 Ravalason H, Jan G, Molle D, Pasco M, Coutinho PM, Lapierre C, Pollet B, Bertaud F, Petit-
Conil M, Grisel S, Sigoillot JC, Asther M, Herpoël-Gimbert I (2008) Secretome analysis of Phanerochaete chrysosporium strain CIRM-BRFM41 grown on softwood. Appl Microbiol Biotechnol 80: 719-733
Sato S, Liu F, Koc H, Tien M (2007) Expression analysis of extracellular proteins from
Phanerochaete chrysosporium grown on different liquid and solid substrates. Microbiol 153:3023-3033
Sato S, Feltus FA, Iyer P, Tien M (2009) The first genome-level transcriptome of the wood-
degrading fungus Phanerochaete chrysosporium grown on red oak. Curr Genet 55: 273-286
Shevchenko A, Wilm M, Vorm O, Mann M (1996) Mass spectrometric sequencing of proteins
silver-stained polyacrylamide gels. Anal Chem 68: 850-858 Shevchenko A, Sunyaev S, Loboda A, Shevchenko A, Bork P, Ens W, Standing KG (2001)
Charting the proteomes of organisms with unsequenced genomes by MALDI-quadrupole time-of-flight mass spectrometry and BLAST homology searching. Anal Chem 73: 1917-1926

106
Spanikova S, Polakova M, Joniak D, Hirsch J, Biely P (2007) Synthetic esters recognized by
glucuronoyl esterase from Schizophyllum commune. Arch Microbiol 188: 185-189 Tanaka H, Yoshida G, Baba Y, Matsumura K, Wasada H, Murata J, Agawa M, Itakura S, Enoki,
A (2007) Characterization of a hydroxyl-radical-producing glycoprotein and its presumptive genes from the white-rot basidiomycete Phanerochaete chrysosporium. J Biotechnol 128: 500-511
Vanden Wymelenberg AV, Sabat G, Martinez D, Rajangam AS, Teeri TT, Gaskell J, Kersten PJ,
Cullen D (2005) The Phanerochaete chrysosporium secretome: Database predictions and initial mass spectrometry peptide identifications in cellulose-grown medium. J. Biotechnol 118: 17-34.
Vanden Wymelenberg A, Minges P, Sabat G, Martinez D, Aerts A, Salamov A, Grigoriev I,
Shapiro H, Putnam N, Belinky P, Dosoretz C, Gaskell J, Kersten P, Cullen D (2006) Computational analysis of the Phanerochaete chrysosporium v2.0 genome database and mass spectrometry identification of peptides in ligninolytic cultures reveal complex mixtures of secreted protein. Fungal Genet Biol 43(5):343-356
Vanden Wymelenberg A, Gaskell J, Mozuch, M., Kersten, P., Sabat, G., Martinez, D., Cullen, D.
(2009) Transcriptome and secretome analyses of Phanerochaete chrysosporium reveal complex patterns of gene expression. Appl Environ Microbiol 75: 4058-4068
Yoshida M, Igarashi K, Wada M, Kaneko S, Suzuki N, Matsumura H, Nakamura N, Ohno H,
Samejima M (2005) Characterization of carbohydrate-binding cytochrome b562 from the white-rot fungus Phanerochaete chrysosporium, Appl Environ Microbiol 71: 4548-4555
Yoshida T, Inoue T, Ichishima E (1993) 1,2-alpha-D-mannosidase from Penicillium citrinum:
molecular and enzymic properties of two isoenzymes. Biochem J 290 ( Pt 2): 349-354

107
Chapter 6: Synthesis and Conclusions
Four hypotheses were primary addressed through this study:
1. Cultivation conditions that promote the expression of lignocellulytic enzymes in P.
chrysosporium will also induce the expression of lignocellulytic enzymes in P. carnosa.
2. P. carnosa will exhibit selective decay of lignin and the extent of this decay will depend on
wood fibre composition.
3. Because of its isolation from softwood species, P. carnosa will efficiently degrade guaiacyl
lignin moiety, which dominates softwood lignin.
4. The extracellular protein activities secreted by P. carnosa to degrade various lignocellulosic
feedbstocks will differ depending on the particular chemistry of the lignocellulosic substrate.
Figure 6.1 describes schematically the synthesis of information flow between the three research
objectives.
To characterize the impact of P.carnosa growth on various lignocellulose substrates and to
understand the mode of lignocellulose decay adopted by P. carnosa, there was a need to initially
establish the growth conditions for P. carnosa. Through a series of experiments, it was
established that P. carnosa grows well at 27˚C and is capable of lignocellulolytic activity
expression under nitrogen limiting conditions. These experimental conditions were then used to
investigate the impact of P. carnosa growth on various wood fibres, specifically to confirm that
P.carnosa decays softwood and if so, then how.
Through FTIR and ToF-SIMS investigations, it was validated that P. carnosa initiates softwood
decay through targeting G-lignin. In a recent transcriptomic analyses, the ability of P. carnosa to
degrade softwood was explained by the relatively high number of MnP genes than LiP genes in
P. carnosa as compared to P. chrysosporium (which is essentially a hardwood degrading white-
rot fungus) (MacDonald et al. 2011). The biochemical assays in Chapter 3 also imply similar
results, where significant MnP activity but no LiP activity was detected on various P. carnosa
cultivations on cellulose.

108
Limitations associated with activity-based enzyme discovery particularly in culture supernatants
that contain multiple enzymes with similar activity, was discussed in Chapter 3. Accordingly, a
proteomic method was used to compare the profiles of enzymes secreted by P. carnosa during
growth on cellulose and spruce wood fibre. Notably, differences in the profile of enzymes
secreted by P. carnosa grown on cellulose and spruce were subtler than initially hypothesized.
In particular, activities detected in spruce cultivations were also identified in cellulose
cultivations, and the profile of enzymes secreted by P. carnosa grown on cellulose was more
complex than that secreted by P. carnosa grown on spurce. This result implies that P. carnosa
might regulate the relative abundances of a similar set of lignocellulose-active enzymes rather
than the presense of particular activities.
Figure 6.1 Conclusions and information flow between the three key objectives of this thesis.
Optimal temperature: 27°C Cellulolytic activity is expressed. MnP activity is expressed. LiP activity is not detected.
G-lignin is degraded. Growth of P. carnosa on balsam fir, lodgepole pine and white spruce reveals similar degradation patterns. P.carnosa selectively degrades lignin prior to wood polysaccharides.
Proteins secreted during growth on spruce were also detected in cellulose cultivations. Cellulase enzymes appear to be more abundant in culture supernatant than β-glucosidases. Glyoxal oxidases appear to be more abundant than lignin-degrading peroxidases. An additional 127 peptides were annonated to predicted CAZymes or FOLymes when the P.carnosa genome was used in the analysis.
