discovery and refinement of loci associated with lipid ... · discovery and refinement of loci...
TRANSCRIPT

GLGC Metabochip Supplement 2013
1
Discovery and Refinement of Loci Associated with Lipid Levels
Supplementary Information
The Global Lipids Genetics Consortium
Correspondence To:
Cristen Willer
Division of Cardiovascular Medicine and Department of Human Genetics
University of Michigan Medical School
Ann Arbor, MI
Karen Mohlke
Department of Genetics
University of North Carolina Medical School
Chapel Hill, NC
Erik Ingelsson
Department of Medical Sciences, Molecular Epidemiology
Uppsala University
Uppsala, Sweden
Gonçalo Abecasis
Department of Biostatistics
University of Michigan School of Public Health
Ann Arbor, MI
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
2
Cristen J. Willer1-4*†; Ellen M. Schmidt2*; Sebanti Sengupta4*; Gina M. Peloso5-7; Stefan Gustafsson8,9; Stavroula Kanoni10;
Andrea Ganna8,9,11; Jin Chen4; Martin L. Buchkovich12; Samia Mora13,14; Jacques S. Beckmann15,16; Jennifer L. Bragg-
Gresham4; Hsing-Yi Chang17; Ayşe Demirkan18; Heleen M. Den Hertog19; Ron Do6; Louise A. Donnelly20; Georg B.
Ehret21,22; Tõnu Esko7,23,24 ; Mary F. Feitosa25; Teresa Ferreira26; Krista Fischer23; Pierre Fontanillas7; Ross M. Fraser27;
Daniel F. Freitag28; Deepti Gurdasani10,28; Kauko Heikkilä29; Elina Hyppönen30; Aaron Isaacs18,31; Anne U. Jackson4; Åsa
Johansson32,33; Toby Johnson34,35; Marika Kaakinen36,37; Johannes Kettunen38,39; Marcus E. Kleber40,41; Xiaohui Li42; Jian'an
Luan43; Leo-Pekka Lyytikäinen44,45; Patrik K.E. Magnusson11; Massimo Mangino46; Evelin Mihailov23,24; May E.
Montasser47; Martina Müller-Nurasyid48-50; Ilja M. Nolte51; Jeffrey R. O'Connell47; Cameron D. Palmer7,52,53; Markus
Perola23,38,39; Ann-Kristin Petersen48; Serena Sanna54; Richa Saxena 55; Susan K. Service56; Sonia Shah57; Dmitry Shungin58-
60; Carlo Sidore4,54,61; Ci Song8,9,11; Rona J. Strawbridge62,63 ; Ida Surakka38,39; Toshiko Tanaka64; Tanya M. Teslovich4;
Gudmar Thorleifsson65; Evita G. Van den Herik19; Benjamin F. Voight66,67; Kelly A. Volcik68; Lindsay L. Waite69; Andrew
Wong70; Ying Wu12; Weihua Zhang71,72; Devin Absher69; Gershim Asiki73; Inês Barroso10,74; Latonya F. Been75; Jennifer L.
Bolton27; Lori L Bonnycastle76; Paolo Brambilla77; Mary S. Burnett78; Giancarlo Cesana79; Maria Dimitriou80; Alex S.F.
Doney20; Angela Döring81,82; Paul Elliott37,83; Stephen E. Epstein78; Gudmundur Ingi Eyjolfsson84; Bruna Gigante85; Mark O.
Goodarzi86; Harald Grallert87; Martha L. Gravito75; Christopher J. Groves88; Göran Hallmans89; Anna-Liisa Hartikainen90;
Caroline Hayward91; Dena Hernandez92; Andrew A. Hicks93; Hilma Holm65; Yi-Jen Hung94; Thomas Illig87,95; Michelle R.
Jones86; Pontiano Kaleebu73; John J.P. Kastelein96; Kay-Tee Khaw97; Eric Kim42; Norman Klopp87,95; Pirjo Komulainen98;
Meena Kumari57; Claudia Langenberg43; Terho Lehtimäki44,45; Shih-Yi Lin99; Jaana Lindström100; Ruth J.F. Loos43,101-103;
François Mach21; Wendy L McArdle104; Christa Meisinger81; Braxton D. Mitchell47; Gabrielle Müller105; Ramaiah
Nagaraja106; Narisu Narisu76; Tuomo V.M. Nieminen107-109; Rebecca N. Nsubuga73; Isleifur Olafsson110; Ken K. Ong43,70;
Aarno Palotie38,111,112; Theodore Papamarkou10,28,113; Cristina Pomilla10,28; Anneli Pouta90,114; Daniel J. Rader115,116;
Muredach P. Reilly115,116; Paul M. Ridker13,14; Fernando Rivadeneira117,118, 119; Igor Rudan27; Aimo Ruokonen120; Nilesh
Samani121,122; Hubert Scharnagl123; Janet Seeley73,124; Kaisa Silander38,39; Alena Stančáková125; Kathleen Stirrups 10; Amy J.
Swift76; Laurence Tiret 126; Andre G. Uitterlinden117-119; L. Joost van Pelt127,128; Sailaja Vedantam7,52,53; Nicholas
Wainwright10,28; Cisca Wijmenga128,129; Sarah H. Wild27; Gonneke Willemsen130; Tom Wilsgaard131; James F. Wilson27;
Elizabeth H. Young10,28; Jing Hua Zhao43; Linda S. Adair132; Dominique Arveiler133; Themistocles L. Assimes134; Stefania
Bandinelli135; Franklyn Bennett136; Murielle Bochud137; Bernhard O. Boehm138,139; Dorret I. Boomsma130; Ingrid B.
Borecki25; Stefan R. Bornstein140; Pascal Bovet137,141; Michel Burnier142; Harry Campbell27; Aravinda Chakravarti22; John C.
Chambers71,72,143; Yii-Der Ida Chen144,145; Francis S. Collins76; Richard S. Cooper146; John Danesh28; George Dedoussis80; Ulf
de Faire85; Alan B. Feranil147; Jean Ferrières148; Luigi Ferrucci64; Nelson B. Freimer56,149; Christian Gieger48; Leif C.
Groop150,151; Vilmundur Gudnason152; Ulf Gyllensten32; Anders Hamsten62,63,153 ; Tamara B. Harris 154; Aroon Hingorani57;
Joel N. Hirschhorn7,52,53; Albert Hofman117,119; G. Kees Hovingh96; Chao Agnes Hsiung155; Steve E. Humphries156; Steven C.
Hunt157; Kristian Hveem158; Carlos Iribarren159; Marjo-Riitta Järvelin36,37,83,114,160; Antti Jula161; Mika Kähönen162; Jaakko
Kaprio29,38,163; Antero Kesäniemi 164; Mika Kivimaki57; Jaspal S. Kooner72,143,165; Peter J. Koudstaal19; Ronald M. Krauss166;
Diana Kuh70; Johanna Kuusisto167; Kirsten O. Kyvik168,169; Markku Laakso167; Timo A. Lakka98,170; Lars Lind171; Cecilia M.
Lindgren26; Nicholas G. Martin172; Winfried März41,123,173; Mark I. McCarthy26,88; Colin A. McKenzie174; Pierre Meneton175;
Andres Metspalu23,24; Leena Moilanen176; Andrew D. Morris20; Patricia B. Munroe34,35; Inger Njølstad131; Nancy L.
Pedersen11; Chris Power30; Peter P. Pramstaller93,177,178; Jackie F. Price27; Bruce M. Psaty179,180; Thomas Quertermous134;
Rainer Rauramaa98,181; Danish Saleheen28,182,183; Veikko Salomaa184; Dharambir K. Sanghera75; Jouko Saramies185; Peter
E.H. Schwarz140,186; Wayne H-H Sheu187; Alan R. Shuldiner47,188; Agneta Siegbahn8,33,171; Tim D. Spector46; Kari
Stefansson65,189; David P. Strachan190; Bamidele O. Tayo146; Elena Tremoli191; Jaakko Tuomilehto100,192-194; Matti
Uusitupa195,196; Cornelia M. van Duijn18,31; Peter Vollenweider197; Lars Wallentin33,171; Nicholas J. Wareham43; John B.
Whitfield172; Bruce H.R. Wolffenbuttel128,198; Jose M. Ordovas199-201; Eric Boerwinkle68; Colin N.A. Palmer20; Unnur
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
3
Thorsteinsdottir65,189; Daniel I. Chasman13,14; Jerome I. Rotter42; Paul W. Franks58,60,202; Samuli Ripatti10,38,39; L. Adrienne
Cupples5,203; Manjinder S. Sandhu10,28; Stephen S. Rich204; Michael Boehnke4*; Panos Deloukas10*; Sekar
Kathiresan6,7,205,206*; Karen L. Mohlke12*†; Erik Ingelsson8,9,26*†; Gonçalo R. Abecasis4*†
* Denotes equal contribution
† Corresponding authors
1. Department of Internal Medicine, Division of Cardiovascular Medicine, University of Michigan, Ann Arbor, Michigan 48109, USA.
2. Department of Computational Medicine and Bioinformatics, University of Michigan, Ann Arbor, Michigan 48109, USA.
3. Department of Human Genetics, University of Michigan, Ann Arbor, Michigan 48109, USA. 4. Center for Statistical Genetics, Department of Biostatistics, University of Michigan, Ann Arbor, Michigan 48109,
USA. 5. Department of Biostatistics, Boston University School of Public Health, Boston, Massachusetts 02118, USA. 6. Center for Human Genetic Research, Massachusetts General Hospital, Boston, Massachusetts 02114, USA. 7. Broad Institute, Program in Medical and Population Genetics, Cambridge, Massachusetts 02142, USA. 8. Department of Medical Sciences, Molecular Epidemiology, Uppsala University, Uppsala, Sweden. 9. Science for Life Laboratory, Uppsala University, Uppsala, Sweden. 10. Wellcome Trust Sanger Institute, Wellcome Trust Genome Campus, CB10 1SA, Hinxton, United Kingdom. 11. Department of Medical Epidemiology and Biostatistics, Karolinska Institutet, Stockholm, Sweden. 12. Department of Genetics, University of North Carolina, Chapel Hill, NC 27599 USA. 13. Division of Preventive Medicine, Brigham and Women's Hospital, 900 Commonwealth Ave., Boston MA 02215,
USA. 14. Harvard Medical School, Boston MA 02115, USA. 15. Service of Medical Genetics, Lausanne University Hospital, Lausanne, Switzerland. 16. Department of Medical Genetics, University of Lausanne, Lausanne, Switzerland. 17. Division of Preventive Medicine and Health Services Research, Institute of Population Health Sciences, National
Health Research Institutes, Zhunan, Taiwan. 18. Genetic Epidemiology Unit, Department of Epidemiology, Erasmus University Medical Center, Rotterdam, The
Netherlands. 19. Department of Neurology, Erasmus Medical Center, Rotterdam, The Netherlands. 20. Medical Research Institute, University of Dundee, Ninewells Hospital and Medical School. Dundee, DD1 9SY,
United Kingdom. 21. Cardiology, Department of Specialities of Medicine, Geneva University Hospital, Rue Gabrielle-Perret-Gentil 4,
1211 Geneva 14, Switzerland. 22. Center for Complex Disease Genomics, McKusick-Nathans Institute of Genetic Medicine, Johns Hopkins
University School of Medicine, Baltimore, MD 21205, USA. 23. Estonian Genome Center of the University of Tartu, Tartu, Estonia. 24. Institute of Molecular and Cell Biology, University of Tartu, Tartu, Estonia. 25. Department of Genetics, Washington University School of Medicine, USA. 26. Wellcome Trust Centre for Human Genetics, University of Oxford, Oxford, OX3 7BN, United Kingdom. 27. Centre for Population Health Sciences, University of Edinburgh, Teviot Place, Edinburgh, EH8 9AG, Scotland,
United Kingdom. 28. Department of Public Health and Primary Care, University of Cambridge, Cambridge, United Kingdom. 29. Hjelt Institute, Department of Public Health, University of Helsinki, Finland. 30. Centre For Paediatric Epidemiology and Biostatistics/MRC Centre of Epidemiology for Child Health, University
College of London Institute of Child Health, London, United Kingdom.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
4
31. Centre for Medical Systems Biology, Leiden, the Netherlands. 32. Department of Immunology, Genetics and Pathology, Uppsala University, Uppsala, Sweden. 33. Uppsala Clinical Research Center, Uppsala University, Uppsala, Sweden. 34. Genome Centre, Barts and The London School of Medicine and Dentistry, Queen Mary University of London,
London, UK. 35. Clinical Pharmacology, NIHR Cardiovascular Biomedical Research Unit, William Harvey Research Institute, Barts
and The London School of Medicine and Dentistry Queen Mary University of London, London, UK. 36. Biocenter Oulu, University of Oulu, Oulu, Finland. 37. Institute of Health Sciences, University of Oulu, Finland. 38. Institute for Molecular Medicine Finland FIMM, University of Helsinki, Finland. 39. Public Health Genomics Unit, National Institute for Health and Welfare, Helsinki, Finland. 40. Department of Internal Medicine II – Cardiology, University of Ulm Medical Centre, Ulm, Germany. 41. Mannheim Institute of Public Health, Social and Preventive Medicine, Medical Faculty of Mannheim, University
of Heidelberg, Ludolf-Krehl-Strasse 7-11, 68167 Mannheim, Germany. 42. Medical Genetics Institute, Cedars-Sinai Medical Center, Los Angeles, CA 90048, USA. 43. MRC Epidemiology Unit, Institute of Metabolic Science, Box 285, Addenbrooke's Hospital, Hills Road, Cambridge,
CB2 0QQ, United Kingdom. 44. Department of Clinical Chemistry, Fimlab Laboratories, Tampere 33520, Finland. 45. Department of Clinical Chemistry, University of Tampere School of Medicine, Tampere 33014, Finland. 46. Department of Twin Research and Genetic Epidemiology, King’s College London, London, United Kingdom. 47. Division of Endocrinology, Diabetes, and Nutrition, Department of Medicine, University of Maryland, School of
Medicine, Baltimore, Maryland. 48. Institute of Genetic Epidemiology, Helmholtz Zentrum München, Neuherberg 85764, Germany. 49. Department of Medicine I, University Hospital Grosshadern, Ludwig-Maximilians University, Munich, Germany. 50. Institute of Medical Informatics, Biometry and Epidemiology, Ludwig-Maximilians-University of Munich, Munich,
Germany. 51. Department of Epidemiology, University of Groningen, University Medical Center Groningen, The Netherlands. 52. Division of Endocrinology, Children's Hospital Boston, Massachusetts 02115, USA. 53. Division of Genetics, Program in Genomics, Children's Hospital, Boston, Massachusetts 02115, USA. 54. Istituto di Ricerca Genetica e Biomedica, CNR, Monserrato, 09042, Italy. 55. Massachusetts General Hospital/ Broad Institute, Harvard University, Cambridge, MA, USA. 56. Center for Neurobehavioral Genetics, The Semel Institute for Neuroscience and Human Behavior, University of
California, Los Angeles, USA. 57. Genetic Epidemiology Group, Deparment of Epidemiology and Public Health, UCL, London WC1E 6BT, United
Kingdom. 58. Department of Clinical Sciences, Genetic & Molecular Epidemiology Unit, Lund University Diabetes Center,
Scania University Hosptial, Malmö, Sweden. 59. Department of Odontology, Umeå University, Umeå, Sweden. 60. Department of Public Health and Primary Care, Unit of Medicine, Umeå University, Umeå, Sweden. 61. Dipartimento di Scienze Biomediche, Universita di Sassari, 07100 SS, Italy. 62. Atherosclerosis Research Unit, Department of Medicine Solna, Karolinska University Hospital, Karolinska
Institutet, Stockholm, Sweden. 63. Center for Molecular Medicine, Karolinska University Hospital, Stockholm, Sweden. 64. Clinical Research Branch, National Institute Health, Baltimore, MD, USA. 65. deCODE Genetics/Amgen, 101 Reykjavik, Iceland. 66. Department of Genetics, University of Pennsylvania - School of Medicine, Philadelphia PA, 19104, USA. 67. Department of Systems Pharmacology and Translational Therapeutics, University of Pennsylvania - School of
Medicine, Philadelphia PA, 19104, USA.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
5
68. Human Genetics Center, University of Texas Health Science Center - School of Public Health, Houston, TX 77030, USA.
69. HudsonAlpha Institute for Biotechnology, Huntsville, AL, USA. 70. MRC Unit for Lifelong Health and Ageing, 33 Bedford Place, London, WC1B 5JU, United Kingdom. 71. Department of Epidemiology and Biostatistics, School of Public Health, Imperial College London, London, United
Kingdom. 72. Ealing Hospital, Southall, Middlesex UB1 3HW, United Kingdom. 73. MRC/UVRI Uganda Research Unit on AIDS, Entebbe, Uganda. 74. University of Cambridge Metabolic Research Laboratories and NIHR Cambridge Biomedical Research Centre,
Level 4, Institute of Metabolic Science Box 289 Addenbrooke’s Hospital Cambridge CB2 OQQ, UK. 75. Department of Pediatrics, University of Oklahoma Health Sciences Center, Oklahoma City, OK, USA. 76. Genome Technology Branch, National Human Genome Research Institute, NIH, Bethesda, MD 20892, USA. 77. Department of Experimental Medicine, University of Milano Bicocca, Italy. 78. MedStar Health Research Institute, 6525 Belcrest Road, Suite 700, Hyattsville, MD 20782, USA. 79. Research Centre on Public Health, University of Milano Bicocca, Italy. 80. Department of Dietetics-Nutrition, Harokopio University, 70 El. Venizelou Str, Athens, Greece. 81. Institute of Epidemiology I, Helmholtz Zentrum München, Neuherberg 85764, Germany. 82. Institute of Epidemiology II, Helmholtz Zentrum München, Neuherberg 85764, Germany. 83. Department of Epidemiology and Biostatistics, MRC Health Protection Agency (HPA) Centre for Environment and
Health, School of Public Health, Imperial College London, UK. 84. The Laboratory in Mjodd, 108 Reykjavik, Iceland. 85. Division of Cardiovascular Epidemiology, Institute of Environmental Medicine, Karolinska Institutet, Stockholm,
Sweden. 86. Division of Endocrinology, Diabetes and Metabolism, Department of Medicine, Cedars-Sinai Medical Center, Los
Angeles, CA 90048, USA. 87. Research Unit of Molecular Epidemiology, Helmholtz Zentrum München, Neuherberg 85764, Germany. 88. Oxford Centre for Diabetes, Endocrinology and Metabolism, University of Oxford, OX3 7LJ, United Kingdom. 89. Department of Public Health and Clinical Medicine, Nutritional research, Umeå University, Umeå, Sweden. 90. Department of Clinical Sciences/Obstetrics and Gynecology, Oulu University Hospital, Oulu, Finland. 91. MRC Human Genetics Unit, Institute of Genetics and Molecular Medicine, Western General Hospital, Edinburgh,
Scotland, United Kingdom. 92. Laboratory of Neurogenetics, National Institute on Aging, Bethesda, MD 20892, USA. 93. Center for Biomedicine, European Academy Bozen/Bolzano (EURAC), Bolzano, Italy - Affiliated Institute of the
University of Lübeck, Lübeck, Germany. 94. Division of Endocrinology & Metabolism, Tri-Service General Hospital, National Defense Medical Center, Taipei,
Taiwan. 95. Hannover Unified Biobank, Hannover Medical School, Hannover 30625, Germany. 96. Department of Vascular Medicine, Academic Medical Center, Amsterdam, The Netherlands. 97. Clinical Gerontology Unit, University of Cambridge, Cambridge, United Kingdom. 98. Kuopio Research Institute of Exercise Medicine, Kuopio, Finland. 99. Division of Endocrine and Metabolism, Department of Internal Medicine, Taichung Veterans General
Hospital, School of Medicine, National Yang-Ming University, Taipei, Taiwan. 100. Diabetes Prevention Unit, National Institute for Health and Welfare, 00271 Helsinki, Finland. 101. The Genetics of Obesity and Related Metabolic Traits Program, The Icahn School of Medicine at Mount Sinai,
New York, USA. 102. The Charles Bronfman Institute for Personalized Medicine, The Icahn School of Medicine at Mount Sinai, New
York, USA. 103. The Mindich Child Health and Development Institute, The Icahn School of Medicine at Mount Sinai, New York.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
6
104. School of Social and Community Medicine, University of Bristol, Oakfield House, Oakfield Grove, Bristol BS8 2BN, United Kingdom.
105. Institute for Medical Informatics and Biometrics, University of Dresden, Medical Faculty Carl Gustav Carus, Fetscherstrasse 74, 01307 Dresden, Germany.
106. Laboratory of Genetics, National Institute on Aging, Baltimore, MD21224, USA. 107. Department of Clinical Pharmacology, University of Tampere School of Medicine, Tampere 33014, Finland. 108. Department of Internal Medicine, Päijät-Häme Central Hospital, Lahti, Finland. 109. Division of Cardiology, Helsinki University Central Hospital, Helsinki, Finland. 110. Department of Clinical Biochemistry, Landspitali University Hospital, 101 Reykjavik, Iceland. 111. Department of Medical Genetics, Haartman Institute, University of Helsinki and Helsinki University Central
Hospital, Helsinki, Finland. 112. Genetic Epidemiology Group, Wellcome Trust Sanger Institute, Hinxton, Cambridge, United Kingdom. 113. Department of Statistical Sciences, University College of London, London, United Kingdom. 114. National Institute for Health and Welfare, Oulu, Finland. 115. Cardiovascular Institute, Perelman School of Medicine at the University of Pennsylvania, 3400 Civic Center Blvd,
Building 421, Translational Research Center, Philadelphia, PA 19104-5158, USA. 116. Division of Translational Medicine and Human Genetics, Perelman School of Medicine at the University of
Pennsylvania, 3400 Civic Center Blvd, Building 421, Translational Research Center, Philadelphia, PA 19104-5158, USA.
117. Department of Epidemiology, Erasmus University Medical Center, Rotterdam, the Netherlands. 118. Department of Internal Medicine, Erasmus University Medical Center, Rotterdam, the Netherlands. 119. Netherlands Genomics Initiative (NGI)-sponsored Netherlands Consortium for Healthy Aging (NCHA), Leiden, The
Netherlands. 120. Department of Clinical Sciences/Clinical Chemistry, University of Oulu, Oulu, Finland. 121. National Institute for Health Research Leicester Cardiovascular Biomedical Research Unit, Glenfield Hospital,
Leicester LE3 9QP, UK. 122. Department of Cardiovascular Sciences, University of Leicester, Glenfield Hospital, Leicester, LE3 9QP, UK 123. Clinical Institute of Medical and Chemical Laboratory Diagnostics, Medical University of Graz, Austria. 124. School of International Development, University of East Anglia, Norwich NR4 7TJ, United Kingdom. 125. University of Eastern Finland and Kuopio University Hospital, 70210 Kuopio, Finland. 126. INSERM UMRS 937, Pierre and Marie Curie University, Paris, France. 127. Department of Laboratory Medicine, University of Groningen, University Medical Center Groningen, The
Netherlands. 128. LifeLines Cohort Study, University of Groningen, University Medical Center Groningen, The Netherlands. 129. Department of Genetics, University of Groningen, University Medical Center Groningen, The Netherlands. 130. Department of Biological Psychology, VU Univ, Amsterdam, The Netherlands. 131. Department of Community Medicine, Faculty of Health Sciences, University of Tromsø, Tromsø, Norway. 132. Department of Nutrition, University of North Carolina, Chapel Hill, NC, USA. 133. Department of Epidemiology and Public Health, EA 3430, University of Strasbourg, Faculty of Medicine,
Strasbourg, France. 134. Department of Medicine, Stanford University School of Medicine, Stanford, CA, USA. 135. Geriatric Unit, Azienda Sanitaria Firenze (ASF), Florence, Italy. 136. Chemical Pathology, Department of Pathology, University of the West Indies, Mona, Kingston 7, Jamaica. 137. Institute of Social and Preventive Medicine (IUMSP), Lausanne University Hospital, Route de la Corniche 10,
1010 Lausanne, Switzerland. 138. Division of Endocrinology and Diabetes, Department of Internal Medicine, Ulm University Medical Centre, Ulm,
Germany. 139. Lee Kong Chian School of Medicine, Nanyang Technological University, Singapore.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
7
140. Department of Medicine III, University of Dresden, Medical Faculty Carl Gustav Carus, Fetscherstrasse 74, 01307 Dresden, Germany.
141. Ministry of Health, Victoria, Republic of Seychelles. 142. Service of Nephrology, Lausanne University Hospital, Lausanne, Switzerland. 143. Imperial College Healthcare NHS Trust, London, United Kingdom. 144. Division of Reproductive Endocrinology, Department of Obstetrics and Gynecology, Cedars-Sinai Medical Center,
Los Angeles, California, USA. 145. Department of Medicine, University of California Los Angeles, Los Angeles, California, USA. 146. Department of Preventive Medicine and Epidemiology, Loyola University Medical School, Maywood, Illinois
60153, USA. 147. Office of Population Studies Foundation, University of San Carlos, Talamban, Cebu City, Philippines. 148. Department of Cardiology, Toulouse University School of Medicine, Rangueil Hospital, Toulouse, France. 149. Department of Psychiatry, University of California, Los Angeles, USA. 150. Department of Clinical Sciences, Lund University, SE-20502, Malmö, Sweden. 151. Department of Medicine, Helsinki University Hospital, FI-00029 Helsinki, Finland. 152. Icelandic Heart Association, Kopavogur, Iceland. 153. Department of Cardiology, Karolinska University Hospital, Stockholm, Sweden. 154. Laboratory of Epidemiology, Demography, and Biometry, National Institute on Ageing, Bethesda, MD, USA. 155. Institute of Population Health Sciences, National Health Research Institutes, Zhunan, Taiwan. 156. Cardiovascular Genetics, BHF Laboratories, Institute Cardiovascular Science, University College London, London,
United Kingdom. 157. Cardiovascular Genetics, University of Utah School of Medicine, Salt Lake City, UT, USA. 158. HUNT Research Centre, Department of Public Health and General Practice, Norwegian University of Science and
Technology, Levanger, Norway. 159. Kaiser Permanente, Division of Research, Oakland, CA, USA. 160. Unit of Primary Care, Oulu University Hospital, Oulu, Finland 161. Department of Chronic Disease Prevention, National Institute for Health and Welfare, Turku, Finland. 162. Department of Clinical Physiology, University of Tampere School of Medicine, Tampere 33014, Finland. 163. Department of Mental Health and Substance Abuse Services, National Institute for Health and Welfare, Helsinki,
Finland. 164. Institute of Clinical Medicine, Department of Medicine, University of Oulu and Clinical Research Center, Oulu
University Hospital, Oulu, Finland. 165. National Heart & Lung Institute, Imperial College London, Hammersmith Hospital, London, United Kingdom. 166. Children's Hospital Oakland Research Institute, 5700 Martin Luther King Junior Way, Oakland, CA 94609, USA. 167. Department of Medicine, University of Eastern Finland and Kuopio University Hospital, 70210 Kuopio, Finland. 168. Institute of Regional Health Services Research, University of Southern Denmark, Odense, Denmark. 169. Odense Patient data Explorative Network (OPEN), Odense University Hospital, Odense, Denmark. 170. Institute of Biomedicine/Physiology, University of Eastern Finland, Kuopio Campus, Finland. 171. Department of Medical Sciences, Uppsala University, Uppsala, Sweden. 172. Queensland Institute of Medical Research, Locked Bag 2000, Royal Brisbane Hospital, Queensland 4029,
Australia. 173. Synlab Academy, Synlab Services GmbH,Gottlieb-Daimler-Straße 25, 68165 Mannheim, Germany. 174. Tropical Metabolism Research Unit, Tropical Medicine Research Institute, University of the West Indies, Mona,
Kingston 7, Jamaica. 175. U872 Institut National de la Santé et de la Recherche Médicale, Centre de Recherche des Cordeliers, 75006
Paris, France. 176. Department of Medicine, Kuopio University Hospital, Kuopio, Finland. 177. Department of Neurology, General Central Hospital, Bolzano, Italy. 178. Department of Neurology, University of Lübeck, Lübeck, Germany.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
8
179. Cardiovascular Health Research Unit, Departments of Medicine, Epidemiology, and Health Services, University of Washington, Seattle, WA, USA.
180. Group Health Research Institute, Group Health Cooperative, Seattle, WA, USA. 181. Department of Clinical Physiology and Nuclear Medicine, Kuopio University Hospital, Kuopio, Finland. 182. Center for Non-Communicable Diseases, Karachi, Pakistan. 183. Department of Medicine, University of Pennsylvania, USA. 184. Unit of Chronic Disease Epidemiology and Prevention, National Institute for Health and Welfare, Helsinki,
Finland. 185. South Karelia Central Hospital, Lappeenranta, Finland. 186. Paul Langerhans Institute Dresden, German Center for Diabetes Research (DZD), Dresden, Germany. 187. Division of Endocrine and Metabolism, Department of Internal Medicine, Taichung Veterans General Hospital,
Taichung, Taiwan. 188. Geriatric Research and Education Clinical Center, Veterans Administration Medical Center, Baltimore, Maryland. 189. Faculty of Medicine, University of Iceland, 101 Reykjavík, Iceland. 190. Division of Population Health Sciences and Education, St George's, University of London, Cranmer Terrace,
London SW17 0RE, United Kingdom. 191. Department of Pharmacological Sciences, University of Milan, Monzino Cardiology Center, IRCCS, Milan, Italy. 192. Centre for Vascular Prevention, Danube-University Krems, 3500 Krems, Austria. 193. King Abdulaziz University, Faculty of Medicine, Jeddah 21589, Saudi Arabia. 194. Red RECAVA Grupo RD06/0014/0015, Hospital Universitario La Paz, 28046. 195. Institute of Public Health and Clinical Nutrition, University of Eastern Finland, Finland. 196. Research Unit, Kuopio University Hospital, Kuopio, Finland. 197. Department of Medicine, Lausanne University Hospital, Switzerland. 198. Department of Endocrinology, University of Groningen, University Medical Center Groningen, The Netherlands. 199. Department of Cardiovascular Epidemiology and Population Genetics, National Center for Cardiovascular
Investigation, Madrid, Spain. 200. IMDEA-Alimentacion, Madrid, Spain. 201. Nutrition and Genomics Laboratory, Jean Mayer-USDA Human Nutrition Research Center on Aging at Tufts
University, Boston, MA, USA. 202. Department of Nutrition, Harvard School of Public Health, Boston, MA, USA. 203. Framingham Heart Study, Framingham, MA, USA. 204. Center for Public Health Genomics, University of Virginia, Charlottesville, VA 22908, USA. 205. Cardiovascular Research Center, Massachusetts General Hospital, Boston, Massachusetts 02114, USA. 206. Department of Medicine, Harvard Medical School, Boston, Massachusetts 02115, USA.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
9
Supplementary Information
Supplementary Tables ....................................................................................................................................................................................... 11
SUPPLEMENTARY TABLE 1: Phenotypic Summary of Samples with Metabochip Genotype Results ...................................................................................... 11
SUPPLEMENTARY TABLE 2. Biological Candidate Genes at Novel Loci based on Literature Search, Nonsynonymous Variants, Gene Expression Levels (eQTLs)
and Pathway Analysis ....................................................................................................................................................................................................... 14
SUPPLEMENTARY TABLE 3. Summary of Joint Meta-analysis Association Results for 95 Previously Discovered Lipid Loci .................................................... 18
SUPPLEMENTARY TABLE 4. Overlap of Novel Loci and Literature ....................................................................................................................................... 21
SUPPLEMENTARY TABLE 5. Pathways that Show Enrichment of Genes at Novel Loci by MAGENTA analysis ........................................................................ 24
SUPPLEMENTARY TABLE 5A. Pathways that Show Enrichment of Genes at Novel HDL Associated Loci by MAGENTA analysis ............................................. 24
SUPPLEMENTARY TABLE 5B. Pathways that Show Enrichment of Genes at Novel LDL Associated Loci by MAGENTA analysis .............................................. 26
SUPPLEMENTARY TABLE 5C. Pathways that Show Enrichment of Genes at Novel Total Cholesterol Associated Loci by MAGENTA analysis .......................... 28
SUPPLEMENTARY TABLE 5D. Pathways that Show Enrichment of Genes at Novel Triglyceride Associated Loci by MAGENTA analysis .................................. 30
