discrimination of geographical song variants by darwin’s finches · discrimination of...
TRANSCRIPT

ANIMAL BEHAVIOUR, 2007, 73, 833e844doi:10.1016/j.anbehav.2006.11.001
Discrimination of geographical song variants by
Darwin’s finches
JEFFREY PODOS
Department of Biology, Graduate Program in Organismic and Evolutionary Biology,
University of Massachusetts, Amherst
(Received 27 April 2006; initial acceptance 19 June 2006;
final acceptance 7 November 2006; published online 26 March 2007; MS. number: A10432)
Geographical variation in the structure of mating signals can influence the processes of mate recognitionand reproductive isolation. In Darwin’s finches of the Galapagos Islands, Ecuador, prior studies of geo-graphical variation in vocal mating signals (songs) have focused on populations on small islands, on whichthere is presumably little opportunity for the evolution of within-island song variation or discriminationabilities. The situation on larger islands, however, is less clear. I characterized aspects of song variation anddiscrimination in the medium ground finch, Geospiza fortis, in the southeastern coastal region of the largeisland of Santa Cruz. Songs from two sites, Bahia Academia and El Garrapatero, separated by approximately11 km, differed in mean minimum frequency and in a corresponding principal component factor, andcould be distinguished with moderate success in discriminant function analysis. Territorial males at El Gar-rapatero showed strong discrimination of songs by site: in response to playback of local songs, birdsshowed higher rates of singing and flying, and significantly closer approaches to the playback speaker. Suc-cessful discrimination of songs by site, in the face of overlap in song structure between these sites, suggeststhat these birds rely on an exemplar-based mechanism of song perception. Moreover, the results supportthe hypothesis that birds in these populations are relatively philopatric, and thus suggest an opportunityfor within-island evolutionary divergence.
2007 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd.
Keywords: Darwin’s finches; geographical variation; Geospiza fortis; medium ground finch; passerine; reproductiveisolation; song acoustics; within-island evolutionary divergence; vocal communication
The evolutionary diversification of many animal taxa isfacilitated by geographical variation in mating signals(West-Eberhard 1983; Wells & Henry 1998; Foster 1999).Divergence in mating signals among localities tends tolimit interpopulation mating, and thus promotes repro-ductive isolation and within-species divergence (Ptacek2000; Irwin et al. 2001; MacDougall-Shackleton &MacDougall-Shackleton 2001; Slabbekoorn & Smith 2002;Lachlan & Servedio 2004). The evolutionary impact ofgeographical variation in signal structure depends criti-cally on patterns of signal perception and discrimination.If animals discriminate among geographical signal varia-tions, then barriers to interpopulation gene flow can beintroduced (Baker et al. 1987b; Wells & Henry 1998;Irwin 2000; Searcy et al. 2002). By contrast, a lack of
Correspondence: J. Podos, Department of Biology, University of Massa-chusetts, Amherst, MA 01003, U.S.A. (email: [email protected]).
80003e3472/07/$30.00/0 2007 The
discrimination among geographical signal variants fosterscontinued gene flow across localities. A comprehensiveunderstanding of the influence of mating signals in repro-ductive isolation thus requires attention to both matingsignal structure and signal discrimination abilities.
Key insights into the origins and consequences ofgeographical variation in mating signals have come fromstudies of birds (e.g. Marler & Tamura 1964; Baptista 1975;Slater 1986; Slabbekoorn & Smith 2002; Nelson & Soha2004; reviewed by Edwards et al. 2005). Geographicalvariation in bird songs evolves as a result of numerous fac-tors, including physical and genetic isolation among pop-ulations, limited dispersal, and divergent trajectories ofsexual selection, natural selection or drift among popula-tions (Nelson & Marler 1990; Podos et al. 2004; Podos &Warren, in press). In oscine songbirds, in which songsare learned, geographical variation in song structure alsoresults from the accumulation of copying inaccuraciesduring cultural transmission, with distinct memes taking
33Association for the Study of Animal Behaviour. Published by Elsevier Ltd.

ANIMAL BEHAVIOUR, 73, 5834
root in partially isolated populations (Marler & Tamura1964; Krebs & Kroodsma 1980; Slater 1989). Songbirdsof some species discriminate local and foreign song types,suggesting opportunities for interpopulation genetic di-vergence (Baker et al. 1987b; Slabbekoorn & Smith 2002).
I characterized aspects of geographical variation in song,and tested for song discrimination by site, in a species ofDarwin’s finch from the Galapagos Islands. Darwin’sfinches have been a central model system in studies ofpopulation evolution and dynamics (Grant 1999; Grant &Grant 2002c). Song is a key factor in territoriality, mate se-lection and hybridization in these birds (Grant & Grant1997, 1998). In a series of playback studies, Ratcliffe &Grant (1985) and Grant & Grant (2002b) found that terri-torial male Geospiza finches responded more strongly tolocal song types than they did to conspecific song typesfrom other islands. Greater responses to local songs wereattributed to geographical isolation and a resulting lackof familiarity with foreign song types (Ratcliffe & Grant1985). These studies were conducted on the relativelysmall islands of Daphne Major and South Plaza, and onthe medium-sized islands of Genovesa and Pinta. All localsong types were presumably familiar to most if not all is-land residents. Moreover, low interisland migration ratesvirtually assured that playback songs from other islandswere unfamiliar. Together these interisland playback stud-ies support an allopatric model of finch divergence andspeciation (Grant & Grant 2002a, 2003).
The situation may be more complicated, however, forfinch populations on larger Galapagos islands such asIsabela, Fernandina, San Cristobal and Santa Cruz. Theseislands presumably could support geographical variationin song structure. Snodgrass & Heller (1904) described mi-nor differences in Geospiza fortis songs among the IsabelaIsland sites of Tagus Cove, Tagus Hill and Iguana Cove.Bowman (1979) and Ratcliffe (1981) described elevationalgradients in song frequency parameters for, respectively,Geospiza fuliginosa of Pinta Island and Camarhynchus par-vulus of Santa Cruz Island. The presence of geographicallydistinct variants would suggest limited recent dispersaltendencies among the populations in question, whereasa lack of geographically distinct variants would suggestwide dispersal and comparatively wide song meme distri-bution (Kroodsma et al. 1985). Similarly, greater responsesto local versus foreign song types would suggest successfuldiscrimination abilities and biased familiarity with localtypes, and thus some degree of philopatry. These factors(dispersal tendencies, geographical variation in matingsignals and signal discrimination abilities) bear upon thepossibility of within-island divergence in Darwin’sfinches, and thus merit closer attention.
The first goal of this study was to quantify possiblestructural variation in songs of the medium ground finch,G. fortis, between two sites, El Garrapatero and BahiaAcademia, separated by approximately 11 km along thesoutheast coastal region of Santa Cruz Island. Birds atboth sites express similar ranges of variation in beak andbody size, yet differ in distributions of these parameters.In particular, birds at the Bahia Academia site show fairlycontinuous variation in beak and body size, whereas birdsat the El Garrapatero site show bimodal variation in these
parameters (Hendry et al. 2006; Huber & Podos 2006),with small- and large-male ‘morphs’ producing songswith distinctive acoustic features (Huber & Podos 2006).At both sites, variation in some song features correspondswith variation in beak morphology, in a manner consis-tent with a hypothesis of morphological constraints onsong production and evolution (Podos 2001; Podos &Nowicki 2004; Huber & Podos 2006). I quantified differ-ences in song features at both sites to determine whetherand how songs vary between sites.
