Download - Binding of soybean agglutinin to glycolipid components of porcine lymphocyte plasma membranes

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 216, No. 2, July, pp. 685-692, 1982
Binding of Soybean Agglutinin to Glycolipid Components of Porcine Lymphocyte Plasma Membranes’
HARVEY J. SAGE,2 LYNN D. YATES,3 AND CURLEY B. HORTON
VA Hospital and Bicchemiatry Departmat, Duke University Medical Center, Durham, North Carolina 27705
Received December 14, 1981, and in revised form February 28, 1982
‘l-Labeled soybean agglutinin binds primarily to glycolipids contained in pig lym- phocyte plasma membranes as measured by in situ “staining” of membranes subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Separation of these gly- colipids by differential extraction, silicic acid chromatography, and high-performance thin-layer chromatography showed that three different species of plasma membrane glycolipid bind this lectin; trihexosyl ceramide, globoside, and ganglioside Gm in order of increasing affinity (over a range of lo- to 20-fold). Trihexosyl ceramide and globoside, major neutral membrane glycolipids, are the major binders; while GY2, a minor acidic glycolipid, is a quantitatively smaller lectin-binding component.
micelle with the tracking dye (3). Soybean
Plasma membranes (PM)* from porcine lymph node lymphocytes contain two dis-
agglutinin (SBA) is unique among re-
tinct classes of covalent carbohydrate complexes; glycopolypeptides and glyco- lipids (1, 2). When PM are dissociated in sodium dodecyl sulfate (SDS) a variety of lectins bind to a large array of PM gly- copolypeptides and to one or more glyco- lipids which, by polyacrylamide gel elec- trophoresis, migrate as a small mixed
ported lectins in binding almost exclu- sively to the glycolipid fraction of PM (3). Because this material appears to be the major lymphocyte surface receptor for SBA (4) we have attempted to identify which particular glycolipids present in PM bind SBA, their relative amounts, and af- finities for the lectin.
MATERIALS AND METHODS
i Part of this work is taken from a thesis by L.D.Y. in partial fulfilment of the Ph.D. degree in Biochem- istry, Duke University, 19’79.
2Author to whom all correspondence should be sent.
a Current address: Department of Microbiology, Duke University Medical Center, Durham, N. C. 27710.
’ Abbreviations used: SBA, soybean agglutinin;
Gni., Gnib, Gm, Gus, and bovine brain mixed gan- gliosides were from Supelco (Bellefonte, Pa). Addi- tional samples of gangliosides were kindly provided
Purified globoside (GL-4), trihexosyl ceramide (GL- 3), glucosyl ceramide (Glc-Cer), gangliosides Gun
by Drs. Eric Bremer, Ellen Grollman, Bernard Kauf- man, and James Rearick.
PM were prepared as described previously (3). PM total lipids (TL), neutral lipids (NL), and gangliosides (GS) were prepared by published procedures (5, 6) with a delipo protein-containing residue (AP). Pro- tein content of PM fractions was measured by mi-
PM, plasma membranes; TL, total lipid fraction; AP, crobiuret analysis (7) with a bovine serum albumin Folch extracted PM residue; NL, neutral lipid frac- standard and carbohydrate content was measured by tion, GS, ganglioside fraction; GL-4, globoside; GL- the anthrone method (8) using a lactose standard. 3, trihexosyl ceramide; SDS, sodium dodecyl sulfate; Lipid content was measured gravimetrically. PBS, phosphate-buffered saline; Glc-Cer, glucosyl PM lipids were fractionated by column chroma- ceramide; TLC, thin-layer chromatography; C-M-W, tography on activated silicic acid (Unisil, Clarkson chloroform:methanol:water. Chemical Co.) by a modification (see Fig. 4 legend
685 0003-9861/82/080685-08$02.00/O Copyright Q 19&X! by Academic Press. Inc. All rights of reproduction in any form reserved.