SUPPLEMENTARY TABLE 6. Overlap Between eQTL Loci and New Lipid Associated Loci ...................................................................................................... 31
SUPPLEMENTARY TABLE 7. Nonsynonymous Variants in Linkage Disequilibrium with Index SNPs at Novel Loci .................................................................. 32
SUPPLEMENTARY TABLE 8. Overlap of SNPs at Known and Novel Lipid Loci with Chromatin States in 9 Different Cell Types ............................................... 33
SUPPLEMENTARY TABLE 9. Overlap with Chromatin States, Histone Marks and Transcription Factor ChIP-Seq in HepG2 Cells ............................................ 34
SUPPLEMENTARY TABLE 10. Overlap of Regulatory Features and Associated SNPs at Novel Lipid Loci ................................................................................ 35
SUPPLEMENTARY TABLE 11. Fine-Mapping Results in Different Ancestries ......................................................................................................................... 38
SUPPLEMENTARY TABLE 12. Novel and Known Lipid Loci Associated with BMI, CAD, DBP, SBP, Fasting Glucose, T2D, and WHR adj BMI ............................. 40
SUPPLEMENTARY TABLE 12A. Novel and Known Lipid Loci with BMI P-value < 0.05 from GIANT ......................................................................................... 40
SUPPLEMENTARY TABLE 12B. Novel and Known Lipid Loci with CAD P-value < 0.05 from CARDIOGRAM+C4D Meta-analysis* ............................................. 41
SUPPLEMENTARY TABLE12C. Novel and Known Lipid Loci with DBP P-value < 0.05 from ICBP* ........................................................................................... 42
SUPPLEMENTARY TABLE12D. Novel and Known Lipid Loci with SBP P-value < 0.05 from ICBP* ........................................................................................... 43
SUPPLEMENTARY TABLE12E. Novel and Known Lipid Loci with Fasting GlucoseP-value < 0.05 from MAGIC*....................................................................... 44
SUPPLEMENTARY TABLE12F. Novel and Known Lipid Loci with T2D P-value < 0.05 from DIAGRAM* ................................................................................... 45
SUPPLEMENTARY TABLE12G. Novel and Known Lipid Loci with WHR adj BMI P-value < 0.05 from GIANT* ......................................................................... 46
SUPPLEMENTARY TABLE 13.Overlap of Lipid Subfractions in Framingham with Novel and Known Lipid Associated Loci (P<1.4x10-5) ................................... 47
SUPPLEMENTARY TABLE 14. Overlap of Sphingolipids with Novel and Known Lipid Loci (P<1.5x10-5) .................................................................................. 48
Supplementary Figures ...................................................................................................................................................................................... 49
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
10
SUPPLEMENTARY FIGURE 1. Study Design ......................................................................................................................................................................... 49
SUPPLEMENTARY FIGURE 2. QQ Plots of Metabochip Meta-analysis P-value Distributions ................................................................................................. 50
SUPPLEMENTARY FIGURE 3. Manhattan Plots of Lipid-specific Association Results ............................................................................................................ 52
SUPPLEMENTARY FIGURE 4. Effect Size vs. Allele Frequency at Lipid Associated Loci .......................................................................................................... 54
SUPPLEMENTARY FIGURE 5. Direct Protein-Protein Interactions from Dapple Analysis ....................................................................................................... 55
SUPPLEMENTARY FIGURE 6. Lipid vs. CAD Effect Sizes ....................................................................................................................................................... 56
SUPPLEMENTARY FIGURE 7. Association with Lipid Subfractions ....................................................................................................................................... 57
Supplementary Note ......................................................................................................................................................................................... 68
Cohort Descriptions .......................................................................................................................................................................................................... 68
European Metabochip Cohorts ...................................................................................................................................................................................................... 68
East Asian Metabochip Cohorts ..................................................................................................................................................................................................... 74
South Asian Metabochip Cohorts .................................................................................................................................................................................................. 75
African Metabochip Cohorts .......................................................................................................................................................................................................... 76
European GWAS Cohorts ............................................................................................................................................................................................................... 77
Cohorts Excluded from Meta-analysis ........................................................................................................................................................................................... 77
Candidate Genes at Novel Loci .......................................................................................................................................................................................... 79
Detailed Acknowledgements ............................................................................................................................................................................................ 84
Full Name Author Contributions ....................................................................................................................................................................................... 98
Supplementary References ............................................................................................................................................................................................. 101
Nature Genetics: doi:10.1038/ng.2797
GLGC Metabochip Supplement 2013
11
Supplementary Tables
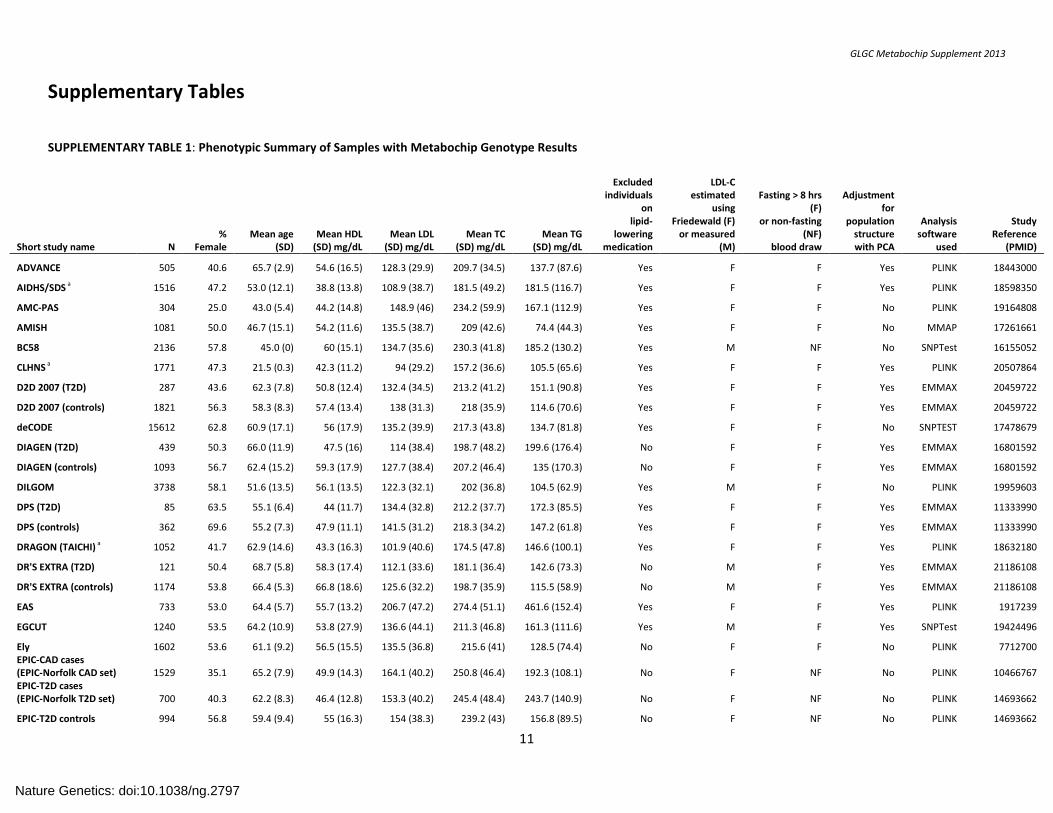
SUPPLEMENTARY TABLE 1: Phenotypic Summary of Samples with Metabochip Genotype Results
Short study name N %
Female Mean age
(SD) Mean HDL
(SD) mg/dL Mean LDL
(SD) mg/dL Mean TC
(SD) mg/dL Mean TG
(SD) mg/dL
Excluded individuals
on lipid-
lowering medication
LDL-C estimated
using Friedewald (F)
or measured (M)
Fasting > 8 hrs (F)
or non-fasting (NF)
blood draw
Adjustment for
population structure with PCA
Analysis software
used
Study Reference
(PMID)
ADVANCE 505 40.6 65.7 (2.9) 54.6 (16.5) 128.3 (29.9) 209.7 (34.5) 137.7 (87.6) Yes F F Yes PLINK 18443000
AIDHS/SDS a 1516 47.2 53.0 (12.1) 38.8 (13.8) 108.9 (38.7) 181.5 (49.2) 181.5 (116.7) Yes F F Yes PLINK 18598350
AMC-PAS 304 25.0 43.0 (5.4) 44.2 (14.8) 148.9 (46) 234.2 (59.9) 167.1 (112.9) Yes F F No PLINK 19164808
AMISH 1081 50.0 46.7 (15.1) 54.2 (11.6) 135.5 (38.7) 209 (42.6) 74.4 (44.3) Yes F F No MMAP 17261661
BC58 2136 57.8 45.0 (0) 60 (15.1) 134.7 (35.6) 230.3 (41.8) 185.2 (130.2) Yes M NF No SNPTest 16155052
CLHNS a 1771 47.3 21.5 (0.3) 42.3 (11.2) 94 (29.2) 157.2 (36.6) 105.5 (65.6) Yes F F Yes PLINK 20507864
D2D 2007 (T2D) 287 43.6 62.3 (7.8) 50.8 (12.4) 132.4 (34.5) 213.2 (41.2) 151.1 (90.8) Yes F F Yes EMMAX 20459722
D2D 2007 (controls) 1821 56.3 58.3 (8.3) 57.4 (13.4) 138 (31.3) 218 (35.9) 114.6 (70.6) Yes F F Yes EMMAX 20459722
deCODE 15612 62.8 60.9 (17.1) 56 (17.9) 135.2 (39.9) 217.3 (43.8) 134.7 (81.8) Yes F F No SNPTEST 17478679
DIAGEN (T2D) 439 50.3 66.0 (11.9) 47.5 (16) 114 (38.4) 198.7 (48.2) 199.6 (176.4) No F F Yes EMMAX 16801592
DIAGEN (controls) 1093 56.7 62.4 (15.2) 59.3 (17.9) 127.7 (38.4) 207.2 (46.4) 135 (170.3) No F F Yes EMMAX 16801592
DILGOM 3738 58.1 51.6 (13.5) 56.1 (13.5) 122.3 (32.1) 202 (36.8) 104.5 (62.9) Yes M F No PLINK 19959603
DPS (T2D) 85 63.5 55.1 (6.4) 44 (11.7) 134.4 (32.8) 212.2 (37.7) 172.3 (85.5) Yes F F Yes EMMAX 11333990
DPS (controls) 362 69.6 55.2 (7.3) 47.9 (11.1) 141.5 (31.2) 218.3 (34.2) 147.2 (61.8) Yes F F Yes EMMAX 11333990
DRAGON (TAICHI) a 1052 41.7 62.9 (14.6) 43.3 (16.3) 101.9 (40.6) 174.5 (47.8) 146.6 (100.1) Yes F F Yes PLINK 18632180
DR'S EXTRA (T2D) 121 50.4 68.7 (5.8) 58.3 (17.4) 112.1 (33.6) 181.1 (36.4) 142.6 (73.3) No M F Yes EMMAX 21186108
DR'S EXTRA (controls) 1174 53.8 66.4 (5.3) 66.8 (18.6) 125.6 (32.2) 198.7 (35.9) 115.5 (58.9) No M F Yes EMMAX 21186108
EAS 733 53.0 64.4 (5.7) 55.7 (13.2) 206.7 (47.2) 274.4 (51.1) 461.6 (152.4) Yes F F Yes PLINK 1917239
EGCUT 1240 53.5 64.2 (10.9) 53.8 (27.9) 136.6 (44.1) 211.3 (46.8) 161.3 (111.6) Yes M F Yes SNPTest 19424496
Ely 1602 53.6 61.1 (9.2) 56.5 (15.5) 135.5 (36.8) 215.6 (41) 128.5 (74.4) No F F No PLINK 7712700 EPIC-CAD cases (EPIC-Norfolk CAD set) 1529 35.1 65.2 (7.9) 49.9 (14.3) 164.1 (40.2) 250.8 (46.4) 192.3 (108.1) No F NF No PLINK 10466767 EPIC-T2D cases (EPIC-Norfolk T2D set) 700 40.3 62.2 (8.3) 46.4 (12.8) 153.3 (40.2) 245.4 (48.4) 243.7 (140.9) No F NF No PLINK 14693662
EPIC-T2D controls 994 56.8 59.4 (9.4) 55 (16.3) 154 (38.3) 239.2 (43) 156.8 (89.5) No F NF No PLINK 14693662
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
12
(EPIC-Norfolk T2D set)
FBPP a 1614 64.0 43.9 (12.5) 53.6 (15.8) 118.4 (37.8) 193.2 (43.5) NA Yes F F No Merlin 11799070
Fenland 3186 53.3 46.9 (7.1) 58.8 (15.1) 129.3 (34.8) 207 (39.5) 103.7 (71.8) No F F No PLINK
FINCAVAS 1201 44.4 58.5 (12.6) 54.6 (17.8) 113 (33.3) 191.2 (38.7) 119.6 (65.6) Yes F F Yes SNPTest 16515696
FRISCII 2963 30.6 66.2 (9.7) 46.7 (15.2) 145.7 (38.2) 228.1 (43.1) 185.7 (119.5) Yes F F No GenABEL 10892758
FUSION2 (T2D) 843 43.3 59.5 (8.5) 51.1 (14.8) 123.1 (36.2) 217.6 (47.3) 163.3 (92.8) Yes F F Yes EMMAX 17463248
FUSION2 (controls) 1880 44.7 56.2 (8.2) 58.1 (15.8) 137.9 (34.9) 225.7 (42.5) 114.8 (68.5) Yes F F Yes EMMAX 17463248
GLACIER 5764b 61.4 b 54.2 (8.0) b 56.3 (13.4) 173 (45.3) 232.5 (47.4) 142.6 (47.4) Yes F F Yes PLINK 20870969
Go-DARTs 6759 46.1 57.8 (10.3) 55 (17) 120.7 (35.2) 216.7 (46.8) 177.2 (126.7) No F F No PLINK 17429603
GXE a 397 76.8 39.8 (8.2) 50.9 (12) 146.1 (41.6) 204.3 (43.7) 84.7 (50.1) Yes F F Yes PLINK 21347282
HALST (TAICHI) a 2375 49.4 68.9 (8.3) 52.6 (13.5) 120.0 (31.8) 197.7 (36.8) 123.1 (76.2) Yes F F Yes PLINK
HUNT (T2D) 588 49.5 69.3 (11.3) 45.8 (14.6) 151.9 (42) 241.7 (51.8) 238.7 (163.4) No F NF Yes EMMAX 22879362
HUNT (controls) 784 49.0 66.3 (14.4) 50.9 (14.3) 163.4 (44.1) 249.7 (48.3) 183.8 (101.1) No F NF Yes EMMAX 22879362
IMPROVE 1769 50.0 64.4 (5.3) 48.8 (14.7) 148.6 (36.4) 224.1 (42.2) 139.1 (96.6) Yes F F Yes PLINK 19952003
KORA F3 2816 52.1 56.3 (12.8) 59.2 (18.2) 129.6 (32.1) 220.2 (39.5) 163.9 (124.9) Yes M NF No PLINK 16032514
KORA F4 2678 53.1 54.5 (13.1) 56.5 (14.7) 138.5 (34.8) 218.3 (39.5) 122.3 (86.8) Yes M F No PLINK 16032514
MRC/UVRIGPCad 1687 56.7 35.0 (19.1) 39.9 (14.3) 78.6 (29) 138.9 (36.8) 104.5 (55.8) No M NF No EMMAX
LURIC (cases) 983 26.4 64.9 (9.7) 38.5 (10.7) 124.5 (33.4) 201.2 (37.3) 170 (99.9) Yes M F No PLINK 11258203
LURIC (controls) 523 46.1 57.9 (12.5) 42.8 (11.5) 121.2 (29.4) 198.8 (35) 154.5 (100.9) Yes M F No PLINK 11258203
MDC 2125 51.3 57.4 (6.0) 53.9 (13.9) 160 (36.6) 235.5 (40.2) 109.2 (50.5) No F F Yes PLINK 8429286
METSIM (T2D) 634 0.0 59.7 (6.8) 52.1 (16.1) 134.6 (36.2) 212.8 (44.4) 167 (111.8) Yes M F Yes EMMAX 19223598
METSIM (controls) 829 0.0 53.7 (5) 57.3 (14.7) 138.4 (29.9) 215.3 (34) 120.1 (76.2) Yes M F Yes EMMAX 19223598
NFBC86 4164 52.0 16.0 (0.4) 54.6 (11.2) 87.1 (22.4) 164.9 (30.2) 73.5 (36.3) No F F No PLINK
NSHD 941 52.8 53.0 63.5 (19.7) 136.2 (38.3) 237.2 (43) 206.4 (154.2) No F NF No PLINK 16204333
PIVUS 854 51.0 70.0 (0.2) 58.1 (15.5) 135.5 (31) 216.7 (38.7) 106.3 (44.3) Yes F F Yes PLINK 18489581
PROMIS a 3385 18.0 52.5 (9.9) 35.5 (9.9) 122.3 (43.7) 192.7 (50.7) 210.2 (128.8) No M NF Yes PLINK 19404752
SAPPHIRe (TAICHI) a 251 49.4 54.6 (10.5) 44.2 (12.6) 127.1 (38.2) 200.4 (43.4) 143.0 (80.0) Yes F F Yes PLINK 22839215
SardiNIA 5378 56.8 43.2 (17.4) 64.3 (14.9) 127.1 (35.5) 208.4 (42.6) 86.9 (68.3) Yes F F No Merlin 16934002
SCARFSHEEP 2973 0.3 58.3 (7.2) 45.7 (13.9) 156.3 (37.5) 230.7 (42.2) 148 (101) Yes F F No PLINK
SEY a 426 54.7 48.7 (14.1) 48.3 (13.1) 141.4 (44.1) 213.3 (49.1) 117.8 (80.1) Yes F F No Merlin 15610228
SPT a 838c 61.8c 46.7 (0.5)c 48.8 (13) 133.7 (41.3) 192.8 (42.1) 89.1 (53.8) Yes F F Yes PLINK 9103091
STR 2543 57.0 75.0 (10.2) 54.2 (15.5) 150.9 (42.6) 243.8 (50.3) 150.6 (79.7) Yes F F No Merlin 19606474
Nature Genetics: doi:10.1038/ng.2797
GLGC Metabochip Supplement 2013
13
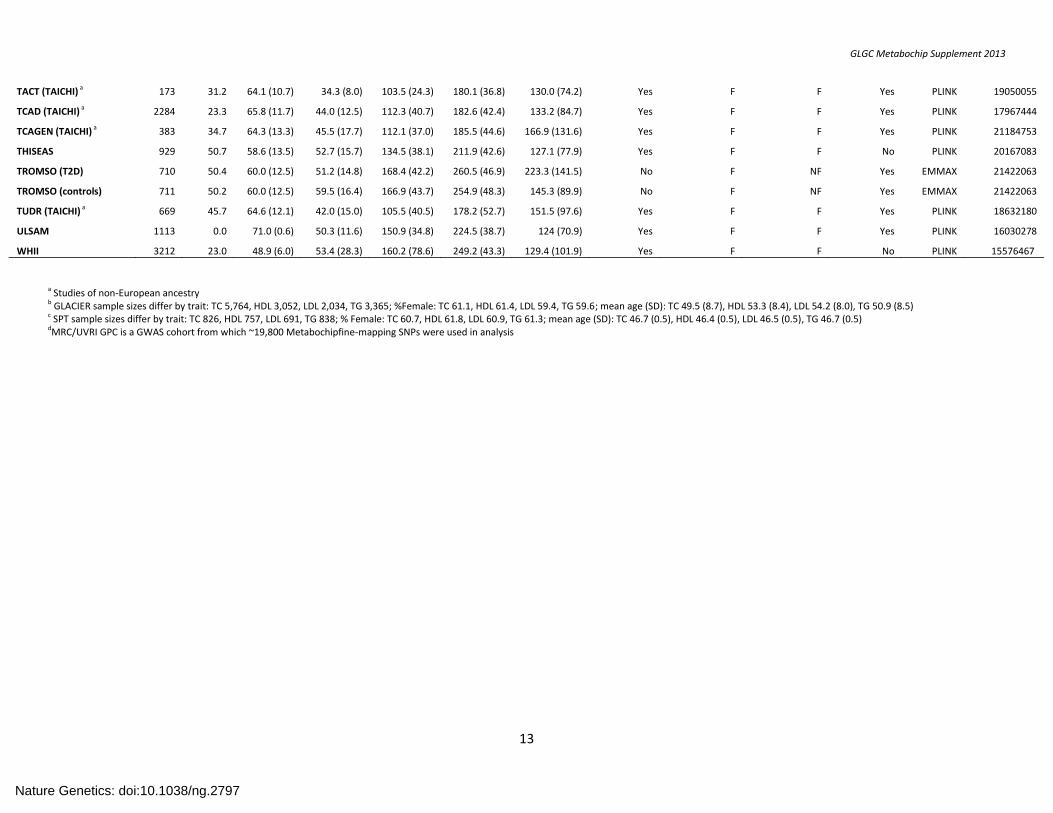
TACT (TAICHI) a 173 31.2 64.1 (10.7) 34.3 (8.0) 103.5 (24.3) 180.1 (36.8) 130.0 (74.2) Yes F F Yes PLINK 19050055
TCAD (TAICHI) a 2284 23.3 65.8 (11.7) 44.0 (12.5) 112.3 (40.7) 182.6 (42.4) 133.2 (84.7) Yes F F Yes PLINK 17967444
TCAGEN (TAICHI) a 383 34.7 64.3 (13.3) 45.5 (17.7) 112.1 (37.0) 185.5 (44.6) 166.9 (131.6) Yes F F Yes PLINK 21184753
THISEAS 929 50.7 58.6 (13.5) 52.7 (15.7) 134.5 (38.1) 211.9 (42.6) 127.1 (77.9) Yes F F No PLINK 20167083
TROMSO (T2D) 710 50.4 60.0 (12.5) 51.2 (14.8) 168.4 (42.2) 260.5 (46.9) 223.3 (141.5) No F NF Yes EMMAX 21422063
TROMSO (controls) 711 50.2 60.0 (12.5) 59.5 (16.4) 166.9 (43.7) 254.9 (48.3) 145.3 (89.9) No F NF Yes EMMAX 21422063
TUDR (TAICHI) a 669 45.7 64.6 (12.1) 42.0 (15.0) 105.5 (40.5) 178.2 (52.7) 151.5 (97.6) Yes F F Yes PLINK 18632180
ULSAM 1113 0.0 71.0 (0.6) 50.3 (11.6) 150.9 (34.8) 224.5 (38.7) 124 (70.9) Yes F F Yes PLINK 16030278
WHII 3212 23.0 48.9 (6.0) 53.4 (28.3) 160.2 (78.6) 249.2 (43.3) 129.4 (101.9) Yes F F No PLINK 15576467
a Studies of non-European ancestry
b GLACIER sample sizes differ by trait: TC 5,764, HDL 3,052, LDL 2,034, TG 3,365; %Female: TC 61.1, HDL 61.4, LDL 59.4, TG 59.6; mean age (SD): TC 49.5 (8.7), HDL 53.3 (8.4), LDL 54.2 (8.0), TG 50.9 (8.5) c SPT sample sizes differ by trait: TC 826, HDL 757, LDL 691, TG 838; % Female: TC 60.7, HDL 61.8, LDL 60.9, TG 61.3; mean age (SD): TC 46.7 (0.5), HDL 46.4 (0.5), LDL 46.5 (0.5), TG 46.7 (0.5) dMRC/UVRI GPC is a GWAS cohort from which ~19,800 Metabochipfine-mapping SNPs were used in analysis
Nature Genetics: doi:10.1038/ng.2797
GLGC Metabochip Supplement 2013
14
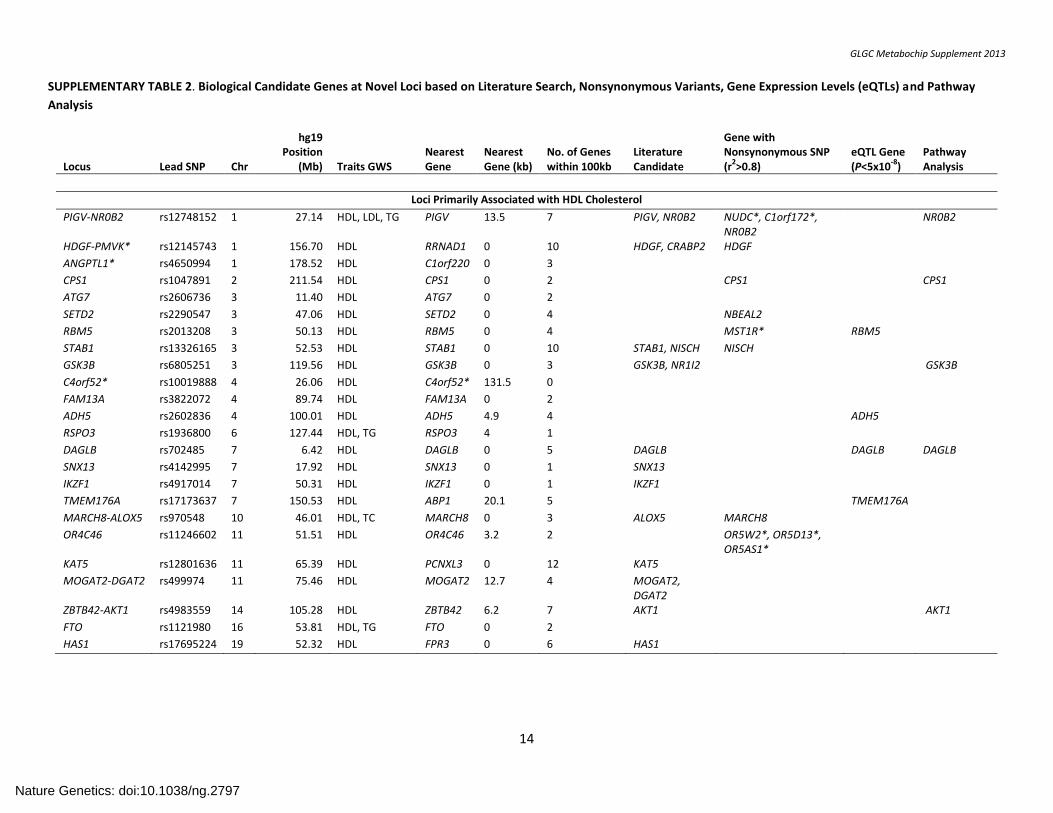
SUPPLEMENTARY TABLE 2. Biological Candidate Genes at Novel Loci based on Literature Search, Nonsynonymous Variants, Gene Expression Levels (eQTLs) and Pathway
Analysis
Locus Lead SNP Chr
hg19 Position
(Mb) Traits GWS
Nearest Gene
Nearest Gene (kb)
No. of Genes within 100kb
Literature Candidate
Gene with Nonsynonymous SNP (r
2>0.8)
eQTL Gene (P<5x10
-8)
Pathway Analysis
Loci Primarily Associated with HDL Cholesterol
PIGV-NR0B2 rs12748152 1 27.14 HDL, LDL, TG PIGV 13.5 7 PIGV, NR0B2 NUDC*, C1orf172*, NR0B2
NR0B2
HDGF-PMVK* rs12145743 1 156.70 HDL RRNAD1 0 10 HDGF, CRABP2 HDGF
ANGPTL1* rs4650994 1 178.52 HDL C1orf220 0 3
CPS1 rs1047891 2 211.54 HDL CPS1 0 2 CPS1 CPS1
ATG7 rs2606736 3 11.40 HDL ATG7 0 2
SETD2 rs2290547 3 47.06 HDL SETD2 0 4 NBEAL2
RBM5 rs2013208 3 50.13 HDL RBM5 0 4 MST1R* RBM5
STAB1 rs13326165 3 52.53 HDL STAB1 0 10 STAB1, NISCH NISCH
GSK3B rs6805251 3 119.56 HDL GSK3B 0 3 GSK3B, NR1I2 GSK3B
C4orf52* rs10019888 4 26.06 HDL C4orf52* 131.5 0
FAM13A rs3822072 4 89.74 HDL FAM13A 0 2
ADH5 rs2602836 4 100.01 HDL ADH5 4.9 4 ADH5
RSPO3 rs1936800 6 127.44 HDL, TG RSPO3 4 1
DAGLB rs702485 7 6.42 HDL DAGLB 0 5 DAGLB DAGLB DAGLB
SNX13 rs4142995 7 17.92 HDL SNX13 0 1 SNX13
IKZF1 rs4917014 7 50.31 HDL IKZF1 0 1 IKZF1
TMEM176A rs17173637 7 150.53 HDL ABP1 20.1 5 TMEM176A
MARCH8-ALOX5 rs970548 10 46.01 HDL, TC MARCH8 0 3 ALOX5 MARCH8
OR4C46 rs11246602 11 51.51 HDL OR4C46 3.2 2 OR5W2*, OR5D13*, OR5AS1*
KAT5 rs12801636 11 65.39 HDL PCNXL3 0 12 KAT5
MOGAT2-DGAT2 rs499974 11 75.46 HDL MOGAT2 12.7 4 MOGAT2, DGAT2
ZBTB42-AKT1 rs4983559 14 105.28 HDL ZBTB42 6.2 7 AKT1 AKT1
FTO rs1121980 16 53.81 HDL, TG FTO 0 2
HAS1 rs17695224 19 52.32 HDL FPR3 0 6 HAS1
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
15
Supplementary Table 2 (continued).
Locus Lead SNP Chr
hg19 Position
(Mb) Traits GWS
Nearest Gene
Nearest Gene (kb)
No. of Genes within 100kb
Literature Candidate
Gene with Nonsynonymous SNP (r
2>0.8)
eQTL Gene (P<5x10
-8)
Pathway Analysis
Loci Primarily Associated with LDL Cholesterol
ANXA9-CERS2 rs267733 1 150.96 LDL ANXA9 0 10 CERS2 ANXA9 ANXA9
EHBP1 rs2710642 2 63.15 LDL EHBP1 0 1 EHBP1
INSIG2 rs10490626 2 118.84 LDL, TC INSIG2 10.2 2 INSIG2 CCDC93 INSIG2
LOC84931 rs2030746 2 121.31 LDL, TC LOC84931 85.6 1
FN1 rs1250229 2 216.30 LDL FN1 3.6 2 FN1 FN1
CMTM6 rs7640978 3 32.53 LDL, TC CMTM6 0 3 DYNC1LI1
ACAD11 rs17404153 3 132.16 LDL, HDL DNAJC13 0 2 ACAD11*
CSNK1G3 rs4530754 5 122.86 LDL, TC CSNK1G3 0 2
MIR148A rs4722551 7 25.99 LDL, TC, TG MIR148A 2.2 1
SOX17 rs10102164 8 55.42 LDL, TC SOX17 48.2 1
BRCA2 rs4942486 13 32.95 LDL BRCA2 0 5 BRCA2
APOH-PRXCA rs1801689 17 64.21 LDL APOH 0 3 APOH, PRKCA APOH APOH
SPTLC3 rs364585 20 12.96 LDL SPTLC3 26.9 1 SPTLC3 SPTLC3
SNX5 rs2328223 20 17.85 LDL SNX5 76.3 2 SNX5
MTMR3 rs5763662 22 30.38 LDL MTMR3 0 2
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
16
Supplementary Table 2 (continued).
Locus Lead SNP Chr
hg19 Position
(Mb) Traits GWS
Nearest Gene
Nearest Gene (kb)
No. of Genes within 100kb
Literature Candidate
Gene with Nonsynonymous SNP (r
2>0.8)
eQTL Gene (P<5x10
-8)
Pathway Analysis
Loci Primarily Associated with Total Cholesterol
ASAP3 rs1077514 1 23.77 TC ASAP3 0 6
ABCB11 rs2287623 2 169.83 TC ABCB11 0 4 ABCB11 ABCB11 ABCB11
FAM117B rs11694172 2 203.53 TC FAM117B 0 2
UGT1A1 rs11563251 2 234.68 TC, LDL UGT1A1 0 12 UGT1A1/3/4/5 UGT1A6/7/8/9
UGT1A1
PXK rs13315871 3 58.38 TC PXK 0 4 PXK PXK
KCNK17 rs2758886 6 39.25 TC KCNK17 15.9 4
HBS1L rs9376090 6 135.41 TC HBS1L 35.2 2
GPR146 rs1997243 7 1.08 TC C7orf50 0 7 GPR146 GPR146
VLDLR rs3780181 9 2.64 TC, LDL VLDLR 0 3 VLDLR VLDLR
VIM-CUBN rs10904908 10 17.26 TC VIM 10.0 3 VIM, CUBN CUBN
PHLDB1 rs11603023 11 118.49 TC PHLDB1 0 7
PHC1-A2ML1 rs4883201 12 9.08 TC PHC1 0 4 A2ML1
DLG4 rs314253 17 7.09 TC, LDL DLG4 1.6 13 ACADVL, CTDNEP1, SLC2A4
DLG4
TOM1 rs138777 22 35.71 TC TOM1 0 4 HMOX1 HMGXB4
PPARA rs4253772 22 46.63 TC, LDL PPARA 0 6 PPARA PPARA
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
17
Supplementary Table 2 (continued).
Locus Lead SNP Chr
hg19 Position
(Mb) Traits GWS
Nearest Gene
Nearest Gene (kb)
No. of Genes within 100kb
Literature Candidate
Gene with Nonsynonymous SNP (r
2>0.8)
eQTL Gene (P<5x10
-8)
Pathway Analysis
Loci Primarily Associated with Triglycerides
LRPAP1 rs6831256 4 3.47 TG, LDL, TC DOK7 0 4 LRPAP1 LRPAP1
VEGFA rs998584 6 43.76 TG, HDL VEGFA 3.7 1 VEGFA VEGFA
MET rs38855 7 116.36 TG MET 0 1
AKR1C4 rs1832007 10 5.25 TG AKR1C4 0 2 AKR1C4 AKR1C4 AKR1C4
PDXDC1 rs3198697 16 15.13 TG PDXDC1 0 4
MPP3 rs8077889 17 41.88 TG MPP3 0 6 MPP3
INSR rs7248104 19 7.22 TG INSR 0 1 INSR PEPD rs731839 19 33.90 TG, HDL PEPD 0 2 CEBPG
Supplementary Table 2 summarizes results of our search for candidates at each locus. The locus label includes a gene used to refer to the locus throughout the text. Except for loci labeled * (PMVK, ANGPTL1 and C4orf52)the locus label always refers to a gene within 100kb of the SNP with strongest association; in these three cases, the gene selected as the locus label was judged to be an especially worthy candidate >100kb or no genes within 100kb of the lead SNP were available. The columns labeled literature candidate, non-synonymous SNP, eQTL and pathway analysis candidate indicate genes flagged in our various searches for candidate genes, further detailed in the text and in supplementary tables. Genes with a non-synonymous SNP in disequilibrium with the lead SNP for the locus but more than 100kb away are also labeled *.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
18
SUPPLEMENTARY TABLE 3. Summary of Joint Meta-analysis Association Results for 95 Previously Discovered Lipid Loci
Nearest gene
MarkerName
Chr
hg19
Position (Mb)
Primary trait, Secondary trait(s)
MAF
Alleles minor/ major
Effect
Joint N (in 1000s)
Joint P-value
Loci Primarily Associated with HDL Cholesterol
PABPC4 rs4660293 1 40.03 HDL .24 G/A -.035 187 3x10-18
ZNF648 rs1689800 1 182.17 HDL .35 G/A -.034 187 5x10-20
GALNT2 rs4846914 1 230.30 HDL,TG .41 G/A -.048/.040 187/178 4x10-41
/7x10-31
COBLL1 rs12328675 2 165.54 HDL .13 C/T .045 187 2x10-15
IRS1 rs2972146 2 227.10 HDL,TG .37 G/T .032/-.028 184/175 2x10-17
/3x10-15
SLC39A8 rs13107325 4 103.19 HDL .08 T/C -.071 179 1x10-15
ARL15 rs6450176 5 53.30 HDL .26 A/G -.025 187 7x10-10
CITED2 rs605066 6 139.83 HDL .42 C/T -.028 94 3x10-8
KLF14 rs4731702 7 130.43 HDL .49 T/C .029 187 5x10-17
PPP1R3B rs9987289 8 9.18 HDL,TC,LDL .10 A/G -.082/-.084/-.071 169/174/160 2x10-41
/2x10-36
/9x10-24
TRPS1 rs2293889 8 116.60 HDL .41 T/G -.031 180102 4x10-17
TTC39B rs581080 9 15.31 HDL,TC .21 G/C -.042/-.038 187/187 1x10-19
/1x10-13
ABCA1 rs1883025 9 107.66 HDL,TC .25 T/C -.07/-.067 186/187 2x10-65
/6x10-53
AMPD3 rs2923084 11 10.39 HDL .18 G/A -.026 187 5x10-8
LRP4 rs3136441 11 46.74 HDL .18 C/T .054 187 7x10-29
PDE3A rs7134375 12 20.47 HDL .43 A/C .021 187 1x10-8
MVK rs7134594 12 110.00 HDL .48 C/T -.035 94 2x10-13
SBNO1 rs4759375 12 123.80 HDL .08 T/C .056 94 3x10-8
ZNF664 rs4765127 12 124.46 HDL,TG .35 T/G .032/-.029 94/91 8x10-10
/2x10-8
SCARB1 rs838880 12 125.26 HDL .34 C/T .048 173 6x10-32
LIPC rs1532085 15 58.68 HDL,TC,TG .40 A/G .107/.054/.031 185/186/176 1x10-188
/7x10-47
/2x10-18
LACTB rs2652834 15 63.40 HDL .21 A/G -.028 186 4x10-11
CETP rs3764261 16 56.99 HDL,LDL,TC,TG .32 A/C .241/-.053/.050/-.040 178/165/177/169 1x10-769/2x10-34/4x10-31/2x10-25
LCAT rs16942887 16 67.93 HDL .14 A/G .083 186 8x10-54
CMIP rs2925979 16 81.53 HDL .31 T/C -.035 186 1x10-19
STARD3 rs11869286 17 37.81 HDL .35 G/C -.032 178 3x10-17
ABCA8 rs4148008 17 66.88 HDL .33 G/C -.028 166 1x10-12
PGS1 rs4129767 17 76.40 HDL .48 G/A -.024 185 2x10-11
LIPG rs7241918 18 47.16 HDL,TC .19 G/T -.09/-.058 93/93 1x10-44
/4x10-18
MC4R rs12967135 18 57.85 HDL .25 A/G -.026 154 4x10-8
ANGPTL4 rs7255436 19 8.43 HDL .47 C/A -.032 93 2x10-8
ANGPTL8 rs737337 19 11.35 HDL .11 C/T -.056 185 5x10-17
LILRA3 rs386000 19 54.79 HDL .26 C/G .048 165 3x10-23
HNF4A rs1800961 20 43.04 HDL,TC .05 T/C -.127/-.106 158/156 2x10-34
/1x10-24
PLTP rs6065906 20 44.55 HDL,TG .19 C/T -.059/.053 186/176 5x10-40
/2x10-34
UBE2L3 rs181362 22 21.93 HDL .23 T/C -.038 178 4x10-18
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
19
Supplementary Table 3 (continued).
Loci Primarily Associated with LDL Cholesterol
PCSK9 rs2479409 1 55.50 LDL,TC .32 G/A .064/.054 173/187 3x10-50
/2x10-39
SORT1 rs629301 1 109.82 LDL,TC .24 G/T -.167/-.134 143/156 5x10-241
/2x10-170
APOB rs1367117 2 21.26 LDL,TC .32 A/G .119/.100 173/187 1x10-182
/3x10-139
ABCG5/8 rs4299376 2 44.07 LDL,TC .31 G/T .081/.079 145/158 4x10-72
/3x10-73
MYLIP rs3757354 6 16.13 LDL,TC .24 T/C -.038/-.035 173/187 2x10-17
/2x10-15
HFE rs1800562 6 26.09 LDL,TC .07 A/G -.062/-.056 171/185 8x10-14
/2x10-12
LPA rs1564348 6 160.58 LDL,TC .18 C/T .048/.049 173/187 3x10-21
/3x10-23
PLEC1 rs11136341 8 145.04 LDL,TC .40 G/A .045/.038 83/87 7x10-12
/6x10-9
ABO rs9411489 9 136.155 LDL,TC .21 T/C .077/.069 119/130 2x10-41
/3x10-35
ST3GAL4 rs11220462 11 126.24 LDL,TC .14 A/G .059/.047 145/157 7x10-21
/6x10-15
NYNRIN rs8017377 14 24.88 LDL .46 A/G .030 173 3x10-15
OSBPL7 rs7206971 17 45.43 LDL,TC .49 A/G .029/.030 81/85 3x10-7
/1x10-7
LDLR rs6511720 19 11.20 LDL,TC .12 T/G -.221/-.185 171/185 4x10-262
/5x10-202
APOE rs4420638 19 45.42 LDL,TC,HDL .19 G/A .225/.197/-.067 93/104/100 2x10-178
/1x10-149
/2x10-21
TOP1 rs6029526 20 39.67 LDL,TC .47 A/T .044/.040 88/93 5x10-18
/1x10-16
Loci Primarily Associated with Total Cholesterol
LDLRAP1 rs12027135 1 25.78 TC,LDL .46 A/T -.027/-.030 178/165 5x10-12
/2x10-14
EVI5 rs7515577 1 93.01 TC .23 C/A -.037 95 2x10-8
MOSC1 rs2642442 1 220.97 TC,LDL .33 C/T -.035/-.036 111/102 3x10-11
/5x10-11
IRF2BP2 rs514230 1 234.86 TC,LDL .48 A/T -.039/-.036 95/90 5x10-14
/9x10-12
RAB3GAP1 rs7570971 2 135.84 TC .35 A/C .030 185 1x10-13
RAF1 rs2290159 3 12.63 TC .23 C/G -.037 94 2x10-9
HMGCR rs12916 5 74.66 TC,LDL .40 C/T .068/.073 183/168 5x10-74
/8x10-78
TIMD4 rs6882076 5 156.39 TC,TG,LDL .36 T/C -.051/-.029/-.046 187/178/173 5x10-41
/2x10-15
/3x10-31
HLA rs3177928 6 32.41 TC,LDL .17 A/G .048/.045 180/166 1x10-21
/3x10-17
C6orf106 rs2814982 6 34.55 TC .12 T/C -.044 187 4x10-15
FRK rs9488822 6 116.31 TC,LDL .36 T/A .034/.031 95/90 1x10-9
/2x10-7
DNAH11 rs12670798 7 21.61 TC,LDL .25 C/T .036/.034 187/173 1x10-16
/5x10-14
NPC1L1 rs2072183 7 44.58 TC,LDL .29 C/G .036/.039 184/170 4x10-15
/7x10-16
CYP7A1 rs2081687 8 59.39 TC,LDL .36 T/C .038/.031 95/90 9x10-12
/1x10-7
GPAM rs2255141 10 113.93 TC,LDL .30 A/G .031/.030 187/173 7x10-16
/1x10-13
SPTY2D1 rs10128711 11 18.63 TC .30 T/C -.031 157 1x10-11
UBASH3B rs7941030 11 122.52 TC,HDL .39 C/T .028/.027 187/187 2x10-14
/1x10-14
BRAP rs11065987 12 112.07 TC,LDL .41 G/A -.031/-.027 187/173 2x10-16
/1x10-11
HNF1A rs1169288 12 121.42 TC,LDL .34 C/A .032/.038 176/163 4x10-17
/6x10-21
HPR rs2000999 16 72.11 TC,LDL .20 A/G .062/.065 186/172 7x10-41
/4x10-41
CILP2 rs10401969 19 19.41 TC,TG,LDL .09 C/T -.137/-.121/-.118 186/176/171 4x10-77
/1x10-69
/3x10-54
FLJ36070 rs492602 19 49.21 TC .47 G/A .031 184 1x10-16
ERGIC3 rs2277862 20 34.15 TC .15 T/C -.035 186 5x10-11
MAFB rs2902940 20 39.09 TC,LDL .30 G/A -.024/-.027 186/172 9x10-10
/2x10-11
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
20
Supplementary Table 3 (continued).
Loci Primarily Associated with Triglycerides
ANGPTL3 rs2131925 1 63.03 TG,LDL,TC .34 G/T -.066/-.049/-.075 178/173/187 3x10-74
/3x10-32
/4x10-80
GCKR rs1260326 2 27.73 TG,TC .39 T/C .115/.051 178/187 2x10-239
/3x10-42
MSL2L1 rs645040 3 135.93 TG .23 G/T -.029 178 2x10-12
KLHL8 rs442177 4 88.03 TG .42 G/T -.031 178 1x10-18
MAP3K1 rs9686661 5 55.86 TG .20 T/C .038 177 3x10-16
TYW1B rs13238203 7 72.13 TG .04 T/C -.059 102 3x10-6
MLXIPL rs17145738 7 72.98 TG,HDL .13 T/C -.115/.041 176/185 9x10-99
/5x10-13
PINX1 rs11776767 8 10.68 TG .37 C/G .022 177 3x10-11
NAT2 rs1495741 8 18.27 TG,TC .26 G/A .040/.032 88/92 3x10-12
/3x10-8
LPL rs12678919 8 19.84 TG,HDL .13 G/A -.170/.155 178/187 2x10-199
/1x10-149
TRIB1 rs2954029 8 126.49 TG,TC,LDL,HDL .47 T/A -.076/-.062/-.056/.040 178/187/173/187 1x10-107/2x10-65/2x10-50/3x10-29
JMJD1C rs10761731 10 65.03 TG .44 T/A -.031 91 8x10-12
CYP26A1 rs2068888 10 94.84 TG .45 A/G -.024 178 2x10-11
FADS1-2-3 rs174546 11 61.57 TG,LDL,TC,HDL .36 T/C .045/-.051/-.048/-.039 178/173/187/187 7x10-38/2x10-39/3x10-37/8x10-28
APOA1 rs964184 11 116.65 TG,TC,HDL,LDL .84 C/G -.234/-.121/.106/-.086 91/95/94/90 7x10-224/3x10-55/6x10-48/2x10-26
LRP1 rs11613352 12 57.79 TG,HDL .26 T/C -.028/.028 178/187 9x10-14
/2x10-13
CAPN3 rs2412710 15 42.68 TG .04 A/G .099 154 2x10-11
FRMD5 rs2929282 15 44.25 TG .07 T/A .072 84 2x10-9
CTF1 rs11649653 16 30.92 TG .40 G/C -.027 90 2x10-7
PLA2G6 rs5756931 22 38.55 TG .40 C/T -.020 174 3x10-8
Nature Genetics: doi:10.1038/ng.2797
GLGC Metabochip Supplement 2013
21
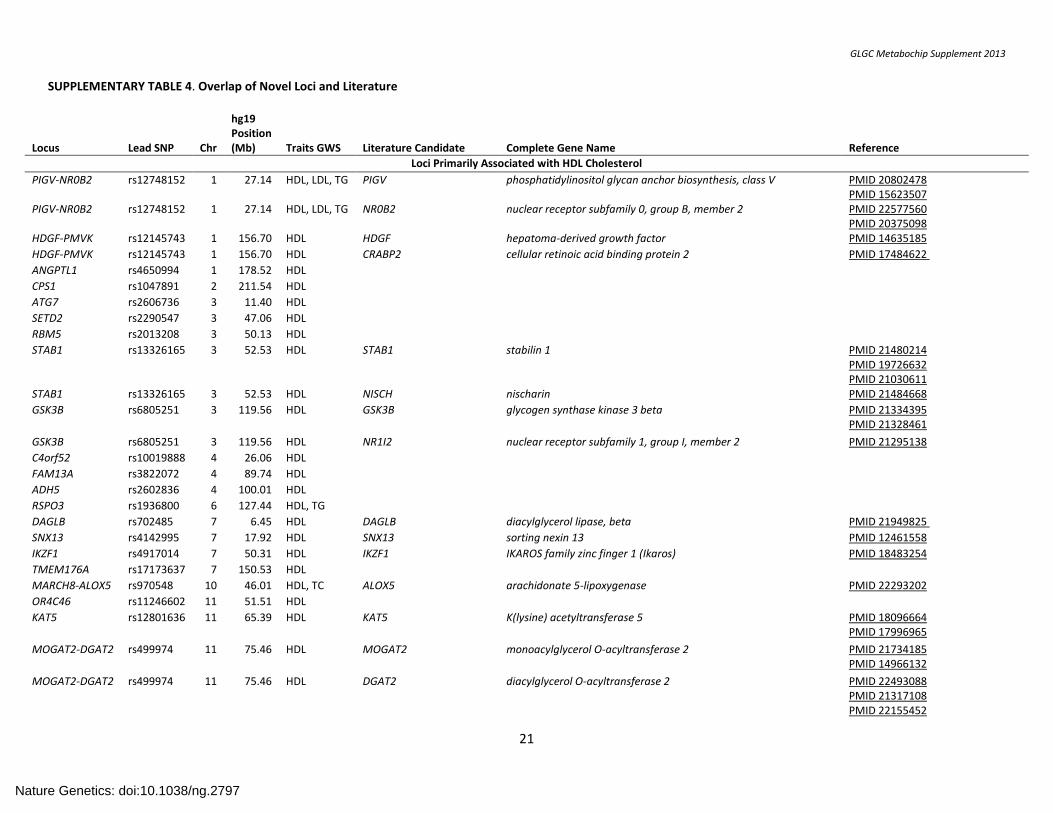
SUPPLEMENTARY TABLE 4. Overlap of Novel Loci and Literature
Locus
Lead SNP
Chr
hg19 Position (Mb)
Traits GWS
Literature Candidate
Complete Gene Name
Reference
Loci Primarily Associated with HDL Cholesterol
PIGV-NR0B2 rs12748152 1 27.14 HDL, LDL, TG PIGV phosphatidylinositol glycan anchor biosynthesis, class V PMID 20802478 PMID 15623507
PIGV-NR0B2 rs12748152 1 27.14 HDL, LDL, TG NR0B2 nuclear receptor subfamily 0, group B, member 2 PMID 22577560 PMID 20375098
HDGF-PMVK rs12145743 1 156.70 HDL HDGF hepatoma-derived growth factor PMID 14635185
HDGF-PMVK rs12145743 1 156.70 HDL CRABP2 cellular retinoic acid binding protein 2 PMID 17484622
ANGPTL1 rs4650994 1 178.52 HDL
CPS1 rs1047891 2 211.54 HDL
ATG7 rs2606736 3 11.40 HDL
SETD2 rs2290547 3 47.06 HDL
RBM5 rs2013208 3 50.13 HDL
STAB1 rs13326165 3 52.53 HDL STAB1 stabilin 1 PMID 21480214 PMID 19726632 PMID 21030611
STAB1 rs13326165 3 52.53 HDL NISCH nischarin PMID 21484668
GSK3B rs6805251 3 119.56 HDL GSK3B glycogen synthase kinase 3 beta PMID 21334395 PMID 21328461
GSK3B rs6805251 3 119.56 HDL NR1I2 nuclear receptor subfamily 1, group I, member 2 PMID 21295138
C4orf52 rs10019888 4 26.06 HDL
FAM13A rs3822072 4 89.74 HDL
ADH5 rs2602836 4 100.01 HDL
RSPO3 rs1936800 6 127.44 HDL, TG
DAGLB rs702485 7 6.45 HDL DAGLB diacylglycerol lipase, beta PMID 21949825
SNX13 rs4142995 7 17.92 HDL SNX13 sorting nexin 13 PMID 12461558
IKZF1 rs4917014 7 50.31 HDL IKZF1 IKAROS family zinc finger 1 (Ikaros) PMID 18483254
TMEM176A rs17173637 7 150.53 HDL
MARCH8-ALOX5 rs970548 10 46.01 HDL, TC ALOX5 arachidonate 5-lipoxygenase PMID 22293202
OR4C46 rs11246602 11 51.51 HDL
KAT5 rs12801636 11 65.39 HDL KAT5 K(lysine) acetyltransferase 5 PMID 18096664 PMID 17996965
MOGAT2-DGAT2 rs499974 11 75.46 HDL MOGAT2 monoacylglycerol O-acyltransferase 2 PMID 21734185 PMID 14966132
MOGAT2-DGAT2 rs499974 11 75.46 HDL DGAT2 diacylglycerol O-acyltransferase 2 PMID 22493088 PMID 21317108 PMID 22155452
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
22
Supplementary Table 4 (continued).
