direct demonstration of a neonatal fc receptor (fcrn)-driven … · sensitive alexa488-labeled...
TRANSCRIPT

Direct demonstration of a neonatal Fc receptor (FcRn)-drivenendosomal sorting pathway for cellular recycling of albuminReceived for publication, May 19, 2017, and in revised form, June 14, 2017 Published, Papers in Press, June 21, 2017, DOI 10.1074/jbc.M117.794248
Esben G. W. Schmidt‡1, Michael L. Hvam§1, Filipa Antunes¶, Jason Cameron¶, Dorthe Viuff‡, Birgitte Andersen‡,Nanna N. Kristensen‡, and Kenneth A. Howard§2
From the ‡Novozymes A/S, 2880 Bagsværd, Denmark, §Interdisciplinary Nanoscience Center (iNANO), Department of MolecularBiology and Genetics, Aarhus University, 8000 Aarhus C, Denmark, and ¶Albumedix Ltd., NG7 1FD Nottingham, United Kingdom
Edited by Luke O’Neill
Albumin is the most abundant plasma protein involved in thetransport of many compounds, such as fatty acids, bilirubin, andheme. The endothelial cellular neonatal Fc receptor (FcRn) hasbeen suggested to play a central role in maintaining high albu-min plasma levels through a cellular recycling pathway. How-ever, direct mapping of this process is still lacking. This workpresents the use of wild-type and engineered recombinant albu-mins with either decreased or increased FcRn affinity in combi-nation with a low or high FcRn-expressing endothelium cell lineto clearly define the FcRn involvement, intracellular pathway,and kinetics of albumin trafficking by flow cytometry, quantita-tive confocal microscopy, and an albumin-recycling assay. Wefound that cellular albumin internalization was proportional toFcRn expression and albumin-binding affinity. Albumin accu-mulation in early endosomes was independent of FcRn-bindingaffinity, but differences in FcRn-binding affinities significantlyaffected the albumin distribution between late endosomes andlysosomes. Unlike albumin with low FcRn-binding affinity,albumin with high FcRn-binding affinity was directed less to thelysosomes, suggestive of FcRn-directed albumin salvage fromlysosomal degradation. Furthermore, the amount of recycledalbumin in cell culture media corresponded to FcRn-bindingaffinity, with a �3.3-fold increase after 1 h for the high FcRn-binding albumin variant compared with wild-type albumin.Together, these findings uncover an FcRn-dependent endo-somal cellular-sorting pathway that has great importance indescribing fundamental mechanisms of intracellular albuminrecycling and the possibility to tune albumin-based therapeuticeffects by FcRn-binding affinity.
Human serum albumin (HSA)3 is the most abundant plasmaprotein involved in transport of a wide range of compoundssuch as fatty acids, bilirubin, and heme, facilitated by multiple
ligand-binding sites and an extended circulatory half-life (1).The binding and transport of endogenous ligands is vital tohuman health; however, a detailed understanding of the intra-cellular route taken by albumin is surprisingly still lacking.
Reports suggest engagement with the cellular recycling neo-natal Fc receptor (FcRn) diverts albumin from lysosomal deg-radation by an endosomal recycling pathway (2). FcRn com-prises a type I transmembrane MHC class I-related heavy �chain that non-covalently associates with a �2-microglobulinlight chain, responsible for Immunoglobulin G (IgG) diversionfrom lysosomal degradation by an endosomal cellular reroutingpathway after FcRn interaction (3). IgG is taken into the cell bypinocytosis and processed within endosomes at a low pH envi-ronment that triggers the binding to FcRn and consequenttransport from the cell either by a transcytosis or recyclingroute dependent on the cell polarized state (4). Exposure tophysiological extracellular pH triggers ligand release into theextracellular milieu (5). Although well described for IgG, onlyindirect evidence is available to apply this recycling pathway toalbumin. The serum level of albumin in mice genetically mod-ified to lack the FcRn expression has been found to be 2–3-foldlower than in a wild-type mice counterpart (2). Furthermore,Andersen et al. (6) found that recombinant human albuminvariants engineered for enhanced FcRn-binding increases theblood circulatory half-life in mice and non-human primates (7).
This work presents the use of wild-type (WT) and engineeredrecombinant albumins with decreased and increased affinity toFcRn in combination with a low and high FcRn-expressing der-mal human microvascular endothelium cell line (HMEC-1 andHMEC-1-FcRn, respectively) to define the role of FcRn in inter-nalization, sorting, and rescuing of albumin from intracellulardegradation. Intracellular trafficking of fluorescent-labeledalbumins is investigated using a combination of confocal andflow cytometric techniques. Furthermore, a recycling assay isused for quantitative dynamic measurement of albumin recy-cling. The findings in this work provide the first direct evidenceshowing an FcRn-dependent endosomal recycling route that isvital to understand albumin’s pivotal role in endogenous ligandtransport and utilization to tune the pharmacokinetics of albu-min-based therapeutics.
Results
Cellular uptake studies were performed in isogenic humanendothelial cells derived from dermal microvasculature (HMEC-1)exhibiting a low (HMEC-1) or high expression (HMEC-1-
The authors declare that they have no conflicts of interest with the contentsof this article.
This article contains supplemental Table S1 and Figs. S1–S5.1 Both authors contributed equally to this work.2 To whom correspondence should be addressed. Tel.: 45-87155831; E-mail:
[email protected] The abbreviations used are: HSA, human serum albumin; FcRn, neonatal Fc
receptor; hFcRn, human FcRn; HMEC, human endothelial cell; qPCR, quan-titative PCR; LB, low binder; HB, high binder; 5FAM, 5-carboxyfluorescein;WFI, water for injection; HBSS, Hanks’ balanced salt solution; LAMP1, lyso-somal-associated membrane protein 1; EEA1, early endosomal antigen-1;RAB7A, Ras-related protein Rab-7a; CF, correction factor.
croARTICLE
13312 J. Biol. Chem. (2017) 292(32) 13312–13322
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

FcRn) of human FcRn that was demonstrated by quantitativePCR (qPCR; 8 –9-fold increase) and Western analysis (supple-mental Fig. 1). Albumins with different FcRn affinity provided atool to specifically investigate the role of FcRn in uptake andtrafficking. FcRn affinity measured by Biolayer Interferometryshowed a 15-fold decrease in affinity for the FcRn low-binding(LB) variant and a 24-fold increase for the FcRn high-binding(HB) variant compared with the WT variant at pH 5.5 (Table 1).Attachment of 5-carboxyfluorescein (5FAM) yielded a slightincrease in binding affinity for WT and LB and a slight reduc-tion for HB (�2.2-, �0.9-, �1.9-fold change for WT, HB, andLB compared with the unlabeled variant, respectively). A simi-lar tendency was observed for Alexa594 (�3.9-, �0.1-, �4.2-fold change for WT, HB, and LB, compared with the unlabeledvariant, respectively). Attachment of Alexa488, however, gen-erally resulted in a decreased FcRn affinity (�0.6-, �0.6-, �0.7-fold change for WT, HB, and LB, compared with the unlabeledvariant, respectively). However, with either of the fluorescenttags, the HB retained significantly higher FcRn-binding affini-ties than WT, and LB exhibits significantly lower affinities toFcRn than WT. The labeling efficiency, determined by the ratioof absorbance from the fluorophore and albumin for the 5FAMLB, WT, and HB was shown to be 1.6, 1.6, and 1.0, for theAlexa488 was 1.2, 1.3, and 1.0, and for the Alexa594 was 1.5, 1.0,and 1.0 (see supplemental Table 1).
Flow cytometric analysis was used to measure the amount ofretained fluorescence in HMEC-1 and HMEC-1-FcRn cellsafter 2 h of exposure to 8 �M Alexa488-labeled albumin (Fig. 1).A greater percentage of albumin was retained within the cellsfor the LB than the WT and HB after 1 h (78%, 46%, and 42%,respectively), and 2 h (53%, 21%, and 21%) in the HMEC-1 cells(Fig. 1, a– c). A similar amount of albumin was retained withinthe HMEC-1-FcRn cells for the LB; however, less WT and HBwas retained within these cells at 1 h (80% LB, 23% WT, and 23%HB) and 2 h (55% LB, 15% WT, and 19% HB). Overall, similarlevels for LB were observed between the two cell types, whereasthe WT and HB were seemingly ejected from the cells more
rapidly from the HMEC-1-FcRn cells (Fig. 1, a– c), suggestive ofFcRn-mediated recycling.
