discovery of serum biomarkers implicated in the onset and progression of serous ovarian cancer in a...
TRANSCRIPT

European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103
Discovery of serum biomarkers implicated in the onset and progression of serousovarian cancer in a rat model using iTRAQ technique
Yiping Huang a,d,1, Xiaoyan Zhang a,d,1, Wei Jiang a,d, Yisheng Wang a,d, Hong Jin b,Xiaohui Liu b, Congjian Xu a,b,c,d,*a Obstetrics and Gynecology Hospital, Fudan University, Shanghai 200032, People’s Republic of Chinab Institutes of Biomedical Sciences, Fudan University, Shanghai 200032, People’s Republic of Chinac Department of Obstetrics and Gynecology of Shanghai Medical School, Fudan University, Shanghai 200032, People’s Republic of Chinad Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Shanghai 200011, People’s Republic of China
A R T I C L E I N F O
Article history:
Received 24 April 2012
Received in revised form 29 May 2012
Accepted 26 June 2012
Keywords:
Serous ovarian cancer
Carcinogenesis
Biomarker
Proteomics
Animal model
A B S T R A C T
Objective: Epithelial ovarian cancer (EOC) is the most lethal gynecologic malignancy, and early tumor
detection is the most promising approach for improving the EOC survival rate. The goal of this study was
to identify the biomarkers underlying ovarian carcinogenesis.
Study design: To mimic the onset and progression of human ovarian cancer, we established a rat model of
ovarian neoplasm by implanting 7,12-dimethylbenz(a)anthracene (DMBA)-coated silk cloth strips onto
the ovaries. Sera collected from rats bearing serous ovarian carcinoma (SOC) at baseline, 12 and 24 weeks
after DMBA treatment and from controls were analyzed using iTRAQ combined with two-dimensional
liquid chromatography and tandem mass spectrometry. The data were analyzed with ProteinPilot
software for peptide matching, protein identification, and protein quantitation. Ingenuity pathway
analysis software was used to identify the canonical pathways and biological interaction networks of
differentially expressed proteins.
Results: The cumulative ovarian tumor incidence rate reached 75% at 32 weeks after DMBA treatment.
Out of all tumors, 94% were EOC, and 51% of the EOC cases were SOC. A total of 225 unique, non-
redundant proteins were identified with 95% confidence. Twenty-seven differentially expressed proteins
were significantly up- or down-regulated during the early or advanced carcinogenesis of SOC. Fifteen
proteins were previously reported to be involved in ovarian cancer, and 12 proteins, including MMRN1,
SERPINC1, TLN1, AHSG, PLG, APOA2, HPX, APOC1, APOC2, FERMT3, FETUB and HBB, were identified for
the first time in our study.
Conclusion: The discovery of these differentially expressed proteins provides valuable clues for
understanding the molecular mechanism underlying the dynamic carcinogenic process of ovarian
cancer. These proteins could be used as diagnostic biomarkers for early detection, disease monitoring
and therapeutic targets.
� 2012 Elsevier Ireland Ltd. All rights reserved.
Contents lists available at SciVerse ScienceDirect
European Journal of Obstetrics & Gynecology andReproductive Biology
jou r nal h o mep ag e: w ww .e lsev ier . co m / loc ate /e jo g rb
1. Introduction
Epithelial ovarian cancer (EOC) is the most lethal gynecologicmalignancy. Most patients suffering from EOC are diagnosed atadvanced stages because of the difficulties of early detection.Patients diagnosed with early-stage EOC have a 5-year survivalrate over 90%, while patients at advanced stages have a less than30% 5-year survival rate [1].
* Corresponding author at: Obstetrics and Gynecology Hospital, Fudan Universi-
ty, Shanghai 200032, People’s Republic of China. Tel.: +86 21 63455050;
fax: +86 21 63455090.
E-mail address: [email protected] (C. Xu).1 These authors contributed equally to this work.
0301-2115/$ – see front matter � 2012 Elsevier Ireland Ltd. All rights reserved.
http://dx.doi.org/10.1016/j.ejogrb.2012.06.031
The most commonly used serum biomarker for early detection ofovarian cancer is cancer antigen 125 (CA125), but CA125 has lowsensitivity and specificity. To improve early EOC detection, manybiomarkers have been identified. Although these biomarkers areoverexpressed in EOC, they might not play key roles in the initiationand progression of carcinogenesis. Thus, identifying biomarkers thatmediate carcinogenesis could be a promising approach for earlydetection and dynamic disease monitoring of EOC.
Studies on early detection often focus on cell lines or tissues,sera and plasma collected from patients, but human specimens atearly stages of carcinogenesis are limited because many patientssuffering from EOC are diagnosed at advanced stages. Also,cultured cells do not reflect the dynamic carcinogenic process.The use of animal tumor models might be an alternative approachthat could represent the onset and progression of cancer [2].

Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103 97
We used a rat model of 7,12-dimethylbenz(a)anthracene(DMBA)-induced ovarian neoplasm to mimic the onset andprogression of human ovarian cancer. To discover potential serumbiomarkers underlying the carcinogenesis process, sera collectedfrom rats at baseline, 12 and 24 weeks post-DMBA treatment andcontrols were analyzed using isobaric tags for relative and absolutequantification (iTRAQ) combined with two-dimensional liquidchromatography and tandem mass spectrometry (2DLC–MS/MS).The comparison sets for early and advanced carcinogenic processeswere then analyzed to determine the proteomic changes duringcarcinogenesis. To the best of our knowledge, this is the first studyto investigate the proteomic changes in the carcinogenic process ofEOC using the iTRAQ technique.
