distinct contribution of the striatum and cerebellum...
TRANSCRIPT

Brain and Cognition 45, 189–211 (2001)doi:10.1006/brcg.2000.1237, available online at http://www.idealibrary.com on
Distinct Contribution of the Striatum andCerebellum to Motor Learning
Robert Laforce, Jr.* and Julien Doyon†
*University of New Brunswick, Saint John, New Brunswick, Canada; and †Rehabilitation ResearchGroup, Centre Francois-Charon, Quebec City, Quebec, Canada
Published online August 3, 2000
The striatum and cerebellum have been shown to be key structures of a distributed systemfor the control of skilled movements. However, the mechanisms under which they operateremain unclear. This study compared the performance of patients with Parkinson’s disease(PD) or with cerebellar damage (CE) to that of age-matched controls. Each group performedtwo visuomotor paradigms: a random variant of the serial reaction time (SRT) task that testedthe subject’s ability to make efficient stimulus–response associations and an adapted versionof the mirror-tracing task that measured their capacity to combine simple movements intocomplex ones. PD patients with bilateral striatal damage showed an impaired learning profileon the SRT task and a normal facilitation effect in the tracing task, while CE patients showedthe reverse pattern. Although further research is needed, the present findings suggest that thestriatum and cerebellum are involved in distinct learning mechanisms. 2001 Academic Press
INTRODUCTION
Over a century of observations have indicated that lesions to the striatum andcerebellum impair cognitive and motor functions in very different ways (see Mars-den & Obeso, 1994; Schmahmann, 1996; Thach, Mink, Goodkin, & Keating, 2000,for reviews). In the motor domain, for example, authors have often noted that basalganglionic deficits are seen during attempts to relax and rest, whereas cerebellar dys-functions are only seen during movements. Not surprisingly, such a distinct profileof cognitive and motor deficits has been supported by neuroanatomical findings whichhave shown that the cortico-basal ganglia-thalamo-cortical and the cortico-cerebello-thalamo-cortical loops constitute two separate neural systems (Asunama, Thach, &Jones, 1983; Middleton & Strick, 1994; Percheron, Francois, Yelnik, Fenelon, &Talbi, 1993; Sakai, Inase, & Tanji, 1996; Yamamoto, Yoshida, Yoshikawa, Kishi-
This work served as partial fulfillment of the requirements for a doctoral degree at the Departmentof Psychology, Laval University, Quebec City, Quebec, Canada. It was supported in part by a scholarshipfrom ‘‘La Fondation de l’Universite Laval’’ to Robert Laforce, Jr. and by a grant from the NaturalSciences and Engineering Research Council of Canada (OPGIN-012) to Dr. Julien Doyon. We are grate-ful to the patients and control subjects who participated in the present study, as well as to Dr. Paul J.Bedard for the opportunity to study his patients and for providing detailed descriptions of the patients’diagnoses. Also, we address special thanks to Joanne Roy for her help in various aspects of this research,Rhonda Amsel for statistical advice, and to all the members of the Groupe de recherche en readaptationphysique, IRDPQ, site Francois-Charon, Quebec City, Quebec, Canada.
Address correspondence and reprint requests to Robert Laforce, Jr., Assistant Professor, Departmentof Psychology, University of New Brunswick, P.O. Box 5050, Saint John (N.B.), Canada E2L 4L5. Fax:(506) 648-5780. E-mail: [email protected].
1890278-2626/01 $35.00
Copyright 2001 by Academic PressAll rights of reproduction in any form reserved.

190 LAFORCE AND DOYON
moto, & Oka, 1992). They have also been corroborated by imaging studies whichhave shown a distinct involvement of these structures in the control of movement(see Jueptner & Weiller, 1998, for a review).
In the motor learning literature, several investigations in both animals and humanshave supported the participation of the striatum and cerebellum in the acquisition ofskilled motor behaviors through practice (see Bloedel & Bracha, 1997; Doyon, 1997;Graybiel & Kimura, 1995; Leiner, Leiner, & Dow, 1995; Moscovitch, Vriezen, &Goshen-Gottstein, 1993; Thach, 1996; for reviews). In humans, pathological degener-ative processes affecting the striatum [as in Parkinson’s (PD) or Huntington’s (HD)diseases], or circumscribed damage to the cerebellum, have been shown to producean impairment on various skill-learning tasks, especially in the visuomotor modality(e.g., Doyon, Gaudreau, Laforce, Castonguay, Bedard, Bedard, & Bouchard, 1997a;Ferraro, Balota, & Connor, 1993; Harrington, York Haaland, Yeo, & Marder, 1990;Heindel, Salmon, Shults, Walicke, & Butters, 1989; Saint-Cyr, Taylor, & Lang, 1988;Sanes, Dimitrov, & Hallett, 1990). These findings have been corroborated by studieswith healthy control subjects using modern brain-imaging techniques such as positronemission tomography (PET) and functional magnetic resonance imaging (fMRI) inwhich hemodynamic changes have been observed in the striatum and/or the cerebel-lum during the incremental acquisition of visuomotor skills (e.g., Doyon, Owen, Pe-trides, Sziklas, & Evans, 1996b; Flament, Ellermann, Kim, Ugurbil, & Ebner, 1996;Grafton, Woods, & Mike, 1994; Jenkins, Brooks, Nixon, Frackowiak, & Passingham,1994; Rauch et al., 1997).
Interestingly, recent neurophysiological studies have shown that cells in both thestriatum and the cerebellum differ with respect to their intrinsic learning properties(e.g., Graybiel & Kimura, 1995; Hikosaka, Rand, Miyachi, & Miyashita, 1995; Ito,1982, 1993; Schultz, Romo, Ljungberg, Mirenowicz, Hollerman, & Dickinson,1995). Behavioral investigations which have compared patients with striatal and cere-bellar damage have identified a different pattern of learning impairment in these pa-tients (Pascual-Leone, Grafman, Clark, Stewart, Massaquoi, Lou, & Hallett, 1993;Sanes et al., 1990). A number of recent theoretical models have emerged in whicha critical role for the incremental acquisition of skills has been allocated to the ba-sal ganglia, the cerebellum, or both (Burton, 1990; Houk & Wise, 1995; Jueptner,Jenkins, Brooks, Frackowiak, & Passingham, 1996; Jueptner & Weiller, 1998; Thach,1996; Willingham, 1998; Wise & Houk, 1994). Altogether, these efforts indicate thatalthough both structures participate in motor learning, their contribution is distinct.Neuroscientists now face the challenge of identifying the mechanisms under whichthe striatum and cerebellum operate. This research is an effort in that direction.
The goal of this study was to identify the preferential learning mechanisms underwhich the striatal and the cerebellar systems operate when individuals acquire visuo-motor skills. We tested the hypothesis that the striatum would be involved in theelaboration of perceptual-motor programs based on stimulus–response (S-R) types ofassociations, while the cerebellum would play a preponderant role in linking togethersimple movements into more complex compound movements (Bloedel, 1992; Inhoff,Diener, Rafal, & Ivry, 1989; Inhoff & Rafal, 1990; Thach, Goodkin, & Keating,1992).
Evidence supporting the role of the striatum in perceptual-motor learning comesfrom physiological and behavioral studies in animals (see Graybiel & Kimura, 1995;Marsden & Obeso, 1994; see White, 1989, 1997, for reviews). For example, Graybieland Kimura (1995) have demonstrated the existence of TANs which undergo electro-physiological changes in responsiveness as nonhuman primates learn to associate aresponse to the presentation of a conditioning stimulus. Using a ‘‘win-stay’’ versionof the eight-arm radial maze, White and his collaborators have also shown that the

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 191
dorsal striatum mediates the generation of reinforced S-R associations (McDonald &White, 1993; Packard, Hirsh, & White, 1989; Packard & White, 1990; see White,1997, for a review). Further evidence that the striatum contributes to this type oflearning comes from two studies in humans (Knowlton, Mangels, & Squire, 1996;Singh, Metz, Gabrieli, Willingham, Dooley, Jiang, Chen, & Cooper, 1993). First,Knowlton et al. (1996) showed that patients with PD failed to learn a probabilisticclassification task in which they were required to predict which of two outcomeswould occur on each trial based on a particular combination of cues presented. Theyconcluded that the striatum plays a critical role in the ability to acquire nonmotordispositions that depend on new stimulus–response associations. Second, in a brain-imaging study with PET, Singh and his colleagues (1993) have reported increasedactivity in the striatum and thalamus while normal control subjects were executingblocks of trials in a random condition of the four-choice serial reaction-time (SRT)task and thus suggested that simple sensori-motor associations may depend on theintegrity of a striato-thalamic circuit.
Likewise, evidence in favor of the cerebellar participation in combining simplemovements into more complex actions is based on the study of cell physiology, whichhas demonstrated that this structure is involved in linking together the constituent,simpler movements that make up volitional compound motor acts (Thach et al., 1992)and in providing an on-line modification of activity in the central motor pathwaysthat are required for optimal coordination of movements (Bloedel, 1992; Ito, 1982,1993). This notion is congruent with a series of studies showing that cerebellar dam-age impairs the ability to translate a programmed motor sequence into action beforethe onset of movement (Inhoff & Bisiacchi, 1990; Inhoff et al., 1989; Inhoff & Rafal,1990). It should be noted that some researchers (Benecke, Rothwell, Dick, Day, &Marsden, 1987; Canavan, Passingham, Marsden, Quinn, Wyke, & Polkey, 1989;Georgiou, Bradshaw, Iansek, Phillips, Mattingley, & Bradshaw, 1994; Harrington &Haaland, 1991; Stelmach, Worringham, & Strand, 1987; Weiss, Stelmach, & Hefter,1997; see Dominey & Jeannerod, 1997, for a review) have previously investigatedthe role of the striatum in the sequencing of movements, by examining the perfor-mance of patients with PD who were required to switch between two completelydifferent types of motions such as elbow extension and hand squeeze (Benecke etal., 1987), or were asked to shift from one step in the sequence to the next by changinghand postures (Harrington & Haaland, 1991), in situations where patients had explicitknowledge of the sequence of movements they had to perform. By contrast, in thisexperiment, the notion first proposed by Flourens (1824), Babinski (1899), andHolmes (1939) that damage to the cerebellum produces a ‘‘decomposition of move-ments’’ was further investigated using a new task that did not require switching be-tween separate movements, but instead measured the patients ability to perform awell-articulated sequence of movements in an implicit fashion.
The present study explored, in the same groups of patients, the distinct contributionof the striatum and the cerebellum in the two types of skilled behaviors describedabove by comparing the performance of patients in early (Stage 1) or advanced stages(Stages 2–3) of Parkinson’s disease (PD), and of a group of patients with damageto the cerebellum (CE), to that of a group of aged (ANC) or young (YNC) matchednormal controls, respectively, on two visuomotor skill-learning tasks. Their abilityto make stimulus–response associations was examined using a version of the SRTtask first used by Willingham and Koroshetz (1993) in which stimuli were presentedat random and the subjects had to respond by pressing a button located to the rightof the stimulus that was displayed. Second, implicit sequencing of learned movementswas tested with an adapted version of the Mirror-Tracing Test in which subjectswere required to trace a series of complex figures which, unbeknown to the subjects,