To further confirm FcRn-driven uptake and trafficking ofalbumin, flow cytometric uptake studies were performed at alower pH (pH 6.0), known to facilitate increased FcRn engage-ment (2) and demonstrated at pH 5.5 in Table 1. For theHMEC-1 cells, no significant differences in albumin uptakewere observed between the three variants at both pH 6.0 and 7.4(Fig. 1d). Albumin uptake in the HMEC-1-FcRn cells, however,showed an FcRn affinity-dependent uptake that was potenti-ated at pH 6.0 for WT and HB (Fig. 1e). Possible surface bindingof albumin to FcRn in combination with retained albumin-FcRn-binding after recycling could explain the higher level ofuptake for the WT and HB. At pH 7.4, only slight differencesbetween the cellular fluorescence were observed between thevariants in the HMEC-1-FcRn cells, although slightly highervalues for the HB and WT compared with LB (14.2, 7.9, and5.9, respectively). Much greater differences, however, wereobserved at pH 6.0 in the HMEC-1-FcRn cells, with the LBremaining constant at 4.9, whereas the HB and WT increased3-fold (43.7 and 23.7, respectively). A trypan blue quenchingassay at 4 °C and 37 °C was used to show the absence of surface-bound fluorescence and that cellular fluorescence was mostprobably due to internalized albumin (supplemental Fig. 2, band c).
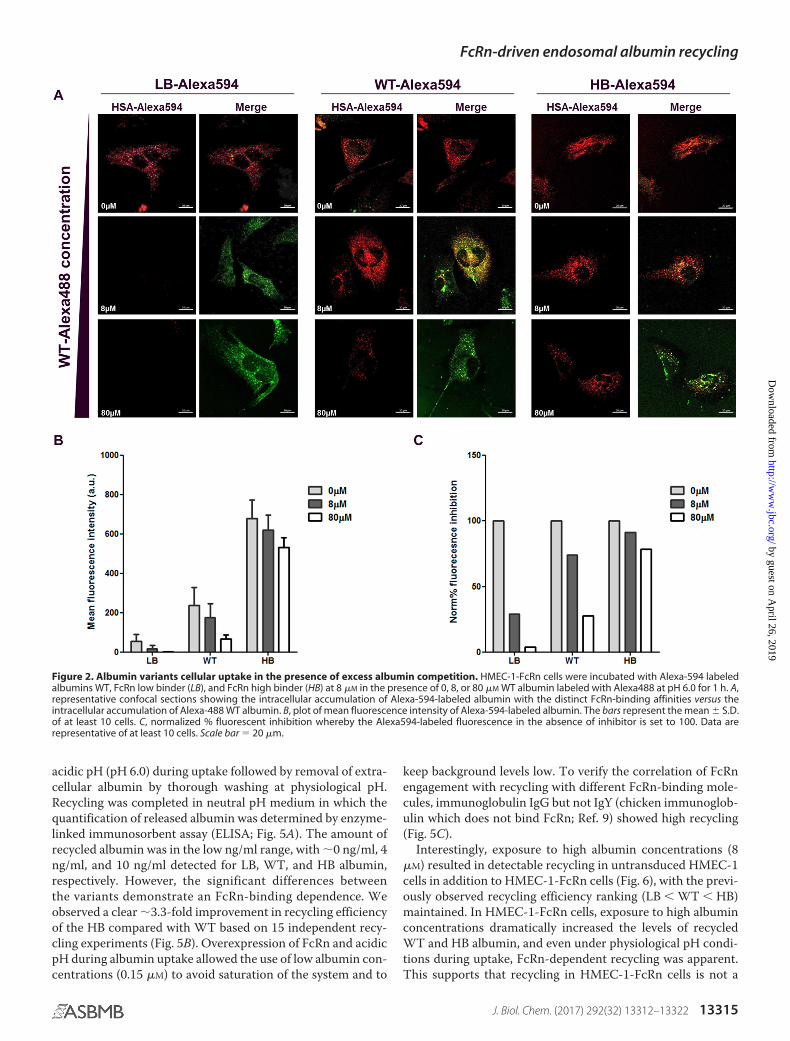
Confocal microscopy was used to further explore the FcRn–albumin affinity-dependent uptake in HMEC-1-FcRn cells.Cells were exposed to Alexa594-labeled LB, WT, and HB albu-mins at a concentration of 8 �M in the presence of equimolar or10-fold excess of Alexa488-labeled WT albumin for 1 h fol-lowed by fixation. Fixed cells were imaged, and the fluorescenceof Alexa594-labeled material was quantified. Again, the intra-cellular fluorescence increased proportionally to FcRn-bindingaffinity (Fig. 2, A and B). The relative drop in intracellular con-centration was lower with increased FcRn-binding affinity (Fig.2C). The HB variant resisted the challenge with only an �20%drop in the presence of 10-fold excess of competing WT albu-min, whereas the LB was nearly 100% blocked. Higher affinityalbumins likely occupy the receptors excluding variants withreduced affinity.
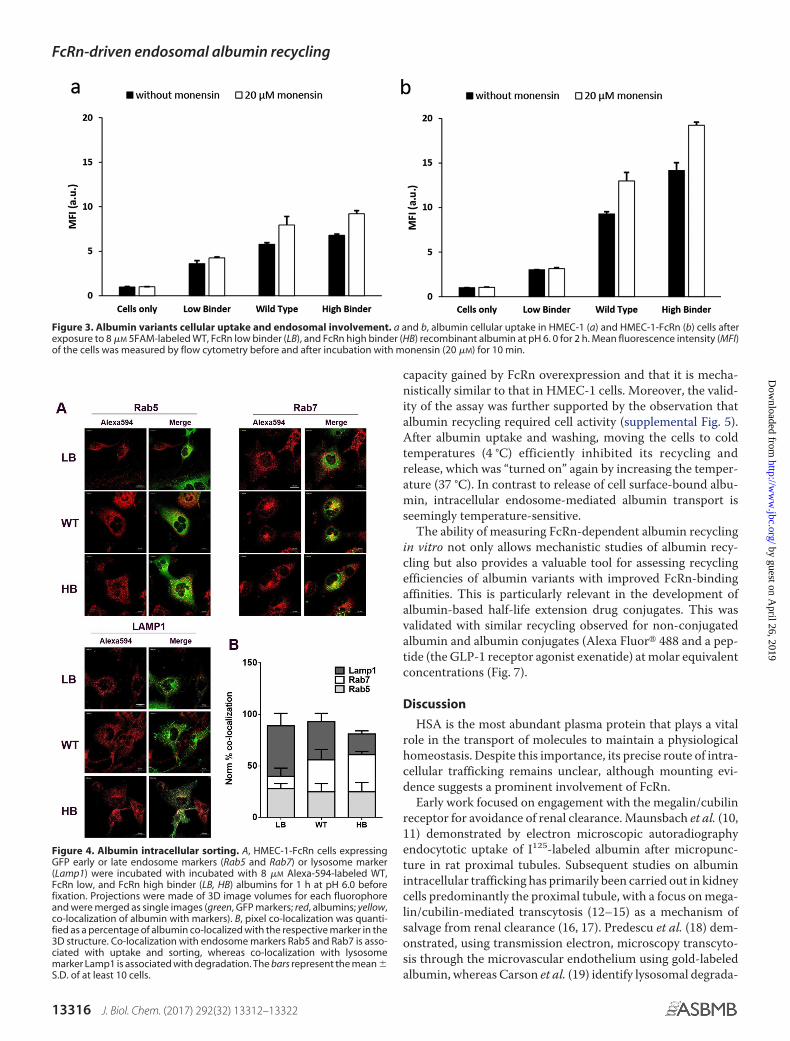
To investigate the involvement of endosomes in the albuminintracellular trafficking process as a consequence of FcRn-bind-ing, fluorescent albumins labeled with the 5FAM were used.The assay is based on the spectral shift and consequent decreasein 5FAM intensity in the endosomal pH range (supplementalFig. 2a), which is restored by cellular addition of the ionophoremonensin that equilibrates the pH gradient between the cyto-plasm and acidic compartments by triggering the exchange ofprotons for potassium ions (8). HMEC-1 and HMEC-1-FcRncells were exposed to 8 �M 5FAM-labeled LB, WT, and HB for2 h followed by assessment of the fluorescence uptake by flowcytometric analysis before and after incubation with monensin(20 �M) for 10 min at room temperature. In HMEC-1 cells, theaddition of monensin resulted in an increase in cellular fluores-cence of 24.4%, 44.8%, and 41.8% for LB, WT, and HB, respec-tively, whereas in HMEC-1-FcRn cells the increase was 4.2%,44.0%, and 38.2% (Fig. 3). No significant differences wereobserved after the addition of monensin using the non-pH-
Table 1Binding of conjugated and non-conjugated albumin variants againsthuman FcRn at pH 5.5 and 7.0Binding kinetics of recombinant albumin WT and FcRn low and high binder albu-min (LB, HB) non-labeled or labeled with the different fluorophores 5FAM, Alex-aFluor488, or AlexaFluor594 against the human FcRn measured at pH 5.5 or pH 7.0.The KD values are the average of three-six measurements, and each measurementconsists of a seven-step dilution series for evaluation of kinetic parameters. NBdenotes no detectable binding.