686 SAGE, YATES, AND HORTON
0.25 -
I
.Ol .02 .03 .04 .05 06 .07 .0.9
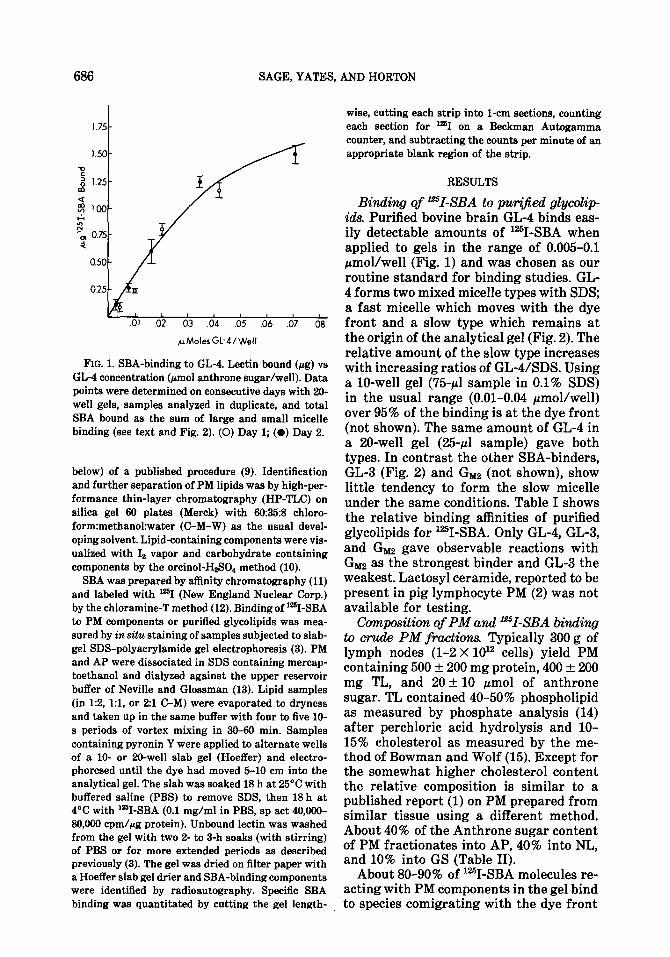
FIG. 1. SBA-binding to GL-4. Lectin bound (Fg) vs GL-4 concentration (am01 anthrone sugar/well). Data points were determined on consecutive days with 20- well gels, samples analyzed in duplicate, and total SBA bound as the sum of large and small micelle binding (see text and Fig. 2). (0) Day 1; (0) Day 2.
below) of a published procedure (9). Identification and further separation of PM lipids was by high-per- formance thin-layer chromatography (HP-TLC) on silica gel 60 plates (Merck) with 60~353 chloro- form:methanol:water (C-M-W) as the usual devel- oping solvent. Lipid-containing components were vis- ualized with Ia vapor and carbohydrate containing components by the orcinol-HaSO, method (10).
SBA was prepared by affinity chromatography (11) and labeled with ‘261 (New England Nuclear Corp.) by the chloramine-T method (12). Binding of ‘%I-SBA to PM components or purified glycolipids was mea- sured by in situ staining of samples subjected to slab- gel SDS-polyacrylamide gel electrophoresis (3). PM and AP were dissociated in SDS containing mercap- toethanol and dialyzed against the upper reservoir buffer of Neville and Glossman (13). Lipid samples (in 1:2, l:l, or 21 C-M) were evaporated to dryness and taken up in the same buffer with four to five lo- s periods of vortex mixing in 30-60 min. Samples containing pyronin Y were applied to alternate wells of a lo- or 20-well slab gel (Hoeffer) and electro- phoresed until the dye had moved 5-10 cm into the analytical gel. The slab was soaked 18 h at 25°C with buffered saline (PBS) to remove SDS, then 18 h at 4°C with ‘%I-SBA (0.1 mg/ml in PBS, sp act 40,000- 36,600 cpm/pg protein). Unbound lectin was washed from the gel with two 2- to 3-h soaks (with stirring) of PBS or for more extended periods as described previously (3). The gel was dried on filter paper with a Hoeffer slab gel drier and SBA-binding components were identified by radioautography. Specific SBA binding was quantitated by cutting the gel length-
wise, cutting each strip into l-cm sections, counting each section for =I on a Beckman Autogamma counter, and subtracting the counts per minute of an appropriate blank region of the strip.