192 LAFORCE AND DOYON
consisted of the juxtaposition of three simple figures that they had either practicedprior to testing (Practiced condition) or had never traced individually before (Unprac-ticed condition). In accordance with the models mentioned above, it was predictedthat, compared to the ANC group, patients in the PD groups would show a deficitin acquiring perceptual-motor associations as reflected by a smaller reduction in reac-tion time across sessions, whereas the performance of patients in the CE group wouldnot differ significantly from that of the YNC group on this task. Based on the resultsof previous studies (Doyon, Karni, Song, Adams, Maisog, & Ungerleider, 1997b;Doyon, Laforce, Bouchard, Gaudreau, Roy, Poirier, Bedard, Bedard, & Bouchard,1998), we hypothesized that this impairment should be more evident in the late (slow)phase as opposed to the early (fast) phase of learning. It would also be more severein patients with a bilateral striatal dysfunction (i.e., patients in Stages 2–3 of PDaccording to the Hoehn and Yahr scale, 1967) versus those with a unilateral motordefect (i.e., Stage 1). By contrast, it was expected that, compared to their respectivecontrol groups, patients in the CE group, but not those in the PD groups, would failto show a facilitation effect when tracing triads that were composed of practicedsimple figures versus those that were made up of unpracticed figures.
METHODS
Subjects
Five groups of subjects participated in this study. All of the patients were recruited via the Departmentof Neurological Sciences and Neuroradiology at the Hopital de l’Enfant-Jesus, Quebec City (Quebec),Canada, whereas aged and young normal control subjects were either acquaintances of the experimentersor volunteers from the community. None of the controls had a positive history of a psychiatric or neuro-logical disorder. Each subject gave informed written consent for their participation in the study, whichwas approved by the Review Ethics Board of the Hopital de l’Enfant-Jesus.
Parkinson’s Disease Groups (PD)
Two groups of patients with a diagnosis of idiopathic Parkinson’s disease (PD) were included in thepresent experiment. The first group was composed of 15 patients (5 female and 10 male) in Stage 1 ofthe disease as assessed by an experienced neurologist (Dr. P. J. Bedard, Hopital de l’Enfant-Jesus) usingHoehn and Yahr’s scale (1967). On average, these patients were 58.7 (SD 5 9.5) years old and had12.8 (SD 5 4.5) years of education (see Table 1). The second group consisted of 15 patients (7 femaleand 8 male) who were in Stages 2–3 of the disease and who were, on average, 59.3 (SD 5 6.3) yearsold and had 13.5 (SD 5 4.2) years of education. All of these patients were taking optimal levels oflevodopa medication at the time of testing (i.e., appropriate levels of levodopa as recommended bytheir neurologist). Patients with drug-induced parkinsonism, multiple system atrophy, cerebro-vasculardisease, epilepsy, history of alcoholism, head injury or tumor, cerebellar disturbances, or disproportionateoculomotor and autonomic dysfunction were excluded from this study.
Cerebellar Group (CE)
A heterogeneous group of 15 patients [mean age (years) 5 40.7, SD 5 11.8; mean level of education(years) 5 11.9, SD 5 3.0] with a radiologically documented lesion to the cerebellum was also tested(see Table 1). Twelve of these had pure cerebellar atrophy (PCA), while the last three had lesionsextending into the brainstem or spinal cord. All of these patients showed signs of dysarthria, ataxia,and/or dysmetria, although the severity of these cerebellar symptoms differed between patients. Becausetwo patients with PCA were able to complete testing on only half of the skill-learning tasks (i.e., theMirror-Tracing task or the random version of the SRT task), two other patients with PCA were includedinto the study to complete the data acquisition on the remaining task. It is important to note that thelatter patients were well matched to the original group of patients so that there were still no significantdifferences when compared to the control subjects with respect to their mean age and mean level ofeducation as well as to the sex distribution. In addition, these new patients showed a pattern of results

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 193
TA
BL
E1
Subj
ects
’C
hara
cter
istic
s
PD:S
tage
1aPD
:St
ages
2–3a
AN
CC
ereb
ella
rY
NC
Var
iabl
e/gr
oup:
(n5
15)
(n5
15)
(n5
15)
(n5
15)
(n5
15)
Age
(yea
rs):
Mea
n(S
D)
58.7
(9.5
)59
.3(6
.3)
53.0
(9.6
)40
.7(1
1.8)
42.5
(9.4
)E
duca
tion
(yea
rs):
Mea
n(S
D)
12.8
(4.5
)13
.5(4
.2)
13.4
(4.0
)11
.9(3
.00)
14.1
(3.3
)Se
x(f
emal
e/m
ale)
5/10
7/8
9/6
8/7
5/10
Dia
gnos
isA
kine
to-r
igid
:2
Aki
neto
-rig
id:
8V
CA
:4
Tre
mor
:2
Tre
mor
:1
OPC
A:
1M
ixed
:11
Mix
ed:
6C
A:
8SP
:2
Dur
atio
nof
the
dise
ase
0–5
year
s:11
0–5
year
s:3
0–5
year
s:4
6–10
year
s:4
6–10
year
s:6
6–10
year
s:5
11–3
0ye
ars:
011
–30
year
s:6
11–3
0ye
ars:
6L
ater
aliz
atio
nL
eft:
7L
eft:
0L
eft:
7R
ight
:8
Rig
ht:
0R
ight
:2
Bila
tera
l:0
Bila
tera
l:15
Bila
tera
l:6
Med
icat
ion
L-D
opa:
15L-D
opa:
15A
ntic
holin
ergi
csA
ntic
holin
ergi
csA
rtan
e:2
Art
ane:
4Pa
rsita
n:0
Pars
itan:
1
Abb
revi
atio
ns:
VC
A,v
ascu
lar
cere
bral
acci
dent
;OPC
A,o
livo-
pont
o-ce
rebe
llar
atro
phy;
CA
,cer
ebel
lar
atro
phy;
SP,s
pino
cere
bella
rat
roph
y.N
umbe
rsin
pare
n-th
eses
repr
esen
tst
anda
rdde
viat
ions
ofth
em
ean.
aH
oehn
and
Yah
r’s
Scal
e(1
967)
.

194 LAFORCE AND DOYON
on the basic neuropsychological assessment that was similar to the overall group of patients in the CEgroup.
Normal Control Groups
Two separate groups of normal control subjects were selected to match the clinical groups with respectto mean age, mean level of education, and sex distribution (see Table 1). They were composed of agroup of 15 aged-normal subjects (ANC) and of a group of 15 young-normal subjects (YNC) that weretested, respectively, as controls for the PD and cerebellar groups. Again, because a few control subjects(n 5 4) were only available to complete half the testing, other subjects were recruited to replace themin order to bring the sample size up to 15 subjects in each group. These subjects were selected to matchthe overall groups’ characteristics with regard to age, education, and sex distribution.
Basic Neuropsychological Assessment
A short battery of neuropsychological tests was administered to the patients in the three clinical groupsin order to eliminate those showing signs of dementia and/or depression. Because of the lengthy skill-learning protocol, it was decided that the neuropsychological battery of tests was designed only to elimi-nate those showing signs of dementia and/or depression. This assessment consisted of the Mini-MentalState Examination (Folstein, 1983); the ‘‘Vocabulary,’’ ‘‘Digit Span,’’ ‘‘Picture Arrangement,’’ and‘‘Block Design’’ subtests of the WAIS-R (Wechsler, 1987); as well as the French version of the BeckDepression Inventory—Revised (BDI; Bourque & Beaudette, 1982). Because we wanted to avoid testingpatients on a ‘‘bad day,’’ all of them provided their own subjective estimate of their health conditionbefore testing began by filling the General Health Status Scale. This homemade, nonvalidated scaleranged from 1 to 3 where patients had to indicate if they felt their condition (including their motorsymptoms) at the time of testing was 1 5 Worse than usual, 2 5 Same as usual, and 3 5 Better thanusual. This measure was administered to ensure that patients were tested under optimal conditions. Nopatients showed clinical evidence of motor deterioration during testing sessions. It should be noted thatPD patients in Stage 1 of the disease showed, on average, a score of 11.5 (SD 5 9.1) on the BDI, whichreflects signs of a mild level of depression in this group. However, these patients were not excludedbecause (1) Taylor, Saint-Cyr, Lang, and Kenny (1986) have demonstrated that such mild depressivestates do not interfere significantly with patients’ performance on cognitive tasks and (2) consistent withthis notion, the results of the present neuropsychological assessment revealed that these patients did notsuffer from an overall deterioration in their level of cognitive functioning. Finally, except for the threeclinical groups who showed an impairment on the Purdue Pegboard task, hence reflecting a deficit infine motor coordination, the results of the patients on the remaining tests of the basic neuropsychologicalevaluation reveal that they did not show any significant cognitive deterioration (see Table 2).
Materials and Procedure
Random Version of the SRT Task: Perceptual-Motor Skill Learning
The subject’s ability to acquire perceptual-motor associations was measured with a random versionof the SRT task. This test was originally developed by Nissen and Bullemer in 1987 and subsequentlymodified by several researchers in the field. The present study used a similar version than Willinghamand Koroshetz (1993). This test was administered using a response box that had four identical lights(stimuli) and four buttons, one below (1.75 cm) each light. The lights and buttons were arranged horizon-tally equidistant from one another. This box was connected to an IBM PC computer that controlledstimulus presentation and recorded the subject’s reaction time (RT) and accuracy on each trial.
Contrary to the original SRT task, in which each block of trials was composed of a repeated 10-itemsequence (Doyon et al., 1997a), this version used a completely random presentation of the stimuli. Thesubjects were instructed to use the middle and index fingers of each hand and to keep one finger on eachof the four keys. Furthermore, they were asked to press as quickly as possible the button corresponding tothe right of the one under which the visual stimulus (light) appeared, while trying to make as few errorsas possible. When the stimulus to the far right was displayed, the subjects were instructed to press thebutton to the far left. This experimental manipulation was used to increase the level of difficulty of thetask, hence reducing the possibility of obtaining floor effects within the six training sessions. The stimulusremained displayed until the subject responded. After the subject’s response, the light went off and wasfollowed 500 ms later by the display of another stimulus. Each subject completed 6 sessions of 4 blocks,each block comprising 100 random trials (total 5 2400 trials). The sessions were separated by pausesvarying between 10 and 20 min, while the blocks of trials within a session were administered 90 s apart.