Compound
pH 5.5
pH 7.0 KD
KDaverage S.D.
FcRn affinity(-fold change comparedto the WT counterpart)
nM nM
Unlabeled WT 633.0 97.0 NB5FAM-WT 293.2 20.9ALEXA488-WT 1157.8 188.9ALEXA594-WT 161.6 25.3Unlabeled HB 26.3 3.7 � 24.1 NB5FAM-HB 30.1 0.6 � 9.7ALEXA488-HB 42.6 2.3 � 27.2ALEXA594-HB 24.1 1.3 � 6.7Unlabeled LB 9633.3 1439.9 �15.2 NB5FAM-LB 4978.0 289.5 �17.0ALEXA488-LB 13126.0 3875.9 �11.3ALEXA594-LB 2276.7 515.8 �14.1
FcRn-driven endosomal albumin recycling
J. Biol. Chem. (2017) 292(32) 13312–13322 13313
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

sensitive Alexa488-labeled variants (supplemental Fig. 2, a, d,and e). The greater fluorescence increase for the WT and HBcompared with LB in HMEC-1-FcRn cells after monensin addi-tion suggests higher endosomal localization facilitated by thehigher FcRn affinity.
Confocal microscopy was used to directly detect endosomalFcRn-driven compartmentalization sorting of Alexa594-la-beled albumin variants after 1 h of incubation in HMEC-1-FcRncells (Fig. 4). The spatial location of trafficking was determinedby colocalization of the labeled albumins with each of the greenfluorescent protein (GFP)-fused markers, Rab5 for early endo-somes, Rab7 for late endosomes, and lysosomal-associatedmembrane protein 1 (LAMP1) for lysosomes. Punctuate fluo-rescence indicative of localization within vesicular compart-ments was observed for the LB, WT, and HB (Fig. 4A). Relativequantification of co-localization with the endosomal/lysosomalmarkers shows that all three albumins share an equal preva-lence in early endosomes (Fig. 4B). For the HB albumin,increased co-localization with late endosomal marker RAB7was observed with minimal presence in the lysosomes, whichsuggests cellular sorting and salvage from lysosomal degrada-tion. In contrast, the LB was highly co-localized with LAMP1lysosomal marker, which suggests subsequent trafficking to alysosomal degradation environment. WT showed a somewhat
intermediate trafficking with its presence in late endosomescomparable with HB but increased accumulation in lysosomes.To further confirm that the distinct compartmentalization wasFcRn-binding– dependent, dextran co-localization with lateendosomes and lysosomes was studied 30 – 60 min afterremoval of dextran. Dextran, as expected, was found almostexclusively within the lysosome compartment (supplementalFig. 3). Together, this supports endosomal trafficking of albu-min with subsequent FcRn-mediated diversion from lysosomaldegradation. In parallel to the GFP-marker transfected cellmethod, endosomal localization in HMEC-1-FcRn cells wasalso observed by immunofluorescence staining in permeabi-lized cells, where almost no albumin was found to localizewithin the lysosomes for the WT and HB (supplemental Fig.4). Both methods indicate the same overall trafficking of thealbumin, and the slight deviations may be a product of cel-lular compartment labeling methods. Having demonstratedthe predominant role of FcRn in albumin recycling and theunderlying intracellular transport route, we aimed at estab-lishing an in vitro assay to quantitate the albumin recyclingdynamics.
FcRn overexpressing HMEC-1-FcRn cells were seeded inculture plates and allowed to reach confluency before the assay.The cells were exposed to either LB, WT, or HB albumin at
Figure 1. Albumin variants cellular uptake. a– c, mean fluorescence intensity (MFI) detected by flow cytometry of low (HMEC-1) and high (HMEC-1-FcRn)FcRn-expressing cells after exposure to Alexa488-labeled recombinant albumin variants. Shown are FcRn low binder (LB) (a), WT (b), and FcRn high binder (HB)(c), at pH 6.0 for 2 h followed by incubation in HBSS (pH 7.4) for 0, 1, or 2 h. MFI was normalized to cells at t � 0 h for each albumin variant. Albumin uptake inHMEC-1 (d) and HMEC-1-FcRn (e) cells after exposure to 8 �M Alexa488-labeled LB, WT, and HB recombinant albumin variants at pH 6.0 or pH 7.4 for 2 h. CellularMFI was determined by flow cytometry and normalized to non-treated cells.
FcRn-driven endosomal albumin recycling
13314 J. Biol. Chem. (2017) 292(32) 13312–13322
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from
acidic pH (pH 6.0) during uptake followed by removal of extra-cellular albumin by thorough washing at physiological pH.Recycling was completed in neutral pH medium in which thequantification of released albumin was determined by enzyme-linked immunosorbent assay (ELISA; Fig. 5A). The amount ofrecycled albumin was in the low ng/ml range, with �0 ng/ml, 4ng/ml, and 10 ng/ml detected for LB, WT, and HB albumin,respectively. However, the significant differences betweenthe variants demonstrate an FcRn-binding dependence. Weobserved a clear �3.3-fold improvement in recycling efficiencyof the HB compared with WT based on 15 independent recy-cling experiments (Fig. 5B). Overexpression of FcRn and acidicpH during albumin uptake allowed the use of low albumin con-centrations (0.15 �M) to avoid saturation of the system and to
keep background levels low. To verify the correlation of FcRnengagement with recycling with different FcRn-binding mole-cules, immunoglobulin IgG but not IgY (chicken immunoglob-ulin which does not bind FcRn; Ref. 9) showed high recycling(Fig. 5C).
Interestingly, exposure to high albumin concentrations (8�M) resulted in detectable recycling in untransduced HMEC-1cells in addition to HMEC-1-FcRn cells (Fig. 6), with the previ-ously observed recycling efficiency ranking (LB � WT � HB)maintained. In HMEC-1-FcRn cells, exposure to high albuminconcentrations dramatically increased the levels of recycledWT and HB albumin, and even under physiological pH condi-tions during uptake, FcRn-dependent recycling was apparent.This supports that recycling in HMEC-1-FcRn cells is not a
Figure 2. Albumin variants cellular uptake in the presence of excess albumin competition. HMEC-1-FcRn cells were incubated with Alexa-594 labeledalbumins WT, FcRn low binder (LB), and FcRn high binder (HB) at 8 �M in the presence of 0, 8, or 80 �M WT albumin labeled with Alexa488 at pH 6.0 for 1 h. A,representative confocal sections showing the intracellular accumulation of Alexa-594-labeled albumin with the distinct FcRn-binding affinities versus theintracellular accumulation of Alexa-488 WT albumin. B, plot of mean fluorescence intensity of Alexa-594-labeled albumin. The bars represent the mean � S.D.of at least 10 cells. C, normalized % fluorescent inhibition whereby the Alexa594-labeled fluorescence in the absence of inhibitor is set to 100. Data arerepresentative of at least 10 cells. Scale bar � 20 �m.
FcRn-driven endosomal albumin recycling
J. Biol. Chem. (2017) 292(32) 13312–13322 13315
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from
capacity gained by FcRn overexpression and that it is mecha-nistically similar to that in HMEC-1 cells. Moreover, the valid-ity of the assay was further supported by the observation thatalbumin recycling required cell activity (supplemental Fig. 5).After albumin uptake and washing, moving the cells to coldtemperatures (4 °C) efficiently inhibited its recycling andrelease, which was “turned on” again by increasing the temper-ature (37 °C). In contrast to release of cell surface-bound albu-min, intracellular endosome-mediated albumin transport isseemingly temperature-sensitive.
