digesón de proteínasy) absorciónde aminoácidos) · chapter 44: digestion and absorption of...
TRANSCRIPT

Diges&ón de proteínas y absorción de aminoácidos

Pepsinógeno Pepsina (endopep&dasa AA alifá&cos, en especial con radicales aromá&cos)
HCl
600 -‐800 AA
25-‐100 AA
Duodeno
Estómago
Fase de vaciado: contracción del antro medio y
relajación del píloro

Si7os de hidrólisis de la cadena pep9dica: residuos aminoácidos hidrofóbicos (preferentemente aromá7cos) en posicíon P1 y P1’. Ejemplo: hidroliza los enlaces pep9dicos Phe1-‐/-‐Val, Gln4-‐/-‐His, Glu13-‐/-‐Ala, Ala14-‐/-‐Leu, Leu15-‐/-‐Tyr, Tyr16-‐/-‐Leu, Gly23-‐/-‐Phe, Phe24-‐/-‐Phe and Phe25-‐/-‐Tyr de la cadena B de la insulina
Pepsina (EC 3.4.23.1) Cerdo
Lugares preferidos de hidrólisis: enlaces pep9dicos cercanos a aminoácidos con cadenas alífá7cas, especialmente los que 7enen radicales aromá7cos (Phe, Tyr, Trp)
Insulina
¿Puede volver a actuar sobre los pép7dos residuales después de la primera hidrólisis? ¿Hay un tamaño mímino de pép7do (N residuos aminoácidos) que la pepsina reconoce?
1 2 3 4 5 6
7 8 9
Preferen7al cleavage: hydrophobic, preferably aroma7c, residues in P1 and P1' posi7ons. Cleaves Phe1-‐/-‐Val, Gln4-‐/-‐His, Glu13-‐/-‐Ala, Ala14-‐/-‐Leu, Leu15-‐/-‐Tyr, Tyr16-‐/-‐Leu, Gly23-‐/-‐Phe, Phe24-‐/-‐Phe and Phe25-‐/-‐Tyr bonds in the B chain of insulin. No cofactors

Chapter 44: Digestion and Absorption of Nutrients 511
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
mammals. Both pepsin and rennin function optimally when the pH is between 2 and 3. The net result is that proteins that were hundreds of amino acids long when they entered the stomach enter the duodenum as fragments that may be 25–100 amino acids long.
As peptides reach the small intestine they activate receptors of enteroendocrine cells lining the duodenum crypts, stimu-lating them to secrete cholecystokinin (CCK). The CCK enters the circulation and reaches the pancreatic acinar cells and myoepithelial cells surrounding each acinus. This triggers secretion of pancreatic enzymes into the upper duodenum via the pancreatic ducts (Figure 44.9). The proteolytic enzymes of the pancreas are produced and secreted into the pancreatic ducts in an inactive form. This prevents autodigestion of the pancreas and pancreatic ducts. The proteolytic proenzymes secreted by the pancreas incude trypsinogen, chymotrypsinogen, pro‐elastase, and pro‐carboxypeptidases A and B.
The CCK secreted by crypt enteroendocrine cells in response to peptides (and fats) entering the duodenum also reaches the villous enterocytes. This causes the enterocytes to secrete an enzyme called enteropeptidase (also called enterokinase) into the lumen of the duodenum. Enteropeptidase seeks out trypsinogen that has entered the duodenum and cleaves off a fragment to form the active proteolytic enzyme trypsin. Trypsin then cleaves off portions of each of the other proteolytic enzymes secreted by the pancreas causing them to become active as well. Trypsin can actually convert trypsinogen to active trypsin in an example of positive feedback regulation. The action of
enteropeptidase quickly causes all the inactive proteolytic enzymes in the pancreatic secretions to become active in the lumen of the gut. Each of these different proteolytic enzymes (trypsin, chymo-trypsin, elastase, and the carboxypeptidases) cleave peptide bonds between specific amino acids so that when the luminal phase of digestion is finished the protein has been converted to peptides that are generally just 1–12 amino acids long.