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 195
TA
BL
E2
Res
ults
ofth
eB
asic
Neu
rops
ycho
logi
cal
Ass
essm
ent
for
the
Clin
ical
Gro
ups
PD:
Stag
e1
PD:
Stag
es2–
3C
ereb
ella
r(n
515
)(n
515
)(n
515
)T
est/
grou
pM
ean
(SD
)M
ean
(SD
)M
ean
(SD
)N
orm
ativ
eda
ta
Min
i-M
enta
lSt
ate
Exa
min
atio
n28
.90
(1.0
)28
.30
(1.4
)27
.30
(1.8
)C
utof
f:,
271
Gen
eral
Hea
lthSt
atus
2.13
(.35
)2.
13(.
35)
2.20
(.56
)C
utof
f:,
2B
eck
Dep
ress
ion
Inve
ntor
y11
.50
(9.1
)*9.
27(5
.1)
7.47
(5.9
)C
utof
f:.
9Pu
rdue
Pegb
oard
(Bot
hH
ands
)9.
13(1
.9)*
8.33
(1.5
)*7.
13(2
.8)*
13.1
(1.3
)2
WA
IS-R
:M
ean
Scal
edSc
ores
(SD
)D
igit
Span
10.5
0(3
.0)
9.33
(2.7
)8.
13(2
.1)
10(3
)3
Voc
abul
ary
10.9
0(2
.8)
11.4
0(1
.8)
9.40
(1.9
)10
(3)3
Pict
ure
Arr
ange
men
t9.
40(3
.3)
9.93
(3.0
)7.
73(2
.7)
10(3
)3
Blo
ckD
esig
n11
.50
(3.3
)12
.00
(3.9
)9.
73(3
.0)
10(3
)3
Not
e.N
umbe
rsin
pare
nthe
ses
repr
esen
tst
anda
rdde
viat
ions
ofth
em
ean.
Ref
eren
ce:
1 Fols
tein
(198
3):
2 Spre
enan
dSt
raus
s(1
991)
;3 W
echs
ler
(198
7).
Not
e.N
umbe
rsin
pare
nthe
ses
repr
esen
tst
anda
rdde
viat
ions
ofth
em
ean.
*Dif
fers
sign
ifica
ntly
from
norm
ativ
eda
ta.

196 LAFORCE AND DOYON
FIG. 1. Examples of the (A) simple figures and (B) complex triads used in the new adapted versionof the Mirror-Tracing Test. In this task, subjects were asked to trace the figures through the reflec-tion of a mirror as quickly as possible, while avoiding touching the contours. The simple figures con-sisted of curved or angled designs, whereas the complex triads were composed of the consecutive juxta-position of three simple figures.
Mirror-Tracing Task: Integration of Practiced Movements
Integration of practiced movements was measured with a new visuomotor skill-learning task that wasdeveloped in our laboratory and was based on the original mirror-tracing test. In this task, the subjectswere required to learn to trace figures of different shapes while viewing their hand and figures throughthe reflection of a mirror. The apparatus consisted of a wooden baseboard (30 3 30 cm) with a rearvertical panel (30 3 30 cm) on which a mirror (23 cm 3 23 cm) was fixed. Perpendicular to this panel,a metal plate was mounted 15 cm above the baseboard to prevent the subject’s direct view of the handand figures. This metal plate could be fixed on either side of the board to allow right- or left-handedsubjects to complete the task adequately using their dominant hand.
In this experiment, the subjects were asked to trace two different types of drawings including simplefigures and complex triads. The simple figures (Fig. 1A) consisted of curved or angled designs, whereasthe triads (Fig. 1B) were composed of the consecutive juxtaposition of three of these simple figures.All of these figures were originally created using the software Autocad (Version 12, Autodesk Inc.).
Paper sheets on which the different simple figures or triads were illustrated were placed directly onthe baseboard at the beginning of each trial. Using a pencil, the subjects were asked to trace the figuresas quickly as possible, while avoiding touching the edges. They were also required to follow as accuratelyas possible the overall shape of the figures. Each trial began by asking the subjects to place their pencilat the starting point displayed on the figure and to start tracing at the ‘‘Go’’ signal. Two dependentmeasures were recorded: The completion time (CT) (i.e., the amount of time in seconds required tocomplete each figure from the beginning to the finishing line) and the number of errors (i.e., the numberof times a subject crossed the borders of a figure).
Each subject completed four phases of testing. In Phase I, the subjects were asked to trace 12 simplefigures in order to familiarize themselves with the mirror-tracing task. In Phase II, the subjects werethen required to trace each of the 12 triads once. These were later divided into two sets and were usedin both the Practiced and Unpracticed conditions of Phase IV. The reasons for this testing phase weretwofold: first, to familiarize the subjects with the task using complex instead of simple figures. Second,although the results of a pilot study (described below) revealed that the triads used in both Practicedand Unpracticed conditions did not differ in terms of their physical characteristics and their level ofdifficulty, this phase was administered to ensure that this was the case in the present experiment. InPhase III (called ‘‘Learning of simple figures’’), the subjects were given 10 blocks of practice in whichthey had to trace 18 new simple figures that were repeatedly presented at random within each block.The aim of this phase was to have the subjects learn a series of simple movements by repeatedly tracingthe same simple figures. Finally, Phase IV (named ‘‘Integration of practiced movements’’) consisted ofthree testing blocks of trials, each block comprising six triads in a Practiced condition and six triadsin an Unpracticed condition. In the Practiced condition, the triads were composed of the consecutivejuxtaposition of three simple figures that were previously practiced in Phase III of testing, while thetriads in the Unpracticed condition were made up of three simple figures that subjects had never tracedindividually before. This phase was administered to explore the ability of the subjects to integrate learned

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 197
simple movements into a fluid sequence by comparing the performance of the triads in the Practiced vsUnpracticed conditions.
It is important to note that the level of difficulty of the different sets of simple figures that were usedin the familiarization and learning phases, as well as for designing the triads of the Practiced and Unprac-ticed conditions of Phase IV, was controlled based on the results of two pilot studies. The first pilotexperiment was carried out with 20 control subjects [mean age (years) 5 25.8, SD 5 7.2; mean levelof education (years) 5 14.3, SD 5 5.6] to determine the mean completion time and the number of errorsthat were made while tracing once the 48 simple figures. On average, these figures were 10.4 cm (SD 52.6) long; the subjects took 6.91 s (SD 5 9.87) and committed 0.73 (SD 5 1.16) errors per figure.Twelve of these figures were selected to be used in Phase I of the new mirror-tracing task discussedabove, whereas the remaining 36 simple figures were then divided randomly into two subsets of 18figures, which were used to create the triads in the Practiced and Unpracticed conditions.
The second pilot experiment was conducted to ensure that the triads used in the Practiced conditionwere equivalent to those in the Unpracticed condition with respect to their physical characteristics andlevel of difficulty (i.e., accuracy and completion time). As expected, the results of a one-way analysisof variance conducted on the performance of a group of 20 new normal control subjects [mean age(years) 5 26.3, SD 5 11.2; mean level of education (years): 13.4, SD 5 9.7] yielded no significantdifference between these two types of complex figures. On average, the triads in the Practiced conditionhad a total length of 33.9 cm (SD 5 1.6), whereas the triads in the Unpracticed condition were 34.4cm (SD 5 0.47) long. Furthermore, the triads in the Practiced condition took a mean time of 15.7 s(SD 5 12.6) to complete, whereas 16.5 s (SD 5 11.1) were required for the triads in the Unpracticedcondition. Finally, there was no significant difference in the mean number of errors committed whentracing both types of triads (Practiced 5 2.06, SD 5 2.69; Unpracticed 5 2.27, SD 5 3.04).
Experimental Design
This study was conducted on 2 separate days of testing. The subjects were first asked to complete ashort neuropsychological assessment. They were then required to execute either the random version ofthe SRT test or the mirror-tracing task on the first of these 2 days, the order of administration of thesetwo visuomotor skill-learning tasks being counterbalanced within each group.
RESULTS
Patients in both PD groups and the CE group were well matched to their respectivenormal control subjects, as separate one-way analyses of variance (ANOVAs) re-vealed no significant difference with respect to either age or level of education. Also,there was no significant difference in sex distribution of the subjects in both clinicaland control groups as measured with the χ2.
In the random version of the SRT task, the dependent measures of interest were(a) the number of correct responses and (b) the mean RT in milliseconds. In theadapted version of the Mirror-Tracing Test, the dependent measures of interest were(a) the number of errors and (b) the mean CT in seconds. Finally, separate statisticalanalyses were conducted comparing the PD and CE groups with their respectivecontrol groups.
Random Version of the Serial Reaction-Time Task: Perceptual-Motor SkillLearning
All five groups (i.e., both PD groups and the ANC, CE, and YNC groups) werevery accurate on this test. The mean percentage of correct responses was 91.6%(SD 5 1.23) for PD patients in Stage 1, 87.4% (SD 5 0.8) for PD patients in Stages2–3, and 90.2% (SD 5 1.35) for the ANC group. CE patients reached an 86.2%(SD 5 9.1) accuracy while YNC got 90.3% (SD 5 4.7). There were no group differ-ences in either cases.
A one-way ANOVA was first conducted in order to determine whether the PDgroups differed from the ANC group with respect to the mean RT on the very first