Chapter 3: Growth on microcrystalline cellulose
Chapter 4: Growth on coniferous wood
Chapter 5: Comparative proteomic analysis of proteins secreted by P.carnosa grown on cellulose and spruce
Optimal
growth
conditions
used
Optimal
growth
conditions
used

109
P. chrysosporium, which is a model white-rot fungus, was used as a benchmark for results obtain
through studies using P. carnosa. Cultivation conditions published for P. chrysosporium were
used to define the initial experimental conditions for P. carnosa (Chapter 3) and the genome
sequence for P. chrysosporium was used to assist with the protein annotation based on sequence
similarity (Chapter 5). While P. carnosa selectively degrades lignin like certain strains of P.
chrysosporium, these organisms differed in terms of optimal growth conditions and expression of
lignin-degrading and carbohydrate-active enzymes. For example, the optimal growth temperature
of P. carnosa is 27˚C, while that of P. chrysosporium is 37˚C. Unlike P. chrysosporium, P.
carnosa’s lignocellulolytic enzyme expression is not severely hampered by high nitrogen and/or
shaking cultivation conditions. P. carnosa is also distinguished by its ability to effectively
degrade softwood lignin. In addition to lignin degradation, the proteomic analyses also suggest
that P. carnosa was actively degrading softwood hemicelluloses. Softwood lignin and
hemicellulose degradation by P. carnosa was supported by the detection of mannosidase and
lignin-degading activities in the secretome of P.carnosa (Chapter 5), which has previously not
been reported for P. chrysosporium.
To conclude, P. carnosa has been established as a softwood degrading white-rot fungus, which is
capable of selectively degrading softwood-lignin. The following list summarizes
recommendations for further research on the lignocellulolytic activity of P. carnosa and wood-
degrading basiodiomycetes in general:
1. Systematic investigation of the effect of growth conditions (rpm, pH, oligosaccharide
addition) on P. carnosa growth and lignocellulolytic activity.
2. Annotation of proteins with unknown function, identified through proteomic analysis of the
secretome of P. carnosa on spruce and micro-crystalline cellulose.
3. Differential proteomics for extracellular enzymes expressed by P.carnosa on other simple
substrates and hardwood species to determine if the conclusion of non-substrate specific
protein expression can be extended.
4. Investigation of changes in fibre chemistry for hardwood species when degraded by P.
carnosa to determine explicitly the differences between conventional white-rot degrading
fungi isolated from hardwoods and P.carnosa.

110
Chapter 7: Engineering Relevance
Growing environmental concerns are requiring that more industrial sectors adopt eco-friendly
processes. Further, with increasing competition from industries located in the Southern
hemisphere, North American and European forest industries are realizing the need to diversify
the range of products that can be derived from wood fibre. Accordingly, there is an opportunity
for technology that harnesses the full potential of biomass for the production of fuels and other
biodegradable materials.
Enzymes are renewable biocatalysts, and several enzymes are already being used by forest
industries to reduce chemical consumption. For instance, approximately 10% of North American
pulp and paper industries use microbial xylanases to boost fibre bleaching processes (Kenealy
and Jeffries 2003; Subramaniyan and Prema 2002). Laccases and manganese peroxidases have
also shown promise for improving pulp bleaching; cellulases have been used during fibre
pulping and to recover fibre from recycled paper products, whereas lipases and pectinases have
been used to remove pitch deposited on paper manufacturing equipment (Bajpai 2004; Kenealy
and Jeffries 2003). The synergistic action of glycosyl hydrolases, including endo and exo-
glucanases and glucosidases have been used to transform cellulosic material to glucose which
can be fermented for production of bioethanol. Enzymes have also been used to alter the
functionality of wood polymers. For example, Gustavsson et al. (2005) used a lipase from
Candida antartica to link chemically modified oligosaccharides to xyloglucan (Gustavsson et al.
2005). The modified xyloglucans hydrogen bond with cellulose, thereby functionalizing
cellulose microfibrils. In this way, chemically reactive and bioactive side groups were grafted to
filter paper.
Despite their potential as industrial catalysts, associated costs have limited the utility of enzymes
for production of commodities. These costs can be attributed to enzyme production, typically
low availability of enzymes to insoluble substrates like plant fibre, and the variable composition
of plant biomass. In particular, the distinct composition and structure of lignocellulosic substrates
contained in plant fibre presents unique challenges for bioconversion technologies. It is possible
that the efficiency of bioconversion technologies could be improved by tailoring enzyme
applications to particular lignocellulosic feedstocks. One approach is to mimic the profiles of

111
enzymes secreted by fungi grown on different lignocellulosic substrates. Accordingly, it is
anticipated that the proteomic analysis performed as part of the current thesis, will help to
formulate ideal enzyme mixtures for softwood conversion. For instance, the various ligninolytic
enzymes expressed by P. carnosa on the softwoods can be eventually purified for pre-treatment
of softwood fibre used to generate mechanical pulp or otherwise sugars for subsequent
fermentation. Alternatively, since P. carnosa has demonstrated ability to selectively remove
lignin from multiple softwood species, this organism could be used for in-situ conversion of
softwood wastes to fermentable sugars for fuels or chemical production.
In addition to potential industrial relevance, contributions to method development that have
broader scientific relevance include:
1. Proteomic Analysis of Uncharacterized Microorganisms: The methodology
established for differential proteomic analysis of P. carnosa can be transferred to other
investigations where the genome sequence for the organism is not available. In addition,
it can also be applied towards metagenomic studies, where microbial consortia are being
studied using incomplete genomic data.
2. Analytical Techniques for Fibre Characterization: The method developed in this
study for investigating fibre properties of wood chips (as opposed to the commonly used
wood powder) through FTIR and ToF-SIMS makes a significant contribution in
monitoring changes in fibre chemistry. This becomes especially relevant since most
industrial processes deal with wood chips. In addition, the method development for ToF-
SIMS using un-embedded samples will also contribute tremendously for ToF-SIMS
based fibre analysis, while preventing unwanted interference with embedding medium.
Also through this thesis, the use of ToF-SIMS for both spectral and spatial analysis of
degraded wood fibre was established. These results have been validated with those from
FTIR and TEM. The use of ToF-SIMS as a single method to obtain information
regarding changes in fibre chemistry and localization of cell wall constituents, will
improve the efficiency and ease of analyzing treated wood fibre.

112
Chapter 7 References Bajpai P (2004) Biological Bleaching of Chemical Pulps. Critical Reviews in Biotechnology 24
(1):1-58 Gustavsson MT, Persson PV, Iversen T, Martinelle M, Hult K, Teeri TT, Brumer H (2005)
Modification of Cellulose Fiber Surfaces by Use of a Lipase and a Xyloglucan Endotransglycosylase. Biomacromolecules 6 (1):196-203.
