diurnality and nocturnality in nonhuman primates - smartsite
TRANSCRIPT

This article was downloaded by: [University of California Davis]On: 10 November 2011, At: 18:06Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Biological Rhythm ResearchPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/nbrr20
Diurnality and nocturnality innonhuman primates: comparativechronobiological studies in laboratoryand natureHans G. Erkert aa University of Tuebingen, Institute for Zoology/Animal Physiology,Auf der Morgenstelle 28, D-72076, Tuebingen, Germany
Available online: 05 Mar 2008
To cite this article: Hans G. Erkert (2008): Diurnality and nocturnality in nonhuman primates:comparative chronobiological studies in laboratory and nature, Biological Rhythm Research, 39:3,229-267
To link to this article: http://dx.doi.org/10.1080/09291010701683391
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.

Diurnality and nocturnality in nonhuman primates: comparative
chronobiological studies in laboratory and nature
Hans G. Erkert*
University of Tuebingen, Institute for Zoology/Animal Physiology, Auf der Morgenstelle 28, D-72076,Tuebingen, Germany
Looking for differences in circadian clock characteristics of diurnal and nocturnalnonhuman primates, this article summarizes results of chronobiological studies carriedout in various nocturnal, diurnal, and cathemeral prosimian and anthropoid primatespecies under controlled laboratory conditions, under seminatural conditions, and in thewild. In almost all circadian parameters investigated, no differences were discerniblebetween the two main chrono-ecotypes, either in circadian period length and theinfluence upon it of after-effects, of light intensity, and ambient temperature, or in thePRC, re-entrainment behavior, rhythm splitting, and internal desynchronization.Diurnal and nocturnal or cathemeral species differed only in the phase of artificial ornatural LDs to which their circadian activity phase was adjusted as well as in thecharacteristics of masking activity upon the rhythms produced by the direct inhibitingor enhancing effects of light. Pronounced lunar periodicity—observed in the activityrhythm of nocturnal neotropical owl monkeys, genus Aotus, in seminatural and naturalenvironments as well as in wild cathemeral Malagasy lemurs, genus Eulemur—is shownto result from masking effects of moonlight. In captive Eulemur fulvus albifrons, achange from dark-active over cathemeral to light-active behavior, without concurrentlychanging the circadian phase-setting of activity to D, was produced by direct maskingeffects of a stepwise reduction of darktime luminosity on an LD 12:12 cycle. Long-termactivity recordings carried out in wild diurnal Malagasy sifakas (Propithecus verreauxi)and cathemeral redfronted lemurs (Eulemur fulvus rufus), as well as in wild nocturnalowl monkeys (Aotus a. azarai) of the North Argentinean Chaco, yielded in all speciesdistinct bimodal long- and short-day activity patterns with pronounced peaks duringdusk and dawn. Applying Pittendrigh’s two-oscillator concept to these results, it ishypothesized that the differences in chrono-ecotype behavior may result from variationsin internal coupling and external phase-setting of morning and evening oscillators (m, e)to dawn and dusk, interacting with direct masking effects of light.
Keywords: circadian rhythms; activity patterns; photic and nonphotic entrainment;phase-response; light masking; lunar periodicity; cathemerality; prosimian andanthropoid primates
Introduction
Extant mammals are generally assumed to have evolved from small arboreal insectivoreswith a nocturnal lifestyle. The diurnality found in a large number of mammalian taxa is
*Email: [email protected]
Biological Rhythm Research
Vol. 39, No. 3, June 2008, 229–267
ISSN 0929-1016 print/ISSN 1744-4179 online
� 2008 Taylor & Francis
DOI: 10.1080/09291010701683391
http://www.informaworld.com
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

thought to have developed polyphyletically from nocturnal ancestors (Crompton et al.1978; Starck 1978). This holds also for nonhuman primates (Charles-Dominique 1975;Fleagle 1988; Martin 1990). Why and how the transition from the nocturnal to a diurnallifestyle might have proceeded in the various taxa is still a matter of speculation (Martin1990; van Schaik & Kappeler 1996; Smale et al. 2003). While evolutionary biology andbehavioral ecology were mainly concerned with the ultimate factors that might havecaused certain members of an order, family, or genus to remain nocturnal or to adopt amore diurnal lifestyle, chronobiologists have almost exclusively focused on thephenomenology and functional analysis of the circadian timing system (CTS) involvedin the temporal regulation of the species’ daily activity and resting phases. Butchronobiologists have often ignored the function of this timing system for the animals’life and survival in nature (DeCoursey 2004; Marques & Waterhouse 2004; Morgan 2004).
Animal models used for the behavioral, physiological, molecular and molecular-genetical approaches followed thus far in the functional analysis of mammalian circadianclockwork were only a few nocturnal rodent species such as laboratory rats, mice, andSyrian hamsters. Corresponding comprehensive studies in other nocturnal or even diurnalmammalian species have been performed to a much lesser extent (Moore-Ede & Sulzman1977, 1982; Sulzman et al. 1979; Edgar et al. 1993; Hut et al. 1999a, 1999b; Caldelas et al.2003). Comparative chronobiological studies in nocturnal and diurnal non-rodentmammalian species, carried out both under strictly controlled laboratory conditions andin nature, are rare (DeCoursey & DeCoursey 1964; Erkert 1974, 1976a, 1976b; Erkert &Groeber 1986; Kappeler & Erkert 2003; Fernandez-Duque & Erkert 2006). For thisreason, not many details of the tissue structure, cellular, molecular, and moleculargenetical mechanisms of the principal central nervous circadian pacemaker located in thehypothalamic suprachiasmatic nuclei (SCN) are known, except in these few nocturnalrodent models, and this ignorance extends to an understanding of their afferent andefferent pathways and the mechanisms of photic and, partly, nonphotic entrainment(Klein et al. 1991; Weaver 1998; King & Takahashi 2000; Shearman et al. 2000; Zordanet al. 2000; Moore & Leak 2001; Schwartz et al. 2001; Lee et al. 2003; Meijer & Schwartz2003). Robust ideas on the chronophysiological and chronoecological basis of mammaliandiurnality and its multiple evolution are still lacking. Another gap in our knowledgeconcerns the diversity of circadian characteristics and their functional and/or adaptivemeaning in mammals of different systematic groups, ecotypes and chronotypes. To get abetter idea of the diversity developed in the phylogenetically ancient CTS during theirevolutionary transition from an originally nocturnal to a more diurnal lifestyle and/orwhile adapting to changed abiotic and biotic time structures of newly conquered ecologicalniches, extensive comparative chronobiological studies are required both under strictlycontrolled laboratory conditions and in the natural environment.
In a first attempt to characterize the originally assumed fundamental differences in theresponse of the CTS in nocturnal (dark-active) and diurnal (light-active) animals tochanges in light intensiy, Aschoff (1964, 1979) formulated his so-called ‘‘circadian rule’’. Itoriginally stated that, in nocturnal animals, the circadian spontaneous period t ispositively correlated with light intensity, while total activity and the ratio of activitytime:resting time (a:r) are negatively correlated with it. Diurnal animals should behave inexactly the opposite manner. As to the circadian effects of ambient temperature, nocturnalspecies were thought to behave in a ‘‘cold-active’’, and diurnal ones in a ‘‘warm active’’,manner. However, while the prediction concerning the light effect on t, nowadays oftencited as ‘‘Aschoff’s rule’’ (Munoz et al. 2005; Daan 2000), could be substantiated in manynocturnal mammals, most diurnal species did not follow it (Aschoff 1979). Also,
230 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

generalizations concerning the temperature effect on circadian rhythmicity could not bevalidated in the endothermic mammals.
Since those early years of chronobiological research, no fundamental differencesbetween the characteristics of the CTS in nocturnal and diurnal mammals could beestablished, either in the shape and amplitude of the photic phase response curve essentialfor the circadian systems’ phase-setting (entrainment) to the environmental 24-h day(Pittendrigh & Daan 1976; Daan 2000; Kas & Edgar 2000; Refinetti 2004), themorphological, immuno-histological, electrophysiological, and metabolic characteristicsof the SCN pacemaker, or its afferent and efferent pathways (for review see Smale et al.2003). For this reason, it has been assumed that the mechanism or ‘‘swich’’ responsible forthe phase-setting of the diurnal mammals’ active phase to the bright part of the day mustbe located downstream from the SCN and/or be a result of direct masking effects due tolight, which were also thought to be mediated independent of the SCN (Mrosovsky 2003;Smale et al. 2003; Redlin & Mrosovsky 2004; Erkert et al. 2006). However, some recentfindings of nocturnal wheel-running in some diurnal African grass rats, Arvicanthusniloticus, have been interpreted as indicating fundamental differences in the molecularmechanism of certain SCN cells in diurnal and nocturnal mammals (Lee 2004; Schwartzet al. 2004). But since this evidence was obtained from a normally diurnal rodent species, afew animals of which behave dark-active only when having access to a running wheel,these results need further substantiation by also analyzing strictly diurnal and nocturnalspecies, including non-rodents.
As in rodents, several other mammalian orders are found that are not clearlynocturnal, diurnal or crepuscular. In primates, for instance, with the exception of thenocturnal South American owl monkey genus Aotus, all haplorrhine or anthropoid speciesare strictly diurnal, while most of the strepsirrhine or prosimian species remainednocturnal. Only a few Malagasy lemurs such as the indris (Indri indri), sifakas(Propithecus), ringtailed lemurs (Lemur catta), and ruffed lemurs (Varecia) have adopteda diurnal lifestyle while some Eulemur and Hapalemur species may be active both duringthe day and night. This mode of behavioral activity or chronotype, which is also knownfrom some other mammalian orders such as Carnivores, Artiodactyla, Perissodactyla, andXenarthra, has recently been designated as ‘‘cathemerality’’ (Tattersall 1987, 2006; Curtis& Rasmussen 2006). Which factors may account for this variability in primate chrono-ecotypes is still in controversial debate among primatologists, though the debate haslargely taken place without considering relevant chronobiological aspects (van Schaik &Kappeler 1996; Curtis et al. 1999; Donati et al. 2001; Curtis & Rasmussen 2002; Kirk2006). Due to this chronotype diversity, nonhuman primates may be of special interest forthe analysis of the chronobiological traits that may have caused or allowed the variousprosimian and simian primate species to become diurnal or cathemeral, or to remainnocturnal. For these reasons, we have carried out extensive comparative chronobiological/chronoecological studies in some nocturnal, diurnal, and cathemeral prosimians as well asin diurnal and nocturnal anthropoid primate species, both under strictly controlledconditions in the laboratory, under semi-natural conditions in outdoor enclosures withinthe species’ habitat, and in the wild. Here we will briefly summarize some of the resultsobtained in laboratory settings and present findings from activity recordings obtainedunder semi-natural conditions and in the wild. To give an adequate overview of currentknowledge on the characteristics of circadian rhythmicity in nonhuman primates, ratherunconventionally, the results of other authors are integrated into the ‘‘results’’ chapter. Abrief description of the methods used seems necessary because some unpublished resultsare presented.
Biological Rhythm Research 231
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Methods
Laboratory studies
In neotropical haplorrhine (anthropoid) primates, chronobiological experiments undercontrolled laboratoryconditionswere conducted in12adult, nocturnalNorthColombianowlmonkeys,Aotus lemurinus griseimembra (sevenmales, five females) as well as in 23 males and26 females of the strictly diurnal common marmoset, Callithrix j. jacchus. Preliminaryexperiments were also carried out in two adult females of the Bolivian owl monkey,A. azaraiboliviensis. Strepsirrhine (prosimian) primates studied under laboratory conditions were thecathemeral, white-fronted lemur, Eulemur fulvus albifrons (three males, two females), thenocturnalmouse lemur,Microcebusmurinus (sixmales), and thenocturnalAfricanbushbabyspecies, Galago senegalensis (five males) andOtolemur garnettii (eight males, one female).
Activity recordings and, in some cases, telemetrical core temperature measurementswere carried out at the Institute for Zoology of the University of Tuebingen, Germany, insound-protected and light-proof program-controlled air-conditioned rooms. All experi-ments were conducted according to German law for animal welfare, with permission andunder control of the relevant veterinary authorities.
Locomotor activity was usually recorded by means of transducer-like microphonesfixed on the wire mesh of the recording cages. If activated by the vibrations which theanimals induced in the wire mesh when moving around, these microphones triggeredelectrical pulses via an amplifier. These pulses were counted over 5 min intervals and thesums printed out and stored on disk by a multiplex counter/PC/printer device. In someAotus and Eulemur animals, body temperature was simultaneously measured by means ofintraperitoneally implanted miniature transmitters (MiniMitter, TM disc), whose signalwas picked up and transformed by a LRR-27 multichannel receiver connected to a PC-bound DATAQUEST III recording system (MiniMitter). For further details see Rappoldand Erkert (1994) and Rauth-Widmann et al. (1996).
Studies under seminatural conditions
Aotus lemurinus griseimembra
The first activity recordings under semi-natural conditions in a primate’s distribution areawere conducted in 1971–1972 in four owl monkey pairs (A. lemurinus griseimembra) kept in afield-laboratory shed in the Central Colombian East Andes, in a clearing of the ‘‘cafetal’’ ofFinca Rhenania, El Colegio/Cundinamarca (48350N, 748270W), about 1000 m above sealevel. Transparent perspex roofs above a large ventilation space on top of the animals’rooms and a broad ventilation slot at the bottom of their outer wall, both covered with wiremesh and flyscreen, ensured that the monkeys were optimally exposed to natural day- andmoonlight as well as to local climatic factors. Rooms were equipped with a sleeping box andseveral climbing bars that were suspended from steel spring/microswitch devices connectedto event recorders and electromechanic multiplex counters that printed out the numbers ofclosed contacts in 30 min intervals. For further details, see Erkert (1974, 1976).
Ateles geoffroyi
Long-term activity records of spider monkeys (Ateles geoffroy) were collected incooperation with Drs. J. Munoz-Delgado (Instituto Mexicano de Psiquiatrıa, MexicoD.F.), M. Corsi-Cabrera (Facultad de Psicologıa de la Universidad Autononoma de
232 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Mexico) and D. Canales-Espinoza (Instituto de Neuroetologıa de la UniversidadVeracruzana) at this institute’s ‘‘Parque de la Flora y Fauna Silvestre Tropical Pipiapan’’(Munoz et al. 2004). This etho-ecological field station is located in Central Mexico, about12 km east of Catemaco at 188270N, 958020W, 330 m above sea level. It is surrounded bysecondary rainforest, has a tropical climate with a rainy season during the northern long-daysummer months, and a dry season, roughly corresponding to the short-day winter months.
Throughout the first recording period, covering a whole annual cycle, the monkeyswere kept in a large wire mesh outdoor enclosure (126663 m [length6width6height]).For actimetry, seven subjects of a large social group were provided with Actiwatch1 AW4accelerometer collars (Cambridge Neurotechnology, UK; http://www.camntech.co.uk).Those collars were fitted on the monkey’s neck while being anesthetized for about 30 minwith 2.5 mg/kg ketamine-hydrochloride administered via blow-pipe darts. In a laterrecording period, some spider monkeys wearing AW4 collars were transferred to a smallforest island of about a quarter hectare surrounded by a high electric fence.
Recordings in the wild
Malagasy lemurs
The first long-term activity registrations in wild nonhuman primates were conducted incooperation with Prof. P. Kappeler (Department of Sociobiology of the German PrimateCenter (DPZ), Goettingen), at this institute’s field station in the Kirindy Forest, WesternMadagascar. From September 1998 to March 2003, records lasting several months wereobtained from a total of four adult cathemeral red-fronted lemurs (Eulemur fulvus rufus;Lemuridae) and eight adult subjects of the considerably larger and strictly diurnalVerreaux’s sifaka (Propithecus v. verreauxi, Indridae). The study site was a deciduous dryforest situated 60 km north of Morondava at 448390E, 208030S, about 60 m above sea level.The local climate is characterized by a hot wet season during the summer in the southernhemisphere from November to March and a cooler dry season from April to October.
Activity data were also collected with AW4 accelerometer collars fixed on the lemur’sneck after having been captured via blow pipe darting by an experienced Malagasytechnician. Formore details, seeKappeler andErkert (2003) andErkert andKappeler (2004).
North Argentinean owl monkeys
The North Argentinean Chaco is the southernmost distribution area of the neotropicalowl monkey, genus Aotus. In the provinces of Formosa and Chaco, Aotus a. azarai mainlyinhabits the gallery forests of the Rio Paraguay and its tributaries. This Aotus subspecieshad been described as showing considerable amounts of activity during the bright portionof the day (Arditi 1992; Rotundo et al. 2000; Fernandez-Duque 2003). For this reason, ithas been considered to behave cathemerally, like some lemurs (Tattersall 1987, 2006;Curtis & Rasmussen 2006; Fernandez-Duque 2003). Since the other Aotus species seem,instead, to behave strictly nocturnally (Moynihan 1964; Erkert 1974; Wright 1978; Garcia& Braza 1987), those North Argentinean owl monkeys were of special interest with respectto the diurnality – nocturnality problem.
Together with Dr. E. Fernandez-Duque (Fundacion ECO, Formosa, Argentina andDepartment of Anthropology, University of Pennsilvania, USA), we also conductedquantitative long-term activity recordings in wild Aotus a. azarai owl monkeys of theArgentinean Chaco. The study site was a small gallery forest of the ‘‘Riacho Pilaga’’
Biological Rhythm Research 233
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