ZBTB42-AKT1 rs4983559 14 105.28 HDL AKT1 v-akt murine thymoma viral oncogene homolog 1 PMID 18054314
PMID 20054340 FTO rs1121980 16 53.81 HDL, TG
HAS1 rs17695224 19 52.32 HDL HAS1 hyaluronan synthase 1 PMID 9933623 Loci Primarily Associated with LDL Cholesterol
ANXA9-CERS2 rs267733 1 150.96 LDL CERS2 ceramide synthase 2 PMID 20940143 PMID 20110363 PMID 19801672
EHBP1 rs2710642 2 63.15 LDL EHBP1 EH domain binding protein 1 PMID 21332221
INSIG2 rs10490626 2 118.84 LDL, TC INSIG2 insulin induced gene 2 PMID 22143767 PMID 20817058 PMID 20090767
LOC84931 rs2030746 2 121.31 LDL, TC
FN1 rs1250229 2 216.30 LDL FN1 fibronectin 1 PMID 16150826
CMTM6 rs7640978 3 32.53 LDL, TC
ACAD11 rs17404153 3 132.16 LDL, HDL
CSNK1G3 rs4530754 5 122.86 LDL, TC
MIR148A rs4722551 7 25.99 LDL, TC, TG
SOX17 rs10102164 8 55.42 LDL, TC
BRCA2 rs4942486 13 32.95 LDL
APOH-PRXCA rs1801689 17 64.21 LDL APOH apolipoprotein H PMID 12740481
APOH-PRXCA rs1801689 17 64.21 LDL PRKCA protein kinase C, alpha PMID 20692055 PMID 12952980
SPTLC3 rs364585 20 12.96 LDL SPTLC3 serine palmitoyltransferase, long chain base subunit 3 PMID 19648650
SNX5 rs2328223 20 17.85 LDL SNX5 sorting nexin 5 PMID 15561769
MTMR3 rs5763662 22 30.38 LDL
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
23
Supplementary Table 4 (continued).
Loci Primarily Associated with Total Cholesterol
ASAP3 rs1077514 1 23.77 TC
ABCB11 rs2287623 2 169.83 TC ABCB11 ATP-binding cassette, sub-family B (MDR/TAP), member 11 PMID 21726512 PMID 19228692
FAM117B rs11694172 2 203.53 TC
UGT1A1 rs11563251 2 234.68 TC, LDL UGT1A1/3/4/5/6/7/8/9/20 UDP glucuronosyltransferase 1 family, polypeptide A1 PMID 17908920
PXK rs13315871 3 58.38 TC PXK PX domain containing serine/threonine kinase PMID 20086096 PMID 17178602
KCNK17 rs2758886 6 39.25 TC
HBS1L rs9376090 6 135.41 TC
GPR146 rs1997243 7 1.08 TC
VLDLR rs3780181 9 2.64 TC, LDL VLDLR very low density lipoprotein receptor PMID 8827514
VIM-CUBN rs10904908 10 17.26 TC VIM vimentin PMID 22535769 PMID 7706405 PMID 1527066
VIM-CUBN rs10904908 10 17.26 TC CUBN cubilin PMID 10371504
PHLDB1 rs11603023 11 118.49 TC
PHC1-A2ML1 rs4883201 12 9.08 TC A2ML1 alpha-2-macroglobulin-like 1 PMID 18648652
DLG4 rs314253 17 7.09 TC, LDL ACADVL acyl-CoA dehydrogenase, very long chain PMID 19889959
DLG4 rs314253 17 7.09 TC, LDL CTDNEP1 CTD nuclear envelope phosphatase 1 PMID 22134922
DLG4 rs314253 17 7.09 TC, LDL SLC2A4 solute carrier family 2, member 4 PMID 16096283
TOM1 rs138777 22 35.71 TC HMOX1 hemeoxygenase (decycling) 1 PMID 22004613
PPARA rs4253772 22 46.63 TC, LDL PPARA peroxisome proliferator-activated receptor alpha PMID 21540177 PMID 21487230
Loci Primarily Associated with Triglycerides
LRPAP1 rs6831256 4 3.47 TG, LDL, TC LRPAP1 low density lipoprotein receptor-related protein associated protein 1 PMID 16973241
VEGFA rs998584 6 43.76 TG, HDL VEGFA vascular endothelial growth factor A PMID 21348596 PMID 18789802
MET rs38855 7 116.36 TG
AKR1C4 rs1832007 10 5.25 TG AKR1C4 aldo-ketoreductase family 1, member C4 PMID 18024509
PDXDC1 rs3198697 16 15.13 TG
MPP3 rs8077889 17 41.88 TG
INSR rs7248104 19 7.22 TG
PEPD rs731839 19 33.90 TG, HDL CEBPG CCAAT/enhancer binding protein (C/EBP), gamma PMID 12177065
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
24
SUPPLEMENTARY TABLE 5. Pathways that Show Enrichment of Genes at Novel Loci by MAGENTA analysis
SUPPLEMENTARY TABLE 5A. Pathways that Show Enrichment of Genes at Novel HDL Associated Loci by MAGENTA analysis
Database Gene Set
Effective Gene Set
Size
No. of Expected
Genes (>95% Cutoff)
No. of Observed
Genes (>95% Cutoff)
FDR P-value
Genome-wide Significant Genes
Novel Known Other Enriched Genes
HDL Cholesterol
Ingenuity FXR RXR activation 54 3 13 6.0x10-4
NR0B2 SCARB1,LIPC,HNF4A,PLTP, APOB,APOE,MLXIPL,APOA1 APOC3,NR1H3,APOC2,NR1I2
GOTERM Cholesterol metabolic process 52 3 11 1.3x10-2
NR0B2 CETP,LCAT,STARD3,APOB, APOE,APOA1 ABCA1,APOA4,APOC3,APOC1
Ingenuity LPS IL-1 mediated inhibition of RXR function 52 3 9 2.0x10
-2 NR0B2
ABCA1,SCARB1,LIPC,CETP, PLTP,APOE APOC2,NR1I2
KEGG Neurotrophin signaling 116 6 16 2.1x10-2
AKT1,GSK3B SORT1
RAC1,RPS6KA1,SH2B3,NFKB1, NTRK1,MAP2K7,RELA,PLCG2, PRKCD,PTPN11,MAP2K2,TP53, MAPK10
Ingenuity PXR RXR activation 45 2 8 2.4x10-2
NR0B2 HNF4A
RELA,NR1I2,ABCB9,GSTM1,
INSR,CPT1A
KEGG Adipocytokine signaling 63 3 11 2.5x10-2
AKT1
AGRP,TRADD,SOCS3,ACSL5, NFKB1,RELA,PTPN11,CHUK, CPT1A,MAPK10
GOTERM Triglyceride lipase activity 15 1 5 2.5x10-2
DAGLB LIPC,LIPG,LPL DAGLA
Ingenuity NFKB signaling 39 2 7 2.9x10-2
GSK3B
RELB,CD40,NFKB1,RELA,
PLCG2,CHUK
Ingenuity PPARaRXRa activation 50 3 8 3.4x10-2
NR0B2 ABCA1,LPL,APOA1 CKAP5,MED1,NCOA6,INSR
GOTERM Enzyme binding 108 5 16 3.8x10-2
AKT1 UBE2L3,SORT1,APOB, APOA1
APOA5,RAC1,CSF3,CD40, PRKCD,PLAUR,DNM2,CBX1, TP53,MIZF,HMGA1
GOTERM Phospholipid binding 47 2 9 4.2x10-2
CPS1 ABCA1,APOB,APOE,APOA1 APOA5,LYPLA3,APOC3, MAP1LC3A
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
25
Supplementary Tables 5A-D.Database and Gene Set define the source of the gene set with evidence for enrichment; Effective Gene Set Size, the number of genes in a pathway
independently assigned a score, after clustering nearby genes and excluding genes in regions with no SNP data; Expected Number of Hits, the number of genes expected to have
a score in the top 5% of all scores given the gene set size; Observed Number of Hits, the number of genes observed in the top 5% of all gene scores; FDR P-value, the false
discovery rate incurred by rejecting the null for this gene set and all others with more extreme enrichmentusing all GWAS+Metabochip results; Genome-wide Significant
Genes,genes in the pathway labeled as hits by MAGENTA (Known and Novel refer to association evidence reported by this study); Other Enriched Genes, genes with scores in the
top 5% of all gene scores but that do not reach genome-wide significance. We show here significant pathways (FDR p< .05) which contain at least one gene from one of the 62
Novel loci.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
26
SUPPLEMENTARY TABLE 5B. Pathways that Show Enrichment of Genes at Novel LDL Associated Loci by MAGENTA analysis
Database Gene Set
Effective Gene Set
Size
No. of Expected
Genes (>95% Cutoff)
No. of Observed
Genes (>95% Cutoff)
FDR P-value
Genome-wide Significant Genes
Novel Known Other Enriched Genes
LDL Cholesterol
GOTERM Cholesterol metabolic process 53 3 18 < 3.3x10-5
NR0B2,INSIG2, VLDLR,CUBN
ABCA1,CETP,PCSK9,APOB, LDLR,APOE,LDLRAP1,HNF1A, ANGPTL3,APOA1 APOC1,APOA4,APOC3,PPARD
Ingenuity FXR RXR activation 54 3 16 < 3.3x10-5
NR0B2,PPARA,VLDLR, ABCB11
HNF4A,APOB,APOE,
CYP7A1,HNF1A,APOA1
PPARG,APOC2,ABCG5,ABCG8,APOC3,
MTTP
GOTERM Lipoprotein metabolic process 15 1 7 1.7x10-4
PPARA PCSK9,NPC1L1 APOC1,APOA5,APOA4,APOC3
GOTERM Lipid transport 61 3 14 1.6x10-3
VLDLR
CETP,APOB,LPA,LDLR,
APOE,APOA1 APOC1,APOC2,APOC4,COL4A3BP,APOA5,APOA4,APOC3
Ingenuity LPS I-1 mediated inhibition of RXR function 53 3 11 2.2x10
-3
NR0B2,PPARA, ABCB11 ABCA1,CETP,APOE,CYP7A1 APOC2,ABCG5,ABCG8,LY96
GOTERM Low-density lipoprotein receptor binding 11 1 5 2.9x10
-3 LRPAP1 PCSK9,APOB,APOE APOA5
GOTERM
Negative regulation of macrophage derived foam cell differentiation 12 1 5 3.4x10
-3 PPARA ABCA1,CETP ITGB3,PPARG
Ingenuity Hepatic cholestasis 58 3 11 3.4x10-3
NR0B2,PPARA, ABCB11
CETP,HNF4A,CYP7A1, HNF1A TIRAP,MAP3K4,LY96,NR1H4
GOTERM Steroid metabolic process 67 3 13 6.5x10-3
INSIG2,VLDLR, CUBN
ABCA1,CETP,PCSK9,APOB, OSBPL7,LDLR,LDLRAP1, NPC1L1,CYP7A1 SORL1
Ingenuity PPAR Signaling 18 1 5 7.9x10-3
NR0B2,PPARA RAF1 PPARG,PPARD
GOTERM Phosphatidylserine binding 10 1 4 1.3x10-2
ANXA9 SCARB1 CPNE1,TRIM72
Ingenuity PXR RXR activation 46 2 8 1.7x10-2
NR0B2,PPARA, ABCB11, UGT1A1 HNF4A,CYP7A1 GSTM1,UGT1A9
GOTERM Lipoprotein transport 10 1 4 1.7x10-2
CUBN APOB PPARG,MTTP
Nature Genetics: doi:10.1038/ng.2797
GLGC Metabochip Supplement 2013
27
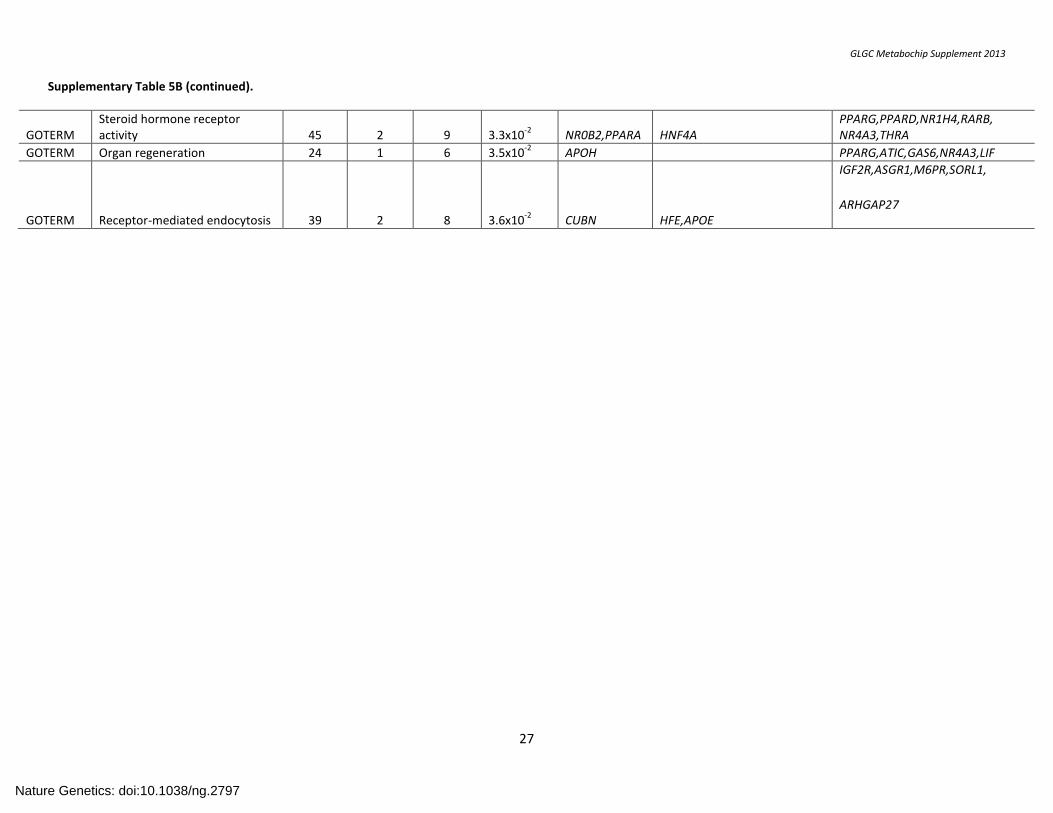
Supplementary Table 5B (continued).
GOTERM Steroid hormone receptor activity 45 2 9 3.3x10
-2 NR0B2,PPARA HNF4A
PPARG,PPARD,NR1H4,RARB, NR4A3,THRA
GOTERM Organ regeneration 24 1 6 3.5x10-2
APOH PPARG,ATIC,GAS6,NR4A3,LIF
GOTERM Receptor-mediated endocytosis 39 2 8 3.6x10-2
CUBN HFE,APOE
IGF2R,ASGR1,M6PR,SORL1,
ARHGAP27
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
28
SUPPLEMENTARY TABLE 5C. Pathways that Show Enrichment of Genes at Novel Total Cholesterol Associated Loci by MAGENTA analysis
Database Gene Set
Effective Gene Set
Size
No. of Expected
Genes (>95% Cutoff)
No. of Observed
Genes (>95% Cutoff)
FDR P-value
Genome-wide Significant Genes
Novel Known Other Enriched Genes
Total Cholesterol
GOTERM Cholesterol metabolic process 52 3 18 < 3.3x10-5
CUBN,INSIG2,VLDLR
ABCA1,CETP,LCAT,PCSK9, APOB,LDLR,APOE,LDLRAP1, HNF1A,ANGPTL3,APOA1 APOC1,APOA4,APOC3,PPARD
Ingenuity FXR RXR activation 54 3 18 < 3.3x10-5
ABCB11,PPARA,VLDLR,NR0B2
SCARB1,LIPC,HNF4A,APOB,
APOE,CYP7A1,HNF1A,
APOA1
APOC2,ABCG5,ABCG8,APOC3,
PPARG,SDC1
Ingenuity LPS IL-1 Mediated Inhibition of RXR Function 53 3 14 6.7x10
-5
ABCB11,PPARA,NR0B2
ABCA1,SCARB1,LIPC,CETP, APOE,CYP7A1
APOC2,ABCG5,ABCG8,LY96, ABCB9
GOTERM Low-density lipoprotein receptor binding 11 1 6 1.6x10
-4 LRPAP1 PCSK9,APOB,APOE APOA5,SNX17
Ingenuity Hepatic cholestasis 56 3 12 7.5x10-4
ABCB11,PPARA,NR0B2 CETP,HNF4A,CYP7A1,HNF1A
LY96,NR1I2,MAP3K4,NR1H4,
HSD3B7
Ingenuity PXR RXR activation 46 2 10 1.1x10-3
ABCB11,PPARA,UGT1A1, NR0B2 HNF4A,CYP7A1 GSTM1,UGT1A9,ABCB9,NR1I2
GOTERM
Negative regulation of macrophage-derived foam cell differentiation 12 1 5 4.2x10
-3 PPARA ABCA1,CETP PPARG,ITGB3
Ingenuity PPAR signaling 18 1 5 7.7x10-3
PPARA,NR0B2 RAF1 PPARG,PPARD
GOTERM Steroid metabolic process 68 3 13 9.8x10-3
CUBN,INSIG2, VLDLR
ABCA1,CETP,LCAT,PCSK9,APOB,LDLR,LDLRAP1,NPC1L1,CYP7A1 NR1I2
GOTERM Gamma-tubulin binding 10 1 4 1.6x10-2
BRCA2 SPATC1,MARK4,BLOC1S2
GOTERM Phosphatidylserine binding 10 1 4 2.0x10-2
ANXA9 SCARB1 CPNE1,TRIM72
Ingenuity NRF2-mediated oxidative stress Response 50 3 8 3.3x10
-2 GSK3B SCARB1,RAF1
HERPUD1,KEAP1,ERP29, HMOX1,FTH1
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
29
Supplementary Table 5C (continued).
Ingenuity Axonal guidance signaling 65 3 9 3.3x10-2
GSK3B RAF1 SDCBP,VASP,PTPN11,CXCR4, ARHGEF15,GDF7,ERBB2
Ingenuity Neuregulin signaling 25 1 5 3.4x10-2
DLG4 RAF1 PTPN11,RPS6,GRB7
Ingenuity Estrogen receptor signaling 30 2 5 4.4x10-2
NR0B2 RAF1 SMARCA4,CARM1,PELP1
Ingenuity PPARaRXRa activation 50 3 7 4.5x10-2
PPARA,NR0B2 ABCA1,RAF1,APOA1 MED24,MED1
GOTERM Receptor-mediated endocytosis 39 2 8 4.7x10-2
CUBN APOE
IGF2R,ASGR1,M6PR,CXCL16,
PLD2,SNX17
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
30
SUPPLEMENTARY TABLE 5D. Pathways that Show Enrichment of Genes at Novel Triglyceride Associated Loci by MAGENTA analysis
Database Gene Set
Effective Gene Set
Size
No. of Expected
Genes (>95% Cutoff)
No. of Observed
Genes (>95% Cutoff)
FDR P-value
Genome-wide Significant Genes
Novel Known Other Enriched Genes
Triglycerides
Ingenuity FXR RXR activation 54 3 15 < 3.3x10-5
NR0B2 SCARB1,LIPC,PLTP,APOB,APOE,CYP7A1,MLXIPL,APOA1
APOC2,APOC3,PPARG,CYP27A1,NR1H3,SLCO1B1
GOTERM Low-density lipoprotein receptor binding 11 1 5 2.3x10
-3 LRPAP1 APOB,APOE SNX17,APOA5
KEGG Primary bile acid biosynthesis 16 1 5 1.0x10-2
AKR1C4 CYP7A1 HSD3B7,CYP27A1,HSD17B4
GOTERM Cholesterol metabolic process 52 3 11 1.2x10-2
NR0B2
ABCA1,CETP,APOB,APOE,
ANGPTL3,APOA1 APOA4,APOC1,APOC3,CYP27A1
Ingenuity PPAR signaling 18 1 5 1.4x10-2
INSR,NR0B2 RAF1 PPARG,NR1H3
GOTERM Cell surface 184 9 23 3.0x10-2
VEGFA,LRPAP1,MPP3 SCARB1
PVRL2,EDG4,BACE1,STX4,BCAM,STRC,FLT3LG,MPP2,TMEM102, PCSK6,DSCAML1,HSPB1,CD6, C9orf127,BMPR2,IGF2R,ITGAL, SDC1,HFE2
Ingenuity LPS IL-1 mediated inhibition of RXR function 51 3 8 5.0x10
-2 NR0B2
ABCA1,LIPC,CETP,PLTP,APOE,CYP7A1 APOC2
Nature Genetics: doi:10.1038/ng.2797
GLGC Metabochip Supplement 2013
31
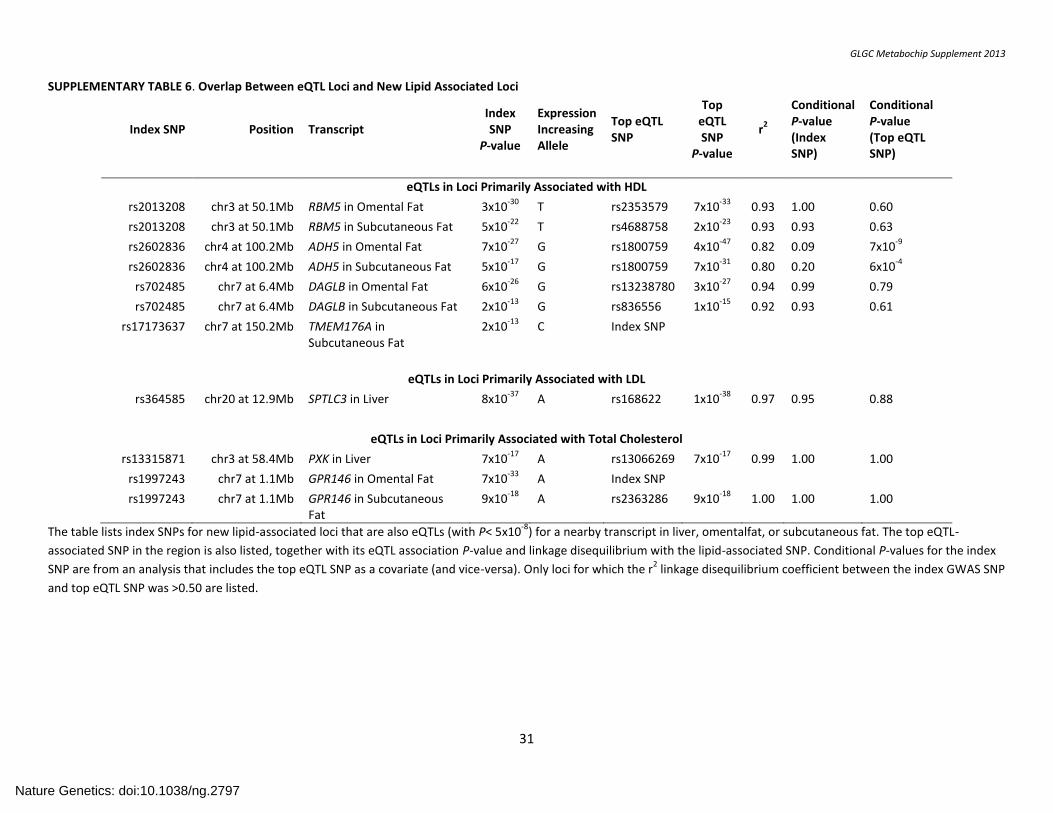
SUPPLEMENTARY TABLE 6. Overlap Between eQTL Loci and New Lipid Associated Loci
Index SNP Position Transcript Index SNP
P-value
Expression Increasing Allele
Top eQTL SNP
Top eQTL SNP
P-value
r2
Conditional P-value (Index SNP)
Conditional P-value (Top eQTL SNP)
eQTLs in Loci Primarily Associated with HDL
rs2013208 chr3 at 50.1Mb RBM5 in Omental Fat 3x10-30
T rs2353579 7x10-33
0.93 1.00 0.60
rs2013208 chr3 at 50.1Mb RBM5 in Subcutaneous Fat 5x10-22
T rs4688758 2x10-23
0.93 0.93 0.63
rs2602836 chr4 at 100.2Mb ADH5 in Omental Fat 7x10-27
G rs1800759 4x10-47
0.82 0.09 7x10-9
rs2602836 chr4 at 100.2Mb ADH5 in Subcutaneous Fat 5x10-17
G rs1800759 7x10-31
0.80 0.20 6x10-4
rs702485 chr7 at 6.4Mb DAGLB in Omental Fat 6x10-26
G rs13238780 3x10-27
0.94 0.99 0.79
rs702485 chr7 at 6.4Mb DAGLB in Subcutaneous Fat 2x10-13
G rs836556 1x10-15
0.92 0.93 0.61
rs17173637 chr7 at 150.2Mb TMEM176A in Subcutaneous Fat
2x10-13
C Index SNP
eQTLs in Loci Primarily Associated with LDL
rs364585 chr20 at 12.9Mb SPTLC3 in Liver 8x10-37
A rs168622 1x10-38
0.97 0.95 0.88
eQTLs in Loci Primarily Associated with Total Cholesterol
rs13315871 chr3 at 58.4Mb PXK in Liver 7x10-17
A rs13066269 7x10-17
0.99 1.00 1.00
rs1997243 chr7 at 1.1Mb GPR146 in Omental Fat 7x10-33
A Index SNP
rs1997243 chr7 at 1.1Mb GPR146 in Subcutaneous Fat
9x10-18
A rs2363286 9x10-18
1.00 1.00 1.00
The table lists index SNPs for new lipid-associated loci that are also eQTLs (with P< 5x10-8
) for a nearby transcript in liver, omentalfat, or subcutaneous fat. The top eQTL-
associated SNP in the region is also listed, together with its eQTL association P-value and linkage disequilibrium with the lipid-associated SNP. Conditional P-values for the index
SNP are from an analysis that includes the top eQTL SNP as a covariate (and vice-versa). Only loci for which the r2 linkage disequilibrium coefficient between the index GWAS SNP
and top eQTL SNP was >0.50 are listed.
Nature Genetics: doi:10.1038/ng.2797
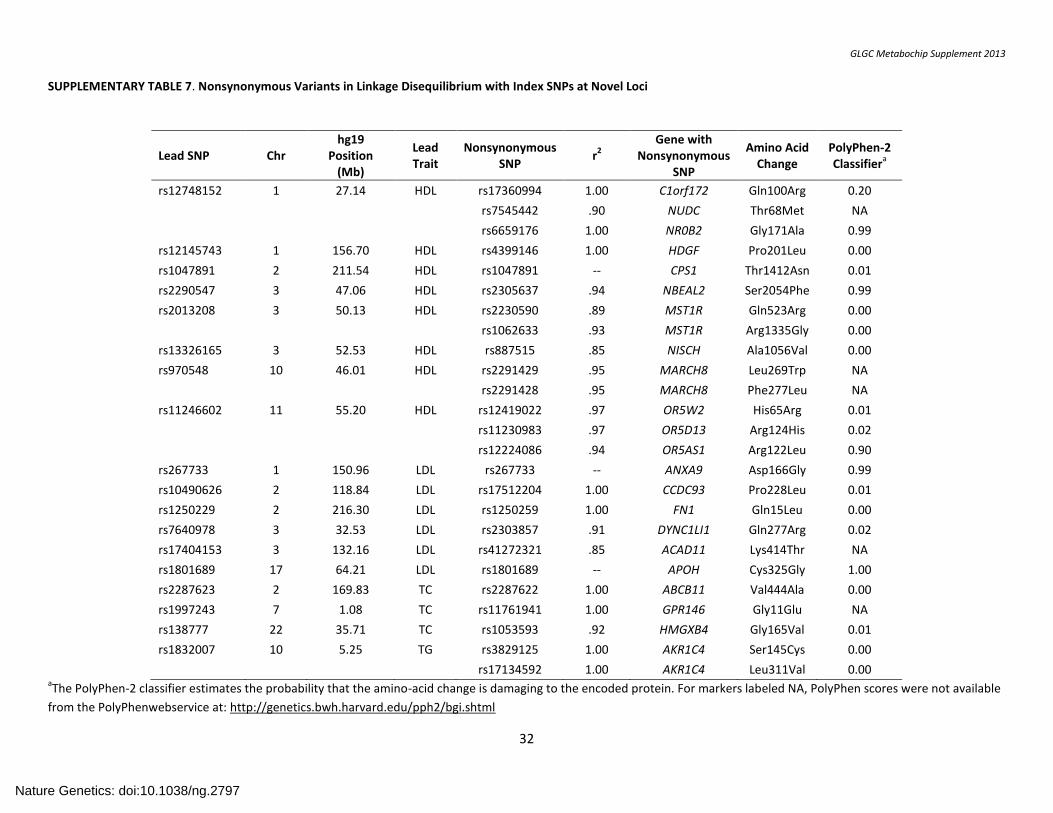
GLGC Metabochip Supplement 2013
32
SUPPLEMENTARY TABLE 7. Nonsynonymous Variants in Linkage Disequilibrium with Index SNPs at Novel Loci
Lead SNP Chr hg19
Position (Mb)
Lead Trait
Nonsynonymous SNP
r2
Gene with Nonsynonymous
SNP
Amino Acid Change
PolyPhen-2 Classifier
a
rs12748152 1 27.14 HDL rs17360994 1.00 C1orf172 Gln100Arg 0.20
rs7545442 .90 NUDC Thr68Met NA
rs6659176 1.00 NR0B2 Gly171Ala 0.99
rs12145743 1 156.70 HDL rs4399146 1.00 HDGF Pro201Leu 0.00
rs1047891 2 211.54 HDL rs1047891 -- CPS1 Thr1412Asn 0.01
rs2290547 3 47.06 HDL rs2305637 .94 NBEAL2 Ser2054Phe 0.99
rs2013208 3 50.13 HDL rs2230590 .89 MST1R Gln523Arg 0.00
rs1062633 .93 MST1R Arg1335Gly 0.00
rs13326165 3 52.53 HDL rs887515 .85 NISCH Ala1056Val 0.00
rs970548 10 46.01 HDL rs2291429 .95 MARCH8 Leu269Trp NA
rs2291428 .95 MARCH8 Phe277Leu NA
rs11246602 11 55.20 HDL rs12419022 .97 OR5W2 His65Arg 0.01
rs11230983 .97 OR5D13 Arg124His 0.02
rs12224086 .94 OR5AS1 Arg122Leu 0.90
rs267733 1 150.96 LDL rs267733 -- ANXA9 Asp166Gly 0.99
rs10490626 2 118.84 LDL rs17512204 1.00 CCDC93 Pro228Leu 0.01
rs1250229 2 216.30 LDL rs1250259 1.00 FN1 Gln15Leu 0.00
rs7640978 3 32.53 LDL rs2303857 .91 DYNC1LI1 Gln277Arg 0.02
rs17404153 3 132.16 LDL rs41272321 .85 ACAD11 Lys414Thr NA
rs1801689 17 64.21 LDL rs1801689 -- APOH Cys325Gly 1.00
rs2287623 2 169.83 TC rs2287622 1.00 ABCB11 Val444Ala 0.00
rs1997243 7 1.08 TC rs11761941 1.00 GPR146 Gly11Glu NA
rs138777 22 35.71 TC rs1053593 .92 HMGXB4 Gly165Val 0.01
rs1832007 10 5.25 TG rs3829125 1.00 AKR1C4 Ser145Cys 0.00
rs17134592 1.00 AKR1C4 Leu311Val 0.00 aThe PolyPhen-2 classifier estimates the probability that the amino-acid change is damaging to the encoded protein. For markers labeled NA, PolyPhen scores were not available
from the PolyPhenwebservice at: http://genetics.bwh.harvard.edu/pph2/bgi.shtml
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
33
SUPPLEMENTARY TABLE 8. Overlap of SNPs at Known and Novel Lipid Loci with Chromatin States in 9 Different Cell Types
Cell Type
Observed Number of Chromatin States* Showing Excess Overlap
with Lipid Loci (of 13 tested, P< 1x10-5)
Chromatin States* Showing Excess Overlap with Lipid Loci (P< 1x10-5)
H1 embryonic stem cells (H1 ES) 2 Transcription Transition (HMM9) P =4x10-10 Transcription Elongation (HMM10) P =5x10-10
B-lymphoblastoid cells (GM12878) 0
Umbilical vein endothelial cells (HUVEC) 2 Transcription Transition (HMM9) P =2x10-7 Transcription Elongation (HMM10) P =6x10-7
Skeletal muscle myoblasts (HSMM) 1 Transcription Elongation (HMM10) P =6x10-8
Mammary epithelial cells (HMEC) 2 Transcription Transition (HMM9) P =6x10-11 Transcription Elongation (HMM10) P =2x10-9
Normal epidermal keratinocytes (NHEK) 2 Transcription Elongation (HMM10) P =2x10-8 Weak Transcription (HMM11) P =3x10-6
Normal lung fibroblasts (NHLF) 2 Transcription Elongation (HMM10) P =2x10-10 Transcription Transition (HMM9) P =8x10-8
Erythrocyticleukaemia cells (K562) 3 Weak Transcription (HMM11) P =1x10-11 Weak Enhancer (HMM7) P =2x10-10 Strong Enhancer (HMM5) P =4x10-8
Hepatocellular carcinoma cells (HepG2) 8
Strong Enhancer (HMM4) P =2x10-25 Weak Enhancer (HMM7) P =4x10-14 Weak Transcription (HMM11) P =2x10-11 Strong Enhancer (HMM5) P =5x10-11 Transcription Elongation (HMM10) P =3x10-10 Weak Enhancer (HMM6) P =1x10-7 Active Promoter (HMM1) P =4x10-7 Weak Promoter (HMM2) P =7x10-7
*Chromatin states were described previously (Ernst J et al. Nature 473, 43-9, 2011) based on hidden Markov models of histone
methylation and acetylation marks from 9 cell types. SNPs in high linkage disequilibrium (r2> 0.8 in 1000 Genomes Project
European ancestry samples) with known or novel lipid loci was compared to matched sets of HapMap SNPs (seeMethods).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
34
SUPPLEMENTARY TABLE 9. Overlap with Chromatin States, Histone Marks and Transcription Factor ChIP-Seq in HepG2 Cells
Known and Novel Lipid Loci (N=157) Only Novel Lipid Loci (N=62)
Observed Number of
Loci with 1 SNP in a
Regulatory Region
Expected Number of Loci
P-value
Observed Number of
Loci with 1 SNP in a
Regulatory Region
Expected Number of
Loci P-value
Overlap with Chromatin States from Ernst et al.* (13 tested)
Strong Enhancer (HMM4) 49 13.7 2x10-25
20 6.2 9x10-10
Weak Enhancer (HMM7) 60 26.9 4x10
-14 25 11.9 3x10
-5
Weak Transcription (HMM11) 99 62.1 2x10-11
41 26.4 9x10-5
Strong Enhancer (HMM5) 34 12.8 5x10
-11 10 5.6 5x10
-2
Transcription Elongation (HMM10)
65 35.4 3x10-10
26 15.4 1x10-3
Weak Enhancer (HMM6) 57 33.5 1x10-7
21 14.5 .013 Active Promoter (HMM1) 39 20.3 4x10
-7 14 8.8 .039
Weak Promoter (HMM2) 45 24.8 7x10-7
15 10.6 .088 Transcription Transition (HMM9)
37 18.7 3x10-5
18 8.0 4x10-4
Overlap with Histone Marks (5 tested)
H3K9ac 97 47.3 3x10-22
37 20.1 6x10-8
H3K27ac 84 39.2 3x10
-20 34 16.7 4x10
-8
H3K4me3 88 47.9 2x10-15
34 20.1 7x10-5
H3K36me3 104 62.3 4x10
-14 41 26.1 2x10
-5
H3K4me2 111 74.3 8x10-12
44 31.1 7x10-5
Overlap with Open Chromatin (2 tested)
FAIRE 51 26.5 5x10-9
19 11.3 8x10-3
DNase hypersensitivity 33 18.3 2x10
-4 12 8.1 .09
Overlap with Transcription Factor ChIP-Seq (11 tested)
HNF4Α 38 16.2 6x10-10
14 7.1 6x10-3
CEBP/Β 40 20.4 1x10-5
16 9.1 .010
CTCF 55 37.6 4x10-4
21 16.2 .055
HSF1 9 2.6 1x10-3
4 1.1 .024
*Chromatin states were described previously (Ernst J et al. Nature 473, 43-9, 2011) based on hidden Markov models of histone
methylation and acetylation marks from 9 cell types.Data for histone marks, open chromatin, and transcription factor ChIP-seq were
obtained from the ENCODE Project (ENCODE Project Consortium, PLoS Biol. 9:e1001046, 2011). SNPs in high linkage disequilibrium
(r2> .8 in 1000 Genomes Project European ancestry samples) with known or novel lipid loci were compared to matched sets of
HapMap SNPs (see Methods). The table lists only regulatory elements that exhibited a significant excess overlap (P< 1x10-3
to
account for 31 HepG2 regulatory elements tested).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
35
SUPPLEMENTARY TABLE 10. Overlap of Regulatory Features and Associated SNPs at Novel Lipid Loci
Hidden Markov model-defined regulatory domains from histone methylation marks Histone methylation marks Markers of open chromatin
Transcription factor binding (ChIP-Seq)
Locus Lead SNP Strong Enhancer (HMM4)
Weak Enhancer (HMM7)
WeakTxn (HMM11)
Strong Enhancer (HMM5)
Txn Elongation (HMM10)
Weak Enhancer (HMM6)
Active Promoter (HMM1)
Weak Promoter (HMM2)
Txn Transition (HMM9)
H3k9ac H3k27ac H3k4me3 H3k36me3 H3k4me2 FAIRE DNase
Hnf4A (Forskolin)
Cebpb (Forskolin)
CTCF Hsf1
Loci Primarily Associated with HDL Cholesterol
PIGV- NR0B2
rs12748152 x x x x x x x x x x x x x x x x x x x x
HDGF- PMVK
rs12145743 x x x x x x x x x x x x x x x x
ANGPTL1 rs4650994 x x x x x x x x x x x x x x
CPS1 rs1047891 x x
ATG7 rs2606736 x x x x x x x x x x x x x
SETD2 rs2290547 x x x x x x x
RBM5 rs2013208 x x x x x x x x x x x x x x x
STAB1 rs13326165 x x
GSK3B rs6805251 x x x x x x x x
C4orf52 rs10019888 x
FAM13A rs3822072 x x x x x x x x x x x x x x
ADH5 rs2602836 x x x x x x x x x x x x
RSPO3 rs1936800 x x x
DAGLB rs702485 x x x x x x x x
SNX13 rs4142995 x x
IKZF1 rs4917014
TMEM176A rs17173637 x x x x x
MARCH8- ALOX5
rs970548 x x x x x x x x x
OR4C46 rs11246602 x x x x x x x
KAT5 rs12801636 x x x x x x x x x x
MOGAT2- DGAT2
rs499974 x x x x x x x x
ZBTB42- AKT1
rs4983559 x x
FTO rs1121980 x x x x x x x x x x x x
HAS1 rs17695224
Nature Genetics: doi:10.1038/ng.2797
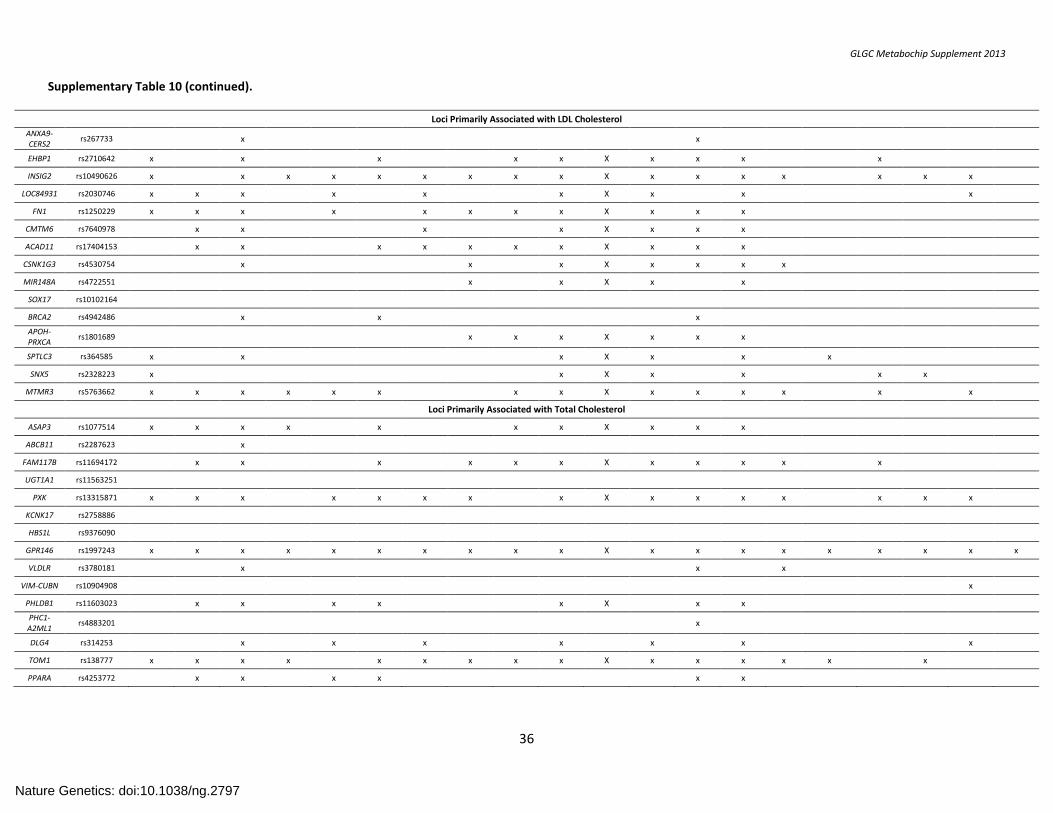
GLGC Metabochip Supplement 2013
36
Supplementary Table 10 (continued).