The second goal of this study was to test whether songdifferences that might occur between these sites arefunctionally significant. Towards this end, I conducteda playback experiment to test the responses of territorialmales at one site, El Garrapatero, to playback of both localsongs and songs recorded at Bahia Academia. The re-ciprocal experiment (i.e. playback of songs from both sitesto males at Bahia Academia) would have been additionallyinstructive but was not feasible because of logisticallimitations (see Methods). In simulated territorial intru-sion tests, high levels of response to song playback aregenerally regarded as evidence that songs are functionallysignificant (i.e. meaningful to the birds in the context ofsocial interactions; Falls 1992; Collins 2004). In particular,I tested an a priori hypothesis that birds would respondmore strongly to local songs from their site (Lemon1967; McGregor 1983; Kroodsma et al. 1985; Baker et al.1987b). A positive result would implicate learning andphilopatry as factors in the evolution of G. fortis song pref-erences and discrimination abilities (Kroodsma et al. 1985;MacDougall-Shackleton & MacDougall-Shackleton 2001),and would also suggest a possible opportunity forwithin-island divergence among populations. A lack ofdiscrimination among geographical song variants, onthe other hand, would be more difficult to interpret butcould result from a combination of factors, includinga lack of discernable site-specific vocal features, low philo-patry, late learning of song preferences, historical inertiain song preferences or weak selection on precise song rec-ognition abilities (Grant & Grant 2002a).
METHODS
Geographical Song Variation
Songs of birds were recorded during February andMarch 1999 and 2001e2004. All songs analysed in thisstudy were from finches that had been captured andbanded with unique combinations of one metal andthree colour bands. Song recordings were made at twolocalities separated by approximately 11 km along thesoutheast coastal region of Santa Cruz Island: Bahia Aca-demia (GPS coordinates 00�4401500e3000 S, 90�1800500e0900
W; N ¼ 18 birds) and El Garrapatero (GPS coordinates00�4002000e4102000 S, 90�1301000e1404000 W; N ¼ 39 birds).Both of these sites are in the arid coastal zone and featuresimilar vegetation, including Opuntia echios, Acacia ma-cracantha, Cordia lutea and Bursera graveolens. This habitatoccurs continuously between the two sites, and G. fortishave been captured at regular intervals along a 4-km

PODOS: DISCRIMINATION OF SONG BY DARWIN’S FINCHES 835
transect extending from Bahia Academia to El Garrapa-tero (A. Gabela & J. Podos, unpublished data). I focusedon finches at El Garrapatero for two reasons. First, birdsat this site tend to be more accessible than those at BahiaAcademia, in part because of a higher density of intro-duced goats and donkeys that have produced a networkof trails. Second, an ongoing study of the population atEl Garrapatero provided necessary background data onthe identity, territory location, territory boundaries andpairing status of potential playback subjects. No suchbackground data were available for the population at Ba-hia Academia at the time of this study. The compositionof background sounds, although not quantified in thisstudy, appeared to be similar between sites. Recordings offinch songs were made using Sony TCD-5M stereo-cassetterecorders and directional (K6/ME66 or MKH70) or omnidi-rectional (K6/ME62 or MKH20) microphones. MKH micro-phones were powered with Sennheiser MZA14 phantompower supplies; omnidirectional microphones weremounted in 22-inch (55.9-cm) diameter Telinga Pro Uni-versal parabolas.
Each male that was recorded sang only a single songtype, as is typical for Darwin’s finches (Bowman 1983;Grant 1999). Up to 10 (mean ¼ 4.5) of the highest-qualitysong recordings of each bird were identified and digitized,using a DART PCMCIA digitizing card and DartDisk 1.0(Beeman 2002). Four acoustic features were measuredfrom each song using Signal 4.0 sound analysis software(Beeman 2002): duration and trill rate were measured (tothe nearest ms) from oscillograms and spectrograms usingan on-screen cursor. Minimum frequency and maximumfrequency were measured from amplitude spectra, bothdetermined at �24 dB relative to peak frequencies(32 000 points fast Fourier transform, FFT, length,smoothed to 100 Hz, the effective frequency resolutionof the analysis; Podos 1997, 2001). A fifth acoustic feature,frequency bandwidth, was calculated as the difference be-tween minimum and maximum frequencies. Values forthe five acoustic features were averaged across multiplesongs for each bird.
I used three approaches to examine differences in songstructure between sites. First, I assessed between-sitevariation in the five song features to determine whetherbasic timing or frequency features reliably distinguishedsongs by site. Values for each of the five acoustic featureswere compared across sites using ManneWhitney U testsin JMP (2002). To account for possible intervariable corre-lations, I also conducted principal component analysis(PCA) on the correlation matrix of these five song featuresand compared PCA factors between sites using ManneWhitney U tests. I assessed the overall statistical signifi-cance of the intersite comparison, based on P values oftests on PC factors only, using the Bernoulli equation(Moran 2003). I did not conduct a similar test on theraw song features because of nonindependence amongsome of these features (Moran 2003). The statistical signif-icance of each song feature and PC factor was assessedwith respect to P value thresholds corrected for multiplecomparisons (Bonferroni method).
Second, I conducted discriminant function analyses(DFA), first on the five raw acoustic features and then on
the first three PC factors, using JMP (2002), to determinewhether any of the input variables allowed correct classi-fication of song exemplars to locality (Kachigan 1991),and to provide an overall view of the feasibility of clas-sifying songs to locality based on the selected combi-nation of acoustic features. The magnitude of anybetween-site song differences is presumably reflected inthe accuracy with which songs can be classified correctlyto site.
Third, I compared the fine-scale acoustic structure ofsong notes both within and between sites using a spectro-gram cross-correlation method (Beeman 2002). Thismethod calculates the structural similarity of any givenpair of notes, in terms of note duration, absolute fre-quency and dynamic changes in frequency and amplitude(Clark et al. 1987). Notes are defined as continuous traceson spectrograms, separated by a minimum of 10 ms.Cross-correlation scores near zero indicate low similarityand suggest that the two notes being compared vary con-siderably in their fine-scale structure, whereas high cross-correlation scores indicate greater similarity in fine-scalestructure between notes. Higher similarity scores withinsites versus between sites would support the presence ofgeographical variation in note structure.
Cross-correlation analyses were conducted as follows. Iextracted a single note from the highest-quality record-ings of each of the 57 songs for this analysis (N ¼ 18 atBahia Academia; N ¼ 39 at El Garrapatero). All songs inthe sample consisted of two or more notes (Fig. 1). Acrossthe sample, I chose the second note for analysis (in songswith three or more notes, the introductory note is oftendistinctive from remaining notes; the second note thuscan be viewed as a more typical note in such songs;personal observation). Comparisons of spectrograms(FFT ¼ 256 points, frequency range 0e10 kHz) of allnote pairs across the sample generated a similarity matrixof 1596 cells, comprising 874 within-site comparisonsand 722 between-site comparisons. Differences in notesimilarity scores for the three types of comparisons(within Bahia Academia, within El Garrapatero, andbetween sites) were assessed using one-way ANOVA( JMP 2002).
Numerous prior studies of vocal geographical variation,especially in species with small repertoire sizes, haveexamined the proportion of distinct song types producedat different localities (e.g. Slater et al. 1984; Baker &Thompson 1985). Such an approach is not feasible forG. fortis of Santa Cruz Island, however, because of the diffi-culty of reliably assigning songs to type. Some birds do in-deed produce songs with similar overall structure. However,putative types often grade into each other, and many birdsproduce songs with unique features (e.g. Fig. 1). The scat-tered nature of data points in a feature-by-feature structuralanalysis (see Results, Fig. 2) supports this impression.
Playback Study
SubjectsPlayback trials were conducted with nine male G. fortis
at El Garrapatero, between 1 and 20 February 2004. Prior

ANIMAL BEHAVIOUR, 73, 5836
Bahia Academia
Time (s)0
8
Freq
uen
cy (
kHz)
EI Garrapatero Subjects
a
b
c
d
e
f f
g
h
i
a
b
c
d
e
g
h
i
A
B
C
D
E
F
G
H
I
Figure 1. Sound spectrograms of the nine G. fortis songs from Bahia Academia and El Garrapatero that were used as playback stimuli and in-cluded in the analyses of geographical variation in song structure. Also shown are sound spectrograms of songs from the nine subjects for
which playback trials were completed. Subjects A, E, F and H were large-morph birds, and subjects B, C, D, G and I were small-morph birds.