Fig. 1. Histopathology of DMBA-induced ovarian tumors in rats (original magnification 20
mucinous carcinoma and (E) sarcoma.
2. Materials and methods
2.1. Animals and treatment
Six-week-old female Wistar rats (purchased from theExperimental Animal Center, Chinese Academy of Science,China) were anesthetized by intraperitoneal injection of chloralhydrate. Then both ovaries were surgically exposed, and aDMBA-coated or DMBA-free silk cloth strip (0.5 cm � 0.5 cm)was placed onto the surfaces of both ovaries and packed withthe surrounding fatty tissues. Each cloth strip containedapproximately 1.5 mg DMBA (Sigma Chemical Co., USA). Afterthe surgery, tumor growth was determined weekly by palpating
0�). (A) Serous carcinoma, (B) endometrioid carcinoma, (C) clear cell carcinoma, (D)

Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–10398
the abdomen and sera were collected at baseline, 12 and 24weeks. The animals were sacrificed at 32 weeks, and theovaries were harvested and subjected to histopathologicalexamination.
2.2. Serum depletion and iTRAQ labeling
Sera collected were pooled into six groups, each containingsera from 5 rats. Serum pools were depleted of high-abundanceproteins using ProteoMiner enrichment kit (Bio-Rad Laborato-ries Inc., USA) according to the manufacturer’s instructions.Then, 100 mg peptides from each sample were labeled withiTRAQ reagents. The serum pools from SOC rats at pre-treatmentand at 12 and 24 weeks after DMBA treatment were labeled withisobaric tags 117, 118 and 119, and the serum pools from thematched controls were labeled with isobaric tags 114, 115 and116, respectively.
2.3. Strong cation exchange liquid chromatography
To reduce sample complexity, labeled peptides were fraction-ated by strong cation exchange (SCX) liquid chromatography usinga PolySulfoethyl A column (PolyLC Inc., USA). The dried peptideswere resuspended in buffer A (10 mM KH2PO4, 25%, v/v,acetonitrile (ACN), pH 2.6) and were loaded onto the SCX column.After washing with buffer A, peptides were eluted by running a
Fig. 2. Multiplex analysis of sera from rats with DMBA-induced SOC and controls. (A) The
pre- and post-DMBA treatment and matched controls were analyzed. Each time point con
115/114 (control: 12 weeks of follow-up versus baseline) and 118/117 (SOC: 12 weeks
advanced carcinogenesis. The ratios for 116/114 (control: 24 weeks of follow-up versus
compared.
5–25% buffer B gradient (10 mM KH2PO4, 350 mM KCl, 25%, v/v,ACN, pH 2.6) for 45 min and then to 100% buffer B.
2.4. Reverse-phase LC–MS/MS analysis
Dried SCX fractions were reconstituted in 100 ml of buffer A (5%,v/v, ACN, 0.1%, v/v, formic acid (FA)) and loaded into a reverse-phase C18 peptide Captrap (Agilent Technologies, USA). Afterdesalting, peptides were eluted by running a 5% to 35% buffer Bgradient (95%, v/v, ACN, 0.1%, v/v, FA) for 55 min and then a 50%buffer B for 15 min. The LC elution was subjected to positive-ionnano-flow electrospray analysis using a Qstar XL MS/MS system(Applied Biosystems Inc., USA). A survey scan was acquired fromm/z 400–1800 for 0.5 s with up to four precursor ions selected fromm/z 100–2000 for MS/MS.
2.5. Database search, protein identification and IPA analysis
The MS/MS data were analyzed with ProteinPilot software 3.0(Applied Biosystems Inc., USA). The International Protein Index(IPI) rat database (IPI.rat.v3.55) was used for all searches. Thecriteria for protein identification were as follows: >95% peptideconfidence (ProtScore �1.3) with a relative quantification P-value <0.05. Ingenuity pathway analysis (IPA) software was usedto identify the canonical pathways and biological interactionnetworks.
design workflow used for iTRAQ based analysis. Sera collected from SOC-bearing rats
tained 5 sera samples. (B) The comparison set for early carcinogenesis. The ratios for
post-DMBA treatment versus baseline) were compared. (C) The comparison set for
baseline) and 119/117 (SOC: 24 weeks post-DMBA treatment versus baseline) were

Table 1Differentially expressed proteins identified in SOC-bearing rats at 12 weeks post-DMBA treatment.