RESULTS
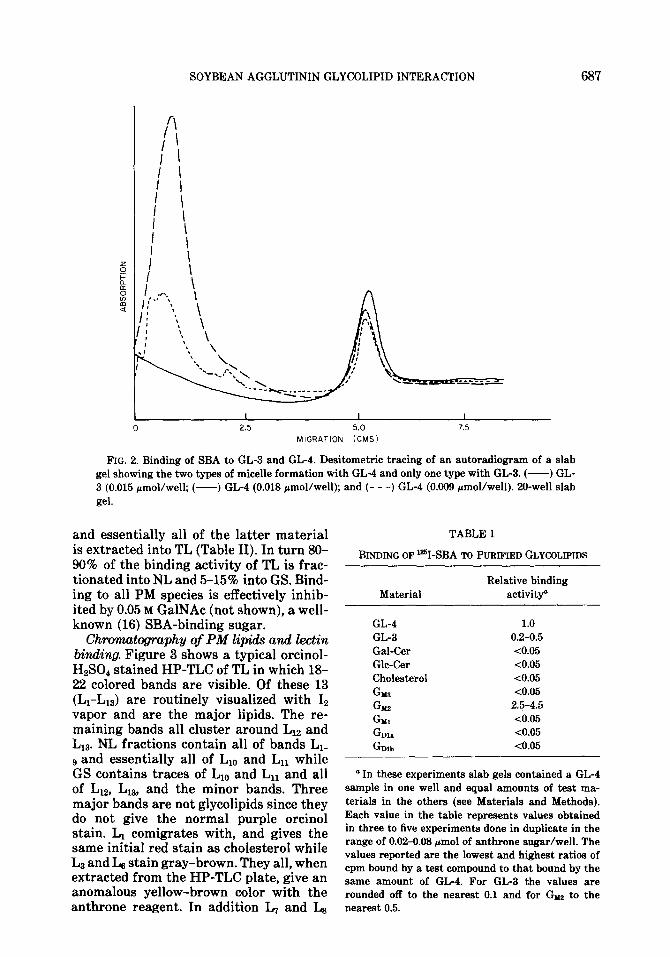
Binding of ‘=I-SBA to pw@ied glycolip ids. Purified bovine brain GL-4 binds eas- ily detectable amounts of ‘%I-SBA when applied to gels in the range of 0.005-0.1 pmol/well (Fig. 1) and was chosen as our routine standard for binding studies. GL- 4 forms two mixed micelle types with SDS; a fast micelle which moves with the dye front and a slow type which remains at the origin of the analytical gel (Fig. 2). The relative amount of the slow type increases with increasing ratios of GL-USDS. Using a lo-well gel (75~1 sample in 0.1% SDS) in the usual range (0.01-0.04 pmol/well) over 95% of the binding is at the dye front (not shown). The same amount of GL-4 in a 20-well gel (25~1 sample) gave both types. In contrast the other SBA-binders, GL-3 (Fig. 2) and Gnaz (not shown), show little tendency to form the slow micelle under the same conditions. Table I shows the relative binding affinities of purified glycolipids for ?-SBA. Only GL-4, GL-3, and Gm gave observable reactions with Glllz as the strongest binder and GL-3 the weakest. Lactosyl ceramide, reported to be present in pig lymphocyte PM (2) was not available for testing.
Composition of PM and ‘%I-SBA binding to crude PM fractions. Typically 300 g of lymph nodes (l-2 X 10” cells) yield PM containing 500 f 200 mg protein, 400 f 200 mg TL, and 20 -t 10 pmol of anthrone sugar. TL contained 40-50% phospholipid as measured by phosphate analysis (14) after perchloric acid hydrolysis and lo- 15% cholesterol as measured by the me- thod of Bowman and Wolf (15). Except for the somewhat higher cholesterol content the relative composition is similar to a published report (1) on PM prepared from similar tissue using a different method. About 40% of the Anthrone sugar content of PM fractionates into AP, 40% into NL, and 10% into GS (Table II).
About 8090% of ‘%I-SBA molecules re- acting with PM components in the gel bind to species comigrating with the dye front
SOYBEAN AGGLUTININ GLYCOLIPID INTERACTION 687
0 2.5 5.0 7.5
MIGRATION (CMS)
FIG. 2. Binding of SBA to GL-3 and GL-4. Desitometric tracing of an autoradiogram of a slab gel showing the two types of micelle formation with GL-4 and only one type with GL-3. (-) GL- 3 (0.015 rmol/well; (-) GL-4 (0.018 pmol/well); and (- - -) GL-4 (0.003 pmol/well). 20-well slab gel.
and essentially all of the latter material is extracted into TL (Table II). In turn 80- 90% of the binding activity of TL is frac- tionated into NL and 5-15% into GS. Bind- ing to all PM species is effectively inhib- ited by 0.05 M GalNAc (not shown), a well- known (16) SBA-binding sugar.