situated about 20 km north of Formosa, Argentina at the Estancia Guaycolec (588110W,258580S), about 60 m above sea level. The climate characteristics of this subtropical opensavanna region, with a mosaic of grasslands and dry and gallery forests, are relatively dryand cold winter months from May–September, with average night-time temperatureminima falling well below 108C and daytime maxima of 21–258C, while the summermonths from October–February are relatively warm and rainy, with average minimum –maximum temperatures ranging from about 15–30 to 20–378C. Activity recordings forseveral months with Actiwatch AW4 accelerometer collars were performed upon a total ofseven male and three female Aotus a. azarai monkeys. For details on the species’ behaviorand ecology, study site, climate conditions, etc., see Fernandez-Duque (2003) andFernandez-Duque and Erkert (2006).
Data processing and evaluation
After each of the activity measurements which lasted up to 200 days, the animals wearingactiwatch collars were recaptured (by the use of a dart) and their collars were replaced orremoved. Activity data were downloaded from the actiwatches to a PC by means of areader interface and the commercial Actiwatch-Sleep1 program of Cambridge Neuro-technology. Thereafter, the data files were transformed into another format and analyzedby an extended periodogram analysis according to Doerrscheidt and Beck (1978). Toobtain comparable data, the individuals’ activity records in five min bins were averagedover five month periods, covering the long-day and short-day halves of the year. Resultingactivity patterns were then used to calculate average activity and resting times by best-fitting a square wave pattern to them (cf. Figure 8). Then the various parameters of thetwo patterns could be determined (cf. Figures 9A, B) and compared within and/or betweenspecies. For statistical comparisions, ANOVA followed by Scheffe tests, or otherparametric or nonparametric tests or correlation analyses, were applied as appropriate. Pvalues of 50.05 were considered to indicate significant differences or correlations. If notindicated otherwise, arithmetic means + SD are given.
Results
Laboratory studies
Circadian period and after-effects
Under constant conditions, all primate species studied showed free-running circadianactivity rhythms (CAR). Average periods (t) shorter and longer than 24 h occurred bothin nocturnal and diurnal, as well as in strepsirrhine and haplorrhine, species (Table 1,Hoban et al. 1985). Thus no clear relationship seems to occur between primate chrono-ecotypes or systematic affiliation and circadian period length. Whether or not in nocturnalprosimians there is a tendency to periods shorter than 24 h, as one might be tempted toinfer from Table 1, requires further studies in other species.
Distinct after-effects on t of preceeding photic entrainment to 24 h were observed inCallithrix, Aotus and Otolemur but not in Eulemur. Callithrix shortened t from 23.6 h to23.2 h on average (Erkert 1989). In Aotus, in a first trial t lengthened from 24.4 + 0.1 to25.3 + 0.7 h (Thiemann-Jaeger 1986), while in a second experiment only a minorlengthening from 24.3 + 0.3 to 24.45 + 0.4 h resulted from exposure to LL 0.1 lux(Rappold & Erkert 1994). Otolemur shortened t in a first experiment over 60–80 days,by 1.2 h on average, while extended recordings carried out later revealed much stronger
234 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Table
1.
Circadianperiodt,
after-effects,andtheeff
ectoflightintensity
onperiodlength
indiurnalandnocturnalprimates.
Species
Act.
nPara-
meter
Luminance
range(lux)
Average
t(h)
Variation
oft(h)
After-
effects
Light-eff
ect
ontmeets
Aschoff’s
rule
Reference
Strepsirrhini(Prosimii)
Microcebusmurinus
no
30
Loc
0–RR
22.5+
0.6
20.7–23.3
(þ)
(þ)
Schillinget
al.1999
Eulemurfulvusalbifr.
cat
5Loc/Tb
0.1–240
25.3+
0.4
24.1–26.5
–n.t.
ErkertandCramer
2006
Galagosenegalensis
no
5Loc
0–0.1
23.6+
0.6
23.4–24.0
(þ)
n.t.
SchanzandErkert1986
Otolemurgarnettii
no
9Loc
0–0.1
22.6+
0.7
21.1–23.9
þþ
n.t.
Erkertet
al.2006
Haplorrhini(A
nthropoidea)
Aotuslemurinusgris.
no
6Loc
0.002–360
25.1+
0.9
24.2–26.2
þ7
/þThiemann-Jaeger
1986
10
Loc/Tb
0.2
24.4+
0.4
23.9–25.6
(þ)
n.t.
Rappold
andErkert1994
Callithrixj.jacchus
di
14
Loc
0.1–480
23.3+
0.3
22.7–24.0
þ(þ
)Erkert1989
6Loc
0.1–430
23.2+
0.4
22.5–24.2
þþ
Wechselberger
1995
8Loc
50.5
23.3+
0.4
a–
n.i.
n.t.
Glass
etal.2001
Saim
irisciureus
di
16
Feed
1–600
25.0+
0.5
a24.3–26.3
n.i.
–Sulzmanet
al.1979
Tb
1–600
24.9+
0.6
a23.3–25.8
n.i.
–Kþ-exc.
1–600
25.1+
0.3
a20.5–29.5
n.i.
–5
Dri
0–600
24.3+
0.1
25.7+
0.3
n.i.
–HobanandSulzman1985
8Loc
0.1–400
25.2+
0.4
24.5–26.2
n.i.
–AschoffandTokura
1986
4Feed/Tb
1–300
24.8+
0.3/.1a,b
25.2/.3+
0.2/.1a,b
n.i.
–Fuller
andEdgar1986
9Dri
0–600
24.5+
0.1
25.6+
0.1
n.i.
–Ferraro
andSulzman1988
Atelesgeoffroyi
di
4Loc
107
23.9+
0.1
23.7–24.0
n.i.
n.t.
Munoz-D
etal.2005
Macaca
mulatta
di
4Loc
270
24.0+
0.3
23.8–24.4
n.i.
n.t.
YellinandHauty
1971
4Loc
0–300
24.1+
0.3
23.8–24.9
n.i.
–Martinez
1972
Macaca
nem
estrina
di
3Loc
0.003–100
23.1+
0.4
22.3–23.8
n.i.
–Tokura
andAschoff1978
3Loc
0.45–450
22.6–25.0
n.i.
–Tokura
andAschoff1983
Macaca
irus
di
2Loc
0–50
24.2
23.7–24.6
n.i.
–HawkingandLobban1970
Pantroglodytes
di
1Feed
1–85
24.4
23.7–25.1
n.i.
–Farrer
andTernes
1969
Act:activitytype/chrono-ecotype:
di:diurnal;no:nocturnal;cat:cathem
eral.
Parameter:Loc:
locomotoractivity;Tb:bodytemperature;Feed:Feeding,Dri:drinking;Kþ-exc:
renalpotassium
excretion.
After-effects:þþ,þ,(þ
):strong,distinct,evidence
ofafter-effects;7:noafter-effects;n.i.:noinform
ation.
Lighteff
ectont:þ,(þ
),þ/7
:is,seem
s,isonly
partly
consistent;7:inconsistentwithAschoff’srule
statingfornocturnalspeciesalengtheningandfordiurnalspeciesa
shorteningoftwithincreasinglightintensity;n.t.:nottested.
RR:constantdim
redlight;+S.D
.orS.E.a;b)Thefirstaveragecorrespondsto
tin
LL1lux,thesecondoneto
tin
LL300lux.
Biological Rhythm Research 235
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

after-effects of Dt ¼ 2.2 h, lasting up to 180 days (Erkert et al. 2006). In Eulemur, nosystematic period changes depending on the duration of constant conditions could bediscerned. Though not checked specifically, the short activity records obtained underconstant conditions (DD, LL 1071 lux) in Microcebus and Galago (Figure 1) seemed toindicate the presence of distinct after-effects in these two prosimians as well. InMicrocebus, after-effects might have been responsible for the differences between therelatively long ts obtained in our short LL experiments and the much shorter periodsfound by Schilling et al. (1999) in longer lasting DD and dim red light (RR) experiments.
Parametric light effects on t (Aschoff’s rules)
Clear luminosity-correlated variations in period length, as predicted by Aschoff’s rule,were not found either in the diurnal marmosets and squirrel monkeys or the nocturnal owlmonkeys (Table 1). In a first study in Callithrix, the effect of LL-luminosity on t within therange of 0.08–480 lux was a negative correlation of rs ¼ 70.29, one that just missed
Figure 1. Photic entrainment and masking of free-running circadian activity rhythms inrepresentatives of three nocturnal prosimian species by 12:12 h LD cycles with different lightintensities during the light and dark portions of the LD, as indicated on the right margin. Shadedareas mark the light portions of the applied LDs. While all animals’ endogenous rhythm wassynchronized both by the LD 102:0 (51077) lux and 102:1071 lux cycles, the LD 1071:0 lux cyclefailed to entrain the two galagos’ free-running activity rhythm, although it produced pronouncedmasking effects. Note that the activity level of the LD-entrained rhythm in all species was higher inLD 102:1071 lux (bottom) than in the LD 102:0 lux (top) cycles.
236 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

statistical significance (Erkert 1989). However, after reducing the LL-luminosity from 430to 0.3 lux, Wechselberger (1995) found t to lengthen significantly by 22 + 13 min, and toshorten by 18 + 14 min after increasing LL luminance several months later from 0.3 to430 lux. Corresponding to Aschoff’s rule, owl monkeys shortened their spontaneousperiod from 26.2 + 0.2 h (in LL 240 lux) to 24.3 + 0.1 h (in LL 0.3 lux) but, in LL 0.002lux, t lengthened again to 24.6 + 0.1 h (Thiemann-Jaeger 1986). Diurnal squirrelmonkeys (Saimiri sciureus) lengthened t of their drinking and activity rhythm significantlyfrom about 24.3–24.5 h in DD (0 lux) to 25.6–26 h in LL 600 lux (Hoban & Sulzman 1985;Aschoff & Tokura 1986; Ferraro & Sulzman 1988). Correspondingly, after reducing LLluminosity from 330 to 1 lux, the period of this species’ body temperature and feedingrhythm shortened significantly from 25.2 to 24.8 h (Fuller & Edgar 1986). Slightlenghthening of t at higher LL intensities (600 vs. 60 and 1 lux) was also found in the free-running circadian rhythms of the squirrel monkeys’ feeding, rectal temperature, and renalpotassium excretion rhythms (Sulzman et al. 1979). Longer periods at higher LLluminosities also occurred in chimpanzees (Pan troglodytes; Farrer & Ternes 1969) andsome macaques (Macaca nemestrina, M. irus; Tokura & Aschoff 1978, 1983; Hawking &Lobban 1970), while, in Macaca mulatta, t did not vary uniformely with LL luminosity(Martinez 1972). Thus, as it had already been realized by Aschoff (1979) himself, (most)diurnal primates do not follow his rules (Table 1). So in primates at least their centralprediction concerning the parametric light effect on period length t does apply at most forthe nocturnal species. Since in most other diurnal mammals studied thus far the observedeffects of light intensity on t are also inconsistent with that prediction of Aschoff’s rule,parametric light effects on t cannot be causally connected with mammalian diurnality ornocturnality, as has originally been assumed by Aschoff.
Characteristics of photic phase response
The phase response curves (PRCs) established in the nocturnal Aotus lemurinus (Rauth-Widmann et al. 1991; Rappold & Erkert 1994) and the diurnal Callithrix jacchus(Wechselberger & Erkert 1994) resembled one another as well as those of nocturnal mouselemurs (Schilling et al. 1999) and diurnal squirrel monkeys (Hoban & Sulzman 1985). Justas in nocturnal and diurnal rodents, in primates also, delay shifts were elicited by lightin the late subjective day/early subjective night while advance shifts resulted fromlight pulses given in the late subjective night/early subjective day. In the marmosets’ andmouse lemurs’ PRCs, the delay section was more pronounced (peaks of about 790and 7150 min, respectively) than the advance section (peak around þ30 min), whereasthe PRCs of squirrel and owl monkeys with longer t’s had a more pronounced advancethan delay section. Thus, in primates as in other animals, no fundamental differencesbetween nocturnal and diurnal species seem to occur in their photic phase responsecharacteristics.
Photic entrainment and masking
Entrainment and re-entrainment. As was expected, in all primate species studied, light –dark cycles were the most potent environmental time cue or Zeitgeber that stablysynchronized the animals’ circadian system with the 24-h time structure in theirenvironment. In diurnal species, the Zeitgeber phase-set the activity time to coincidewith the light, and in nocturnal species with the dark, portion of the light – dark cycle(Figure 1; Hoban & Sulzman 1985; Erkert & Thiemann 1983). With respect to someZeitgeber parameters, however, marked species-specific differences seemed to exist, and
Biological Rhythm Research 237
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