Kenealy WR, Jeffries TW (2003) Wood deterioration and preservation: advances in our
changing world. In: Society AC (ed), Washington, DC, Oxford University Press, pp 210-239
Subramaniyan S, Prema P (2002) Biotechnology of microbial xylanases: enzymology, molecular
biology, and application. Critical Reviews in Biotechnology 22 (1):33-64

113
Appendix 1: B3 Medium
10X Buffer Recipe (per 1L) 20 g KH2PO4 5g MgSO4-7H2O 1g CaCl2-2H2O pH to 4.5 add milliQ water up to 1L autoclave for 30 min Mineral Elixir (per 1L): 1.5 g nitrilotriacetate 0.5 g MnSO4 1g NaCl 100 mg FeSO4-7H2O 100mg CoSO4 (= 0.645 mM ≡ 180 mg CoSO4-7H2O 100 mg ZnSO4 (= 0.619 mM ≡ 178 mg ZnSO4-7H2O 10 mg CuSO4-5H2O 10mg AlK(SO4)2 10 mg H3BO3 10mg NaMoO4
autoclave 30 min Vitamins 10 mg/ml thiamine HCl filter sterilize Nitrogen Source 1g/10ml ammonium tartrate filter sterilize Carbon Source
0.5 g/ml selected carbon source (if not wood chips), ρglycerol = 1.27 g/ml autoclave 30 min Other 0.73 g/50 ml 2,2-dimethylsuccinic acid (added as a filter sterilized solution adjusted to pH 4.5 using NaOH)
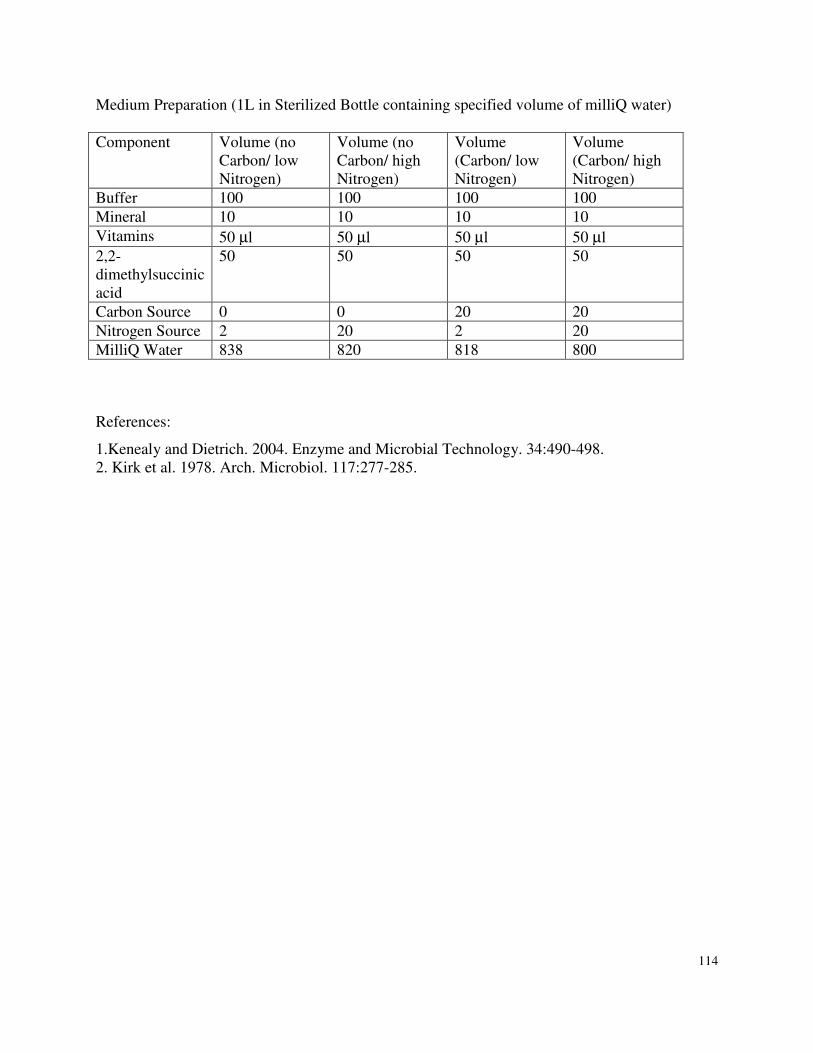
114
Medium Preparation (1L in Sterilized Bottle containing specified volume of milliQ water) Component Volume (no
Carbon/ low Nitrogen)
Volume (no Carbon/ high Nitrogen)
Volume (Carbon/ low Nitrogen)
Volume (Carbon/ high Nitrogen)
Buffer 100 100 100 100 Mineral 10 10 10 10 Vitamins 50 µl 50 µl 50 µl 50 µl 2,2-dimethylsuccinic acid
50 50 50 50
Carbon Source 0 0 20 20 Nitrogen Source 2 20 2 20 MilliQ Water 838 820 818 800
References:
1.Kenealy and Dietrich. 2004. Enzyme and Microbial Technology. 34:490-498. 2. Kirk et al. 1978. Arch. Microbiol. 117:277-285.
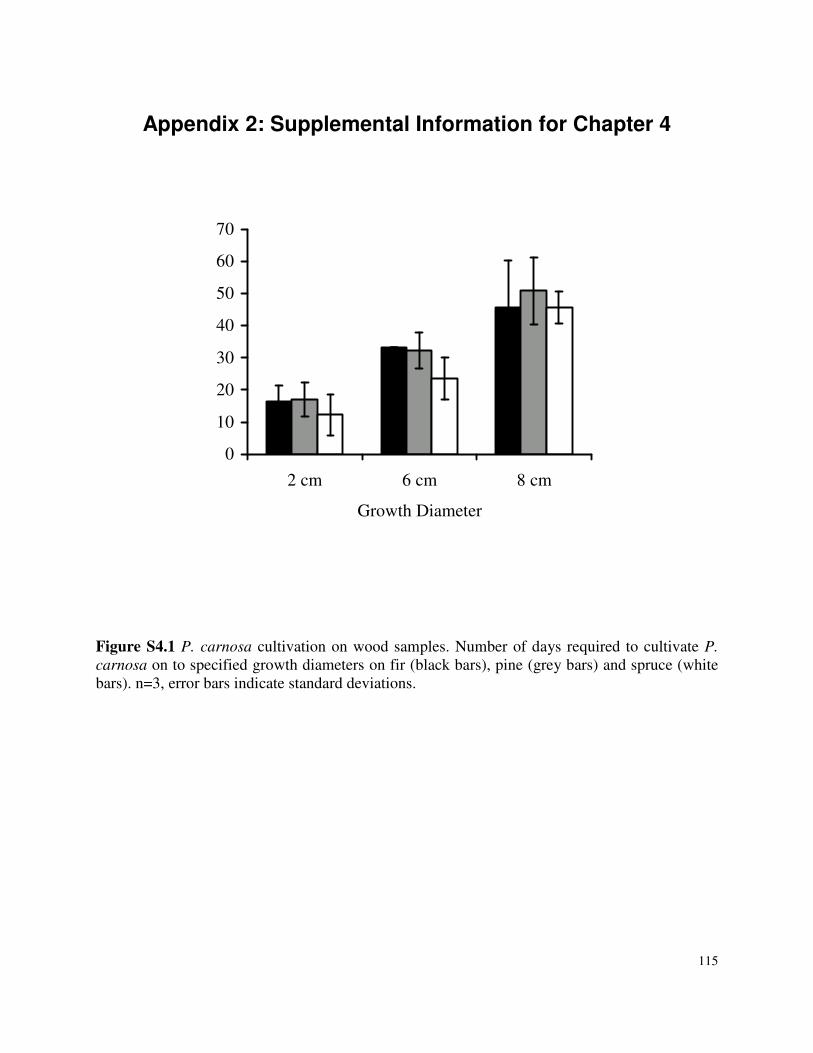
115
Appendix 2: Supplemental Information for Chapter 4
0
10
20
30
40
50
60
70
2 cm 6 cm 8 cm
Growth Diameter
Figure S4.1 P. carnosa cultivation on wood samples. Number of days required to cultivate P.
carnosa on to specified growth diameters on fir (black bars), pine (grey bars) and spruce (white bars). n=3, error bars indicate standard deviations.