Accession Gene symbol Protein name Ratio
115/114a
P-value Ratio
118/117b
P-value No. of
peptidesc
IPI00372372.2 SERPINC1 Serine peptidase inhibitor, clade C, member 1 0.163 0.000 8.954 0.004 42
IPI00476292.3 CP Ceruloplasmin 1.738 0.001 8.872 0.000 38
IPI00202610.1 F7 Coagulation factor VII precursor 3.048 0.007 7.112 0.016 5
IPI00422076.1 THBS1 Thrombospondin 1 NI 4.246 0.001 65
IPI00197700.1 APOA2 Apolipoprotein A-II precursor 8.954 0.002 4.093 0.000 87
IPI00393058.4 MMRN1 Similar to multimerin 1 NI 3.908 0.038 6
IPI00188541.3 ITIH4 Inter-alpha (globulin) inhibitor H4 5.248 0.000 2.333 0.000 378
IPI00363974.3 GSN Gelsolin precursor 1.941 0.000 1.803 0.001 23
IPI00188225.1 CRP C-reactive protein precursor NI 1.690 0.002 5
IPI00195516.6 HPX Hemopexin precursor NI 0.525 0.020 8
IPI00679202.2 TF Serotransferrin precursor NI 0.425 0.002 14
IPI00209113.3 MYH9 Myosin-9 0.506 0.025 0.409 0.015 6
IPI00362014.5 TLN1 Talin 1 0.167 0.000 0.347 0.000 99
IPI00194097.5 GC Vitamin D-binding protein precursor NI 0.331 0.005 26
IPI00764461.1 ACTG1 Similar to actin, cytoplasmic 2 NI 0.227 0.024 13
IPI00327469.1 AHSG Alpha-2-HS-glycoprotein precursor NI 0.196 0.002 45
IPI00197703.2 APOA1 Apolipoprotein A-I precursor NI 0.185 0.000 131
IPI00454431.1 ACTN1 Brain-specific alpha actinin 1 isoform 0.334 0.002 0.177 0.002 11
IPI00206780.1 PLG Plasminogen precursor NI 0.149 0.000 53
IPI00778265.1 CFH Complement factor H NI 0.108 0.034 114
IPI00231757.12 PSMA2 Proteasome subunit alpha type-2 NI 0.097 0.024 2
IPI00362927.1 TUBA4A Tubulin alpha-4A chain 0.273 0.020 NI 19
IPI00561922.2 HRG Histidine-rich glycoprotein 1.585 0.015 NI 24
IPI00231772.3 ECM1 Extracellular matrix protein 1 precursor 4.742 0.008 NI 10
IPI00214905.3 TPM4 Tropomyosin alpha-4 chain 0.100 0.007 NI 22
NI: not identified.a Ratio of the fold change of proteins in controls at 12 weeks of follow-up versus baseline.b Ratio of the fold change of proteins in SOC-bearing rats at 12 weeks post-DMBA treatment versus baseline.c The peptides identified with 95% confidence in the reporter ion quantification.
Fig. 3. Potential biomarkers in the initiation process of DMBA-induced SOC. The bar
graph represents the log ratios of the proteins (P < 0.05) identified in SOC-bearing
rats at 12 weeks post-DMBA treatment.
Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103 99
3. Results
3.1. Ovarian neoplasia induced by DMBA in rats
At 32 weeks, the cumulative incidence reached 75% (96/128).The majority of induced tumors (94%, 90/96) were diagnosed asEOC, and the remaining tumors (6%, 6/96) were diagnosed asovarian sarcoma. In DMBA-induced EOC, 46 cases (46/90) wereserous carcinoma, 28 cases (28/90) were endometrioid carcinoma,3 cases (3/90) were clear cell carcinoma, 2 cases (2/90) weremucinous carcinoma, and 11 cases (11/90) were mixed epithelialcarcinomas (Fig. 1). As serous ovarian carcinoma (SOC) is the mostcommon type of EOC, the sera from rats bearing induced SOC wereanalyzed.
3.2. Multiplex analysis of sera from rats with DMBA-induced serous
ovarian carcinoma
The sera from rats with DMBA-induced SOC and controls atdifferent time points were analyzed. The experimental design isshown in Fig. 2A. A total of 225 unique, non-redundant proteinswere identified with 95% confidence. To determine the proteomicchanges during DMBA-induced SOC, two comparison sets weredesigned. In the comparison set for early carcinogenesis, the iTRAQratios for 115/114 (control: 12 weeks of follow-up versus baseline)and 118/117 (SOC: 12 weeks post-DMBA treatment versusbaseline) were compared (Fig. 2B). In the comparison set foradvanced carcinogenesis, iTRAQ 115 was replaced with 116(control: 24 weeks of follow-up), and 118 was replaced with119 (SOC: 24 weeks post-DMBA treatment) (Fig. 2C).
3.3. Differentially expressed proteins in the initiation process of
DMBA-induced serous ovarian cancer
Twenty-five proteins were differentially expressed in the first12 weeks after DMBA exposure (Table 1). As shown in Fig. 3, levels
of serine peptidase inhibitor C1 (SERPINC1), ceruloplasmin (CP),coagulation factor VII (F7), thrombospondin 1 (THBS1), multimerin1 (MMRN1) and C-reactive protein (CRP) increased in DMBA-induced carcinogenesis, while levels of hemopexin (HPX),
Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103100
serotransferrin (TF), vitamin D-binding protein (GC), actin cyto-plasmic 2 (ACTG1), alpha-2-HS-glycoprotein (AHSG), apolipopro-tein A-I (APOA1), actinin alpha 1 (ACTN1), plasminogen (PLG),complement factor H (CFH) and proteasome subunit alpha type-2(PSMA2) decreased. In addition, apolipoprotein A-II (APOA2) andinter-alpha globulin inhibitor H4 (ITIH4) levels were elevated overtime, although their levels in SOC-bearing rats were lower than incontrols. Although the level of talin 1 (TLN1) decreased in all ratsover time, the level in SOC-bearing rats was higher than in controls.