Loci Primarily Associated with LDL Cholesterol
ANXA9- CERS2
rs267733 x x
EHBP1 rs2710642 x x x x x X x x x x
INSIG2 rs10490626 x x x x x x x x x X x x x x x x x
LOC84931 rs2030746 x x x x x x X x x x
FN1 rs1250229 x x x x x x x x X x x x
CMTM6 rs7640978 x x x x X x x x
ACAD11 rs17404153 x x x x x x x X x x x
CSNK1G3 rs4530754 x x x X x x x x
MIR148A rs4722551 x x X x x
SOX17 rs10102164
BRCA2 rs4942486 x x x
APOH- PRXCA
rs1801689 x x x X x x x
SPTLC3 rs364585 x x x X x x x
SNX5 rs2328223 x x X x x x x
MTMR3 rs5763662 x x x x x x x x X x x x x x x
Loci Primarily Associated with Total Cholesterol
ASAP3 rs1077514 x x x x x x x X x x x
ABCB11 rs2287623 x
FAM117B rs11694172 x x x x x x X x x x x x
UGT1A1 rs11563251
PXK rs13315871 x x x x x x x x X x x x x x x x
KCNK17 rs2758886
HBS1L rs9376090
GPR146 rs1997243 x x x x x x x x x x X x x x x x x x x x
VLDLR rs3780181 x x x
VIM-CUBN rs10904908 x
PHLDB1 rs11603023 x x x x x X x x
PHC1- A2ML1
rs4883201 x
DLG4 rs314253 x x x x x x x
TOM1 rs138777 x x x x x x x x x X x x x x x x
PPARA rs4253772 x x x x x x
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
37
Supplementary Table 10 (continued).
Loci Primarily Associated with Triglycerides
LRPAP1 rs6831256 x x x x
VEGFA rs998584 x x
MET rs38855 x x x
AKR1C4 rs1832007 x x x x X x x x x x
PDXDC1 rs3198697 x x
MPP3 rs8077889 x x x x x x x X x x x x x x x
INSR rs7248104 x x x x
PEPD rs731839 x x x x X x x x x x x x
Supplementary Table 10 annotates overlap (denoted as x) between regulatory features and either the index SNP or a variant in high linkage disequilibrium (r2>0.8) with the index
SNP. Regulatory features were obtained from Ernst J et al. Nature 473, 43-9, 2011, and the ENCODE Project (ENCODE Project Consortium, PLoS Biol. 9:e1001046, 2011). The
corresponding BED file available on the UCSC Genome Browser for each regulatory feature is listed below.
Strong Enhancer (HMM4): wgEncodeBroadHmmHepg2HMM.bed.4_Strong_Enhancer.bed Weak Enhancer (HMM7): wgEncodeBroadHmmHepg2HMM.bed.7_Weak_Enhancer.bed WeakTxn (HMM11): wgEncodeBroadHmmHepg2HMM.bed.11_Weak_Txn.bed Strong Enhancer (HMM5): wgEncodeBroadHmmHepg2HMM5_Strong_Enhancer.bed Txn Elongation (HMM10): wgEncodeBroadHmmHepg2HMM.bed.10_Txn_Elongation.bed Weak Enhancer (HMM6): wgEncodeBroadHmmHepg2HMM.bed.6_Weak_Enhancer.bed Active Promoter (HMM1): wgEncodeBroadHmmHepg2HMM1_Active_Promoter.bed Weak Promoter (HMM2): wgEncodeBroadHmmHepg2HMM.bed.2_Weak_Promoter.bed Txn Transition (HMM9): wgEncodeBroadHmmHepg2HMM.bed.9_Txn_Transition.bed H3k9ac: wgEncodeBroadChipSeqPeaksHepg2H3k9ac.bed H3k27ac: wgEncodeBroadChipSeqPeaksHepg2H3k27ac.bed H3k4me3: wgEncodeBroadChipSeqPeaksHepg2H3k4me3.bed H3k36me3: wgEncodeBroadChipSeqPeaksHepg2H3k36me3.bed H3k4me2: wgEncodeBroadChipSeqPeaksHepg2H3k4me2.bed FAIRE: wgEncodeUncFAIRESeqPeaksHepg2V3.bed DNase: wgEncodeUwDnaseSeqPeaksRep1Hepg2.bed Hnf4a-Forskln: wgEncodeYaleChIPseqPeaksHepg2Hnf4aForskln.narrowPeak Cebpb-Forskln: wgEncodeYaleChIPseqPeaksHepg2CebpbForskln.narrowPeak CTCF:wgEncodeBroadChipSeqPeaksHepg2Ctcf HSF1:wgEncodeYaleChIPseqPeaksHepg2Hsf1Forskln.narrowPeak
Nature Genetics: doi:10.1038/ng.2797
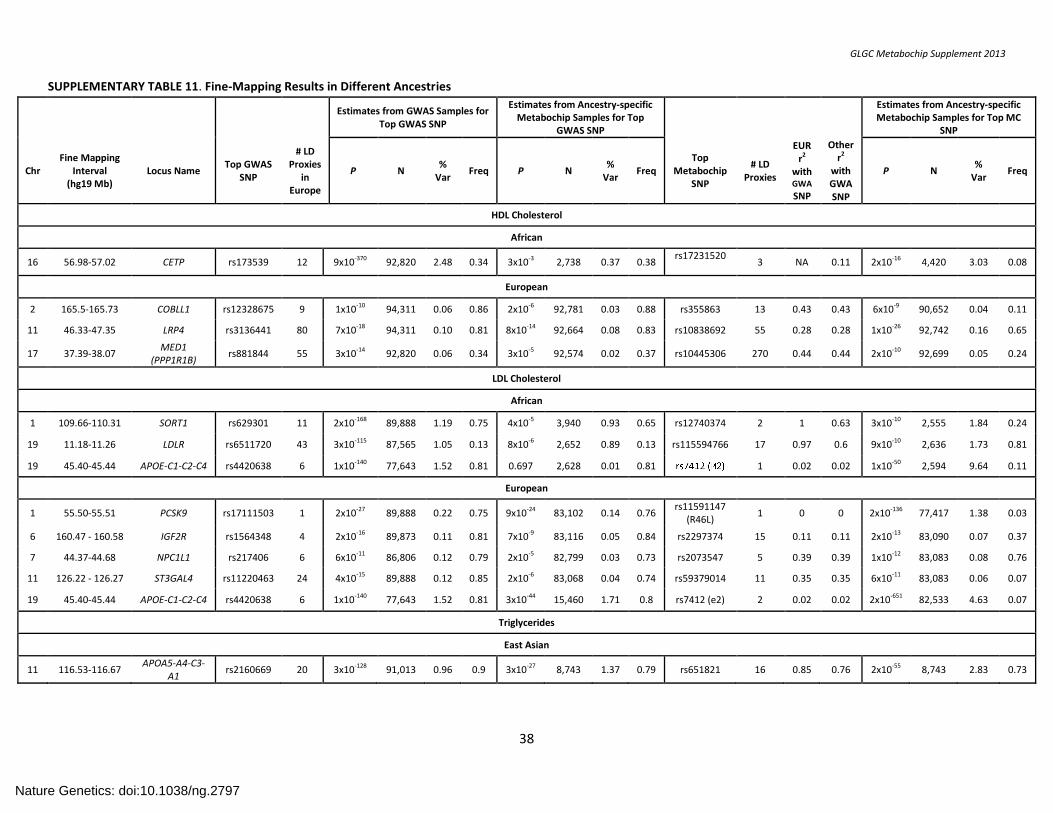
GLGC Metabochip Supplement 2013
38
SUPPLEMENTARY TABLE 11. Fine-Mapping Results in Different Ancestries
Estimates from GWAS Samples for
Top GWAS SNP
Estimates from Ancestry-specific Metabochip Samples for Top
GWAS SNP
Estimates from Ancestry-specific Metabochip Samples for Top MC
SNP
Chr Fine Mapping
Interval (hg19 Mb)
Locus Name Top GWAS
SNP
# LD Proxies
in Europe
P N %
Var Freq P N
% Var
Freq Top
Metabochip SNP
# LD Proxies
EUR r2
with GWA
SNP
Other r2
with GWA SNP
P N %
Var Freq
HDL Cholesterol
African
16 56.98-57.02 CETP rs173539 12 9x10-370 92,820 2.48 0.34 3x10-3 2,738 0.37 0.38 rs17231520
3 NA 0.11 2x10-16 4,420 3.03 0.08
European
2 165.5-165.73 COBLL1 rs12328675 9 1x10-10 94,311 0.06 0.86 2x10-6 92,781 0.03 0.88 rs355863 13 0.43 0.43 6x10-9 90,652 0.04 0.11
11 46.33-47.35 LRP4 rs3136441 80 7x10-18 94,311 0.10 0.81 8x10-14 92,664 0.08 0.83 rs10838692 55 0.28 0.28 1x10-26 92,742 0.16 0.65
17 37.39-38.07 MED1
(PPP1R1B) rs881844 55 3x10-14 92,820 0.06 0.34 3x10-5 92,574 0.02 0.37 rs10445306 270 0.44 0.44 2x10-10 92,699 0.05 0.24
LDL Cholesterol
African
1 109.66-110.31 SORT1 rs629301 11 2x10-168 89,888 1.19 0.75 4x10-5 3,940 0.93 0.65 rs12740374 2 1 0.63 3x10-10 2,555 1.84 0.24
19 11.18-11.26 LDLR rs6511720 43 3x10-115 87,565 1.05 0.13 8x10-6 2,652 0.89 0.13 rs115594766 17 0.97 0.6 9x10-10 2,636 1.73 0.81
19 45.40-45.44 APOE-C1-C2-C4 rs4420638 6 1x10-140 77,643 1.52 0.81 0.697 2,628 0.01 0.81 1 0.02 0.02 1x10-50 2,594 9.64 0.11
European
1 55.50-55.51 PCSK9 rs17111503 1 2x10-27 89,888 0.22 0.75 9x10-24 83,102 0.14 0.76 rs11591147
(R46L) 1 0 0 2x10-136 77,417 1.38 0.03
6 160.47 - 160.58 IGF2R rs1564348 4 2x10-16 89,873 0.11 0.81 7x10-9 83,116 0.05 0.84 rs2297374 15 0.11 0.11 2x10-13 83,090 0.07 0.37
7 44.37-44.68 NPC1L1 rs217406 6 6x10-11 86,806 0.12 0.79 2x10-5 82,799 0.03 0.73 rs2073547 5 0.39 0.39 1x10-12 83,083 0.08 0.76
11 126.22 - 126.27 ST3GAL4 rs11220463 24 4x10-15 89,888 0.12 0.85 2x10-6 83,068 0.04 0.74 rs59379014 11 0.35 0.35 6x10-11 83,083 0.06 0.07
19 45.40-45.44 APOE-C1-C2-C4 rs4420638 6 1x10-140 77,643 1.52 0.81 3x10-44 15,460 1.71 0.8 rs7412 (e2) 2 0.02 0.02 2x10-651 82,533 4.63 0.07
Triglycerides
East Asian
11 116.53-116.67 APOA5-A4-C3-
A1 rs2160669 20 3x10-128 91,013 0.96 0.9 3x10-27 8,743 1.37 0.79 rs651821 16 0.85 0.76 2x10-55 8,743 2.83 0.73
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
39
Supplementary Table 11: Locus labels are from Teslovichet al. (2010). # LD Proxies in Europe, the number of SNPs r2> 0.7 with GWAS SNP in 1000 Genomes European Ancestry
samples; # Ancestry-Specific LD Proxies,number of SNPs r2> 0.7 with top Metabochip SNP in the relevant ancestry group; EUR r
2, LD between top GWAS SNP and top Metabochip
SNP in European ancestry samples; Other r2, LD between top GWAS SNP and top Metabochip SNP in the relevant ancestry group.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
40
SUPPLEMENTARY TABLE 12. Novel and Known Lipid Loci Associated with BMI, CAD, DBP, SBP, Fasting Glucose, T2D, and WHR adj
BMI
In silico Association Results (P< .05) at Lipid Associated Loci for A. Body Mass Index (BMI), B. Coronary Artery Disease (CAD), C.
Diastolic Blood Pressure (DBP), D. Systolic Blood Pressure (SBP), E. Fasting Glucose (FG), F. Type 2 Diabetes (T2D), and G. Waist-Hip
Ratio adjusted for BMI (WHRadjBMI)
SUPPLEMENTARY TABLE 12A. Novel and Known Lipid Loci with BMI P-value < 0.05 from GIANT*
Locus SNP Chr hg19 Pos (Mb) Type Trait A1/A2
Lipid Direction Lipid N
Lipid P-value
BMI Direction BMI N
BMI P-value
FTO rs1121980 16 53.81 novel HDL A/G - 185,524 6.8x10-9
+ 123,845 1.8x10-57
MC4R rs12967135 18 57.85 known HDL A/G - 153,533 3.6x10-8
+ 123,864 5.3x10-22
SLC39A8 rs13107325 4 103.19 known HDL T/C - 179,316 1.1x10-15
+ 123,348 1.4x10-7
ARL15 rs6450176 5 53.3 known HDL A/G - 187,132 6.9x10-10
- 123,861 7.7x10-5
BRAP rs11065987 12 112.07 known TC A/G + 187,309 2.1x10-16
+ 123,855 1.2x10-4
HMGCR rs12916 5 74.66 known TC T/C - 182,530 4.6x10-74
+ 123,863 1.5x10-4
UBASH3B rs7941030 11 122.52 known TC T/C - 187,106 2.4x10-14
- 123,819 6.6x10-4
JMJD1C rs10761731 10 65.03 known TG A/T + 91,013 8.4x10-12
+ 123,863 9.9x10-4
RBM5 rs2013208 3 50.13 novel HDL T/C + 169,708 8.9x10-12
- 123,864 1.4x10-3
ZNF664 rs4765127 12 124.46 known HDL T/G + 94,198 7.8x10-10
+ 123,737 1.7x10-3
RAB3GAP1 rs7570971 2 135.84 known TC A/C + 184,956 1.2x10-13
- 123,850 3.3x10-3
HPR rs2000999 16 72.11 known TC A/G + 185,692 6.8x10-41
+ 123,673 4.9x10-3
PDE3A rs7134375 12 20.47 known HDL A/C + 187,088 1.1x10-8
+ 123,830 4.9x10-3
PEPD rs731839 19 33.9 novel TG A/G - 176,161 2.7x10-9
+ 123,854 5.2x10-3
PGS1 rs4129767 17 76.4 known HDL A/G + 185,469 2.1x10-11
- 123,798 6.1x10-3
IRS1 rs2972146 2 227.1 known HDL T/G - 184,044 1.9x10-17
- 123,855 7.2x10-3
TOP1 rs6029526 20 39.67 known LDL A/T + 88,433 4.8x10-18
- 123,862 7.2x10-3
FRMD5 rs2929282 15 44.25 known TG A/T - 83,616 2.0x10-9
+ 122,284 1.1x10-2
ZBTB42-AKT1 rs4983559 14 105.28 novel HDL A/G - 183,672 9.6x10-9
+ 119,958 1.6x10-2
KCNK17 rs2758886 6 39.25 novel TC A/G + 187,266 3.0x10-8
- 123,863 1.6x10-2
LRP1 rs11613352 12 57.79 known TG T/C - 177,799 9.4x10-14
+ 123,865 1.9x10-2
EHBP1 rs2710642 2 63.15 novel LDL A/G + 172,994 6.1x10-9
+ 123,853 2.1x10-2
TRPS1 rs2293889 8 116.6 known HDL T/G - 180,102 4.3x10-17
+ 123,863 2.1x10-2
C6orf106 rs2814982 6 34.55 known TC T/C - 187,263 3.7x10-15
+ 123,848 3.1x10-2
VEGFA rs998584 6 43.76 novel TG A/C + 174,573 3.4x10-15
- 119,481 3.3x10-2
COBLL1 rs12328675 2 165.54 known HDL T/C - 187,092 2.1x10-15
- 123,856 3.4x10-2
CTF1 rs11649653 16 30.92 known TG C/G + 89,449 1.6x10-7
+ 123,819 3.6x10-2
PDXDC1 rs3198697 16 15.13 novel TG T/C - 175,934 2.2x10-8
+ 123,669 3.7x10-2
SETD2 rs2290547 3 47.06 novel HDL A/G - 187,142 3.7x10-9
- 118,647 3.8x10-2
UBE2L3 rs181362 22 21.93 known HDL T/C - 178,283 4.3x10-18
+ 123,910 4.7x10-2
LRP4 rs3136441 11 46.74 known HDL T/C - 186,975 6.8x10-29
+ 123,866 4.7x10-2
FLJ36070 rs492602 19 49.21 known TC A/G - 184,180 1.1x10-16
+ 120,451 4.9x10-2
*Speliotes EK et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet. 2010;42, 937-948
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
41
SUPPLEMENTARY TABLE 12B. Novel and Known Lipid Loci with CAD P-value < 0.05 from CARDIOGRAM+C4D Meta-analysis*
Locus SNP Chr hg19 Pos (Mb) Type Trait A1/A2
Lipid Direction Lipid N
Lipid P-value
CAD Direction CAD N
CAD P-value
APOA1 rs964184 11 116.65 known TG C/G - 90,991 6.6x10-224
- 110,492 4.8x10-11
SORT1 rs629301 1 109.82 known LDL T/G + 142,643 5.4x10-241
+ 82,222 6.1x10-10
BRAP rs11065987 12 112.07 known TC A/G + 187,308 2.1x10-16
- 73,578 2.4x10-9
LDLR rs6511720 19 11.2 known LDL T/G - 170,607 3.9x10-262
- 86,870 1.2x10-7
ABCG5/8 rs4299376 2 44.07 known LDL T/G - 144,861 3.9x10-72
- 106,016 3.7x10-7
HNF1A rs1169288 12 121.42 known TC A/C - 175,774 3.9x10-17
- 80,633 3.5x10-6
NAT2 rs1495741 8 18.27 known TG A/G - 87,977 2.7x10-12
- 109,804 1.2x10-5
TRIB1 rs2954029 8 126.49 known TG A/T + 177,729 1.0x10-107
+ 81,977 2.8x10-5
LPL rs12678919 8 19.84 known TG A/G + 177,749 1.8x10-199
+ 111,065 4.7x10-5
RBM5 rs2013208 3 50.13 novel HDL T/C + 169,708 8.9x10-12
- 82,470 7.0x10-5
LPA rs1564348 6 160.58 known LDL T/C - 172,988 2.8x10-21
- 108,431 1.8x10-4
APOE rs4420638 19 45.42 known LDL A/G - 93,103 1.5x10-178
- 36,066 2.1x10-4
CILP2 rs10401969 19 19.41 known TC T/C + 185,666 4.1x10-77
+ 81,644 2.4x10-4
IRS1 rs2972146 2 227.1 known HDL T/G - 184,044 1.9x10-17
+ 82,540 3.8x10-4
CMTM6 rs7640978 3 32.53 novel LDL T/C - 172,227 9.8x10-9
- 81,843 4.1x10-4
C6orf106 rs2814982 6 34.55 known TC T/C - 187,262 3.7x10-15
+ 99,096 1.6x10-3
ACAD1 rs17404153 3 132.16 novel LDL T/G - 172,898 1.8x10-9
- 83,225 1.8x10-3
CETP rs3764261 16 56.99 known HDL A/C + 177,533 1.4x10-769
- 83,626 2.2x10-3
FRMD5 rs2929282 15 44.25 known TG A/T - 83,616 2.0x10-9
- 81,446 2.8x10-3
MAP3K1 rs9686661 5 55.86 known TG T/C + 177,050 2.5x10-16
+ 81,234 3.2x10-3
KLF14 rs4731702 7 130.43 known HDL T/C + 187,085 4.8x10-17
- 99,195 3.2x10-3
ZNF664 rs4765127 12 124.46 known HDL T/G + 94,198 7.8x10-10
- 83,532 3.6x10-3
SPTY2D1 rs10128711 11 18.63 known TC T/C - 157,199 1.1x10-11
- 80,934 3.9x10-3
CAPN3 rs2412710 15 42.68 known TG A/G + 153,909 1.7x10-11
+ 79,267 5.3x10-3
HMGCR rs12916 5 74.66 known TC T/C - 182,529 4.6x10-74
- 81,050 5.3x10-3
CYP26A1 rs2068888 10 94.84 known TG A/G - 177,712 1.7x10-11
- 83,627 7.2x10-3
ST3GAL4 rs11220462 11 126.24 known LDL A/G + 145,030 6.6x10-21
+ 109,031 7.4x10-3
VEGFA rs998584 6 43.76 novel TG A/C + 174,573 3.4x10-15
+ 66,823 9.0x10-3
PCSK9 rs2479409 1 55.5 known LDL A/G - 172,970 2.5x10-50
- 83,207 1.1x10-2
PINX1 rs11776767 8 10.68 known TG C/G + 177,360 2.9x10-11
- 81,760 1.2x10-2
CITED2 rs605066 6 139.83 known HDL T/C + 94,311 2.8x10-8
- 81,709 1.5x10-2
ABCA8 rs4148008 17 66.88 known HDL C/G + 165,732 1.1x10-12
- 96,645 2.0x10-2
HBS1L rs9376090 6 135.41 novel TC T/C + 187,263 2.6x10-9
+ 81,664 2.1x10-2
APOB rs1367117 2 21.26 known LDL A/G + 173,007 9.5x10-183
+ 79,823 2.3x10-2
IKZF1 rs4917014 7 50.31 novel HDL T/G - 186,868 1.0x10-8
+ 111,434 3.4x10-2
KAT5 rs12801636 11 65.39 novel HDL A/G + 187,099 3.2x10-8
- 74,817 3.8x10-2
HPR rs2000999 16 72.11 known TC A/G + 185,692 6.8x10-41
+ 97,651 4.1x10-2
GALNT2 rs4846914 1 230.3 known HDL A/G + 186,995 3.5x10-41
- 84,068 4.1x10-2
ASAP3 rs1077514 1 23.77 novel TC T/C + 184,079 6.4x10-9
+ 84,078 4.4x10-2
KLHL8 rs442177 4 88.03 known TG T/G + 177,798 1.3x10-18
+ 82,034 4.6x10-2
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
42
Supplementary Table 12B: *Schunkert H et al. Large-scale association analysis identifies 13 new susceptibility loci for coronary artery disease. Nat Genet. 2011;43(4):333-8 *Coronary Artery Disease (C4D) Genetics Consortium. A genome-wide association study in Europeans and South Asians identifies five new loci for coronary artery disease. Nat Genet. 2011; 43(4):339-44
SUPPLEMENTARY TABLE12C. Novel and Known Lipid Loci with DBP P-value < 0.05 from ICBP*
Locus SNP Chr hg19 Pos (Mb) Type Trait A1/A2
Lipid Direction Lipid N Lipid P-value
DBP Direction DBP N DBP P-value
BRAP rs11065987 12 112.07 known TC A/G + 187,309 2.1x10-16
- 62,481 3.4x10-12
SLC39A8 rs13107325 4 103.19 known HDL T/C - 179,316 1.1x10-15
- 58,926 7.5x10-7
VEGFA rs998584 6 43.76 novel TG A/C + 174,573 3.4x10-15
+ 49,589 1.6x10-4
HFE rs1800562 6 26.09 known LDL A/G - 171,209 8.3x10-14
+ 65,399 3.2x10-4
CITED2 rs605066 6 139.83 known HDL T/C + 94,311 2.8x10-8
- 68,145 9.9x10-4
LACTB rs2652834 15 63.4 known HDL A/G - 185,613 3.6x10-11
+ 61,977 1.4x10-3
PABPC4 rs4660293 1 40.03 known HDL A/G + 187,027 2.9x10-18
- 69,815 1.8x10-3
LOC55908 rs737337 19 11.35 known HDL T/C + 185,432 4.6x10-17
- 61,569 2.1x10-3
PDE3A rs7134375 12 20.47 known HDL A/C + 187,088 1.1x10-8
- 63,231 2.3x10-3
FAM13A rs3822072 4 89.74 novel HDL A/G - 187,115 4.1x10-12
+ 66,600 2.8x10-3
FADS1-2-3 rs174546 11 61.57 known TG T/C + 177,785 7.4x10-38
+ 69,718 6.9x10-3
RSPO3 rs1936800 6 127.44 novel HDL T/C - 187,111 3.1x10-10
- 67,494 7.3x10-3
JMJD1C rs10761731 10 65.03 known TG A/T + 91,013 8.4x10-12
+ 68,336 8.3x10-3
PINX1 rs11776767 8 10.68 known TG C/G + 177,360 2.9x10-11
- 68,201 1.2x10-2
TOM1 rs138777 22 35.71 novel TC A/G + 185,274 4.7x10-8
+ 67,303 1.4x10-2
KAT5 rs12801636 11 65.39 novel HDL A/G + 187,099 3.2x10-8
- 62,171 1.7x10-2
KCNK17 rs2758886 6 39.25 novel TC A/G + 187,266 3.0x10-8
+ 69,242 1.9x10-2
ABCA1 rs1883025 9 107.66 known HDL T/C - 186,365 1.5x10-65
+ 61,161 1.9x10-2
FTO rs1121980 16 53.81 novel HDL A/G - 185,524 6.8x10-9
- 67,121 2.7x10-2
MAFB rs2902940 20 39.09 known TC A/G + 185,716 8.8x10-10
+ 67,497 2.7x10-2
SBNO1 rs4759375 12 123.8 known HDL T/C + 94,311 3.0x10-8
+ 62,022 2.9x10-2
APOE rs4420638 19 45.42 known LDL A/G - 93,103 1.5x10-178
+ 43,118 3.2x10-2
ARL15 rs6450176 5 53.3 known HDL A/G - 187,132 6.9x10-10
+ 65,297 3.3x10-2
KLF14 rs4731702 7 130.43 known HDL T/C + 187,085 4.8x10-17
- 68,636 3.5x10-2
PEPD rs731839 19 33.9 novel TG A/G - 176,161 2.7x10-9
- 62,641 3.9x10-2
CYP26A1 rs2068888 10 94.84 known TG A/G - 177,712 1.7x10-11
- 56,303 3.9x10-2
MTMR3 rs5763662 22 30.38 novel LDL T/C + 162,777 1.2x10-8
+ 58,243 4.4x10-2
TYW1B rs13238203 7 72.13 known TG T/C - 101,951 3.1x10-6
- 34,202 4.6x10-2
CMIP rs2925979 16 81.53 known HDL T/C - 185,553 1.3x10-19
+ 65,526 4.7x10-2
*International Consortium for Blood Pressure Genome-Wide Association Studies, Ehret GB et al. Genetic variants in novel pathways
influence blood pressure and cardiovascular disease risk. Nature. 2011;478(7367):103-9
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
43
SUPPLEMENTARY TABLE12D. Novel and Known Lipid Loci with SBP P-value < 0.05 from ICBP*
Locus SNP Chr hg19 Pos (Mb) Type Trait A1/A2
Lipid Direction Lipid N Lipid P-value
SBP Direction SBP N SBP P-value
BRAP rs11065987 12 112.07 known TC A/G + 187,309 2.1x10-16
- 62,444 2.1x10-8
SLC39A8 rs13107325 4 103.19 known HDL T/C - 179,316 1.1x10-15
- 58,910 2.6x10-7
VEGFA rs998584 6 43.76 novel TG A/C + 174,573 3.4x10-15
+ 49,585 9.3x10-5
CITED2 rs605066 6 139.83 known HDL T/C + 94,311 2.8x10-8
- 68,154 1.1x10-3
LACTB rs2652834 15 63.4 known HDL A/G - 185,613 3.6x10-11
+ 61,931 2.6x10-3
KCNK17 rs2758886 6 39.25 novel TC A/G + 187,266 3.0x10-8
+ 69,249 2.9x10-3
PABPC4 rs4660293 1 40.03 known HDL A/G + 187,027 2.9x10-18
- 69,821 3.0x10-3
KAT5 rs12801636 11 65.39 novel HDL A/G + 187,099 3.2x10-8
- 62,173 3.7x10-3
KLF14 rs4731702 7 130.43 known HDL T/C + 187,085 4.8x10-17
- 68,646 7.0x10-3
MTMR3 rs5763662 22 30.38 novel LDL T/C + 162,777 1.2x10-8
+ 58,275 1.8x10-2
DAGLB rs702485 7 6.45 novel HDL A/G - 186,974 6.5x10-12
+ 67,622 2.2x10-2
RSPO3 rs1936800 6 127.44 novel HDL T/C - 187,111 3.1x10-10
- 67,485 2.5x10-2
PLEC1 rs11136341 8 145.04 known LDL A/G - 82,810 7.1x10-12
+ 45,602 2.7x10-2
TOM1 rs138777 22 35.71 novel TC A/G + 185,274 4.7x10-8
+ 67,285 3.0x10-2
CSNK1G3 rs4530754 5 122.86 novel LDL A/G + 173,003 3.6x10-12
- 69,174 3.3x10-2
HFE rs1800562 6 26.09 known LDL A/G - 171,209 8.3x10-14
+ 65,402 3.3x10-2
MVK rs7134594 12 110 known HDL T/C + 94,311 1.8x10-13
+ 69,719 3.9x10-2
PEPD rs731839 19 33.9 novel TG A/G - 176,161 2.7x10-9
- 62,643 4.1x10-2
LOC55908 rs737337 19 11.35 known HDL T/C + 185,432 4.6x10-17
- 61,587 4.1x10-2
PDE3A rs7134375 12 20.47 known HDL A/C + 187,088 1.1x10-8
- 63,215 4.3x10-2
*International Consortium for Blood Pressure Genome-Wide Association Studies, Ehret GB et al. Genetic variants in novel pathways
influence blood pressure and cardiovascular disease risk. Nature. 2011;478(7367):103-9
Nature Genetics: doi:10.1038/ng.2797
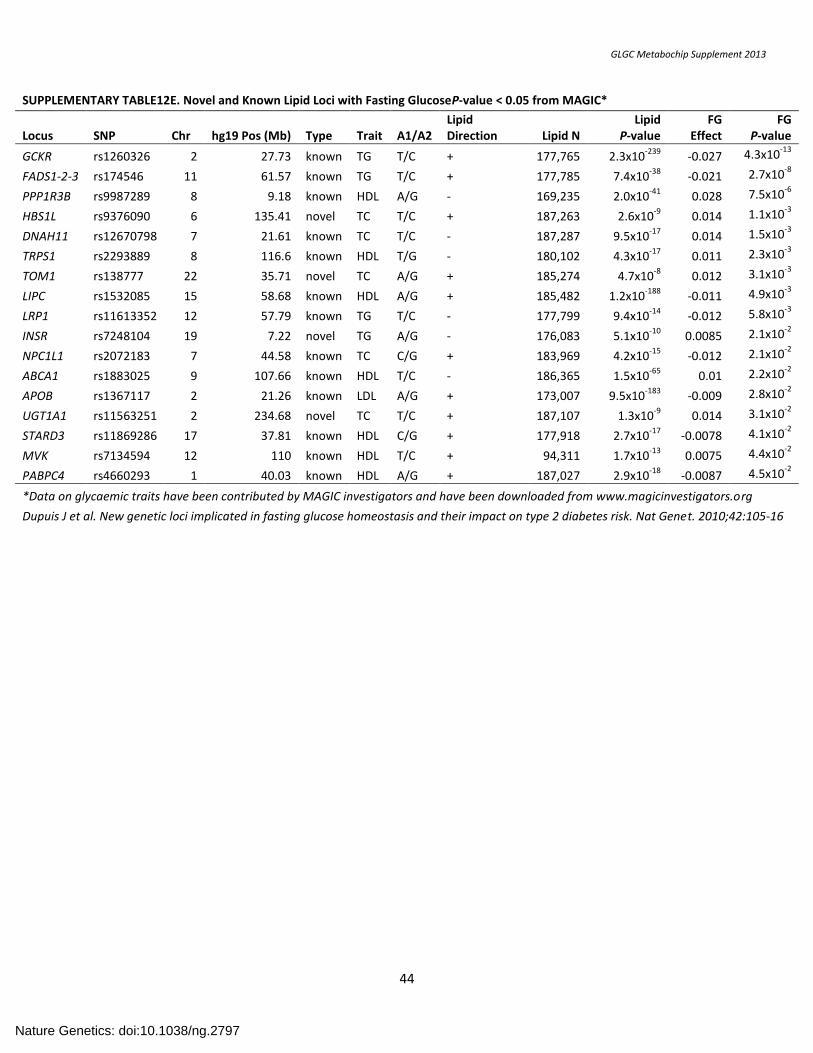
GLGC Metabochip Supplement 2013
44
SUPPLEMENTARY TABLE12E. Novel and Known Lipid Loci with Fasting GlucoseP-value < 0.05 from MAGIC*
Locus SNP Chr hg19 Pos (Mb) Type Trait A1/A2 Lipid Direction Lipid N
Lipid P-value
FG Effect
FG P-value
GCKR rs1260326 2 27.73 known TG T/C + 177,765 2.3x10-239
-0.027 4.3x10-13
FADS1-2-3 rs174546 11 61.57 known TG T/C + 177,785 7.4x10-38
-0.021 2.7x10-8
PPP1R3B rs9987289 8 9.18 known HDL A/G - 169,235 2.0x10-41
0.028 7.5x10-6
HBS1L rs9376090 6 135.41 novel TC T/C + 187,263 2.6x10-9
0.014 1.1x10-3
DNAH11 rs12670798 7 21.61 known TC T/C - 187,287 9.5x10-17
0.014 1.5x10-3
TRPS1 rs2293889 8 116.6 known HDL T/G - 180,102 4.3x10-17
0.011 2.3x10-3
TOM1 rs138777 22 35.71 novel TC A/G + 185,274 4.7x10-8
0.012 3.1x10-3
LIPC rs1532085 15 58.68 known HDL A/G + 185,482 1.2x10-188
-0.011 4.9x10-3
LRP1 rs11613352 12 57.79 known TG T/C - 177,799 9.4x10-14
-0.012 5.8x10-3
INSR rs7248104 19 7.22 novel TG A/G - 176,083 5.1x10-10
0.0085 2.1x10-2
NPC1L1 rs2072183 7 44.58 known TC C/G + 183,969 4.2x10-15
-0.012 2.1x10-2
ABCA1 rs1883025 9 107.66 known HDL T/C - 186,365 1.5x10-65
0.01 2.2x10-2
APOB rs1367117 2 21.26 known LDL A/G + 173,007 9.5x10-183
-0.009 2.8x10-2
UGT1A1 rs11563251 2 234.68 novel TC T/C + 187,107 1.3x10-9
0.014 3.1x10-2
STARD3 rs11869286 17 37.81 known HDL C/G + 177,918 2.7x10-17
-0.0078 4.1x10-2
MVK rs7134594 12 110 known HDL T/C + 94,311 1.7x10-13
0.0075 4.4x10-2
PABPC4 rs4660293 1 40.03 known HDL A/G + 187,027 2.9x10-18
-0.0087 4.5x10-2
*Data on glycaemic traits have been contributed by MAGIC investigators and have been downloaded from www.magicinvestigators.org
Dupuis J et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat Genet. 2010;42:105-16
Nature Genetics: doi:10.1038/ng.2797
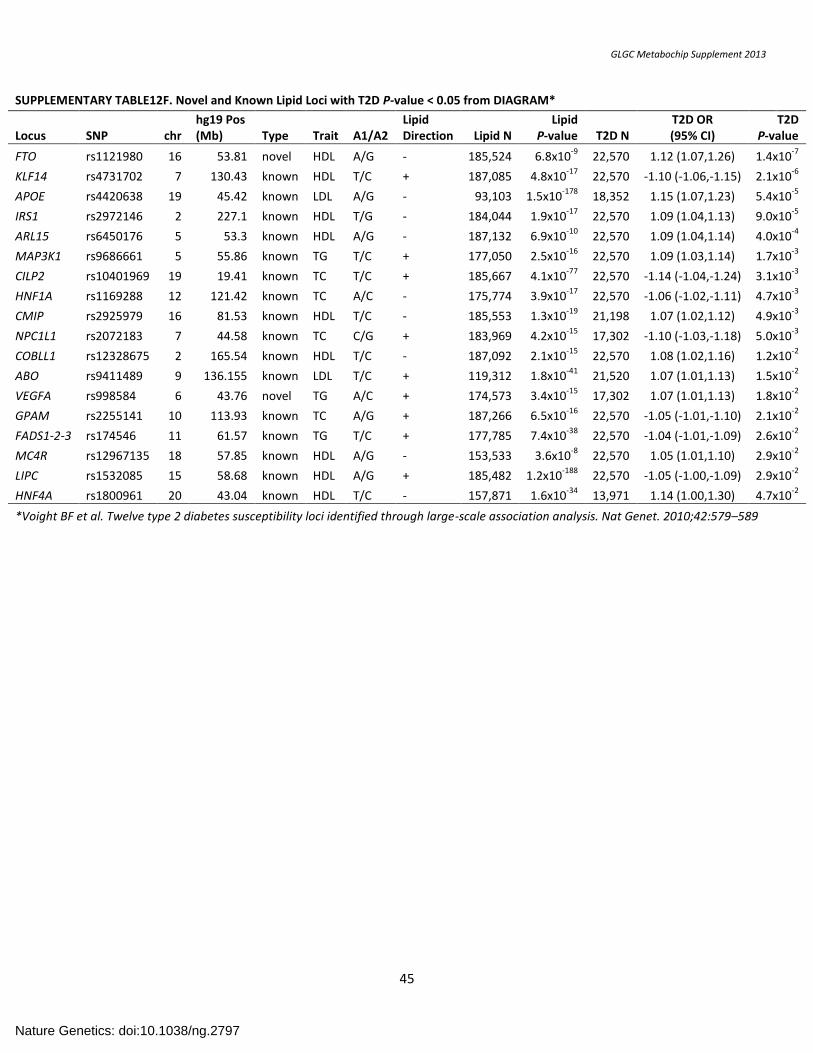
GLGC Metabochip Supplement 2013
45
SUPPLEMENTARY TABLE12F. Novel and Known Lipid Loci with T2D P-value < 0.05 from DIAGRAM*
Locus SNP chr hg19 Pos (Mb) Type Trait A1/A2
Lipid Direction Lipid N
Lipid P-value T2D N
T2D OR (95% CI)
T2D P-value
FTO rs1121980 16 53.81 novel HDL A/G - 185,524 6.8x10-9
22,570 1.12 (1.07,1.26) 1.4x10-7
KLF14 rs4731702 7 130.43 known HDL T/C + 187,085 4.8x10-17
22,570 -1.10 (-1.06,-1.15) 2.1x10-6
APOE rs4420638 19 45.42 known LDL A/G - 93,103 1.5x10-178
18,352 1.15 (1.07,1.23) 5.4x10-5
IRS1 rs2972146 2 227.1 known HDL T/G - 184,044 1.9x10-17
22,570 1.09 (1.04,1.13) 9.0x10-5
ARL15 rs6450176 5 53.3 known HDL A/G - 187,132 6.9x10-10
22,570 1.09 (1.04,1.14) 4.0x10-4
MAP3K1 rs9686661 5 55.86 known TG T/C + 177,050 2.5x10-16
22,570 1.09 (1.03,1.14) 1.7x10-3
CILP2 rs10401969 19 19.41 known TC T/C + 185,667 4.1x10-77
22,570 -1.14 (-1.04,-1.24) 3.1x10-3
HNF1A rs1169288 12 121.42 known TC A/C - 175,774 3.9x10-17
22,570 -1.06 (-1.02,-1.11) 4.7x10-3
CMIP rs2925979 16 81.53 known HDL T/C - 185,553 1.3x10-19
21,198 1.07 (1.02,1.12) 4.9x10-3
NPC1L1 rs2072183 7 44.58 known TC C/G + 183,969 4.2x10-15
17,302 -1.10 (-1.03,-1.18) 5.0x10-3
COBLL1 rs12328675 2 165.54 known HDL T/C - 187,092 2.1x10-15
22,570 1.08 (1.02,1.16) 1.2x10-2
ABO rs9411489 9 136.155 known LDL T/C + 119,312 1.8x10-41
21,520 1.07 (1.01,1.13) 1.5x10-2
VEGFA rs998584 6 43.76 novel TG A/C + 174,573 3.4x10-15
17,302 1.07 (1.01,1.13) 1.8x10-2
GPAM rs2255141 10 113.93 known TC A/G + 187,266 6.5x10-16
22,570 -1.05 (-1.01,-1.10) 2.1x10-2
FADS1-2-3 rs174546 11 61.57 known TG T/C + 177,785 7.4x10-38
22,570 -1.04 (-1.01,-1.09) 2.6x10-2
MC4R rs12967135 18 57.85 known HDL A/G - 153,533 3.6x10-8
22,570 1.05 (1.01,1.10) 2.9x10-2
LIPC rs1532085 15 58.68 known HDL A/G + 185,482 1.2x10-188
22,570 -1.05 (-1.00,-1.09) 2.9x10-2
HNF4A rs1800961 20 43.04 known HDL T/C - 157,871 1.6x10-34
13,971 1.14 (1.00,1.30) 4.7x10-2
*Voight BF et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat Genet. 2010;42:579–589
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
46
SUPPLEMENTARY TABLE12G. Novel and Known Lipid Loci with WHR adj BMI P-value < 0.05 from GIANT*
Locus SNP Chr hg19 Pos (Mb) Type Trait A1/A2
Lipid Direction Lipid N
Lipid P-value
WHR Direction WHR N
WHR P-value
RSPO3 rs1936800 6 127.44 novel HDL T/C - 187,111 3.1x10-10
+ 77,164 5.0x10-14
VEGFA rs998584 6 43.76 novel TG A/C + 174,573 3.4x10-15
+ 72,804 3.2x10-9
ZNF664 rs4765127 12 124.46 known HDL T/G + 94,198 7.8x10-10
- 77,048 1.8x10-5
COBLL1 rs12328675 2 165.54 known HDL T/C - 187,092 2.1x10-15
+ 77,160 2.8x10-5
C4orf52 rs10019888 4 26.06 novel HDL A/G + 187,077 4.9x10-8
- 77,165 5.1x10-5
MAP3K1 rs9686661 5 55.86 known TG T/C + 177,050 2.5x10-16
+ 77,164 8.0x10-5
CITED2 rs605066 6 139.83 known HDL T/C + 94,311 2.8x10-8
- 77,164 1.3x10-4
TOM1 rs138777 22 35.71 novel TC A/G + 185,274 4.7x10-8
+ 77,218 2.8x10-4
GCKR rs1260326 2 27.73 known TG T/C + 177,765 2.3x10-239
+ 77,128 3.4x10-4
FAM13A rs3822072 4 89.74 novel HDL A/G - 187,115 4.1x10-12
+ 77,163 3.5x10-4
FN1 rs1250229 2 216.3 novel LDL T/C - 173,032 3.1x10-8
+ 77,155 6.6x10-4
APOE rs4420638 19 45.42 known LDL A/G - 93,103 1.5x10-178
+ 69,832 8.5x10-4
STAB1 rs13326165 3 52.53 novel HDL A/G + 187,134 9.0x10-11
- 77,168 1.0x10-3
CILP2 rs10401969 19 19.41 known TC T/C + 185,666 4.1x10-77
- 77,160 2.5x10-3
ERGIC3 rs2277862 20 34.15 known TC T/C - 185,738 5.3x10-11
- 77,165 9.9x10-3
TOP1 rs6029526 20 39.67 known LDL A/T + 88,433 4.8x10-18
+ 77,165 1.0x10-2
KCNK17 rs2758886 6 39.25 novel TC A/G + 187,266 3.0x10-8
+ 77,167 1.2x10-2
CMIP rs2925979 16 81.53 known HDL T/C - 185,553 1.3x10-19
+ 77,164 1.5x10-2
ARL15 rs6450176 5 53.3 known HDL A/G - 187,131 6.9x10-10
- 77,165 1.8x10-2
ACAD1 rs17404153 3 132.16 novel LDL T/G - 172,898 1.8x10-9
- 77,166 1.9x10-2
PPP1R3B rs9987289 8 9.18 known HDL A/G - 169,234 1.9x10-41
+ 77,170 2.0x10-2
MYLIP rs3757354 6 16.13 known LDL T/C - 172,986 2.1x10-17
- 72,863 2.6x10-2
HBS1L rs9376090 6 135.41 novel TC T/C + 187,263 2.6x10-9
- 77,165 3.3x10-2
ANXA9-CERS2 rs267733 1 150.96 novel LDL A/G + 164,562 5.3x10-9
- 77,162 4.1x10-2
LRPAP1 rs6831256 4 3.47 novel TG A/G - 177,494 1.6x10-12
- 77,141 4.1x10-2
NAT2 rs1495741 8 18.27 known TG A/G - 87,977 2.7x10-12
+ 77,166 4.6x10-2
TTC39B rs581080 9 15.31 known HDL C/G + 186,937 1.0x10-19
+ 77,165 4.9x10-2
*Heid IM et al. Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nat Genet 2010;42, 949-960.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
47
SUPPLEMENTARY TABLE 13.Overlap of Lipid Subfractions in Framingham with Novel and Known Lipid Associated Loci (P<1.4x10-5
)
Locus SNP Lipid Subfraction Trait* A1/A2 N MAF Beta P-value
Novel Lipid Locus
Lipid P-value
Overlap of Lipid Subfractions with HDL Loci
LIPC rs1532085 HDL2 cholesterol subfraction A/G 2,900 0.38 0.13 2x10-06
N 1x10-188
LIPC rs1532085 HDL size A/G 2,742 0.38 0.17 4x10-09
N 1x10-188
LIPC rs1532085 Large particles of HDL A/G 2,742 0.38 0.16 6x10-08
N 1x10-188
CETP rs3764261 Intermediate density lipoprotein A/C 2,742 0.31 -0.16 9x10-08
N 1x10-769
CETP rs3764261 HDL2 cholesterol subfraction A/C 2,900 0.31 0.18 1x10-09
N 1x10-769
CETP rs3764261 LDL size A/C 2,742 0.31 0.17 7x10-08
N 1x10-769
CETP rs3764261 Large particles of LDL A/C 2,742 0.31 0.14 9x10-06
N 1x10-769
CETP rs3764261 HDL size A/C 2,742 0.31 0.19 6x10-10
N 1x10-769
CETP rs3764261 Large particles of HDL A/C 2,742 0.31 0.22 4x10-13
N 1x10-769
CETP rs3764261 HDL3 cholesterol subfraction A/C 2,900 0.31 0.23 1x10-14
N 1x10-769
CETP rs3764261 Apoliprotein AI concentration A/C 2,885 0.31 0.19 4x10-10
N 1x10-769
LIPG rs7241918 Apoliprotein AI concentration G/T 2,885 0.17 -0.19 2x10-07
N 1x10-44
PLTP rs6065906 Large particles of HDL C/T 2,742 0.18 -0.18 1x10-06
N 5x10-40
PLTP rs6065906 Medium particles of HDL C/T 2,742 0.18 0.35 1x10-21
N 5x10-40
Overlap of Lipid Subfractions with LDL Loci
SORT1 rs629301 Apolipoprotein B concentration G/T 2,821 0.21 -0.19 2x10-08
N 5x10-241
ApoE rs4420638 ApoE concentration G/A 2,260 0.16 -0.62 9x10-10
N 2x10-178
Overlap of Lipid Subfractions with Triglyceride Loci
GCKR rs1260326 Apolipoprotein CIII concentration T/C 2,484 0.45 0.18 2x10-10
N 2x10-239
LPL rs12678919 Apoliprotein AI concentration G/A 2,885 0.1 0.2 1x10-05
N 2x10-199
APOA1 rs964184 Medium particles of VLDL G/C 2,742 0.14 0.26 2x10-10
N 7x10-224
APOA1 rs964184 Remnant like particles expressed as triglycerides G/C 2,385 0.14 0.2 5x10-06
N 7x10-224
APOA1 rs964184 Remnant like particles expressed as cholesterol G/C 2,468 0.14 0.19 7x10-06
N 7x10-224
APOA1 rs964184 Apolipoprotein B concentration G/C 2,821 0.14 0.23 4x10-09
N 7x10-224
*LDL=low density lipoprotein, HDL=high density lipoprotein, VLDL=very low density lipoprotein
The threshold used for significance is1.4x10-5
. This corresponds to a Bonferroni correction for 23subfractions and 151SNPsfound in
the lipid subfraction dataset (0.05/(23*151)).