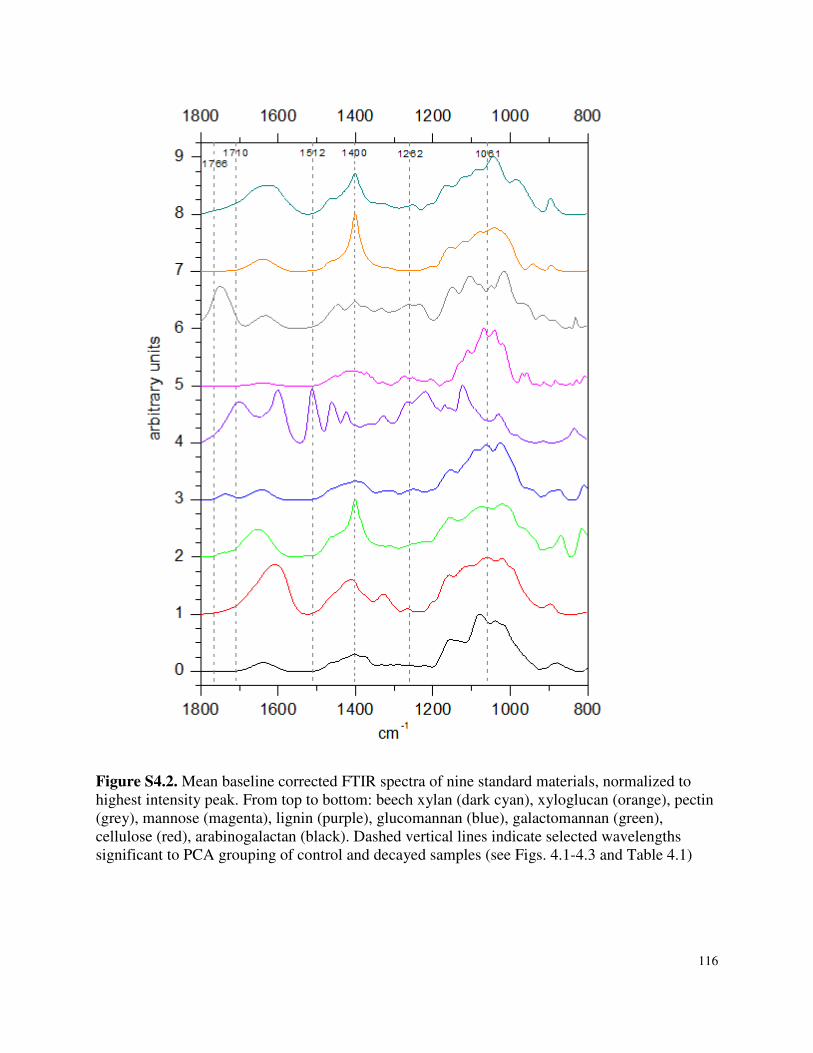
116
Figure S4.2. Mean baseline corrected FTIR spectra of nine standard materials, normalized to highest intensity peak. From top to bottom: beech xylan (dark cyan), xyloglucan (orange), pectin (grey), mannose (magenta), lignin (purple), glucomannan (blue), galactomannan (green), cellulose (red), arabinogalactan (black). Dashed vertical lines indicate selected wavelengths significant to PCA grouping of control and decayed samples (see Figs. 4.1-4.3 and Table 4.1)
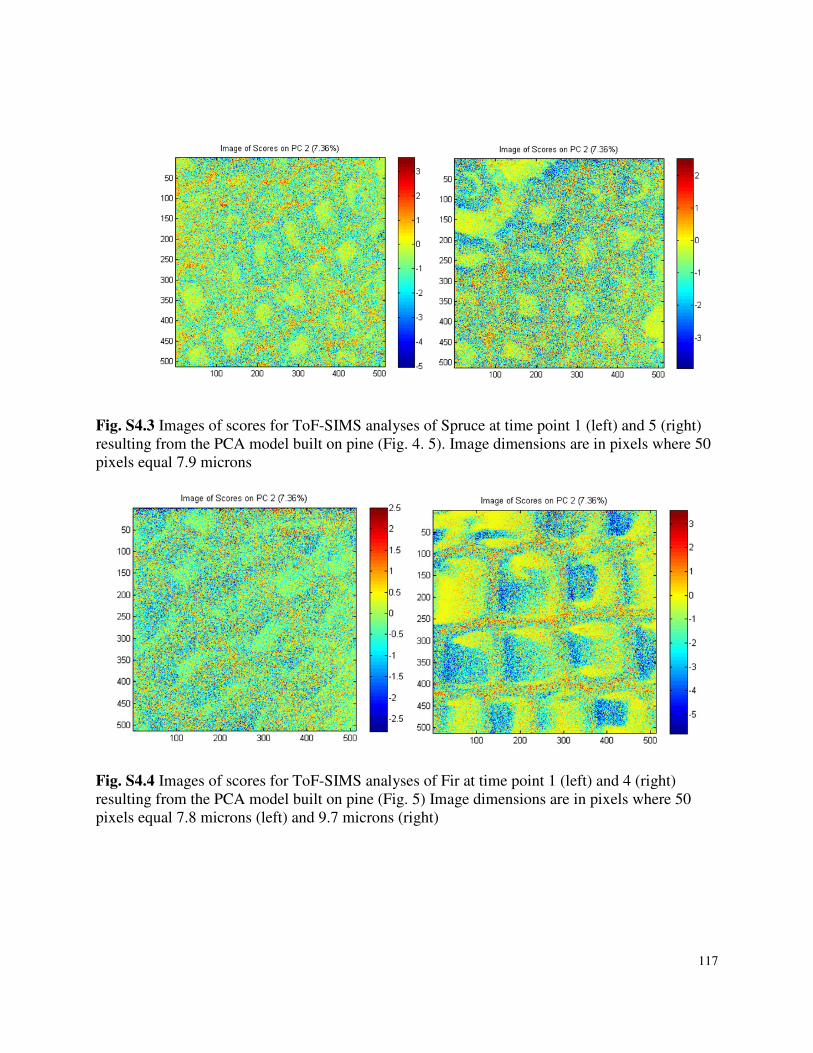
117
Fig. S4.3 Images of scores for ToF-SIMS analyses of Spruce at time point 1 (left) and 5 (right) resulting from the PCA model built on pine (Fig. 4. 5). Image dimensions are in pixels where 50 pixels equal 7.9 microns
Fig. S4.4 Images of scores for ToF-SIMS analyses of Fir at time point 1 (left) and 4 (right) resulting from the PCA model built on pine (Fig. 5) Image dimensions are in pixels where 50 pixels equal 7.8 microns (left) and 9.7 microns (right)

118
Table S4.1. The Ratio of Bromine Concentration in the Middle Lamella to Cell Wall in Spruce
Samples Analyzed by TEM-EDXA
Ratio of Time point 1 Time point 5 1. cell wall: middle lamella 1.63 1.38
1.25 2.42 1.33 1.33
2. cell corner: middle lamella 1.88 2.25 1.39 1.23 1.23 0.852
* t-test was used to determine the statistical significance; p-value = 0.6590 (1) and 0.4876 (2)
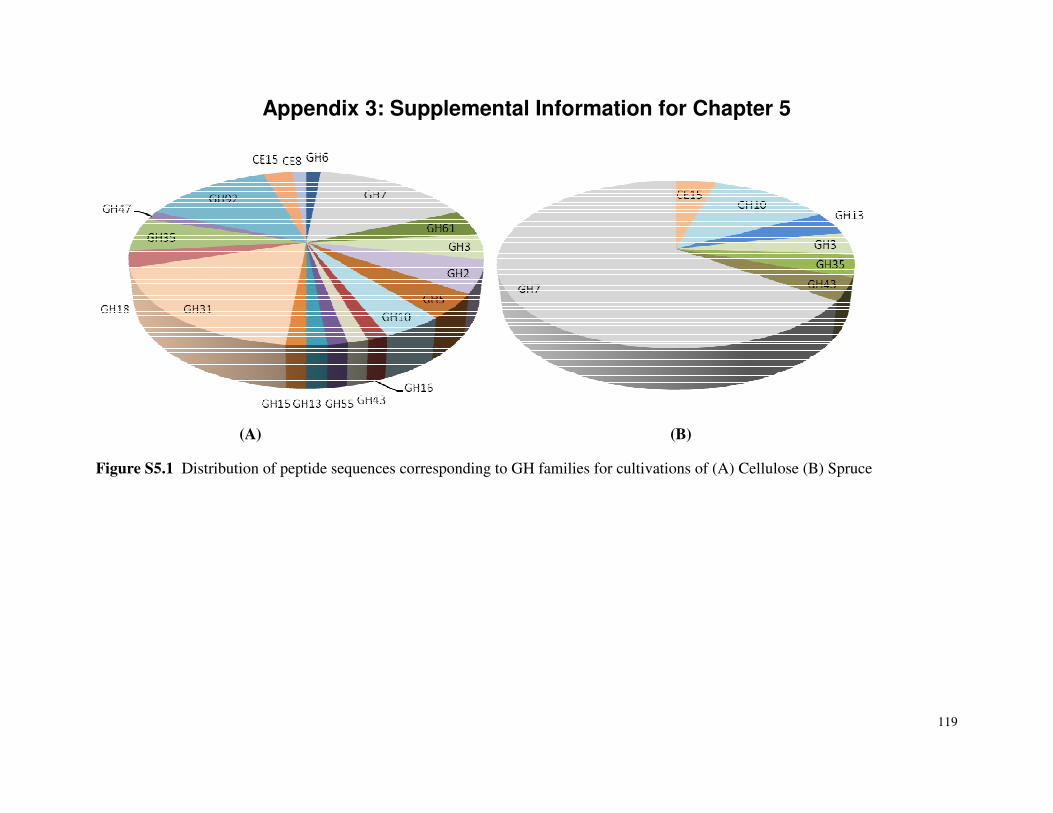
119
Appendix 3: Supplemental Information for Chapter 5
(A) (B)
Figure S5.1 Distribution of peptide sequences corresponding to GH families for cultivations of (A) Cellulose (B) Spruce