Each bird received the two corresponding songs as playback stimuli (e.g. subject ‘A’ received the two ‘a’ stimuli).
observations indicated that territorial behaviour in groundfinches at this site is associated most directly with defenceof nests and nesting locations, and not with defence offood resources. Furthermore, prior observations suggestedthat territorial behaviour is strongest in males that arepaired with a female. For these reasons, I chose as playbacksubjects males that had nests and that were observed to beactively paired with a female (i.e. engaged in courtship,mating, or parental behaviour). Confirmation of ‘paired’status for playback subjects was based on observations ofcourtship feeding, back-and-forth calling, copulation and
incubation (S. K. Huber & J. Podos, unpublishedobservations).
Playback stimuliI selected a subset of nine songs of high quality from
each locality that provided a good sample of the diversityof songs. In addition, I limited my selection of songs fromEl Garrapatero to those of only one morph, the large-malemorph, to simplify the experimental design and tofacilitate preparation of playback stimuli. Males might be
PODOS: DISCRIMINATION OF SONG BY DARWIN’S FINCHES 837
expected to respond more strongly to songs of their ownmorphs, given the presence of acoustic differences bymorph in this population (Huber & Podos 2006), and ourobservation that males tend to engage in territorial dis-putes more often with males from their own morphthan with males of the other morph. In the context ofthe hypothesis being tested here, which is that local songsshould elicit greater responses than ‘foreign’ songs, any re-duction in response of small males to the playback of locallarge-male songs would decrease the likelihood of detect-ing a site-specific bias in responsiveness. Thus, the deci-sion to use songs from only one morph was conservativewith respect to the hypothesis being tested. No subjectswere tested with their own songs.
I prepared 18 playback tapes, one for each of thestimulus songs, using Signal 4.0 (Beeman 2002). Eachtape consisted of 18 repetitions of the song played every10 s for 3 min. Stimulus sets (one local song, one foreignsong) were assigned at random to playback birds. Playbacksongs were standardized for amplitude and were high-passfiltered (above 1 kHz) to reduce background noise(Beeman 2002).
2(a)
(b)
1
0
–1
–2
–3
2.5
1.5
0.5
–0.5
–1.5
–2.5
–2.5 –1.5 –0.5 0.5 1.5
PC1
PC3
PC2
2.5 3.5 4.5
–3.5
–4.5
A,g
b
f
I
g i
g
e
acb
f
h
D
G
i
Be
Ea
Fh
H
dc
c
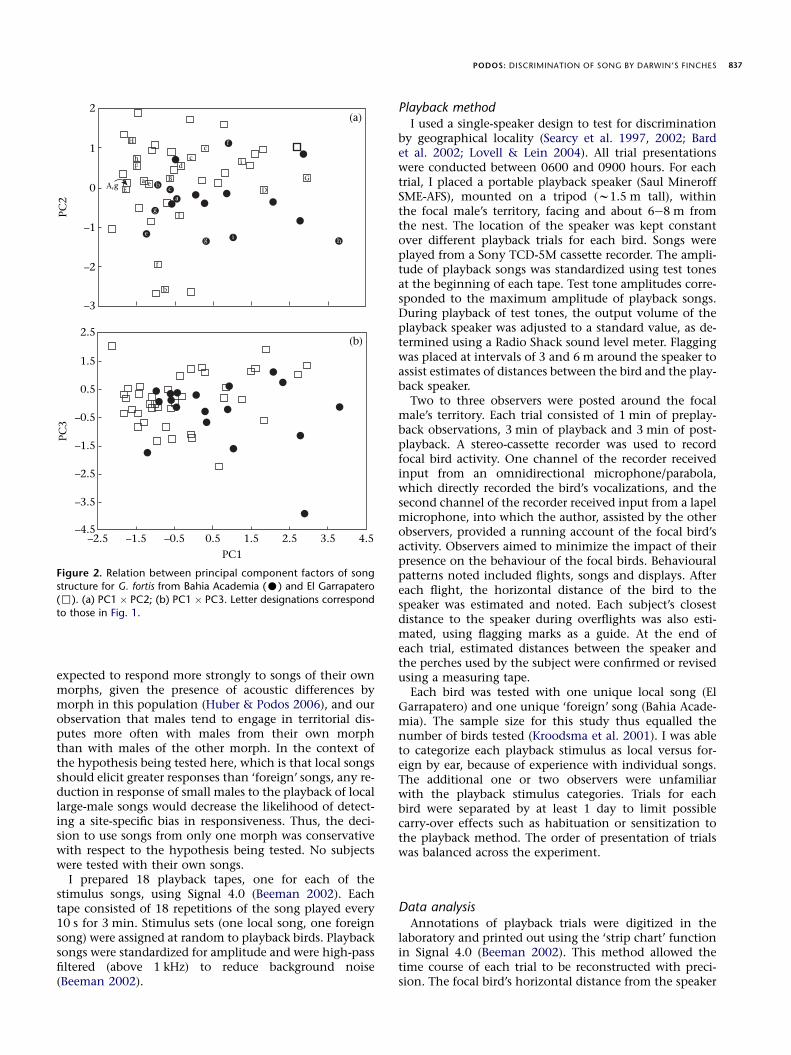
Figure 2. Relation between principal component factors of songstructure for G. fortis from Bahia Academia (C) and El Garrapatero
(,). (a) PC1 � PC2; (b) PC1 � PC3. Letter designations correspond
to those in Fig. 1.
Playback methodI used a single-speaker design to test for discrimination
by geographical locality (Searcy et al. 1997, 2002; Bardet al. 2002; Lovell & Lein 2004). All trial presentationswere conducted between 0600 and 0900 hours. For eachtrial, I placed a portable playback speaker (Saul MineroffSME-AFS), mounted on a tripod (w1.5 m tall), withinthe focal male’s territory, facing and about 6e8 m fromthe nest. The location of the speaker was kept constantover different playback trials for each bird. Songs wereplayed from a Sony TCD-5M cassette recorder. The ampli-tude of playback songs was standardized using test tonesat the beginning of each tape. Test tone amplitudes corre-sponded to the maximum amplitude of playback songs.During playback of test tones, the output volume of theplayback speaker was adjusted to a standard value, as de-termined using a Radio Shack sound level meter. Flaggingwas placed at intervals of 3 and 6 m around the speaker toassist estimates of distances between the bird and the play-back speaker.
Two to three observers were posted around the focalmale’s territory. Each trial consisted of 1 min of preplay-back observations, 3 min of playback and 3 min of post-playback. A stereo-cassette recorder was used to recordfocal bird activity. One channel of the recorder receivedinput from an omnidirectional microphone/parabola,which directly recorded the bird’s vocalizations, and thesecond channel of the recorder received input from a lapelmicrophone, into which the author, assisted by the otherobservers, provided a running account of the focal bird’sactivity. Observers aimed to minimize the impact of theirpresence on the behaviour of the focal birds. Behaviouralpatterns noted included flights, songs and displays. Aftereach flight, the horizontal distance of the bird to thespeaker was estimated and noted. Each subject’s closestdistance to the speaker during overflights was also esti-mated, using flagging marks as a guide. At the end ofeach trial, estimated distances between the speaker andthe perches used by the subject were confirmed or revisedusing a measuring tape.
Each bird was tested with one unique local song (ElGarrapatero) and one unique ‘foreign’ song (Bahia Acade-mia). The sample size for this study thus equalled thenumber of birds tested (Kroodsma et al. 2001). I was ableto categorize each playback stimulus as local versus for-eign by ear, because of experience with individual songs.The additional one or two observers were unfamiliarwith the playback stimulus categories. Trials for eachbird were separated by at least 1 day to limit possiblecarry-over effects such as habituation or sensitization tothe playback method. The order of presentation of trialswas balanced across the experiment.