Nature Genetics: doi:10.1038/ng.2797
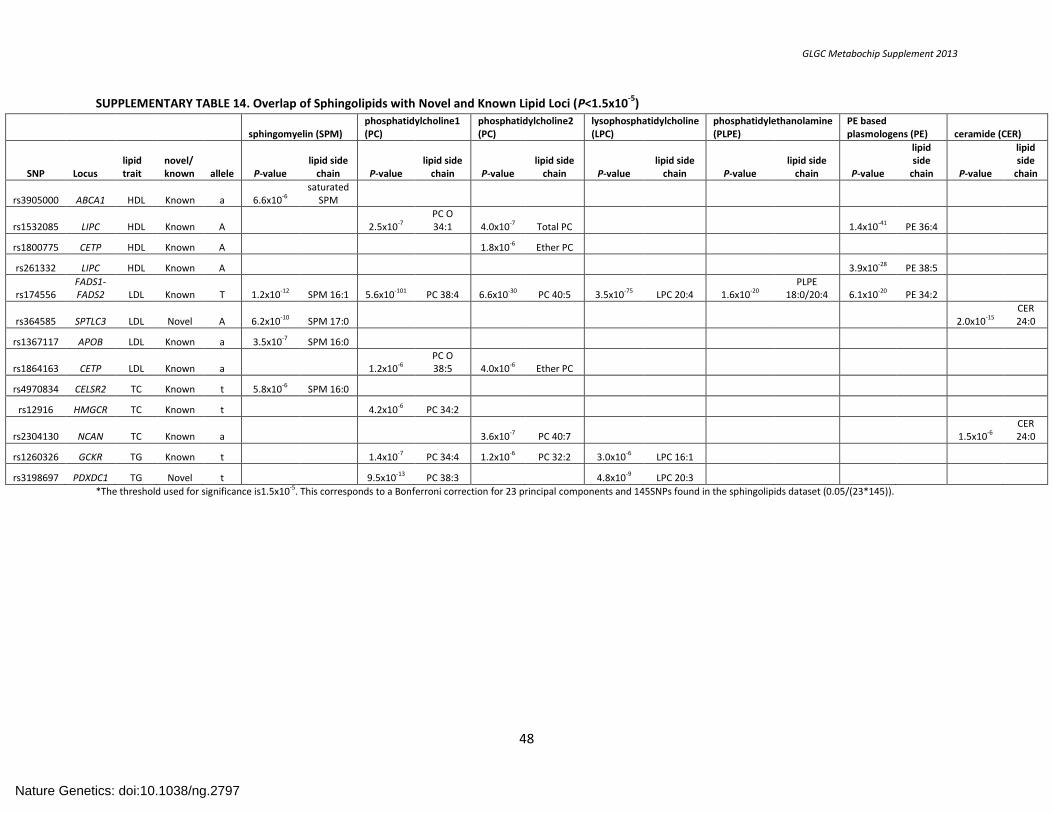
GLGC Metabochip Supplement 2013
48
SUPPLEMENTARY TABLE 14. Overlap of Sphingolipids with Novel and Known Lipid Loci (P<1.5x10-5
)
sphingomyelin (SPM) phosphatidylcholine1 (PC)
phosphatidylcholine2 (PC)
lysophosphatidylcholine (LPC)
phosphatidylethanolamine (PLPE)
PE based plasmologens (PE) ceramide (CER)
SNP Locus lipid trait
novel/ known allele P-value
lipid side chain P-value
lipid side chain P-value
lipid side chain P-value
lipid side chain P-value
lipid side chain P-value
lipid side
chain P-value
lipid side
chain
rs3905000 ABCA1 HDL Known a 6.6x10-6 saturated
SPM
rs1532085 LIPC HDL Known A 2.5x10-7 PC O 34:1 4.0x10-7 Total PC 1.4x10-41 PE 36:4
rs1800775 CETP HDL Known A 1.8x10-6 Ether PC
rs261332 LIPC HDL Known A 3.9x10-28 PE 38:5
rs174556 FADS1-FADS2 LDL Known T 1.2x10-12 SPM 16:1 5.6x10-101 PC 38:4 6.6x10-30 PC 40:5 3.5x10-75 LPC 20:4 1.6x10-20
PLPE 18:0/20:4 6.1x10-20 PE 34:2
rs364585 SPTLC3 LDL Novel A 6.2x10-10 SPM 17:0 2.0x10-15 CER 24:0
rs1367117 APOB LDL Known a 3.5x10-7 SPM 16:0
rs1864163 CETP LDL Known a 1.2x10-6 PC O 38:5 4.0x10-6 Ether PC
rs4970834 CELSR2 TC Known t 5.8x10-6 SPM 16:0
rs12916 HMGCR TC Known t 4.2x10-6 PC 34:2
rs2304130 NCAN TC Known a 3.6x10-7 PC 40:7 1.5x10-6 CER 24:0
rs1260326 GCKR TG Known t 1.4x10-7 PC 34:4 1.2x10-6 PC 32:2 3.0x10-6 LPC 16:1
rs3198697 PDXDC1 TG Novel t 9.5x10-13 PC 38:3 4.8x10-9 LPC 20:3
*The threshold used for significance is1.5x10-5. This corresponds to a Bonferroni correction for 23 principal components and 145SNPs found in the sphingolipids dataset (0.05/(23*145)).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
49
Supplementary Figures
SUPPLEMENTARY FIGURE 1. Study Design
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
50
SUPPLEMENTARY FIGURE 2. QQ Plots of Metabochip Meta-analysis P-value Distributions
Quantile-quantile plots of metabochip meta-analysis observed association –log10 p-values plotted
against expected –log10 p-values. Points in blue represent the p-value distribution after removing ± 1MB
of previously known lipid loci. There is reduced inflation of p-values after removing ± 1MB of all
genome-wide significant loci (shown in green). Lambda values for all Metabochip SNPs were between
1.19 (triglyceride levels) and 1.28 (HDL cholesterol) and reflect the enrichment of associated SNPs in the
genotyping array. After removing SNPs within 1 Megabase of previously reported associated variants,
the lambda values ranged from 1.00 (LDL cholesterol) to 1.10 (HDL cholesterol). After removing SNPs in
newly genome-wide significant loci, lambda values reached 1.00 for two traits (LDL cholesterol and
triglyceride levels) but were at 1.05 for total cholesterol and 1.07 for HDL cholesterol. The
interpretation of genomic control values from this experiment is complex because MetaboChip SNPs are
heavily concentrated on regions associated with lipids and other cardiovascular traits. The initial
genomic control values likely reflect this enrichment; the modestly high genomic control value after
excluding confirmed regions of association could reflect a combination of polygenic effects, additional
loci to discover, or population stratification.
Nature Genetics: doi:10.1038/ng.2797
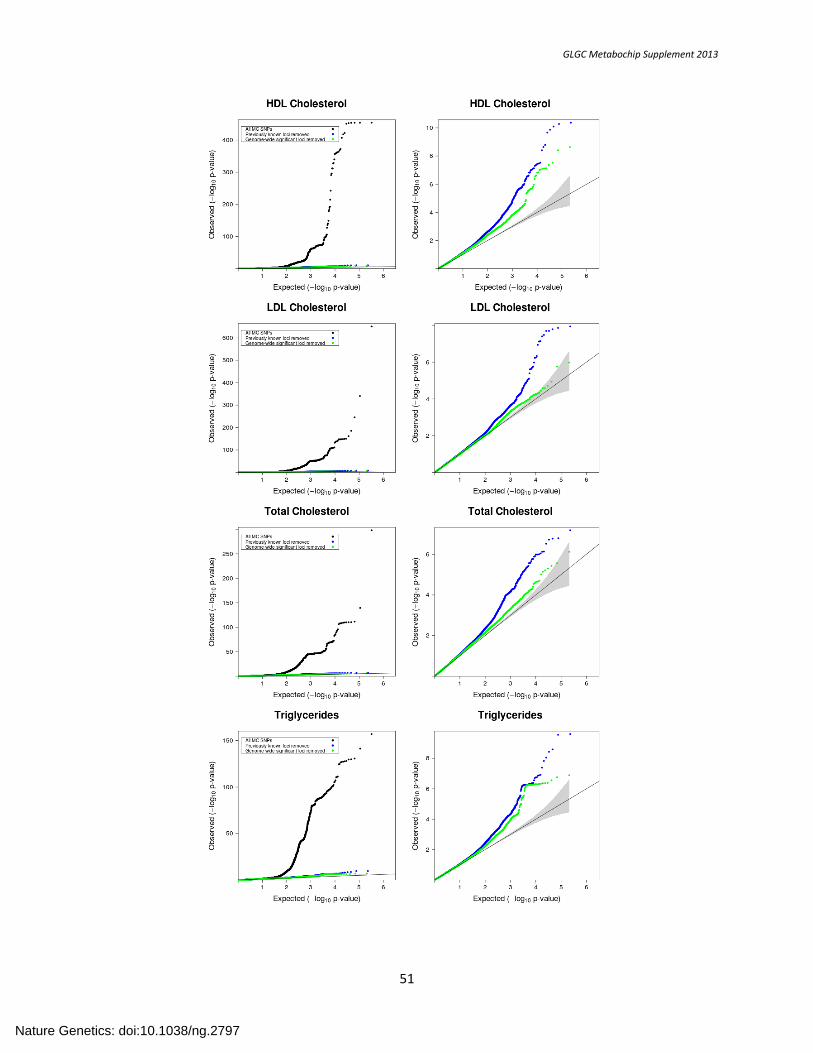
GLGC Metabochip Supplement 2013
51
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
52
SUPPLEMENTARY FIGURE 3. Manhattan Plots of Lipid-specific Association Results
Manhattan plots highlight significant SNP associations for each trait (P< 5x10-8). Trait-specific novel loci
are shown in red. Association results for known markers previously reported to be associated with lipid
traits are shown in dark blue(when primary trait is the same trait) and light blue (when primary trait is a
different lipid trait).
Nature Genetics: doi:10.1038/ng.2797
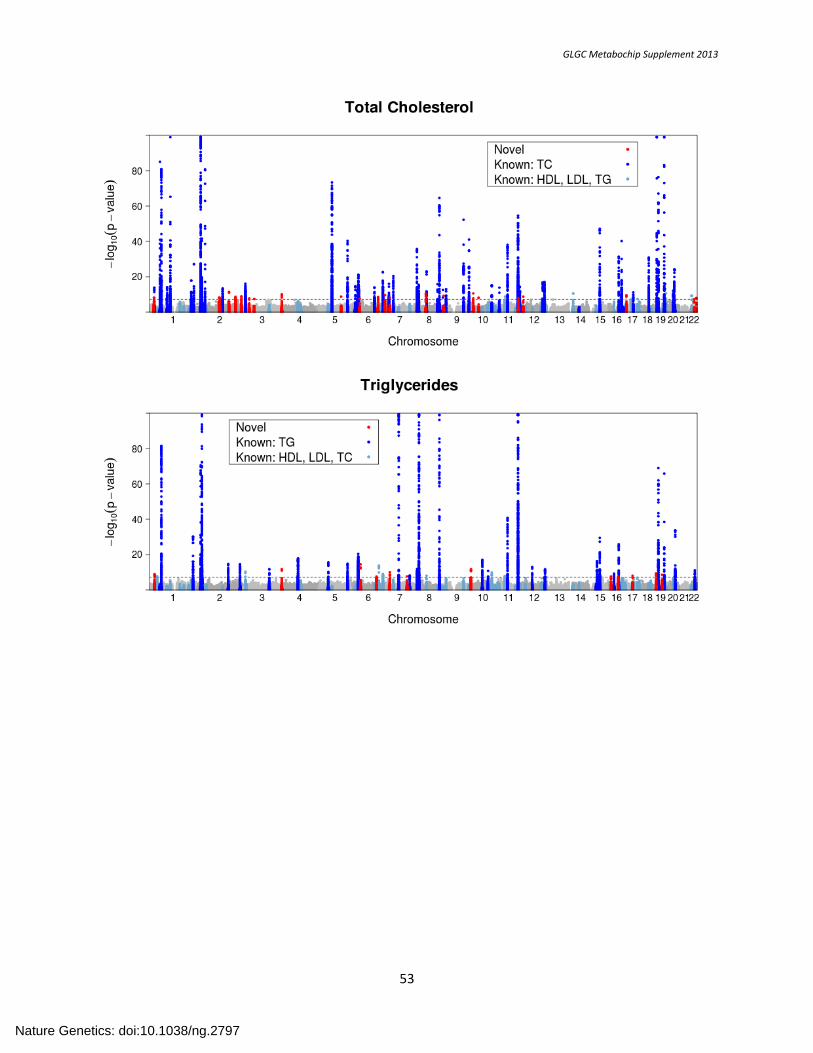
GLGC Metabochip Supplement 2013
53
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
54
SUPPLEMENTARY FIGURE 4. Effect Size vs. Allele Frequency at Lipid Associated Loci
Lipid effect sizes of SNPs in the GWAS + Metabochip meta-analysis are shown in red (novel lipid loci) in
comparison to SNPs discovered by previous GWAS efforts (shown in blue and green). Dotted lines
represent power curves for the minimum effect sizes that could be identified for a given effect-allele
frequency with 10%, 50%, and 90% power, assuming sample size 200,000 and alpha level 5x10-8.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
55
SUPPLEMENTARY FIGURE 5. Direct Protein-Protein Interactions from Dapple Analysis
Direct protein-protein interactions for A. HDL-C (8 interactions, P = .002), B. LDL-C (10 interactions, P =
.0002), C. total cholesterol (6 interactions, P = .017), and D. triglycerides (2 interactions, P = .27) show
connections between novel and known genes in the same pathways. We tested genes near previously
known and new loci.
A. HDL B. LDL
C. TC D. TG
ABCA1
PABPC4
HNF4A
GSK3B
AKT1
CETP
LPL
PLTP
SCAR3B1
APOE
APOB
LIPC
CETP
LPL
APOB
APOE
PLEC1
HNF1A
HNF4A
ABCG5
ABCG8
ZNF259
TOP1 VIM
PCSK9
APOB LDLR
TOP1
ZNF259
LIPC
APOE
LPL
VLDLR
CETP
PCSK9
PLTP
APOB
LDLR
Nature Genetics: doi:10.1038/ng.2797
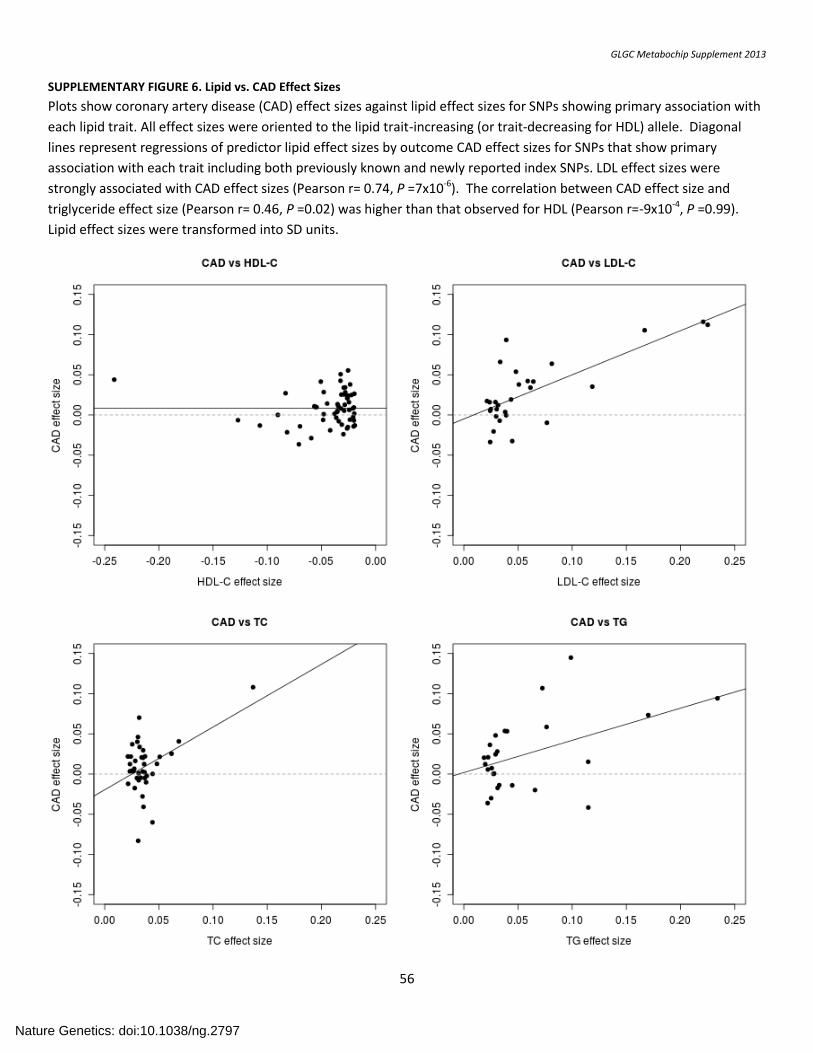
GLGC Metabochip Supplement 2013
56
SUPPLEMENTARY FIGURE 6. Lipid vs. CAD Effect Sizes
Plots show coronary artery disease (CAD) effect sizes against lipid effect sizes for SNPs showing primary association with
each lipid trait. All effect sizes were oriented to the lipid trait-increasing (or trait-decreasing for HDL) allele. Diagonal
lines represent regressions of predictor lipid effect sizes by outcome CAD effect sizes for SNPs that show primary
association with each trait including both previously known and newly reported index SNPs. LDL effect sizes were
strongly associated with CAD effect sizes (Pearson r= 0.74, P =7x10-6). The correlation between CAD effect size and
triglyceride effect size (Pearson r= 0.46, P =0.02) was higher than that observed for HDL (Pearson r=-9x10-4, P =0.99).
Lipid effect sizes were transformed into SD units.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
57
SUPPLEMENTARY FIGURE 7. Association with Lipid Subfractions
Heatmaps show effect sizes for association (P < 0.10) with 23 lipid subfractions in Framingham offspring with respect to
the trait-decreasing allele of (A) HDL-C and trait-increasing allele of (B) LDL-C, (C) TC, and (D) TG. Significant association
(P < 0.05) of lipid-associated SNPs with coronary artery disease (CAD) is annotated on the y-axis at both known and
novel genetic loci primarily associated with each trait. Dendrogram clustering of loci (y-axis) and lipid subfraction
phenotypes (x-axis) based on the effect sizes (beta) are also shown. (E) shows a heatmap of correlations for the 23 lipid
subfractions in Framingham. F-I show results from Women’s Genome Health Study1. (J) shows a heatmap of lipid
subfraction correlations in WGHS.
A.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
58
B.
*The beta for the strongest association observed, rs4420638 at the APOE locus and Lapoeser5apc (beta = -0.62), is displayed as the
minimum (-0.3) so that the color scale for the heatmap is more comparable to the heatmaps from the other 3 lipid traits.
Nature Genetics: doi:10.1038/ng.2797
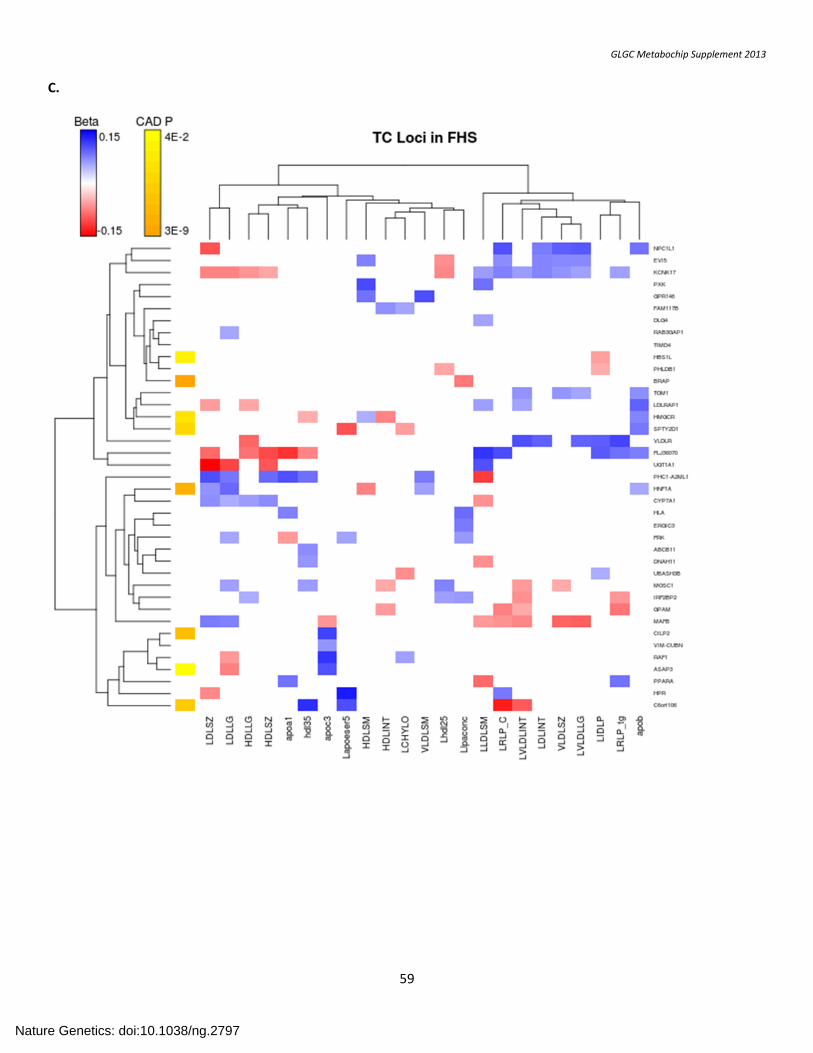
GLGC Metabochip Supplement 2013
59
C.
Nature Genetics: doi:10.1038/ng.2797
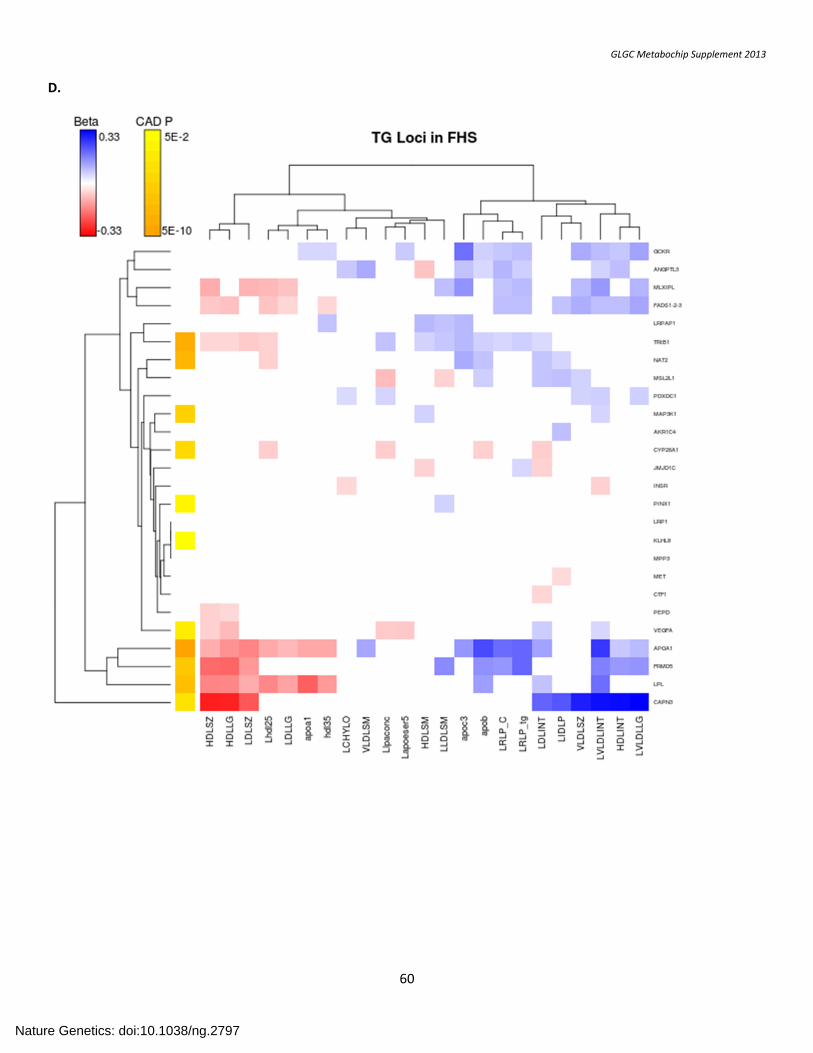
GLGC Metabochip Supplement 2013
60
D.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
61
E.
HDLLG: Large particles of high density lipoproteinconcentrations determined by NMR, Exam 4 HDLSM: Small particles of high density lipoproteinconcentrations determined by NMR, Exam 4 HDLSZ: Weighted average for HDL size based on measurements of HDLP1 through HDLP6, Exam 4 Lapoeser5
*: ApoE concentrations in mg/dL using immunochemical technique by Servia, Exam 5
LCHYLO*: Chylomicron particles size >220 nm (expressed as TG concentrations in mg/dl) and determined using NMR, Exam 4
LDLINT: Medium particles of low density lipoprotein determined by NMR, Exam 4 LDLLG: Large particles of low density lipoprotein determined by NMR, Exam 4 LDLSZ: Weighted average for LDL size based on measurements of LDLP1 through LDLP6 determined by NMR, Exam 4
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
62
Lhdl25*: HDL2 cholesterol subfractions after chemical precipitation
LIDLP*: Intermediate density lipoprotein determined by NMR, Exam 4
LLDLSM*: Small particles of low density lipoprotein determined by NMR, Exam 4
Llpaconc*:Lipoprotein(a) concentration, Exam 3
LRLP_C*: Remnant like particles measured using selective immunoseparation of lipoproteins using the Otsuka kit. Expressed as
cholesterol in mg/dL, Exam 4 LRLP_tg
*: Remnant like particles measured using selective immunoseparation of lipoproteins using the Otsuka kit. Expressed as
triglycerides in mg/dL, Exam 4 LVLDLINT
*: Medium particles of very low density lipoprotein determined by NMR, Exam 4
LVLDLLG*: Large particles of very low density lipoprotein determined by NMR, Exam 4
VLDLSM: Small particles of very low density lipoprotein determined by NMR, Exam 4 VLDLSZ: Weighted average for VLDL size based on measurements of VLDLP1 through VLDLP6 determined by NMR, Exam 4 *log transformed
All models were adjusted for age, sex and PCs. Low-, high-, intermediate- and very low-density lipoprotein particle concentrations were measured by nuclear magnetic resonance.
Nature Genetics: doi:10.1038/ng.2797
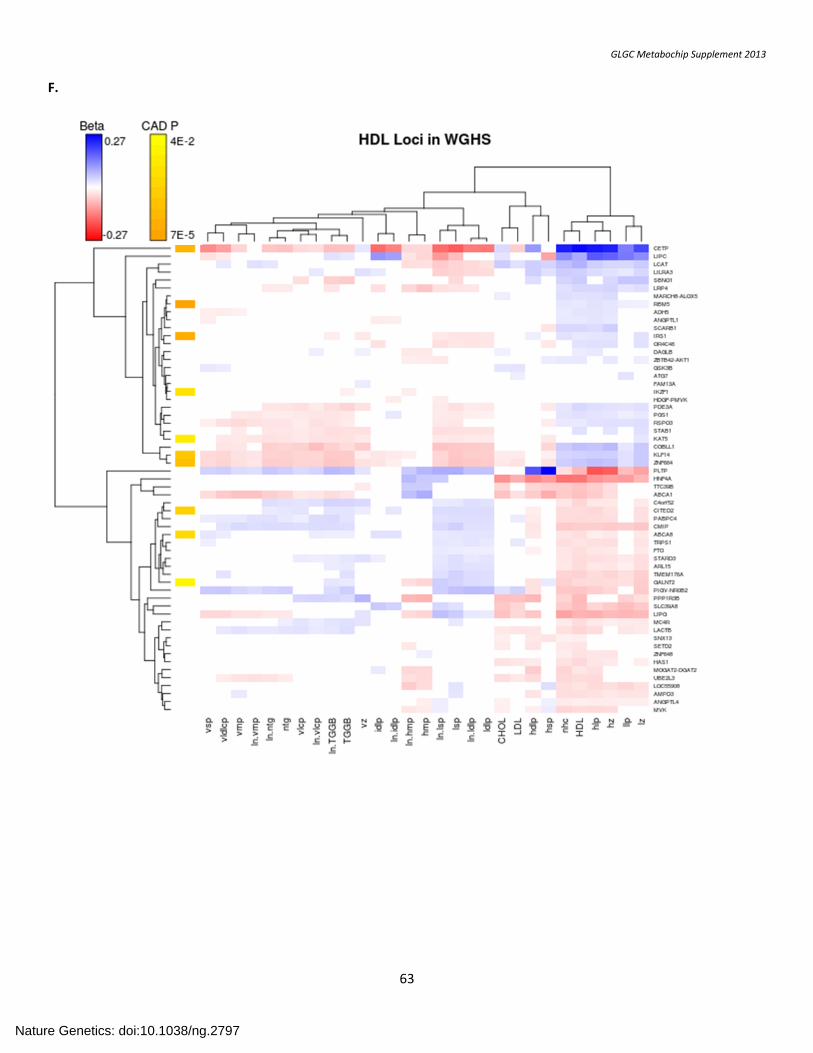
GLGC Metabochip Supplement 2013
63
F.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
64
G.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
65
H.
Nature Genetics: doi:10.1038/ng.2797
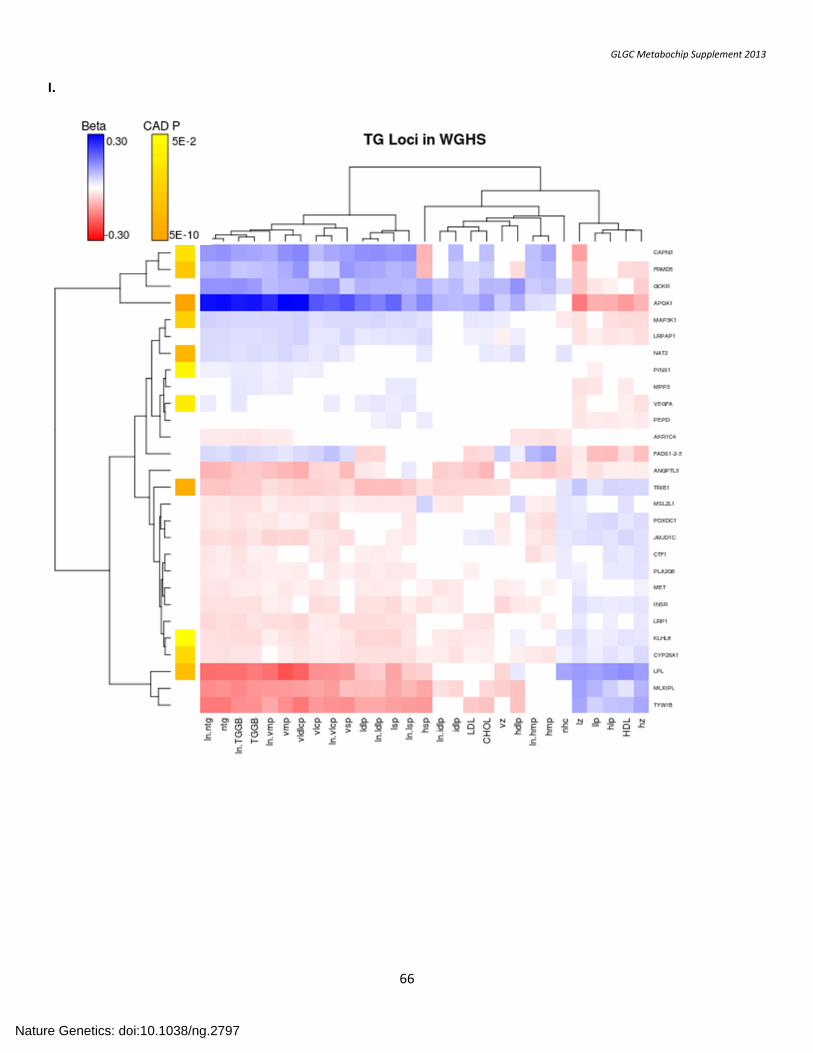
GLGC Metabochip Supplement 2013
66
I.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
67
J.
llp: LDL large hmp: HDL medium vsp: VLDL small lsp: LDL small ln.hmp: ln [HDL medium] vz: VLDL mean size ln.lsp: ln [LDL small] hsp: HDL small ntg: TG by NMR lz: LDL mean size hz: HDL mean size ln.ntg: ln [TG by NMR] idlp: IDL total nhc: HDL-C by NMR TGGB: TG assay ln.idlp: ln [IDL total] HDL: HDL-C assay ln.TGGB: ln[TG assay] ldlp: LDL total vldlcp: VLDL total CHOL: Total Cholesterol ln.ldlp: ln [LDL total] vlcp: VLDL large LDL: LDL-C assay ln.vlcp: ln [VLDL large] hdlp: HDL total vmp: VLDL medium hlp: HDL large ln.vmp: ln [VLDL medium]
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
68
Supplementary Note
Cohort Descriptions
European Metabochip Cohorts
1. ADVANCE (Atherosclerotic Disease, VAscular FuNction, & GenetiC Epidemiology study) ADVANCE is an epidemiological case-control study of genetic and non-genetic determinants of coronary artery disease (CAD) that started in 2000 as a collaborative effort between researchers at Stanford University and Kaiser Permanente of Northern California (KPNC)2. Between October 2001 and December 2003, a total of 3,179 subjects from KPNC were enrolled into three case groups and two control groups. A multi-ethnic subset of ~520 cases underwent GWAS in 2007 along with a set of controlsfrequency-matched to cases by age and sex. All remaining older onset cases (n = 974) and older controls (n =705) of white/European ancestry underwent Metabochipgenotyping in 2010. Included in this analysis are all controls not on lipid altering therapy at the time of determination of serum lipid levels.
2. AMC-PAS (The Academic Medical Center of Amsterdam Premature Atherosclerosis Cohort) AMC-PAS is aprospective study cohort recruited from the Academic Medical Centre Amsterdam that consists of patients with Premature AtheroSclerosis (PAS), defined as symptomatic CAD before the age of 51 years, or MI, coronary revascularization, or evidence of at least 70% stenosis in a major epicardial artery3. The total sample size at time of genotyping was 1,089; only cases with sufficient good quality DNA were included.
3. AMISH
The Old Order Amish individuals included in this study were participants of several ongoing studies of cardiovascular health carried out at the University of Maryland among relatively healthy volunteers from the Old Order Amish community of Lancaster County, PA and their family members4,5. The 1,200 subjects on whom the Metabochip was genotyped were recruited between 2002 and 2006 and examined at the Amish Research Clinic in Strasburg, PA.
4. BC58 (1958 British Birth Cohort)
The 1958 British Birth Cohort was initiated as a study of perinatal mortality focusing on just over 17,000 births in England, Wales, and Scotland, during one week in 1958, with the aim of identifying social and obstetric factors linked to stillbirth and neonatal death6-8. The individuals were followed into adult life, with a biomedical survey of the cohort conducted when they were aged 44–45 years. The survey included anthropometric measurements, blood sampling for lipids, and blood pressure measurements. The sample comprises 5,639 individuals of white European ethnicity, of which 2,136 were analyzed (http://www.b58cgene.sgul.ac.uk/collection.php).