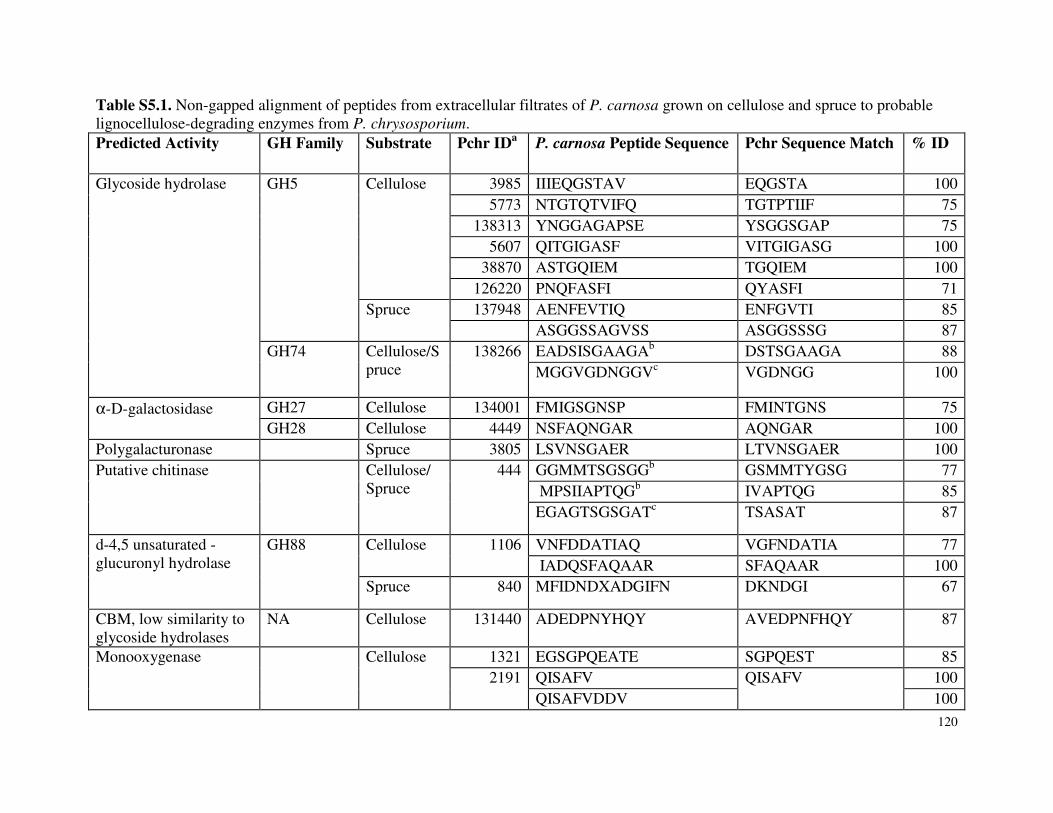
120
Table S5.1. Non-gapped alignment of peptides from extracellular filtrates of P. carnosa grown on cellulose and spruce to probable lignocellulose-degrading enzymes from P. chrysosporium. Predicted Activity GH Family Substrate Pchr IDa P. carnosa Peptide Sequence Pchr Sequence Match % ID
Glycoside hydrolase
GH5
Cellulose
3985 IIIEQGSTAV EQGSTA 100 5773 NTGTQTVIFQ TGTPTIIF 75
138313 YNGGAGAPSE YSGGSGAP 75 5607 QITGIGASF VITGIGASG 100
38870 ASTGQIEM TGQIEM 100 126220 PNQFASFI QYASFI 71
Spruce
137948 AENFEVTIQ ENFGVTI 85 ASGGSSAGVSS ASGGSSSG 87
GH74
Cellulose/Spruce
138266
EADSISGAAGAb DSTSGAAGA 88 MGGVGDNGGVc VGDNGG 100
α-D-galactosidase
GH27 Cellulose 134001 FMIGSGNSP FMINTGNS 75 GH28 Cellulose 4449 NSFAQNGAR AQNGAR 100
Polygalacturonase Spruce 3805 LSVNSGAER LTVNSGAER 100 Putative chitinase
Cellulose/ Spruce
444
GGMMTSGSGGb GSMMTYGSG 77 MPSIIAPTQGb IVAPTQG 85 EGAGTSGSGATc TSASAT 87
d-4,5 unsaturated -glucuronyl hydrolase
GH88
Cellulose
1106
VNFDDATIAQ VGFNDATIA 77 IADQSFAQAAR SFAQAAR 100
Spruce 840 MFIDNDXADGIFN DKNDGI 67
CBM, low similarity to glycoside hydrolases
NA Cellulose 131440 ADEDPNYHQY AVEDPNFHQY 87
Monooxygenase
Cellulose
1321 EGSGPQEATE SGPQEST 85 2191
QISAFV QISAFV
100
QISAFVDDV 100
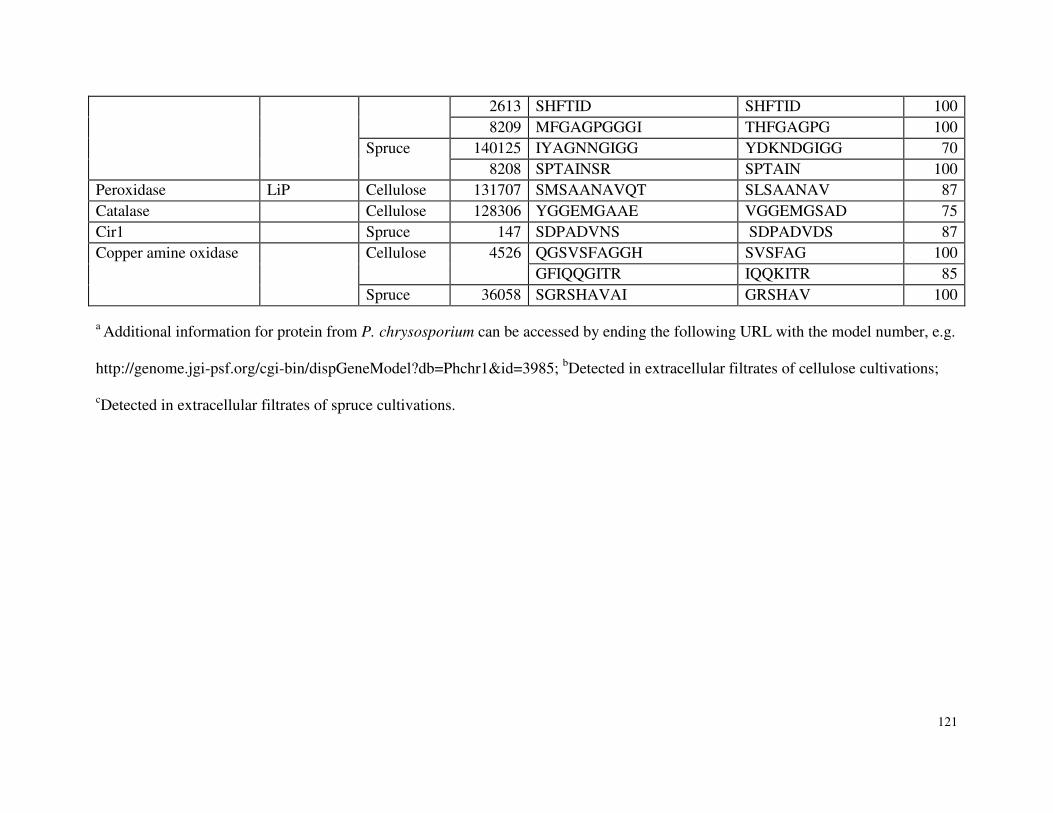
121
2613 SHFTID SHFTID 100 8209 MFGAGPGGGI THFGAGPG 100
Spruce
140125 IYAGNNGIGG YDKNDGIGG 70 8208 SPTAINSR SPTAIN 100
Peroxidase LiP Cellulose 131707 SMSAANAVQT SLSAANAV 87 Catalase Cellulose 128306 YGGEMGAAE VGGEMGSAD 75 Cir1 Spruce 147 SDPADVNS SDPADVDS 87 Copper amine oxidase
Cellulose
4526