these indicate differences in the circadian systems’ susceptibility to this Zeitgeber. While12:12 h LD cycles of 0.1 lux (full moon luminosity) to 1074 lux or physiological darkness(about 1076/1077 lux) set the activity phase of the free-running circadian activity – restrhythm (CAR) of owl monkeys and mouse lemurs to the dark time (Figure 1 left, Erkert &Thiemann 1982), such LDs failed to entrain the endogenous rhythm in the two (alsostrictly nocturnal) bush baby species, Galago senegalensis and Otolemur garnettii (Figure 1;Erkert et al. 2006). Thus even in nocturnal primates/prosimians there are distinct speciesdifferences in the circadian systems’ thresholds for photic entrainment. In Otolemur, thistheshold was established in the range of about 3–30 lux, and it has been hypothesized thatsuch high thesholds for photic entrainment in nocturnal mammals may be an adaptivetrait preventing the circadian mechanism of photic phase-setting of circadian rhythmicityfrom being affected by moonlight (Erkert et al. 2006). Unfortunately, in no other primateshave the thresholds for photic entrainment yet been determined, but this is required inorder to find out whether this circadian parameter might be related to diurnal andnocturnal behavior.
Besides the variability of t (including its dependence on light intensity) and the PRC,the range of photic entrainment and the time needed for re-entrainment (tr) after phaseshifts of an entraining Zeitgeber may also be used to estimate the degree of plasticity of aspecies’ circadian system. Ranges of photic entrainment have been determined in theneotropical anthropoids, Callithrix jacchus and Aotus lemurinus, as well as in theprosimian species, Galago senegalensis, Otolemur garnettii, and Eulemur fulvus. In femalemarmosets, Haerter and Erkert (1993) found the range of entrainment for LDs of 345:0.03lux with corresponding L and D times between 22/23 and 26/27 h. In Aotus lemurinus, forLDs of 240:0.3 lux, the lower limit of photic entrainment was established at about 20–22 hand the upper one around 30 h (Thiemann-Jaeger 1986). For LD cycles with only 3 h L of100 lux and 20–23 h D of 0.2 lux, Rappold and Erkert (1994) determined the range ofentrainment between 23/23.5 and 25.5/26 h (LD 3:20 to LD 3:23). Relatively shortsynchronization experiments with LDs of about 100:0.1 lux yielded in Galago senegalensisa range of entrainment between 22/23 and 26/28 h and, in Otolemur garnettii, of 20/22 to25/28 h (Erkert 1984). It is still unknown, however, whether or not, at the limits of therange of photic entrainment of those species studied, forced internal desynchronizationbetween the activity rhythm and other circadian functions might occur, as has been foundin squirrel monkeys (Moore-Ede et al. 1982).
Both single light pulse photoperiods and various skeleton photoperiods entrained thefree-running CAR to 24 h in diurnal Callithrix and nocturnal Aotus. Phase positions of theentrained rhythms always corresponded to those expected from a knowledge of therespective spontaneous periods and PRCs (Rauth-Widmann et al. 1991; Wechselberger1995).
The time needed for re-entrainment (tr) after sudden phase-shifts of synchronizing LDcycles may differ depending on species, circadian parameter, direction (advance or delay)and amount of shift, as well as on the L:D luminance and time relationship, i.e. theZeitgeber strength. In Table 2 this is documented for some nocturnal and diurnal primatespecies. For instance, after 8-h advances and delays (DF þ/7 8 h) of an LD 12:12 cycle(56:0.07 lux), nocturnal Galago senegalensis re-entrained on average within about 9.2 and 5days, respectively, while they took about 7.4 and 5.6 transient cycles to re-adapt tocorresponding shifts of an LD 8:16 cycle. Otolemur garnettii showed an opposite directioneffect in re-entrainment. The subjects required less transient cycles to re-adapt to an 8-hadvance than to compensate for an 8-h delay of the LD 12:12 (Schanz & Erkert 1987). InA. lemurinus griseimembra, re-entrainment times of 5.3 and 2.7 days were determined after
238 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Table2.
Re-entrainmentbehaviourofcircadianrhythmsin
diurnalandnocturnalprimates.Tim
esneeded
forre-entrainment(tr)
after
advance
anddelay
shifts
(DFþ/7
)ofsynchronizinglight–dark
cycles.
Species
Act.
nParameter
L:D
cycle
h(lux)
Shift
DF
(h)
trafter
DFþ
(advance;d)
trafter
DF7
(delay;d)
Reference
Strepsirrhini(Prosimii)
Microcebusmurinus
no
6LA
12:12(100:0)
88
3.7
SchanzandErkert,
unpub.
Eulemurfulvusalbifr.
cat
5LA
12:12(140:0.1)
87
3.7
SchanzandErkert,
unpub.
Galagosenegalensis
no
5LA
12:12(56:0.07)
89.2
+1.6
5.0
+0.7
SchanzandErkert1987
8:16(56:0.07)
87.4
+0.8
5.6
+0.5
SchanzandErkert1987
Otolemurgarnettii
no
3LA
12:12(56:0.07)
86.3
+1.5
10.0
+1.0
SchanzandErkert1987
Haplorrhini(A
nthropoidea)
Aotuslemurinusgris.
no
5LA
12:12(243:0.3)
85.3
+0.6
2.7
+0.6
Thiemann-Jaeger
1986
5LA
8:16(243:0.3)
811.0
+1.0
5.0
+1.0
8LA
3:21(100:0.2)
810.2
+1.9
7.6
+1.8
Rappold
andErkert
1994
Callithrixj.jacchus
di
14
LA-on
12:12(280:0.2)
86.6
+0.9
6.4
+0.7
Erkert1989
LA-end
12:12(280:0.2)
86.8
+0.7
8.6
+1.3
5LA-on
12:12(410:0.2)
85.4
+1.5
(4.8
+1.6)
Fischer
andErkert,
unpub.1
LA-end
12:12(410:0.2)
86.8
+0.8
(6.0
+1.2)
Saim
irisciureus
di
4LA
12:12(60:0)
85.8
+1.2
a2.9
+0.9
aWexlerand
Moore-Ede1986
Tb
12:12(60:0)
85.2
+0.5
a4.1
+0.5
a
4A,F,D,Tb12:12(n.i.)
82–6
Moore-Edeet
al.1977
K,Na,UV
12:12(n.i.)
86–12
6Tb
12.3:11.7
(500:0.001)
83.8
+0.7
5.4
+1.0
Bouloset
al.1996
Act:activitytype:
di:diurnal;no:nocturnal;cat:cathem
eral.
Parameter:LA:locomotoractivity;A:motoractivity,Tb:bodytemperature;F:Feeding;D:drinking;K,Na:renalpotassium
andsodium
excretion;UV:urinevolume.
tr:
aithmetic
means+
S.D
.orS.E.a.
1trafter
onesingledose
of50ml
10%
ethanolwithoutandwith(inbrackets)250mg
melatonin
administeredorally(inmealworm
s)3hbefore
lights-offonthefirst8-h
advanced
LD
cycle.
Biological Rhythm Research 239
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

8-h advance and delay shifts of an LD 12:12 (240:0.3 lux) cycle, respectively, and times of11 + 1 and 5 + 1 d after corresponding shifts of an LD 8:16 (240:0.3 lux) cycle(Thiemann-Jaeger 1986). To 8-h advance and delay shifts of an LD 3:21 (100:0.2 lux)cycle, they adjusted within 10.2 and 7.6 d, respectively (Rappold & Erkert 1994). Indiurnal marmosets, tr for activity onset was slightly different from that of the end ofactivity, both after 8-h advance and delay shifts of an LD 12:12 (280:0.2 lux) cycle (Table2, Erkert 1989).
Different re-entrainment times for the onset (evening) and end (morning) componentof the activity pattern were observed in an Otolemur garnetti. Following a 6-h delay of anLD 12:12 (450:0.3 lux) cycle, the activity onset component of the CAR re-entrainedorthodromically (by delaying) while the offset component adjusted antidromically, in thislatter case taking as long as about 40 days (Erkert 2006).
In chair-restrained squirrel monkeys, the circadian rhythms of activity, feeding,drinking, and body temperature were found to re-entrain significantly faster after an 8-hdelay shift of an LD-Zeitgeber than the rhythms of urinary K and Na excretion and urinevolume (Moore-Ede et al. 1977). After 8-h advance and delay shifts of an LD 12:12 (60:0lux) cycle, the squirrel monkeys’ body temperature (Tb) rhythm was found to re-entrainwithin about five and four days, respectively, while the activity rhythm needed about sixand three days (Wexler & Moore-Ede 1986). In both cases, differing tr’s of the recordedcircadian parameters indicated transient internal desynchronization. After 8-h advanceand delay shifts of an LD 12:12 (500:0.001 lux) cycle, with and without interposed twilight,Boulos et al. (1996) found a significant dependence of tr in the Tb-rhythm of Saimiri onthe direction of the Zeitgeber shift but not on the presence or absence of twilight.
Thus, in all primates tested so far, more or less pronounced direction effects occurredin the times needed for re-entrainment after advance and delay shifts of LD cycles(Table 2). Only in part of the species the direction effect was related to the circadian period(t5/424 h). In the others the phase-response characteristics might have played a morecrucial role for the circadian system’s re-entrainment behavior, which did not differbetween diurnal and nocturnal species (c.f. Table 2).
Masking direct effects of light. In all primate species studied thus far, pronounced directinhibitory or enhancing effects of high or low light intensities, respectively, have beenshown that masked free-running and/or entrained circadian functions such as theactivity – rest or body temperature rhythm (Figures 2–4; Aschoff et al. 1982; Gander &Moore-Ede 1983; Erkert & Groeber 1986; Rappold & Erkert 1994; Erkert et al. 2006). Indiurnal species, the magnitude of the inhibitory (negative) photic masking effects on thecircadianly programmed activity level is usually negatively correlated with light intensity.Nocturnal primates, however, often show strong negative light masking both at high andvery low luminosities. For instance, in Aotus lemurinus, an optimum function for thedependence of total daily activity on darktime luminosity (with an LD 12:12 cycle withabout 100 lux in L and stepwise reductions on D-time luminosities) was established(Figure 5). The luminosity of about 0.1–0.5 lux, at which highest activity level occurred,corresponded to full moonlight (Erkert 1976b). Due to this strong dependence of thelocomotor activity on darktime luminosity in Aotus, all kinds of activity patterns could beproduced in a predictable manner, simply by providing adequate patterns of darktimeluminosity (Erkert & Groeber 1986). Since the inhibitory effect of short-term reductions ofluminosity during the night seems to depend to a small degree on the circadian phase, itmay be concluded that primates might also show some circadian modulation of theirsusceptibility to the inhibitory effects of low light intensities (cf. Mrosovsky 1999).
240 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Largely in parallel with the light-induced variation of the activity pattern, the owlmonkeys’ core temperature rhythm was also modulated. Though this may largely beattributed to the inhibitory effects of light on motor activity and muscle tone, additionalmasking direct effects of low light intensities on the circadian level of Tb seem to occur(Erkert & Groeber 1986). Studying the impact of continuous 2:2 h short-term LD cycles of60:0 lux on the activity and body temperature rhythm of diurnal squirrel monkeys, Ganderand Moore-Ede (1983) came to a similar conclusion. However, since they coulddemonstrate that these LD-related short-term modulations in both circadian functionsoccurred during the monkeys’ resting time also, though to a lesser extent, the relativelydim light of 60 lux in this case did not merely have a permissive effect but also exerted anenhancing direct effect, i.e. positive masking. Analyzing the effect of short-term light –dark cycles (LD 2:2; 200:0 lux) on circadian functions in Saimiri, Robinson and Fuller(1999) showed not only that the circadian Tb rhythm but also the patterns of heatproduction and heat loss (both of which contribute to a determination of the Tb rhythm)were negatively masked by darkness.
Figure 1 shows that, in these three nocturnal prosimians also, due to a strong negativemasking effect of very low light intensities, the darktime activity level in the LD 12:12(100:0.1 lux) cycle is considerably higher (bottom) than in LDs with physiological (total)darkness in the dark (top). In Galago and Otolemur, the same holds for the free-runningCAR, as shown in an LD 12:12 cycle with phsiological darkness in D and dim light of 0.1
Figure 2. Averaged patterns (n ¼ 10 d each) of the free-running circadian body temperaturerhythm in four owl monkeys (Aotus lemurinus griseimembra) kept in LL of 0.1 lux at constantambient temperatures of 30 and 208C (upper and lower curves, respectively). Note that the lower Ta
not only led to a generally lower Tb level but, in three animals, also to a distinct ultradianmodulation of the circadian Tb pattern.
Biological Rhythm Research 241
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

lux in L (Figure 1, middle), which, for these species, is well below the threshold for photicentrainment. The fact that in Otolemur the period of the free-running CAR, despite thestrong masking effects of the LD 12:12 (0.1:0 lux) cycle, does not normally show anyvariation, that is, a sign of relative coordination, indicates that masking by light, at least inthis case, is not mediated via the circadian pacemaker in the SCN but at some pointdownstream from it (Erkert et al. 2006).
Figure 3. Upper panel: Average activity pattern of Eulemur fulvus albifrons (n ¼ 5) in an LD 12:12(140:0.1 lux) cycle. Shaded areas indicate the light time. Lower panel: changing activity pattern of anE. fulvus albifrons pair in an LD 12:12 cycle with 140 lux in L and stepwise reductions in luminosityduring the dark phase, as indicated on the right margin. Note that the lemurs behaved as dark-activewith 0.1 lux and light-active with physiological darkness in the D phase, and that, in the subsequentconstant dim light of 0.1 lux, the rhythm’s activity phase started to free-run from the dark portion ofthe preceding LD cycle. For further information, see text. From Erkert HG, Cramer B: FoliaPrimatol 2006; 77: 87–103, S. Karger AG, Basel.
242 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011
Particularly strong direct inhibitory effects of low dark-time luminosities that totallymasked the animals’ CAR occurred in the white-fronted lemur Eulemur fulvus albifrons(Figure 3; Erkert & Cramer 2006). In nature, the cathemeral E. f. rufus and E. f.mayottensis are much more active throughout the day than the night (Donati et al. 2001;Kappeler and Erkert 2003). Under laboratory conditions (LD 12:12 of 145:0.15 lux,ambient temperature ¼ 238C, 60% relative humidity, food and water ad libitum),however, E. f. albifrons became dark-active and developed, on average, 81.8 + 5.5% of itsdaily total activity during the dark time (Figure 3, top). Reduction of dark-time luminosityto 0.002 or 0.0002 lux caused the animals to become ‘‘cathemeral’’, in that they nowdeveloped an average of around 40–60% of total daily activity during the light. With 1075
lux or physiological darkness in the dark, the animals became light-active, showing up to90% of their total daily activity in the light phase. Despite this, on a subsequent LL 0.15lux regimen, their circadian activity phase started free-running from the previous darkphase. This indicates that activity had remained in phase with the dark time of the LDcycle (Figure 3, bottom). Corresponding results were obtained in another trial in which thedark-time luminosity (of 0.15 lux) of an LD 12:12 cycle was suddenly switched off for two
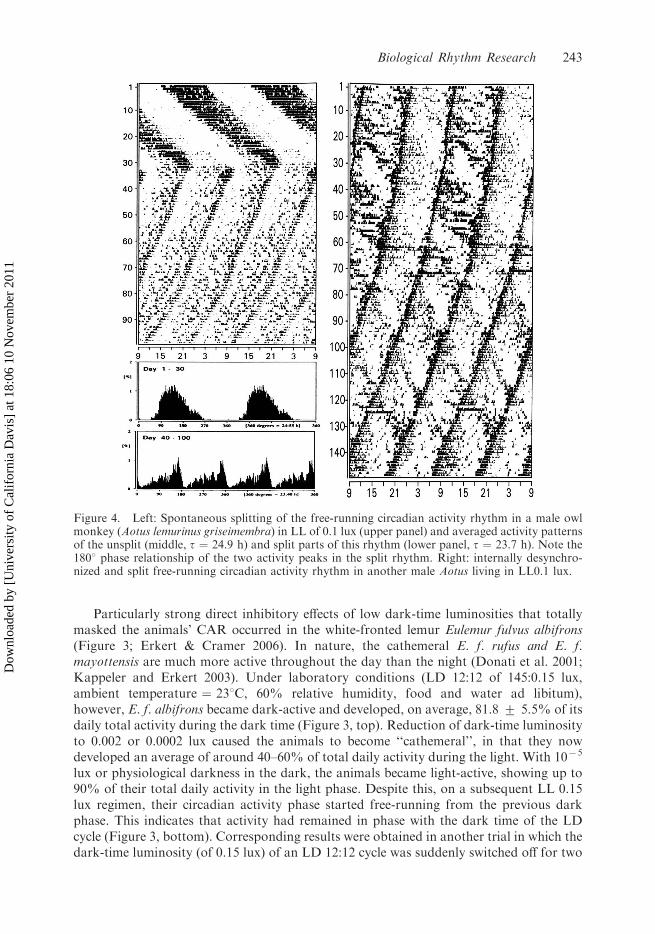
Figure 4. Left: Spontaneous splitting of the free-running circadian activity rhythm in a male owlmonkey (Aotus lemurinus griseimembra) in LL of 0.1 lux (upper panel) and averaged activity patternsof the unsplit (middle, t ¼ 24.9 h) and split parts of this rhythm (lower panel, t ¼ 23.7 h). Note the1808 phase relationship of the two activity peaks in the split rhythm. Right: internally desynchro-nized and split free-running circadian activity rhythm in another male Aotus living in LL0.1 lux.
Biological Rhythm Research 243
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Figure 5. Owl monkey activity patterns under semi-natural conditions, in laboratory settings and inthe wild. Left panel: lunar periodicity of nocturnal activity in an Aotus lemurinus griseimembra pairkept under semi-natural conditions in Central Colombia. Hourly activity counts every other day overa period of about two months. Second column: lower panel: average activity patterns of another A.lemurinus pair (n ¼ 13months) at four phases of the moon (new, waxing, full and waningmoon;+1 deach) and time course of night-time luminosity (curves) measured during corresponding nights duringone lunar cycle (from Erkert 1974). Upper panel: effect of dark-time luminosity of an LD 12:12 cyclewith about 100 lux in L on the total daily activity of two A. lemurinus pairs (from Erkert 1976). Thirdcolumn: average activity patterns of A. lemurinus (n ¼ 7) in rectangular LD cycles simulating thenocturnal lighting conditions at the four phases of the moon, as indicated by the curves on top(luminosity is indicated by the y-axes at the top right of the diagrams; pd ¼ physiological darknessassumed below 1076/77 lux). From Erkert and Groeber 1986. Right panel: activity rhythm in a wildfemale A. azarai owl monkey of the Argentinean Chaco showing similar lunar periodicity asA.lemurinus (30 min counts of 100–1000 impulses are plotted). For more details, see text.
weeks and the lemurs thereafter were subjected to LL 0.15 lux for some days (Erkert &Cramer 2006). In parallel to the light-induced phase reversal of the lemurs’ activity – restrhythm, the two peaks of their Tb rhythm were only slightly phase-delayed and advanced,respectively. When starting to free-run in the subsequent LL 0.15 lux protocol, the twotemperature peaks immediately re-adopted their former phase position.
From these results, it may be concluded that, although the lemurs became cathemeralor even light-active, their circadian systems’ phase-setting mechanism behaved like that ofa ‘‘nocturnal’’ species. Hence the observed ‘‘cathemeral’’ and ‘‘diurnal’’ behavior in theselemurs resulted mainly from strong, direct activity-inhibiting effects of low light intensities,presumably in combination with higher tolerance of and/or a greater activity-enhancingeffect of the higher luminosity during the light phase. That is, it seems to have been theresult of extreme ‘‘light masking’’ (Erkert & Cramer 2006).
Nonphotic entrainment and masking
Ambient temperature (Ta). The daily variations of Ta and relative humidity generallyprovide much more imprecise and unpredictable time cues for circadian systems than thelight – dark cycle. Since Ta rhythms, in addition, often have considerably smaller
244 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