198 LAFORCE AND DOYON
FIG. 2. Random version of the serial reaction-time task: perceptual motor skill learning. Mean reac-tion times of the six training sessions for both groups of Parkinson’s disease (PD) patients and the groupof aged normal control subjects (ANC). Error bars represent standard errors of the mean.
practice session. This analysis yielded no main effect of Group, hence suggesting thatthe subjects’ ability to perform the random version of the SRT task was equivalent atthe beginning of the training sessions.
A repeated-measures ANOVA with trend analysis was then conducted on the RTdata of Session 1 to Session 6 (see Fig. 2). The results only revealed a main effectof Group, F(2, 42) 5 7.65, p , .001, PD patients in Stages 2–3 of the disease beingoverall significantly slower to respond (501.43 ms, SD 5 122.83) to the stimuli thanthe two other groups (ANC:377.94 ms, SD 5 67.99; PD Stage 1:425.71 ms, SD 584.69), and a main effect of Session, F(5, 210) 5 80.54, p , .0001, all groupsimproving their performance across training sessions. Such results suggests that, asa whole, the performance of PD patients did not diverge significantly form controlsubjects in their learning ability on this version of the SRT task.
Because it was predicted that the severity of the disease might affect performanceon learning of S-R associations, subsequent ANOVAs with trend analysis were car-ried out separately between the three groups to further investigate their rate of learn-ing. The contrast analysis conducted between the ANC group and PD patients inStage 1 did not reach significance. The contrast analysis between the performanceof the ANC group and the PD patients in Stages 2–3 did reveal, however, a significantquadratic Group 3 Session interaction, F(1, 28) 5 4.72, p , .05. Both groups ofpatients did not differ with respect to each other. Post hoc pairwise tests (using New-man–Keuls procedure) used to further investigate between-sessions changes in per-formance showed that significant improvement was seen up to Session 2 in the PDStages 2–3 group (Sessions 1–2: q 5 8.99, p , .05), up to Session 3 in the PD Stage1 group (Sessions 2–3: q 5 3.28, p , .05), and to Session 4 in the ANC group(Sessions 2–3: q 5 3.26, p , .05; Sessions 3–4: q 5 3.19, p , .05). Altogether,these findings suggest that the three groups showed some learning of the S-R associa-tions, but that patients with a bilateral striatal dysfunction differed in their patternof acquisition of the task across training sessions. These findings are consistent withthe notion that the severity of the deficit in this type of learning mechanism is depen-dent upon the extent of striatal dysfunction.
Figure 3 shows the mean RT data for both the CE and YNC groups on the six

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 199
FIG. 3. Random version of the serial reaction-time task: perceptual motor skill learning. Mean reac-tion times of the six training sessions for the group of patients with damage to the cerebellum and thegroup of young normal control subjects (YNC). Error bars represent standard errors of the mean.
training sessions. A one-way ANOVA conducted on the results of Session 1 onlyrevealed a significant effect of Group, F(1, 28) 5 25.78, p , .0001, suggesting thatcontrary to the PD groups, the patients with a lesion to the cerebellum were signifi-cantly slower than their matched normal controls at the beginning of testing. AnANOVA for repeated measures using the mean RT of Sessions 1 to 6 showed thatthe CE group (627.24 ms, SD 5 182.57) was also significantly slower to respond,F(1, 28) 5 31.20, p , .0001, than subjects in the YNC group (344.94 ms, SD 593.22). There was also a main effect of Session, F(5, 140) 5 34.53, p , .0001,indicating that the RT of the two groups decreased from Session 1 to Session 6.Contrary to the results of both PD groups, the Group 3 Session interaction was notsignificant, F(5, 140) 5 .75, p 5 .58, thereby suggesting that both the CE and theYNC groups did not differ in their ability to acquire perceptual-motor associations.
Correlational Analyses
To determine whether the deficit in visuomotor skill learning observed in both PDgroups could be attributed to a cognitive dysfunction, a mood disturbance, or to theseverity of motor symptoms, separate Pearson’s product–moment correlations werecarried out between the patients’ level of learning (as measured by subtracting themean RT of Session 6 from that of Session 1) and several neuropsychological mea-sures such as the Mini-Mental State Examination, the General Health Status scale,the Beck Depression Inventory, the Purdue Pegboard, and the results obtained withthe different subtests from the WAIS-R (‘‘Vocabulary,’’ ‘‘Digit Span,’’ ‘‘PictureArrangement,’’ and ‘‘Block Design’’; Wechsler, 1987) gathered before testing began.None of the correlations were significant, hence suggesting that the perceptual-motorlearning deficit in both PD groups could not be attributed to those variables.
Functional Dissociation
Thus far, the distinct contribution of the striatum and the cerebellum to the learningof S-R associations was tested by comparing the performance of patients in early

200 LAFORCE AND DOYON
FIG. 4. Mirror-tracing task: learning of simple figures. Mean completion times for both PD groupsand the ANC group. Error bars represent standard errors of the mean.
(Stage 1) or advanced stages (Stages 2–3) of PD, and of a group of patients withdamage to the cerebellum, to that of groups of aged and young matched normalcontrols, respectively. The use of healthy subjects as controls for the clinical groupswas planned a priori because it was expected that patients in both PD and CE groupswould differ significantly with respect to age and that this type of design would bethe most valid approach to address our hypothesis. Out of curiosity though, and inorder to explore the hypothesis of a functional dissociation between these structures,additional ANOVAs were conducted by comparing directly the performance of thetwo subgroups of patients with PD (Stage 1 and Stages 2–3) to that of the group ofpatients with a lesion to the cerebellum (CE). These analyses did not reveal a signifi-cant trend, hence suggesting that the three clinical groups did not differ in their learn-ing ability on this version of the SRT task.
Mirror-Tracing Task: Integration of Practiced Movements
Phase I and II: Familiarization to the tracing of simple and complex figures. Re-sults obtained on the familiarization phases indicated that PD patients in Stage 1produced, on average, fewer tracing errors than the PD patients in Stages 2–3 andthe ANC group. CE and YNC subjects had similar accuracy levels. Figures in bothPracticed and Unpracticed conditions were traced with the same accuracy by allgroups. Finally, the clinical groups did not differ from their respective control groupsin terms of the mean time to complete the simple or complex figures.
Phase III: Learning of simple figures. Participants made very few errors (i.e.,less than one per figure on average) while tracing the simple figures in this phase.The mean CT in seconds required to trace the simple figures for both PD groups andthe ANC group across the 10 training Blocks of trials are shown in Fig. 4. A repeated-measures ANOVA yielded a Group effect, F(2, 42) 5 15.68, p , .0001, as PDpatients in Stages 2–3 were slower to trace the simple figures (4.11, SD 5 0.15) thanthe other groups (ANC: 2.25, SD 5 0.07; PD Stage 1: 3.83, SD 5 0.10). The resultsalso showed a main effect of Block, F(9, 378) 5 29.23, p , .0001, as all groupsimproved their performance across training trials. Finally, there was no significantGroup 3 Block interaction, hence suggesting that both PD groups and the ANCgroup showed a similar amount of learning to trace the simple figures.