The ability of measuring FcRn-dependent albumin recyclingin vitro not only allows mechanistic studies of albumin recy-cling but also provides a valuable tool for assessing recyclingefficiencies of albumin variants with improved FcRn-bindingaffinities. This is particularly relevant in the development ofalbumin-based half-life extension drug conjugates. This wasvalidated with similar recycling observed for non-conjugatedalbumin and albumin conjugates (Alexa Fluor� 488 and a pep-tide (the GLP-1 receptor agonist exenatide) at molar equivalentconcentrations (Fig. 7).
Discussion
HSA is the most abundant plasma protein that plays a vitalrole in the transport of molecules to maintain a physiologicalhomeostasis. Despite this importance, its precise route of intra-cellular trafficking remains unclear, although mounting evi-dence suggests a prominent involvement of FcRn.
Early work focused on engagement with the megalin/cubilinreceptor for avoidance of renal clearance. Maunsbach et al. (10,11) demonstrated by electron microscopic autoradiographyendocytotic uptake of I125-labeled albumin after micropunc-ture in rat proximal tubules. Subsequent studies on albuminintracellular trafficking has primarily been carried out in kidneycells predominantly the proximal tubule, with a focus on mega-lin/cubilin-mediated transcytosis (12–15) as a mechanism ofsalvage from renal clearance (16, 17). Predescu et al. (18) dem-onstrated, using transmission electron, microscopy transcyto-sis through the microvascular endothelium using gold-labeledalbumin, whereas Carson et al. (19) identify lysosomal degrada-
Figure 4. Albumin intracellular sorting. A, HMEC-1-FcRn cells expressingGFP early or late endosome markers (Rab5 and Rab7) or lysosome marker(Lamp1) were incubated with incubated with 8 �M Alexa-594-labeled WT,FcRn low, and FcRn high binder (LB, HB) albumins for 1 h at pH 6.0 beforefixation. Projections were made of 3D image volumes for each fluorophoreand were merged as single images (green, GFP markers; red, albumins; yellow,co-localization of albumin with markers). B, pixel co-localization was quanti-fied as a percentage of albumin co-localized with the respective marker in the3D structure. Co-localization with endosome markers Rab5 and Rab7 is asso-ciated with uptake and sorting, whereas co-localization with lysosomemarker Lamp1 is associated with degradation. The bars represent the mean �S.D. of at least 10 cells.
Figure 3. Albumin variants cellular uptake and endosomal involvement. a and b, albumin cellular uptake in HMEC-1 (a) and HMEC-1-FcRn (b) cells afterexposure to 8 �M 5FAM-labeled WT, FcRn low binder (LB), and FcRn high binder (HB) recombinant albumin at pH 6. 0 for 2 h. Mean fluorescence intensity (MFI)of the cells was measured by flow cytometry before and after incubation with monensin (20 �M) for 10 min.
FcRn-driven endosomal albumin recycling
13316 J. Biol. Chem. (2017) 292(32) 13312–13322
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

tion of albumin within podocytes (19). Recent experimentaldata implicates a role for FcRn in the intracellular trafficking ofalbumin (14, 20). The intracellular FcRn-mediated albuminrecycling pathway has not been studied in cells, but due to non-competitive shared FcRn-binding (21, 22) was thought to followthe well described route of IgG (4, 23–26). The recycling andsalvage from lysosomal degradation of albumin by a singularreceptor was originally proposed by Schultze and Heremans(27) as it was for IgG by Brambell (28). More recently, engi-neered recombinant variants of human albumin with enhancedFcRn-binding (6) were found to increase the blood circulatoryhalf-life in mice and non-human primates thought as a conse-quence of FcRn recycling (7).
Although FcRn expression has been demonstrated in varioustissues, the microvascular endothelium is believed to be themain site of FcRn-mediated IgG recycling (29, 30). In this studywe, therefore, used isogenic human dermal microvascularendothelial (HMEC-1) cells with a low (HMEC-1) or high(HMEC-1-FcRn) FcRn-expression. Furthermore, we used apanel of recombinant human serum albumin variants (non-fluorescent or fluorescent-labeled) with different FcRn-bind-ing affinities, engineered by single-point mutations in theC-terminal domain III, which is known to represent the majorFcRn-binding site within albumin (7). An in vitro cell-based
albumin recycling assay combined with flow cytometric andconfocal microscopic techniques allows precise investigationsinto the role of FcRn-binding in albumin uptake and intracel-lular trafficking. The application of the HMEC-1-FcRn linewith the isogenic parental cell line HMEC-1 should allow anydifferences in albumin trafficking to be attributed to FcRnexpression rather than intercellular differences. The HMEC-1-FcRn line, but not the HMEC-1 cell line, however, was main-tained under puromycin and G418 antibiotic selection condi-tions to maintain stable expression of FcRn in the former.
LB albumin was shown by flow cytometric analysis to bemore retained in HMEC-1 cells than both WT and HB variants,an effect that was potentiated in HMEC-1-FcRn cells exhibitingoverexpression of the FcRn (Fig. 1, a– c). This suggests FcRn-mediated recycling similar to IgG (4, 24). Brülisauer et al. (20),however, observed none or only a slight decrease in the totalcell-associated fluorescence in HUVEC and Caco-2, respec-tively, up to 3 h after pulse-wash of fluorescent wild-type albu-min using a flow cytometric analysis. The authors suggestedcellular retention may be due to lower expression of FcRn inHUVEC than Caco2 cells.
Furthermore, we show an albumin-FcRn affinity-dependentuptake in HMEC-1-FcRn cells at both physiological pH and pH6.0 with increased uptake for HB and reduced uptake for LB
Figure 5. Endothelial cellular recycling of albumin. A, WT, FcRn high binder (HB), and FcRn low binder (LB) albumin variants were recycled inHMEC-1-FcRn cells. HMEC-1-FcRn cells were exposed to 0.15 �M albumin at pH 6.0 for 1 h to allow cellular uptake. After thorough washing of the cells,fresh pH 7.4 medium was added in which recycled albumin was quantified by ELISA after 1 h incubation. One representative of three experiments isshown. Student’s t test was used for comparison of LB and HB albumin variants to WT. ***, p � 0.001. Error bars, S.D. B, 15 independent recyclingexperiments with WT and HB albumin were performed as in A, and data are presented as recycling efficiency relative to WT. Error bar, S.E. C, IgG and IgYwere recycled as in A and measured by respective immunoglobulin specific ELISA. One representative of two experiments is shown. BD, Below detection.Error bars, S.D.
Figure 6. The effect of FcRn expression, pH, and albumin concentration on albumin recycling. WT, low (LB), and high FcRn binder (HB) albumin variantswere recycled in HMEC-1-FcRn (A) and HMEC-1 (B) cells as described under “Experimental Procedures.” All exposure cells received either 0.15 �M or 8 �M
albumin where the pH was either 6.0 or 7.4. During recycling and release of albumin pH was 7.4 in all cases. The figures show concentrations of albuminmeasured in supernatants, and one representative of two experiments performed is shown. Student’s t test was used to compare LB and HB albumin variantsto WT. ***, p � 0.001; *, p � 0.05. BD, below detection. NS, non-significant. Error bars, S.D.
FcRn-driven endosomal albumin recycling
J. Biol. Chem. (2017) 292(32) 13312–13322 13317
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

compared with the WT. Greater uptake observed at pH 6.0seemingly reflects the higher FcRn engagement shown previ-ously at low pH by surface plasmon resonance (5). In contrast towhat we observed in HMEC-1-FcRn, we observed minimal dif-ferences between the three albumin variants in the low FcRn-expressing HMEC-1 cells. Together these findings indirectlydemonstrate an FcRn-dependent uptake and clearance of albu-min from the cells.
Flow cytometric uptake studies using the pH-sensitive5FAM in combination with monensin revealed a greater fluo-rescence increase for the WT and HB after monensin addi-tion in the HMEC-1-FcRn cells, which supports localizationin a low pH environment suggestive of endosomal compart-mentalization, �6.0 – 6.8 (31), facilitated by the higher FcRnaffinity.