3.4. Differentially expressed proteins in the advanced process of
DMBA-induced serous ovarian cancer
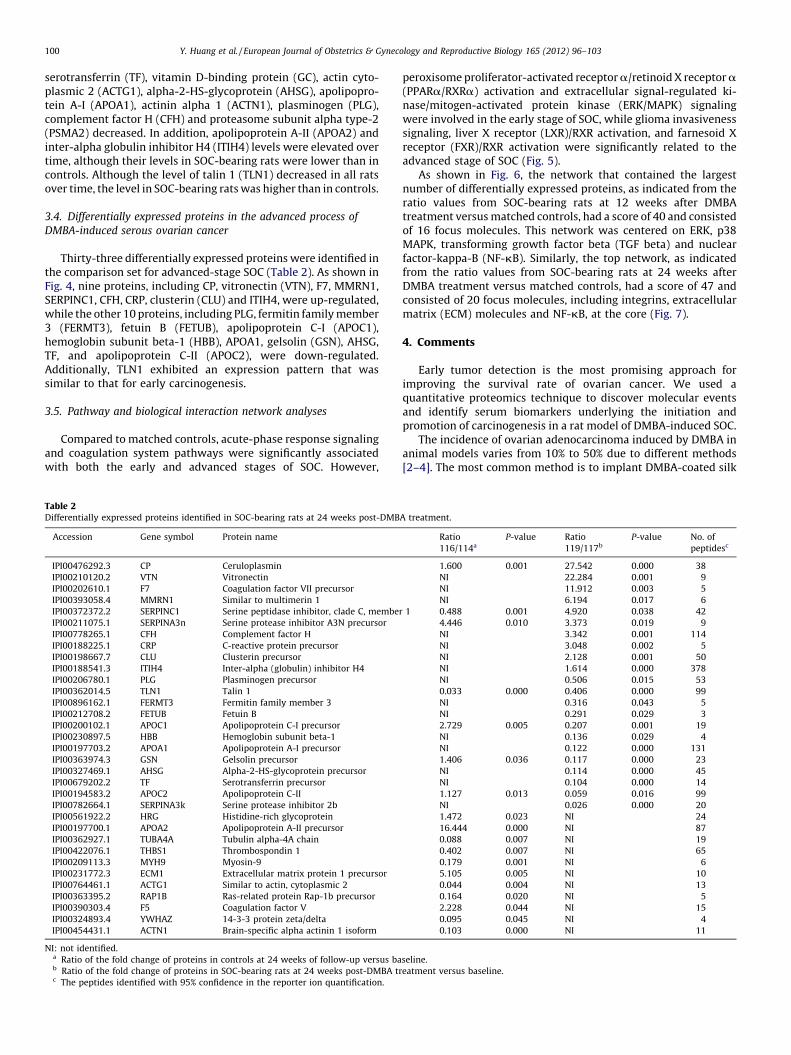
Thirty-three differentially expressed proteins were identified inthe comparison set for advanced-stage SOC (Table 2). As shown inFig. 4, nine proteins, including CP, vitronectin (VTN), F7, MMRN1,SERPINC1, CFH, CRP, clusterin (CLU) and ITIH4, were up-regulated,while the other 10 proteins, including PLG, fermitin family member3 (FERMT3), fetuin B (FETUB), apolipoprotein C-I (APOC1),hemoglobin subunit beta-1 (HBB), APOA1, gelsolin (GSN), AHSG,TF, and apolipoprotein C-II (APOC2), were down-regulated.Additionally, TLN1 exhibited an expression pattern that wassimilar to that for early carcinogenesis.
3.5. Pathway and biological interaction network analyses
Compared to matched controls, acute-phase response signalingand coagulation system pathways were significantly associatedwith both the early and advanced stages of SOC. However,
Table 2Differentially expressed proteins identified in SOC-bearing rats at 24 weeks post-DMB
Accession Gene symbol Protein name
IPI00476292.3 CP Ceruloplasmin
IPI00210120.2 VTN Vitronectin
IPI00202610.1 F7 Coagulation factor VII precursor
IPI00393058.4 MMRN1 Similar to multimerin 1
IPI00372372.2 SERPINC1 Serine peptidase inhibitor, clade C, member
IPI00211075.1 SERPINA3n Serine protease inhibitor A3N precursor
IPI00778265.1 CFH Complement factor H
IPI00188225.1 CRP C-reactive protein precursor
IPI00198667.7 CLU Clusterin precursor
IPI00188541.3 ITIH4 Inter-alpha (globulin) inhibitor H4
IPI00206780.1 PLG Plasminogen precursor
IPI00362014.5 TLN1 Talin 1
IPI00896162.1 FERMT3 Fermitin family member 3
IPI00212708.2 FETUB Fetuin B
IPI00200102.1 APOC1 Apolipoprotein C-I precursor
IPI00230897.5 HBB Hemoglobin subunit beta-1
IPI00197703.2 APOA1 Apolipoprotein A-I precursor
IPI00363974.3 GSN Gelsolin precursor
IPI00327469.1 AHSG Alpha-2-HS-glycoprotein precursor
IPI00679202.2 TF Serotransferrin precursor
IPI00194583.2 APOC2 Apolipoprotein C-II
IPI00782664.1 SERPINA3k Serine protease inhibitor 2b
IPI00561922.2 HRG Histidine-rich glycoprotein
IPI00197700.1 APOA2 Apolipoprotein A-II precursor
IPI00362927.1 TUBA4A Tubulin alpha-4A chain
IPI00422076.1 THBS1 Thrombospondin 1
IPI00209113.3 MYH9 Myosin-9
IPI00231772.3 ECM1 Extracellular matrix protein 1 precursor
IPI00764461.1 ACTG1 Similar to actin, cytoplasmic 2
IPI00363395.2 RAP1B Ras-related protein Rap-1b precursor
IPI00390303.4 F5 Coagulation factor V
IPI00324893.4 YWHAZ 14-3-3 protein zeta/delta
IPI00454431.1 ACTN1 Brain-specific alpha actinin 1 isoform
NI: not identified.a Ratio of the fold change of proteins in controls at 24 weeks of follow-up versus bab Ratio of the fold change of proteins in SOC-bearing rats at 24 weeks post-DMBA trc The peptides identified with 95% confidence in the reporter ion quantification.