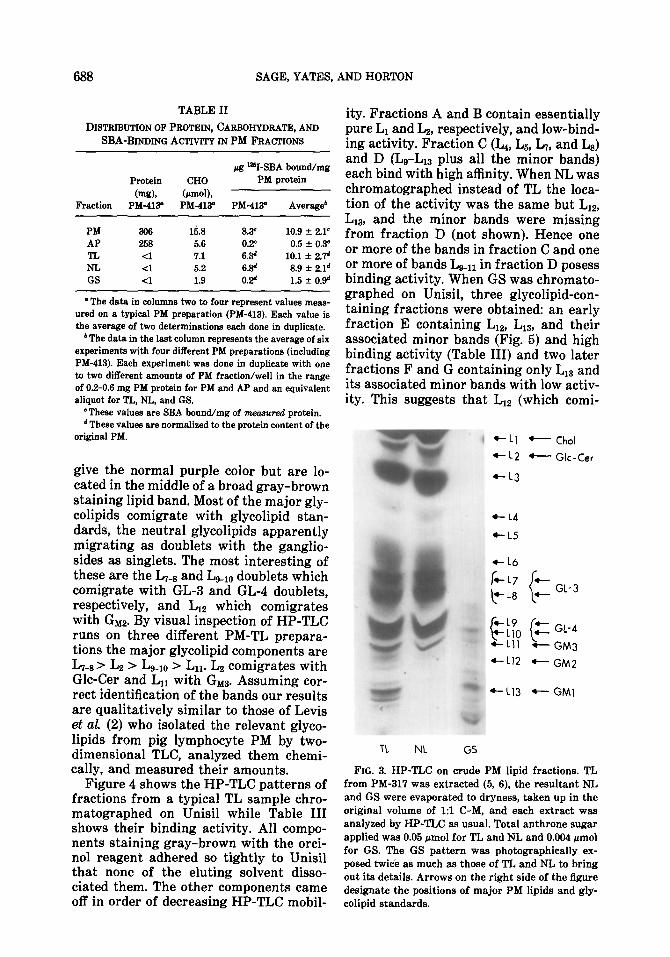
Chromatography of PM lipids and l&in binding. Figure 3 shows a typical orcinol- HzS04 stained HP-TLC of TL in which 18- 22 colored bands are visible. Of these 13 (L1-L13) are routinely visualized with I2 vapor and are the major lipids. The re- maining bands all cluster around LIZ and L13. NL fractions contain all of bands L1- 9 and essentially all of Li,, and Ln while GS contains traces of L1,, and Ln and all of Liz, L13, and the minor bands. Three major bands are not glycolipids since they do not give the normal purple orcinol stain. L1 comigrates with, and gives the same initial red stain as cholesterol while L3 and Ls stain gray-brown. They all, when extracted from the HP-TLC plate, give an anomalous yellow-brown color with the anthrone reagent. In addition L, and Ls
TABLE I
BINDING OF l%I-SBA TO PURIFIED GLYCOLIPIDS
Material Relative binding
activity
GL-4 GL-3 Gal-Cer Glc-Cer Cholesterol GM8 Gd2 G Ml G D1a Gmb
1.0 0.2-0.5 <0.05 <0.05 <0.05 <0.05
2.5-4.5 10.05 <0.05 <0.05
a In these experiments slab gels contained a GL-4 sample in one well and equal amounts of test ma- terials in the others (see Materials and Methods). Each value in the table represents values obtained in three to five experiments done in duplicate in the range of 0.02-0.08 pmol of anthrone sugar/well. The values reported are the lowest and highest ratios of cpm bound by a test compound to that bound by the same amount of GL-4. For GL-3 the values are rounded off to the nearest 0.1 and for Gm to the nearest 0.5.
SAGE, YATES, AND HORTON
TABLE II
DISTRIBUTION OF PROTEIN, CARBOHYDRATE, AND SBA-BINDING ACTIVITY IN PM FRACTIONS
ag “I-SBA bound/mg Protein CHO PM protein
(mf3). hoi), Fraction PM-4lb PM-413” PM-4W Average*
PM 306 15.8 8.3” 10.9 + 2.1” AP 258 5.6 02 0.5 * 03 TL <l 7.1 6.@ 10.1 2 2.74 NL <I 5.2 6.e 8.9 + 2.1d GS <l 1.9 0.P 1.5 f 0.9
o The data in columns two to four represent values meas- ured on a typical PM preparation (PM-413). Each value is the average of two determinations each done in duplicate.
b The data in the last column represents the average of six experiments with four different PM preparations (including PM-413). Each experiment was done in duplicate with one to two different amounts of PM fraction/well in the range of 0.2-0.6 mg PM protein for PM and AP and an equivalent aliquot for TL, NL, and GS.
c These values are SBA bound/mg of measured protein. 4 These values are normalized to the protein content of the
original PM.
give the normal purple color but are lo- cated in the middle of a broad gray-brown staining lipid band. Most of the major gly- colipids comigrate with glycolipid stan- dards, the neutral glycolipids apparently migrating as doublets with the ganglio- sides as singlets. The most interesting of these are the LTe8 and Lgdl,, doublets which comigrate with GL-3 and GL-4 doublets, respectively, and LIZ which comigrates with Gm. By visual inspection of HP-TLC runs on three different PM-TL prepara- tions the major glycolipid components are L,-s> L2 > L+lo > Ln. Lz comigrates with Glc-Cer and Ln with GMas. Assuming cor- rect identification of the bands our results are qualitatively similar to those of Levis et al. (2) who isolated the relevant glyco- lipids from pig lymphocyte PM by two- dimensional TLC, analyzed them chemi- cally, and measured their amounts.