Confocal analysis using fluorescently labeled albumin andcellular markers allowed direct detection of the cellular loca-tion site of albumin. Determination of co-localization with cel-lular compartments was investigated by both endogenousexpressing fluorescent cellular compartment markers and im-munofluorescence staining. No significant differences in thetrafficking pathway were found in both non-permeabilized cellsor permeabilized cells. Punctuated fluorescence within the cellsindicated cellular vesicular compartmentalization. The LB,WT, and HB were shown to co-localize within early endo-somes, in agreement with the previous reports suggesting cel-lular uptake by endocytosis (10, 18, 20). All three variants wereshown to localize with late endosomes that supports an endo-cytotic pathway; however, only the LB exhibited significant lys-osome co-localization. This gives direct support for the neces-
sity for FcRn-mediated salvage from lysosomal degradation.This is in contrast to the findings by Carson et al. (19) thatFITC-labeled albumin is degraded within lysosomes inpodocytes. This discrepancy may be due to labeling used anddifferences between cell types and different FcRn expressionlevels. Previously, Brülisauer et al. (20) demonstrated thatmodified WT albumin is found in RAB7-positive vesiclesand exposed to a non- or only poorly bio-reducing environ-ment in epithelial (Caco-2) and endothelial cells (HUVEC);however, the FcRn dependence was not addressed. We showalbumin located in late endosomes; however, using the albu-min variants, we show that reducing the FcRn affinity resultsin lysosomal accumulation. Albumin cellular traffickingwould also be expected to be influenced by alternative albu-min receptors reported in the literature such as the gp18,gp30 (32, 33), gp60 (34), and megalin/cubilin; however, thefocus of this work was to investigate the role of FcRn inendosomal recycling.
By the use of the HMEC-1-FcRn cells we have developed anovel assay to quantitatively measure the cellular recycling ofalbumin. Similar assays have been demonstrated for IgG butwere primarily based on epithelium and focusing on transcyto-sis (9, 35–39). We show the crucial role of FcRn during albuminrecycling in vitro by the increased levels of albumin recycled byHMEC-1-FcRn cells compared with non-transduced HMEC-1cells. Moreover, we demonstrate a significant increase in recy-cling efficiency of an albumin variant with improved FcRn-binding. These findings are in contrast to a previous reportshowing no effect of increased rat FcRn expression in Madin-Darby canine kidney (MDCK) cells on rat albumin transcytosisand recycling (40).
The application of albumin as a drug carrier represents anattractive approach for drug circulatory half-life extension andefficacy improvement (41– 43). Hence, the in vitro quantifica-tion of albumin recycling provides a useful tool for predicting invivo half-life of albumin variants and their drug conjugatedcounterparts. Here, we demonstrate the unaltered in vitro recy-cling efficiencies of albumin conjugated to either a small chem-ical molecule or a peptide. Although various in vivo clearancemechanisms exist, these findings suggest that our recyclingassay could be used as a screening tool of albumin-drug mole-cules for maintained FcRn affinity and recycling efficiency and,furthermore, demonstrate the potential benefits of albuminvariants with improved FcRn-binding as a drug half-life exten-sion technology.
In this work a panel of recombinant albumins with alteredFcRn-binding affinities in combination with isogenic low andhigh FcRn-expressing HMEC-1 cells provided a unique tool kitto investigate the role of FcRn in the intracellular trafficking ofalbumin. The combination of flow cytometric and confocalmicroscopic analysis together with an in vitro cellular recyclinginvestigation allowed direct evidence for the first time of anFcRn-mediated endocytotic recycling pathway that is vital tounderstanding albumin’s pivotal role in endogenous ligandtransport and utilization to tune the pharmacokinetics of albu-min-based therapeutics.
Figure 7. FcRn-dependent recycling of albumin variants conjugated to asmall chemical molecule or peptide. WT, low (LB), and high FcRn binder (HB)albumin variants conjugated to Alexa Fluor 488 (A), exenatide (B), or non-conjugated were recycled in HMEC-1-FcRn cells and detected by ELISA asdescribed in Fig. 5. Results are presented as relative recycling with WT albu-min set as 100%. Error bars, S.D.
FcRn-driven endosomal albumin recycling
13318 J. Biol. Chem. (2017) 292(32) 13312–13322
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Experimental procedures
Albumin production and labeling
HSA WT and albumin variants LB (with low-binding affinityto hFcRn; HSA K500A) and HB (with high-binding affinity tohFcRn; HSA K573P) were produced in yeast followed by a two-step purification using AlbuPure� matrix and diethylaminoeth-yl weak anion exchange Sepharose Fast Flow (GE Healthcare)matrix as described previously (44). Alexa488, Alexa594, and5FAM dyes with a maleimide-reactive group (MolecularProbes) were conjugated to albumin’s Cys-34 free thiol residue.The conjugation reactions were performed in PBS (phosphate-buffered saline, Medicago, Sweden) at pH 7.1 at an �1:1.3molar ratio (albumin to fluorescence-maleimide compound).The reaction was performed overnight at room temperaturewith gentle shaking, wrapped in foil to protect against light, andterminated by the addition of 0.5 volume WFI (water for injec-tion) followed by a pH adjustment to 5.3 with acetic acid. Theunreacted fluorescence-maleimide compound was removed bytwo cycles of concentration and dilution with WFI in VivaSpin-30. The conjugation was stabilized by hydrolysis (ring-opening)of the maleimide ring (45) by the addition of PBS and pH adjust-ment to 9.0. The hydrolysis was performed for 20 h at roomtemperature and terminated by neutralizing to pH 7.0. Becausethe overall yield after hydrolysis was �50%, the material wassubjected to another cycle of conjugation and hydrolysis asdescribed above. After the second round of hydrolysis, unre-acted and hydrolyzed fluorescence-maleimide compound wereremoved by ultra/diafiltration in VivaSpin-30 with repeatedcycles of concentration and dilution with PBS (pH 7.0).
The degree of labeling is estimated based on the manufactu-rer’s instructions (46) using the following parameters: 5FAM(493 nm) � � 68,000, CF280 � 0.3; Alexa488 (488 nm) � �73,000; CF280 � 0.11; Alexa594 � � 92000, CF280 � 0.56.
FcRn-binding affinity was assessed by Biolayer Interferome-try on an Octet Red96 system (Pall/Fortebio) to evaluate thebinding integrity of the fluorescent albumins against humanFcRn at pH 5.5 and 7.0 essentially as described previously (47).
Cell lines
Human dermal microvascular endothelial cells (HMEC-1)(obtained from the Center for Disease Control) were culturedin MCDB131 medium (without L-glutamine) (Thermo FisherScientific) supplemented with 10% fetal bovine serum (FBS;Sigma), 2 mM L-glutamine (Lonza, Basel, Switzerland), 10 ng/mlhuman epidermal growth factor (EGF) (Peprotech), 1 ng/mlfibroblast growth factor (Peprotech), 50 �g/ml gentamicin(Sigma), and 0.25 �g/ml Fungizone (Thermo Fisher Scientific)at 37 °C in a humidified atmosphere with 5% CO2. HMEC-1cells were engineered to stably overexpress hFcRn (HMEC-1-FcRn) by lentiviral transduction (transduction performed bySirion Biotech, Martinsried, Germany). In brief, HMEC-1 cellswere transduced by replicative-deficient lentivirus bearing vec-tors with hFcRn and human �2-microglobulin sequences underthe control of constitutive cytomegalovirus and the ubiquitin Cpromoters, respectively. Vector inclusion of neo and puro resis-tance genes enabled antibiotic selection using puromycin(Thermo Fisher Scientific) and G418 (Sigma).
Quantitative PCR
qPCR was performed to confirm the increased expression ofFcRn in the engineered HMEC-1-FcRn cells. qPCR was carriedout using a LightCycler480 (Roche Applied Science) using theLightCycler480 SYBR Green I Master (Roche Applied Science)reagent system. The sequences of the primers used wereFcRn-Fw (5�-AAACCTGGAGTGGAAGGAGC) and FcRn-Rv(5�-GGTAGAAGGAGAAGGCGCTG) for amplification ofFcRn expression and GAPDH-Fw (5�-GTCAGCCGCATC-TTCTTTTG and GAPDH-Rv (5�-GCGCCCAATACGAC-CAAATC) for amplification of the reference gene GAPDH(DNA Technology, Risskov, Denmark).