These single amino acids and longer peptides then move to the brush border. They are very soluble in water and have no problem crossing the unstirred water layer and entering the glycocalyx, adhering to the microvilli forming the brush border of the villous enterocytes. Several intestinal peptidases project from the brush border into the glycocalyx, but these enzymes are not released into the lumen of the intestine. These intestinal peptidases hydrolyze the peptide bonds, reducing the length of the peptides to no more than three amino acids in length. The next obstacle to their absorption is moving across the apical membrane of the villous enterocyte.
Crossing the apical membrane of villous enterocytesThe single amino acids develop a large concentration gradient above the apical membrane of duodenal and jejunal villous cells following ingestion of a meal. Thanks to the secretory efforts of the crypt enterocytes, high amounts of sodium are also found above the apical membrane. At least four facilitated carriers are known to exist in the apical membrane of the villous cells. These transporters seem to be specific for the basic, acidic, or neutral amino acids.
CCK
Pancreas
Trypsinogen Trypsin
Inactive proteolytic enzymes
Enteropeptidase
EEC
Fat, AA
Duodenum
Enzymes
Active enzymes
Figure 44.9 Many of the enzymes secreted by the pancreas are in an inactive form. Activation begins when an enteroendocrine cell (EEC) secretes cholecystokinin (CCK) in response to the presence of fats or amino acids (AA) in the duodenum. The CCK stimulates the pancreas to secrete enzymes, many of which are in an inactive form. The CCK also acts on nearby villous enterocytes and causes them to secrete enteropeptidase. Enteropeptidase converts pancreatic trypsinogen to the active enzyme trypsin. Trypsin then cleaves off portions of all the other inactive pancreatic enzymes allowing them to become active in the intestinal lumen. Trypsin can also cleave trypsinogen to form more trypsin enzyme.
Procarboxipep&dasa A y B Proelastasa Quimotripsinógeno
Tripsinógeno
Enteropep7dasa
Tripsina
Enzimas ac&vas
Grasas, aminoácidos
Duodeno
Páncreas
Enzimas

512 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Proline seems to have its own unique carrier. All these single amino acid carriers are facilitated carriers that bind the amino acid and a Na+ atom (Figure 44.10). The combined force of the amino acid moving down its concentration gradient and Na+ moving down its electrical and concentration gradient helps drive the large amino acid molecule across the apical membrane.
The dipeptides and tripeptides at the brush border can be absorbed by active transport mechanisms. They are so large that it takes the force supplied by an ATP molecule along with a trans-porter protein to pump them across the apical membrane. Research suggests that the bulk of amino acids is transported across the apical membrane in the form of dipeptides and tripep-tides. Once these arrive in the cytosol of the villous enterocyte they are hydrolyzed to single amino acids by intracellular peptidases.
Crossing the basolateral membrane of the villous enterocyteAs the single amino acids accumulate at the basolateral side of the enterocytes, their concentration becomes much higher than that of free amino acids in the extracellular fluid. Transporters unique to the basolateral membrane facilitate the diffusion of amino acids across the basolateral membrane, independent of Na+. The amino acids enter the extracellular fluid and are trans-ported in the portal circulation to the liver. The Na+ ions that accompanied the single amino acids across the apical mem-brane are pumped into the extracellular fluid by the 3Na+/2K+
electrogenic pump residing in the basolateral membrane at the expense of an ATP. The sodium may be removed from the blood by crypt secretory cells and returned to the lumen to assist facilitated transport of other amino acids by villous cells.
Absorption of intact proteinsIn rare instances some very specific proteins can be absorbed intact across the intestinal villous cells. The most important of these are the colostral antibodies that provide passive immunity for the neonatal mammal. In the case of colostral immunoglobu-lins, the antibodies found in colostrum have unique properties that allow them to resist degradation by stomach acid and the proteolytic enzymes. Neonatal proteolytic enzyme secretion and activation processes do not seem to be fully developed, which also helps the protein avoid digestion. Villous cells of the neonate have specific receptors that recognize the immunoglobulins. Once the immunoglobulin binds its receptor, it activates endocy-tosis of the immunoglobulin: it is enclosed in a section of the apical membrane, transported to the basolateral membrane, and released into the extracellular fluid by exocytosis. The presence of these receptors on the neonatal villous enterocytes is short‐lived: most mammals lose these immunoglobulin receptors and stop absorbing immunoglobulin within 24 hours of birth.