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 201
FIG. 5. Mirror-tracing task: learning of simple figures. Mean completion times for the CE and YNCgroups. Error bars represent standard errors of the mean.
Figure 5 shows the mean CT required to trace the simple figures for the CE andYNC groups across the 10 training Blocks of trials. The repeated-measures ANOVArevealed a Group effect, F(1, 28) 5 29.68, p , .0001, the YNC subjects being fasterto trace the simple figures (2.12, SD 5 0.91) than the CE group (4.44, SD 5 2.77).The results also showed a main effect of Block, F(9, 252) 5 9.95, p , .0001, becauseboth groups improved their performance across training trials. By contrast, their wasno significant Group 3 Block interaction, suggesting that these two groups did notdiffer in their learning ability to trace simple figures.
Phase IV: Integration of practiced movements. Data obtained in Phase IV wereanalyzed using a three-way ANOVA for repeated measures. Results of the PD–ANCanalyses revealed a main effect of Group, F(2, 42) 5 4.26, p , .05, where PD patientsin Stages 2–3 produced significantly more errors (1.79, SD 5 2.18) than the ANCgroup (.86, SD 5 1.32) and the PD group in Stage 1 (.73, SD 5 1.11). There wasalso a main effect of Condition, F(1, 42) 5 18.45, p , .0001, as the triads in thePracticed condition were traced with fewer errors (.95, SD 5 1.35) than those in theUnpracticed condition (1.31, SD 5 1.72), and a main effect of Block, F(2, 84) 57.33, p , .005, as all groups improved their performance across the three blocks oftrials. The CE–YNC analyses only yielded a main effect of Block, F(2, 56) 5 4.65,p , .05. In both cases, no significant Group 3 Condition interaction was noted,hence suggesting that all groups showed the same level of precision in tracing thetriads in both conditions.
The results of a three-way repeated-measures ANOVA conducted on the mean CTof the three blocks of trials for the two PD groups and the ANC group are presentedin Fig. 6. The results showed a main effect of Group, F(2, 42) 5 4.72, p , .05, asthe ANC subjects were significantly faster (8.38, SD 5 4.76) than the two PD groups(PD Stage 1: 12.13, SD 5 11.73; PD Stages 2–3: 12.6, SD 5 7.48). There was amain effect of Condition, F(1, 42) 5 79.65, p , .0001, the triads in the Practicedcondition being traced significantly faster (9.76, SD 5 4.79) than the triads in theUnpracticed condition (12.32, SD 5 7.28). The results also showed a learning effectin tracing the complex figures from Block 1 to Block 3, F(2, 84) 5 13.78, p , .0001.More importantly, however, the Group 3 Condition interaction did not reach signifi-cance, suggesting that the three groups showed a similar facilitation effect in the
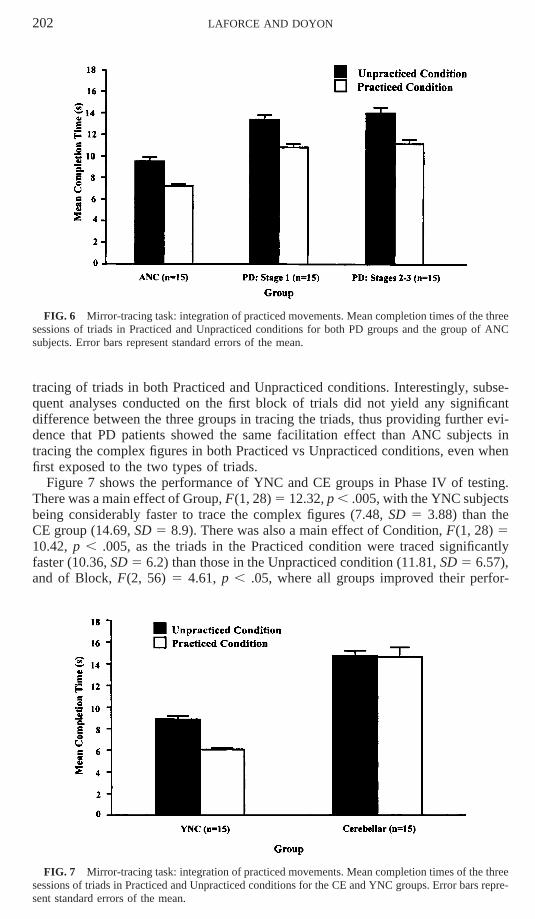
202 LAFORCE AND DOYON
FIG. 6 Mirror-tracing task: integration of practiced movements. Mean completion times of the threesessions of triads in Practiced and Unpracticed conditions for both PD groups and the group of ANCsubjects. Error bars represent standard errors of the mean.
tracing of triads in both Practiced and Unpracticed conditions. Interestingly, subse-quent analyses conducted on the first block of trials did not yield any significantdifference between the three groups in tracing the triads, thus providing further evi-dence that PD patients showed the same facilitation effect than ANC subjects intracing the complex figures in both Practiced vs Unpracticed conditions, even whenfirst exposed to the two types of triads.
Figure 7 shows the performance of YNC and CE groups in Phase IV of testing.There was a main effect of Group, F(1, 28) 5 12.32, p , .005, with the YNC subjectsbeing considerably faster to trace the complex figures (7.48, SD 5 3.88) than theCE group (14.69, SD 5 8.9). There was also a main effect of Condition, F(1, 28) 510.42, p , .005, as the triads in the Practiced condition were traced significantlyfaster (10.36, SD 5 6.2) than those in the Unpracticed condition (11.81, SD 5 6.57),and of Block, F(2, 56) 5 4.61, p , .05, where all groups improved their perfor-
FIG. 7 Mirror-tracing task: integration of practiced movements. Mean completion times of the threesessions of triads in Practiced and Unpracticed conditions for the CE and YNC groups. Error bars repre-sent standard errors of the mean.

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 203
mance across the three Blocks of trials. As predicted and contrary to the profile ofthe PD patients, this analysis revealed a significant Group 3 Condition interaction,F(1, 28) 5 9.05, p , .01. This suggests that the CE group did not show the samelevel of facilitation in tracing the triads in both Practiced and Unpracticed conditions.Such an impairment is particularly revealing, as both groups had shown evidence of asimilar degree of learning to trace the simple figures in Phase III of testing. Moreover,subsequent analyses conducted on the first block of trials yielded a similar Group 3Condition interaction, F(1, 28) 5 7.24, p , .05, suggesting that the difference inperformance between Practiced and Unpracticed conditions could also be seen at thebeginning of testing when subjects were first exposed to the complex figures. Consid-ered together, these results suggest that, compared to the PD patients, those withdamage to the cerebellum showed an impairment in the integration of learned simplefigures.
In order to eliminate the possibility that the impairment in the tracing of triads inboth Practiced and Unpracticed conditions found in the cerebellar group was due tothe presence of additional extracerebellar damage, further analyses were carried outcomparing the performance of the control group to that of a subgroup of patients(n 5 12) in which the lesions were circumscribed to the cerebellum. Interestingly,these analyses revealed a very similar pattern of findings to that of the group as awhole. Indeed, the results showed a significant effect of Group, F(1, 25) 5 8.50,p , .01, Condition, F(1, 25) 5 14.15, p , .005, and again, the Group 3 Conditioninteraction reached significance, F(1, 25) 5 5.46, p , .05. Thus, as for the resultsof the whole group of patients with damage to the cerebellum, these findings indicatethat patients with lesions restricted to the cerebellum also show a lack of facilitationeffect in tracing complex triads of figures in the Practiced condition.
Correlational Analyses
To determine whether the impairment in the integration of learned movementsfound in the CE group could be due to cognitive or mood defects, separate Pearson’sproduct–moment correlations were carried out between the patients’ level of learning(as measured by subtracting the mean CT of the three Blocks of triads in the Practicedcondition from that of the three Blocks of triads in the Unpracticed condition) andthe dependent measures that were gathered during the basic neuropsychological eval-uation. Because the importance of assessing the severity of the cerebellar dysfunctionhas been stressed in previous studies (e.g., Inhoff et al., 1989; Kish, El-Awar, Stuss,Nobrega, Currier, Aita, Schut, Zoghbi, & Freedman, 1994), a rating of the motorsigns in the upper limbs based on adiadokokinesia, dysmetria, and intention tremorswas included in the present analysis. The latter rating was performed by an experi-enced neurologist, using scores between 0 and 5, where 0 indicates a relatively normallevel of functioning and 5 indicates a severe dysfunction (Inhoff et al., 1989). Theresults of these analyses revealed no significant correlation, therefore suggesting thatthe deficit in the integration of practiced movements observed in the CE group couldnot be attributed to a cognitive dysfunction, a mood disturbance, nor to the severityof the motor symptoms.
Functional Dissociation
Again, the performance of the two PD subgroups and the CE group was compareddirectly to test further the functional dissociation with regard to the integration ofmovements in Phase IV of testing. Interestingly, these analyses revealed a similarpattern of findings, and the Group 3 Condition interaction was highly significant,F(2, 132) 5 11.50, p , .0001. Thus, these findings suggest that only patients with

204 LAFORCE AND DOYON
lesions to the cerebellum show a lack of facilitation effect in tracing complex triadsof figures in the Practiced condition.
DISCUSSION
The goal of this study was to explore the potentially distinct mechanisms underwhich the striatum and the cerebellum operate when learning a visuomotor skill.More specifically, this investigation aimed at exploring the effects of damage to thesestructures on learning mechanisms thought to be dependent upon the striatum (Gray-biel & Kimura; 1995; Knowlton et al., 1996; Marsden & Obeso, 1994; McDonald &White, 1993; Singh et al., 1993) and the cerebellum (Bloedel, 1992; Gilbert & Thach,1977; Ito, 1993; Thach et al., 1992). The results showed that PD patients with bilateralstriatal damage had a different learning profile on the random version of the SRTtask when compared to the ANC group, whereas no significant difference was foundbetween the performance of the CE and the YNC groups. By contrast, patients inthe CE group, but not in the PD groups, failed to show a facilitation effect whentracing triads of figures in the Practiced vs Unpracticed conditions of the mirror-tracing task. Further correlational analyses revealed that the respective impairmentsin both skill-acquisition tests were not related to a general decline in cognitive func-tioning, to mood disturbances, or to a motor limitation per se. Taken together, theresults of this study suggest that motor functions of the striatum and the cerebellumare potentially dissociable in humans and that these two structures may play distinctroles when acquiring new visuomotor skilled behaviors.
Random Version of the SRT Task: Perceptual-Motor Skill Learning
The results showed that, compared to controls, patients with PD in Stages 2–3 ofthe disease (but not those with damage to the cerebellum) were impaired on a randomversion of the SRT task, as reflected by a flattening of their reaction times startingas soon as in the second session of testing. One possible interpretation of these results,is that this deficit might be attributed to difficulties in ancillary cognitive processesand not to a learning impairment per se. Given that the SRT task requires both atten-tional and visuospatial processing abilities and that deficits in each of these functionshave been reported in patients with a striatal dysfunction (Boller, Passafiume, Keefe,Rogers, Morrow, & Kim, 1984; Doyon, Bourgeois, & Bedard, 1996a; see Brown &Marsden, 1990; Dubois, Boller, Pillon, & Agid, 1991; Ogden, 1990, for reviews),one could argue that the impairment is due to a deficit in these processes. However,the results of the present study suggest that this is not the case because if a problemin attentional and/or visuospatial functions was the source of the impairment, groupdifferences should have been readily observed on the first session of training. Thefact that no difference in performance was noted at the beginning of testing is thusinconsistent with such interpretation.
Another possible interpretation of the learning deficit observed in the PD Stages2–3 group is that their performance was due to their incapacity to produce a speededresponse and not to a learning deficit per se. Indeed, because such patients are knownto have poor motor abilities due to the nature of their disorder, one could suggestthat their performance reflected the fact that they reached their limit earlier thancontrols in executing motor responses rather than their difficulty in learning the stimu-lus–response associations. However, none of the correlational analyses performedon the patients’ level of learning and a measure of fine motor coordination (i.e.,