5. D2D 2007 The purpose of the study is to gather information about prevalence of diabetes and cardiovascular diseases and of the risk factors associated with these within the Finnish population9. The survey assists in the evaluation of the effects of the national type 2 diabetes prevention plan. The study sample consists of 4,500 people randomly selected from the Finnish population register between the ages of 45 and 74 years and living in one of the three hospital districts chosen for the study: South Ostrobothnia, Central Finland, and Pirkanmaa.
6. deCODE
The deCODE lipid study includes lipid measurements from Icelanders recruited through various genetic studies at deCODE, primarily cardiovascular studies10. The measurements were done between the years 1987 and 2010. For the current analysis we excluded individuals using lipid lowering drugs. Genotypes and lipidmeasurements were available for 15,612 Icelanders. The study was approved by the Icelandic Data Protection Commission and the
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
69
National Bioethics Committee. All study participants signed informed consent and donated blood samples. Personal identities were encrypted by a third party system provided by the Icelandic Data Protection Commission.
7. DIAGEN (The DIAbetes GENetic Study)
DIAGEN is a large, prospective study of diabetes genetics that began in 1997 and seeks to understand the genetic causes of diabetes development; 6,282 individuals have been studied to date11. DIAGEN recruits and phenotypes individuals at high risk of T2D; 3,611 individuals from German families with a history of T2D, obesity, or dyslipoproteinemia have been studied to date from Dresden and surrounding areas.
8. DILGOM (The Dietary, Lifestyle, and Genetic determinants of Obesity and Metabolic syndrome study)
DILGOM consists of 5,025 individuals who took part in the larger FINRISK 2007 collection12. The DILGOM sample was collected to study in greater detail the components affecting obesity and metabolic syndrome.
9. DPS (The Finnish Diabetes Prevention Study) DPS is a prospective randomized controlled trial aimed at preventing the progression from IGT to diabetes13. The original DPS was initiated in 1993. A total of 522 middle-aged, overweight subjects with IGT at baseline were randomized into either a lifestyle intervention or a standard-care control group. They were followed for occurrence of diabetes until the year 2000, when the first interim analysis of the data was carried out as originally planned. At this point, the randomized trial was prematurely terminated due to markedly lower diabetes incidence rate in the lifestyle intervention group as compared to the control group. Since the termination of the randomized phase of the DPS, the original cohorts are no longer offered different treatments. However, all participants are monitored with yearly visits for long-term development of type 2 diabetes and complications.
10. DR'S EXTRA (The Dose Responses to Exercise Training Study)
DR's EXTRA is a 4-year randomized controlled trial on the health effects of aerobic and resistance exercise training and a diet with lowsaturated fat, highunsaturatedfat, and highfiber in a population sample of middle-aged and older men and women14. The target population was a representative sample of 3,000 individuals (1,500 men, 1,500 women) who lived in the city of Kuopio in Finland and who were 55-74 years of age in 2002, when they were randomly selected from the national population register. Of these individuals, 2,062 were willing to participate and 1,479 (72%) participated in the baseline examinations in 2005-2006. 1,410 individuals were randomly allocated into one of the six study groups, each of which included about 235 persons.
11. EAS (Edinburgh Artery Study)
EASis a prospective study of 1,592 men and women whose age at baseline ranged from 55 to 74 years15. The population was selected at random, in 5-year age bands, from 10 general practices spread socioeconomically across the city of Edinburgh, Scotland. A large number of biomarkers and phenotypes, focusing predominantly on putative vascular risk factors, have been measured. To identify all deaths, participant records were flagged at the UK NHS Central Registry. To obtain details of possible non-fatal events, information was sought from general practitioners, hospitals, the Information Services Division of NHS Scotland and by annual questionnaire to the subjects themselves. All cardiovascular events and deaths were further investigated using hospital or general practitioner records. The study was approved by the Lothian Health Board Ethics Committee and informed consent was obtained from each participant.
12. EGCUT (Estonian Genome Center of University of Tartu) The Estonian cohort is from the population-based biobank of the Estonian Genome Project of University of Tartu (EGCUT)16. The project is conducted according to the Estonian Gene Research Act, and all participants have signed the broad informed consent. The current cohort size is >51,515, 18 years of age and older, which reflects closely the age distribution in the adult Estonian population. Subjects are recruited by the general practitioners (GP) and physicians in the hospitals were randomly selected from individuals visiting GP offices or hospitals. Each participant
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
70
filled out a computer-assisted personal interview during 1-2 hours at a doctor’s office, including personal data (place of birth, place(s) of living, nationality. etc.), genealogical data (three generation family history), educational and occupational history, and lifestyle data (physical activity, dietary habits, smoking, alcohol consumption, women’s health, quality of life). For the current study, 2,770 individuals were genotyped with Metabochip; 1,072 of these had lipids data available.
13. Ely
The Ely study commenced in 1990 as a prospective population-based cohort study of the etiology and pathogenesis of type 2 diabetes and related metabolic disorders. Study aims are: 1) understanding the pathogenesis of type 2 diabetes in a Caucasian population through longitudinal examination of people free of diabetes at baseline; 2) determining early markers of future risk of progression of glucose intolerance and providing the means to identify populations at high risk of diabetes; and 3) quantifying and specifying the role of key exposures in adult life, particularly diet and physical activity17.
14. EPIC-CAD and EPIC-T2D (The European Prospective Investigation into Cancer and Nutrition)
EPIC was designed to investigate the relationships between diet, nutritional status, lifestyle and environmental factors and the incidence of cancer and other chronic diseases. EPIC is a large study of diet and health having recruited 520,000 individuals18,19.
15. Fenland Fenland is a population-based study of lifestyle and health. The study will investigate the influence of diet, lifestyle, and genetic factors on the development of diabetes and obesity. The participants are from the general population in the East Cambridgeshire and Fenland area and were born between 1950 and 1975.
16. FINCAVAS (The Finnish Cardiovascular Study)
The purpose of the FINCAVAS is to construct a risk profile - using genetic, haemodynamic, and electrocardiographic (ECG) markers - of individuals at high risk of cardiovascular diseases, events, and deaths20. All patients scheduled for an exercise stress test at Tampere University Hospital and willing to participate were recruited between October 2001 and December 2007. The final number of participants is 4,567. In addition to repeated measurement of heart rate and blood pressure, digital high-resolution ECG at 500 Hz is recorded continuously during the entire exercise test, including the resting and recovery phases. About 20% of the patients are examined with coronary angiography. Genetic variations known or suspected to alter cardiovascular function or pathophysiology are analyzed to elucidate the effects and interactions of these candidate genes, exercise, and commonly used cardiovascular medications. Genotyping has been done with the Metabochip for 2,795 participants. After exclusions, both genotype and lipid data were available for 1,201 participants.
17. FRISCII (Fragmin and Fast Revascularization during Instability in Coronary Artery Disease) CAD patients were randomly assigned an early invasive or non-invasive treatment strategy with placebo-controlled long-term low-molecular-mass heparin (dalteparin) for 3 months as part of a clinical trial21. Total sample size of cases is 3,489. Healthy controls (N=500) were selected to match in age and sex to the cases. In total, 2,963 individuals have been analyzed (N cases = 2,552 N controls = 411).
18. FUSION2 FUSION2 includes subjects chosen from the following studies: Dehko 2D (D2D) 2004: a population-based study to screen individuals regarding T2D risk and to prevent T2D development; Finrisk 1987: an early round of the 5-yearly Finrisk national population-based health surveys; Finrisk 2002: a population-based survey of non-communicable diseases in >13,000 individuals aged 25-74 years living in 80 communities of Finland; Action LADA: a study of latent autoimmune diabetes in adults (LADA)22. Action LADA investigators screened individuals aged 30-69 years with recently-diagnosed diabetes and identified 373 T2D cases who agreed to participate in FUSION; Health 2000: a
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
71
population-based study of people aged ≥30 years from throughout Finland; Savitaipale Diabetes Study: a study of diabetes in the town of Savitaipale in eastern Finland.
19. GLACIER (Gene x Lifestyle interactions And Complex traits Involved in Elevated disease Risk)
GLACIER is an ongoing population-based prospective cohort study comprising 19,547 adults from the northern Swedish county of Västerbotten, nested within the Northern Sweden Health and Disease Study23. All GLACIER participants underwent detailed health and lifestyle examinations as part of the Västerbotten Intervention Programme (VIP), an ongoing population-wide project focused on type 2 diabetes, cardiovascular disease, and common cancers. Since 1985, all residents of the county aged 40, 50, and 60 years have been invited to visit their primary health care centers for clinical examinations according to a standardized protocol. Baseline examinations of GLACIER participants were undertaken from 1985 through 2004. In the present study, 5,764 individuals were included in the analysis.
20. Go-DARTs (Genetics of Diabetes and Audit Research Tayside) A high quality resource, initially funded by the Wellcome Trust and supported by Diabetes UK, has been created with successful recruitment of consented patients with type 2 diabetes and matching controls (non diabetics) throughout Tayside24.
21. HUNT (Nord-Trøndelag Health Study 2) The HUNT study is a large population-based health study in Nord-Trøndelag county, central Norway (population 127,000). HUNT 2 was carried out in 1995-1997; data were collected through questionnaires, clinical exam, and blood and urine samples. 74,000 individuals (71%) participated.
22. IMPROVE
IMPROVE is a cohort of European subjects with ≥3 classical CVD risk factors, recruited from seven centers in five countries25. Of 3,418 subjects with genotyping and detailed ultrasonic phenotyping of carotid inter-media thickness, 1,769 subjects were included in this study.
23. KORA F3 (Cooperative Research in the Region of Augsburg)
KORA is a series of independent population-based epidemiological surveys and follow-up studies of participants living in the region of Augsburg, Southern Germany26. All participants are residents of Germany. In KORA S3, 4,856 subjects have been examined. 3,184 subjects participated in a 10-year follow-up examination of S3 in 2004/05. The present study includes data of 2,816 individuals of the follow-up study KORA F3. All participants gave signed informed consent. The local ethics committee has approved the studies.
24. KORA F4 (Cooperative Research in the Region of Augsburg) KORAis a series of independent population-based epidemiological surveys and follow-up studies of participants living in the region of Augsburg, Southern Germany26. All participants are residents of Germany. In the KORA S4 study, 4,261 subjects have been examined. 3,080 subjects participated in a 10-year follow-up examination of S4 in 2006-2008. The present study includes data of 2,678 individuals of the follow-up study KORA F4. All participants gave signed informed consent. The local ethics committee has approved the studies.
25. LURIC (Ludwigshafen Risk and Cardiovascular Health Study) LURIC includes 3,316 consecutive white patients of Caucasian origin (17 to 92 years of age) hospitalized for coronary angiography between June 1997 and May 200127. Clinical indications for angiography were chest pain or non-invasive tests consistent with myocardial ischemia. To limit clinical heterogeneity, individuals suffering from acute illness other than acute coronary syndromes, chronic non-cardiac diseases and a history of malignancy within the five past years were excluded. For this study, 1,506 samples were included in the analysis.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
72
26. MDC (Malmo Diet and Cancer Study) Prospective observational population-based study with a baseline examination 1991-1996 (n = 28,098) and follow ups of mortality and morbidity by means of registries28. The main focus is on cancer and cardiovascular disease.
27. METSIM (METabolic Syndrome In Men)
The METSIM study aims to investigate the metabolic syndrome, type 2 diabetes, cardiovascular disease, and cardiovascular risk factors29. It is an ongoing study of men aged 50 to 70 years, randomly selected from the population registry of the town of Kuopio, in Eastern Finland.
28. NFBC1986 (Northern Finland Birth Cohort 1986) NFBC1986 is a longitudinal one-year birth cohort study from an unselected population. The cohort included all the mothers (N = 9,362) with children whose expected date of birth fell between July 1, 1985 and June 30, 1986 in the two northernmost provinces of Finland (Oulu and Lapland). Altogether 9,479 children were born into the cohort, 9,432 of them live-born. In the 15/16-year follow-up study, blood samples were taken from 6,400 individuals of whom 4,900 were genotyped.
29. NSHD (MRC National Survey of Health and Development)
The Medical Research Council National Survey of Health and Development (NSHD), MRC 1946 Study30. This is an ongoing prospective birth cohort study consisting of all births in England, Scotland, and Wales in one week in March 1946. The sample includes single, legitimate births whose fathers were in non-manual or agricultural occupations and a randomly selected one in four of all others, whose fathers were in manual labor. The original cohort, now 62 years of age, comprised 2,547 women and 2,815 men who have been followed up >20 times since birth. The data collected to date include cognitive function, physical, lifestyle and anthropomorphic measures as well as blood analytes, and other measures. Through MRC Unit funding, the cohort is currently undergoing a particularly intensive phase of clinical assessment and biological sampling, with blood and urine sampling and analysis, cardiac and vascular imaging (http:www.nshd.mrc.ac.uk).
30. PIVUS (Prospective Investigation of the Vasculature in Uppsala Seniors)
Participants were randomly sampled from all men and women aged 70 years living in Uppsala County in 2001 (www.medsci.uu.se/PIVUS)31. Of the 2,025 individuals invited, 1,016 participated. Participants underwent a medical examination including a detailed questionnaire on lifestyle and socioeconomic factors, fasting blood sampling, blood pressure measurement and anthropometric measurements, as previously described.18 Blood and plasma samples were frozen until analysis, and blood tests performed included a wide variety of traditional and more recent CVD risk factors, along with DNA extraction. In addition, individuals have undergone extensive phenotyping including whole body MRI, echocardiography, endothelial function measurements, carotid ultrasound, DXA, and spirometry.
31. SardiNIA (SardinNIA Study on Aging) The SardiNIA study is a cohort of 6,148 participants, aged 14–102 years, from four clustered towns in Sardinia; it includes 34,469 relative pairs32. Phenotype data are available on hundreds of traits. GWAS data has been collected on ~4,700 individuals and the entire sample has been genotyped with the Metabochip.
32. SCARFSHEEP
A case-control study recruited from the Stockholm region of Sweden. Controls subjects were matched (for age and geographical location) to subjects with MI. Of a total of 3,400 subjects, 2,973 were included in this study.
33. STR (Swedish Twin Register) Four sub-studies within the Swedish Twin Register were utilized in current study, including Sex differences in health and aging (GENDER), Individual differences among the oldest-old (OCTO-Twin), and Swedish Adoption/Twin Study of Aging (SATSA)33. Similar examinations with both questionnaires and blood sampling were done in each study.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
73
Information has been updated annually on mortality and morbidity using national registries, such as the Cause of Death Registry, the National Patient Registry, and the Prescribed Drug Registry. Different biomarkers have been measured, such as markers of inflammation, lipoproteins, fatty acid composition, glucose and insulin metabolism. We required overnight fasting before blood draw except for OCTO-Twin study.
34. THISEAS (The Hellenic study of Interactions between SNPs and Eating in Atherosclerosis Susceptibility)
THISEAS is a case-control study designed to investigate the association between genetic and lifestyle environmental factors and the risk of coronary artery disease in adults34,35. Study participants were recruited from 8 hospitals and from Open Care Centers for the elderly in the region of Athens. Cases were subjects presenting with either ACS or stable CAD defined as >50% stenosis in at least one of the three main coronary vessels assessed by coronary angiography. ACS was defined as acute MI or unstable angina corresponding to class III of the Braunwald classification. ACS patients have also undergone coronary angiography examination that verified the presence of significant stenosis. Controls were subjects with negative coronary angiography findings, or negative stress test, or subjects without symptoms of disease that were admitted at the same hospitals as cases and were free of any cardiovascular disease, cancer, or inflammatory diseases. Exclusion criteria for both study groups were renal or hepatic disease. The bioethics committee of Harokopio University approved the study and all participants gave their informed consent. Hematological, biochemical, and anthropometric measurements were conducted for all participants. Dietary assessment through a semi-quantitative food frequency questionnaire and physical activity data were collected through face-to-face interview by well trained scientists.
35. TROMSO (Tromsø 4)
The Tromsø study is a prospective population-based study in the city of Tromsø in Northern Norway (population 64,000)36. There have been five study waves to date. The participation rate was >75% each time; 38,164 adults have participated at least once. For Tromsø 4 (1994), all inhabitants aged ≥25 years were invited and 27,159 (77%) participated.
36. ULSAM (Uppsala Longitudinal Study of Adult Men) ULSAM was initiated as a health investigation focused on identifying metabolic risk factors for cardiovascular disease, to which all 50-year-old men living in Uppsala County, Sweden, in 1970-74 were invited37. Of these, 82% (2,322 men) participated in the investigation. The men have been investigated at the ages of 50, 60, 71, 77, 82, and 88 years. The examination performed at age 71 (in 1991-1995), when the DNA was collected, included medical examination, questionnaire, cognitive function testing, 7-day dietary assessment, anthropometric measurements, blood sampling (after an overnight fast), blood pressure measurement, 24-hour ambulatory blood pressure measurement, electrocardiography, echocardiography, OGTT, muscle biopsy, and euglycemic insulin clamp. Outcome data have been updated annually on mortality and morbidity using national registries, such as the Cause of Death Registry, the National Patient Registry, and the Prescribed Drug Registry. Many biomarkers have been measured, such as markers of inflammation/oxidation, lipoproteins, fatty acid composition, and glucose and insulin metabolism. Insulin sensitivity was determined using the euglycemic insulin clamp technique, with a slight modification; insulin was infused at a constant rate of 56 instead of 40 mU/(min*m2) to achieve nearly complete suppression of hepatic glucose output.
37. Whitehall II
The Whitehall II Study recruited 10,308 participants (70% men) between 1985 and 1989 and involved 20 London-based civil service departments38. In this longitudinal study, clinical data were collected in phase 1 (1985-1988), phase 3 (1991-1993), phase 5 (1997-1999), and phase 7 (2003-2004). DNA was stored from phase 7 from >6,000 participants. The study individuals are all well phenotyped for cardiovascular and other ageing related health outcomes.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
74
East Asian Metabochip Cohorts
1. CLHNS (Cebu Longitudinal Health and Nutrition Survey) CLHNSis an on-going community-based study that began in 198339. The baseline survey randomly recruited 3,327 mother-child pairs from 17 urban and 16 rural areas from the Metropolitan Cebu area, the Philippines. Overnight fasting blood samples for biomarkers and DNA were obtained at the 2005 survey. In this study, 1,771 offspring samples were included in analysis.
2. TAICHI (TAIwanmetaboCHIp consortium)cohorts
The TaiChi consortium was formed through a collaborative effort between investigators based in the U.S. and Taiwan. The consortium’s primary aim is to identify genetic determinants of atherosclerosis and diabetes related traits in East Asians and to fine-map validated loci identified in other race/ethnic groups.The main academic sites in Taiwan include Taipei and Taichung Veteran’s General Hospitals (VGH), National Health Research Institute (NHRI), Tri-Service General Hospital (TSGH), and National Taiwan University Hospital (NTUH). The main U.S academic sites participating in the TaiChi consortium include Stanford University School of Medicine in Stanford, California; Hudson-Alpha Biotechnology Institute in Huntsville, Alabama; and Cedars-Sinai Medical Center (CSMC) in Los Angeles, California. There are 7 principal Taiwan based cohorts that make up the current TaiChi bio-resource with a total of 11,859 subjects in the current study. Each cohort is described in more detail in the supplementary text.
a. Taiwan DRAGON (Taiwan Diabetes and RelAted Genetic COmplicatioN) study is a cohort study with T2DM at
the Veteran’s General Hospital in Taichung, Taiwan (Taichung VGH). Participants include individuals with either
newly diagnoses or established diabetes who visit the diabetes outpatient clinic on a regular basis. Subjects with
hyperglycemia who do not meet criteria for Type 2 DM defined by IDF are not included. Individuals participate in
a health examination program at Taichung VGH are also interviewed. Specialized tests include an oral glucose
tolerance tests (OGTT) in subjects without an established diagnosis of diabetes.
b. HALST (Healthy Aging Longitudinal Study in Taiwan) is a population based multi-site cohort study of ambulatory
adults aged > 55 years living in 7 major geographic regions of Taiwan, established by the NHRI. The aim of the
study is to investigate themultidimensional determinants, including lifestyle, genetic, metabolic, and
inflammatory factors, of an older Asian population.These 7 locations include both urban and rural areas: two are
in the north (Taipei’s Shilin District and Taoyuan County’s Yangmei Township), two in central Taiwan (Miaoli City
in Miaoli County and Changhua City in Changhua County), two in the south (Puzi, Chiayi County, and Kaohsiung’s
Lingya District), and one in the east (Hualien City/County). The only exclusion criteria are presence of highly
contagious diseases, advanced illnesses with limited life span or bedridden status, dementia, other advanced
neurological deficit, severe hearing loss, and institutionalization in a chronic care facility for any reason. Over
5000 subjects are being recruited over a five-year period (2008-2012) from seven recruitment sites across the
country. Following completion of recruitment of the initial HALST cohort, follow-up in person visits will be
scheduled during a second 5-year study cycle scheduled to begin in 2013 (~1000 subjects / year). Within each
wave, participants are to be followed up by telephone contact every year for vital status and for updates on
health-related conditions. Medical records are requested to confirm the development of any new health
conditions. Vital status, health claims, health care utilization data are being collected for the cohort on a regular
basis by linking to the National Death Registry DatabaseandtheNational Health Insurance Database.
c. SAPPHIRe (Stanford-Asian Pacific Program in Hypertension and Insulin Resistance)is a family based study
established in 1995 with an initial goal of identifying major genetic loci underlying hypertension and insulin
resistance through linkage in East Asian populations. SAPPHIRe was also one of four networks participating the
NHLBI’s Family Blood Pressure Program (FBPP). At the outset, SAPPHIRe involved recruitment sites in the San
Francisco Bay Area, Hawaii, and Taiwan. However, a majority of the ~1,700 sibpairs in SAPPHIRe were recruited
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
75
from 3 centers in Taiwan (NTUH, Taipei VGH and Taichung VGH) with NHRI being the DCC. Sibpairs were either
highly concordant or discordant for blood pressure and a subset underwent an insulin suppression test. Many
metabolicvariables associated with blood pressure and insulin resistance were examined in the first 5-year
investigative cycle funded by the NIH (1995-2000). Further extensive phenotyping through return visits and
regular follow ups occurred between 2001 and 2008 in the Taiwanese SAPPHIRe participants which included
echocardiographic and multi-detector row CT imaging procedures. During this time, SAPPHIRe was referred to as
the HIRGs study in Taiwan.
d. TACT (TAiwan Coronary and Transcatheter intervention) cohort study enrolled patients with angina pectoris
and objective documentation of myocardial ischemia who underwent diagnostic coronary angiography and/or
revascularization any time after October 2000 at the National Taiwan University Hospital (NTUH). This cohort is
very similar to TCAGEN but was collected independently. Participants provided clinically relevant information
including use of cardiovascular related medication through a standardized questionnaire. Clinically relevant
information is also available through a comprehensive electronic medical records database that includes
information on drug use and surgical interventions. Fasting blood samples were collected before cardiac
catheterization.
e. TCAD (Taichung CAD study) includes patients with a variety of cardiovascular diseases receiving care at the
Taichung Veterans General Hospital. Specifically, individuals who were hospitalized for diagnostic and
interventional coronary angiography examinations and treatment are included in TAI CHI. Also included in TAI
CHI are subjects with a history of myocardial infarction or revascularizationof any type (percutaneous coronary
intervention or coronary artery bypass).
f. TCAGEN (Taiwan Coronary Artery Disease GENetic) study is an ongoing cohort study that has been enrolling
patients undergoing coronary angiography or percutaneous intervention at the National Taiwan University
Hospital (NTUH) in the setting of either stable angina pectoris or prior myocardial infarction. Participants are not
only from the north of Taiwan where the main NTU medical school/hospital is located, but also from Yulin
branch of NTUH, located in south/central Taiwan. The hospital uses an elaborate electronic medical record
system that provides access to clinic visit notes, diagnostic codes of clinic encounters, prescriptions, and
laboratory data in a searchable form. Fasting blood samples were collected before cardiac catheterization while
10 ml of peripheral blood was collected in the catheter lab specifically for buffy coat isolation and DNA
extraction.
g. TUDR (Taiwan USA Diabetes Retinopathy) is a cohort that enrolled subjects with T2DM receiving care at
Taiching Veteran’s General Hospital, a small number of subjects were included from Tri-Service General
Hospital(TSGH). All TUDR subjects underwent a complete fundoscopic examination to carefully document the
presence and extent of retinopathy. To date, a total of 2222 unrelated type 2 DM subjects with and without
retinopathy were ascertained and have undergone metabochip genotyping.
South Asian Metabochip Cohorts
1. AIDHS/SDS (Asian Indian Diabetic Heart Study/Sikh Diabetes Study) A total of 3,958 participants from AIDHS are from two different cohorts comprising 2,902 individuals from the Sikh Diabetes Study (SDS) and 1,056 migrant Asian Indians living in the US40,41. Of these, 1,516 subjects (800 males/ 716 females) available with genome-wide genotyping data (llumina’s 660W-Quad BeadChip) were used in the present study. These individuals were recruited through public advertisements from Northern states of India including Punjab, Haryana, and Delhi. Both men and women aged 17-90 years participated. Individuals with non-Punjabi ancestry and non-Asian Indian ancestry were not enrolled. Normoglycemic control subjects were random individuals recruited from the same Asian Indian community as the patients, and matched for ethnicity and geographic location.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
76
Diagnoses of type 2 diabetes were confirmed by reviewing medical records for symptoms, use of medication, and measuring fasting glucose levels following the guidelines of the American Diabetes Association (2004)42. A medical record indicating either (1) a fasting plasma glucose level ≥7.0 mmol/L or ≥126 mg/dL after a minimum 12h fast or (2) a 2h post-glucose level (2h oral glucose tolerance test) ≥11.1mmol/L or ≥200 mg/dL on more than one occasion, combined with symptoms of diabetes confirmed the diagnosis. Impaired fasting glucose (IFG) is defined as a fasting blood glucose level ≥100 mg/dL (5.6 mmol/L) but ≤126 mg/dL (7.0 mmol/L). Impaired glucose tolerance (IGT) is defined as a 2h OGTT > 140mg dL (7.8mmol/L) but < 200mg /dL(11.1mmol/L).Subjects with IFG or IGT were excluded from this study. The 2h OGTTs were performed following the criteria of the World Health Organizations (WHO) (75 g oral load of glucose). The selection of normoglycemic controls was based on a fasting glycemia <100.8 mg/dL or a 2h glucose <141.0 mg/dL. Serum lipids (total cholesterol, LDL cholesterol, HDL cholesterol, VLDL cholesterol, and triglycerides) were quantified using standard enzymatic methods (Roche, Basel, Switzerland).All quantitative parameters were determined by following manufacturer’s instructions using a Hitachi 902 auto-analyzer (Roche, Basel, Switzerland). All blood samples were obtained at the baseline visits. All participants signed a written informed consent for the investigations. The study was reviewed and approved by the University of Oklahoma Health Sciences Center’s Institutional Review Board, as well as the Human Subject Protection Committees at the participating hospitals and institutes in India.
3. PROMIS (The Pakistan Risk Of Myocardial Infarction Study)
PROMIS is an ongoing case-control study of acute myocardial infarction (MI) in urban Pakistan, which by mid-2009 included 5,500 MI cases and 5,500 controls43. Cases have typical ECG characteristics, a positive troponin test, and MI symptoms within the previous 24 hours. Controls are individuals frequency-matched to cases by sex and age (in 5 year bands) and identified in the same hospitals as the index cases. Controls have been recruited in the following order of priority: (i) visitors of patients attending the out-patient department; (ii) patients attending the out-patient department for routine non-cardiac complaints, or (iii) non-blood related visitors of index MI cases. A locally-piloted and validated epidemiological questionnaire has been administered to participants by medically qualified research officers that seeks >200 items of information in relation to: ethnicity (eg, personal and paternal ethnicity, spoken language, place of birth and any known consanguinity); demographic characteristics; lifestyle factors (eg, tobacco and alcohol consumption, dietary intake, and physical activity); personal and family history of cardiovascular disease; and medication usage. PROMIS has received approval by the relevant research ethics committee of each of the institutions involved in participant recruitment. Informed consent has been obtained from each participant recruited into the study, including consent to use the samples in genetic, biochemical, and other analyses.
African Metabochip Cohorts
1. FBPP (Family Blood Pressure Project GenNet and HyperGEN studies) The FBPP GenNet study recruited European-American (N=1,497) and African American (N=1,101) participants at two field centers between 1995 and 2003, based on a hypertensive proband44. Non-Hispanic white subjects were recruited from Tecumseh, Michigan, and African American subjects were recruited from Maywood, Illinois. Probands were defined as individuals aged 18-50 years with blood pressures in the upper 20th to 25th percentile of the age/gender specific blood pressure distribution. Once the proband was identified, an attempt was made to enroll all siblings and parents of the proband, irrespective of their blood pressure or hypertension treatment status. Blood pressure measurements were carried out according to standard procedures in a sitting position after a resting period. Subjects were not allowed to smoke or drink coffee before the visit. The average of two manual BP measurements was used as the phenotype. DNA was available for 1,381 European American and 848 African-American participants (www.biostat.wustl.edu/fbpp/FBPP.shtml). The Hypertension Genetic Epidemiology Network recruited two types of participants (hypertensive sibships and random samples of subjects) in European American and African American samples. Recruitment of the study participants, including the hypertensive probands, was carried out at five field centers based largely on ongoing population based studies. For European Americans, HyperGEN recruited and characterized a total of 1,142 hypertensive subjects from 480 sibships, yielding a total of 992 self-reported sib-pairs, and a random sample of 472 biologically unrelated participants. For African Americans,
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
77
HyperGEN recruited and characterized a total of 1,261 hypertensive subjects from 596 sibships yielding a total of 826 self-reported sib-pairs, and a random sample of 446 biologically unrelated African Americans (www.biostat.wustl.edu/fbpp/FBPP.shtml). Blood pressure measurements were carried out according to standard procedures in a sitting position after a resting period.
2. GXE (Kingston GXE; subset of International Collaborative Study of Hypertension in Blacks (ICSHIB)) The Kingston GXE cohort was obtained from a survey conducted inKingston, Jamaica as part of a larger project to examine gene by environment interactions in the determination of blood pressure among adults 25-74 years45. The principal criterion for eligibility was a body mass index in either the top or bottom third of BMI for the Jamaican population. Participants were identified principally from the records of the Heart Foundation of Jamaica, a non-governmental organization based in Kingston, which provides low-cost screening services (height and weight, blood pressure, glucose, cholesterol) to the general public. Other participants were identified from among participants in family studies of blood pressure at the Tropical Metabolism Research Unit (TMRU) and from among staff members at the University of the West Indies, Mona.
3. MRC/UVRI GPC (General Population Cohort (GPC) Study, Kyamulibwa, Uganda)
MRC/UVRI GPC is a population-based cohort study of >18,000 people living within the Kyamulibwa sub-county of Kalungu District in rural south-west Uganda. The cohort was established in 1989 by the Medical Research Council (MRC) Programme on AIDS in Uganda to describe trends in the prevalence and incidence of HIV infection and their determinants in the general population. Today the study also aims to estimate the prevalence and distribution of cardiometabolic risks factors and diseases in the population. This cohort has GWAS data but only the SNPs overlapping with metabochip were included.
4. SEY (Seychelles Tandem Study)
SEY recruited 494 participants, aged over 18 years, from 76 families of East African descent from the Republic of Seychelles (Indian Ocean), collected for the primary purpose of a candidate gene study of arterial hypertension46. Families were selected from a national register that includes all patients with hypertension. Families were selected if there were two or more full siblings with hypertension (defined as being on current antihypertensive treatment or having an average (6 measures) office systolic/diastolic blood pressure greater than or equal to 140/90 mm Hg). Participants were recruited from July 1999 until January 2002. Blood was collected in the morning between 7 am and noon after overnight fasting.
5. SPT (Spanishtown; subset of International Collaborative Study of Hypertension in Blacks (ICSHIB))
Participants were recruited from Spanish Town, a stable, residential urban area neighboring the capital city of Kingston, Jamaica as part of the International Collaborative Study of Hypertension in Blacks (ICSHIB)47. A stratified random sampling scheme was used to recruit adult males and females aged 25–74 years from the general population. Spanish Town was chosen because its demographic make-up was broadly representative of Jamaica as a whole.
European GWAS Cohorts
Descriptions for the European GWAS cohorts have been published previously48.
Cohorts Excluded from Meta-analysis
Metabochip data were received from PARC and MORGAM, but were excluded from the meta-analysis because of sample overlap and failure to pass quality control checks. 1. PARC (Pharmacogenomics and Risk of Cardiovascular Disease)
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
78
Samples for analysis were from the Cholesterol and Atherosclerosis Pharmacogenetics (CAP) Study49. CAP was a clinical trial designed to identify genetic influences responsible for inter-individual variation in response to simvastatin. CAP participants were 944 Caucasiansand African Americans, aged 30 and above, who received open label 40 mg simvastatin daily for 6 weeks. They were recruited on the basis of having serum total cholesterol levels of 4.14-10.36 mmol/L (160-400 mg/dL). Complete phenotypes of lipids before and after statin treatment and Metabochip data are available on 530 Caucasian CAP participants.
2. MORGAM (MOnica Risk, Genetics, Archiving and Monograph) MORGAM is a multinational collaborative study exploring the relationships between the development of cardiovascular diseases, their classic and genetic risk factors and biomarkers50.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
79
Candidate Genes at Novel Loci The list of notable genes in newly identified loci, below, is meant to provide an overview of the diverse set of loci associated with
blood lipids in our study. Although the list can provide a starting point for exploration of these loci and help motivate follow-up
studies and/or hypotheses, the list should not be considered exhaustive.
ABCB11 (ATP-binding cassette, sub-family B, member 11) is involved in the ATP-dependent secretion of bile salts (MIM 603201).
Hepatic overexpression of Abcb11 in mice increased absorption of cholesterol and promoted diet-induced obesity and
hypercholesterolemia51
. G6PC2 encodes a glucose-6-phosphatase catalytic subunit (MIM 608058). Variants at this locus have been
implicated in liver enzyme and fasting glucose levels52,53
.
ACAD11 (acyl-CoA dehydrogenase family, member 11) is involved in the β-oxidation of long-chain fatty acids in muscle and heart
(MIM 614288).
ADH5 (alcohol dehydrogenase 5 (class III), chi polypeptide) encodes a protein involved in oxidation of long-chain primary alcohols
and which catalyzes a step in the elimination of formaldehyde (MIM 103710).
AKR1C4 (aldo-keto reductase family 1, member C4) encodes a protein that produces intermediates in bile acid biosynthesis and
inactivates circulating steroid hormones (MIM 600451). AKR1C4 is expressed exclusively in the liver and is transcriptionally regulated
by LXRA.
ANGPTL1 (angiopoietin-like 1 gene) is a member of the angiopoietin family involved in angiogenesis, and widely expressed in highly
vascularized tissues (MIM 603874).
ANXA9 (annexin A9) and CERS2 (ceramide synthase 2). ANXA9 is a calcium-dependent phospholipid-binding protein (MIM 603319).
CERS2 is involved in regulation of long acyl chain and sphingolipid metabolism (MIM 606920).
APOH (Apolipoprotein H, also known as beta-2 glycoprotein I) and PRKCA (protein kinase C, alpha). APOH is a glycoprotein that is
involved in the activation of lipoprotein lipase and which neutralizes negatively charged phospholipids (MIM 138700). PRKCA is
activated by APOA1 and diacylglycerol during cholesterol mobilization (MIM 176960)54
.
ASAP3 (ArfGAP with SH3 domain, ankyrin repeat and PH domain 3) is a GTPase-activating protein that promotes cell differentiation
and migration and has been implicated in cancer cell invasion55
.
ATG7 (autophagy related 7) encodes a protein that is part of the autophagy machinery (MIM 608760). Dysfunction in autophagy
canimpact systems related to intracellular energy utilization andpromote apoptotic cell death.
BRCA2 (breast cancer 2, early onset) is involved in maintenance of genome stability, specifically the homologous recombination
pathway for repair of double stranded DNA. Variants in the region can increase risk of breast and other types of cancer (MIM
600185).
C4orf52 (chromosome 4 open reading frame 52). The nearest gene to the lead signal is an uncharacterized gene with unknown
function, and there are no other obvious candidate genes in the locus.
CMTM6 (CKLF-like MARVEL). This gene belongs to the chemokine-like factor gene superfamily, but the exact function of the
encoded protein is unknown (MIM 607889).
CPS1 (carbamoyl-phosphate synthase 1, mitochondrial) encodes a mitochondrial enzyme that catalyzes the first committed step of
the urea cycle (MIM 608307). The lead variant encodes a threonine to asparagine substitution previously associated with levels of
homocysteine and fibrinogen 56,57
.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
80
CSNK1G3 (casein kinase 1, gamma 3) encodes a serine/threonine-protein kinase that is involved in a number of cellular processes
including DNA repair, cell division, nuclear localization and membrane transport (MIM 604253).
DAGLB (diacylglycerol lipase, beta) catalyzes the hydrolysis of diacylglycerol (DAG) to 2-arachidonoyl-glycerol, an abundant
endocannabinoid (MIM 614016). Endocannabinoids function signaling molecules, regulate axonal growth, and drive adult
neurogenesis58
.
DLG4 (discs, large homolog 4) encodes a membrane-associated guanylate kinase and may function at postsynaptic sites (MIM
602887). Nearby,DVL2 may also play a role in signal transduction (MIM 602151) and CTDNEP1 is involved in a phosphatase cascade
regulating nuclear membrane biogenesis (MIM 610684)59
. SLC2A4 is an insulin-regulated glucose transporter (MIM 138190). The
variant identified here was previously associated with alkaline phosphatase levels in plasma52
.
EHBP1 (EH domain binding protein 1). The mouse homologue of EHBP1 was down-regulated in a transgenic Pcsk9 mouse model and
up-regulated in a Pcsk9 knockout mouse60
.
FAM13A (family with sequence similarity 13, member A). FAM13A has a putative role in signal transduction, and gene expression
has been shown to be increased in response to hypoxia in cell lines from several tissues (MIM 613299).
FAM117B (family with sequence similarity 117, member B) is an uncharacterized protein. Nearby,BMPR2 encodes a bone
morphogenetic protein receptor (MIM 600799). Defects in BMPR2 cause primary pulmonary hypertension.
FN1 (fibronectin 1) is a glycoprotein involved in cell adhesion and migration processes including embryogenesis, wound healing,
blood coagulation, host defense, and metastasis (MIM 135600). Fibronectin is one of the first extracellular matrix proteins deposited
at atherosclerosis-prone sites, and is central in the formation of atherosclerotic lesions61
.
FTO (fat mass and obesity associated) contributes to the regulation of the global metabolic rate, energy expenditure and energy
homeostasis (MIM 610966). Variants in this gene have been repeatedly associated with obesity-related phenotypes, and it may act
through hypothalamic regulation of food intake62,63
.
GPR146 (G protein-coupled receptor 146) is an orphan G protein-coupled receptor. While no ligand has yet been identified,
knockout mice exhibit reduced cholesterol levels (U. S. Patent Filing 20090036394). The adjacent gene, GPER encodes the
intracellular G protein-coupled estrogen receptor 1 (MIM 601805).
GSK3B (glycogen synthase kinase 3 beta) encodes a kinase involved in energy metabolism, neuronal cell development, and body
pattern formation (MIM 605004). In mice, Gsk3b activity regulates pancreatic islet beta cell growth64
. Nearby, NR1I2 encodes a
nuclear receptor that can form a heterodimer with retinoic acid receptor RXR and involved with homeostasis of numerous
metabolites, including lipids (MIM 603065).
HAS1 (hyaluronan synthase 1) is one of three isozymes that synthesize hyaluronic acid, produced during wound healing and tissue
repair to provide a framework for growth of blood vessels and fibroblasts (MIM 601463). The nearest gene, FPR3 (formyl peptide
receptor 3) is involved in host defense and inflammation (MIM 136539).
HBS1L (HBS1-like, S. cerevisiae) encodes a member of the GTP-binding elongation factor family (MIM 612450)65
. Variants at this
locus regulate persistence of fetal hemoglobinin adults and other haematological traits66,67
.
HDGF (hepatoma derived growth factor) and PMVK (phosphomevalonate kinase). HDGFis a growth factor that may be involved in
cell proliferation and differentiation (MIM 600339). PMVK catalyzes the fifth reaction of the cholesterol biosynthetic pathway (MIM
607622). Nearby, CRABP2 (cellular retinoic acid binding protein 2) encodes a cytosol-to-nuclear shuttling protein involved in the
retinoid signaling pathway (MIM 180231)68
.
IKZF1 (IKAROS family zinc finger 1) is a transcription factor that regulates the low-density lipoprotein receptor in certain cell types69
.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
81
INSIG2 (insulin induced gene 2). INSIG2 influences cholesterol metabolism, lipogenesis, and glucose homeostasis in diverse tissues
(MIM 608660).
INSR (insulin receptor) is a transmembrane tyrosine kinase receptor that binds insulin and stimulates glucose uptake (MIM 147670).
The receptor activates several downstream pathways.
LOC84931 (uncharacterized gene). The nearest gene to the lead signal is an uncharacterized gene with unknown function, and there
are no obvious candidate genes in the region.
LRPAP1 (low density lipoprotein receptor-related protein associated protein 1) encodes a chaperone for the lipoprotein receptor-
related proteins (MIM 104225). Lrpap1 knockout mice exhibit impaired export of LRP2 and VLDL receptors from the endoplasmic
reticulum.
KAT5 (K(lysine) acetyltransferase 5). KAT5 is a positive regulator of PPARG transcription involved in adipogenesis70
.
KCNK17 (potassium channel, subfamily K, member 17) passes outward current under physiological potassium concentrations (MIM
607370). Variants ~50 kb away at KCNK16 have been implicated in type 2 diabetes71
.
MARCH8 (membrane-associated ring finger (C3HC4) 8, E3 ubiquitin protein ligase) and ALOX5 (arachidonate 5-lipoxygenase).