peroxisome proliferator-activated receptor a/retinoid X receptor a(PPARa/RXRa) activation and extracellular signal-regulated ki-nase/mitogen-activated protein kinase (ERK/MAPK) signalingwere involved in the early stage of SOC, while glioma invasivenesssignaling, liver X receptor (LXR)/RXR activation, and farnesoid Xreceptor (FXR)/RXR activation were significantly related to theadvanced stage of SOC (Fig. 5).
As shown in Fig. 6, the network that contained the largestnumber of differentially expressed proteins, as indicated from theratio values from SOC-bearing rats at 12 weeks after DMBAtreatment versus matched controls, had a score of 40 and consistedof 16 focus molecules. This network was centered on ERK, p38MAPK, transforming growth factor beta (TGF beta) and nuclearfactor-kappa-B (NF-kB). Similarly, the top network, as indicatedfrom the ratio values from SOC-bearing rats at 24 weeks afterDMBA treatment versus matched controls, had a score of 47 andconsisted of 20 focus molecules, including integrins, extracellularmatrix (ECM) molecules and NF-kB, at the core (Fig. 7).
4. Comments
Early tumor detection is the most promising approach forimproving the survival rate of ovarian cancer. We used aquantitative proteomics technique to discover molecular eventsand identify serum biomarkers underlying the initiation andpromotion of carcinogenesis in a rat model of DMBA-induced SOC.
The incidence of ovarian adenocarcinoma induced by DMBA inanimal models varies from 10% to 50% due to different methods[2–4]. The most common method is to implant DMBA-coated silk
A treatment.
Ratio
116/114a
P-value Ratio
119/117b
P-value No. of
peptidesc
1.600 0.001 27.542 0.000 38
NI 22.284 0.001 9
NI 11.912 0.003 5
NI 6.194 0.017 6
1 0.488 0.001 4.920 0.038 42
4.446 0.010 3.373 0.019 9
NI 3.342 0.001 114
NI 3.048 0.002 5
NI 2.128 0.001 50
NI 1.614 0.000 378
NI 0.506 0.015 53
0.033 0.000 0.406 0.000 99
NI 0.316 0.043 5
NI 0.291 0.029 3
2.729 0.005 0.207 0.001 19
NI 0.136 0.029 4
NI 0.122 0.000 131
1.406 0.036 0.117 0.000 23
NI 0.114 0.000 45
NI 0.104 0.000 14
1.127 0.013 0.059 0.016 99
NI 0.026 0.000 20
1.472 0.023 NI 24
16.444 0.000 NI 87
0.088 0.007 NI 19
0.402 0.007 NI 65
0.179 0.001 NI 6
5.105 0.005 NI 10
0.044 0.004 NI 13
0.164 0.020 NI 5
2.228 0.044 NI 15
0.095 0.045 NI 4
0.103 0.000 NI 11
seline.
eatment versus baseline.