Data analysisAnnotations of playback trials were digitized in the
laboratory and printed out using the ‘strip chart’ functionin Signal 4.0 (Beeman 2002). This method allowed thetime course of each trial to be reconstructed with preci-sion. The focal bird’s horizontal distance from the speaker

ANIMAL BEHAVIOUR, 73, 5838
was tabulated on a per-second basis across the entire trial.The onset of songs and flights was also noted.
I tallied the following four dependent variables as theyoccurred during preplayback, playback and postplaybacksegments of trials: (1) flight rate (flights/min), (2) song rate(songs/min), (3) closest approach to speaker, based onlyon perched locations, and (4) closest approach to speaker,based on perched locations and estimated distancesduring overflights of the speaker. For trial segments thatdid not include overflights, values for (3) and (4) wereidentical. Within the playback trial, I also calculated (5)latency (s) to first flight, and (6) latency (s) to first song.Overall response levels were calculated using principalcomponent analysis, PCA (JMP 2002). Response parame-ters were compared between stimuli (local versus foreign),using Wilcoxon two-tailed matched-pairs signed-rankstests, on pre-, post- and playback trial segments separately.To determine which specific response parameters variedby locality, I compared raw P values to P value thresholdscorrected for multiple comparisons (Bonferroni method).
RESULTS
Geographical Song Variation
Songs from the two localities did not differ in trill rate,duration, maximum frequency or frequency bandwidth(Table 1), but songs from Bahia Academia had significantlylower minimum frequencies than those from EI Garrapa-tero. Differences in song structure between localitieswere also evident in the PCA results; loadings for PC2 dif-fered significantly between sites, according to Bonferronicorrected P values (Table 1). The probability of two ofthree PC factors differing across sites (uncorrected Pvalues) by chance is approximately 0.0086 (Bernoulliequation; Moran 2003). PC1 was best represented by du-ration and maximum frequency, and PC2 by trill rateand minimum frequency (Table 2). Songs from thetwo localities showed considerable overlap in plots ofPC1 � PC2 and of PC1 � PC3 (Fig. 2).
Discriminant function analyses (DFA) on the raw songvariables classified 12 Bahia Academia songs correctly tosite, but misclassified an additional six Bahia Academiasongs to El Garrapatero. Furthermore, 33 El Garrapatero
songs were correctly classified to site, but an additional sixsongs were misclassified to Bahia Academia. The DFA onPC factors was slightly less accurate, misclassifying thesame songs as in the raw song variable DFA, but alsomisclassifying an additional two songs from Bahia Acade-mia and an additional three songs from El Garrapatero.
Spectrogram cross-correlation scores differed signifi-cantly between the three types of comparisons (X� SD:within Bahia Academia ¼ 0.452 � 0.102; within El Garr-apatero ¼ 0.404 � 0.107; between sites ¼ 0.402 � 0.105;F2,1593 ¼ 16.393, P < 0.0001). Post hoc tests revealed thatcross-correlation scores were similar for comparisonswithin El Garrapatero and between sites (Tukey test:P > 0.05), but higher for comparisons within Bahia Acade-mia (Tukey tests: P < 0.001).
Playback Study
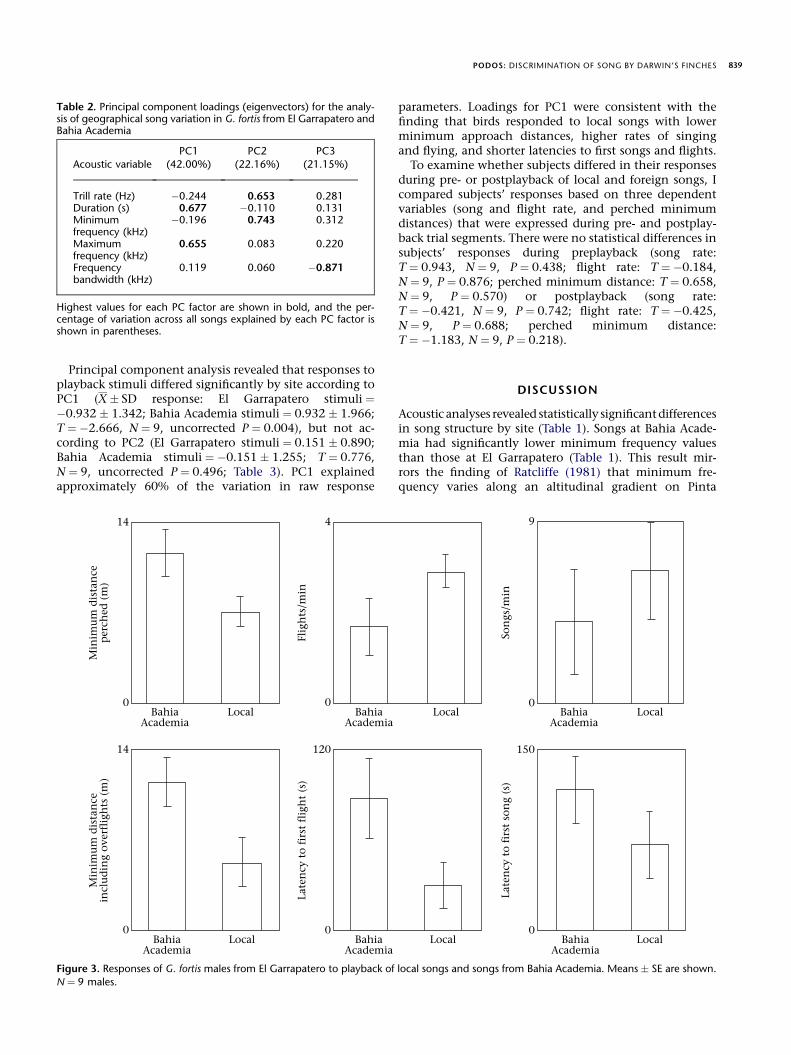
During song playback, birds at El Garrapatero respondedmore strongly to local songs than to foreign songs fromBahia Academia, for all six raw response variables exam-ined (Fig. 3). Birds flew significantly closer to the speakerduring playback of local song than they did during play-back of foreign song when overflights were included(Wilcoxon matched-pairs signed-ranks test: T ¼ �2.524,N ¼ 9, uncorrected P ¼ 0.008, significance retained afterBonferroni correction), but not when overflights were ex-cluded (perched distances: T ¼ �2.316, N ¼ 9, uncorrectedP ¼ 0.016). Differences in the birds’ responses to local andforeign songs for the other four variables showed trends inthe same direction, although none was significant afterBonferroni correction. Birds flew more often duringplayback of local song (T ¼ 1.622, N ¼ 9, uncorrectedP ¼ 0.055), and showed a shorter latency to first flight af-ter playback onset (T ¼ �1.365, N ¼ 9, uncorrectedP ¼ 0.102). Similar results were found for song rates(T ¼ 1.866, N ¼ 9, uncorrected P ¼ 0.039) and latency tofirst song (T ¼ �1.521, N ¼ 9, uncorrected P ¼ 0.078).The level of response to local versus foreign playbacksongs did not differ between small-morph subjects(N ¼ 5, birds B, C, D, G and I; Fig. 1) and large-morph sub-jects (N ¼ 4, birds A, E, F and H; Fig. 1; all ManneWhitneyU values < 1.15, all P > 0.26).
Table 1. Descriptive statistics (means � SD) for acoustic features and principal components (PC1ePC3) of G. fortis song from two sites, ElGarrapatero and Bahia Academia
Acoustic feature
Locality Between-site comparison
El Garrapatero (N¼39) Bahia Academia (N¼18) ManneWhitney U score Uncorrected P value
Trill rate (Hz) 2.29�0.61 2.06�0.60 �0.876 0.381Duration (s) 0.84�0.14 0.88�0.24 �0.241 0.810Minimum frequency (kHz) 2.21�0.23 1.92�0.24 �4.080 0.001*Maximum frequency (kHz) 4.27�1.02 4.63�0.91 1.743 0.081Frequency bandwidth (kHz) 2.07�0.99 2.71�1.04 2.258 0.024PC1 �0.31�1.32 0.66�1.52 2.481 0.013PC2 0.18�1.13 �0.39�0.76 �2.721 0.007*PC3 0.14�0.93 �0.31�1.18 �1.039 0.299
*Statistically significant at P < 0.05 after Bonferroni correction.