MARCH8 induces the internalization of several membrane glycoproteins (MIM 613335). ALOX5 is a lipid metabolism enzyme that
catalyzes the conversion of arachidonic acid to leukotrienes, inflammatory mediators implicated in atherosclerosis and several
cancers (MIM 152390).
MET (met proto-oncogene (hepatocyte growth factor receptor)) encodes a receptor tyrosine kinase that regulates hepatocytecell
proliferation, migration and survival (MIM 164860)72,73
.
MIR148A (microRNA 148a). MicroRNAs are short non-coding RNAs involved in post-transcriptional regulation of gene expression.
miR-148a has been implicated in several cancers (MIM 613786)74,75
.
MOGAT2 (monoacylglycerol O-acyltransferase 2) and DGAT2 (diacylglycerol O-acyltransferase 2). MOGAT2 plays a central role in
absorption of dietary fat in the small intestine76
. DGAT2 encodes one of two enzymes that catalyze the final reaction in the synthesis
of triglycerides, in which diacylglycerol is covalently bound to long chain fatty acyl-CoA (MIM 606983).
MPP3 (membrane protein, palmitoylated 3) is a membrane-associated guanylate kinase that regulates trafficking and processing of
cell-cell adhesion molecule nectin-1α (MIM 601114).
MTMR3 (myotubularin related protein 3) encodes a phosphatase that binds to phosphoinositide lipids (MIM 603558).
OR4C46 (olfactory receptor, family 4, subfamily C, member 46). This signal is located in a cluster of G-protein-coupled olfactory
receptors, including OR5W2, OR5D13, and OR5AS1 (MIM 614273).
PDXDC1 (pyridoxal-dependent decarboxylase domain containing 1). Little is known about this decarboxylase (MIM 614244).
Variants at this locus have been shown previously to be associated with circulating sphingolipid levels77
. About 300 kb away,
PLA2G10 encodes a protein that releases arachidonic acid from cell membrane phospholipids (MIM 603603).
PEPD (peptidase D) encodes an enzyme that hydrolyzes peptides with C-terminal proline or hydroxyproline residues and helps
recycle proline (MIM 613230). Also at this locus are the genes encoding transcription factors CCAAT/enhancer binding protein alpha
and gamma (CEBPA (MIM 116897), CEBPG (MIM 138972)), involved in adipogenesis. Variants in this locus are associated with
adiponectin levels and type 2 diabetes in East Asians71,78
.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
82
PHC1 (polyhomeotic homolog 1) and A2ML1 (alpha-2-macroglobulin-like 1) is required to maintain the transcriptionally repressed
state of many genes (MIM 602978). A2ML1 is an inhibitor for several proteases and binds to low density lipoprotein receptor-related
protein 1 (MIM 610627)79
.
PHLDB1 (pleckstrin homology-like domain, family B, member 1). PHLDB1 is an insulin-responsive protein that enhances Akt
activation, and PHLDB1 expression is increased during adipocyte differentiation (MIM 612834)80
.
PIGV (phosphatidylinositol glycan anchor biosynthesis, class V) and NR0B2 (nuclear receptor subfamily 0, group B, member 2).
PIGV is a mannosyltransferase that plays a role in multiple cellular processes, including protein sorting and signal transduction (MIM
610274). NR0B2 is a transcriptional regulator involved in cholesterol, bile acid, and fatty acid metabolism and glucose-energy
homeostasis.
PPARA (peroxisome proliferator activated receptor alpha) encodes a nuclear transcription factor that regulates fatty acid synthesis,
and oxidation and gluconeogenesis (MIM 170998). PPARA regulates the expression of lipoprotein receptors and cholesterol
transporters involved in the reverse cholesterol transport pathway.
PXK (PX domain containing serine/threonine kinase) plays a critical role in epidermal growth factor receptor trafficking by
modulating ubiquitination of the receptor (MIM 611450)81
.
RBM5 (RNA binding motif protein 5) is an hypothetical tumour suppressor gene encoding a nuclear RNA binding protein involved in
the induction of cell cycle arrest and apoptosis (MIM 606884). Nearby, MST1R encodes macrophage stimulating 1 receptor and is
involved in host defense (MIM 600168).
RSPO3 (R-spondin 3). RSPO3 encodes a protein that regulates beta-catenin signaling, promotes angiogenesis and vascular
development (MIM 610574). In mouse, Rspo3 is required for Vegf expression and endothelial cell proliferation82
. Variants in this
locus are associated with waist-hip ratio83
, bone mineral density84
and renal traits85
.
SETD2 (SET domain containing 2) encodes a histone methyltransferase specific for lysine-36 of histone H3, a mark associated with
active chromatin (MIM 612778). Nearby, NBEAL2 encodes neurobeachin-like 2, which may play a role in megakaryocyte alpha-
granule biogenesis (MIM 614169).
SNX5 (sorting nexin 5) encodes a protein that binds to phosphatidylinositol 4,5-bisphosphate and is involved in intracellular
transport of cargo receptors from endosomes to the trans-Golgi network (MIM 605937) 86
.
SNX13 (sorting nexin 13). This gene belongs to the sorting nexin (SNX) family and the regulator of G protein signaling (RGS) family
(MIM 606589). It may be involved in several stages of intracellular trafficking.
SOX17 (SRY (sex determining region Y)-box 17) encodes a transcription regulator that plays a key role in the regulation of
embryonic development and is required for normal looping of the embryonic heart tube(MIM 610928).
SPTLC3 (serine palmitoyltransferase, long chain base subunit 3). SPTLC3 catalyzes the rate-limiting step of the de novo synthesis of
sphingolipids (MIM 611120). Variants at this locus are associated with circulating sphingolipid levels87
.
STAB1 (stabilin 1) encodes a large, transmembrane receptor involved in angiogenesis, lymphocyte homing, cell adhesion, and
receptor scavenging (MIM 608560). STAB1 mediates endocytosis of various ligands, including low-density lipoprotein88
. Variants at
this locus have been associated with waist-hip ratio83
.
TMEM176A (transmembrane protein 176A) is a transmembrane protein (MIM 610334).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
83
TOM1 (target of myb1). TOM1 shares its N-terminal domain in common with proteins associated with vesicular trafficking at the
endosomes (MIM 604700). Nearby, HMOX1 encodes an essential enzyme in heme catabolism (MIM 141250). Hmox1 knockout mice
have low plasma triglycerides and altered composition of HDL89
.
UGT1A1 (UDP glucuronosyltransferase 1 family, polypeptide A1). This complex locus encodes several glycosyltransferases that
transform small lipophilic molecules, such as steroids, bilirubin, hormones, and drugs, into water-soluble excretable metabolites
(MIM 191740). Variants at this locus are associated with serum bilirubin levels.
VEGFA (vascular endothelial growth factor A) encodes a growth factor active in angiogenesis and endothelial cell growth,
promoting cell migration, and inhibiting apoptosis (MIM 192240). Variants in this locus are associated with waist-hip ratio83
.
VIM (vimentin) and CUBN (cubilin, intrinsic factor-cobalamin receptor). VIM is an intermediate filament that controls the transport
of LDL-derived cholesterol from a lysosome to the site of esterification (MIM 193060)90
. CUBN is a receptor for high-density
lipoproteins/apolipoprotein A-I, intrinsic factor-vitamin B12, and albumin (MIM 602997).
VLDLR (very low density lipoprotein receptor) binds VLDL and other lipoproteins and transports them into cells (MIM 192977).
VLDLR is expressed on the capillary endothelium of skeletal muscle, heart, and adipose tissue91
.
ZBTB42 (zinc finger and BTB domain containing 42) and AKT1 (v-akt murine thymoma viral oncogene homolog 1). ZBTB42is a DNA-
binding transcriptional repressor (MIM 613915). AKT1 is a serine-threonine protein kinase that is activated by platelet-derived
growth factor (MIM 164730). The Akt signaling pathway controls multiple cellular functions in the cardiovascular system, and murine
Akt1 has an atheroprotective role92
.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
84
Detailed Acknowledgements
ADVANCE
The ADVANCE study was supported by a grant from the Reynold's Foundation and NHLBI grant HL087647.
AGES Reykjavik study
Age, Gene/Environment Susceptibility--Reykjavik Study has been funded by NIH contract N01-AG-12100, Hjartavernd
(the Icelandic Heart Association), and the Althingi (the Icelandic Parliament).
AIDHS/SDS
This work is supported by the NIH grants: KO1 TW006087, funded by the Fogarty International Center, R01 DK082766,
funded by National Institute of Diabetes and Digestive and Kidney Diseases, and a seed grant from University of
Oklahoma Health Sciences Center, Oklahoma City, USA.
AMC-PAS and THISEAS
Recruitment for THISEAS was partially funded by a research grant (PENED 2003) from the Greek General Secretary of
Research and Technology; we thank all the dieticians and clinicians for their contribution to the project. We thank the
members of the Wellcome Trust Sanger Institute's Genotyping Facility for genotyping and Wellcome Trust grants
083948/B/07/Z and 077016/Z/05/Z for funding.
Amish
The Amish studies are supported by grants and contracts from the NIH, including R01 AG18728, R01 HL088119, U01
GM074518, U01 HL072515-06, U01 HL84756, R01 DK54261,the University of Maryland General Clinical Research Center,
grant M01 RR 16500, the Mid-Atlantic Nutrition Obesity Research Center grant P30 DK72488, the Baltimore Diabetes
Research and Training Center grant P60DK79637, and by the T32 training grant AG000219 . In addition, this project was
supported by National Research Initiative Competitive Grant no. 2007-35205-17883 from the USDA National Institute of
Food and Agriculture. We gratefully thank our Amish community and research volunteers for their long-standing
partnership in research, and acknowledge the dedication of our Amish liaisons, field workers and the Amish Research
Clinic staff, without which these studies would not have been possible.
ARIC
The Atherosclerosis Risk in Communities Study is carried out as a collaborative study supported by National Heart, Lung,
and Blood Institute contracts N01-HC-55015, N01-HC-55016, N01-HC-55018, N01-HC-55019, N01-HC-55020, N01-HC-
55021, N01-HC-55022, R01HL087641, R01HL59367, R01HL086694 and RC2 HL102419; National Human Genome
Research Institute contract U01HG004402; and National Institutes of Health contract HHSN268200625226C. The
authors thank the staff and participants of the ARIC study for their important contributions. Infrastructure was partly
supported by Grant Number UL1RR025005, a component of the National Institutes of Health and NIH Roadmap for
Medical Research.
British 1958 cohort Metabochip (B58C-Metabochip)
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
85
DNA collection was funded by MRC grant G0000934 and cell-line creation by Wellcome Trust grant 068545/Z/02. This
research used resources provided by the Type 1 Diabetes Genetics Consortium, a collaborative clinical study sponsored
by the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK), National Institute of Allergy and
Infectious Diseases, National Human Genome Research Institute, National Institute of Child Health and Human
Development, and Juvenile Diabetes Research Foundation International (JDRF) and supported by U01 DK062418. This
study makes use of data generated by the Wellcome Trust Case-Control Consortium. A full list of investigators who
contributed to generation of the data is available from the Wellcome Trust Case-Control Consortium website. Funding
for the project was provided by the Wellcome Trust under award 076113. Dr Sue Ring and Dr Wendy McArdle
(University of Bristol), and Mr Jon Johnson (Centre for Longitudinal Studies, Institute of Education, London) are thanked
for help with data linkage. Work was undertaken at Great Ormond Street Hospital /University College London, Institute
of Child Health which received a proportion of funding from the Department of Health's National Institute of Health
Research ('Biomedical Research Centres' funding). The Medical Research Council provides funds for the MRC Centre of
Epidemiology for Child Health. C.L. is funded by WT086596/Z/08/Z (Wellcome Trust Research Career Development
Fellowship).
British 1958 cohort GWAS (B58C-WTCCC & B58C-T1DGC)
We acknowledge use of phenotype and genotype data from the British 1958 Birth Cohort DNA collection, funded by the
Medical Research Council grant G0000934 and the Wellcome Trust grant 068545/Z/02.
(http://www.b58cgene.sgul.ac.uk/). Genotyping for the B58C-WTCCC subset was funded by the Wellcome Trust grant
076113/B/04/Z. The B58C-T1DGC genotyping utilized resources provided by the Type 1 Diabetes Genetics Consortium, a
collaborative clinical study sponsored by the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK),
National Institute of Allergy and Infectious Diseases (NIAID), National Human Genome Research Institute (NHGRI),
National Institute of Child Health and Human Development (NICHD), and Juvenile Diabetes Research Foundation
International (JDRF) and supported by U01 DK062418. B58C-T1DGC GWAS data were deposited by the Diabetes and
Inflammation Laboratory, Cambridge Institute for Medical Research (CIMR), University of Cambridge, which is funded by
Juvenile Diabetes Research Foundation International, the Wellcome Trust and the National Institute for Health Research
Cambridge Biomedical Research Centre; the CIMR is in receipt of a Wellcome Trust Strategic Award (079895).
BLSA
The BLSA was supported in part by the Intramural Research Program of the NIH, National Institute on Aging. A portion
of that support was through a R&D contract with MedStar Research Institute.
BRIGHT
The BRIGHT study is supported by the Medical Research Council of Great Britain (G9521010D) and the British Heart
Foundation (BHF) (PG/02/128). Genotyping was funded by the Wellcome Trust (grant number; 076113/B/04/Z) as part
of The Wellcome Trust Case Control Consortium. TJ was supported by the Wellcome Trust (grant number
093078/Z/10/Z). The BRIGHT study is extremely grateful to all the patients who participated in the study and the BRIGHT
nursing team. BRIGHT investigators are also supported by BHF grants (RG/07/005/23633 and SP/08/005/25115) and EU
Ingenious HyperCare Consortium: Integrated Genomics, Clinical Research and Care in Hypertension (LSHM-C7-2006-
037093). This work forms part of the research themes contributing to the translational research portfolio of Barts
Cardiovascular Biomedical Research Unit which is supported and funded by the National Institute for Health Research.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
86
CHS
Cardiovascular Health Study: This CHS research was supported by NHLBI contracts N01-HC-85239, N01-HC-85079
through N01-HC-85086; N01-HC-35129, N01 HC-15103, N01 HC-55222, N01-HC-75150, N01-HC-45133 and NHLBI grants
HL080295, HL075366, HL087652, HL105756 with additional contribution from NINDS. Additional support was provided
through AG-023629, AG-15928, AG-20098, and AG-027058 from the NIA. See also http://www.chs-nhlbi.org/pi.htm.
DNA handling and genotyping was supported in part by National Center for Research Resources CTSI grant UL
1RR033176 and National Institute of Diabetes and Digestive and Kidney Diseases grant DK063491 to the Southern
California Diabetes Endocrinology Research Center, and Cedars-Sinai Board of Governors' Chair in Medical Genetics (JIR).
CLHNS
We thank the Office of Population Studies Foundation research and data collection teams and the study participants
who generously provided their time for this study. This work was supported by National Institutes of Health grants
DK078150, TW05596, HL085144, HD054501, RR20649, ES10126, and DK56350.
CoLaus
The CoLaus study was supported by research grants from GlaxoSmithKline, the Faculty of Biology and Medicine of
Lausanne, Switzerland and the Swiss National Science Foundation (grant no: 33CSCO-122661 and FN 33CSC0-139468).
The authors also express their gratitude to the participants in the Lausanne CoLaus study and to the investigators who
have contributed to the recruitment, in particular Yolande Barreau, Anne-Lise Bastian, Binasa Ramic, Martine
Moranville, Martine Baumer, Marcy Sagette, Jeanne Ecoffey and Sylvie Mermoud, the nurses of the CoLaus study
(CHUV, Lausanne, Switzerland) for data collection.
CROATIA-Vis
The CROATIA-Vis study in the Croatian island of Vis was supported through the grants from the Medical Research
Council UK and Ministry of Science, Education and Sport of the Republic of Croatia (number 108-1080315-0302) and the
European Union framework program 6 EUROSPAN project (contract no. LSHG-CT-2006-018947). We would like to
acknowledge the invaluable contributions of the recruitment team (including those from the Institute of Anthropological
Research in Zagreb) in Vis, the administrative teams in Croatia and Edinburgh and the people of Vis.
deCODE
The deCODE authors would like to thank participants in deCODE cardiovascular- and obesity studies and collaborators for their cooperation. We would also like to acknowledge the staff at the Clinical Research Centre (Iceland) and the deCODE Genetics biological materials and genotyping facilities for their work. The research performed at deCODE Genetics was in part funded through the European Community's Seventh Framework Programme (FP7/2007-2013), ENGAGE project, grant agreement HEALTH-F4-2007- 201413 and the NIH grant, National Heart, Lung and Blood Institute R01HL089650‐02. DIAGEN
The DIAGEN study was supported by the Commission of the European Communities funded project “CEED3 -
Collaborative European Effort to Develop Diabetes Diagnostics”, Grant Agreement number - 223211. We are grateful to
all of the patients who cooperated in this study and to their referring physicians and diabetologists in Saxony.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
87
DILGOM
DILGOM Study was funded in part by the Academy of Finland (grant number 118065). S.R was supported by the
Academy of Finland Center of Excellence in Complex Disease Genetics (213506 and 129680), Academy of Finland
(251217), the Finnish foundation for Cardiovascular Research and the Sigrid Juselius Foundation. MP is partly financially
supported for this work by the Finnish Academy SALVE program ‘‘Pubgensense’’ 129322, by the Finnish Foundation for
Cardiovascular Research and by the University of Tartu (SP1GVARENG).
DPS
The DPS has been financially supported by grants from the Academy of Finland (117844 and 40758, 211497, and
118590; The EVO funding of the Kuopio University Hospital from Ministry of Health and Social Affairs (5254), Finnish
Funding Agency for Technology and Innovation (40058/07), Nordic Centre of Excellence on Systems biology in controlled
dietary interventions and cohort studies, SYSDIET (070014), The Finnish Diabetes Research Foundation, Yrjö Jahnsson
Foundation (56358), Sigrid Juselius Foundation, Juho Vainio Foundation and TEKES grants 70103/06 and 40058/07.
DR's EXTRA
The DR.s EXTRA Study was supported by grants by the Ministry of Education and Culture of Finland (627;2004-2011),
Academy of Finland (102318; 123885; 211119), Kuopio University Hospital , Finnish Diabetes Association, Finnish Heart
Association, Päivikki and Sakari Sohlberg Foundation and by grants from European Commission FP6 Integrated Project
(EXGENESIS); LSHM-CT-2004-005272, City of Kuopio and Social Insurance Institution of Finland (4/26/2010).
EAS
The Edinburg Artery Study was supported by the British Heart Foundation. Genotyping was funded by a project grant
from the Chief Scientist Office, Scotland (CZB/4/672).
EGCUT
EGCUT received financing by FP7 grants (201413, 245536), also received targeted financing from Estonian Government
SF0180142s08, direct funding from the ministries of research and science and social affairs and through 3.2.0304.11-
0312, EXCEGEN and by the Development Fund of the University of Tartu (grant:SP1GVARENG).
ELY
The Ely Study was funded by the MRC and Diabetes UK. We are grateful to all the volunteers, and to the staff of St.
Mary’s Street Surgery, Ely and the study team. The analyses in this paper were supported by the Medical Research
Council (MC_U106179471, G0701863).
ENGAGE
The European Network for Genetic and Genomic Epidemiology consortium has received funding from the European
Community's Seventh Framework Programme (FP7/2007-2013)/grant agreement HEALTH-F4-2007-201413. The
GenomEUtwin project was supported by the European Commission under the 5th Framework Programme 'Quality of
Life and Management of the Living Resources' of 5th Framework Programme (no. QLRT-2001-01254; QLG2-CT-2002-
01254). The Swedish Twin Cohort within GenomEUtwin was also supported by the Swedish Research Council and
Swedish Foundation for Strategic Research. The Danish Twin Registry has been supported by the Danish Medical
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
88
Research Council, the Danish Diabetes Foundation, the Danish Heart Association, and the Novo Nordic Foundation. The
TWINSUK study was funded by the Wellcome Trust (Grant ref. 079771); European Community’s Framework 6 Project
EUroClot. The study also receives support from the National Institute for Health Research (NIHR) comprehensive
Biomedical Research Centre award to Guy's & St Thomas' NHS Foundation Trust in partnership with King's College
London. Timothy D Spector is an NIHR senior Investigator and is holder of an ERC Advanced Principal Investigator
award. Genotyping was performed by The Wellcome Trust Sanger Institute, support of the National Eye Institute via an
NIH/CIDR genotyping project. The Australian Twin cohort was funded by the Australian National Health and Medical
Research Council (241944, 339462, 389927, 389875, 389891, 389892, 389938, 442915, 442981, 496739, 552485,
552498), the Australian Research Council (A7960034, A79906588, A79801419, DP0770096, DP0212016, DP0343921) and
the U.S. National Institutes of Health (AA07535, AA10248, AA11998, AA13320, AA13321, AA13326, AA14041, AA17688,
DA12854, MH66206). Netherlands Twin Registry (NTR) has been supported by the Netherlands Scientific Organization
(911-09-032, 480-04-004, CMSB NWO Genomics, NWO-SPI 56-464-1419, NBIC/BioAssist/RK/2008.024), BBMRI-NL,
Institute for Health and Care Research (EMGO+) , the European Union (EU/WLRT-2001-01254, ERC Advanced 230374).
The FinnTwin cohorts were also supported by Academy of Finland Center of Excellence in Complex Disease Genetics
(grant numbers: 213506, 129680), the Finnish Foundation for Cardiovascular research and the Finnish Academy (grant
numbers 129494, 139635, and 251217) and the Sigrid Juselius Foundation.
Special Population Research Network (EUROSPAN) was supported by European Commission FP6 STRP grant number
018947 (LSHG-CT-2006-01947). In South Tyrol, the MICROS Study was supported by the Ministry of Health of the
Autonomous Province of Bolzano and the South Tyrolean Sparkasse Foundation.The Vis Study in the Croatian island of
Vis was supported by the Medical research Council UK and Ministry of Science, Education and Sport of the Republic of
Croatia. (number 108-1080315-0302). Erasmus Ruchpen Family (ERF) was supported by grants from The Netherlands
Organization for Scientific Research (NOW; Pionier Grant), Erasmus MC, and the Netherlands Genomics Initiative (NGI)-
sponsored Center for Medical Systems Biology (CMSB). The Northern Swedish Population Health Study (NSPHS) was
funded by the Swedish Medical Research Council (Project Number K2007-66X-20270-01-3) and the Foundation for
Strategic Research (SSF).
The Northern Finland Birth Cohort 1966 received financial support from the Academy of Finland (project grants 104781,
120315, 129269 (SALVE), 114194, and Center of Excellence in Complex Disease Genetics), University Hospital Oulu,
Biocenter, University of Oulu, Finland, NHLBI grant 5R01HL087679 through the STAMPEED program (1RL1MH083268-
01), the Medical Research Council (grant G0500539, centre grant G0600705, PrevMetSyn), and the Wellcome Trust
(project grant GR069224), UK. The genotyping of NFBC1966 was funded by NHLBI grant 5R01HL087679, the Academy of
Finland and Biocentrum Helsinki.
The generation and management of GWAS genotype data for the Rotterdam Study is supported by the Netherlands
Organisation of Scientific Research NWO Investments (nr. 175.010.2005.011, 911-03-012). This study is funded by the
Research Institute for Diseases in the Elderly (014-93-015; RIDE2), the Netherlands Genomics Initiative
(NGI)/Netherlands Organisation for Scientific Research (NWO) project nr. 050-060-810. We thank Pascal Arp, Mila
Jhamai, Marijn Verkerk, Lizbeth Herrera and Marjolein Peters for their help in creating the GWAS database, and Karol
Estrada and Maksim V. Struchalin for their support in creation and analysis of imputed data. The Rotterdam Study is
funded by Erasmus Medical Center and Erasmus University, Rotterdam, Netherlands Organization for the Health
Research and Development (ZonMw), the Research Institute for Diseases in the Elderly (RIDE), the Ministry of Education,
Culture and Science, the Ministry for Health, Welfare and Sports, the European Commission (DG XII), and the
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
89
Municipality of Rotterdam. The authors are grateful to the study participants, the staff from the Rotterdam Study and
the participating general practitioners and pharmacists.
GENMETS was supported through funds from The European Community's Seventh Framework Programme (FP7/2007-
2013), BioSHaRE Consortium, grant agreement 261433. GWAS genotyping was supported by the Wellcome Trust Sanger
Institute.
FINRISK was funded by the Finnish National Institute for Health and Welfare, supplemented by grants from the Finnish
Foundation for Cardiovascular Research and from the Academy of Finland.
Veikko Salomaa was supported by the Academy of Finland (grant numbers 129 494 and 139 635) and the Finnish
Foundation for Cardiovascular Disease.
EPIC
EPIC-Norfolk is supported by the Medical Research Council programme grants (G0401527,G1000143) and Cancer
Research UK programme grants (C864/A8257, C864/A14136). The analyses in this paper were supported by the Medical
Research Council (MC_U106179471, G0701863, G0801566) and the British Heart Foundation (PG/08/094/26019). The
authors would like to acknowledge the contribution of the staff and participants of the EPIC-Norfolk Study. I.B.
acknowledges support from EU FP6 funding (contract no LSHM-CT-2003-503041) and by the Wellcome Trust.
ERF Study
Erasmus Rucphen Family (ERF) was supported by grants from The Netherlands Organization for Scientific Research
(NWO; Pionier Grant), Erasmus MC, and the Netherlands Genomics Initiative (NGI)-sponsored Center of Medical Systems
Biology (CMSB).
EUROSPAN
EUROSPAN (European Special Populations Research Network) was supported by European Commission FP6 STRP grant
number 018947 (LSHG-CT-2006-01947). Lipidomic analysis was supported by the European Commission FP7 grant
LipidomicNet (2007-202272). Analyses of lipidomics data were run on the Genetic Cluster Computer, Amsterdam
(http://www.geneticcluster.org).
Family Heart Study (FHS)
The Family Heart Study is supported in part by NHLBI grants 5R01HL087700 and 5R01HL088215 (M. Provice) and NIDDK
grants 5R01DK075681 and 1R01DK8925601 (I. Borecki).
FBPP
We would like to acknowledge and thank all participants in the GenNet and HyperGENE studies. The Family Blood
Pressure Program studies are supported in part by NHLBI grant 2R01HL086694 (A. Chakravarti).
FENLAND
The Fenland Study is funded by the Medical Research Council (MC_U106179471 and G0701863,) and the Wellcome
Trust. We are grateful to all the volunteers for their time and help, and to the General Practitioners and practice staff for
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
90
help with recruitment. We thank the Fenland Study coordination team and the Field Epidemiology team of the MRC
Epidemiology Unit for recruitment and clinical testing. We also thank the NIHR Cambridge Biomedical Research Centre,
Cambridge, U.K. for biochemical analyses.
FINCAVAS
This work was supported by the Competitive Research Funding of the Tampere University Hospital (Grant 9M048 and
9N035), the Finnish Cultural Foundation, the Finnish Foundation for Cardiovascular Research, the Emil Aaltonen
Foundation, Finland, and the Tampere Tuberculosis Foundation. The authors thank the staff of the Department of
Clinical Physiology for collecting the exercise test data.
FIN-D2D 2007
The FIN-D2D study has been financially supported by the hospital districts of Pirkanmaa, South Ostrobothnia, and
Central Finland, the Finnish National Public Health Institute (current National Institute for Health and Welfare), the
Finnish Diabetes Association, the Ministry of Social Affairs and Health in Finland, the Academy of Finland (grant number
129293),Commission of the European Communities, Directorate C-Public Health (grant agreement no. 2004310) and
Finland’s Slottery Machine Association.
Framingham
The Framingham Heart Study is supported by National Heart, Lung, and Blood Institute grant HL-54776 and contracts 53-
K06-5-10 and 58-1950-9-001 from the USDA Research Service.The Framingham Heart Study SHARe Project was partially
supported by the NHLBI Framingham Heart Study (Contract No. N01-HC-25195) and its contract with Affymetrix, Inc for
genotyping services (Contract No. N02-HL-6-4278). DNA isolation and biochemistry were partly supported by NHLBI HL-
54776. A portion of this research utilized the Linux Cluster for Genetic Analysis (LinGA-II) funded by the Robert Dawson
Evans Endowment of the Department of Medicine at Boston University School of Medicine and Boston Medical Center.
FRISCII
This part of the FRISCII study was supported by the Swedish Research Council and the Swedish Heart and Lung
Foundation.
FUSION
Support for FUSION was provided by NIH grants R01-DK062370 (to M.B.), R01-DK072193 (to K.L.M.), and intramural
project number 1Z01-HG000024 (to F.S.C.). Genome-wide genotyping was conducted by the Johns Hopkins University
Genetic Resources Core Facility SNP Center at the Center for Inherited Disease Research (CIDR), with support from CIDR
NIH contract no. N01-HG-65403.
GLACIER
The GLACIER Study was funded by grants from the Swedish Diabetes Association, Swedish Heart-Lung Foundation,
Swedish Research Council, Medical Research Foundation of Umeå University, and Novo Nordisk (all to PWF). We thank
the participants for there outstanding contributions to VIP and the GLACIER Study. We also thank the staff of the Umeå
Medical Biobank, especially Göran Hallmans, Åsa Agren, John Hutilainen, and Ann-Marie Ahren for data reteival and
organisation and Kerstin Enqusit and Tore Johansson for expert assistance with DNA extraction and plating. The GLACIER
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
91
Study is nested within the Västerbottens Intervention Project (VIP); we thank the staff of the VIP Study for phenotype
data collection, particularly Lars Wennehall who leads the VIP Study. Inês Barroso acknowledges funding from the
Wellcome Trust grant 098051, United Kingdom NIHR Cambridge Biomedical Research Centre and the MRC Centre for
Obesity and Related Metabolic Diseases. We would like to thank Sarah Edkins, Emma Gray, Douglas Simpkin, Sarah Hunt
and staff of the WTSI Sample Logistics, Genotyping and Variation Informatics Facilities.
Go-DARTS
We acknowledge the support of the Health Informatics Centre, University of Dundee for managing and supplying the
anonymised data and NHS Tayside, the original data owner. We are grateful to all the participants who took part in the
Go-DARTS study, to the general practitioners, to the Scottish School of Primary Care for their help in recruiting the
participants, and to the whole team, which includes interviewers, computer and laboratory technicians, clerical workers,
research scientists, volunteers, managers, receptionists, and nurses.
GxE/Spanish Town
Principally, our thanks go to the participants in the Spanish Town and GxE studies. We also thank Nurse Orgen Brown,
Mrs Stacey Chin, Mr Windsor Cuffe, and other past and present members of the labs at TMRU for their assistance in
carrying out these studies. This work was supported by NIH Grant R01HL53353.
HUNT2
The Nord-Trøndelag Health Study (The HUNT Study) is a collaboration between HUNT Research Centre (Faculty of
Medicine, Norwegian University of Science and Technology NTNU), Nord-Trøndelag County Council, Central Norway
Health Authority, and the Norwegian Institute of Public Health.
IMPROVE
The authors wish to express their deep and sincere appreciation to all members of the IMPROVE group for their time
and extraordinary commitment. The IMPROVE study was funded by the European Commission (Contract number:
QLG1-CT-2002-00896), the Academy of Finland (Grant #110413), the British Heart Foundation (RG2008/008), the Italian
Ministry of Health (Ricerca Corrente), the Swedish Heart-Lung Foundation, the Swedish Research Council (8691 and
09533), the Knut and Alice Wallenberg Foundation, the Foundation for Strategic Research, the Torsten and Ragnar
Söderberg Foundation, the Strategic Cardiovascular and Diabetes Programmes of Karolinska Institutet, and the
Stockholm County Council (560183, 562183).
InCHIANTI
The InCHIANTI study baseline (1998-2000) was supported as a "targeted project" (ICS110.1/RF97.71) by the Italian
Ministry of Health and in part by the U.S. National Institute on Aging (Contracts: 263 MD 9164 and 263 MD 821336).
KORA
The KORA Augsburg studies were financed by the Helmholtz Zentrum München, German Research Center for
Environmental Health, Neuherberg, Germany and supported by grants from the German Federal Ministry of Education
and Research (BMBF). Part of this work was financed by the German National Genome Research Network (NGFN). Our
research was supported within the Munich Center of Health Sciences (MC Health) as part of LMUinnovativ.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
92
LIFELINES
The LifeLines Cohort Study, and generation and management of GWAS genotype data for the LifeLines Cohort Study is
supported by the Netherlands Organization of Scientific Research NWO (grant 175.010.2007.006), the Economic
Structure Enhancing Fund (FES) of the Dutch government, the Ministry of Economic Affairs, the Ministry of Education,
Culture and Science, the Ministry for Health, Welfare and Sports, the Northern Netherlands Collaboration of Provinces
(SNN), the Province of Groningen, University Medical Center Groningen, the University of Groningen, Dutch Kidney
Foundation and Dutch Diabetes Research Foundation.
We thank Behrooz Alizadeh, Annemieke Boesjes, Marcel Bruinenberg, Noortje Festen, Pim van der Harst, Ilja Nolte, Lude
Franke, Mitra Valimohammadi for their help in creating the GWAS database, and Rob Bieringa, Joost Keers, René
Oostergo, Rosalie Visser, Judith Vonk for their work related to data-collection and validation. The authors are grateful to
the study participants, the staff from the LifeLines Cohort Study and Medical Biobank Northern Netherlands, and the
participating general practitioners and pharmacists.
LifeLines Scientific Protocol Preparation: Rudolf de Boer, Hans Hillege, Melanie van der Klauw, Gerjan Navis, Hans Ormel,
Dirkje Postma, Judith Rosmalen, Joris Slaets, Ronald Stolk, Bruce Wolffenbuttel.
LifeLines GWAS Working Group: Behrooz Alizadeh, Marike Boezen, Marcel Bruinenberg, Lude Franke, Pim van der Harst,
Gerjan Navis, Dirkje Postma, Harold Snieder, Cisca Wijmenga, Bruce Wolffenbuttel.
LOLIPOP
LOLIPOP (London Life Sciences Prospective Population Cohort) - The LOLIPOP study is supported by the National
Institute for Health Research (NIHR) Comprehensive Biomedical Research Centre Imperial College Healthcare NHS Trust;
the NIHR Cardiovascular Biomedical Research Unit of Royal Brompton and Harefield NHS Foundation Trust; the British
Heart Foundation (SP/04/002); the Medical Research Council (G0601966,G0700931); the Wellcome Trust
(084723/Z/08/Z); the NIHR (RP-PG-0407-10371); European Union FP7 (EpiMigrant, 279143) and Action on Hearing Loss
(G51). PE is an NIHR Senior Investigator. The funders had no role in study design, data collection and analysis, decision to
publish, or preparation of the manuscript. We thank the participants and research teams involved in LOLIPOP. We thank
GSK for support and genotyping of the Affymetrix data.
LURIC
We extend appreciation to the participants of the Ludwigshafen Risk and Cardiovascular Health Study. We thank the
LURIC study team either temporarily or permanently involved in patient recruitment, sample and data handling, and the
laboratory staff at the Ludwigshafen General Hospital and the Universities of Freiburg (Germany), Ulm (Germany), and
Graz (Austria). LURIC has received funding from the 6th Framework Program (integrated project Bloodomics, grant
LSHM-CT-2004-503485) and 7th of Framework Program (integrated project AtheroRemo, Grant Agreement number
201668) of the European Union.
Medstar
Medstar was funded through grants from Leducq Fondation and NIH (grant no. HL089309).
METSIM
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
93
The METSIM study was funded by the Academy of Finland (grants no. 77299 and 124243).
MICROS
For the MICROS study, we thank the primary care practitioners in the villages of the Val Venosta and the personnel of
the Hospital of Silandro (Department of Laboratory Medicine) for their participation and collaboration in the research
project. In South Tyrol, the study was supported by the Ministry of Health and Department of Educational Assistance,
University and Research of the Autonomous Province of Bolzano, and the South Tyrolean Sparkasse Foundation.
MORGAM
The MORGAM study was part funded through the European Community's Sixth Framework Programme Cardiogenics
project, grant agreement LSHM-CT-2006-037593 and Seventh Framework Programme ENGAGE project, grant agreement
HEALTH-F4-2007-201413. We would like to acknowledge the staff from the Genotyping Facilities at the Wellcome Trust
Sanger Institute for genotyping the data.
Sites and key personnel of contributing MORGAM Centres:
Finland
FINRISK, National Institute for Health and Welfare, Helsinki: V. Salomaa (principal investigator), A. Juolevi, E. Vartiainen,
P. Jousilahti; ATBC, National Institute for Health and Welfare, Helsinki: J. Virtamo (principal investigator), H. Kilpeläinen;
MORGAM Data Centre, National Institute for Health and Welfare, Helsinki: K. Kuulasmaa (responsible person), Z.
Cepaitis, A. Haukijärvi, B. Joseph, J. Karvanen, S. Kulathinal, M. Niemelä, O. Saarela; MORGAM Central Laboratory,
National Institute for Health and Welfare, Helsinki: M. Perola (responsible person), P. Laiho, M. Sauramo. The ATBC
Study was supported by US Public Health Service contracts N01-CN-45165, N01-RC-45035 and N01-RC-37004 from the
National Cancer Institute.
France
National Coordinating Centre, National Institute of Health and Medical Research (U258), Paris: P. Ducimetière (national
coordinator), A. Bingham; PRIME/Strasbourg, Department of Epidemiology and Public Health, EA 3430, University of
Strasbourg, Faculty of Medicine, Strasbourg: D. Arveiler (principal investigator), B. Haas, A. Wagner; PRIME/Toulouse,
Department of Epidemiology, Toulouse University School of Medicine, Toulouse: J. Ferrières (Principal Investigator), J-B.
Ruidavets, V. Bongard, D. Deckers, C. Saulet, S. Barrere; PRIME/Lille, Department of Epidemiology and Public Health,
INSERM U744-Université Lille Nord de France – Institut Pasteur de Lille: P. Amouyel (principal investigator), M. Montaye,
B. Lemaire, S. Beauchant, D. Cottel, C. Graux, N. Marecaux, C. Steclebout, S. Szeremeta; MORGAM Laboratory, INSERM
U937, Paris: F. Cambien (responsible person), L. Tiret, V. Nicaud.
Italy
Centro Ricerche EPIMED - Epidemiologia e Medicina Preventiva, Dipartimento di Medicina Sperimentale. Università
degli Studi dell'Insubria, Varese: M. Ferrario (principal investigator), G. Veronesi. Research Centre on Public Health,
University of Milano-Bicocca, Monza, Italy: Giancarlo Cesana. This study was supported by the Health Administration of
Regione Lombardia [grant numbers 9783/1986, 41795/1993, 31737/1997 and 17155/2004], for the baseline
examinations and the follow-up.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
94
United Kingdom
PRIME/Belfast, Queen's University Belfast, Belfast, Northern Ireland: F. Kee (principal investigator) A. Evans (former
principal investigator), J. Yarnell, E. Gardner; MORGAM Coordinating Centre, Queen's University Belfast, Belfast,
Northern Ireland: A. Evans (MORGAM coordinator), S. Cashman, F Kee.
MORGAM Management Group: A. Evans (chair, Belfast, UK), S. Blankenberg (Hamburg, Germany), F. Cambien (Paris,
France), M. Ferrario (Varese, Italy), K. Kuulasmaa (Helsinki, Finland), A. Palotie (Cambridge, UK), M. Perola (Helsinki,
Finland), A. Peters (Neuherberg, Germany), V. Salomaa (Helsinki, Finland), H. Tunstall-Pedoe (Dundee, Scotland), P.G.
Wiklund (Umeå, Sweden); Previous members: K. Asplund (Stockholm, Sweden), L. Peltonen (Helsinki, Finland), D.
Shields (Dublin, Ireland), B. Stegmayr (Umeå, Sweden).
MRC/UVRI GPC GWAS
The MRC/UVRI GPC study is supported by the Medical Research Council (MRC/UVRI Uganda Research Unit on AIDS core
funding, Grants G0901213-92157 and G0801566) and the Wellcome Trust Sanger Institute.
MRC National Survey of Health & Development
This work was supported by the Medical Research Council [U123092720]. We are very grateful to the members of this
birth cohort for their continuing interest and participation in the study.
NFBC1986
We thank Professor Paula Rantakallio (launch of NFBC1986 and initial data collection), Ms Sarianna Vaara (data
collection), Ms Tuula Ylitalo (administration), Mr Markku Koiranen (data management), Ms Outi Tornwall and Ms Minttu
Jussila (DNA biobanking). This work was supported by the Academy of Finland [project grants 104781, 120315, 129418,
add the following : 129269, 24300796, 1114194, Center of Excellence in Complex Disease Genetics and Public Health
Challenges Research Program (SALVE)], University Hospital Oulu, Biocenter, University of Oulu, Finland (75617), the
European Commission [EURO-BLCS, Framework 5 award QLG1-CT-2000-S3601643], The National Heart, Lung and Blood
Institute [5R01HL087679-02] through the SNP Typing for Association with Multiple Phenotypes from Existing
Epidemiologic Data (STAMPEED) program [1RL1MH083268-01], The National Institute of Health/The National Institute
of Mental Health [5R01MH63706:02], European Network of Genomic and Genetic Epidemiology (ENGAGE) project and
grant agreement [HEALTH-F4-2007-201413], and the Medical Research Council, UK [G0500539, G0600705, PrevMetSyn/
Public Health Challenges Research Program (SALVE)].
NSPHS
The Northern Swedish Population Health Study (NSPHS) was funded by the Swedish Medical Research Council (project
number K2007-66X-20270-01-3), and the Foundation for Strategic Research (SSF). The NSPHS as part of EUROSPAN
(European Special Populations Research Network) was also supported by European Commission FP6 STRP grant number
01947 (LSHG-CT-2006-01947). This work was also supported by the Swedish Society for Medical Research (ÅJ). The
authors are grateful for the contribution of district nurse Svea Hennix for data collection and Inger Jonasson for logistics
and coordination of the health survey. Finally, the authors thank all the community participants for their interest and
willingness to contribute to the study.