QGSVSFAGGH SVSFAG 100 GFIQQGITR IQQKITR 85
Spruce 36058 SGRSHAVAI GRSHAV 100
a Additional information for protein from P. chrysosporium can be accessed by ending the following URL with the model number, e.g.
http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db=Phchr1&id=3985; bDetected in extracellular filtrates of cellulose cultivations;
cDetected in extracellular filtrates of spruce cultivations.
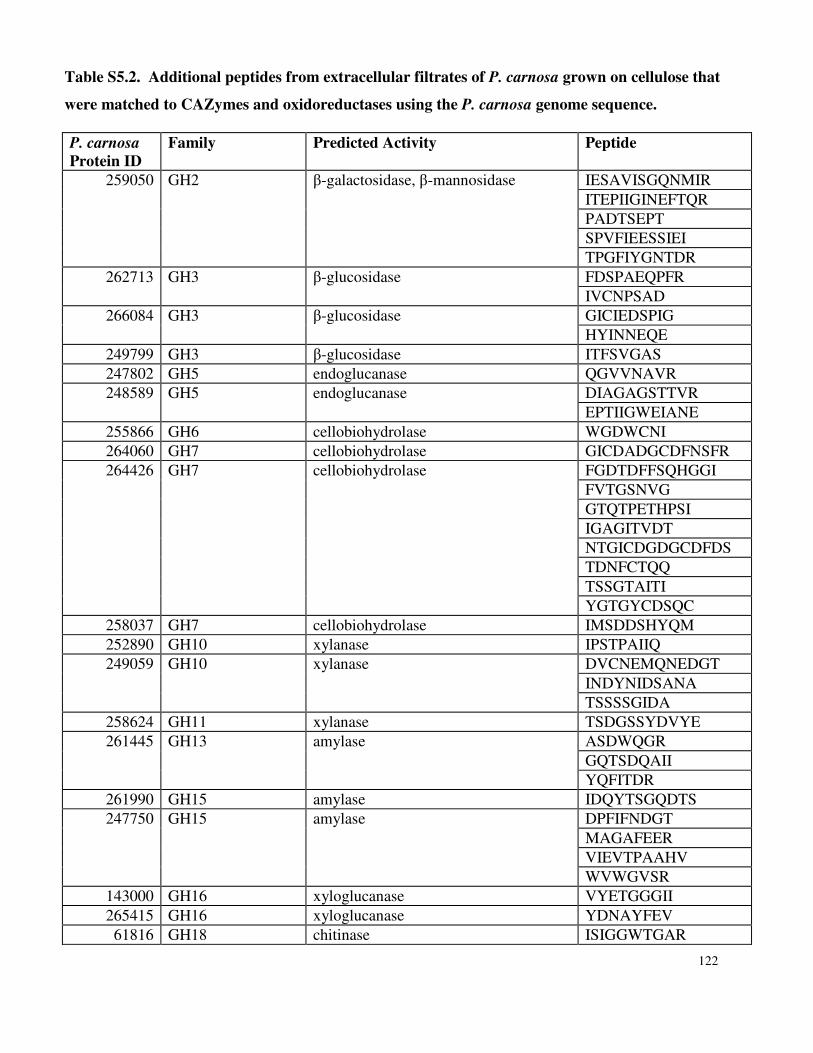
122
P. carnosa Protein ID
Family Predicted Activity Peptide
259050 GH2 β-galactosidase, β-mannosidase IESAVISGQNMIR ITEPIIGINEFTQR PADTSEPT SPVFIEESSIEI TPGFIYGNTDR
262713 GH3 β-glucosidase FDSPAEQPFR IVCNPSAD
266084 GH3 β-glucosidase GICIEDSPIG HYINNEQE
249799 GH3 β-glucosidase ITFSVGAS 247802 GH5 endoglucanase QGVVNAVR 248589 GH5 endoglucanase DIAGAGSTTVR
EPTIIGWEIANE 255866 GH6 cellobiohydrolase WGDWCNI 264060 GH7 cellobiohydrolase GICDADGCDFNSFR 264426 GH7 cellobiohydrolase FGDTDFFSQHGGI
FVTGSNVG GTQTPETHPSI IGAGITVDT NTGICDGDGCDFDS TDNFCTQQ TSSGTAITI YGTGYCDSQC
258037 GH7 cellobiohydrolase IMSDDSHYQM 252890 GH10 xylanase IPSTPAIIQ 249059 GH10 xylanase DVCNEMQNEDGT
INDYNIDSANA TSSSSGIDA
258624 GH11 xylanase TSDGSSYDVYE 261445 GH13 amylase ASDWQGR
GQTSDQAII YQFITDR
261990 GH15 amylase IDQYTSGQDTS 247750 GH15 amylase DPFIFNDGT
MAGAFEER VIEVTPAAHV WVWGVSR
143000 GH16 xyloglucanase VYETGGGII 265415 GH16 xyloglucanase YDNAYFEV
61816 GH18 chitinase ISIGGWTGAR
Table S5.2. Additional peptides from extracellular filtrates of P. carnosa grown on cellulose that
were matched to CAZymes and oxidoreductases using the P. carnosa genome sequence.