PODOS: DISCRIMINATION OF SONG BY DARWIN’S FINCHES 839
Principal component analysis revealed that responses toplayback stimuli differed significantly by site according toPC1 (X� SD response: El Garrapatero stimuli ¼�0.932 � 1.342; Bahia Academia stimuli ¼ 0.932 � 1.966;T ¼ �2.666, N ¼ 9, uncorrected P ¼ 0.004), but not ac-cording to PC2 (El Garrapatero stimuli ¼ 0.151 � 0.890;Bahia Academia stimuli ¼ �0.151 � 1.255; T ¼ 0.776,N ¼ 9, uncorrected P ¼ 0.496; Table 3). PC1 explainedapproximately 60% of the variation in raw response
Table 2. Principal component loadings (eigenvectors) for the analy-sis of geographical song variation in G. fortis from El Garrapatero andBahia Academia
Acoustic variable
PC1
(42.00%)
PC2
(22.16%)
PC3
(21.15%)
Trill rate (Hz) �0.244 0.653 0.281Duration (s) 0.677 �0.110 0.131Minimumfrequency (kHz)
�0.196 0.743 0.312
Maximumfrequency (kHz)
0.655 0.083 0.220
Frequencybandwidth (kHz)
0.119 0.060 �0.871
Highest values for each PC factor are shown in bold, and the per-centage of variation across all songs explained by each PC factor isshown in parentheses.
parameters. Loadings for PC1 were consistent with thefinding that birds responded to local songs with lowerminimum approach distances, higher rates of singingand flying, and shorter latencies to first songs and flights.
To examine whether subjects differed in their responsesduring pre- or postplayback of local and foreign songs, Icompared subjects’ responses based on three dependentvariables (song and flight rate, and perched minimumdistances) that were expressed during pre- and postplay-back trial segments. There were no statistical differences insubjects’ responses during preplayback (song rate:T ¼ 0.943, N ¼ 9, P ¼ 0.438; flight rate: T ¼ �0.184,N ¼ 9, P ¼ 0.876; perched minimum distance: T ¼ 0.658,N ¼ 9, P ¼ 0.570) or postplayback (song rate:T ¼ �0.421, N ¼ 9, P ¼ 0.742; flight rate: T ¼ �0.425,N ¼ 9, P ¼ 0.688; perched minimum distance:T ¼ �1.183, N ¼ 9, P ¼ 0.218).
DISCUSSION
Acoustic analyses revealed statistically significant differencesin song structure by site (Table 1). Songs at Bahia Acade-mia had significantly lower minimum frequency valuesthan those at El Garrapatero (Table 1). This result mir-rors the finding of Ratcliffe (1981) that minimum fre-quency varies along an altitudinal gradient on Pinta
0Bahia
AcademiaBahia
AcademiaBahia
Academia
BahiaAcademia
BahiaAcademia
BahiaAcademia
Local Local Local
LocalLocalLocal
Min
imu
m d
ista
nce
per
ched
(m
)M
inim
um
dis
tan
cein
clu
din
g ov
erfl
igh
ts (
m)
Flig
hts
/min
Son
gs/m
inLa
ten
cy t
o fi
rst
son
g (s
)
Late
ncy
to
firs
t fl
igh
t (s
)
14
0
4
0
9
0
150
0
12014
0
Figure 3. Responses of G. fortis males from El Garrapatero to playback of local songs and songs from Bahia Academia. Means � SE are shown.
N ¼ 9 males.

ANIMAL BEHAVIOUR, 73, 5840
Island for G. fuliginosa. Ratcliffe (1981) suggested thatthe G. fuliginosa song gradient might result from selec-tion for lower-frequency songs at higher elevations onPinta, where vegetation is more dense, because songtransmission in dense acoustic environments tends tobe optimized at lower frequencies (e.g. Hunter & Krebs1979; Slabbekoorn & Smith 2002). However, this expla-nation seems unlikely to account for differences in songstructure between Bahia Academia and EI Garrapatero,given the similarity in vegetation between the two sites.
The DFA showed moderate success in classifying songsto site (78.9% for raw acoustic parameters and 70.2% forPC factors as input variables). It therefore seems that birdscould reliably distinguish a majority of songs from the twosites based on the combination of song features examined.This interpretation is complemented in part by the resultsof spectrogram cross-correlation analyses. Songs from oneof the sites, Bahia Academia, showed higher similarity innote structure to other Bahia Academia songs than tosongs from El Garrapatero. This indicates that birds atBahia Academia tend to share songs with comparativelysimilar note structures. This difference in similarity scoreswas minor, however, and notes at El Garrapatero were nomore similar to each other than they were to notes fromBahia Academia.
Songs from both sites showed broad overlap in raw andPC song parameters (Fig. 2, Table 1). Structural overlap insongs between the two sites can be explained in part bythe wide diversity of song variants at each site (Fig. 1;see also Bowman 1983). In species with sharp geographi-cal differences in song between sites, such as white-crowned sparrows, Zonotrichia leucophrys, and cornbuntings, Miliaria calandra, individuals normally produceonly a single song type, and tend to share the same dis-tinctive type with their neighbours (Baptista 1975; McGre-gor 1983). For example, the trills of white-crownedsparrows in Berkeley, California, U.S.A., contain fewernotes of descending frequencies than do trills of othernearby populations (Marler & Tamura 1962). Limitedwithin-site variation invariably enhances between-sitecontrasts, thus making geographical variants easier to de-tect, for both bird and ornithologist. In contrast, for Geo-spiza fortis of southeastern Santa Cruz Island, the widerange of song patterns found at each site enhances struc-tural overlap of songs between sites. Diversity in songstructure for Santa Cruz G. fortis also appears to be
Table 3. Principal component loadings (eigenvectors) for playbackresponse parameters
Response parameter PC1 (59.79%) PC2 (18.97%)
Minimum distance (perched) 0.466 �0.238Minimum distance(with overflights)
0.486 �0.181
Flights/min �0.388 0.469Latency to first flight 0.376 �0.080Songs/min �0.379 �0.512Latency to first song 0.334 0.650
The percentage of variation across all songs explained by each PCfactor is shown in parentheses.
correlated with the wide range of beak morphologies pres-ent, in a manner consistent with a vocal tract resonancemodel of song production and evolution (Podos 2001;Huber & Podos 2006).
Territorial males at El Garrapatero responded morestrongly to playback of local songs than to playback ofBahia Academia songs, particularly in terms of the closestapproach to the playback speaker (Fig. 3). Distance ofapproach is generally viewed as a reliable measure ofstrength of response to playback. The validity of the play-back results is reinforced by comparison to prior playbackstudies with Geospiza finches (Ratcliffe 1981; Ratcliffe &Grant 1985; Grant & Grant 2002b). Birds in these studiesand in the present study showed similar responses to play-back in terms of flight and song rates, flight and song la-tencies, and nearest approach. Geospiza fortis males at ElGarrapatero thus appear to show strong song discrimina-tion by site despite the similarities in song structure be-tween the two sites. This result is striking given that thetwo sites are separated by a mere 11 km. Prior demonstra-tions of differential response by locality have been reportedfor populations separatedby greater distances (e.g. w40 km:McGregor 1983; w540 km: Searcy et al. 2002), with the ex-ception of the study by Leader et al. (2002) on orange-tuftedsunbirds, Nectarinia osea, which showed discrimination ofsongs from adjacent localities in an urban setting. To myknowledge, this is the first demonstration of discriminationof within-island geographical song variants.