amplitudes and may vary substantially with the habitat’s latitude, altitude, geographic andgeomorphologic characteristics, as well as with vegetation, they are a priori less suitable tobecome reliable Zeitgebers for circadian entrainment in endothermic mammals. Inprimates, the effects of constant Tas and temperature cycles (TCs) on the parameters ofcircadian rhythmicity have been studied in Macaca nemestrina, Saimiri sciureus, Callithrixjacchus and Aotus lemurinus (Tokura & Aschoff 1983; Aschoff & Tokura 1986; Erkert1991; Palkova et al. 1999).
No significant Ta effects on t were found in owl monkeys (Figure 2) and commonmarmosets (tested in the range 20–308C; Erkert 1991; Palkova et al. 1999) as well as insquirrel monkeys (range 15–328C; Aschoff & Tokura 1986). In the much larger pigtailedmacaque, however, in LL of 100 and 450 lux, t was significantly longer at 328C than at178C, while, at lower LL luminosities, no significant temperature effect on t could beestablished (Tokura & Aschoff 1983).
Trapezoidal TCs of 20–308C failed to entrain the free-running circadian activity andTb rhythms in owl monkeys or to modulate their period length, while, in somemarmosets, it led to entrainment and in others to period changes indicating relativecoordination (Erkert 1991; Palkova et al. 1999). Squirrel monkeys and pig-tailedmacaques responded similarly to 24 h TCs of 17–328C. Most individuals showed moreor less pronounced relative coordination while a few others were entrained (Aschoff &Tokura 1986; Tokura & Aschoff 1983). While in TC-entrained marmosets and macaquesthe animals’ active phase was set to coincide with the warm fraction of the externaltemperature cycle, it coincided with the cold fraction in the TC-entrained squirrelmonkeys. According to the species’ average t (of mainly less or more than 24 h), inCallithrix and Macaca, the entrained activity time, a, adopted a more positive phaserelationship (advance) to the warm fraction (Palkova et al. 1999; Tokura & Aschoff1983) and, in Saimiri, a more negative one (delay) to the cold fraction of the temperaturecycle (Aschoff & Tokura 1986).
Except for Macaca nemestrina, all other primate species studied showed more or lessstrong direct masking effects of Ta on locomotor activity. Lower ambient temperatureusually caused higher activity levels, both with constant Ta and with TCs. In Aotusmonkeys, the Tb level also varied with changing Tas, but, despite higher activity at 208Cthan at 308C, it was higher at 308C (Figure 2). Though the ts of the activity and Tb
rhythms were not influenced by the applied TC of 20–308C, both circadian functions werestrongly masked by it: activity increased and core temperature decreased as soon as, andfor as long as, the circadian activity phase coincided with the cold fraction of the TC. Insome animals at 208C, the circadian Tb rhythm showed pronounced short-term variations(Figure 2, left) which were not obseved in the activity rhythm. This observation, and thatshowing that ultradian modulations of Tb also occurred during the cold fraction of theapplied TC 20–308C, points to their thermoregulatory origin.
Other environmental factors
Social entrainment by daily periodic contact with conspecifics has been evidenced in thediurnal common marmoset Callithrix jacchus (Erkert & Schardt 1991). Acoustic socialcontact produced entrainment in a few animals, while, in others, both acoustic and visualsocial contact merely led to rather weak relative coordination, and full social contact wasrequired for real entrainment. Thus, daily periodic social cues exert, at best, a very weakZeitgeber effect on marmoset/primate circadian systems.
By inducing locomotor activity and/or increased arousal—by presenting 2-h playbacksof agitated vocalizations of conspecifics in combination with a mirror—Wechselberger
Biological Rhythm Research 245
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

(1994, 1995) was able to elicit phase responses and to entrain the marmosets’ free-runningCAR in LL of 430 lux. Phase responses and entrainment—by 1-h activity pulsesstimulated by agitation of the cages and intermittent sprinkling with water (while lightswere off)—were also obtained in Callithrix monkeys kept isolated in dim LL of less than0.1 lux that obviously had strong inhibiting effects on these monkeys’ spontaneouslocomotor activity (Glass et al. 2001). In contrast to the marmosets living in LL 430 lux,however, in those kept in dim light, induced activity led to phase responses andentrainment characteristics corresponding to those produced in nocturnal hamsters bysocial stimuli and cage agitation (Reebs & Mrosovsky 1989). The discrepancy between theresults obtained in the two settings may be attributed to the fact that in Wechselberger’sexperiment, with high LL luminosity, the phase-shifting light effects may have dominatedthose of induced activity and/or arousal.
Evidence for social masking was found in the free-running circadian activity and Tb
rhythm in a pair of the gregarious Eulemur fulvus albifrons when living in LL 0.15 lux,either in isolation or with different kinds of social contact with each other and/or aneighboring family group of conspecifics (Erkert & Cramer 2006).
Entraining effects of time-restricted food availability (eating and fasting, EF cycles)have been tested repeatedly in diurnal squirrel monkeys, with inconsistent results. Inchair-restrained animals, Sulzman et al. (1977, 1978) found that the colonic tempera-ture, drinking and urinary excretion rhytms became synchronized to a 24-h EF cyclewith 3 h of food availability. Aschoff and von Goetz (1986), however, observed thatEF cycles induced in Saimiri, at most, weak relative coordination, while Boulos et al.(1989) found entrainment to an EF cycle in only one of 10 monkeys, and relativecoordination in one other. In both studies, the EF cycle produced distinct maskingeffects. Taken together, EF cycles may exert, at most, a very weak Zeitgeber effect on themonkeys’ CTS.
Splitting and internal desynchronization
Circadian phenomena pointing to the existence of a two- or multi-oscillator systemunderlying circadian rhythmicity, such as rhythm splitting and internal desynchronization,have been described both in nocturnal and diurnal primates. Splitting occurred in theactivity and Tb rhythm of Aotus lemurinus (cf. Figure 4, Rappold & Erkert 1994) as well asin the activity rhythm of Otolemur garnettii (Erkert et al. 2006) and Callithrix jacchus(Schardt et al. 1989; Wechselberger 1995; Palkova et al. 1999). Long-lasting spontaneousinternal desynchronization of the circadian activity and feeding rhythm has beendocumented in a male Aotus (Erkert 2000). In another Aotus, internal desynchronizationwas observed between two activity components which had periods of 23.75 and 24.75 h(Figure 4). Spontaneous, forced, and transient internal desynchronization of variousphysiological and behavioral parameters occurred in up to about 25% of the diurnalsquirrel monkeys studied by Moore-Ede and Sulzman (1977), Sulzman et al. (1977b) andHoban and Sulzman (1985).
Activity rhythms under seminatural conditions
Aotus lemurinus griseimembra
Continuous activity recordings lasting 6–13 months and carried out 1971–1972 in CentralColombia in four pairs of the North Colombian owl monkey species, Aotus lemurinus
246 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

griseimembra (Karyotype II-IV, 2n ¼ 52–54 chromosomes), clearly proved this species tobehave strictly nocturnally. Its mostly bimodal activity pattern, with peaks during duskand dawn, showed pronounced lunar periodic variations (Figure 5). Around the newmoon, the owl monkeys largely limited their locomotor activity to the twilight periods.With the waxing moon, their nocturnal activity time extended in parallel to the moonlitfraction of the night until, at the full moon, it finally covered the whole night. With thewaning moon, the animals’ active phase shortened in parallel to the gradual reduction ofthe nocturnal moonlight. The largely coincident time course of activity and night-timeluminosity at the four phases of the lunar cycle indicated that this pronounced lunarvariation of the owl monkeys’ activity rhythm was mainly an effect of masking by light(Figure 5, Erkert 1974). This hypothesis was substantiated by laboratory results showingin Aotus a darktime luminosity-dependent activity optimum at about 0.1 lux, whichcorresponds to the luminance on cloudless nights with a full moon (Figure 5, Erkert1976b). Furthermore, by simulating the nightly time course of luminosity at the fourphases of the lunar cycle with corresponding rectangular light cycles, similar activitypatterns as observed under semi-natural conditions could be produced (Figure 5, Erkert &Groeber 1986).
In addition to the variation of the owl monkeys’ activity pattern, significant lunarvariations also occurred in the phase position of the onset (Co) and end (Ce) of activity inrelation to sunset and sunrise, respectively. Around the new moon, the monkeys begantheir locomotor activity earliest (average Co ¼ þ43 min) and terminated it latest(Ce ¼ þ7 min), while, on nights with a full moon, they became active latest (Co ¼ þ13min) and entered the resting phase earliest (Ce ¼ þ23 min). For further details, see Erkert(1976a).
Ateles geoffroyi
Quantitative long-term activity recordings carried out in seven Ateles geoffroyi kept undersemi-natural conditions in the Central Mexican tropics clearly showed that this largeneotropical anthropoid behaves strictly diurnally (Figure 6, Munoz-Delgado et al. 2004).Throughout the half of the year associated with short daylengths, the spider monkeysdeveloped, on average, 91 + 1.5% and, throughout the half of the year associated withlong daylengths, 93.5 + 3.5% of their total daily activity during the light phase(t12 ¼ 71.41, p ¼ 0.18). Almost all animals had bimodal activity patterns with a firstactivity peak (in the morning) more than twice the size of the second peak (Figure 6). Peakactivity height remained largely constant throughout the year, averaging 1.41 + 0.62% ofdaily total activity/5 min during the short-day months and 1.36 + 0.62%/5 min duringthe long-day months. The average trough of the bimodal activity phase amounted to0.43 + 0.11%/5 min in the short day and 0.34 + 0.08%/5 min during the long day halfof the year (t11 ¼ 1.5, p ¼ 0.16).
Activity time changed with day length and amounted, on average, to 11.6 + 0.4 hthroughout half of the year with short days and 13.4 + 0.7 h during the half with longdays (t12 ¼ 6.2, p 5 0.001). Accordingly, the mean activity:resting time ratio (a:r) wassignificantly larger during the summer than the winter months (1.27 + 0.16 vs0.94 + 0.06; t12 ¼ 75.3, p 5 0.001). Mean peak-to peak intervals (PPI) amounted to5.63 + 0.92 h during the short days and to 7.61 + 0.52 h during the long days(t12 ¼ 74.87, p 5 0.001).
After the transition from winter to summer time, and vice versa, gradual advance anddelay shifts, respectively, of about 1-h in the first activity peak were observed. From these
Biological Rhythm Research 247
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