ORCADES
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
95
ORCADES was supported by the Chief Scientist Office of the Scottish Government, the Royal Society, the MRC Human
Genetics Unit, Arthritis Research UK and the European Union framework program 6 EUROSPAN project (contract no.
LSHG-CT-2006-018947). DNA extractions were performed at the Wellcome Trust Clinical Research Facility in Edinburgh.
We would like to acknowledge the invaluable contributions of Lorraine Anderson and the research nurses in Orkney, the
administrative team in Edinburgh and the people of Orkney.
PARC
This work was funded by National Institutes of Health grant U01 HL69757. DNA handling and genotyping at Cedars-Sinai
Medical Center was supported in part by the National Center for Research Resources, grant UL1RR033176, and is now at
the National Center for Advancing Translational Sciences, CTSI grant UL1TR000124; in addition to the National Institute
of Diabetes and Digestive and Kidney Disease grant DK063491 to the Southern California Diabetes Endocrinology
Research Center.
PennCath
PennCath was funded through grants from Leducq Fondation and NIH (grant no. HL089309).
PIVUS
This project was supported by grants from the Swedish Research Council, the Swedish Heart-Lung Foundation, the
Swedish Foundation for Strategic Research, the Royal Swedish Academy of Sciences, Swedish Diabetes Foundation,
Swedish Society of Medicine, and Novo Nordisk Fonden. Genotyping was performed by the SNP&SEQ Technology
Platform in Uppsala (www.genotyping.se). We thank Tomas Axelsson, Ann-Christine Wiman and Caisa Pöntinen for their
excellent assistance with genotyping. The SNP Technology Platform is supported by Uppsala University, Uppsala
University Hospital and the Swedish Research Council for Infrastructures.
SardiNIA
We thank all the volunteers who generously participated in this study, Monsignore Piseddu, Bishop of Ogliastra and the
mayors and citizens of the Sardinian towns (Lanusei, Ilbono, Arzana, and Elini). This work was supported by the
Intramural Research Program of the National Institute on Aging (NIA), National Institutes of Health (NIH). The SardiNIA
(“Progenia”) team was supported by Contract NO1-AG-1–2109 from the NIA; the efforts of GRA were supported in part
by contract 263-MA-410953 from the NIA to the University of Michigan and by research grant HG002651 and HL084729
from the NIH (to GRA).
SCARFSHEEP
The SCARFSHEEP study was funded by the Swedish Heart-Lung Foundation, the Swedish Research Council (8691 and
09533), the Knut and Alice Wallenberg Foundation, the Foundation for Strategic Research, the Torsten and Ragnar
Söderberg Foundation, the Strategic Cardiovascular and Diabetes Programmes of Karolinska Institutet, and the
Stockholm County Council (560183, 562183).
SEYCHELLES
The Seychelles study was financed by grants from the Swiss National Science Foundation (TANDEM No 31–51115.97;
PROSPER 3200BO-111362/1, 3233BO-111361/1 ; 33CM30-124087/1). We thank the Ministry of Health of the Republic of
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
96
Seychelles for continued support of this epidemiologic research and Air Seychelles and SkyChef for their logistic support
in transporting equipment and samples. Murielle Bochud is supported by the Swiss School of Public Health Plus (SSPH+).
SUVIMAX
This work was supported by the Institut National de la Santé et de la Recherche Médicale, the Institut National de la
Recherche Agronomique, the Université Paris 13, the Centre National de Génotypage and the Commissariat à L'Energie
Atomique.
Swedish Twin Registry
This work was supported by grants from the US National Institutes of Health (AG028555, AG08724, AG04563, AG10175,
AG08861), the Swedish Research Council, the Swedish Heart-Lung Foundation, the Swedish Foundation for Strategic
Research, and the Royal Swedish Academy of Science., and ENGAGE (within the European Union Seventh Framework
Programme, HEALTH-F4-2007-201413). Genotyping was performed by the SNP&SEQ Technology Platform in Uppsala
(www.genotyping.se). We thank Tomas Axelsson, Ann-Christine Wiman and Caisa Pöntinen for their excellent assistance
with genotyping. The SNP Technology Platform is supported by Uppsala University, Uppsala University Hospital and the
Swedish Research Council for Infrastructures.
TAICHI
TAICHI consoritum study was supported by the National Institutes of Health, USA [HL087647, HL088457, EY014684,
DK079888], ARRA Supplement [EY014684-03S1, -04S1], the National Health Research Institutes, Taiwan [PH-100-SP-01,
BS-094-PP-01, PH-100-PP-03], the National Science Council, Taiwan [NSC 98-2314-B-075A-002-MY3, NSC 96-2314-B-002-
151, NSC 96-2314-B-002-152, NSC 98-2314-B-002-122-MY2, NSC 100-2314-B-002-115, NSC 101-2325-002-078], the
Taichung Veterans General Hospital, Taiwan [TCVGH-1013001C, TCVGH-1013002D], and the National Taiwan University
Hospital, Taiwan [NTUH 98-N1266, NTUH 100-N1775, NTUH 101-N2010, NTUH 101-N, VN101-04, NTUH 101-S1784].
Tromsø
The work was supported by the University of Tromsø, Norwegian Research Council (project number 185764), and the
National Health Screening Service (current Norwegian Institute of Public Health).
TWINGENE
This work was supported by grants from the Ministry for Higher Education, the Swedish Research Council (M-2005-1112
and 2009-2298), GenomEUtwin (EU/QLRT-2001-01254; QLG2-CT-2002-01254),and NIH grant DK U01-066134, The
Swedish Foundation for Strategic Research (SSF; ICA08-0047). We thank Tomas Axelsson, Ann-Christine Wiman and
Caisa Pöntinen for their excellent assistance with genotyping. The SNP Technology Platform is supported by Uppsala
University, Uppsala University Hospital and the Swedish Research Council for Infrastructures.
ULSAM
This project was supported by grants from the Swedish Research Council, the Swedish Heart-Lung Foundation, the
Swedish Foundation for Strategic Research, the Royal Swedish Academy of Sciences, Swedish Diabetes Foundation,
Swedish Society of Medicine, and Novo Nordisk Fonden. Genotyping was performed by the SNP&SEQ Technology
Platform in Uppsala (www.genotyping.se). We thank Tomas Axelsson, Ann-Christine Wiman and Caisa Pöntinen for their
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
97
excellent assistance with genotyping. The SNP Technology Platform is supported by Uppsala University, Uppsala
University Hospital and the Swedish Research Council for Infrastructures.
WGHS
The WGHS is supported by HL 043851 and HL080467 from the National Heart, Lung, and Blood Institute and CA 047988
from the National Cancer Institute, the Donald W. Reynolds Foundation and the Fondation Leducq, with collaborative
scientific support and funding for genotyping provided by Amgen.
Whitehall II
The Whitehall II study has been supported by grants from the Medical Research Council; Economic and Social Research
Council; British Heart Foundation RG/10/12/28456); Health and Safety Executive; Department of Health; National Heart
Lung and Blood Institute (HL36310), US, NIH: National Institute on Aging (AG13196), US, NIH; Agency for Health Care
Policy Research (HS06516); and the John D and Catherine T MacArthur Foundation Research Networks on Successful
Midlife Development and Socio-economic Status and Health.
MISCELLANEOUS
Ellen Schmidt is supported by the National Science Foundation Open Data IGERT Grant 0903629. Cristen Willer is supported by HL094535 and HL109946.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
98
Full Name Author Contributions
Writing and Analysis Group
Gonçalo R Abecasis, Michael Boehnke, L Adrienne Cupples, Panos Deloukas, Paul W Franks, Sekar Kathiresan, Karen L Mohlke, Erik
Ingelsson, Gina M Peloso, Stephen S Rich, Samuli Ripatti, Manjinder S Sandhu, Ellen M Schmidt, Sebanti Sengupta, Cristen J Willer
(Lead). Ellen M Schmidt and Sebanti Sengupta performed meta-analysis and Ellen M Schmidt, Sebanti Sengupta, Gina M Peloso,
Martin L Buchkovich, Jin Chen, Stefan Gustafsson, Andrea Ganna, and Stavroula Kanoni performed bioinformatics analyses. Ellen M
Schmidt and Sebanti Sengupta prepared the tables, figures and supplemental material. Cristen J Willer led the analysis and
bioinformatics efforts. Erik Ingelsson and Karen L Mohlke led the biological interpretation of results. Cristen J Willer and Gonçalo R
Abecasis wrote the manuscript. All analysis and writing group authors extensively discussed the analysis, results, interpretation and
presentation of results.
All authors contributed to the research and reviewed the manuscript.
Design, management and coordination of contributing cohorts
(ADVANCE) Themistocles L Assimes; (AGES Reykjavik study) Tamara B Harris, Vilmundur Gudnason; (AIDHS/SDS) Dharambir K
Sanghera; (AMC-PAS) Panos Deloukas, G Kees Hovingh; (Amish GLGC) Alan R Shuldiner; (ARIC) Eric Boerwinkle; (B58C-WTCCC &
B58C-T1DGC) David P Strachan; (B58C-Metabochip) Cecilia M Lindgren, Chris Power, Mark I McCarthy; (BLSA) Luigi Ferrucci;
(BRIGHT) Patricia B Munroe; (CARDIOGRAM) Nilesh Samani; (CHS) Bruce M Psaty, Jerome I Rotter; (CLHNS) Alan B Feranil, Karen L
Mohlke, Linda S Adair; (CoLaus) Peter Vollenweider; (CROATIA-Vis) Caroline Hayward, Igor Rudan; (deCODE) Kari Stefansson, Unnur
Thorsteinsdottir; (DIAGEN) Peter EH Schwarz, Stefan R Bornstein; (DILGOM) Samuli Ripatti; (DPS) Matti Uusitupa; (DR's EXTRA)
Rainer Rauramaa; (EAS) Jackie F Price; (EGCUT (Estonian Genome Center of University of Tartu)) Andres Metspalu; (ELY) Nicholas
Wareham; (ENGAGE) Nelson B Freimer; (EPIC) Nicholas Wareham, Kay-Tee Khaw; (EPIC_N_OBSET GWAS) Elizabeth H Young; (ERF)
Cornelia M van Duijn; (ESS (Erasmus Stroke Study)) Peter J Koudstaal; (Family Heart Study FHS) Ingrid B Borecki; (FBPP) Aravinda
Chakravarti, Richard S Cooper, Steven C Hunt; (FENLAND) Ruth Loos, Nicholas Wareham; (FIN-D2D 2007) Antero Kesäniemi, Leena
Moilanen; (FINCAVAS) Mika Kähönen; (Framingham) L Adrienne Cupples, Sekar Kathiresan, Jose M Ordovas; (FRISCII) Agneta
Siegbahn, Lars Wallentin; (FUSION GWAS) Karen Mohlke, Michael Boehnke; (FUSION stage 2) Francis S Collins, Jaakko Tuomilehto,
Jouko Saramies; (GenomEUTwin) John B Whitfield, Nicholas G Martin, Kirsten O Kyvik, Veikko Salomaa, Jaakko Kaprio, Antti Jula,
Dorret I Boomsma, Nancy Pedersen, Tim D Spector; (GLACIER) Paul W Franks, Göran Hallmans; (Go-DARTS) Andrew D Morris, Colin
NA Palmer; (GxE/Spanish Town) Bamidele O Tayo, Colin A McKenzie, Franklyn Bennett, Joel N Hirschhorn, Richard S Cooper;
(HUNT2) Kristian Hveem; (IMPROVE) Ulf de Faire, Anders Hamsten, Elena Tremoli, Steve E Humphries; (InCHIANTI) Stefania
Bandinelli; (KORAF4) Christian Gieger;(LifeLines)Bruce HR Wolffenbuttel; (LOLIPOP) Jaspal S Kooner, John C Chambers; (LURIC)
Bernhard O Boehm; Winfried März; (MDC) Leif C Groop, Sekar Kathiresan; (MEDSTAR) Mary S Burnett, Stephen E Epstein; (METSIM)
Johanna Kuusisto, Markku Laakso; (MICROS) Peter P Pramstaller; (MORGAM) Dominique Arveiler, Jean Ferrières; (MRC/UVRI GPC
GWAS) Pontiano Kaleebu, Gershim Asiki, Janet Seeley, Elizabeth H. Young; (MRC National Survey of Health & Development) Diana
Kuh; (NFBC1986) Marjo-Riitta Jarvelin; (NSPHS) Ulf Gyllensten; (ORCADES) Harry Campbell; (PARC) Yii-Der Ida Chen, Ronald M
Krauss, Jerome I Rotter; (PennCath) Daniel J Rader, Muredach P Reilly; (PIVUS) Erik Ingelsson, Lars Lind; (PROMIS) John Danesh,
Panos Deloukas, Danish Saleheen; (Rotterdam Study) Albert Hofman, Andre G Uitterlinden; (SardiNIA) Gonçalo R Abecasis;
(SCARFSHEEP) Anders Hamsten, Ulf de Faire; (SEYCHELLES) Michel Burnier, Murielle Bochud; Pascal Bovet; (SUVIMAX) Pierre
Meneton; (Swedish Twin Reg.) Erik Ingelsson, Nancy L Pedersen; (TAICHI) Themistocles L Assimes, Yii-Der Ida Chen, Chao Agnes
Hsiung, Thomas Quertermous, Jerome I Rotter, Wayne H-H Sheu; (THISEAS) George Dedoussis, Panos Deloukas; (Tromsø) Inger
Njølstad; (TWINGENE) Ulf de Faire, Erik Ingelsson; (ULSAM) Erik Ingelsson; (Whitehall II) Aroon Hingorani, Mika Kivimaki.
Genotyping of contributing cohorts
(ADVANCE) Devin Absher; (AIDHS/SDS) Latonya F Been, Martha L Gravito; (AMC-PAS) Panos Deloukas, G Kees Hovingh; (B58C-
WTCCC & B58C-T1DGC) Wendy L McArdle; (B58C-Metabochip) Mark I McCarthy; (BLSA) Dena Hernandez; (BRIGHT) Patricia B
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
99
Munroe; (CHS) Jerome I Rotter; (DIAGEN) Narisu Narisu, Gabrielle Müller; (DILGOM) Aarno Palotie; (DR's EXTRA) Timo A Lakka; (EAS)
James F Wilson; (EGCUT (Estonian Genome Center of University of Tartu)) Tõnu Esko; (EPIC) Panos Deloukas; (EPIC_N_SUBCOH
GWAS) Inês Barroso; (ERF) Cornelia M van Duijn; (ESS (Erasmus Stroke Study) ) Cornelia M Van Duijn; (FBPP) Aravinda Chakravarti,
Georg B Ehret; (FENLAND) Manjinder S. Sandhu; (FIN-D2D 2007) Amy J Swift; (FINCAVAS) Terho Lehtimäki; (Framingham) Jose M
Ordovas; (FUSION stage 2) Lori L Bonnycastle; (GLACIER) Inês Barroso; (Go-DARTS) Christopher Groves, Colin NA Palmer, Mark I
McCarthy; (IMPROVE) Anders Hamsten; (KORAF3) Harald Grallert, Thomas Illig; (KORAF4) Norman Klopp; (LifeLines) Cisca Wijmenga;
(LOLIPOP) Jaspal S Kooner, John C Chambers; (LURIC) Marcus E Kleber; (MDC) Benjamin F Voight, Ron Do; (MICROS) Andrew A Hicks;
(MORGAM) Laurence Tiret, Paolo Brambilla; (MRC/UVRI GPC GWAS) Manjinder S. Sandhu; (MRC National Survey of Health &
Development) Andrew Wong, Diana Kuh, Ken K Ong; (NFBC1986) Anna-Liisa Hartikainen, Marjo-Riitta Jarvelin, Mark McCarthy, Paul
Elliott, Sailaja Vedantam; (NSPHS and FRISCII) Åsa Johansson; (ORCADES) Harry Campbell; (PARC) Mark O Goodarzi, Michelle R Jones,
Jerome I Rotter; (PIVUS) Erik Ingelsson, Lars Lind; (PROMIS) Panos Deloukas, Kathleen Stirrups; (Rotterdam Study) Andre G
Uitterlinden, Fernando Rivadeneira; (SardiNIA) Ramaiah Nagaraja; (SCARFSHEEP) Bruna Gigante, Rona J Strawbridge; (SEYCHELLES)
François Mach, Georg B Ehret; (Swedish Twin Reg.) Erik Ingelsson, Nancy L Pedersen; (TAICHI) Devin Absher, Themistocles L Assimes,
Eric Kim, Thomas Quertermous, Lindsay L Waite; (THISEAS) Panos Deloukas; (TWINGENE) Anders Hamsten, Erik Ingelsson; (ULSAM)
Erik Ingelsson; (WGHS) Daniel I. Chasman, Samia Mora, Paul M Ridker; (Whitehall II) Aroon Hingorani, Claudia Langenberg, Meena
Kumari, Mika Kivimaki.
Phenotype definition of contributing cohorts
(ADVANCE) Carlos Iribarren; (AGES Reykjavik study) Tamara B Harris, Vilmundur Gudnason; (AIDHS/SDS) Latonya F Been; (AMC-PAS)
John J.P. Kastelein; (Amish GLGC) Alan R Shuldiner, Braxton D Mitchell; (B58C-WTCCC & B58C-T1DGC) David P Strachan; (B58C-
Metabochip) Chris Power; Elina Hyppönen; (BRIGHT) Patricia B Munroe; (CHS) Bruce M Psaty; (CoLaus) Peter Vollenweider;
(deCODE) Gudmundur Ingi Eyjolfsson, Hilma Holm, Isleifur Olafsson; (DIAGEN) Gabrielle Müller; (DILGOM) Kaisa Silander; (DPS)
Jaana Lindström; (DR's EXTRA) Pirjo Komulainen; (EAS) Jennifer L Bolton; (EGCUT (Estonian Genome Center of University of Tartu))
Andres Metspalu; (EGCUT (Estonian Genome Center of University of Tartu)) Krista Fischer; (ERF and Rotterdam Study) Albert
Hofman; (ERF) Cornelia M van Duijn; (ESS (Erasmus Stroke Study) ) Evita G Van den Herik, Heleen M Den Hertog, Peter J Koudstaal;
(FBPP) Aravinda Chakravarti, Richard S Cooper, Steven C Hunt; (FINCAVAS) Tuomo VM Nieminen; (Framingham) Sekar Kathiresan,
Jose M Ordovas; (GenomEUTwin: MZGWA) John B Whitfield; (GenomEUTwin-FINRISK) Veikko Salomaa; (GenomEUTwin-FINTWIN)
Jaakko Kaprio, Kauko Heikkilä; (GenomEUTwin-GENMETS) Antti Jula; (GenomEUTwin-NLDTWIN) Gonneke Willemsen; (Go-DARTS)
Alex SF Doney, Andrew D Morris, Colin NA Palmer, Louise A Donnelly; (GxE/Spanish Town) Colin A McKenzie, Franklyn Bennett;
(IMPROVE) Ulf de Faire; Anders Hamsten, Elena Tremoli; (KORAF3) Christa Meisinger; (KORAF4) Angela Döring; (LifeLines)L Joost van
Pelt; (LOLIPOP) Jaspal S Kooner, John C Chambers; (LURIC) Hubert Scharnagl; (MDC) Leif C Groop; (METSIM) Alena Stančáková;
(MORGAM) Giancarlo Cesana; (MRC/UVRI GPC GWAS) Rebecca N Nsubuga; (MRC National Survey of Health & Development) Diana
Kuh; (NFBC1986) Aimo Ruokonen, Anna-Liisa Hartikainen, Anneli Pouta, Marjo-Riitta Jarvelin; (NSPHS and FRISCII) Åsa Johansson;
(NSPHS) Ulf Gyllensten; (ORCADES) Sarah H Wild; (PARC) Yii-Der Ida Chen, Ronald M Krauss; (PIVUS) Erik Ingelsson, Lars Lind;
(PROMIS) Daniel F. Freitag; (Rotterdam Study) Albert Hofman; (SCARFSHEEP) Ulf de Faire, Bruna Gigante; (SEYCHELLES) Michel
Burnier, Murielle Bochud, Pascal Bovet; (Swedish Twin Reg.) Erik Ingelsson, Nancy L Pedersen; (TAICHI) Hsing-Yi Chang, Chao Agnes
Hsiung, Yi-Jen Hung, Eric Kim, Shih-Yi Lin, Wayne H-H Sheu; (THISEAS) George Dedoussis, Maria Dimitriou; (Tromsø) Tom Wilsgaard;
(TWINGENE) Ulf de Faire, Erik Ingelsson; (ULSAM) Erik Ingelsson; (WGHS) Paul M Ridker; (Whitehall II) Meena Kumari.
Primary analysis from contributing cohorts
(ADVANCE) Lindsay L Waite; (AIDHS/SDS) Richa Saxena; (AMC-PAS) Stavroula Kanoni; (Amish GLGC) Jeff R O'Connell, May E
Montasser; (ARIC) Kelly A Volcik; (B58C-Metabochip) Cecilia M Lindgren, Elina Hyppönen, Teresa Ferreira; (B58C-WTCCC & B58C-
T1DGC) David P Strachan; (BLSA) Toshiko Tanaka; (BRIGHT) Toby Johnson; (CLHNS) Ying Wu; (CoLaus) Jacques S Beckmann;
(deCODE) Gudmar Thorleifsson; (DIAGEN) Anne U Jackson; (DILGOM) Markus Perola; (EAS) Ross M Fraser; (DPS) Anne U Jackson;
(DR’S EXTRA) Anne U Jackson; (EGCUT (Estonian Genome Center of University of Tartu)) Evelin Mihailov, Krista Fischer, Tõnu Esko;
(ELY) Deepti Gurdasani; (EPIC) Kathy Stirrups, Deepti Gurdasani; (EPIC_N_OBSET GWAS) Elizabeth Young, Claudia Langenberg;
(EPIC_N_SUBCOH GWAS) Nicholas Wainwright; (ERF) Aaron Isaacs; (ESS (Erasmus Stroke Study) ) Cornelia M Van Duijn, Evita G Van
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
100
den Herik; (EUROSPAN) Ayşe Demirkan; (Family Heart Study FHS) Ingrid B Borecki, Mary F Feitosa; (FBPP) Aravinda Chakravarti,
Georg B Ehret; (FENLAND) Theodore Papamarkou, Cristina Pomilla; (FENLAND GWAS) Jing Hua Zhao, Jian'an Luan; (FIN-D2D 2007)
Anne U Jackson; (FINCAVAS) Leo-Pekka Lyytikäinen; (Framingham) L Adrienne Cupples, Gina M Peloso; (FRISCII and NSPHS) Åsa
Johansson; (FUSION stage 2) Tanya M Teslovich; (GenomEUTwin-FINRISK) Johannes Kettunen; (GenomEUTwin-FINTWIN) Kauko
Heikkilä; (GenomEUTwin-GENMETS) Ida Surakka; (GenomEUTwin-SWETWIN) Patrik KE Magnusson; (GenomEUTwin-UK-TWINS)
Massimo Mangino; (GLACIER) Dmitry Shungin; (GLACIER) Paul W Franks; (Go-DARTS) Colin NA Palmer, Louise A Donnelly;
(GxE/Spanish Town) Cameron D Palmer; (HUNT) Anne U Jackson; (IMPROVE) Rona J Strawbridge; (InCHIANTI) Toshiko Tanaka;
(KORAF3) Martina Müller-Nurasyid; (KORAF4) Ann-Kristin Petersen; (LifeLines) Ilja M Nolte; (LOLIPOP) Weihua Zhang; (LURIC)
Marcus E Kleber; (MDC) Benjamin F Voight; (MDC) Pierre Fontanillas, Ron Do; (METSIM) Anne U Jackson; (MRC/UVRI GPC GWAS)
Rebecca N Nsubuga; (MRC National Survey of Health & Development) Andrew Wong, Jian'an Luan; (NFBC1986) Marika Kaakinen, Ida
Surakka, Susan K Service; (NSPHS and FRISCII) Åsa Johansson; (PARC) Xiaohui Li; (PIVUS) Ci Song, Erik Ingelsson; (PROMIS) John
Danesh, Daniel F. Freitag, Kathleen Stirrups; (Rotterdam Study) Aaron Isaacs; (SardiNIA) Carlo Sidore, Jennifer L Bragg-Gresham,
Serena Sanna; (SCARFSHEEP) Rona J Strawbridge; (SEYCHELLES) Georg B Ehret, Murielle Bochud; (SUVIMAX) Toby Johnson; (Swedish
Twin Reg.) Ci Song, Erik Ingelsson; (TAICHI) Devin Absher, Themistocles L Assimes, Hsing-Yi Chang, Mark Goodarzi, Chao Agnes
Hsiung, Thomas Quertermous, Lindsay L Waite; (THISEAS) Stavroula Kanoni; (Tromsø) Anne U Jackson; (TWINGENE) Andrea Ganna,
Erik Ingelsson; (ULSAM) Ci Song, Erik Ingelsson, Stefan Gustafsson; (WGHS) Daniel I. Chasman; (Whitehall II) Sonia Shah.
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
101
Supplementary References
1. Chasman, D.I. et al. Forty-three loci associated with plasma lipoprotein size, concentration, and cholesterol content in genome-wide analysis. PLoS Genet5, e1000730 (2009).
2. Assimes, T.L. et al. Susceptibility locus for clinical and subclinical coronary artery disease at chromosome 9p21 in the multi-ethnic ADVANCE study. Hum Mol Genet17, 2320-8 (2008).
3. Samani, N.J. et al. Large scale association analysis of novel genetic loci for coronary artery disease. Arterioscler Thromb Vasc Biol29, 774-80 (2009).
4. Mitchell, B.D. et al. The genetic response to short-term interventions affecting cardiovascular function: rationale and design of the Heredity and Phenotype Intervention (HAPI) Heart Study. Am Heart J155, 823-8 (2008).
5. Post, W. et al. Determinants of coronary artery and aortic calcification in the Old Order Amish. Circulation115, 717-24 (2007).
6. Power, C. & Elliott, J. Cohort profile: 1958 British birth cohort (National Child Development Study). Int J Epidemiol35, 34-41 (2006).
7. Strachan, D.P. et al. Lifecourse influences on health among British adults: effects of region of residence in childhood and adulthood. Int J Epidemiol36, 522-31 (2007).
8. Atherton, K., Fuller, E., Shepherd, P., Strachan, D.P. & Power, C. Loss and representativeness in a biomedical survey at age 45 years: 1958 British birth cohort. J Epidemiol Community Health62, 216-23 (2008).
9. Kotronen, A. et al. Non-alcoholic and alcoholic fatty liver disease - two diseases of affluence associated with the metabolic syndrome and type 2 diabetes: the FIN-D2D survey. BMC Public Health10, 237 (2010).
10. Helgadottir, A. et al. A common variant on chromosome 9p21 affects the risk of myocardial infarction. Science316, 1491-3 (2007).
11. Schwarz, P.E. et al. Hypoadiponectinemia is associated with progression toward type 2 diabetes and genetic variation in the ADIPOQ gene promoter. Diabetes Care29, 1645-50 (2006).
12. Vartiainen, E. et al. Thirty-five-year trends in cardiovascular risk factors in Finland. Int J Epidemiol39, 504-18 (2010).
13. Tuomilehto, J. et al. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N Engl J Med344, 1343-50 (2001).
14. Kouki, R. et al. Diet, fitness and metabolic syndrome - The DR's EXTRA Study. Nutr Metab Cardiovasc Dis (2010). 15. Fowkes, F.G. et al. Edinburgh Artery Study: prevalence of asymptomatic and symptomatic peripheral arterial
disease in the general population. Int J Epidemiol20, 384-92 (1991). 16. Nelis, M. et al. Genetic structure of Europeans: a view from the North-East. PLoS One4, e5472 (2009). 17. Williams, D.R. et al. Undiagnosed glucose intolerance in the community: the Isle of Ely Diabetes Project. Diabet
Med12, 30-5 (1995). 18. Day, N. et al. EPIC-Norfolk: study design and characteristics of the cohort. European Prospective Investigation of
Cancer. Br J Cancer80 Suppl 1, 95-103 (1999). 19. Harding, A.H. et al. Dietary fat and the risk of clinical type 2 diabetes: the European prospective investigation of
Cancer-Norfolk study. Am J Epidemiol159, 73-82 (2004). 20. Nieminen, T. et al. The Finnish Cardiovascular Study (FINCAVAS): characterising patients with high risk of
cardiovascular morbidity and mortality. BMC Cardiovasc Disord6, 9 (2006). 21. Wallentin, L. et al. Outcome at 1 year after an invasive compared with a non-invasive strategy in unstable
coronary-artery disease: the FRISC II invasive randomised trial. FRISC II Investigators. Fast Revascularisation during Instability in Coronary artery disease. Lancet356, 9-16 (2000).
22. Scott, L.J. et al. A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science316, 1341-5 (2007).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
102
23. Renstrom, F. et al. Genetic predisposition to long-term nondiabetic deteriorations in glucose homeostasis: Ten-year follow-up of the GLACIER study. Diabetes60, 345-54 (2011).
24. Kimber, C.H. et al. TCF7L2 in the Go-DARTS study: evidence for a gene dose effect on both diabetes susceptibility and control of glucose levels. Diabetologia50, 1186-91 (2007).
25. Baldassarre, D. et al. Cross-sectional analysis of baseline data to identify the major determinants of carotid intima-media thickness in a European population: the IMPROVE study. Eur Heart J31, 614-22 (2010).
26. Wichmann, H.E., Gieger, C. & Illig, T. KORA-gen--resource for population genetics, controls and a broad spectrum of disease phenotypes. Gesundheitswesen67 Suppl 1, S26-30 (2005).
27. Winkelmann, B.R. et al. Rationale and design of the LURIC study--a resource for functional genomics, pharmacogenomics and long-term prognosis of cardiovascular disease. Pharmacogenomics2, S1-73 (2001).
28. Berglund, G., Elmstahl, S., Janzon, L. & Larsson, S.A. The Malmo Diet and Cancer Study. Design and feasibility. J Intern Med233, 45-51 (1993).
29. Stancakova, A. et al. Changes in insulin sensitivity and insulin release in relation to glycemia and glucose tolerance in 6,414 Finnish men. Diabetes58, 1212-21 (2009).
30. Wadsworth, M., Kuh, D., Richards, M. & Hardy, R. Cohort Profile: The 1946 National Birth Cohort (MRC National Survey of Health and Development). Int J Epidemiol35, 49-54 (2006).
31. Ingelsson, E., Hulthe, J. & Lind, L. Inflammatory markers in relation to insulin resistance and the metabolic syndrome. Eur J Clin Invest38, 502-9 (2008).
32. Pilia, G. et al. Heritability of cardiovascular and personality traits in 6,148 Sardinians. PLoS Genet2, e132 (2006). 33. Reynolds, C.A. et al. A survey of ABCA1 sequence variation confirms association with dementia. Hum Mutat30,
1348-54 (2009). 34. Theodoraki, E.V. et al. Fibrinogen beta variants confer protection against coronary artery disease in a Greek
case-control study. BMC Med Genet11, 28 (2010). 35. Theodoraki, E.V. et al. ROS1 Asp2213Asn polymorphism is not associated with coronary artery disease in a Greek
case-control study. Clin Chem Lab Med47, 1471-3 (2009). 36. Jacobsen, B.K., Eggen, A.E., Mathiesen, E.B., Wilsgaard, T. & Njolstad, I. Cohort profile: The Tromso Study. Int J
Epidemiol (2011). 37. Ingelsson, E., Sundstrom, J., Arnlov, J., Zethelius, B. & Lind, L. Insulin resistance and risk of congestive heart
failure. JAMA294, 334-41 (2005). 38. Marmot, M. & Brunner, E. Cohort Profile: the Whitehall II study. Int J Epidemiol34, 251-6 (2005). 39. Adair, L.S. et al. Cohort profile: the Cebu longitudinal health and nutrition survey. Int J Epidemiol40, 619-25
(2011). 40. Sanghera, D.K. et al. Impact of nine common type 2 diabetes risk polymorphisms in Asian Indian Sikhs: PPARG2
(Pro12Ala), IGF2BP2, TCF7L2 and FTO variants confer a significant risk. BMC Med Genet9, 59 (2008). 41. Been, L.F. et al. Variants in KCNQ1 increase type II diabetes susceptibility in South Asians: a study of 3,310
subjects from India and the US. BMC Med Genet12, 18 (2011). 42. Hammond, C.J., Andrew, T., Tat Mak, Y. & Spector, T.D. A Susceptibility Locus for Myopia in the Normal
Population Is Linked to the PAX6 Gene Region on Chromosome 11: A Genomewide Scan of Dizygotic Twins. Am J Hum Genet75, 294-304 (2004).
43. Saleheen, D. et al. The Pakistan Risk of Myocardial Infarction Study: a resource for the study of genetic, lifestyle and other determinants of myocardial infarction in South Asia. Eur J Epidemiol24, 329-38 (2009).
44. Multi-center genetic study of hypertension: The Family Blood Pressure Program (FBPP). Hypertension39, 3-9 (2002).
45. Lettre, G. et al. Genome-wide association study of coronary heart disease and its risk factors in 8,090 African Americans: the NHLBI CARe Project. PLoS Genet7, e1001300 (2011).
46. Bochud, M. et al. Heritability of renal function in hypertensive families of African descent in the Seychelles (Indian Ocean). Kidney Int67, 61-9 (2005).
47. Cooper, R. et al. The prevalence of hypertension in seven populations of west African origin. Am J Public Health87, 160-8 (1997).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
103
48. Teslovich, T.M. et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature466, 707-13 (2010).
49. Simon, J.A. et al. Phenotypic predictors of response to simvastatin therapy among African-Americans and Caucasians: the Cholesterol and Pharmacogenetics (CAP) Study. Am J Cardiol97, 843-50 (2006).
50. Evans, A. et al. MORGAM (an international pooling of cardiovascular cohorts). Int J Epidemiol34, 21-7 (2005). 51. Henkel, A.S. et al. Hepatic overexpression of abcb11 promotes hypercholesterolemia and obesity in mice.
Gastroenterology141, 1404-11, 1411 e1-2 (2011). 52. Chambers, J.C. et al. Genome-wide association study identifies loci influencing concentrations of liver enzymes
in plasma. Nature Genetics43, 1131-8 (2011). 53. Chen, W.M. et al. Variations in the G6PC2/ABCB11 genomic region are associated with fasting glucose levels.
Journal of Clinical Investigation118, 2620-8 (2008). 54. Ito, J., Li, H., Nagayasu, Y., Kheirollah, A. & Yokoyama, S. Apolipoprotein A-I induces translocation of protein
kinase C[alpha] to a cytosolic lipid-protein particle in astrocytes. Journal of Lipid Research45, 2269-76 (2004). 55. Ha, V.L. et al. ASAP3 is a focal adhesion-associated Arf GAP that functions in cell migration and invasion. Journal
of Biological Chemistry283, 14915-26 (2008). 56. Pare, G. et al. Novel associations of CPS1, MUT, NOX4, and DPEP1 with plasma homocysteine in a healthy
population: a genome-wide evaluation of 13 974 participants in the Women's Genome Health Study. Circ Cardiovasc Genet2, 142-50 (2009).
57. Danik, J.S. et al. Novel loci, including those related to Crohn disease, psoriasis, and inflammation, identified in a genome-wide association study of fibrinogen in 17 686 women: the Women's Genome Health Study. Circ Cardiovasc Genet2, 134-41 (2009).
58. Bisogno, T. et al. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. Journal of Cell Biology163, 463-8 (2003).
59. Kim, Y. et al. A conserved phosphatase cascade that regulates nuclear membrane biogenesis. Proceedings of the National Academy of Sciences of the United States of America104, 6596-601 (2007).
60. Denis, N. et al. Quantitative proteomic analysis of PCSK9 gain of function in human hepatic HuH7 cells. J Proteome Res10, 2011-26 (2011).
61. Rohwedder, I. et al. Plasma fibronectin deficiency impedes atherosclerosis progression and fibrous cap formation. EMBO Mol Med4, 564-76 (2012).
62. Frayling, T.M. et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science316, 889-94 (2007).
63. Fischer, J. et al. Inactivation of the Fto gene protects from obesity. Nature458, 894-8 (2009). 64. Liu, Y. et al. Conditional ablation of Gsk-3beta in islet beta cells results in expanded mass and resistance to fat
feeding-induced diabetes in mice. Diabetologia53, 2600-10 (2010). 65. Wallrapp, C. et al. The product of the mammalian orthologue of the Saccharomyces cerevisiae HBS1 gene is
phylogenetically related to eukaryotic release factor 3 (eRF3) but does not carry eRF3-like activity. FEBS Letters440, 387-92 (1998).
66. Uda, M. et al. Genome-wide association study shows BCL11A associated with persistent fetal hemoglobin and amelioration of the phenotype of beta-thalassemia. Proceedings of the National Academy of Sciences of the United States of America105, 1620-5 (2008).
67. Soranzo, N. et al. A genome-wide meta-analysis identifies 22 loci associated with eight hematological parameters in the HaemGen consortium. Nature Genetics41, 1182-90 (2009).
68. Majumdar, A., Petrescu, A.D., Xiong, Y. & Noy, N. Nuclear translocation of cellular retinoic acid-binding protein II is regulated by retinoic acid-controlled SUMOylation. Journal of Biological Chemistry286, 42749-57 (2011).
69. Loeper, S., Asa, S.L. & Ezzat, S. Ikaros modulates cholesterol uptake: a link between tumor suppression and differentiation. Cancer Research68, 3715-23 (2008).
70. van Beekum, O. et al. The adipogenic acetyltransferase Tip60 targets activation function 1 of peroxisome proliferator-activated receptor gamma. Endocrinology149, 1840-9 (2008).
Nature Genetics: doi:10.1038/ng.2797

GLGC Metabochip Supplement 2013
104
71. Cho, Y.S. et al. Meta-analysis of genome-wide association studies identifies eight new loci for type 2 diabetes in east Asians. Nature Genetics44, 67-72 (2012).
72. Yu, X. et al. Hepatocyte growth factor protects endothelial progenitor cell from damage of low-density lipoprotein cholesterol via the PI3K/Akt signaling pathway. Molecular Biology Reports37, 2423-9 (2010).
73. Zou, C. et al. Lack of Fas antagonism by Met in human fatty liver disease. Nature Medicine13, 1078-85 (2007). 74. Zhou, X. et al. Altered expression of miR-152 and miR-148a in ovarian cancer is related to cell proliferation.
Oncology Reports27, 447-54 (2012). 75. Zheng, B. et al. MicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in
gastric cancer. Clinical Cancer Research17, 7574-83 (2011). 76. Cao, J. et al. A predominant role of acyl-CoA:monoacylglycerol acyltransferase-2 in dietary fat absorption
implicated by tissue distribution, subcellular localization, and up-regulation by high fat diet. Journal of Biological Chemistry279, 18878-86 (2004).
77. Demirkan, A. et al. Genome-wide association study identifies novel loci associated with circulating phospho- and sphingolipid concentrations. PLoS Genet8, e1002490 (2012).
78. Dastani, Z. et al. Novel loci for adiponectin levels and their influence on type 2 diabetes and metabolic traits: a multi-ethnic meta-analysis of 45,891 individuals. PLoS Genet8, e1002607 (2012).
79. Galliano, M.F. et al. Binding of alpha2ML1 to the low density lipoprotein receptor-related protein 1 (LRP1) reveals a new role for LRP1 in the human epidermis. PLoS ONE3, e2729 (2008).
80. Zhou, Q.L. et al. A novel pleckstrin homology domain-containing protein enhances insulin-stimulated Akt phosphorylation and GLUT4 translocation in adipocytes. Journal of Biological Chemistry285, 27581-9 (2010).
81. Takeuchi, H., Takeuchi, T., Gao, J., Cantley, L.C. & Hirata, M. Characterization of PXK as a protein involved in epidermal growth factor receptor trafficking. Molecular and Cellular Biology30, 1689-702 (2010).
82. Kazanskaya, O. et al. The Wnt signaling regulator R-spondin 3 promotes angioblast and vascular development. Development135, 3655-64 (2008).
83. Heid, I.M. et al. Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nature Genetics42, 949-60 (2010).
84. Duncan, E.L. et al. Genome-wide association study using extreme truncate selection identifies novel genes affecting bone mineral density and fracture risk. PLoS Genet7, e1001372 (2011).
85. Kim, Y.J. et al. Large-scale genome-wide association studies in East Asians identify new genetic loci influencing metabolic traits. Nature Genetics43, 990-5 (2011).
86. Koharudin, L.M., Furey, W., Liu, H., Liu, Y.J. & Gronenborn, A.M. The phox domain of sorting nexin 5 lacks phosphatidylinositol 3-phosphate (PtdIns(3)P) specificity and preferentially binds to phosphatidylinositol 4,5-bisphosphate (PtdIns(4,5)P2). Journal of Biological Chemistry284, 23697-707 (2009).
87. Hicks, A.A. et al. Genetic determinants of circulating sphingolipid concentrations in European populations. PLoS Genet5, e1000672 (2009).
88. Li, R. et al. Role of liver sinusoidal endothelial cells and stabilins in elimination of oxidized low-density lipoproteins. Am J Physiol Gastrointest Liver Physiol300, G71-81 (2011).
89. Ishikawa, K., Navab, M. & Lusis, A.J. Vasculitis, Atherosclerosis, and Altered HDL Composition in Heme-Oxygenase-1-Knockout Mice. Int J Hypertens2012, 948203 (2012).
90. Sarria, A.J., Panini, S.R. & Evans, R.M. A functional role for vimentin intermediate filaments in the metabolism of lipoprotein-derived cholesterol in human SW-13 cells. Journal of Biological Chemistry267, 19455-63 (1992).
91. Wyne, K.L., Pathak, K., Seabra, M.C. & Hobbs, H.H. Expression of the VLDL receptor in endothelial cells. Arteriosclerosis, Thrombosis, and Vascular Biology16, 407-15 (1996).
92. Ding, L. et al. Akt3 deficiency in macrophages promotes foam cell formation and atherosclerosis in mice. Cell Metab15, 861-72 (2012).
Nature Genetics: doi:10.1038/ng.2797