Digestion and absorption of nonstructural carbohydrates
Plant starches and glycogen from ingestion of muscle and liver comprise large numbers of glucose molecules linked together by
bonds at the α(1→4) or α(1→6) position. The most common plant sugar in the diet is sucrose, a disaccharide composed of a glucose molecule linked to a fructose molecule. Milk sugar or lactose is a disaccharide of glucose and galactose linked in the β(1→4) position. Digestion of plant structural carbohydrates (cellulose and hemicellulose) is critical to the survival of rumi-nants and hindgut fermenters and is discussed in Chapter 45.
On ingestion, starches in the diet will begin to be broken down by salivary α‐amylase. This process breaks some of the α(1→4) linkages, but the enzyme does not have time to break down very much of the starch molecule before the bolus enters
Peptides
BBPeptidase
Dipeptides andtripeptides
Dipeptides andtripeptides
Aminoacid
Aminoacid
Aminoacid
Aminoacid
ATP
Na+
Na+
Cytosol
Peptidase
Figure 44.10 Brush border digestion and absorption of proteins and amino acids. A brush border peptidase (BB peptidase) of villous cells can break down any peptides greater than three amino acids that reach the glycocalyx. Single amino acids then use one four known types of Na+/amino acid cotransporters to cross the apical membrane. These facilitated diffusion carriers utilize the driving forces provided by the high concentration of amino acid in the lumen following a meal and high lumen Na+ provided by crypt cell secretions to move the large charged amino acids across the brush border. Dipeptides and tripeptides can be transported by special active transport proteins that do not require Na+, but do expend an ATP to move such large molecules across the membrane. Once inside the cell the dipeptides and tripeptides are converted to single amino acids by intracellular peptidases. Transporters unique to the basolateral membrane then facilitate diffusion of the amino acids into the extracellular fluid.
1 What is the difference between starch, glucose, fructose, and lactose?
2 Where does amylase come from and what does it do?
3 Can disaccharides cross the apical membrane?
4 Which enzymes are found within the glycocalyx attached to the brush border membrane? What is their primary function?
5 How do single molecules of galactose and glucose cross the apical cell membrane? How does fructose cross the apical membrane of villous cells.
6 Does sugar absorption from the intestinal tract require insulin?
Carboxipep&dasa A y B Elastasa
Quimotripsina Tripsina
aminoácidos ácidos aminoácidos básicos aminoácidos neutros prolina
Transportadores de AA
Pép7dos Dipép7dos triép7dos
Dipép7dos triép7dos
Aminoácidos
Aminoácidos
Aminoácidos
Pep7dasa
Pep7dasa Pep&dasa
RC
Aminoácidos
Na+
Na+
ATP
ADP
Pep&dasa RC: pep&dasa del ribete en cepillo
3 Na+
2 K+
ATP ADP
¿?
¿Gasto de energía asociado al movimiento de AA?

Diges&ón y absorción de carbohidratos

Chapter 44: Digestion and Absorption of Nutrients 513
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
the stomach where the amylase will be destroyed by the low pH. Stomach acids and proteolytic enzymes have no effect on starches and they continue into the duodenum. The osmolarity change caused by entry of starch and acids into the duodenum causes vagal parasympathetic stimulation of pancreatic secre-tion. The pancreas secretes α‐amylase in an active form that attacks the α(1→4) links between glucose molecules and is highly efficient. The bulk of starch is broken down to maltose (two glucoses) and maltotriose (three glucoses) and limit dex-trins (α1→6 linked glucoses) within minutes of entry to the duodenum. These starch breakdown products then move to the brush border: they are very water‐soluble and have no problem crossing the unstirred water layer to reach the glycocalyx. A wide variety of enzymes extend into the glycocalyx from the brush border and these complete the digestive process (Figure 44.11). Some of these enzymes include sucrase, which converts sucrose to glucose and fructose; maltase and maltotriase, which convert maltose and maltotriose to their constitutive glucose molecules; and lactase, which converts milk lactose to glucose and galactose. Lactase is found on the villous enterocyte brush border of all mammalian neonates, but often disappears after the animal is weaned. Sucrase, on the other hand, is often lacking in neonates and is expressed only after the animal is several weeks old. Trehalase is an enzyme that breaks down trehalose, a sugar found in insect bodies. The brush border also has its own form of α‐amylase to degrade any starch that fails to be broken down by pancreatic α‐amylase . An
α‐dextrinase is also found in the brush border to break down the α(1→6) links between glucose molecules in the limit dex-trins liberated during starch digestion by pancreatic α‐amylase in the lumen of the intestine. In the typical monogastric diet about 80% of ingested nonstructural carbohydrate is glucose, and the rest is fructose (or galactose in young milk‐fed animals).