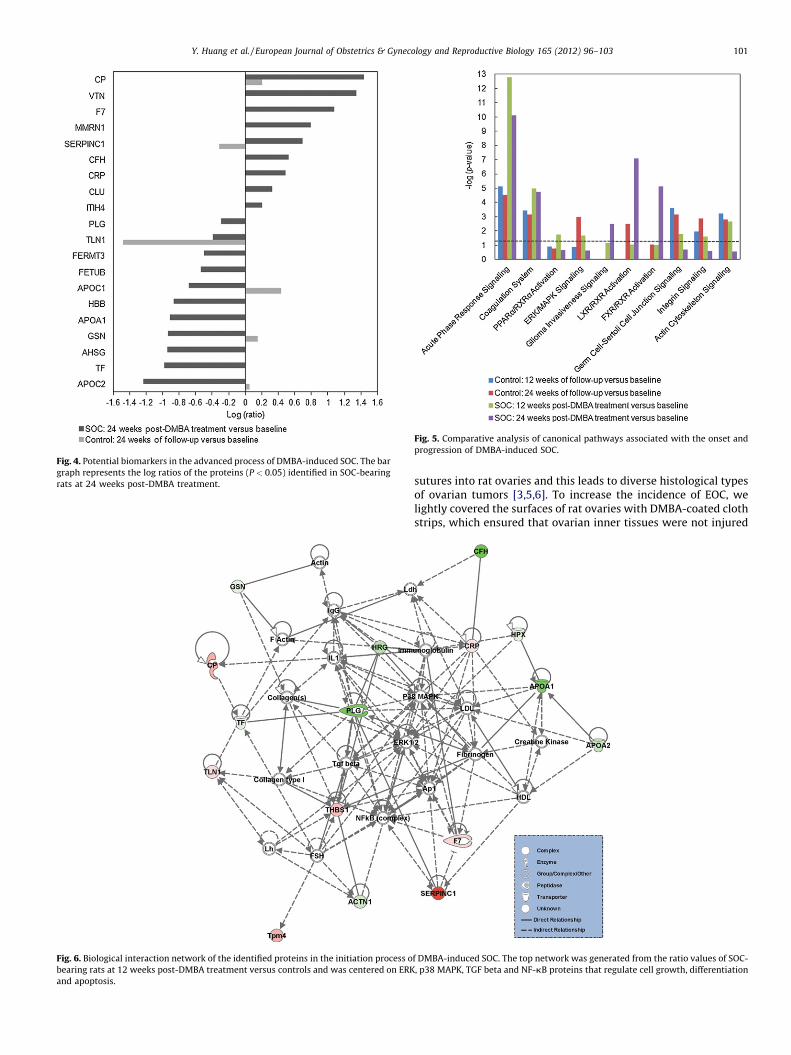
Fig. 6. Biological interaction network of the identified proteins in the initiation process o
bearing rats at 12 weeks post-DMBA treatment versus controls and was centered on ERK
and apoptosis.
Fig. 4. Potential biomarkers in the advanced process of DMBA-induced SOC. The bar
graph represents the log ratios of the proteins (P < 0.05) identified in SOC-bearing
rats at 24 weeks post-DMBA treatment.
Fig. 5. Comparative analysis of canonical pathways associated with the onset and
progression of DMBA-induced SOC.
Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103 101
sutures into rat ovaries and this leads to diverse histological typesof ovarian tumors [3,5,6]. To increase the incidence of EOC, welightly covered the surfaces of rat ovaries with DMBA-coated clothstrips, which ensured that ovarian inner tissues were not injured
f DMBA-induced SOC. The top network was generated from the ratio values of SOC-
, p38 MAPK, TGF beta and NF-kB proteins that regulate cell growth, differentiation

Fig. 7. Biological interaction network of the identified proteins in the advanced process of DMBA-induced SOC. The top network was generated from the ratio values of SOC-
bearing rats at 24 weeks post-DMBA treatment versus controls and was centered on integrins, extracellular matrix (ECM) molecules and NF-kB proteins that regulate cell
motility, attachment and cytoskeletal reorganization.
Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103102
by DMBA. In our study, the cumulative incidence of ovarian tumorsreached 75%, 94% of which were EOC. It suggested that our methodresulted in a higher incidence of EOC and could constitute a morestable rat model for EOC studies.
Because SOC is the most common type of EOC, the sera collectedfrom rats with or without SOC were analyzed. A total of 27 proteinswere significantly altered during carcinogenesis. Six proteins (CP,CRP, F7, MMRN1, SERPINC1 and TLN1) were up-regulated in bothearly and advanced stages of SOC. THBS1 levels increased duringearly carcinogenesis only, while the levels of CFH, CLU, ITIH4 andVTN were elevated until the advanced stage. Conversely, APOA1,AHSG, TF and PLG levels were down-regulated throughout DMBA-induced carcinogenesis. Eight proteins, including ACTG1, ACTN1,APOA2, CFH, GC, HPX, ITIH4 and PSMA2, were reduced during earlycarcinogenesis only, while the reductions of APOC1, APOC2,FERMT3, FETUB, HBB and GSN were only found in the advancedstages of SOC. Among all the proteins identified, 15 werepreviously reported to be involved in ovarian cancer, and 12proteins, including MMRN1, SERPINC1, TLN1, AHSG, PLG, APOA2,HPX, APOC1, APOC2, FERMT3, FETUB and HBB, were identified forthe first time in this study.
Angiogenesis is an essential process in tumor growth andprogression. Cancer cells can produce angiogenic stimulatoryfactors and antiangiogenic factors. The cleaved conformation ofSERPINC1, one of the serine protease inhibitors, has potentantiangiogenic and antitumor roles [7]. Its reduction has beenobserved in hepatocellular carcinoma and intrahepatic cholangio-carcinoma [8,9], although we observed the up-regulation ofSERPINC1 during both early and advanced SOC. It could bespeculated that the up-regulation of SERPINC1 might be caused bythe host defense against carcinogenesis.
MMRN1 is found in platelets and the endothelium of bloodvessels. Its down-regulation has been identified in lung cancer,which could contribute to blood vessel leakage and consequentlyprovide access to oxygen and nutrients for cancer tissue [10]. Wedemonstrated that MMRN1 was up-regulated during early andadvanced SOC in our DMBA-induced rat model. Further studies areneeded to investigate the relationship between MMRN1 andovarian cancer.