Mechanism of Song Discrimination
A number of playback studies have shown that Darwin’sfinches discriminate between conspecific and sympatricheterospecific songs (Lack 1945; Ratcliffe 1981; Bowman1983; Ratcliffe & Grant 1985; Grant 1999). In one suchstudy, Ratcliffe & Grant (1985) found that South Plaza Is-land G. fortis and G. fuliginosa expressed strong species-level discrimination in song playback, although songs ofthe two species were statistically indistinguishable. To ex-plain this apparent paradox, Ratcliffe & Grant (1985, page303) hypothesized that these two species successfully dis-criminate each other’s songs using ‘subtle (as yet uniden-tified) common cues’ (see also Grant 1999). A similarexplanation might apply to the results of the presentstudy. Songs from El Garrapatero and Bahia Academiamay have one or more song features that are diagnosticbut that were overlooked in the present analysis. (Theonly variable that differed statistically in the presentstudy, minimum frequency, showed extensive overlapbetween sites and thus cannot be regarded as diagnostic.)According to this view, birds may learn one or more diag-nostic features of different song stimulus classes, and sub-sequently categorize novel stimuli based on the presenceor absence of such features. Success in geographical songdiscrimination would thus be contingent on the abilityof birds to form and apply precise categorical distinctions(e.g. Harnad 1987).
An alternative mechanism of geographical song dis-crimination worth considering, alluded to by Ratcliffe &Grant (1985), is exemplar based (e.g. Trainor et al. 2004).

PODOS: DISCRIMINATION OF SONG BY DARWIN’S FINCHES 841
According to this view, birds may be able to recognize spe-cific sets of song types that they hear and memorize, andperhaps only when these songs are heard during the sen-sory phase of song learning. Subsequent responses tosongs, such as during territorial interactions, would thendepend on the relative similarity of song stimuli to previ-ously memorized song exemplars. Experience with partic-ular song exemplars, such as in association with territorialdisputes, may give familiar songs greater communicativesalience. Recent experimental support for familiarity asa factor in geographical song discrimination comes fromO’Loghlen & Beecher (1999), who found that femalesong sparrows, Melospiza melodia, displayed with greaterintensity during playback of familiar than of unfamiliarsong types, even when both kinds of song types were re-corded from strangers. Given the extensive degrees ofwithin-site variation and between-site overlap detectedin the present study, local song discrimination seemsmost consistent with an exemplar-based model of songperception, even though discrete song type categoriesare not readily apparent.
Evolution of Geographical Variation in Song
Geographical variation in song, and the ability of birdsto discriminate songs by locality, often evolves in songbirdpopulations in which at least three criteria are met(Lemon 1975; Krebs & Kroodsma 1980; Nelson & Soha2004). The first is that vocal learning occurs mainlythrough imitation rather than improvization (e.g. Marler& Tamura 1964; Kroodsma 1996). Imitative learning en-ables particular song types to persist in given locations.The second criterion is that vocal learning is sufficientlyplastic to allow for the occurrence and retention of minorinaccuracies in imitation (Marler & Peters 1988). ‘Muta-tions’ in song memes can thus be retained and integratedinto vocal repertoires, and geographical variation gener-ated. The third criterion is that song learning occurs inlocations where birds eventually breed. Nelson (1999),for example, has shown that early song learning inwhite-crowned sparrows facilitates geographical song dif-ferentiation in sedentary populations, but may impedegeographical divergence in song structure in populationsthat engage in long-distance migrations away from theirnatal grounds (Nelson 1998). Geographical variation insong also evolves in populations that learn their songslater in life on their breeding territories following dispersal(e.g. Rothstein & Fleischer 1987; Warren 2002).
The present description of vocal geographical variationand discrimination suggests that these three criteria aremet to some extent in Santa Cruz G. fortis. In Darwin’sground finches, as in most emberizids, song learning relieslargely on imitation, and occurs early in life (Bowman1983). The observation that young males normally learntheir songs from their fathers is most likely explained byearly learning (Grant & Grant 1996, 1997, 1998). An earlywindow for vocal imitation in G. fortis, coupled with thepresent observation of geographical song variation anddiscrimination, suggests that these birds breed on their na-tal grounds (i.e. that they show some degree of
philopatry). The actual degree to which G. fortis on south-eastern Santa Cruz are philopatric is presently unknown,although three lines of evidence from our researchprogram support a hypothesis of philopatry (S. Huber,A. Herrel, A. Hendry & J. Podos, unpublished data). First,only one of the approximately 1000 birds banded at thetwo study sites (w500 at each site) between 1999 and2006 has been recaptured or resighted between localities.Second, the between-year recapture/resighting percentageof G. fortis at El Garrapatero, the site of our most intensivecurrent field effort, is substantial (at least 40% from 2003to 2004, and again from 2004 to 2005). Third, birds withknown breeding sites have regularly been observed in thesame or adjacent territories over subsequent years. Thisevidence refers most directly to breeding philopatry; lessis known about natal philopatry for these populations.
Within-island Divergence?
Geographical variation in song, between-site songdiscrimination, early song learning, and indications ofphilopatry, together suggest an opportunity for localadaptation and divergence among G. fortis populationson southeastern Santa Cruz. The results of playback testssuggest that males dispersing to neighbouring localitiesenjoy fewer challenges from other males during territoryacquisition and maintenance. However, females at thesesites presumably also fail to respond to the songs of new-comers, because of a parallel failure to recognize, and thusprefer to mate with local males (e.g. Searcy et al. 2002). Fe-male songbirds tend to show stronger discrimination ofsong variants than do males, perhaps as a result of theirgenerally greater reproductive and parental investment(Ratcliffe & Otter 1996). Searcy et al. (2002), for example,showed that male song sparrows tend to be more catholicthan females in the range of geographical song variants towhich they respond. For Darwin’s finches, data from theresponses of males to simulated territorial intrusionshave been used to draw inferences about speciation, inpart following the assumption that stringency in re-sponses expressed by males will likely carry over, and per-haps be amplified, in females (Grant & Grant 2002a, b).Tests of geographical discrimination in females (e.g. usinga copulation solicitation display assay; Baker et al. 1987a;Searcy 1992), would ultimately provide more direct in-sights into the role of geographical song variation as a po-tential deterrent in cross-population breeding.
Actual probabilities of interpopulation divergence de-pend on numerous additional factors not yet character-ized. Interpopulation divergence may be counterbalancedby even low levels of gene flow between finch populations(Hendry et al. 2002; Emelianov et al. 2004; see also Petrenet al. 2005). To test the possibility of local adaptive diver-gence, it might be best to start with an assessment of geneflow between such populations. Geographical differencesin genetic structure, however, even if present and identi-fied, may not necessarily persist over time. Studies ofG. conirostris (large cactus finch) on Genovesa Island indi-cate that initial within-island divergence in Darwin’s finchmorphology and song can be counteracted by subsequent

ANIMAL BEHAVIOUR, 73, 5842
nonassortative mating and resulting interpopulation geneflow (reviewed by Grant & Grant 1979, 1989). The possi-bility of within-island divergence has thus been largelydiscounted for Darwin’s finches (Grant 1999; Grant &Grant 2003). A recent study by Kleindorfer et al. (2006),however, provides quantitative evidence for within-islandmorphological divergence in one of Darwin’s groundfinches, G. fuliginosa, between upland and coastal siteson Santa Cruz Island.
The conclusions of the present playback study are alsonecessarily tempered by limits on the study design, whichdid not include the reciprocal experiment (playback of ElGarrapatero songs at Bahia Academia), and which usedonly large-morph songs from El Garrapatero as playbackstimuli. Future progress in evaluating probabilities ofintersite divergence in Santa Cruz G. fortis will also requirefurther studies on song variation, perception and discrim-ination at multiple sites (e.g. Grant & Grant 1989; Grantet al. 2004; Patten et al. 2004). Information about geneticdiversity and gene flow would be well complemented bystudies of dispersal and philopatry, insofar as high philo-patry would suggest a greater opportunity for within-island divergence.