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 205
Purdue Pegboard) were significant, hence suggesting that the deficits could not beattributed to the severity of motor symptoms.
Taken together, these results support evidence that the striatum, but not the cerebel-lum, contributes to the development of perceptual-motor programs based on S-Rassociations. They are consistent with evidence from physiological and behavioralstudies in animals (Aosaki, Graybiel, & Kimura, 1995; McDonald & White, 1993;Packard et al., 1989; Packard & White, 1990; see Graybiel, 1995; Graybiel & Kimura,1995; Marsden & Obeso, 1994; White, 1989, 1997, for reviews) and from clinicalas well as imaging investigations in humans (Knowlton et al., 1996; Singh et al.,1993). It is possible that this impairment may be due to a difficulty in the automatiza-tion of S-R associations acquired with practice, as all groups showed a significantincrease in performance from Session 1 to Session 2, but the PD group in Stages 2–3 stopped improving as early as in the second session. The latter findings are consis-tent with other studies which suggest that a striatal dysfunction does not affect thelearning of an incremental perceptual-motor skill at the very beginning (i.e., Session1, fast learning stage), but does so in the later (i.e., slow learning phase) stages ofthe acquisition process (Doyon et al., 1997a, 1997b, 1998, 1996b). Thus, this suggeststhat PD does not impair performance during the fast learning stage in which consider-able improvement in performance can be seen within a single training session, but,instead, that it produces a deficit in the slow learning stage during which further gainsare usually observed across several sessions of practice (Karni, 1996; Karni, Meyer,Rey-Hipolito, Jezzard, Adams, Turner, & Ungerleider, 1998).
Mirror-Tracing Task: Integration of Practiced Movements
Consistent with our predictions, the results reveal that patients with damage to thecerebellum, but not those with PD, are impaired on the mirror-tracing task, as theydid not show any facilitation effect when tracing the triad figures in the Practicedcompared to the Unpracticed conditions. It is important to note that this lack of facili-tation effect was observed even though the triads used in the Practiced and Unprac-ticed conditions did not differ in the level of complexity and that patients in the CEgroup showed evidence of learning in tracing the simple figures in Phase III of testing.These results imply that patients in the CE group failed to benefit from earlier practicein tracing the individual simple figures before they were juxtaposed to produce morecomplex designs and thus demonstrate that the integrity of the cerebellum, but notof the striatum, is critical for bridging implicitly together simple learned movementsinto an integrated compound movement. This notion is supported by work fromThach and colleagues, who recently proposed (based on both lesion and neurophysio-logical studies in monkeys) that the cerebellum plays a major role in combiningsimpler (single-joint) elements of movements into more complex (multijoint) coordi-nated acts (Thach, 1996; Thach et al., 1992). These researchers have suggested thatsuch motor control could be mediated by the actions of multiple Purkinje cells (com-bined together via long parallel fibers) onto the body representations found in cerebel-lar nuclei and by their subsequent effects on downstream executive centers such asthe thalamus and motor cortex, the anatomically related brainstem nuclei, and thespinal cord (Thach, 1996). Because normal performance in our mirror-drawing taskrequires that subjects acquired good coordination of the hand, arm, and shoulderwhen learning to trace the simple figures (and not only the triads), our results reveal,however, that the cerebellum would not only be critical for combining actions ofseveral joints, but that it would also be involved in linking movements of multiplebody parts into a well-articulated sequence of movements.
The fact that patients with PD showed a significant facilitation effect, hence sug-

206 LAFORCE AND DOYON
gesting a preserved ability in combining acquired movements into a well-articulatedaction, may appear to be at variance with the results of recent clinical studies whichsuggest that PD impairs the smooth transition between two successive movements(Benecke et al., 1987; Canavan et al., 1989; Georgiou et al., 1994; Harrington &Haaland, 1991; Stern, Mayeux, Rosen, & Ilson, 1983; Weiss et al., 1997; see Domi-ney & Jeannerod, 1997, for a review). Several reasons may explain this apparentdivergence of findings. First, the difference could be related to the type of movementsthat were required. Indeed, in previous studies, a deficit in patients with PD wasobserved, for example, when they were asked to switch between two completelydifferent types of movements such as elbow extension and hand squeeze (Beneckeet al., 1987) or when transitions from one step in the sequence to the next requiredchanges in hand posture (Harrington & Haaland, 1991). By contrast, no clear demar-cation between the movements to perform while tracing the three juxtaposed simplefigures in a triad was required, and thus our task did not elicit as much of the patients’capacity to switch from one motor program (which was learned in Phase III of testing)to another.
Second, the inconsistency may be due to the explicit vs implicit nature of thesequencing of movements that the patients had to execute. In the other studies, pa-tients with PD acquired declarative knowledge of the sequence of movements theyhad to perform because (1) they were asked either to memorize the sequence (Rafal,Inhoff, Friedman, & Bernstein, 1987; Stelmach et al., 1987) or to practice it beforethe experimental testing began (Benecke et al., 1987; Georgiou et al., 1994; Georgiou,Bradshaw, Phillips, Bradshaw, & Chiu, 1995; Georgiou, Iansek, Bradshaw, Phillips,Mattingley, & Bradshaw, 1993; Jones, Phillips, Bradshaw, Iansek, & Bradshaw,1992; Rafal et al., 1987) or (2) they were allowed to refer to a written version of thesequence they had to produce at all times during the experiment (Harrington & Haa-land, 1991). In this study, however, the subjects were unaware that the triads in thePracticed condition were made of the consecutive juxtaposition of the simple figuresthey had practiced in Phase III, nor that the triads in the Unpracticed condition werecomposed of novel simple figures that they had never traced individually before.Therefore, it is possible that the difference between our results and those from earlierstudies is due to the fact that the patients’ ability to combine learned movements intoa sequence was measured in an implicit instead of an explicit fashion.
Third, evidence from animal (Brotchie, Iansek, & Horne, 1991a, 1991b), clinical(Doyon et al., 1997a, 1998; Georgiou et al., 1994, 1995, 1993), and neuroimaginginvestigations (Doyon et al., 1997b, 1996b; Grafton et al., 1994; Jenkins et al., 1994;Seitz, Roland, Bohm, Greitz, & Stone-Elander, 1990) has demonstrated that the stria-tum is critically involved in the late phases of learning where automatization of a skillis thought to occur. Consistent with such a notion is the absence of deficit observed inboth PD groups that may result from the fact that, in the present study, subjects hadnot achieved automatization of the task. It is thus possible that, if a greater numberof trials had been given until subjects achieved asymptotic performance, a deficitfollowing striatal dysfunctions could have been elicited as well.
Finally, from a methodological point of view, the normal performance of the PDgroups could also be explained by the procedure that was used in our study. Contraryto the original version of this task (Blakemore, 1977), in which subjects started totrace directly between the contours of the figure (i.e., star), our version of this taskalways comprised a constant gap of 1 cm between the starting point and the beginningof the simple and complex figures. Given that the patient’s performance was notrecorded before he/she crossed this entry line, it is possible that such a gap helpedby allowing them to set their motor program before they started to trace the figures.However, this seems highly unlikely because the distance of the gap was the same

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 207
for all of the simple and complex figures and also because trials in which subjectswere seen to move around and to take an unusually long time before beginning thetrial were very rarely encountered during the experiment. Nevertheless, we believethat future studies using this paradigm should make use of recent technologies thatallow direct assessment of the pen path during tracing, as this would provide betterinformation about the different motor parameters involved in this type of incrementallearning (e.g., velocity and force).
Last, together with the results of Inhoff and colleagues (Inhoff & Bisiacchi, 1990;Inhoff et al., 1989; Inhoff & Rafal, 1990), as well as those from Thach et al. (1992)using a prism adaptation task, our findings suggest that the cerebellum is not onlyinvolved in the translation of a motor program into action before the onset of a move-ment, but that it plays an important role in combining the movements during theexecution of a sequence of actions as well. They are also consistent with the classicview that the main symptom associated with cerebellar lesions is the decompositionof movements (Holmes, 1939).
CONCLUDING COMMENTS
In conclusion, the present findings are consistent with a distinct contribution ofthe striatum and the cerebellum in human motor learning. They provide additionalevidence in support of a role for the striatum in building, with practice, a repertoireof motor actions that can be triggered in response to appropriate environmental stim-uli. They further suggest that the cerebellum plays a more important role in combininglearned movements together to produce a well-executed motor skilled behavior. Suchfindings are consistent with studies which have found differences in striatal and cere-bellar functions by comparing patients with PD or with a cerebellar lesion 6 yearsapart (Agostino, Sanes, & Hallett, 1996) or by investigating the performance of pa-tients with PD or with cerebellar damage on the SRT task (Pascual-Leone et al.,1993). The current study is, however, limited by an inability to compare both mecha-nisms under similar conditions (i.e., our tasks do not involve the same number oftrials) and also by the fact that more research work needs to be done in order to fullyunderstand the nature of the mechanisms that are measured. Future studies shouldattempt to address these issues.
REFERENCES
Agostino, R., Sanes, J. N., & Hallett, M. (1996). Motor skill learning in Parkinson’s disease. Journalof Neurological Science, 139, 218–226.
Aosaki, T., Graybiel, A. M., & Kimura, M. (1995). Temporal and spatial characteristics of tonicallyactive neurons of the primate’s striatum. Journal of Neurophysiology, 73, 1234–1252.
Asunama, C., Thach, W. T., & Jones, E. G. (1983). Anatomical evidence for segregated focal groupingsof efferent cells and their terminal ramifications in the cerebellothalamic pathway of the monkey.Brain Research Reviews, 5, 267–297.
Babinski, J. (1899). De l’asynergie cerebelleuse. Revue Neurologique, 7, 806–816.
Benecke, R., Rothwell, K. C., Dick, J. P. R., Day, B. L., & Marsden, C. D. (1987) Disturbance ofsequential movements in patients with Parkinson’s disease. Brain, 110, 361–379.
Blakemore, C. (1977). Mechanics of the mind. Cambridge: Cambridge Univ. Press.
Bloedel, J. R. (1992). Functional heterogeneity with structural homogeneity: How does the cerebellumoperate? Behavioral and Brain Sciences, 15, 666–678.
Bloedel, J. R., & Bracha, V. (1997). Duality of cerebellar motor and cognitive functions. In J. D. Schmah-mann (Ed.), The cerebellum and cognition: International review of neurobiology (Vol. 41, pp. 613–630). New York: Academic Press.