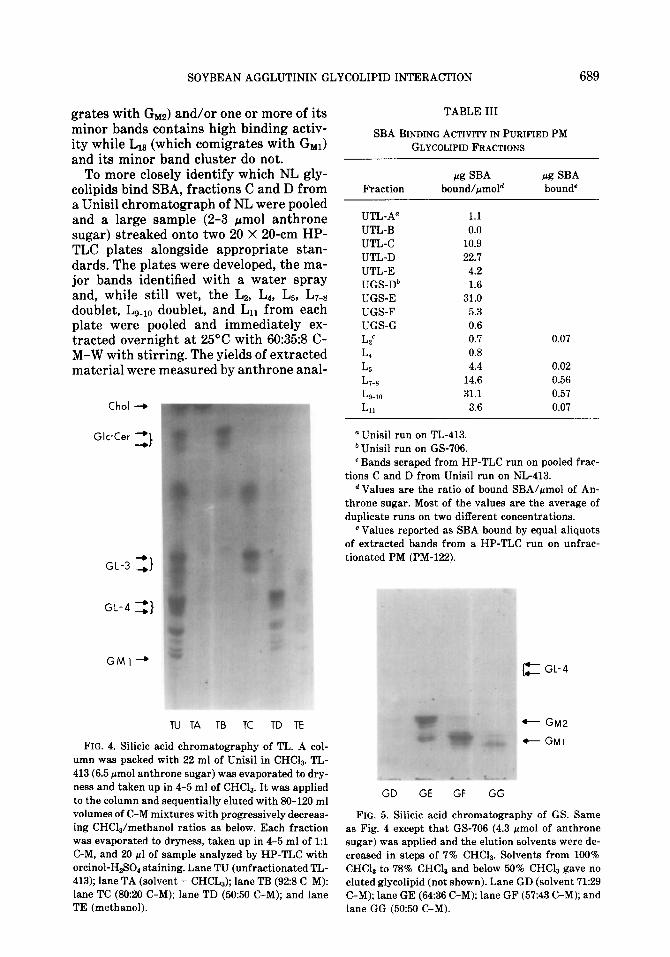
Figure 4 shows the HP-TLC patterns of fractions from a typical TL sample chro- matographed on Unisil while Table III shows their binding activity. All compo- nents staining gray-brown with the orci- no1 reagent adhered so tightly to Unisil that none of the eluting solvent disso- ciated them. The other components came off in order of decreasing HP-TLC mobil-
ity. Fractions A and B contain essentially pure L1 and Lz, respectively, and low-bind- ing activity. Fraction C (L4, Lg, L,, and L8) and D (Lg-Lls plus all the minor bands) each bind with high affinity. When NL was chromatographed instead of TL the loca- tion of the activity was the same but LIZ, Ln,, and the minor bands .were missing from fraction D (not shown). Hence one or more of the bands in fraction C and one or more of bands Lgen in fraction D posess binding activity. When GS was chromato- graphed on Unisil, three glycolipid-con- taining fractions were obtained: an early fraction E containing LIZ, L13, and their associated minor bands (Fig. 5) and high binding activity (Table III) and two later fractions F and G containing only L13 and its associated minor bands with low activ- ity. This suggests that LIZ (which comi-
t Chol t Glc-Csr
GL-3
GL-4
GM3
GM2
GMI
TL NL GS
FIG. 3. HP-TLC on crude PM lipid fractions. TL from PM-317 was extracted (5, 6), the resultant NL and GS were evaporated to dryness, taken up in the original volume of 19 C-M, and each extract was analyzed by HP-TLC as usual. Total anthrone sugar applied was 0.05 pmol for TL and NL and 0.004 amol for GS. The GS pattern was photographically ex- posed twice as much as those of TL and NL to bring out its details. Arrows on the right side of the figure designate the positions of major PM lipids and gly- colipid standards.
SOYBEAN AGGLUTININ GLYCOLIPID INTERACTION 689
grates with G& and/or one or more of its minor bands contains high binding activ- ity while L13 (which comigrates with Gm) and its minor band cluster do not.