Flow cytometric cellular uptake and trafficking studies
HMEC-1 and HMEC-1-FcRn cells were seeded in 48-wellplates (Sarstedt, Numbrecht, Germany) at a density of 8.0 105
cells/well in complete HMEC-1 or complete HMEC-1-FcRnmedia in 0.5 ml for 24 h. Cells were exposed to 8 �M fluorescentalbumin labeled with Alexa488 or 5FAM in a total volume of125 �l of Hanks’ balanced salt solution (HBSS) without phenolred at pH 7.4 or adjusted with 1.0 M MES solution to pH 6.0 for0.5– 4 h followed by subsequent media removal and a 3 washin ice-cold HBSS. Cells were harvested by trypsin treatment andcentrifuged at 300 g for 5 min at 4 °C followed by an addi-tional wash, and cells were resuspended in 500 �l of sterile-filtrated PBS containing 2% bovine serum albumin (BSA), 0.1%NaN3. For determination of albumin retention as an indicatorof recycling, cells were exposed to 8 �M fluorescent albuminlabeled with Alexa488 in a total volume of 125 �l of HBSSadjusted with 1 M MES solution to pH 6.0 for 2 h followed bysubsequent media removal and 3 wash in ice-cold HBSS.Cells were then incubated for 1 or 2 h in HBSS (pH 7.4) beforethe cells were harvested by trypsin treatment and centrifuged at300 g for 5 min at 4 °C followed by an additional wash, andcells were resuspended in 500 �l of sterile-filtrated PBS flowbuffer containing 2% BSA, 0.1% NaN3.
The samples were analyzed by flow cytometry using a Galliosflow cytometer (Beckman Coulter) equipped with a 488-nmlaser and the 525/40-nm filter (FL1). All samples were mea-sured before and after incubation with 20 �M monensin in99.8% ethanol (VWR) for 10 min. Data were processed, andmean fluorescence intensity was determined using Kaluza 1.2software (Beckman Coulter).
Transfection-based detection of endosomal/lysosomallocalization by confocal microscopy of non-permeabilized cells
Cells were transfected with baculovirus (BacMam, Molecu-lar Probes) for transient expression of GFP-fused Ras-relatedprotein Rab-5 (Rab5) as an early endosome marker, Ras-relatedprotein Rab-7a (Rab7) as a late endosome marker, and LAMP1as a lysosome marker according to the manufacturer’s instruc-tions. Cells were transfected 24 – 48 h before the albuminuptake experiments. Alexa647-labeled dextran (MolecularProbes) was added to the cells 16 h before imaging at a finalconcentration of 50 �g/ml preceding the experiment, whereascells were maintained in the cell culture incubator. Dextranmedia was replaced with fresh dextran-free media before imag-ing. Confocal images were captured using a Zeiss confocal
FcRn-driven endosomal albumin recycling
J. Biol. Chem. (2017) 292(32) 13312–13322 13319
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

microscope LSM 780 (Carl Zeiss MicroImaging GmbH, Jena)equipped with lasers providing excitations at 488, 594, and 647nm, a motorized xyz stage, and an incubator for temperatureand CO2 control for live cell imaging. Images were capturedusing a Plan-Apochromat, 63 NA 1.4, differential interfer-ence contrast (DIC) oil immersion objective. For Z-series,0.24-�m slice spacing was used. All system settings were main-tained identically for experiments where quantitative compar-isons were performed.
Immunofluorescence detection of endosomal/lysosomallocalization confocal microscopy of permeabilized cells
Rabbit primary antibodies for early endosomal antigen-1(EEA1), Ras-related protein Rab-7a (RAB7A), and LAMP1 werepurchased from Abcam (Cambridge, UK). Secondary goat anti-rabbit antibodies labeled with Alexa647 were purchased fromLife Technologies. HBSS without phenol red, formalin, Dulbec-co’s PBS, Triton X-100, BSA, Hoechst 33342, (MES, NaN3, andmonensin were purchased from Sigma.
Confocal images were captured using a Zeiss Confocalmicroscope LSM 700 (Carl Zeiss MicroImaging) equipped withlasers providing excitations at 405, 488, 555, and 647 nm, amotorized xyz stage. Images were captured using an alpha Plan-Apochromat, 100 /1.46 oil DIC M27 immersion objective. Allsystem settings were maintained identically for experimentswhere quantitative comparisons were performed.
Image processing
Images were processed using Zeiss Zen Black 2012 edition.Whenever possible, signal saturation was avoided, and cellswith saturated pixels were eliminated from analysis. Imagesshown in the figures were contrast-stretched to enhance thevisibility of dim structures, but all images to be compared werecontrast-enhanced identically. For quantification, the fluores-cence signal in each image plane was corrected for background.A region of interest was drawn around individual cells, and theaverage pixel intensity was recorded for each fluorescent probein the cell. Coefficients of co-localization were obtained fromtri-dimensional images using Zen Black edition software pack-age (Zeiss).
Cellular uptake of albumin
Uptake and co-localization of albumin with endosomal andlysosome markers was investigated using HMEC-1 and HMEC-1-FcRn cells overexpressing GFP-fused endocytic markers(non-permeabilized cells) or by staining the endogenous mark-ers with fluorescently labeled antibodies (permeabilized cells).In both cases the cells were exposed to fluorescently labeledalbumin compounds. Cells were seeded in tissue culture plateschambers (Nunc, Thermo Fisher Scientific, or Sarstedt) withcoverslips to 80% cell confluency (0.8 106 cells/well). Cellswere exposed to 8 �M fluorescent albumin in HBSS for 1–2 hbefore fixation. Upon removal of the media containing thealbumins, the cells were washed 3 with ice-cold PBS and fixedfor 20 min in 4% paraformaldehyde and mounted; alternatively,for antibody staining, the cells were fixed in 10% formalin fol-lowed by a 3 wash in PBS. Cells were permeabilized by incu-bation with 0.5% Triton X-100 in PBS for 10 min followed by
blocking in 5% BSA in PBS for 30 min. Cells were washed 3and incubated with primary antibodies EEA1 (diluted 1:200),RAB7A (1:100), and LAMP1 (1:200) in PBS. Upon incubationwith primary antibody for 1 h at room temperature and fol-lowed by a 3 wash in PBS, the cells were then incubated withAlexa647-labeled goat anti-rabbit secondary antibody dilutedto 4 �g/ml. Cells were incubated with the secondary antibodyfor 1 h at room temperature followed by a 3 wash in PBS.Actin filaments of the cells were stained by incubation with0.5 units of ATTO565-labeled phalloidin (Atto-Tec, Siegen,Germany), diluted in 1% BSA in PBS for 20 min at roomtemperature, and followed by a 3 wash in PBS. The nucleiof the cells were stained with Hoechst 33342 diluted 1:2000in PBS for 10 min at room temperature followed by a 3wash in PBS.
Finally, the cells were mounted in fluorescent mountingmedia (Dako, Glostrup, DK) under a high precision coverglass(VWR) and sealed with transparent nail polish. Images wereacquired using confocal microscopy as described above. ForAlexa488-labeled albumin variants and GFP-fused markers(excitation (Ex), 488 nm; emissions (Em), 530/30), Hoechst33342-stained nuclei (Ex, 405 nm; Em, 445/70), ATTO565-la-beled phalloidin (Ex, 555 nm; Em, 580/40), Alexa647-labeledsecondary antibodies and labeled albumins (Ex, 638; Em, 670/50) filter settings were used. Images were acquired with thesame exposure times and laser power at all times.
Albumin cellular recycling assay
1 105 cells/well HMEC-1-FcRn cells were seeded in48-well plates (Corning) precoated with GelTrex� (ThermoFisher Scientific) using the Thin Layer Method (non-gelling)coating procedure according to the manufactures protocol.The cells received fresh complete medium every other day andwere cultured for at least 10 days to reach a confluent cellmonolayer before experimental use. On the day of recycling,the cells were washed twice in prewarmed PBS (Thermo FisherScientific) followed by the addition of 0.15 �M albumin (300�l/well) in HBSS (Sigma) adjusted to pH 6.0 by 1 M MES buffer(Sigma). For recycling of immunoglobulins, cells were exposedto 0.07 �M IgG or IgY (Jackson ImmunoResearch Laboratories).After a 1-h incubation at 37 °C, 5% CO2, the cells were washed5 times in ice-cold PBS to remove all the extracellular albuminthen received 160 �l/well complete medium without FBS andwere incubated for another 1 h to allow recycling and release ofinternalized albumin. Finally, supernatants were collected andanalyzed for albumin by ELISA.