TLN1 plays a significant role in cell adhesion and motility.Recent reports indicate that TLN1 overexpression enhancesprostate and oral cancer invasion and metastasis by activatingsurvival signals, and TLN1 might be a potential cancer-driving genein oral cancer [11,12]. Our study showed that the levels of TLN1 inboth early and advanced stages of SOC were higher than matchedcontrols, suggesting that TLN1 has important roles in ovariancancer development.
PLG and AHSG were down-regulated throughout DMBA-induced carcinogenesis in our study. PLG is converted to plasminmainly by plasminogen activator. The components of PLGactivation system have been reported to be up-regulated in tissue,plasma or serum from patients with malignant tumors, includingovarian cancer, and may be involved in tumor invasion andmetastasis [13,14]. Our results support the speculation that thePLG activation system plays important roles in tumor progression.AHSG has been shown to be involved in breast, colorectal and lungcancer [15], but its functions during tumor progression arecontroversial [16,17]. The results in this study suggest that AHSGmight inhibit SOC progression, which is consistent with the reportby Swallow, which illustrated that AHSG inhibits intestinal tumorprogression by suppressing TGF-beta signal transduction and thetherapeutic enhancement of AHSG could benefit patients [18].

Y. Huang et al. / European Journal of Obstetrics & Gynecology and Reproductive Biology 165 (2012) 96–103 103
Apolipoproteins serve as lipid transfer carriers, enzymecofactors and receptor ligands. Recently, reports indicated arelationship between cancer and apolipoproteins. The down-regulation of serum APOA1 has been reported in ovarian cancerand it can be used as a biomarker for early detection and atherapeutic target [19]. APOA1 was also found to be down-regulated in our study. Abnormal alterations of APOA2, APOC1 andAPOC2 have been found in pancreatic, breast, papillary thyroid,colorectal, lung and stomach cancers [20–25] but there is noevidence of their changes in ovarian cancer. In our study, areduction in APOA2 was found during early-stage SOC, whilereductions in APOC1 and APOC2 were found during advanced-stage disease. These apolipoproteins might have potential roles incancer progression and be biomarkers for disease diagnosis andmonitoring.
HPX is a heme-binding glycoprotein that protects the body fromoxidative damage. It is elevated in serum or plasma from patientswith oral squamous cell carcinomas and colorectal cancer [26,27]but is down-regulated in samples from patients with lung cancer[28]. Our data indicate that HPX might negatively correlate withovarian cancer development.
FERMT3, FETUB and HBB are found to be down-regulated in thesera from SOC-bearing rats until the advanced stage of carcino-genesis. Little is known about the implication of FERMT3 andFETUB on cancer. A decrease in HBB has only been reported inthyroid cancer [29,30]. The mechanism and clinical significance ofthese proteins in ovarian cancer require further investigation.
In conclusion, we identified multiple differentially expressedproteins involved in the initiation and progression of SOC, whichcould help us understand molecular mechanisms associated withthe dynamic process of ovarian carcinogenesis, and these proteinscould be used as diagnostic biomarkers for early detection, diseasemonitoring and therapeutic targets. Although further validation andevaluation in cancer patients are needed, our findings provide usefulinformation for basic and translational research in ovarian cancer.
Acknowledgments
This work was supported by funding from Shanghai LeadingAcademic Discipline Project (No. B117), National High-tech R&DProgram (No. 2006AA02Z342), Shanghai Science Committee em-phasis project (No. 07JC14006), National Natural Sciences Founda-tion of China (No. 30901588), and Rising-Star Program of ShanghaiScience and Technology Commission (No. 10QA1401000).
References
[1] Lutz AM, Willmann JK, Drescher CW, et al. Early diagnosis of ovarian carcino-ma: is a solution in sight? Radiology 2011;259:329–45.
[2] Sekiya S, Endoh N, Kikuchi Y, et al. In vivo and in vitro studies of experimentalovarian adenocarcinoma in rats. Cancer Research 1979;39:1108–12.
[3] Nishida T, Sugiyama T, Kataoka A, Ushijima K, Yakushiji M. Histologic charac-terization of rat ovarian carcinoma induced by intraovarian insertion of a 7,12-dimethylbenz[a]anthracene-coated suture: common epithelial tumors of theovary in rats? Cancer 1998;83:965–70.
[4] Crist KA, Zhang Z, You M, et al. Characterization of rat ovarian adenocarcino-mas developed in response to direct instillation of 7,12-dimethylbenz[a]an-thracene (DMBA) coated suture. Carcinogenesis 2005;26:951–7.
[5] Tunca JC, Erturk E, Erturk E, Bryan GT. Chemical induction of ovarian tumors inrats. Gynecologic Oncology 1985;21:54–64.
[6] Kim KK, Shim JC, Kim JR. Overexpression of p21, cyclin E and decreasedexpression of p27 in DMBA (7,12-dimethylbenzanthracene)-induced rat ovar-ian carcinogenesis. Pathology International 2003;53:291–6.
[7] O’Reilly MS, Pirie-Shepherd S, Lane WS, Folkman J. Antiangiogenic activity ofthe cleaved conformation of the serpin antithrombin. Science 1999;285:1926–8.