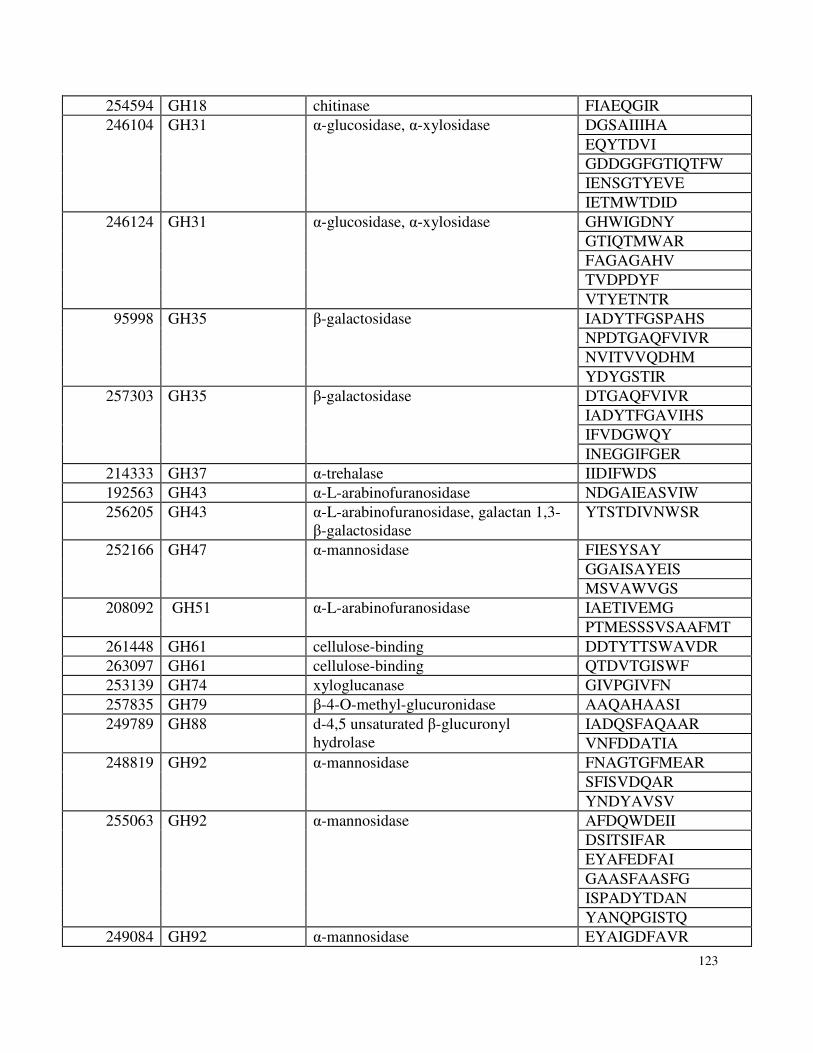
123
254594 GH18 chitinase FIAEQGIR 246104 GH31 α-glucosidase, α-xylosidase DGSAIIIHA
EQYTDVI GDDGGFGTIQTFW IENSGTYEVE IETMWTDID
246124 GH31 α-glucosidase, α-xylosidase GHWIGDNY GTIQTMWAR FAGAGAHV TVDPDYF VTYETNTR
95998 GH35 β-galactosidase IADYTFGSPAHS NPDTGAQFVIVR NVITVVQDHM YDYGSTIR
257303 GH35 β-galactosidase DTGAQFVIVR IADYTFGAVIHS IFVDGWQY INEGGIFGER
214333 GH37 α-trehalase IIDIFWDS 192563 GH43 α-L-arabinofuranosidase NDGAIEASVIW 256205 GH43 α-L-arabinofuranosidase, galactan 1,3-
β-galactosidase YTSTDIVNWSR
252166 GH47 α-mannosidase FIESYSAY GGAISAYEIS MSVAWVGS
208092 GH51 α-L-arabinofuranosidase IAETIVEMG PTMESSSVSAAFMT
261448 GH61 cellulose-binding DDTYTTSWAVDR 263097 GH61 cellulose-binding QTDVTGISWF 253139 GH74 xyloglucanase GIVPGIVFN 257835 GH79 β-4-O-methyl-glucuronidase AAQAHAASI 249789 GH88 d-4,5 unsaturated β-glucuronyl
hydrolase IADQSFAQAAR VNFDDATIA
248819 GH92 α-mannosidase FNAGTGFMEAR SFISVDQAR YNDYAVSV
255063 GH92 α-mannosidase AFDQWDEII DSITSIFAR EYAFEDFAI GAASFAASFG ISPADYTDAN YANQPGISTQ
249084 GH92 α-mannosidase EYAIGDFAVR
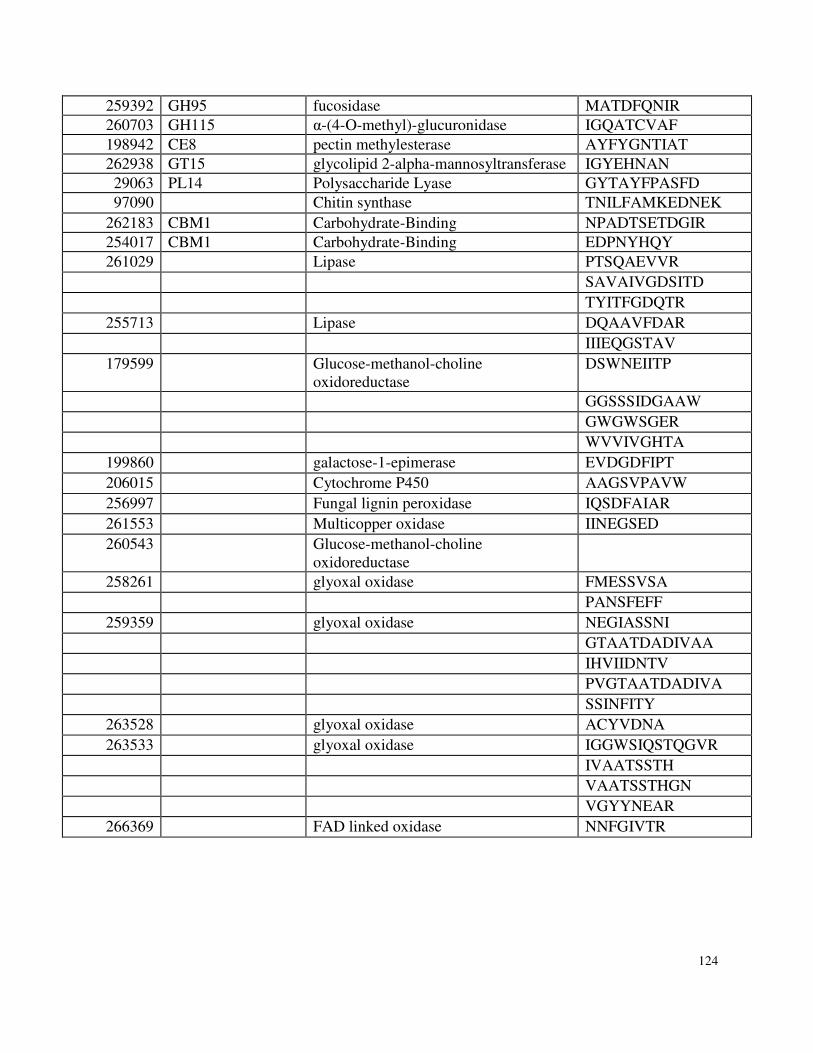
124
259392 GH95 fucosidase MATDFQNIR 260703 GH115 α-(4-O-methyl)-glucuronidase IGQATCVAF 198942 CE8 pectin methylesterase AYFYGNTIAT 262938 GT15 glycolipid 2-alpha-mannosyltransferase IGYEHNAN
29063 PL14 Polysaccharide Lyase GYTAYFPASFD 97090 Chitin synthase TNILFAMKEDNEK
262183 CBM1 Carbohydrate-Binding NPADTSETDGIR 254017 CBM1 Carbohydrate-Binding EDPNYHQY 261029 Lipase PTSQAEVVR
SAVAIVGDSITD TYITFGDQTR
255713 Lipase DQAAVFDAR IIIEQGSTAV
179599 Glucose-methanol-choline oxidoreductase
DSWNEIITP
GGSSSIDGAAW GWGWSGER WVVIVGHTA
199860 galactose-1-epimerase EVDGDFIPT 206015 Cytochrome P450 AAGSVPAVW 256997 Fungal lignin peroxidase IQSDFAIAR 261553 Multicopper oxidase IINEGSED 260543 Glucose-methanol-choline
oxidoreductase
258261 glyoxal oxidase FMESSVSA PANSFEFF
259359 glyoxal oxidase NEGIASSNI GTAATDADIVAA
IHVIIDNTV PVGTAATDADIVA SSINFITY
263528 glyoxal oxidase ACYVDNA 263533 glyoxal oxidase IGGWSIQSTQGVR
IVAATSSTH VAATSSTHGN VGYYNEAR
266369 FAD linked oxidase NNFGIVTR
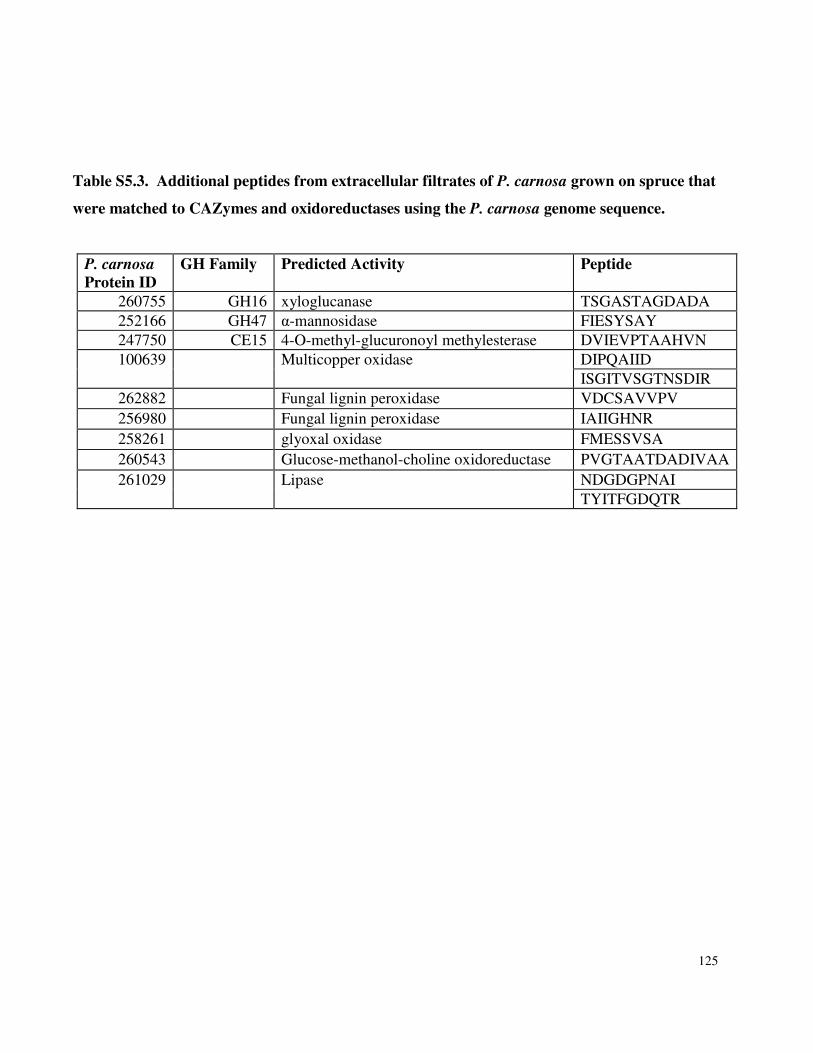
125