observations, it has been deduced that the relatively late occurrence of the morningactivity peak mainly resulted from the late feeding times which reflected this artificial 1-hshift in the animals’ environment (Munoz-Delgado et al. 2004). Individual ratios betweenthe trough values of a and r, which in species with bimodal activity patterns may be usedas an additional indicator for the degree of diurnality, varied from 11–30 in the short-dayfraction of the year and from 10–39 throughout the long-day fraction (average 18.5 + 6.3vs. 22.4 + 12.9, p 4 0.05).
The transfer of some animals from the wire-mesh enclosure to a small forest islandenclosure resulted in a slight extension of their activity time, a, and a more pronouncedevening activity peak (Figure 6). From this result, an even more pronounced bimodality ofactivity, in combination with an earlier morning activity peak, would be expected to occurin wild Ateles.
Figure 6. Upper panel: activity rhythm of a female diurnal spider monkey (Ateles geoffroyi) kept ina small forest island in Cental Mexico (double-plotted original recording over two months; 5-minbins, each of 5–200 counts). Lower panel: mean activity pattern as averaged over this two-monthrecord (5-min values expressed as % of average total daily activity) and activity time calculated by abest-fitting rectangular pattern (dotted).
248 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Long-term activity recordings in the wild
As a further example of the efficiency of comparatively long-term activity recordings withan actiwatch in wild diurnal and nocturnal primates, Figure 7 shows (double plotted) 150-day records in one Malagasy Verreaux’s sifaka and red-fronted lemur as well as in anAotus a. azarai owl monkey of the North Argentinean Chaco. These original data, andthe mean activity patterns, obtained by averaging over the whole registration period(Figure 8), provide evidence that the three species represent different chrono-ecotypes.A common trait in their activity patterns is a distinct bimodal time course with verypronounced peaks during dawn/early morning and late afternoon/dusk.
Propithecus undoubtedly behaved strictly diurnally. Throughout the short-day period,the sifakas exhibited, on average, 85.4 + 2.5% of their total daily activity within the lightportion of the day and, throughout the long-day period, 91.5 + 1.4%. The average aamounted to 11.1 + 0.7 h during the short-day months and to 14.8 + 0.4 h during thelong day months (t11 ¼ 711.72, p 5 0.001). Correspondingly, the average a:r-ratiodiffered significantly between the two halfs of the year (t11 ¼ 712.73, p 5 0.001; cf.Figure 10). Statistically significant differences between the two half-year periods occurredalso in the amplitudes of the morning and evening activity peaks, the trough betweenthem, and in the time of the evening peak, but not in that of the morning peak or the timeof the trough (Figures 9A, B). Analyzing the monthly averages of those parameters forannual variations by ANOVA yielded consonant results (cf. Erkert & Kappeler 2004, p.180, Table 3).
Eulemur fulvus rufus also showed most of its total daily activity during the light portionof the natural light – dark cycle. During the short-day period, the lemurs developed, on
Table 3. Comparision of average activity pattern parameters between the wild Malagasy lemurspecies Propithecus v. verreauxi (P; diurnal) and Eulemur f. fulvus (E; cathemeral) and the NorthArgentinean owl monkey Aotus a. azarai (A; nocturnal) by post-hoc ANOVA and Scheffe tests.
Parameter
Short-day period Long-day period
F14 p - sign.Scheffe-Test
sign. F16 p - sign.Scheffe-Test
sign.
A/a 0.45 0.654 7 6.29 0.012 þ E-Aa/50 0.46 0.644 7 11.27 0.001 þþþ P-A; E-Aa/r 55.08 50.001 þþþ P-E; P-A; E-A 3.33 0.066 (7)a:r 30.37 50.001 þþþ P-A; E-A 3.51 0.058 (7)a-a/50 1.41 0.282 7 9.86 0.002 þþ P-A; E-Aa-r/50 9.78 0.003 þþ P-E; E-A 27.51 50.001 þþþ P-E; E-Aa-a:A/50 122.28 50.001 þþþ P-E; P-A 39.28 50.001 þþþ P-E; P-Aa-r:A/50 16.37 50.001 þþþ P-E; P-A; E-A 35.90 50.001 þþþ P-E; P-A; E-Aa-a:a-r/50 66.86 50.001 þþþ P-E; P-A 40.94 50.001 þþþ P-E; P-Ap1-a 6.70 0.011 þ E-A 30.65 50.001 þþþ P-A; E-Ap2-a 1.89 0.193 7 15.86 50.001 þþþ P-A; E-Atr-a 58.31 50.001 þþþ P-E; P-A 4.92 0.024 þ E-Atr-r 4.06 0.045 þ P-A 6.85 p.008 þþ E-Apt1-a 370.86 50.001 þþþ P-A; E-A 597.52 50.001 þþþ P-E; P-A; E-Apt2-a 653.82 50.001 þþþ P-E; P-A; E-A 1253.79 50.001 þþþ P-E; P-A; E-Atrt-a 52.91 50.001 þþþ P-A; E-A 546.85 50.001 þþþ P-A; E-ATrt-r 16.65 50.001 þþþ P-A; E-A 57.59 50.001 þþþ P-E; P-A; E-A
For parameter abbreviations see legend to Figure 9B; p: probability of error; sign.: significance: 7, (7), þ, þþ,þþþ: not, just not, just, highly, very highly significant.
Biological Rhythm Research 249
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

average, 69.4 + 5.3% and, during the long-day period, 80.2 + 4.1% of their total dailyactivity during a (t7 ¼ 73.47, p ¼ 0.001). Statistically significant differences between thetwo halves of the year were also present in daily total activity, the activity and restingtimes, the a:t-ratio, the amplitude and time of the evening activity peak, the troughbetween the two activity peaks, and in the activity level throughout a (cf. Figures 9A, B).
Because the lemurs also developed considerable amounts of locomotor activity duringthe brighter moonlit parts of the night (Figure 5 right, Figure 7), the ratio between theiractivity level (% of daily total activity/h) throughout the day/twilight span from 05:00 to20:00 and the dark night hours from 20:00 to 05:00 varied with a lunar periodicity. Overthe course of a year, this ratio amouted to 2.72 on nights with a new moon and to only1.46 on nights with a full moon (long-day months: 3.32:1.73; short-day months: 2.19:1.21;Kappeler & Erkert 2003). Such relationships between the activity levels during day andnight may justify the classification of Eulemur fulvus as a cathemeral species.
Aotus azarai monkeys of the North Argentinean Chaco were noticeably more activethroughout the night than the day, but their nocturnal activity usually showed a much
Figure 7. Double-ploted, original long-term activity records from one wild Verreaux’s sifaka(Propithecus v. verreauxi), one red-fronted lemur (Eulemur fulvus rufus) and one Argentinean owlmonkey (Aotus a. azarai). Records obtained by Actiwatch1 actimetry (5-min counts) throughout afive month period during the long-day half of the Austral year. Note the pronounced bimodalpattern in all species, that Propithecus and Eulemur behave mainly diurnally while Aotus shows mostactivity during the night, and that, in both Eulemur and Aotus, a distinct lunar periodic componentoccurs with lowest activity during the nights of the new moon (indicated by closed circles at the rightmargin) and highest activity around the time of the full moon.
250 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

more pronounced lunar periodicity than that observed in the red-fronted lemurs (Figure 7,Fernandez-Duque & Erkert 2006). Throughout the short-day period, the Aotus developed,on average, 83.6 + 7.2% and, during the long-day period, 86.3 + 2.7% of the total dailyactivity within the activity time (t8 ¼ 70.77, p ¼ 0.46), which, around the new moon,usually extended into the late morning hours. Average relationships between the activitylevels during a and r were 40.6 + 9.9:15.9 + 4.5 and 37.6 + 9.5:0.1 + 2.3 counts/5 minduring the short- and long-day halves of the year, respectively (Figure 9A). Significantdifferences between the parameters of the average activity patterns from the two halves ofthe year were found only in the amplitude of the evening peak, the trough between the twoactivity peaks, and in the times of peaks (Figures 9A, B).
From Figure 9(A, B), which compares relevant parameters of the three species’bimodal activity patterns during the short-day and long-day period of the year, it can also
Figure 8. Bimodal, long-day activity patterns in Propithecus verreauxi, Eulemur fulvus, and Aotusazarai, obtained by averaging the 5-min activity records shown in Figure 8. Ordinate values are givenas % of average total daily activity. Dotted rectangles indicate calculated average activity and restingtimes (a, r). Note the generally low night-time activity level in the clearly diurnal Propithecus and therelatively high night-time activity of Eulemur, the peak of which surpasses the daytime trough atnoon. That the Aotus’ morning activity bout extends far into the morning hours is due mainly to acompensatory lengthening of a around the time of the new moon (cf. Figure 7). Since Argentineanofficial time is one hour phase-advanced in relation to local time, the owl monkey activity patternmust be correspondingly advanced in order to become directly comparable with the other twopatterns. It then becomes clearer that all three patterns might have been derived originally from morecrepuscular patterns.
Biological Rhythm Research 251
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

be seen that, between the two halves of the year in the owl monkeys, some of theparameters, such as total activity/day, a, r, and a:t, and activity levels during activity andresting time (a-a and a-r/5min), varied in opposite directions to those values seen in thetwo Malagasy lemurid species. These results also suggest the characteristics of the owlmonkeys’ CAR more in keeping with a nocturnal response. On the other hand, the highestactivity levels during the resting time and the lowest quotients between the activity levelsduring a and r indicate that Eulemur actually exhibited the most cathemeral behavior ofthe three species when studied quantitatively in the wild.
Table 3 summarizes the results of statistical comparisions (using ANOVA followed byScheffe tests) of the individual parameters of the averaged activity patterns in the three
Figure 9A.
252 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Figure 9B. Comparison of various parameters of the bimodal short day (sd) and long day (ld) activitypatterns in wild Malagasy lemurids (Propithecus v. verreauxi:P.v., n¼ 6/7; Eulemur fulvus rufus:E.f.,n¼ 4/5) and North Argentinean Aotus a. azarai owl monkeys (A.a., n¼ 5). Arithmetic means+SE.Panel A: A/d: total activity/day (kc¼ kilo [1000] counts); a-a: percentage of daily total activityoccurring during calculated activity times; alpha (a): calculated activity time; rho (r): calculated restingtime; a-a/50:average activity level during a (counts/5 min); a-r: average activity level during r (counts/5 min); a-a:A/50: activity level during a related to the overall mean of activity/5 min; a-r:A/50: restingtime activity level related to the 5-min overall activity level. Panel B: a:r:activity time:resting timequotient; a-a:a-r/50: relationship between the activity levels during a and r; p1-a, p2-a: height of the firstand second activity peaks (as % of average total activity/day); tr-a, tr-r: height of activity minimum(trough) during activity and resting time; pt1-a, pt2-a: time of first and second activity peak of thebimodal activity patterns; trt-a, trt-r: daytimes of lowest activity levels during activity and resting time.Note that peak numbering refers to the diurnal and nocturnal activity pattern rather than to morningand evening.
Biological Rhythm Research 253
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

species for the short-day and long-day period. In almost all cases there were significantspecies differences. For both the short- and long-day halves of the year, the differenceswere least between Propithecus and Eulemur (n ¼ 7); on the other hand, during the short-day periods, the largest differences were found between Eulemur and Aotus (n ¼ 13) and,during the long-day periods, between Propithecus and Aotus (n ¼ 11 parameters).
Discussion
Laboratory studies
Functional characteristics of primate circadian systems as compared to those of rodents
Primate circadian systems do not differ fundamentally from those of rodents. From lesionexperiments carried out in rhesus and squirrel monkeys, the hypothalamic suprachias-matic nuclei (SCN) have also been identified as the main central nervous pacemaker(Reppert et al. 1981; Fuller et al. 1981; Moore-Ede et al. 1982; Albers et al. 1982, 1984a,1984b; Edgar et al. 1993). The location and appearance of the monkeys’ SCN (Macaca,Saimiri, Callithrix), as well as the neurotransmitters present, largely resemble those ofnocturnal rodents. Retinal afferents from the retino-hypothalamic tract (RHT) alsoterminate in the ventral part of the SCN but, other than in rodents, most RHT fibers stemfrom the ipsilateral retina. Furthermore, the monkeys’ intergeniculate leaflet (IGL), whichalso receives retinal input and projects back to the SCN via geniculo-hypothalamic tract(GHT) fibers, is described as being a much larger component of the geniculate complexthan in rodents, and the efferent projections of the SCN seem more widely distributed(Lydic et al. 1981; Reppert et al. 1987; Murakami & Fuller 1990; Moore 1993; Costa et al.1998). Since comparative immuno-histological studies in other primate species, especiallyin nocturnal prosimians, are still lacking, it is not yet clear whether or not these differencesare specific to primates in general and may be of functional significance for the CTS.Rhythmic clock gene expression (Bmal1, Cry, Per1), documented in the adrenal gland ofMacaca mulatta, indicates that non-human primates have a similar set of clock genes as dorodents and that the adrenal gland may possess an intrinsic circadian clock (Lemos et al.2006).
Taken together, the response characteristics of primate circadian systems, such asperiod variability, shape and amplitude of PRCs, re-entrainment behavior after phaseshifts of LD-Zeitgebers, and ranges of entrainment, do not differ substantially from thosedescribed in rodents. The exception to this general statement might be the large deviationof the circadian period from 24 h that is found in the nocturnal prosimians Microcebusmurinus (Schilling et al. 1999) and Otolemur garnettii (Erkert et al. 2006). Compared withthe large species-specific variation in the plasticity of the CTS response characteristicsestablished in nocturnal chiroptera (cf. Erkert 1982), we may state that non-humanprimates generally possess a well-developed and relatively precise circadian system ofmedium plasticity.
Circadian response characteristics in diurnal and nocturnal species
Comparisions of the response characteristics of the various circadian parameters inprosimian and simian, as well as in diurnal and nocturnal, species reveal no fundamentaldifferences, either between the two primate suborders or the two chrono-ecotypes. Bothgroups comprise species/subspecies with periods longer and shorter than 24 h, though allthree nocturnal prosimians studied had relatively short spontaneous periods. As in most
254 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

other diurnal mammals, parametric light effects on t in anthropoid primates do not follow‘‘Aschoff’s rule’’, except in Callithrix (Aschoff 1964, 1979; Wechselberger 1995). Beingvalid only for (most) nocturnal mammals (Aschoff 1979), this rule has not achieved thediscriminatory power originally hoped for. Therefore, the parametric light effects on t areas unsuitable as the PRC for evaluating if a given species’ CTS shows genuine diurnal ornocturnal response characteristics; the best method for this still remains to carry outphotic entrainment/re-entrainment experiments (cf. Refinetti 2006). These strictures alsoapply to the rather weak and unpredictable (often even lacking) parametric and entrainingeffects of Ta and TCs on the CTS of diurnal and nocturnal species.
In non-human primates, diurnal or nocturnal behavior, i.e. photic phase-setting of theactive phase to the bright or dark portion of the day, can be explained by the PRC nomore satisfactorily than in rodents (cf. Smale et al. 2003; Refinetti 2004). Between primateand rodent representatives of those two chrono-ecotypes studied so far, no real substantialdifferences seem to exist with respect to shape and amplitude of the photic PRC. It is alsodifficult to imagine that the small differences between the PRCs of nocturnal and diurnalmammals, found by Daan (2000) when comparing averaged curves of a number of diurnaland nocturnal mammalian species (mostly rodents), might account for the opposite phase-setting in diurnal and nocturnal species of the active phase of the endogenous rhythms today or night. Whether additional photic period responses may produce such phasedifferences seems just as doubtful. Furthermore, no obvious differences between diurnaland nocturnal primates are discernible thus far concerning the species-specific variationsof circadian paramaters that are indicative of the CTS’ degree of plasticity andadaptability—such as period variability, range of photic entrainment, or re-entrainmenttime. Whether or not there are characteristic differences between the thresholds for photicentrainment in diurnal and nocturnal primates remains to be tested. From the relativelylarge threshold differences found in nocturnal primates (50.1 lux inMicrocebus and Aotusas compared to 43–30 lux in Otolemur), it is impossible to give a firm estimate for thethresholds to be expected in diurnal primate species.
Masking direct effects of light
Masking of circadian rhythms by direct activity-inhibiting and activity-enhancing/disinhibitory effects of high or low environmental luminosity seems to be much morepronounced in primates than in most other mammalian species (cf. Figures 1, 3, 5, 7). Thisresult might, at least partly, be related to the fact that primates in general are more visuallyoriented than most other mammals. Most of them live and move (climb/jump) in a three-dimensional world, in which effective visual long-distance detection of suitable food (suchas ripe fruits, blossoms or young leaves), of potential social partners or competitors, andof dangerous aereal or terrestrial/arboreal predators may be an essential prerequisite forsurvival and thus of selection benefit. Impaired vision at reduced ambient luminositymight therefore, a priori, be expected to also have negative, i.e. inhibitory, consequenceson these animals’ locomotion and muscle activity or tone, and, in turn, also upon heatproduction and the body temperature rhythm. But the fact that both diurnal andnocturnal prosimian and simian primates are equally affected by low luminances in theirfamiliar cages indicates that a more basic mechanism is also involved in the strongmasking effects of light in primates.
While inhibitory masking effects of light in diurnal primates are usually negativelycorrelated with ambient luminosity and increase with decreasing light intensity, innocturnal species they often obey an optimum (‘‘inverted U’’) function (Figure 5).
Biological Rhythm Research 255
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Luminances above and below a certain value, which may correspond to full moonlight atabout 0.1–0.5 lux, as in Aotus, or be below that, as in Galago, Otolemur, or Microcebus,produce increasing inhibition of behavioral activities. Comparable functions in maskingeffects of ambient luminosity, though with maxima at different luminance levels, are alsoknown to exist in some nocturnal rodents, a few bat species, and owls (Erkert 1982;Aschoff 1988; Mrosovsky 1994, 1999). However, speaking here of ‘‘paradoxical positive ornegative masking’’, as proposed by Mrosovsky (1999), seems inappropriate. None of theseterms has been unequivocally defined. Instead, they refer to comparisions between quitevariable stages of activity that inhibit or enhance/disinhibit effects of given ambientfactors—such as luminance or temperature—on the circadianly pre-set activity level of ananimal or species. For instance, Aotus monkeys usually show maximal spontaneouslocomotor activity in the dark with luminosities of about 0.1–0.5 lux that correspond tofull moonlight (Figure 5). Both rising and lowering luminosity in the dark cause increasinginhibition of activity, i.e. ‘‘negative masking’’ according to some authors (Aschoff et al.1982; Aschoff 1988; Mrosovsky 1994, 1999). In the case of entrainment by an LD 12:12cycle of about 100:0.0001 lux, for instance, interposed dim light of 0.001 or 0.01 lux duringthe dark phase would immediately induce increased activity. According to Mrosovsky’s(1999) specified masking terminology, in both cases, the direct activity-enhancing or, moreprobably, disinhibiting effects of light would have to be called ‘‘paradoxical positivemasking’’. In an LD cycle with 0.1 lux in D, however, the pronounced immediate drop ofactivity caused by temporarily lowering the D-time luminosity to 0.01 or 0.001 lux wouldhave to be called ‘‘paradoxical negative masking’’. Thus, depending on the startingconditions, identical luminance levels in one case would produce ‘‘paradoxical positive’’and in the other ‘‘paradoxical negative’’ masking. This is unsatisfactory and quiteconfusing. Such terminology appears inadequate and should not be adopted, not even inchronobiology with its ‘‘clock-centered’’ approach. Aschoff et al. (1982) introduced theterm ‘‘masking’’ for all non-circadianly mediated direct (acute) effects of physical andbiotic environmental factors on circadianly (co-)regulated behavioral and physiologicalfunctions. However, many or even most of such direct effects of environmental factors onphysiology and behavior, i.e. on the various effector systems, though not mediated by thebody clock, do in fact represent a further (probably even primary) regulatory mechanism.This mechanism may be as important as the CTS for an organism’s optimal time-nicheoccupation and adaptation to the manifold challenges of its physically and biotically time-structured environment. We should not ignore its role in most of life’s timing processes indiurnal and nocturnal mammals by considering it merely as ‘‘masking’’ of circadianrhythms, i.e. as some kind of white noise, which has to be removed or minimized(‘‘purified’’, Minors et al. 1966) before a chosen marker rhythm may be chronobiologicallystudied adequately. That’s why we, like Rensing (1989), who also asked ‘‘Is masking anappropriate term?’’, generally prefer to speak of ‘‘direct effects’’, of ‘‘rhythm-maskingdirect effects’’ or, probably the best, of circadian and non-circadian effects of light,ambient temperature, etc. (Haeussler & Erkert 1978; Erkert & Groeber 1986). Forecologists, behaviorists, sensory physiologists and non-chronobiologic neuroscientistswho, for many decades, have been familiar with this phenomenon, a less clock-centeredbut more generally applicable term would be more intelligible.
Activity rhythms under seminatural conditions and in the wild
During the past two decades, numerous field studies have been carried out on behavioralecology and sociobiology in diurnal and nocturnal mammals, including primates. Many of
256 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