208 LAFORCE AND DOYON
Boller, F., Passafiume, D., Keefe, N. C., Rogers, K., Morrow, L., & Kim, Y. (1984). Visuospatial impair-ment in Parkinson’s disease. Archives of Neurology, 41, 485–490.
Bourque, P., & Beaudette, D. (1982). Etude psychometrique du questionnaire de depression de Beckaupres d’un echantillon d’etudiants universitaires francophones. Revue Canadienne des Sciencesdu Comportement, 14, 211–218.
Brotchie, P., Iansek, R., & Horne, M. K. (1991a). Motor function of the monkey globus pallidus. I.Neuronal discharge and parameters of movement. Brain, 114, 1667–1683.
Brotchie, P., Iansek, R., & Horne, M. K. (1991b). Motor function of the monkey globus pallidus. II.Cognitive aspects of movement and phasic neuronal activity. Brain, 114, 1685–1702.
Brown, R. G., & Marsden, C. D. (1990). Cognitive functions in Parkinson’s disease: From descriptionto theory. Trends in Neurosciences, 13, 21–28.
Burton, P. G. (1990). A search for explanation of the brain and learning: Elements of the psychonomicinterface between psychology and neurophysiology. II. Early behavior and its control, the originof consciousness, and the rise of symbolic thought. Psychobiology, 18, 162–194.
Canavan, A. G. M., Passingham, R. E., Marsden, C. D., Quinn, N., Wyke, M., & Polkey,C. E. (1989). Sequencing ability in parkinsonians, patients with frontal lobe lesions, and patientswho have undergone unilateral temporal lobectomies. Neuropsychologia, 27, 787–798.
Dominey, P. F., & Jeannerod, M. (1997). Contribution of frontostriatal function to sequence learningin Parkinson’s disease: Evidence for dissociable systems. NeuroReport, 8, 3–9.
Doyon, J. (1997). Skill learning. In J. D. Schmahmann (Ed.), The cerebellum and cognition: Internationalreview of neurobiology (Vol. 41, pp. 273–294). New York: Academic Press.
Doyon, J., Bourgeois, C., & Bedard, P. (1996a). Deficits visuo-spatiaux associes a la maladie de Parkin-son. Journal International de Psychologie, 31, 161–175.
Doyon, J., Gaudreau, D., Laforce, R. Jr., Castonguay, M., Bedard, P. J., Bedard, F., & Bouchard, G.(1997a). Role of the striatum, cerebellum and frontal lobes in the learning of a visuomotor skill.Brain and Cognition, 34, 218–245.
Doyon, J., Karni, A., Song, A. W., Adams, M. M., Maisog, J. M., Ungerleider, L. G. (1997b). Dynamicchanges in the motor cortical areas, striatum and cerebellum during explicit and implicit learningof a visuomotor skill: A fMRI study. NeuroImage, 5.
Doyon, J., Laforce, R., Jr., Bouchard, G., Gaudreau, D., Roy, J., Poirier, M., Bedard, P. J., Bedard, F., &Bouchard, J.-P. (1998). Role of the striatum, cerebellum and frontal lobes in the automatization ofa repeated visuomotor sequence of movements. Neuropsychologia, 36, 625–641.
Doyon, J., Owen, A. M., Petrides, M., Sziklas, V., & Evans, A. C. (1996b). Functional anatomy ofvisuomotor skill learning examined with positron emission tomography. European Journal ofNeuroscience, 8, 637–648.
Dubois, B., Boller, F., Pillon, B., & Agid, Y. (1991). Cognitive deficits in Parkinson’s disease. In F.Boller & J. Grafman (Eds.), Handbook of neuropsychology (Vol. 5, pp. 195–240). Amsterdam:Elsevier.
Ferraro, R. F., Balota, D. A., & Connor, L. T. (1993). Implicit memory and the formation of new associa-tions in nondemented Parkinson’s disease individuals and individuals with senile dementia of theAlzheimer type: A serial reaction time (SRT) investigation. Brain and Cognition, 21, 163–180.
Flament, D., Ellermann, J. M., Kim, S.-G., Ugurbil, K., & Ebner, T. J. (1996). Functional magneticresonance imaging of cerebellar activation during the learning of a visuomotor dissociation task.Human Brain Mapping, 4, 210–226.
Flourens, P. (1824). Recherches experimentales sur les proprietes et les fonctions du systeme nerveuxdans les animaux vertebres. Paris: Crevot.
Folstein, M. F. (1983). The mini-mental state examination. In T. Crook, S. Ferris, & R. Bartus (Eds.),Assessment in geriatric psychopharmacology (pp. 47–51). Connecticut: Mark Powley Associates.
Georgiou, N., Bradshaw, J. L., Iansek, R., Phillips, J. G., Mattingley, J. B., & Bradshaw, J. A. (1994).Reduction in external cues and movement sequencing in Parkinson’s disease. Journal of Neurology,Neurosurgery, and Psychiatry, 57, 368–370.
Georgiou, N., Bradshaw, J. L., Phillips, J. G., Bradshaw, J. A., & Chiu, E. (1995). Advance informationand movement sequencing in Gilles de la Tourette’s syndrome. Journal of Neurology, Neurosurgery,and Psychiatry, 58, 184–191.
Georgiou, N., Iansek, R., Bradshaw, J. L., Phillips, J. G., Mattingley, J. B., & Bradshaw, J. A. (1993).An evaluation of the role of internal cues in the pathogenesis of parkinsonian hypokinesia. Brain,116, 1575–1587.

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 209
Gilbert, P. F. C., & Thach, W. T. (1977). Purkinje cell activity during motor learning. Brain Research,128, 309–328.
Grafton, S. T., Woods, R. P., & Mike, T. (1994). Functional imaging of procedural motor learning:Relating cerebral blood flow with individual subject performance. Human Brain Mapping, 1, 221–234.
Graybiel, A. M. (1995). Building action repertoires: Memory and learning functions of the basal ganglia.Current Opinion in Neurobiology, 5, 733–741.
Graybiel, A. M., & Kimura, M. (1995). Adaptative neural networks in the basal ganglia. In J. C. Wise,J. C. Davis, & D. G. Beiser (Eds.), Models of information processing in the basal ganglia (pp.103–116). London: MIT Press.
Harrington, D. L., & Haaland, K. Y. (1991). Sequencing in Parkinson’s disease. Brain, 114, 99–115.
Harrington, D. L., York Haaland, K., Yeo, R. A., & Marder, E. (1990). Procedural memory in Parkinson’sdisease: Impaired motor but not visuoperceptual learning. Journal of Clinical and ExperimentalNeuropsychology, 12, 323–339.
Heindel, W. C., Salmon, D. P., Shults, C. W., Walicke, P. A., & Butters, N. (1989). Neuropsychologicalevidence for multiple implicit memory systems: A comparison of Alzheimer’s, Huntington’s, andParkinson’s disease patients. The Journal of Neuroscience, 1989, 582–587.
Hikosaka, O., Rand, M. K., Miyachi, S., & Miyashita, K. (1995). Learning of sequential movements inthe monkey: Process of learning and retention of memory. Journal of Neurophysiology, 74, 1652–1661.
Hoehn, M. M., & Yahr, M. D. (1967). Parkinsonism onset, progression and mortality. Neurology, 17,427–442.
Holmes, G. (1939). The cerebellum of man. Brain, 62, 1–30.
Houk, J. C., & Wise, S. P. (1995). Distributed modular architectures linking basal ganglia, cerebellum,and cerebral cortex: Their role in planning and controlling action. Cerebral Cortex, 2, 95–110.
Inhoff, A. W., & Bisiacchi, P. (1990). Unimanual tapping during concurrent articulation: Examining therole of cortical structures in the execution of programmed movement sequences. Brain and Cogni-tion, 13, 59–76.
Inhoff, A. W., Diener, H. C., Rafal, R. D., & Ivry, R. (1989). The role of cerebellar structures in theexecution of serial movements. Brain, 112, 565–581.
Inhoff, A. W., & Rafal, R. (1990). Cerebellar structures and the programming of movement sequences.Behavioural Neurology, 3, 87–97.
Ito, M. (1982). Cerebellar control of the vestibulo-ocular reflex—Around the flocculus hypothesis. An-nual Review of Neuroscience, 5, 275–296.
Ito, M. (1993). Synaptic plasticity in the cerebellar cortex and its role in motor learning. The CanadianJournal of Neurological Sciences, 20, 70–74.
Jenkins, I. H., Brooks, D. J., Nixon, P. D., Frackowiak, R. S. J., & Passingham, R. E. (1994). Motorsequence learning: A study with positron emission tomography. The Journal of Neuroscience, 14,3775–3790.
Jones, D. L., Phillips, J. G., Bradshaw, J. L., Iansek, R., & Bradshaw, J. A. (1992). Impairment inbilateral alternating movements in Parkinson’s disease? Journal of Neurology, Neurosurgery, andNeuropsychiatry, 55, 503–506.
Jueptner, M., Jenkins, I. H., Brooks, D. J., Frackowiak, R. S. J., & Passingham, R. E. (1996). The sensoryguidance of movement: A comparison of the cerebellum and basal ganglia. Experimental BrainResearch, 112, 462–474.
Jueptner, M., Frith, C. D., Brooks, D. J., Frackowiak, R. S. J., & Passingham, R. E. (1997). The anatomyof motor learning. II. Subcortical structures and learning by trial and error. Journal of Neurophysiol-ogy, 77, 1325–1337.
Jueptner, M., & Weiller, C. (1998). A review of differences between basal ganglia and cerebellar controlof movements as revealed by functional imaging studies. Brain, 121, 1437–1449.
Karni, A. (1996). The acquisition of perceptual and motor skills: A memory system in the adult humancortex. Brain Research and Cognition, 5, 39–48.
Karni, A., Meyer, G., Rey-Hipolito, C., Jezzard, P., Adams, M. M., Turner, R., & Ungerleider, L. G.(1998). The acquisition of skilled motor performance: Fast and slow experience-driven changes inprimary motor cortex. Proceedings of the National Academy of Sciences, 95, 861–868.
Kish, S. J., El-Awar, M., Stuss, D., Nobrega, J., Currier, R., Aita, J. F., Schut, L., Zoghbi. H. Y., &