Crossing the apical membrane of villous enterocytesThe hexose (e.g., glucose and galactose) and pentose (e.g., fruc-tose) sugars liberated by the brush border enzymes are too large to cross the apical membrane easily. Their concentration over the apical membrane rises following a meal so there is a concentration gradient that can help them cross into the cytosol. Glucose and galactose can be transported using a hexose trans-porter molecule (SGLT‐1) in the apical membrane (Figure 43.11). This protein binds hexose sugars and also binds a Na+ ion (provided by secretions of the crypt enterocytes). The combined force provided by the hexose concentration gradient and the electrochemical force of Na+ moving into the cytosol can push the glucose to the interior of the cell. Fructose is also a six‐carbon sugar but its ketone group gives it slightly different properties. Fructose can be absorbed with the help of a pentose transporter protein (GLUT‐5) in the apical and basolateral membranes, allowing it to cross into the cytosol by facilitated diffusion independent of Na+.
Crossing the basolateral membrane of villous enterocytesThe concentration of hexoses and pentoses will increase within the cytosol of the enterocytes and at the basolateral membrane to concentrations that exceed the concentration in the extracel-lular fluid. Both hexoses and fructose diffuse across the baso-lateral membrane into the extracellular fluid facilitated by a transporter molecule (GLUT‐2 for hexoses, GLUT‐5 for fruc-tose). The Na+ ions that accompanied the hexose sugars across the apical membrane are pumped into the extracellular fluid by the 3Na+/2K+ electrogenic pump residing in the basolateral membrane at the expense of an ATP. The Na+ may be removed from the blood by crypt secretory cells and returned to the lumen to assist facilitated transport of other hexoses and amino acids by villous cells.
It is important to consider why the intestine does not digest all dietary starch and disaccharides to their constituent hexoses and pentoses in the lumen. The reason digestion is completed at the brush border is to prevent the osmolarity of the lumen contents from rising too high and drawing excessive amounts of water into the lumen. By liberating the hexoses and pentoses in the brush border they can be absorbed almost as soon as they are liberated, preventing a rise in osmolarity of the lumen contents. It is also important to note that SGLT‐1, GLUT‐2, and GLUT‐5 are insulin‐independent transporters: the intestinal cells absorb sugars during insulin deficiency as well as they do during insulin sufficiency.
Dietdisaccharides
Fructose
BBEnzymes
Fructose
Hexose
Hexose
HexoseNa+
Na+
Hexose
Diet starch Pancreaticamylase
Fructose
Fructose
SGLT-1 GLUT-5
Figure 44.11 Starch is converted to glucose, maltose, and limit dextrins by amylase in the lumen of the intestine. Brush border enzymes (BB enzymes) such as maltase, lactase, sucrase, and dextrinase convert dietary disaccharides (e.g., maltose, lactose, sucrose) and limit dextrins to single hexose (glucose and galactose) or pentose (fructose) molecules. Hexoses in the brush border are brought into the cell using a Na+‐linked glucose transporter (SGLT‐1). Pentose sugars use a Na+‐independent facilitated transporter (GLUT‐5). At the basolateral membrane, both hexose (GLUT‐2 transporter) and pentose sugars use facilitated transport diffusion to enter the extracellular fluid down their concentration gradient.