those studies, besides analyzing the species’ time budgets, also dealt in detail with thetiming and daily patterns of their various behavioral activities. Findings of such studies onMalagasy Eulemur and Hapalemur species led to the differentiation of a further activitytype in addition to the generally distinguished chrono-ecotypes of diurnal, nocturnal,crepuscular, and non-circadian (arrhythmic or ultradian) species: the ‘‘cathemeral’’animals (Tattersall 1987, 2006; Colquhoun 1998; Curtis & Rasmussen 2002, 2006; Curtiset al. 1999; van Schaik & Kappeler 1996; Donati et al. 2001; Donati & Borgognini-Tarli2006; Fernandez-Duque 2003). These animals are neither arrythmic nor do they show aregular ultradian periodicity, but they develop a clear diel rhythm (Halle 2006) regulatedby a strong CTS (Erkert & Cramer 2006). The cathemeral species differ from diurnal,nocturnal, and crepuscular species in that they do not restrict daily activity to the bright ordark portion of the day but extend it, in addition to the twilight periods in which usuallypronounced peaks occur, variably into large amounts of the day and night hours (Figures7–9). Chronobiologists may tend to assign this activity pattern observed in nature to thecrepuscular chronotype rather than to any other, or to create a new category of activity.However, its introduction in primatology has been of great heuristic value in that it hasstimulated further field work on related species as well as an intense discussion onnocturnality and diurnality and their evolution in non-human primates (for review seeCurtis & Rasmussen 2006).
Nevertheless, due to methodological restrictions, many of these observational fieldstudies were of only limited value to chronobiology. For reasons of practicality, it isimpossible to conduct in nature the uninterrupted long-term behavioral observations thatare required to produce chronobiologically relevant datasets. Even so, the use ofactiwatches has proved to be a suitable tool for obtaining such long-term activityrecordings in wild, medium-sized and larger mammals (Erkert 2003). It is advisable,however, to carry out these recordings in collaboration with experienced field workers ofbehavioral ecology (cf. Kappeler & Erkert 2003; Fernandez-Duque & Erkert 2006), whohave access to suitable study sites and animal groups and know their species’ behavior andecology in detail. Such field workers are usually not fully familiar with the data-collectionand data-evaluation methods of chronobiology, nor with its relevant theories, specificapproaches and results.
The comparative studies carried out in captive diurnal and nocturnal primates undercontrolled laboratory conditions and in semi-natural environments, as well as in wild-living subjects in their natural habitat, show that all three approaches yield essential resultsthat complement and explain each other and contribute to a better understanding ofbehavioural, ecological, and evolutionary aspects of biological timing processes in nature,including underlying mechanisms. From the good correspondence between the resultsobtained in Aotus lemurinus kept under seminatural conditions and those obtained fromthe wild A. azarai of the Argentinean Chaco (cf. Figure 5, left and right panels), it may beinferred that recordings made under semi-natural conditions can also provide reliable dataon a species’ CAR in nature and its response to changing physical and bioticenvironmental factors. Furthermore these environments offer the opportunity for simpleexperimentation, such as to check the role of food abundance, quality or dispersal onactivity patterns as well as influences from other environmental factors.
Aotus
The most prominent feature of the owl monkeys’ activity rhythm is its pronounced lunarperiodicity, found in A. lemurinus under semi-natural lighting and climate conditions as
Biological Rhythm Research 257
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

well as in the wild A. azarai of the Argentinean Chaco (Erkert 1974, 1976; Fernandez-Duque 2003; Fernandez-Duque & Erkert 2006). Since indications of lunar periodicity inbehavioral activities have also been observed in other Aotus (sub)species (Wright 1978,1989), the strong dependence of the owl monkeys’ nocturnal activity on moonlight may beconsidered a genus-specific characteristic. Thus it seems justified to assume, further, thatthe laboratory finding that the pronounced activity inhibiting direct effect of low lightintensities (50.1 lux) on the activity level of A. lemurinus, which is thought to be mainlyresponsible for the lunar variation of these monkeys’ activity rhythm (Figure 5, Erkert1974; Erkert & Groeber 1986), would apply to the whole genus.
Despite the pronounced lunar periodicity under natural lighting conditions, inlaboratory studies no evidence of an endogenous lunadian or lunar, i.e. circalunadian orcircalunar component, has been found. At most, the apparently internally desynchronizedand split free-running CAR of the male A. lemurinus, shown in Figure 4 (right panel), mayintuitively be interpreted to indicate that an approximately lunadian component of 24.75 h(the lunar period is 24.8 h) might have been superimposed upon a main circadiancomponent of 23.75 h. However, since no other clues were found pointing to acircalunadian or circalunar component in the owl monkeys’ activity rhythm—either inthe free-running CAR in LL or in rhythms entrained by single light pulse, skeleton, orcomplete photoperiods—this still isolated result should not be over-interpreted. A circa-semilunar rhythm of peak total daily activity, with an average period of 14.0 + 2.3 d,shown to be present in the activity rhythms of five adult Aotus females while free-runningin LL 0.1 lux or entrained by skeleton photoperiods, has been interpreted as an effect ofoestrous (Rauth-Widmann et al. 1996) because its period corresponded approximately tothe species’ endocrinologically-determined ovarian cycle length of 15.5 + 0.6 d (Bonneyet al. 1980).
Recent neurobiological studies have shown that, in cases of behavioral rhythmsplitting, either the left and right SCN or ‘‘core-like’’ and ‘‘shell-like’’ neuron populationsof the SCN may oscillate circadianly and in antiphase to one another (de la Iglesia et al.2000; Yan et al. 2005). These antiphase rhythms have been associated with the morning(m) and evening (e) oscillators (Jagota et al. 2000; Daan et al. 2001; Yan et al. 2005),hypothesized long ago to underlie bimodal circadian activity patterns, rhythm splittingunder constant conditions, and photoperiodic time measurement (Pittendrigh & Daan1976). Taking this into account, from the pure phenomenology of the owl monkeys’ lunar-modulated activity rhythm under natural conditions, one might be tempted to speculatethat this conspicious phenomenon might also probably (co-)result from separate,decoupled oscillators of the bilaterally organized circadian pacemaker system in theSCN, these oscillators being differentially entrained by sunlight and moonlight. Such anassumption would also explain both the desynchonized split activity rhythm shown inFigure 4 (right panel) and the temporary extension of activity into the later morning hoursaround the new moon (cf. Figures 5, right, and 7, lower panel). However, the experimentalresults render this quite improbable (Erkert & Groeber 1986); the most reasonable andparsimonious hypothesis is still the assumption of a photically entrained circadian activityrhythm which shows lunadian masking due to strong inhibitory effects of low night-timeluminance. Significant falls in activity at the time of the full moon, observed during lunareclipses (Fernandez-Duque et al., in preparation), support this hypothesis.
Both the increased late morning activity around the newmoon and the irregular bouts ofdaytime activity observed throughout the year in theAotus azarai of the Argentinean Chaco(Fernandez-Duque 2003; Fernandez-Duque & Erkert 2006) might best be interpreted ascompensatory responses to detrimental nightly lighting, temperature and/or foraging
258 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

conditions, presumably ‘‘allowed’’ by an evolutionarily-acquired increased (day)lighttolerance in conjunction with a reduced predator pressure in those southernmost Aotushabitats (Wright 1989; Fernandez-Duque 2003; Fernandez-Duque & Erkert 2006).
Eulemur
The cathemeral wild red-fronted lemurs’ activity rhythm differed considerably from thatrecorded in E. fulvus albifrons under laboratory conditions. E. f. rufus showed mostactivity during the day/twilight period as well as a distinct lunar variation in nocturnalactivity throughout the year (Figure 7, Kappeler & Erkert 2003) while, depending onluminosity during the dark phase, E. f. albifrons changed from showing dark-active,through cathemeral, to light-active behavior (Figure 3). Since the circadian activityremained in phase with the dark, this phenotypical phase-reversal was interpreted as acompensatory response to the increasing inhibitory masking effects due to stepwisereductions of D-time luminosity (Erkert & Cramer 2006). The minor delays and advances,respectively, observed in the two peaks of a male’s and female’s Tb rhythms—while theiractivity rhythm was inverted after switching off D-lighting of 0.1 lux, and two weeks laterafter switching it on again—agree with this interpretation. But the results may also providesome indication of the underlying mechanism. Starting from Pittendrigh’s two-oscillatorconcept (Pittendrigh & Daan 1976) of coupled m and e oscillator systems phase-set todawn and dusk, respectively, the minor phase shifts of the two Tb-peaks may beinterpreted as due to opposite phase-shifts of m and e. Together with the characteristics ofdirect activity-inhibiting and/or activity-enhancing masking effects of low and high lightintensities, such opposite shifts of m and e oscillators and of the activity bouts coupled tothem may have resulted in the light activity observed in these lemurs. Figure 10 illustratesthis in a simple diagram.
Figure 10. Rough sketch illustrating how minor opposite phase shifts of putative morning (m) andevening (e) oscillator-coupled activity components would lead to a phase reversal from a more ‘‘darkactive’’ (nocturnal) to a more ‘‘light active’’ (diurnal) bimodal activity pattern (middle and lower rowof triangles, respectively). The black and white bar on top indicates the LD cycle to which the twooscillators are phase-set. Considering further the specific activity-inhibiting and activity-enhancing(disinhibiting) non-circadian direct effects of light, the observed luminosity during the dark phaseinduced an inversion of the lemurs’ activity pattern (Figure 2), in spite of there being a lack of phase-reversal in the underlying circadian rhythmicity.
Biological Rhythm Research 259
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Direct activity-inhibiting effects of low light intensities may also be assumed to beresponsible for the lunar activity component of the otherwise predominantly diurnalactivity pattern shown in nature in the subspecies E. f. rufus (Figures 7 and 8). In addition,their cathemeral behavior may also be interpreted to result from similar ‘‘negative maskingeffects’’ of low night-time luminances and/or from disinhibition of nocturnal activity bymoonlight. Though yet to be established, it is assumed that the lemurs’ CTS also indicatesbehavior of a genuinely nocturnal species, in that their ‘‘circadian activity phase’’ in natureis also phase-set to the dark fraction of the 24-h day, in spite of predominantly behaving asdiurnal animals.
A common feature of the three species’ activity rhythms in nature is a very pronouncedbimodal activity pattern (Figure 8). Considering that the study sites in Madagascar andSouth America were located at almost equal latitudes and that Argentinean official time isadvanced about one hour in relation to local time, the morning activity peak occurredearliest in nocturnal Aotus, later in the cathemeral Eulemur, and latest in the diurnalPropithecus—while the sequence of evening peak times was reversed, and daytime troughscoincided around noon (Figure 9B). Applying Pittendrigh’s two-oscillator model(Pittendrigh & Daan 1976) to these species activity patterns, one might hypothesize thatonly minor differences in the phase-setting of the m and e oscillators to dawn and duskmay ultimately be responsible for the differences in crepuscular peak times, and thus, inconjunction with the species’ characteristics of masking by light, also for their morenocturnal, cathemeral, or diurnal behavior in nature, in spite of a generally ‘‘nocturnal’’phase-setting characteristic of the CTS. An internally-coupled dual circadian oscillatorsystem with oscillators set to dawn and dusk might not only be a suitable tool for thetime measurement underlying photoperiodic regulation of annual events such asreproduction (which also occurs in lemurs, van Horn & Eaton 1979), migration andhibernation, but also for the induction of annual activity phase reversals as describedin various vertebrates including microtine rodents (Erkinaro 1969; Pittendrigh & Daan1976, Halle 1995). It might also provide the substrate for the species’ evolutionarily-acquired nocturnal (mainly), diurnal, crepuscular, or ‘‘cathemeral’’ behavior. In that case,the lemurs’ cathemerality, which seems closest related to a crepuscular lifestyle, mightindeed be considered rather to represent an original trait than being derived fromnocturnal or diurnal ancestors, as has recently been proposed by Curtis and Rasmussen(2006).
Evolutionary aspects
Without adequate answers to the yet unsettled question of how, i.e. by which centralnervous mechanism(s), diurnality may be produced or enabled in the various mammaliantaxa, chronobiology will hardly be able to contribute substantially to the problem of‘‘why’’ given species may have changed from nocturnality to diurnality and cathemeralityor vice versa. Thus the evolutionary background of chronotype differentiation, i.e thequestion of the ultimate factors that may have caused or allowed the various mammaliantaxa including primates to adopt and/or maintain a more or less strictly nocturnal,diurnal, crepuscular, or cathemeral lifestyle, cannot be cleared up by a purechronobiological approach. However, systematic extension of long-term activity record-ings in wild-living primates to certain closely related species and to other mammalian taxaof different chrono-ecotypes would provide both a better overview of chronotype diversityand clearer indications of the underlying evolutionary processes and their adaptive value.With regard to our data presented above, of special interest would be comparative studies
260 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