[8] Wang AG, Yoon SY, Oh JH, et al. Identification of intrahepatic cholangiocarci-noma related genes by comparison with normal liver tissues using expressedsequence tags. Biochemical and Biophysical Research Communications2006;345:1022–32.
[9] Yoon SY, Kim JM, Oh JH, et al. Gene expression profiling of human HBV- and/orHCV-associated hepatocellular carcinoma cells using expressed sequence tags.International Journal of Oncology 2006;29:315–27.
[10] Valk K, Vooder T, Kolde R, et al. Gene expression profiles of non-small cell lungcancer: survival prediction and new biomarkers. Oncology-Basel2010;79:283–92.
[11] Sakamoto S, McCann RO, Dhir R, Kyprianou N. Talin-1 promotes tumorinvasion and metastasis via focal adhesion signaling and anoikis resistance.Cancer Research 2010;70:1885–95.
[12] Lai MT, Hua CH, Tsai MH, et al. Talin-1 overexpression defines high risk foraggressive oral squamous cell carcinoma and promotes cancer metastasis.Journal of Pathology 2011;224:367–76.
[13] Kwaan HC, McMahon B. The role of plasminogen–plasmin system in cancer.Cancer Treatment and Research 2009;148:43–66.
[14] Murthi P, Barker G, Nowell CJ, et al. Plasminogen fragmentation and increasedproduction of extracellular matrix-degrading proteinases are associated withserous epithelial ovarian cancer progression. Gynecologic Oncology2004;92:80–8.
[15] Dowling P, Clarke C, Hennessy K, et al. Analysis of acute phase proteins, AHSG,C3, CLI, HP and SAA reveals distinctive expression patterns associated withbreast, colorectal and lung cancer. International Journal of Cancer2012;131:911–23.
[16] Guillory B, Sakwe AM, Saria M, et al. Lack of fetuin-A (alpha2-HS-glycoprotein)reduces mammary tumor incidence and prolongs tumor latency via thetransforming growth factor-beta signaling pathway in a mouse model ofbreast cancer. American Journal of Pathology 2010;177:2635–44.
[17] Kundranda MN, Henderson M, Carter KJ, et al. The serum glycoprotein fetuin-Apromotes Lewis lung carcinoma tumorigenesis via adhesive-dependent andadhesive-independent mechanisms. Cancer Research 2005;65:499–506.
[18] Swallow CJ, Partridge EA, Macmillan JC, et al. Alpha2hs-glycoprotein, anantagonist of transforming growth factor beta in vivo, inhibits intestinal tumorprogression. Cancer Research 2004;64:6402–9.
[19] Su F, Kozak KR, Imaizumi S, et al. Apolipoprotein A-I (apoA-I) and apoA-Imimetic peptides inhibit tumor development in a mouse model of ovariancancer. Proceedings of the National Academy of Sciences of the United Statesof America 2010;107:19997–20002.
[20] Fan Y, Shi L, Liu Q, et al. Discovery and identification of potential biomarkers ofpapillary thyroid carcinoma. Molecular Cancer 2009;8:79.
[21] Fan Y, Wang J, Yang Y, et al. Detection and identification of potential biomark-ers of breast cancer. Journal of Cancer Research and Clinical Oncology2010;136:1243–54.
[22] Engwegen JY, Depla AC, Smits ME, et al. Detection of colorectal cancer byserum and tissue protein profiling: a prospective study in a population at risk.Biomark Insights 2008;3:375–85.
[23] Yang Y, Zhao S, Fan Y, et al. Detection and identification of potential biomark-ers of non-small cell lung cancer. Technology in Cancer Research and Treat-ment 2009;8:455–66.
[24] Cohen M, Yossef R, Erez T, et al. Serum apolipoproteins C-I and C-III arereduced in stomach cancer patients: results from MALDI-based peptidomeand immuno-based clinical assays. PLoS ONE 2011;6(1):e14540.
[25] Chen J, Anderson M, Misek DE, Simeone DM, Lubman DM. Characterization ofapolipoprotein and apolipoprotein precursors in pancreatic cancer serumsamples via two-dimensional liquid chromatography and mass spectrometry.Journal of Chromatography A 2007;1162:117–25.
[26] Bijian K, Mlynarek AM, Balys RL, et al. Serum proteomic approach for theidentification of serum biomarkers contributed by oral squamous cell carci-noma and host tissue microenvironment. Journal of Proteome Research2009;8:2173–85.
[27] Ang CS, Rothacker J, Patsiouras H, Burgess AW, Nice EC. Murine fecal proteo-mics: a model system for the detection of potential biomarkers for colorectalcancer. Journal of Chromatography A 2010;1217:3330–40.
[28] Dowling P, O’Driscoll L, Meleady P, et al. 2-D difference gel electrophoresis ofthe lung squamous cell carcinoma versus normal sera demonstrates consis-tent alterations in the levels of ten specific proteins. Electrophoresis2007;28:4302–10.
[29] Onda M, Akaishi J, Asaka S, et al. Decreased expression of haemoglobin beta(HBB) gene in anaplastic thyroid cancer and recovery of its expression inhibitscell growth. British Journal of Cancer 2005;92:2216–24.
[30] Jarzab B, Wiench M, Fujarewicz K, et al. Gene expression profile of papillarythyroid cancer: sources of variability and diagnostic implications. CancerResearch 2005;65:1587–97.