210 LAFORCE AND DOYON
Freedman, M. (1994). Neuropsychological test performance with dominantly inherited spinocerebel-lar ataxia: Relationship to ataxia severity. Neurology, 44, 1738–1746.
Knowlton, B. J., Mangels, J. A., & Squire, L. R. (1996). A neostriatal habit learning system in humans.Science, 273, 1399–1402.
Leiner, H. C., Leiner, A. L., & Dow, R. S. (1995). The underestimated cerebellum. Human Brain Map-ping, 2, 244–254.
Marsden, C. D., & Obeso, J. A. (1994). The functions of the basal ganglia and the paradox of stereotaxicsurgery in Parkinson’s disease. Brain, 117, 877–897.
McDonald, R., & White, N. M. (1993). A triple dissociation of memory systems: Hippocampus, amyg-dala, and dorsal striatum. Behavioral Neuroscience, 107, 3–22.
Middleton, F. A., & Strick, P. (1994). Anatomical evidence for cerebellar and basal ganglia involvementin higher cognitive function. Science, 266, 458–461.
Moscovitch, M., Vriezen, E., & Goshen-Gottstein, Y. (1993). Implicit tests of memory in patients withfocal lesions or degenerative brain disorders. In F. Boller & J. Grafman (Eds.), Handbook of neuro-psychology (Vol. 8, pp. 133–173). New York: Elsevier.
Nissen, M. J., & Bullemer, P. (1987). Attentional requirements of learning: Evidence from performancemeasures. Cognitive Psychology, 19, 1–32.
Ogden, J. A. (1990). Spatial abilities and deficits in aging and related disorders. In F. Boller & J. Grafman(Eds.), Handbook of neuropsychology (Vol. 4, pp. 265–278). New York: Elsevier.
Packard, M. G., Hirsh, R., & White, N. M. (1989). Differential effects of fornix and caudate nucleuslesions on two radial maze tasks: Evidence for multiple memory systems. The Journal of Neurosci-ence, 9, 1465–1472.
Packard, M. G., & White, N. W. (1990). Lesions of the caudate nucleus selectively impair ‘‘ReferenceMemory’’ acquisition in the radial maze. Behavioral and Neural Biology, 53, 39–50.
Pascual-Leone, A., Grafman, J., Clark, K., Stewart, M., Massaquoi, S., Lou, J.-S., & Hallett, M. (1993).Procedural learning in Parkinson’s disease and cerebellar degeneration. Annals of Neurology, 34,594–602.
Percheron, G., Francois, C., Yelnik, J., Fenelon, G., & Talbi, B. (1993). Neuromorphologie informa-tionnelle du systeme cortico-ponto-cerebello-thalamo-cortical chez le primate. Revue Neurologique,149, 678–691.
Rafal, R. D., Inhoff, A. W., Friedman, J. H., & Bernstein, E. (1987). Programming and execution ofsequential movements in Parkinson’s disease. Journal of Neurology, Neurosurgery, and Psychiatry,50, 1267–1273.
Rauch, S. L., Whalen, P. J., Savage, C. R., Curran, T., Kendrick, A., Brown, H. D., Bush, G., Breiter,H. C., & Rosen, B. R. (1997). Striatal recruitment during an implicit sequence learning task asmeasured by functional magnetic resonance imaging. Human Brain Mapping, 5, 124–132.
Saint-Cyr, J. A., Taylor, A. E., & Lang, A. E. (1988). Procedural learning and neostriatal dysfunctionin man. Brain, 111, 941–959.
Sakai, S. T., Inase, M., & Tanji, J. (1996). Comparison of cerebellothalamic and pallidothalamic projec-tions in the monkey (Macaca fuscata): A double anterograde labeling study. The Journal of Compar-ative Neurology, 368, 215–228.
Sanes, J. N., Dimitrov, B., & Hallett, N. (1990). Motor learning in patients with cerebellar dysfunction.Brain, 13, 103–120.
Schmahmann, J. D. (1996). From movement to thought: Anatomic substrates of the cerebellar contribu-tion to cognitive processing. Human Brain Mapping, 4, 174–198.
Schultz, W., Romo, R., Ljungberg, T., Mirenowicz, J., Hollerman, J. R., & Dickinson, A. (1995). Reward-related signals carried by dopamine neurons. In J. C. Houk, J. C. Davis, & D. G. Beiser (Eds.),Models of information processing in the basal ganglia (pp. 233–248). London: MIT Press.
Seitz, R. J., Roland, P. E., Bohm, C., Greitz, T., & Stone-Elander, S. (1990). Motor learning in man:A positron emission tomography study. NeuroReport, 1, 57–66.
Singh, J., Metz, J., Gabrieli, J. D. E., Willingham, D. B., Dooley, D., Jiang, M., Chen, C.-T., & Cooper,M. (1993). Changes in regional cerebral blood flow associated with a non-patterned proceduralmemory task. Proceeding Abstracts of the Society for Neuroscience, 19, 1001.
Spreen, O., & Strauss, E. (1991). A compendium of neuropsychological tests: Administration, norms,and commentary. New York: Oxford Univ. Press.
Stelmach, G. E., Worringham, C. J., & Strand, E. A. (1987). The programming and execution of move-ment sequences in Parkinson’s disease. International Journal of Neuroscience, 36, 55–65.

STRIATUM VS CEREBELLUM IN MOTOR LEARNING 211
Stern, Y., Mayeux, R., Rosen, J., & Ilson, J. (1983). Perceptual motor dysfunction in Parkinson’s disease:A deficit in sequential and predictive voluntary movement. Journal of Neurology, Neurosurgery,and Psychiatry, 46, 145–151.
Taylor, A. E., Saint-Cyr, J. A., Lang, A. E., & Kenny, F. F. (1986). Parkinson’s disease and depression:A critical reevaluation. Brain, 109, 279–292.
Thach, W. T. (1996). On the specific role of the cerebellum in motor learning and cognition: Clues fromPET activation and lesion studies in man. Behavioral and Brain Sciences, 19, 411–431.
Thach, W. T., Goodkin, H. P., & Keating, J. G. (1992). Cerebellum and the adaptative coordination ofmovement. Annual Review of Neuroscience, 15, 403–442.
Thach, W. T., Mink, J. W., Goodkin, H. P., & Keating, J. G. (2000). Combining versus gating motorprograms: Differential roles for cerebellum and basal ganglia. In M. S. Gazzaniga (Ed.), Cognitiveneuroscience: A reader (pp. 366–375). Oxford: Blackwell.
Wechsler, D. (1987). Wechsler Adult Intelligence Scale—Revised. New York: Psychological Corpora-tion.
Weiss, P., Stelmach, G. E., & Hefter, H. (1997). Programming of a movement sequence in Parkinson’sdisease. Brain, 120, 91–102.
White, N. M. (1989). Reward or reinforcement: What’s the difference? Neuroscience BiobehavioralReview, 13, 181–186.
White, N. M. (1997). Mnemonic functions of the basal ganglia. Current Opinion in Neurobiology, 7,164–169.
Willingham, D. B., & Koroshetz, W. J. (1993). Evidence for dissociable motor skills in Huntington’sdisease patients. Psychobiology, 21, 173–182.
Willingham, D. B. (1998). A neuropsychological theory of motor skill learning. Psychological Review,105, 558–584.
Wise, S., & Houk, J. C. (1994). Modular neuronal architecture for planning and controlling motor behav-ior. Biological Communications of the Danish Royal Academy of Sciences, 43, 21–33.
Yamamoto, T., Yoshida, K., Yoshikawa, H., Kishimoto, Y., & Oka, H. (1992). The medial dorsal nucleusis one of the thalamic relays of the cerebellocerebral response to the frontal association cortex inthe monkey: Horseradish peroxidase and fluorescent dye double staining study. Brain Research,579, 315–320.