of some of the more nocturnal northern owl monkey species as well as of some otherMalagasy lemurids such as the nocturnal indriid Avahi, the cathemeral Hapalemur and thediurnal Lemur catta and Varecia, which have also been reported to show some night-timeactivity temporarily (Tattersall 1987; Curtis & Rasmussen 2006).
As indicated by the variability of the activity patterns in the wild owl monkeys andlemurs, or, more conspiciously, by the well-known examples of wild boars and roe deers(which, according to local hunting pressure, may be either diurnal or nocturnal),mammalian chrono-ecotypes are obviously not to be regarded as fixed and rigid clock-driven systems squeezed once and permanently into a certain temporal niche. Theyrepresent, rather, relatively flexible open systems whose behavioral activities are, by meansof their photically entrained CTS and the various direct effects of abiotic and bioticenvironmental factors, roughly phase-positioned (but, in relation to their needs and givenenvironmental conditions in their habitat, nevertheless, optimally so) within thecontinuum of accidental and regularly recurring, and more or less beneficial and/ordetrimental, external conditions and events. Of course, as recently shown by DeCoursey(2003, 2004) in SCN-lesioned fossorial sciurid rodents, a functioning circadian clock mightbe beneficial for an individual’s survival in nature, and should thus not be neglected inmammalian behavioral ecology. However, on account of the very small number of animalsinvolved in this study and of the possibility, therefore, of a non-representative outcome,further studies with sufficiently large numbers of test animals should be performed beforeone may generalize these results and ascribe to the mammalian CTS a really crucial rolefor the animals’, and hence also the species’, survival in nature. Besides their crucialfunction in coordinating internal timing processes, mammalian circadian clocks arecertainly also essential tools for the timing of annual events (such as reproduction,hibernation, or migration), for the timing of day-time related behavioral activities (such asonset and offset of activity and resting times, especially for cave dwelling and fossorialspecies) and for the time – place learning ability (necessary to avoid or visit particularplaces at a certain time of day, in order to reduce or rise the chance of predator, prey, rivalor mate encounter, or to re-visit certain nectar sources, offered from many flowers onlythroughout certain time windows during the day—e.g. for humming birds—or night—e.g.nectarivorous bats). Despite this, however, their importance for the development ormodification of a certain chrono-ecotype type behavior should not be overestimated.
Driving forces and limiting factors for the transition to another temporal lifestyle, suchas that from nocturnality or crepuscularity to cathemerality or diurnality (as shown inextant Malagasy lemuriform primates and other mammalian taxa) or vice versa (as hasbeen assumed in Aotus, Wright (1989), Martin (1990)) have to be looked for primarilyamong relevant ecological and fitness factors, like food availability and competition,predation pressure, general and extreme climate conditions, metabolic, thermoregulatory,sensory, and motor capacities, etc. As compared to these factors, circadian clockcharacteristics and mechanisms seem of secondary significance. Speculations about theevolution of a certain chrono-ecotype observed in nature (Horton 2001) will have toconsider that this usually requires a whole bundle of co-evolutionary changes, some ofwhich may really be essential while others may be only permissive for the adoptedtemporal lifestyle, or merely tolerated by it. Finally, the controversial discussion inprimatology as to which one of the various proposed factors might have caused lemurs tobecome cathemeral (for review see Curtis & Rasmussen 2006) may be pointless, becauseeach of the facors may have contributed to a greater or lesser extent. Benefits andconstraints associated with chrono-ecotype transition will have to be analyzed separatelyin each species.
Biological Rhythm Research 261
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Acknowledgements
I am very grateful to my colleagues Drs. Peter Kappeler (German Primate Center), EduardoFernandez-Duque (Fundacion Eco, Argentina), Jairo Munoz-Delgado (Instituto Mexicano dePsiquiatrıa, Mexico), and Domingo Canales (Universidad Veracruzana, Mexico) for quite a pleasantand highly productive cooperation in carrying out long-term activity recordings in wild lemurs andowl monkeys, as well as in spider monkeys kept under seminatural conditions. Many thanks also toAngelika Scheideler (Maximilian University, Munich) for her help with the statistics and diagramproduction. Furthermore, I would like to thank Drs. Jim Waterhouse and Roberto Refinetti for theirinvitation to contribute to this special issue of Biological Rhythm Research and for kindly reviewingand improving the manuscript. Last but not least, I’m very much obliged to all my students for theirhighly appreciated contributions to our comparative approach in primate chronobiology. All studieswere supported by grants from the Deutsche Forschungsgemeinschaft.
References
Albers HE, Lydic R, Gander PH. 1982. Gradual decay of circdadian drinking organization
following lesions of the suprachiasmatic nuclei in primates. Neuros Lett. 27:119–124.Albers HE, Lydic R, Moore-Ede MC. 1984a. Role of the suprachiasmatic nuclei in the circadian
timing system of the squirrel monkey. I. The generation of rhythmicity. Brain Res. 300:275–
284.Albers HE, Lydic R, Moore-Ede MC. 1984b. Role of the suprachiasmatic nuclei in the circadian
timing system of the squirrel monkey. II. Light-dark cycle entrainment. Brain Res. 300:285–293.
Arditi SI. 1992. Variaciones estacionales en la actividad y dieta de Aotus azarai y Alouatta caraya enFormosa, Argentina. Boletin Primatologico Latinoamericano. 3:11–30.
Aschoff J. 1964. Die Tagesperiodik licht- und dunkelaktiver Tiere. Rev Susse Zool. 71:528–558.
Aschoff J. 1979. Circadian rhythms:influences of internal and external factors on the periodmeasured in constant conditions. Z Tierpsychol. 49:225–249.
Aschoff J, Daan S, Honma K-I. 1982. Vertebrate circadian systems – structure and physiology. New
York: Springer-Verlag, Heidelberg. Zeitgebers, entrainment, and masking:some unsettledquestions. p. 13–24.
Aschoff J, Tokura H. 1986. Circadian activity rhythms in squirrel monkeys:entrainment bytemperature cycles. J Biol Rhythms. 2:91–99.
Aschoff J, von Goetz C. 1986. Effects of feeding cycles on circadian rhythms in squirrel monkeys. JBiol Rhythms. 1:267–276.
Bonney RC, Dixson AF, Fleming D. 1980. Plasma concentration of oestradiol-17b, oestrone,
progesterone and testosterone during ovarian cycle of the owl monkey (Aotus trivirgatus). JReprod Fert. 60:101–107.
Boulos Z, Frim DM, Dewey LK, Moore-Ede MC. 1989. Effects of feeding schedules on circadian
organization in squirrel monkeys. Physiol Behav. 45:507–515.Boulos Z, Machi M, Terman M. 1996. Effects of twilights on circadian entrainment patterns and
reentrainment rates in squirrel monkeys. J Comp Physiol A. 179:687–694.
Caldelas I, Poirel V-J, Sicard B, Pevet P, Challet E. 2003. Circadian profile and photic regulation ofclock genes in the suprachiasmatic nucleus of a diurnal mammal (Arvicanthus ansorgei).Neuroscience 116:583–591.
Charles-Dominique P. 1975. Phylogeny of primates. New York: Plenum Press. p. 69–90.
Nocturnality and diurnality. An ecological interptetation of these two modes of life by ananalysis of the higher vertebrate fauna in tropical forest ecosystems.
Colguhoun IC. 1998. Cathemeral behaviour of Eulemur macaco macaco at Ambato Massif,
Madagascar. Folia Primatol. 69:22–34.Costa MSMO, Moreira LF, Alones V, Lu J, Santee UR, Cavalcante JS, Moraes PRA, Britto LRG,
Menaker M. 1998. Characterization of the circadian system of monkey (Callithrix jacchus):Im-
munohistochemical analysis and retinal projections. Biol Rhytm Res. 29:510–520.
262 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Crompton AW, Taylor CR, Jagger JA. 1978. Evolution of homeothermy in mammals. Nature.
272:333–336.Curtis DJ, Zaramody A, Martin RD. 1999. Cathemerality in the mongoose lemur, Eulemur mongoz.
Am J Primatol. 47:279–298.
Curtis DJ, Rasmussen MA. 2006. The evolution of cathemerality in primates and other mammals:acomparative and chronoecological approach. Folia Primatol. 77:178–193.
Curtis DJ, Zaramody A, Martin D. 1999. Cathemerality in the mongoose lemur, Eulemur mongoz.Am J Primatol. 47:279–298.
Daan S. 2000. Colin Pittendrigh, Jurgen Aschoff, and the natural entrainment of circadian systems. JBiol Rhythms. 15:195–207.
Daan S, Albrecht U, van der Horst GT, Illnerova H, Roenneberg T, Wehr TA, Schwartz WJ. 2001.
Assembling a clock for all seasons:are there M and E oscillators in the genes? J Biol Rhythms.16:105–116.
DeCoursey PJ. 2003. Chronobiology: Biological timekeeping. Sunderland (MA): Sinauer Associates,
Inc.. p. 144–178. The behavioral ecology and evolution of biological timing systems.DeCoursey PJ. 2004. Diversity of function of SCN pacemakers in behavior and ecology of three
species of sciurid rodents. Biol Rhythm Res. 35:13–33.DeCoursey PJ, DeCoursey G. 1964. Adaptive aspects of activity rhythms in bats. Biol Bull. 126:14–
27.De la Iglesia HO, Meyer J, Carpino A, Jr., Schwartz WJ. 2000. Antiphase oscillation of the left and
right suprachiasmatic nuclei. Science. 290:799–801.
Doerrscheidt GJ, Beck L. 1975. Advanced method for evaluating characteristic parameters (tau,alpha, rho) of circadian rhythms. J Math Biol. 2:107–121.
Donati G, Borgognini-Tarli SM. 2006. Influence of abiotic factors on cathemeral activity:
a case of Eulemur fulvus collaris in the littoral forest of Madagascar. Folia Primatol. 77:104–122.
Donati G, Lunardini A, Kappeler PM, Borgonini Tarli S. 2001. Nocturnal activity in the cathemeral
red-fronted lemur (Eulemur fulvus rufus), with observations during a lunar eclipse. Am JPrimatol. 53:69–78.
Edgar DM, Dement WC, Fuller CA. 1993. Effects of SCN lesions on sleep in squirrelmonkeys:Evidence for opponent processes in sleep-wake regulation. J Neurosci. 13:1065–1079.
Erkert HG. 1974. Der Einfluss des Mondlichtes auf die Aktivitaetsperiodik nachtaktiver Saeugetiere.Oecologia. 14:269–287.
Erkert HG. 1976a. Lunarperiodic variation of the phase-angle difference in nocturnal animals under
natural Zeitgeber conditions near the equator. Int J Chronobiol. 4:125–138.Erkert HG. 1976b. Beleuchtungsabhaengiges Aktivitaetsoptimum bei Nachtaffen (Aotus trivirgatus).
Folia Primatol. 25:186–192.
Erkert HG. 1982. Ecology of bats. New York, London: Plenum Press. p. 201–242. Ecological aspectsof bat activity rhythms.
Erkert HG. 1989. Characteristics of the circadian activity rhythm in common marmosets (Callithrix
j. jacchus). Am J Primatol. 17:271–286.Erkert HG. 1991. Primatology today. Amsterdam, New York, Oxford: Elsevier Science Publishers.
p. 435–438. Influence of ambient temperature on circadian rhythms in Colombian owl monkeys,Aotus lemurinus griseimembra.
Erkert HG. 2003. Field and laboratory methods in primatology. Cambridge: Cambridge UniversityPress. p. 252–270. Chronobiological aspects of primate research.
Erkert HG, Cramer B. 2006. Chronobiological background to cathemerality:circadian rhythms in
Eulemur fulvus albifrons (Prosimii) and Aotus azarai boliviensis (Anthropoidea). Folia Primatol.77:87–103.
Erkert HG, Gburek V, Scheideler A. 2006. Photic entrainment and masking of prosimian circadian
rhythms (Otolemur garnettii, Primates). Physiol Behav. 88:39–46.Erkert HG, Groeber J. 1986. Direct modulation of activity and body temperature of owl monkeys
(Aotus lemurinus griseimembra) by low light intensities. Folia Primatol. 47:171–188.
Biological Rhythm Research 263
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Erkert HG, Kappeler PM. 2004. Arrived in the light:diel and seasonal activity patterns in wild
Verreaux’s sifakas (Propithecus v. verreauxi, Primates:Indriidae). Behav Ecol Sociobiol. 57:174–186.
Erkert HG, Schardt U. 1991. Social entrainment of circadian activity rhythms in common
marmosets, Callithrix j. jacchus (Primates). Ethology. 87:189–202.Erkert HG, Thiemann A. 1983. Dark switch in the entrainment of circadian activity rhythms in night
monkeys, Aotus trivirgatus HUMBOLDT. Comp Biochem Physiol. 74A:307–310.Erkinaro E. 1969. Der Phasenwechsel der lokomotorischen Aktivitaet bei Microtus agrestis (L.), M.
arvalis (Pall.) und M. oeconomicus (Pall.). Aquilo Ser Zool. 13:1–31.Farrer DN, Ternes JW. 1969. Circadian rhythms in nonhuman primates. Basel: S. Karger. p. 1–7.
Bibl Primat 9, Illumination intensity and behavioural circadian rhythms.
Fernandez-Duque EF. 2003. Influences of moonlight, ambient temperature and food availablility onthe diurnal and nocturnal activity of owl monkeys (Aotus azarai). Behav Ecol Sociobiol. 54:431–440.
Fernandez-Duque EF, Erkert HG. 2006. Cathemerality and lunar periodicity of activity rhythms inowl monkeys of the Argentinean chaco. Folia Primatol. 77:123–138.
Ferraro JE, Sulzman FM. 1988. The effects of feedback lighting on the circadian drinking rhythm inthe diurnal new world primate Saimiri sciureus. Am J Primatol. 15:143–155.
Fleagle JG. 1988. Primate adaptation and evolution. San Diego, New York: Academic Press, Inc.. p.474.
Fuller CA, Edgar DM. 1986. Effects of light intensity on the circadian temperature and feeding
rhythms in the squirrel monkey. Physiol Behav. 36:687–691.Fuller CA, Lydic R, Sulzman FM, Albers HE, Tepper B, Moore-Ede MC. 1981. Circadian rhythm
of body temperature persists after suprachiasmatic lesions in the squirrel monkey. Am J Physiol.
241:R385–R391.Gander PH, Moore-Ede MC. 1983. Light-dark masking of circadian temperature and activity
rhythm in squirrel monkeys. Am J Physiol. 245:R927–R934.
Garcia JE, Braza. 1987. Activity rhythms and use of space of a group of Aotus azarae in Boliviaduring the rainy season. Primates. 28:337–342.
Glass JD, Tardiff SD, Clements R, Mrosovsky N. 2001. Photic and nonphotic circadian phaseresetting in a diurnal primate, the common marmoset. Am J Physiol. 280:R191–R197.
Haerter L, Erkert HG. 1993. Alteration of circadian period length does not influence the ovariancycle length in common marmosets, Callithrix j. jacchus (Primates). Chronobiol Int. 10:165–175.
Haeussler U, Erkert HG. 1978. Different direct effects of light intensity on the entrained activityrhythm in neotropical bats (Chiroptera, Phyllostomidae). Behav Proc. 3:223–239.
Halle S. 1995. Diel patterns of locomotor activity in populations of root voles, Microtus
oeconomicus. J Biol Rhythms. 10:211–224.Halle S. 2006. Polyphasic activity patterns in small mammals. Folia Primatol. 77:15–26.Hawking F, Lobban MC. 1970. Circadian rhythms in macaca monkeys (physical activity,
temperature, urine and microfilarial levels). J Interdisc Cycle Res. 1:267–290.Hoban TM, Levine AH, Shane RB, Sulzman FM. 1985. Circadian rhythms of drinking and body
temperature rhythms in night monkeys, Aotus trivirgatus HUMBOLDT. Comp BiochemPhysiol. 74A:307–310.
Hoban TM, Sulzman FM. 1985. Light effects on the circadian timing system of a diurnal primate,the squirrel monkey. Am J Physiol. 249:R274–280.
Horton TH. 2001. Handbook of Neurobiology 12:Circadian clocks. New York: Kluwer Academic
Press/Plenum Publishers. p. 45–57. Conceptual issues in the ecology and evolution of circadianrhythms.
Hut RA, van Oort BEH, Daan S. 1999a. Natural entrainment without dawn and dusk: The case of
the European ground squirrel. J Biol Rhythms. 14:290–299.Hut RA, Mrosovsky N, Daan S. 1999b. Nonphotic entrainment in a diurnal mammal, the European
ground squirrel (Spermophilus citellus). J Biol Rhythms. 14:409–419.
264 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Jagota A, de la Iglesia HO, Schwartz WJ. 2000. Morning and evening circadian oscillations in the
suprachiasmatic nucleus in vitro. Nat Neurosci. 3:372–376.Kappeler PM, Erkert HG. 2003. On the move around the clock:correlates and determinants of
cathemeral activity in wild redfronted lemurs (Eulemur fulvus fulvus). Behav Ecol Sociobiol.
54:359–369.Kas MJH, Edgar DM. 2000. Photic phase response curve in Octodon degus:assessment as a function
of activity phase preference. Am J Physiol. 278:R1385–R1389.King DP, Takahashi JS. 2000. Molecular genetics of circadian rhythms in mammals. Ann Rev
Neurosci. 23:713–742.Kirk EC. 2006. Eye morphology in cathemeral lemurids and other mammals. Folia Primatol. 77:27–
49.
Klein DC, Moore RY, Reppert SM. editors. 1991. Suprachiasmatic nucleus. The mind’s clock. NewYork (Oxford): Oxford University Press. p. 467.
Lee HS, Billings HJ, Lehman MN. 2003. The suprachiasmatic nucleus:a clock of multiple
components. J Biol Rhythms. 18:435–449.Lee TM. 2004. Growing evidence that some aspects of SCN function differ in nocturnal and diurnal
rodents. Am J Phsiol Regul Integr Comp Physiol. 286:R814–R815.Lemos DR, Downs JL, Urbanski HF. 2006. Twenty-four-hour rhythmic gene expression in the
rhesus macaque adrenal gland. Mol Endocrinol. 20:1164–1176.Lydic R, Albers HE, Tepper B, Moore-Ede MC. 1981. Three-dimensional structure of mammalian
suprachiasmatic nuclei:a comparative study of five species. J Comp Neurol. 204:225–237.
Marques MD, Waterhouse J. 2004. Rhythms and Ecology – do chronobiologists still remembernature? Biol Rhythm Res. 35:1–2.
Martin RD. 1990. Primate origins and evolution. London: Chapman and Hall.
Martinez JL. 1972. Effects of selected illumination levels on circadian periodicity in the rhesusmonkey (Macaca mulatta). J Interdisc Cycle Res. 3:47–59.
Meijer JH, Schwartz WJ. 2003. In search of the pathways for light-induced pacemaker resetting the
suprachiasmatic nucleus. J Biol Rhythms. 18:235–249.Minors D, Waterhouse J, Rietveld W. 1996. Constant routine and ‘‘purification’’ methods:do they
measure the same thing? Biol Rhythm Res. 27:166–174.Moore RY. 1993. Organization of the primate circadian system. J Biol Rhythms. 8(Suppl):3–9.
Moore RY, Leak RK. 2001. Suprachiasmatic nucleus. In: Takahashi JS, Turek FW, Moore RY,editors. Handbook of behavioral neurobiology, Vol. 12:Circadian clocks. New York: KluwerAcademic/Plenum Publishers. p. 141–179.
Moore-Ede MC, Sulzman FM. 1977. The physiological basis of circadian timekeeping in primates.Physiologist. 20:17–25.
Moore-Ede MC, Kass DA, Herd JA. 1977. Transient circadian internal desynchronization after
light-dark shifts in monkeys. Am J Physiol. 232:R31–37.Moore-Ede MC, Sulzman FM, Fuller. 1982. The clock that time us. Physiology of the circadian
system. Cambridge Mass and London: Harvard University Press. p. 448.
Morgan E. 2004. Ecological significance of biological clocks. Biol Rhythm Res. 2004:3–12.Moynihan M. 1964. Some behavior patterns of platyrrhine monkeys. I. The night monkey (Aotus
trivirgatus). Smithsonian Misc Coll. 146:1–84.Mrosovsky N. 1994. In prais of masking:behavioral responses of retinally degenerate mice to dim
light. Chronobiol Int. 11:343–348.Mrosovsky N. 1999. Masking:History, definitions, and measurement. Chronobiol Int. 16:415–429.Mrosovsky N. 2003. Beyond the suprachiasmatic nucleus. Chronobiol Int. 20:1–8.
Munoz M, Peirson SN, Hankins MW, Foster RG. 2005. Long-term constant light inducesconstitutive elevated expression of mPER2 protein in the murine SCN:A molecular basis forAschoff’s rule? J Biol Rhythms. 20:3–14.
Munoz-Delgado J, Corsi-Cabrera M, Canales-Espinoza D, Santillan-Doherty AM, Erkert HG.2004. Astronomicl and meteorological parameters and rest-activity rhythm in the spider monkeyAteles geoffroyi. Physiol Behav. 83:107–117.
Biological Rhythm Research 265
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Munoz-Delgado J, Fuentes-Pardo B, Euler Baum A, Lanzagorta N, Arenas-Rosas R, Santillan-
Doherty AM, Guevara MA, Corsi-Cabrera M. 2005. Presence of a circadian rhythm in thespider monkey’s (Ateles geoffroyi) motor activity. Biol Rhythm Res. 36:115–121.
Murakami DM, Fuller CA. 1990. The retinohypothalamic projection and oxidative metabolism in
the suprachiasmatic nucleus of primates and tree shrews. Brain Behav Evol. 35:302–312.Palkova M, Sigmund L, Erkert HG. 1999. Effect of ambient temperature on the circadian
activity rhythm in common marmosets, Callithrix j. jacchus (Primates). Chronobiol Int. 16:149–161.
Pittendrigh CS, Daan S. 1976. A functional analysis of circadian pacemakers in nocturnal rodents.V. Pacemaker structure:a clock for all seasons. J Comp Physiol. 106:333–355.
Rappold I, Erkert HG. 1994. Re-entrainment, phase-response and range of entrainment of
circadian rhythms in owl monkeys (Aotus lemurinus g.) of different age. Biol Rhythm Res.25:133–152.
Rauth-Widmann B, Fuchs E, Erkert HG. 1996. Infradian alteration of circadian rhythms in owl
monkeys (Aotus lemurinus griseimembra):An effect of estrous? Physiol Behav. 59:11–18.Rauth-Widmann B, Thiemann-Jager A, Erkert HG. 1991. Significance of nonparametric light effects
in entrainment of circadian rhythms in owl monkeys (Aotus lemurinus griseimembra) by light-dark cycles. Chronobiol Int. 8:251–266.
Redlin U, Mrosovsky N. 2004. Nocturnal activity in a diurnal rodent (Arvicanthis niloticus):Theimportance of masking. J Biol Rhythms. 19:58–67.
Refinetti R. 2004. Parameters of photic resetting of the circadian system of a diurnal rodent, the nile
grass rat. Acta Sci Vet. 32:1–7.Refinetti R. 2006. Variability of diurnality in laboratory rodents. J Comp Physiol A. 192:701–
714.
Reebs SG, Mrosovsky N. 1989. Effects of induced wheel running on the circadian activity rhythms ofSyrian hamsters:entrainment and phase response curve. J Biol Rhythms. 4:39–48.
Rensing L. 1989. Is ‘‘masking’’ an appropriate term? Chronobiol Int. 6:297–300.
Reppert SM, Perlow MJ, Ungerleider LG, Mishkin M, Tamarkin L, Orloff DG, Hoffman HJ, KleinDC. 1981. Effects of damage to the suprachiasmatic area of the anterior hypothalamus on thedaily melatonin and cortisol rhythms in the rhesus monkey. J Neurosci. 1:1414–1425.
Reppert S, Schwartz W, Uhl G. 1987. Arginine vasopressin:A novel peptide rhythm in cerebrospinal
fluid. Trends Neurosci. 10:76–80.Robinson EL, Fuller CA. 1999. Light masking of circadian rhythms of heat production, heat loss,
and body temperature in squirrel monkeys. Am J Physiol. 276:R298–R307.
Rotundo M, Sloan C, Fernandez-Duque E. 2000. Cambios estacionales en el ritmo de actividad delmono mirikina (Aotus azarai) en Formosa Argentina. In: Cabrera E, Mercolli C, Resquin R(eds.) Manejo de fauna silvestre en Amazonia y Latinoamerica, pp. 413–417. Asuncion,
Fundacion Moises Bertoni.Schanz F, Erkert HG. 1987. Resynchronisationsverhalten der Aktivitaetsperiodik von Galagos
(Galago senegalensis, Galago crassicaudatus garnettii). Z Saeugetierk. 52:218–226.
Schardt U, Wilhelm I, Erkert HG. 1989. Splitting of the circadian activity rhythm in commonmarmosets (Callithrix j.jacchus; Primates). Experientia. 45:1112–1115.
Schilling A, Richard J-P, Serviere J. 1999. Duration of activity and period of circadian activity-restrhythm in a photoperiod-dependent primate, Microcebus murinus. CR Acad Sci Paris, Life Sci.
322:759–770.Schwartz MD, Nunez AA, Smale L. 2004. Differences in the suprachisamatic nucleus and lower
subparaventricular zone of diurnal and nocturnal rodents. Neusoscience. 127:13–23.
Schwartz WJ, de la Iglesia HO, Zlomanczuk P, Illnerova H. 2001. Encoding le quattro stagioniwithin the mammalian brain: Photoperiodic orchestration through the suprachiasmatic nucleus.J Biol Rhythms. 16:302–311.
Shearman LP, Siriam S, Weaver DR, Maywood ES, Chavez I, Zheng B, Kume K, Lee CC, van derHorst GTJ, Hastings MH, Reppert SM. 2000. Interacting molecular loops in the mammaliancircadian clock. Science. 288:1013–1019.
266 H.G. Erkert
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011