ELISA detection of albumin and immunoglobulin G
In vitro recycled albumin was detected in a human albumin-specific sandwich ELISA. Maxisorp plates (Nunc) were coatedovernight at 4 °C with a polyclonal goat anti-human albuminantibody (A-7544) (Sigma) (1:1000 diluted in PBS). Coatedplates were blocked for 1 h with casein buffer (Sigma) beforewashing with PBS � 0.05% Tween and loading of the albuminstandard and samples. Binding was allowed for 1 h followed bywashing and incubation with a horseradish peroxidase (HRP)-conjugated secondary polyclonal sheep anti-human albuminantibody (ab8941) (Abcam) (1:5000 diluted in 10% casein
FcRn-driven endosomal albumin recycling
13320 J. Biol. Chem. (2017) 292(32) 13312–13322
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

buffer) for 1 h. After washing color reaction was achieved by3,3�,5,5�-tetramethylbenzidine (TMB) (KemEnTec, Taastrup,Denmark), and enzymatic reactivity was stopped by H2SO4before reading the plate at 450 nm.
Commercially available ELISA kits (GenWay Biotech) wereused for measurement of human IgG (GWB-A04A50) and IgY(GWB-374Z14) and used according to the manufacturer’s pro-tocol. Briefly, supernatants were transferred to anti-IgY or anti-IgG antibody-coated plates and incubated at room temperaturefor 30 min (IgY) or 60 min (IgG). Enzyme-antibody conjugateswere used for detection and 3,3�,5,5�-tetramethylbenzidine wasused for color development. Reaction was stopped with H2SO4,and plates were read at 450 nm.
Author contributions—K. A. H., J. C., and D. V. conceived theresearch. K. A. H., M. L. H., E. G. W. S., D. V., F. A., B. A., andN. N. K. planned the studies. E. G. W. S., M. L. H., F. A., J. C., B. A.,and N. N. K. performed the experiments and analyzed and inter-preted the data. K. A. H., M. L. H., E. G. W. S., and F. A. drafted themanuscript. All authors read and approved the final manuscript.
References1. Peters, T. (1996) All about Albumin: Biochemistry, Genetics, and Medical
Applications. Academic Press, San Diego, CA2. Chaudhury, C., Mehnaz, S., Robinson, J. M., Hayton, W. L., Pearl, D. K.,
Roopenian, D. C., and Anderson, C. L. (2003) The major histocompatibil-ity complex-related Fc receptor for IgG (FcRn) binds albumin and pro-longs its lifespan. J. Exp. Med. 197, 315–322
3. Roopenian, D. C., Christianson, G. J., Sproule, T. J., Brown, A. C., Akilesh,S., Jung, N., Petkova, S., Avanessian, L., Choi, E. Y., Shaffer, D. J., Eden,P. A., and Anderson, C. L. (2003) The MHC class I-like IgG receptorcontrols perinatal IgG transport, IgG homeostasis, and fate of IgG-Fc-coupled drugs. J. Immunol. 170, 3528 –3533
4. Ward, E. S., Zhou, J., Ghetie, V., and Ober, R. J. (2003) Evidence to supportthe cellular mechanism involved in serum IgG homeostasis in humans.Int. Immunol. 15, 187–195
5. Chaudhury, C., Brooks, C. L., Carter, D. C., Robinson, J. M., and Anderson,C. L. (2006) Albumin binding to FcRn: distinct from the FcRn-IgG inter-action. Biochemistry 45, 4983– 4990
6. Andersen, J. T., Dalhus, B., Cameron, J., Daba, M. B., Plumridge, A., Evans,L., Brennan, S. O., Gunnarsen, K. S., Bjørås, M., Sleep, D., and Sandlie, I.(2012) Structure-based mutagenesis reveals the albumin-binding site ofthe neonatal Fc receptor. Nat. Commun 3, 610
7. Andersen, J. T., Dalhus, B., Viuff, D., Ravn, B. T., Gunnarsen, K. S., Plum-ridge, A., Bunting, K., Antunes, F., Williamson, R., Athwal, S., Allan, E.,Evans, L., Bjørås, M., Kjærulff, S., Sleep, D., Sandlie, I., and Cameron, J.(2014) Extending serum half-life of albumin by engineering neonatal Fcreceptor (FcRn) binding. J. Biol. Chem. 289, 13492–13502
8. Ehrhardt, C., and Kim, K. J. (2008) Drug Absorption Studies: in Situ, inVitro, and in Silico Models, pp. 640 – 662, Springer, New York
9. Dickinson, B. L., Badizadegan, K., Wu, Z., Ahouse, J. C., Zhu, X., Simister,N. E., Blumberg, R. S., and Lencer, W. I. (1999) Bidirectional FcRn-depen-dent IgG transport in a polarized human intestinal epithelial cell line.J. Clin. Invest. 104, 903–911
10. Maunsbach, A. B. (1966) Albumin absorption by renal proximal tubulecells. Nature 212, 546 –547
11. Maunsbach, A. B. (1966) Absorption of I-125-labeled homologous albu-min by rat kidney proximal tubule cells: a study of microperfused singleproximal tubules by electron microscopic autoradiography and histo-chemistry. J. Ultrastruct. Res. 15, 197–241
12. Park, C. H., and Maack, T. (1984) Albumin absorption and catabolism byisolated perfused proximal convoluted tubules of the rabbit. J. Clin. Invest.73, 767–777
13. Russo, L. M., Sandoval, R. M., McKee, M., Osicka, T. M., Collins, A. B.,Brown, D., Molitoris, B. A., and Comper, W. D. (2007) The normal kidneyfilters nephrotic levels of albumin retrieved by proximal tubule cells: re-trieval is disrupted in nephrotic states. Kidney Int. 71, 504 –513
14. Tenten, V., Menzel, S., Kunter, U., Sicking, E. M., van Roeyen, C. R.,Sanden, S. K., Kaldenbach, M., Boor, P., Fuss, A., Uhlig, S., Lanzmich, R.,Willemsen, B., Dijkman, H., Grepl, M., Wild, K., Kriz, W., et al. (2013)Albumin is recycled from the primary urine by tubular transcytosis. J. Am.Soc. Nephrol. 24, 1966 –1980
15. Birn, H., Fyfe, J. C., Jacobsen, C., Mounier, F., Verroust, P. J., Orskov, H.,Willnow, T. E., Moestrup, S. K., and Christensen, E. I. (2000) Cubilin is analbumin binding protein important for renal tubular albumin reabsorp-tion. J. Clin. Invest. 105, 1353–1361
16. Amsellem, S., Gburek, J., Hamard, G., Nielsen, R., Willnow, T. E., Devuyst,O., Nexo, E., Verroust, P. J., Christensen, E. I., and Kozyraki, R. (2010)Cubilin is essential for albumin reabsorption in the renal proximal tubule.J. Am. Soc. Nephrol. 21, 1859 –1867
17. Verroust, P. J., and Christensen, E. I. (2002) Megalin and cubilin-the storyof two multipurpose receptors unfolds. Nephrol. Dial. Transplant 17,1867–1871
18. Predescu, D., Vogel, S. M., and Malik, A. B. (2004) Functional and mor-phological studies of protein transcytosis in continuous endothelia. Am. J.Physiol. Lung Cell Mol. Physiol. 287, L895–L901
19. Carson, J. M., Okamura, K., Wakashin, H., McFann, K., Dobrinskikh, E.,Kopp, J. B., and Blaine, J. (2014) Podocytes degrade endocytosed albuminprimarily in lysosomes. PloS ONE 9, e99771
20. Brülisauer, L., Valentino, G., Morinaga, S., Cam, K., Thostrup Bukrinski, J.,Gauthier, M. A., and Leroux, J. C. (2014) Bio-reduction of redox-sensitivealbumin conjugates in FcRn-expressing cells. Angew. Chem. Int. Ed. Engl.53, 8392– 8396
21. Andersen, J. T., Dee Qian, J., and Sandlie, I. (2006) The conserved histidine166 residue of the human neonatal Fc receptor heavy chain is critical forthe pH-dependent binding to albumin. Eur. J. Immunol. 36, 3044 –3051
22. Oganesyan, V., Damschroder, M. M., Cook, K. E., Li, Q., Gao, C., Wu, H.,and Dall’Acqua, W. F. (2014) Structural insights into neonatal Fc receptor-based recycling mechanisms. J. Biol. Chem. 289, 7812–7824