Sucrasa Matasa Maltotriasa Lactasa Amilasa Dextrinasa
Enzimas
Dieta Almidón
Fructosa
Amilasa pancreá&ca
Amilasa salival
Hexosa
Hexosa
Hexosa
Fructosa
Fructosa
Fructosa Hexosa
dextrina limite ( alfa 1-‐6)
sacarosa
GLUT-‐5
GLUT-‐5
3 Na+
2 K+
ATP ADP
Na+
Na+
SGLT-‐1
¿Gasto de energía asociado al movimiento de hexosas?
maltosa
matotriosa Glucosa
¿?

Diges&ón y absorción de lípidos

Lípidos (Trigliceridos)
Píloro
(¿Lipasa?)
Mezclado gástrico CCK
Acinos Pancreá&cos Higado
CCK: Colecistoquinina; CEE: Célula del sistema endocrino entérico

514 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on
, A
bso
rpti
on
, an
d M
etab
olis
m
Digestion and absorption of fat
Dietary fats are generally in the form of triglycerides. Their diges-tion may begin in the mouth as lingual glands produce pharyn-geal lipase that converts triglycerides to fatty acids, monoglycerides, and diglycerides. This enzyme is relatively stable in acid and is thought to play a role in digestion of milk fat by neonates that may not be producing the full complement of pancreatic enzymes and liver bile. In general, the amount of fat that is digested by pharyn-geal lipase has a negligible effect in normal fat digestion. The first step in fat digestion takes place in the stomach where the dietary fats are subjected to the churning action of stomach contractions. This causes dietary fats to form an emulsion with water – a suspension of fine droplets of fat in water. This is often aided by incorporation of dietary phospholipids into the emulsion. The emulsified fat droplets then enter the duodenum (Figure 44.12). The presence of fats in the duodenum elicits secretion of CCK by crypt enteroendocrine cells. CCK causes the pancreas to secrete
enzymes and also causes contraction of the gallbladder (not pre-sent in the horse or rat). Several critical enzymes can be found in pancreatic secretions: pancreatic lipase, secreted in an active form; colipase, secreted in an inactive pro‐colipase form; phos-pholipases, secreted in an inactive form (cleave phospholipids of cell membranes); and cholesterol esterase, secreted in the active form. Just as with proteolytic enzymes, the inactive lipid‐digest-ing enzymes are activated in the lumen of the duodenum on cleavage by the enzyme trypsin. Bile contains the bile salts that play several roles in fat digestion.
The emulsified fat droplet entering the duodenum is too hydrophobic and too large for pancreatic lipase to access the droplet and begin to break down the triglycerides. Bile salts are essentially detergents produced in the liver by combining cholesterol with an amino acid. One end of the molecule, com-posed of the cholesterol moiety, is hydrophobic which allows it to form ionic bonds with hydrophobic fatty acids. The other end is very hydrophilic due to the amino acid component. Bile salts surround the emulsified fat droplet and break it into smaller fat droplets suspended in the water in the lumen. This increases the surface area available for enzymatic degradation of the triglycerides to occur. Colipase must bind to pancreatic lipase to allow it to be fully active and this complex then begins to digest triglycerides to monoglycerides and two fatty acids at the surface of the small fat droplet created by the bile salts. Cholesterol esterase may free cholesterol from the droplet and phospholipase liberates fatty acids and monoglyceride from phospholipids. As the lipolytic action of these enzymes prog-resses, more bile salts surround the liberated fatty acids, mono-glycerides, cholesterol, and fat‐soluble vitamins (e.g., vitamins A, D, and E) that were in the diet to form small bile salt‐covered structures known as micelles.
1 What happens to fat in the stomach?
2 Colipase is secreted from the pancreas as pro‐colipase. What is its function and why is it not secreted in an active form?