Smale L, Lee T, Nunez AA. 2003. Mammalian diurnality:some facts and gaps. J Biol Rhythms.
18:356–366.Starck D. 1978. Vergleichende Anatomie der Wirbeltiere auf evolutionsbiologischer Grundlage. Vol.
1:Theoretische Grundlagen, Stammesgeschichte und Systematik unter Beruecksichtigung der
niederen Chordata. Berlin, Heidelberg, New York: Springer-Verlag. p. 274.Sulzman FM, Fuller CA, Moore-Ede MC. 1977a. Feeding time synchronizes primate circadian
rhythms. Physiol Behav. 18:775–779.Sulzman FM, Fuller CA, Moore-Ede MC. 1977b. Spontaneous internal desynchronization of
circadian rhythms in the squirrel monkey. Comp Biochem Physiol. 58A:63–67.Sulzman FM, Fuller FA, Moore-Ede MC. 1978. Comparison of synchronization of primate
circadian rhythms by light and food. Am J Physiol. 234:R130–R135.
Sulzman FM, Fuller CA, Moore-Ede. 1979. Tonic effects of light on the circadian system of thesquirrel monkey. J Comp Physiol. 129:43–50.
Tattersall I. 1987. Cathemeral activity in primates:a definition. Folia Primatol. 49:200–202.
Tattersall I. 1979. Patterns of activity in the Mayotte lemur. J Mammal. 60:314–323.Tattersall I. 2006. The concept of cathemerality: history and definition. Folia Primatol. 77:7–14.Thiemann-Jaeger A. 1986. Charakteristika der circadianen Aktivitaetsperiodik von Nachtaffen (Aotus
trivirgatus, HUMBOLDT 1811). Germany: University of Tuebingen. p. 79. Doctoral thesis.
Tokura H, Aschoff J. 1978. Circadian activity rhythms of the pig-tailed macaque, Macacanemestrina, under constant conditions. Pfluegers Archiv. 376:241–243.
Tokura H, Aschoff J. 1983. Effects of temperature on the circadian rhythm of pig-tailed macaques,
Macaca nemestrina. Am J Physiol. 245:R800–804.Van Horn RN, Eaton GG. 1979. The Study of prosimian behaviour. New York: Academic Press. p.
79–122. Reproductive physiology and behavior in prosimians.
Van Schaik CP, Kappeler PM. 1996. The social systems of gregarious lemurs:lack of convergencewith anthropoids due to evolutionary disequilibrium? Ethology. 102:915–941.
Weaver DR. 1998. The suprachiasmatic nucleus:a 25-year retrospective. J Biol Rhythms. 13:100–
112.Wechselberger E. 1994. Current Primatology Vol. III. Strasbourg: Universite Louis Pasteur. p. 223–
226. Phase-shifting effects of arousal on circadian rhythms in Callithrix j. jacchus.Wechselberger E. 1995. Charakteristik und Mechanismen der Synchronisation der Circadianper-
iodik des Weissbuesschelaeffchens, Callithrix j. jacchus. Germany: University of Tuebingen. p.113. Doctoral thesis.
Wechselberger E, Erkert HG. 1994. Characteristics of the light-induced phase response of circadian
activity rhythms in common marmosets, Callithrix j. jacchus [Primates-Cebidae]. Chronobiol Int.11:275–284.
Wexler DB, Moore-Ede MC. 1985. Circadian sleep-wake cycle organization in squirrel monkeys.
Am J Physiol. 248:R353–R362.Wexler DB, Moore-Ede MC. 1986. Resynchronization of circadian sleep-wake and temperature
cycles in the squirrel monkey following phase shifts of the environmental light-dark cycle. Aviat
Space Environ Med. 57:1144–1149.Wright PC. 1978. Home range, activity patterns, and agonistic encounters of a group of night
monkeys (Aotus trivirgatus) in Peru. Folia Primatol. 29:43–55.Wright PC. 1989. The nocturnal primate niche in the New World. J Hum Evol. 18:635–658.
Yan L, Foley NC, Bobula JM, Kriegsfeld LJ, Silver R. 2005. Two antiphase oscillationsoccur in each suprachiasmatic nucleus of behaviorally split hamsters. J Neurosci. 28:9017–9026.
Yellin AM, Hauty GT. 1971. Activity cycles of the rhesus monkey (Macaca mulatta) under severalexperimental conditions both in isolation and in a group situation. J Interdiscipl Cycle Res.2:475–490.
Zordan M, Costa R, Macino G, Fukuhara C, Tosini G. 2000. Circadian clocks: what makes themtick? Chronobiol Int. 17:433–451.
Biological Rhythm Research 267
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
18:
06 1
0 N
ovem
ber
2011