To more closely identify which NL gly- colipids bind SBA, fractions C and D from a Unisil chromatograph of NL were pooled and a large sample (2-3 pmol anthrone sugar) streaked onto two 20 X 20-cm HP- TLC plates alongside appropriate stan- dards. The plates were developed, the ma- jor bands identified with a water spray and, while still wet, the LB, L4, LE., LT-8 doublet, LgmIO doublet, and Ln from each plate were pooled and immediately ex- tracted overnight at 25°C with 60:35:8 C- M-W with stirring. The yields of extracted material were measured by anthrone anal-
Chol +
Glc-Cer =}
GL-3 3
GL-4 3
GMI-,
TU TA TB TC TD TE
FIG. 4. Silicic acid chromatography of TL. A col- umn was packed with 22 ml of Unisil in CHCI,. TL- 413 (6.5 pmol anthrone sugar) was evaporated to dry- ness and taken up in 4-5 ml of CHCls. It was applied to the column and sequentially eluted with SO-120 ml volumes of C-M mixtures with progressively decreas- ing CHCla/methanol ratios as below. Each fraction was evaporated to dryness, taken up in 4-5 ml of 1:l C-M, and 20 pl of sample analyzed by HP-TLC with orcinol-HeSO staining. Lane TU (unfractionated TL- 413); lane TA (solvent = CHCL,); lane TB (92:s C-M): lane TC (80:20 C-M); lane TD (50:50 C-M); and lane TE (methanol).
TABLE III
SBA BINDING ACTIVITY IN PURIFIED PM GLYCOLIPID FRACTIONS
Fraction a SBA
bound/pmold a .%A bound”
UTL-A” 1.1 UTL-B 0.0 UTL-c 10.9 UTL-D 22.7 UTL-E 4.2 UGS-Db 1.6 UGS-E 31.0 UGS-F 5.3 UGS-G 0.6
LC 0.7 0.07
L* 0.8
Lb 4.4 0.02
L8 14.6 0.56
~9.IO 31.1 0.57
L11 3.6 0.07
a Unisil run on TL-413. b Unisil run on GS-706. c Bands scraped from HP-TLC run on pooled frac-
tions C and D from Unisil run on NL-413. d Values are the ratio of bound SBA/amol of An-
throne sugar. Most of the values are the average of duplicate runs on two different concentrations.
e Values reported as SBA bound by equal aliquots of extracted bands from a HP-TLC run on unfrac- tionated PM (PM-122).
t
t
GL-4
GM2 GMI
GD GE GF GG
FIG. 5. Silicic acid chromatography of GS. Same as Fig. 4 except that GS-706 (4.3 rmol of anthrone sugar) was applied and the elution solvents were de- creased in steps of 7% CHCla. Solvents from 100% CHCla to 78% CHCI, and below 50% CHCIa gave no eluted glycolipid (not shown). Lane GD (solvent 71:29 C-M); lane GE (64:36 C-M); lane GF (57:43 C-M); and lane GG (5050 C-M).

690 SAGE, YATES, AND HORTON
ysis on aliquots of each extract and resi- due, with values ranging from 30 to 60%. Equal amounts (0.02 and 0.06 pmol/well) were tested in duplicate for lar’I-SBA bind- ing. Table III shows that highest specific binding activity was associated with Lg_iO while L7-* showed about half the specific activity, with the other bands showing very little activity. The small amount of activity associated with Ln and L5 is prob- ably due to slight contamination with L1,, and L,, respectively. Because of the large amount of sample applied to the plates individual bands were broader and reso- lution was poorer than with a typical an- alytical HP-TLC run (not shown). Two similar experiments done with a different NL but with no quantitation of eluted gly- colipids gave a similar result; high activity associated with L7-8 and Lg-io and little or none with the others. Two similar studies were done with crude NL. However the presence of components giving the anom- alous anthrone color in several of the gly- colipid bands made quantitation imprac- tical. Instead equal aliquots from each band were simply compared directly for binding activity. The results of one exper- iment are shown in Table III; high activity was associated with L,-s and Lg-iO and little activity with the others.
DISCUSSION
In situ gel binding assay has been used by several workers (3, 17-19) to identify lectin-binding membrane glycopolypep- tides. This study extends the technique to membrane glycolipids. Qualitatively it has
5 Glycolipid st7ucture.s
Lactosyl ceramide
been used to identify SBA-binding PM gly- colipids and semiquantitatively compare their affinities for the lectin under a fixed set of conditions. The assay is less con- venient and considerably less sensitive than the microtiter plate assay (20, 21). However our ultimate goal in using this procedure is to compare the binding of SBA to PM glycopolypeptides and glyco- lipids under identical conditions and the microtiter assay doesn’t appear to be suit- able for this purpose. The gel assay could be readily modified to be more rapid and sensitive. Another major drawback to the gel assay is that it doesn’t separate the PM glycolipids. Silicic acid chromatogra- phy gives reasonable separation of several TL components. Silica gel chromatogra- phy also separates PM glycolipids (not shown) and we have obtained essentially pure L-r0 by a combination of the two pro- cedures using smaller eluting solvent steps (22). Magnani et al (23) have reported a direct in situ binding assay in which the glycolipids are separated on a film-backed TLC with a bonded resin prior to incu- bation with a radiolabeled ligand. This procedure has some obvious advantages but it cannot be used with glycopolypep- tides.