Acknowledgments
I thank Marcos Rossi-Santos, Diego Ruiz, J. Patrick Kelleyand Sarah Huber for assistance over several field seasons indesigning and refining the song playback protocol usedhere. Additional field assistance was provided by LuisFernando de Leon, Andrew Hendry, and Ana Gabela.Charles Henry, Paige Warren, Sarah Huber, David Lahti,Dan Ardia, Andrew Hendry, Mike Webster, Bill Searcy andseveral anonymous referees offered valuable feedback onearlier versions of this manuscript. Permits and supportwere kindly provided by the Charles Darwin ResearchStation and the Galapagos National Park Service. Fundedby National Science Foundation grant IBN-0347291.
References
Baker, M. C. & Thompson, D. B. 1985. Song dialects of sparrows:
historical processes inferred from patterns of geographic variation.Condor, 87, 127e141.
Baker, M. C., Bjerke, T. K., Lampe, H. U. & Espmark, Y. O. 1987a.Sexual response of female yellowhammers to differences in regional
song dialects and repertoire sizes. Animal Behaviour, 35, 395e401.
Baker, M. C., Spitler-Nabors, K. J., Thompson, A. D., Jr. &Cunningham, M. A. 1987b. Reproductive behaviour of female
white-crowned sparrows: effect of dialects and synthetic hybrid
songs. Animal Behaviour, 35, 1766e1774.
Baptista, L. F. 1975. Song dialects and demes in sedentary popula-
tions of the white-crowned sparrow (Zonotrichia leucophrys nut-talli). University of California Publications in Zoology, 105, 1e52.
Bard, S. C., Hau, M., Wikelski, M. & Wingfield, J. C. 2002. Vocaldistinctiveness and response to conspecific playback in the spotted
antbird, a Neotropical suboscine. Condor, 104, 387e394.
Beeman, K. 2002. SIGNAL 4.0. Belmont, Massachusetts: Engineer-
ing Design.
Bowman, R. I. 1979. Adaptive morphology of song dialects in
Darwin’s finches. Journal fur Ornithologie, 120, 353e389.
Bowman, R. I. 1983. The evolution of song in Darwin’s finches.
In: Patterns of Evolution in Galapagos Organisms (Ed. by R. I.
Bowman, M. Berson & A. E. Leviton), pp. 237e537. SanFrancisco: American Association for the Advancement of Sci-
ence, Pacific Division.
Clark, C. W., Marler, P. & Beeman, K. 1987. Quantitative analysis
of animal vocal phonology: an application to swamp sparrow
song. Ethology, 76, 101e115.
Collins, S. 2004. Vocal fighting and flirting: the functions of
birdsong. In: Nature’s Music: the Science of Birdsong (Ed. by
P. Marler & H. Slabbekoorn), pp. 39e79. Amsterdam: ElsevierAcademic Press.
Edwards, S. V., Kingan, S. B., Calkins, J. D., Balakrishnan, C. N.,Jennings, W. B., Swanson, W. J. & Sorenson, M. D. 2005.
Speciation in birds: genes geography, and sexual selection.
Proceedings of the National Academy of Sciences, U.S.A., 102,
6550e6557.
Emelianov, I., Marec, F. & Mallet, J. 2004. Genomic evidence for
divergence with gene flow in host races of the larch budmoth.Proceedings of the Royal Society of London, Series B, 271,
97e105.
Falls, J. B. 1992. Playback: a historical perspective. In: Playback and
Studies of Animal Communication (Ed. by P. K. McGregor), pp. 11e
33. New York: Plenum.
Foster, S. A. 1999. The geography of behaviour: an evolutionary
perspective. Trends in Ecology and Evolution, 14, 190e195.
Grant, B. R. & Grant, P. R. 1979. Darwin finches: population varia-
tion and sympatric speciation. Proceedings of the National Academy
of Sciences, U.S.A., 76, 2359e2363.
Grant, B. R. & Grant, P. R. 1989. Evolutionary Dynamics of a Natural
Population: the Large Cactus Finch of the Galapagos. Chicago:University of Chicago Press.
Grant, B. R. & Grant, P. R. 1996. Cultural inheritance of song and itsrole in the evolution of Darwin’s finches. Evolution, 50, 2471e
2487.
Grant, B. R. & Grant, P. R. 1998. Hybridization and speciation in
Darwin’s finches: the role of sexual imprinting on a culturally
transmitted trait. In: Endless Forms: Species and Speciation (Ed. by
D. J. Howard & S. H. Berlocher), pp. 404e422. Oxford: OxfordUniversity Press.
Grant, B. R. & Grant, P. R. 2002a. Lack of premating isolation at thebase of a phylogenetic tree. American Naturalist, 160, 1e19.
Grant, B. R. & Grant, P. R. 2002b. Simulating secondary contact inallopatric speciation: an empirical test of premating isolation.
Biological Journal of the Linnean Society, 76, 545e556.
Grant, B. R. & Grant, P. R. 2003. What Darwin’s finches can teach
us about the evolutionary origin and regulation of biodiversity.
Bioscience, 53, 965e975.
Grant, P. R. 1999. Ecology and Evolution of Darwin’s Finches. Prince-
ton, New Jersey: Princeton University Press.
Grant, P. R. & Grant, B. R. 1997. Hybridization, sexual imprinting,
and mate choice. American Naturalist, 149, 1e28.
Grant, P. R. & Grant, B. R. 2002c. Unpredictable evolution in
a 30-year study of Darwin’s finches. Science, 296, 707e711.
Grant, P. R., Grant, B. R., Markert, J. A., Keller, L. F. & Petren, K.2004. Convergent evolution of Darwin’s finches caused by intro-
gressive hybridization and selection. Evolution, 58, 1588e1599.
Harnad, S. 1987. Categorical Perception: the Groundwork of Cogni-
tion. New York: Cambridge University Press.
Hendry, A. P., Taylor, E. B. & McPhail, J. D. 2002. Adaptive di-
vergence and the balance between selection and gene flow:
lake and stream stickleback in the misty system. Evolution, 56,1199e1216.
Hendry, A. P., Grant, P. R., Grant, B. R., Ford, H., Brewer, M. &Podos, J. 2006. Possible human impacts on adaptive radiation:

PODOS: DISCRIMINATION OF SONG BY DARWIN’S FINCHES 843
beak size bimodality in Darwin’s finches. Proceedings of the Royal
Society, Series B, 273, 1887e1894.
Huber, S. K. & Podos, J. 2006. Beak morphology and song features
covary in a population of Darwin’s finches (Geospiza fortis).
Biological Journal of the Linnean Society, 88, 489e498.
Hunter, M. L. & Krebs, J. R. 1979. Geographical variation in the
song of the great tit (Parus major) in relation to ecological factors.Journal of Animal Ecology, 48, 759e785.
Irwin, D. E. 2000. Song variation in an avian ring species. Evolution,54, 998e1010.
Irwin, D. E., Bensch, S. & Price, T. D. 2001. Speciation in a ring.Nature, 409, 333e337.
JMP 2002. Version 5.1. Cary, NC: SAS Institute Inc.
Kachigan, S. K. 1991. Multivariate Statistical Analysis: a Conceptual
Introduction. New York: Radius.
Kleindorfer, S., Chapman, T. W., Winkler, H. & Sulloway, F. 2006.
Adaptive divergence in contiguous populations of Darwin’s small
ground finch (Geospiza fuliginosa). Evolutionary Ecology Research,8, 357e372.
Krebs, J. R. & Kroodsma, D. E. 1980. Repertoires and geographicalvariation in birdsong. Advances in the Study of Behavior, 11, 143e
177.
Kroodsma, D. E. 1996. Ecology of passerine song development.