23. Ober, R. J., Martinez, C., Lai, X., Zhou, J., and Ward, E. S. (2004) Exocytosisof IgG as mediated by the receptor, FcRn: an analysis at the single-mole-cule level. Proc. Natl. Acad. Sci. U.S.A. 101, 11076 –11081
24. Ward, E. S., Martinez, C., Vaccaro, C., Zhou, J., Tang, Q., and Ober, R. J.(2005) From sorting endosomes to exocytosis: association of Rab4 andRab11 GTPases with the Fc receptor, FcRn, during recycling. Mol. Biol.Cell 16, 2028 –2038
25. Gan, Z., Ram, S., Vaccaro, C., Ober, R. J., and Ward, E. S. (2009) Analysesof the recycling receptor, FcRn, in live cells reveal novel pathways forlysosomal delivery. Traffic. 10, 600 – 614
26. Weflen, A. W., Baier, N., Tang, Q. J., Van den Hof, M., Blumberg, R. S.,Lencer, W. I., and Massol, R. H. (2013) Multivalent immune complexesdivert FcRn to lysosomes by exclusion from recycling sorting tubules. Mol.Biol. Cell 24, 2398 –2405
27. Schultze, H. E., and Heremans, J. F. (1966) Molecular Biology of HumanProteins, with Special Reference to Plasma Proteins. Elsevier Pub. Co., NewYork
28. Brambell, F. W. R. (1970) The Transmission of Passive Immunity fromMother to Young, pp. 1–385, North-Holland Publishing Co., AmericanElsevier, Amsterdam
29. Borvak, J., Richardson, J., Medesan, C., Antohe, F., Radu, C., Simionescu,M., Ghetie, V., and Ward, E. S. (1998) Functional expression of the MHCclass I-related receptor, FcRn, in endothelial cells of mice. Int. Immunol.10, 1289 –1298
30. Akilesh, S., Christianson, G. J., Roopenian, D. C., and Shaw, A. S. (2007)Neonatal FcR expression in bone marrow-derived cells functions to pro-tect serum IgG from catabolism. J. immunol. 179, 4580 – 4588
31. Mellman, I. (1996) Endocytosis and molecular sorting. Annu. Rev. CellDev. Biol. 12, 575– 625
32. Schnitzer, J. E., and Bravo, J. (1993) High affinity binding, endocytosis, anddegradation of conformationally modified albumins: potential role of gp30and gp18 as novel scavenger receptors. J. Biol. Chem. 268, 7562–7570
FcRn-driven endosomal albumin recycling
J. Biol. Chem. (2017) 292(32) 13312–13322 13321
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

33. Schnitzer, J. E., Sung, A., Horvat, R., and Bravo, J. (1992) Preferential in-teraction of albumin-binding proteins, gp30 and gp18, with conforma-tionally modified albumins: presence in many cells and tissues with apossible role in catabolism. J. Biol. Chem. 267, 24544 –24553
34. Schnitzer, J. E., and Oh, P. (1994) Albondin-mediated capillary perme-ability to albumin: differential role of receptors in endothelial transcy-tosis and endocytosis of native and modified albumins. J. Biol. Chem.269, 6072– 6082
35. Claypool, S. M., Dickinson, B. L., Yoshida, M., Lencer, W. I., and Blum-berg, R. S. (2002) Functional reconstitution of human FcRn in Madin-Darby canine kidney cells requires co-expressed human beta 2-micro-globulin. J. Biol. Chem. 277, 28038 –28050
36. Claypool, S. M., Dickinson, B. L., Wagner, J. S., Johansen, F. E., Venu, N.,Borawski, J. A., Lencer, W. I., and Blumberg, R. S. (2004) Bidirectionaltransepithelial IgG transport by a strongly polarized basolateral mem-brane Fc�-receptor. Mol. Biol. Cell 15, 1746 –1759
37. Tzaban, S., Massol, R. H., Yen, E., Hamman, W., Frank, S. R., Lapierre,L. A., Hansen, S. H., Goldenring, J. R., Blumberg, R. S., and Lencer, W. I.(2009) The recycling and transcytotic pathways for IgG transport by FcRnare distinct and display an inherent polarity. J. Cell Biol. 185, 673– 684
38. Liu, X., Ye, L., Christianson, G. J., Yang, J. Q., Roopenian, D. C., and Zhu,X. (2007) NF-kappaB signaling regulates functional expression of theMHC class I-related neonatal Fc receptor for IgG via intronic bindingsequences. J. Immunol. 179, 2999 –3011
39. Leitner, K., Ellinger, I., Grill, M., Brabec, M., and Fuchs, R. (2006) Efficientapical IgG recycling and apical-to-basolateral transcytosis in polarizedBeWo cells overexpressing hFcRn, Placenta 27, 799 – 811
40. Tesar, D. B., Tiangco, N. E., and Bjorkman, P. J. (2006) Ligand valencyaffects transcytosis, recycling and intracellular trafficking mediated by theneonatal Fc receptor. Traffic 7, 1127–1142
41. Xie, D., Yao, C., Wang, L., Min, W., Xu, J., Xiao, J., Huang, M., Chen, B.,Liu, B., Li, X., and Jiang, H. (2010) An albumin-conjugated peptide exhibitspotent anti-HIV activity and long in vivo half-life. Antimicrob. AgentsChemother 54, 191–196
42. Wu, Y. L., Huang, J., Xu, J., Liu, J., Feng, Z., Wang, Y., Lai, Y., and Wu, Z. R.(2010) Addition of a cysteine to glucagon-like peptide-1 (GLP-1) conju-gates GLP-1 to albumin in serum and prolongs GLP-1 action in vivo.Regul. Pept. 164, 83– 89
43. Baggio, L. L., Huang, Q., Cao, X., and Drucker, D. J. (2008) An albumin-exendin-4 conjugate engages central and peripheral circuits regulatingmurine energy and glucose homeostasis. Gastroenterology 134,1137–1147
44. Evans, L., Hughes, M., Waters, J., Cameron, J., Dodsworth, N., Tooth, D.,Greenfield, A., and Sleep, D. (2010) The production, characterisation andenhanced pharmacokinetics of scFv-albumin fusions expressed in Saccha-romyces cerevisiae. Protein Expr. Purif. 73, 113–124
45. Smith, M. E., Caspersen, M. B., Robinson, E., Morais, M., Maruani, A.,Nunes, J. P., Nicholls, K., Saxton, M. J., Caddick, S., Baker, J. R., and Chu-dasama, V. (2015) A platform for efficient, thiol-stable conjugation toalbumin’s native single accessible cysteine. Org. Biomol. Chem. 13,7946 –7949
46. Johnson, I. (2010) Molecular Probes Handbook: A Guide to Fluores-cent Probes and Labeling Technologies. 11th Ed., ThermoFisherScientific
47. Viuff, D., Antunes, F., Evans, L., Cameron, J., Dyrnesli, H., Thue Ravn, B.,Stougaard, M., Thiam, K., Andersen, B., Kjærulff, S., and Howard, K. A.(2016) Generation of a double transgenic humanized neonatal Fc receptor(FcRn)/albumin mouse to study the pharmacokinetics of albumin-linkeddrugs. J. Control Release 223, 22–30
FcRn-driven endosomal albumin recycling
13322 J. Biol. Chem. (2017) 292(32) 13312–13322
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Birgitte Andersen, Nanna N. Kristensen and Kenneth A. HowardEsben G. W. Schmidt, Michael L. Hvam, Filipa Antunes, Jason Cameron, Dorthe Viuff,
pathway for cellular recycling of albuminDirect demonstration of a neonatal Fc receptor (FcRn)-driven endosomal sorting
doi: 10.1074/jbc.M117.794248 originally published online June 21, 20172017, 292:13312-13322.J. Biol. Chem.
10.1074/jbc.M117.794248Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2017/06/21/M117.794248.DC1
http://www.jbc.org/content/292/32/13312.full.html#ref-list-1
This article cites 42 references, 18 of which can be accessed free at
by guest on April 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from