The highest affinities of SBA for Gm and GL-4 are only partly explained by its known higher affinity for GalNAc over Gal (16) and the assignment (24) of a free hy- droxyl hydrogen at the C-6 and C-4 posi- tion of the pyranose ring as being impor- tant contact points for binding. GIld2 and GL-4 both contain nonreducing terminal GalNAc.5 GM1 and GDlb each contain a ter-
Gal(j3 1-4)Glc(l-l*)-Cer
GL-3 Gal(a l-4)Gal@l-4)Glc(l-l*)-Cer
GL-4 GalNAc(p l-3)Gal(ct l-4)Gal(@ 1-4)Glc(l-I*)-Cer
GM8 Gal@ 1-4)Glc(l-l*)-Cer 3 (4 2
GM2
NANA
GalNAc(@ l-4)Gal(j3 l+Glc(l-I*)-Cer 3
(i) 2 NANA

SOYBEAN AGGLUTININ GLYCOLIPID INTERACTION 691
minal Gal-linked j31 - 3 to a penultimate GalNAc with free C-6 and C-4 hydroxyls, yet both are poor binders. This suggests that the GalNAc or Gal residue in an SBA- binding saccharide must be at the termi- nus either because the C-3 hydroxyl is a contact point or because of steric hin- drance. The greater reactivity of GL-3 than all the other glycolipids containing a terminal Gal (Gal-Cer, Gul, Gnu,, and probably Lac-Cer) is puzzling. GL-3 is the only one containing an a-linked Gal ter- minus but Hammarstrbm et al. (24) have reported no difference in the binding of (Y- and p-methyl glycosides of Gal or GalNAc to SBA. The negative reactions with the other glycolipids are probably not due to their being poorly taken up into the SDS-polyacrylamide gel electrophoresis solvent or diffusing out of the gel during the lectin-binding procedure. We (22) have subjected all the glycolipids to a mock binding experiment (no lectin present in the assay) and observed all of them pres- ent in comparable amounts at the dye front with Schiff periodate stain. We have also tested whether the samples were poorly soluble in the solvent by looking for antrhone color in the empty sample tubes. None was detected. Although we have not tested an authentic lactosyl-ceramide for binding, L4 and/or L5 are most likely PM lactosyl-ceramides and both have little or no SBA-binding activity. The assignment of L1z as being a PM Guz is not as well established as are the other SBA binders. We have looked (4) for a Gu.Jike PM sub- stance by HP-TLC on GS in two solvents which give better resolution of ganglio- sides; (a) 60:40:9 C-M-NHIOH and (b) 60:35:8 C-M-0.25% CaClz. The band cor- responding to LIZ comigrated with Gbl12 in both solvents and when the presumptive GY2 band with solvent (b) was scraped from the plate and subjected to mild Dowex-50 catalyzed acid hydrolysis the resultant material comigrated with au-
thentic asialo Ganz by HP-TLC in 60:35:8 C-M-W (4).
The reaction of SBA with GL-3, GL-4, and GY2, and their presence in PM doesn’t prove that they are surface receptors for SBA. In a future paper we will give evi- dence that, based on a comparison of the binding of SBA and chemically crosslinked SBA to lymphocyte surfaces and PM com- ponents, ‘70-80s of pig lymphocyte SBA receptors are glycolipids. The question of whether these receptors have second-mes- senger or ligand internalization roles is not answered. There is an increasing in- terest in surface gangliosides as receptors for interferon (25), thyrotrophin (26), cholera toxin (2’7), other hormones and toxins (for a review see (28)), and as sig- nals for differentiation (29). Furthermore a recent publication (30) suggests that they are involved in lymphocyte mitogen- esis. Little, if any, attention has been fo- cused on neutral glycolipids as surface re- ceptors although they are present in surface membranes in larger amounts than are the gangliosides. The rather small amounts of Gu12 in most tissues and the restricted specificity of SBA for GL-3 and GL-4 makes it potentially very useful for studies of the physiological activities of these major membrane glycolipids.
ACKNOWLEDGMENTS
We would like to thank Dr. Bernard J. Kaufman for his invaluable advice and generosity. Supported by VA General Medical Research.