In: Ecology and Evolution of Acoustic Communication in Birds
(Ed. by D. E. Kroodsma & E. H. Miller), pp. 3e19. Ithaca,
New York: Cornell University Press.
Kroodsma, D. E., Baker, M. C., Baptista, L. F. & Petrinovich, L.1985. Vocal ‘dialects’ in Nuttall’s white-crowned sparrow. CurrentOrnithology, 2, 103e133.
Kroodsma, D. E., Byers, B. E., Goodale, E., Johnson, S. & Liu,W. C. 2001. Pseudoreplication in playback experiments, revisited
a decade later. Animal Behaviour, 61, 1029e1033.
Lachlan, R. F. & Servedio, M. R. 2004. Song learning accelerates
allopatric speciation. Evolution, 58, 2049e2063.
Lack, D. 1945. The Galapagos finches (Geospizinae): a study in var-
iation. Occasional Papers of the California Academy of Sciences, 21,
1e159.
Leader, N., Wright, J. & Yom-Tov, Y. 2002. Dialect discrimination
by male orange-tufted sunbirds (Nectarinia osea): reactions to own
vs. neighbor dialects. Ethology, 108, 367e376.
Lemon, R. E. 1967. The response of cardinals to songs of different
dialects. Animal Behaviour, 15, 538e545.
Lemon, R. E. 1975. How birds develop song dialects. Condor, 77,
385e406.
Lovell, S. F. & Lein, M. R. 2004. Neighborestranger discrimination
by song in a suboscine bird, the alder flycatcher, Empidonax alno-rum. Behavioral Ecology, 15, 799e804.
MacDougall-Shackleton, E. A. & MacDougall-Shackleton, S. A.2001. Cultural and genetic evolution in mountain white-crowned
sparrows: song dialects are associated with population structure.
Evolution, 55, 2568e2575.
McGregor, P. K. 1983. The response of corn buntings to playback of
dialects. Zeitschrift fur Tierpsychologie, 62, 256e260.
Marler, P. & Peters, S. 1988. The role of song phonology and syn-
tax in vocal learning preferences in the song sparrow, Melospiza
melodia. Ethology, 77, 125e149.
Marler, P. & Tamura, M. 1962. Song ‘dialects’ in three populations
of white-crowned sparrows. Condor, 64, 368e377.
Marler, P. & Tamura, M. 1964. Culturally transmitted patterns of
vocal behavior in sparrows. Science, 146, 1483e1486.
Moran, M. D. 2003. Arguments for rejecting the sequential Bonfer-
roni in ecological studies. Oikos, 100, 403e405.
Nelson, D. A. 1998. Geographic variation in song of Gambel’s
white-crowned sparrow. Behaviour, 135, 321e342.
Nelson, D. A. 1999. Ecological influences on vocal development in
the white-crowned sparrow. Animal Behaviour, 58, 21e36.
Nelson, D. A. & Marler, P. 1990. The perception of birdsong and an
ecological concept of signal space. In: Comparative Perception.
Vol. 2, Complex Signals (Ed. by W. C. Stebbins & M. A. Berkley),pp. 443e478. New York: J. Wiley.
Nelson, D. A. & Soha, J. A. 2004. Perception of geographical var-iation in song by male Puget Sound white-crowned sparrows,
Zonotrichia leucophrys pugetensis. Animal Behaviour, 68, 395e
405.
O’Loghlen, A. L. & Beecher, M. D. 1999. Mate, neighbour and
stranger songs: a female song sparrow perspective. Animal Behav-
iour, 58, 13e20.
Patten, M. A., Rotenberry, J. T. & Zuk, M. 2004. Habitat selection,
acoustic adaptation, and the evolution of reproductive isolation.Evolution, 58, 2114e2155.
Petren, K., Grant, P. R., Grant, B. R. & Keller, L. F. 2005. Compar-ative landscape genetics and the adaptive radiation of Darwin’s
finches: the role of peripheral isolation. Molecular Ecology, 14,
2943e2957.
Podos, J. 1997. A performance constraint on the evolution of trilled
vocalizations in a songbird family (Passeriformes: Emberizidae).
Evolution, 51, 537e551.
Podos, J. 2001. Correlated evolution of morphology and vocal signal
structure in Darwin’s finches. Nature, 409, 185e188.
Podos, J. & Nowicki, S. 2004. Beaks, adaptation, and vocal evolu-
tion in Darwin’s finches. BioScience, 54, 501e510.
Podos, J. & Warren, P.S., In press. The evolution of geographic var-
iation in bird songs. Advances in the Study of Behavior.
Podos, J., Huber, S. K. & Taft, B. 2004. Bird song: the interface of
evolution and mechanism. Annual Review of Ecology, Evolution,and Systematics, 35, 55e87.
Ptacek, M. B. 2000. The role of mating preferences in shaping inter-
specific divergence in mating signals in vertebrates. BehaviouralProcesses, 51, 111e134.
Ratcliffe, L. M. 1981. Species recognition in Darwin’s groundfinches (Geospiza Gould). Ph.D. thesis, McGill University.
Ratcliffe, L. M. & Grant, P. R. 1985. Species recognition in Darwin’sfinches (Geospiza Gould). III. Male responses to playback of differ-
ent song types, dialects, and heterospecific songs. Animal Behav-
iour, 33, 290e307.
Ratcliffe, L. M. & Otter, K. 1996. Sex differences in song recogni-
tion. In: Ecology and Evolution of Acoustic Communication in Birds
(Ed. by D. E. Kroodsma & E. H. Miller), pp. 339e355. Ithaca,New York: Cornell University Press.
Rothstein, S. I. & Fleischer, R. C. 1987. Vocal dialects and their pos-sible relation to honest status signalling in the brown-headed cow-
bird. Condor, 89, 1e23.
Searcy, W. A. 1992. Measuring responses of female birds to male
song. In: Playback and Studies of Animal Communication (Ed. by
P. K. McGregor), pp. 175e189. New York: Plenum.
Searcy, W. A., Nowicki, S. & Hughes, M. 1997. The response of
male and female song sparrows to geographic variation in song.
Condor, 99, 651e657.
Searcy, W. A., Nowicki, S., Hughes, M. & Peters, S. 2002. Geo-
graphic song discrimination in relation to dispersal distances insong sparrows. American Naturalist, 159, 221e230.
Slabbekoorn, H. & Smith, T. B. 2002. Bird song, ecology andspeciation. Philosophical Transactions of the Royal Society of London,
Series B, 357, 493e503.
Slater, P. J. B. 1986. The cultural transmission of bird song. Trends in
Ecology and Evolution, 1, 94e97.
Slater, P. J. B. 1989. Bird song learning: causes and consequences.
Ethology, Ecology and Evolution, 1, 19e46.

ANIMAL BEHAVIOUR, 73, 5844
Slater, P. J. B., Clements, F. A. & Goodfellow, D. J. 1984. Local and
regional variations in chaffinch song and the question of dialects.
Behaviour, 88, 76e97.
Snodgrass, R. E. & Heller, E. 1904. Papers from the Hopkinse
Stanford Galapagos expedition, 1898e1899. XVI. Birds. Proceed-ings of the Washington Academy of Science, 5, 231e372.
Trainor, L. J., Wu, L. A. & Tsang, C. D. 2004. Long-term memoryfor music: infants remember tempo and timbre. Developmental
Science, 7, 289e296.
Warren, P. S. 2002. Geographic variation and dialects in the flight whis-
tle of the bronzed cowbird (Molothrus aeneus). Auk, 119, 349e361.
Wells, M. M. & Henry, C. S. 1998. Songs, reproductive isolation
and speciation in cryptic species of insects: a case study using
green lacewings. In: Endless Forms: Species and Speciation (Ed.by D. Howard & S. Berlocher), pp. 217e233. New York: Oxford
University Press.
West-Eberhard, M. J. 1983. Sexual selection, social competition,
and speciation. Quarterly Review of Biology, 58, 155e183.