P. carnosa Protein ID
GH Family Predicted Activity Peptide
260755 GH16 xyloglucanase TSGASTAGDADA 252166 GH47 α-mannosidase FIESYSAY 247750 CE15 4-O-methyl-glucuronoyl methylesterase DVIEVPTAAHVN 100639 Multicopper oxidase DIPQAIID
ISGITVSGTNSDIR 262882 Fungal lignin peroxidase VDCSAVVPV 256980 Fungal lignin peroxidase IAIIGHNR 258261 glyoxal oxidase FMESSVSA 260543 Glucose-methanol-choline oxidoreductase PVGTAATDADIVAA 261029 Lipase NDGDGPNAI
TYITFGDQTR
Table S5.3. Additional peptides from extracellular filtrates of P. carnosa grown on spruce that
were matched to CAZymes and oxidoreductases using the P. carnosa genome sequence.

126
Appendix 4: Substrate recognition and hydrolysis by a fungal xyloglucan-specific family 12 hydrolase
Justin Powlowski, Sonam Mahajan, Matthieu Schapira and Emma R. Master
Abstract
Biochemical studies to elucidate the structural basis for xyloglucan specificity among GH12
xyloglucanases are lacking. Accordingly, the substrate specificity of a GH12 xyloglucanase from
Aspergillus niger (AnXEG12A) was investigated using pea xyloglucan and 12 xylogluco-
oligosaccharides, and data were compared to a structural model of the enzyme. The specific
activity of AnXEG12A with pea xyloglucan was 113 µmol min−1 mg−1, and apparent kcat and Km
values were 49 s−1 and 0.54 mg mL−1, respectively. These values are similar to previously
published results using xyloglucan from tamarind seed, and suggest that substrate fucosylation
does not affect the specific activity of this enzyme. AnXEG12A preferred xylogluco-
oligosaccharides containing more than six glucose units, and with xylose substitution at the −3
and +1 subsites. The specific activities of AnXEG12A on 100 µM XXXGXXXG and 100 µM
XLLGXLLG were 60 ± 4 and 72 ± 9 µmol min−1 mg−1, respectively. AnXEG12A did not
hydrolyze XXXXXXXG, consistent with other data that demonstrate the requirement for an
unbranched glucose residue for hydrolysis by this enzyme.
Graphical abstract
Keywords: Xyloglucan; Oligosaccharides; Xyloglucanase; Glycoside hydrolase; Kinetics
Abbreviations: MALDI, matrix-assisted laser desorption ionization; XEG, xyloglucan endoglucanase; Glc, glucose