REFERENCES
1. ALLAN, D., AND CRUMPTON, M. J. (1970) Bioch,em J. 120.133-143.
2. LEVIS, G. M., EVANGELATOS, G. P., AND CRUMP- TON, M. J. (1976) Bimhem. J. 156, 103-110.
3. ALEXANDER, 5. S., LIVINGSTONE, L. R., YATES, L. D., AND SAGE, H. J. (1978) Biochim Biophys Ada 512, 350-365.
4. YATES, L. D. (1979) Interactions of Mitogenic and
G bn Gal@ 1-S)GalNAc(@ l-4)Gal@ 1-4)Glc(l-l*)-Cer
NANA

692 SAGE, YATES, AND HORTON
Non-mitegenic Soybean Agglutinins with Por- cine Lymphocytes, Ph.D. Thesis, Duke Uni- versity.
5. FOLC!H, J., LEES, M., AND STANLEY, G. H. S. (1957) J. Biol Chem 226.497-509.
6. KANFER, J. N., AND SPIELVOGEL, C. (1973) J. Neu- rochem. 20,1483-1485.
7. ZAMENHOF, S. (1957) in Methods in Enzymology (Celowick, S. P., and Kaplan, N. O., eds.), Vol. 3, pp. 696-704, Academic Press, New York.
8. CARROLL, K. K. (1960) J. Lipid Res. 1, 171-178. 9. KAUFMAN, B. J., AND BASU, S. (1966) in Methods
in Enzymology (Neufeld, E. F., and Ginsburg, V., eds.), Vol. 8, pp. 365-366, Academic Press, New York.
10. WHERRETT, J. R., AND HAKOMORI, S. I. (1973) J. BioL Chem. 243,3046-3651.
11. VRETBLAD, P. (1976) Biochim Biophys Acta. 434, 169-176.
12. HUNTER, W. M. (1967) Handbook of Experimental Immunology (Weir, D. M., ed.), p. 608, Black- well, Oxford.
13. NEVILLE, D. M., AND GLOSSMAN, H. (1974) in Methods in Enzymology (Fleischer, S., and Lester, L., eds.), Vol. 32, pp. 92-102, Academic Press, New York.
14. KOPACZYK, K., PERDUE, J., AND GREEN, D. E. (1966) Arch Biochem Bioph?/s 115,215~225.
15. BOWMAN, R. E., AND WOLF, R. C. (1962) C&n. Chem 8,302-309.
16. LOTAN, R., SIEGELMAN, H. W., LIS, H., AND SHARON, N. (1974) J. BioL Chem 249, 1219- 1224.
17. TANNER, M. J. A., AND ANSTEE, D. J. (1976) Bie them. J. 153.265-270.
18. ROBINSON, P. J., BULL, S. G., ANDERTON, B. N., AND ROITT, I. M. (1975) FEBS Lett. 58, 330- 333.
19. BURRIDGE, K. (1976) Prec Nat. Ad Sci USA 73, 4457-4461.
20. YOUNG, W. W., MACDONALD, E. M. S., NOWINSKI, R. C., AND HAKOMURI, S. I. (1979) J. Exp. Med 150,1008-1019.
21. SMOLARSKY, M. (1980) J. Immunol Methods 38, 85-93.
22. SAGE, H. J. Unpublished observations. 23. MAGNANI, J. L., BROCKHAUS, M., SMITH, D. F.,
GINSBURG, V., BLASZCZYK, M., MITCHELL, K. F., STEPLEWSKI, Z., AND KOPROWSKI, H. (1981) Science 212, 55-56.
24. HAMMARSTR~M, S., MURPHY, L. A., GOLDSTEIN, I. J., AND ETZLER, M. E. (1977) Bioch&q 16,2756-2755.
25. BESANCON, F., ANKEL, H., AND BASU, S. (1976) Nature (London) 259,576-578.
26. MULLIN, B. R., FISHMAN, P. H., LEE, G., &OJ, S. M., LEDLEY, F. D., WINAND, R. J., KOHN, L. D., AND BRADY, R. 0. (1976) &WC. Nat. Acd Sci USA 73,842-846.
27. JOSEPH, K. C., STIEBER, A., AND GONATAS, N. K. (1979) J. CeU BioL 81,543-554.
28. KOHN, L. D., LEE, G., GROLLMAN, E. F., LEDLEY, F. D., MIJLLIN, B. R., FRIEDMAN, R. I$, MEL- DOLESI, M. F., AND ALOJ, S. M. (1978) Cell Sur- face Carbohydrate Chemistry (Harmon, R. E., ed.), pp. 103-133, Academic Press, New York.
29. ROISEN, F. J., BARTFIELD, H., NAGELE, R., AND YORKE, G. (1981) Science 214, 577-578.
30. SPIEGEL, S., AND WILCHEK., M. (1981) J. ZmmuneL 127, 572-575.