3 What are the end products of lipase digestion of fat?
4 What is the function of bile salts in the process of fat digestion? How do they function in fat absorption?
5 What happens after the micelle contacts the apical surface of the villous cell?
6 Why are monoglycerides and fatty acids converted back to triglycerides inside enterocytes?
7 What is an apolipoprotein and what does it do?
8 What is HDL?
CCK
Stomach
Emulsi!edfats
Dietfats
Enzymes Gallbladder
Colipase TrypsinLipase
Hydrophilic
Lipophilic
BileSalt
Fatty acids andmonoglyceridesand cholesteroland bile salts
MicelleEEC
Inactivepro-colipase
Fatty acids andmonoglyceridesand cholesterol
TriglycerideCholesterol
Figure 44.12 Luminal phase of fat digestion. Churning of the stomach emulsifies dietary lipids. As these fats enter the duodenum they stimulate enteroendocrine cells (EEC) to secrete cholecystokinin (CCK). CCK stimulates the pancreas to release digestive enzymes including lipase and pro‐colipase needed for fat digestion. CCK also stimulates the gallbladder to contract causing secretion of bile salts into the lumen. Pro‐colipase is cleaved by trypsin to form active colipase, which is a cofactor needed for full activity of lipase. Lipase, colipase, and the bile salts work together on the emulsified fat to convert the triglycerides to monoglycerides and free fatty acids. The liberated fatty acids and monoglycerides, as well as cholesterol and fat‐soluble vitamins, are surrounded by bile salts to form micelles. Micelles are several hundred fold smaller than the emulsified fat droplet.
Grasas dietarias
514 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Digestion and absorption of fat
Dietary fats are generally in the form of triglycerides. Their diges-tion may begin in the mouth as lingual glands produce pharyn-geal lipase that converts triglycerides to fatty acids, monoglycerides, and diglycerides. This enzyme is relatively stable in acid and is thought to play a role in digestion of milk fat by neonates that may not be producing the full complement of pancreatic enzymes and liver bile. In general, the amount of fat that is digested by pharyn-geal lipase has a negligible effect in normal fat digestion. The first step in fat digestion takes place in the stomach where the dietary fats are subjected to the churning action of stomach contractions. This causes dietary fats to form an emulsion with water – a suspension of fine droplets of fat in water. This is often aided by incorporation of dietary phospholipids into the emulsion. The emulsified fat droplets then enter the duodenum (Figure 44.12). The presence of fats in the duodenum elicits secretion of CCK by crypt enteroendocrine cells. CCK causes the pancreas to secrete
enzymes and also causes contraction of the gallbladder (not pre-sent in the horse or rat). Several critical enzymes can be found in pancreatic secretions: pancreatic lipase, secreted in an active form; colipase, secreted in an inactive pro‐colipase form; phos-pholipases, secreted in an inactive form (cleave phospholipids of cell membranes); and cholesterol esterase, secreted in the active form. Just as with proteolytic enzymes, the inactive lipid‐digest-ing enzymes are activated in the lumen of the duodenum on cleavage by the enzyme trypsin. Bile contains the bile salts that play several roles in fat digestion.
The emulsified fat droplet entering the duodenum is too hydrophobic and too large for pancreatic lipase to access the droplet and begin to break down the triglycerides. Bile salts are essentially detergents produced in the liver by combining cholesterol with an amino acid. One end of the molecule, com-posed of the cholesterol moiety, is hydrophobic which allows it to form ionic bonds with hydrophobic fatty acids. The other end is very hydrophilic due to the amino acid component. Bile salts surround the emulsified fat droplet and break it into smaller fat droplets suspended in the water in the lumen. This increases the surface area available for enzymatic degradation of the triglycerides to occur. Colipase must bind to pancreatic lipase to allow it to be fully active and this complex then begins to digest triglycerides to monoglycerides and two fatty acids at the surface of the small fat droplet created by the bile salts. Cholesterol esterase may free cholesterol from the droplet and phospholipase liberates fatty acids and monoglyceride from phospholipids. As the lipolytic action of these enzymes prog-resses, more bile salts surround the liberated fatty acids, mono-glycerides, cholesterol, and fat‐soluble vitamins (e.g., vitamins A, D, and E) that were in the diet to form small bile salt‐covered structures known as micelles.
1 What happens to fat in the stomach?
2 Colipase is secreted from the pancreas as pro‐colipase. What is its function and why is it not secreted in an active form?
3 What are the end products of lipase digestion of fat?
4 What is the function of bile salts in the process of fat digestion? How do they function in fat absorption?
5 What happens after the micelle contacts the apical surface of the villous cell?
6 Why are monoglycerides and fatty acids converted back to triglycerides inside enterocytes?
7 What is an apolipoprotein and what does it do?
8 What is HDL?
CCK
Stomach
Emulsi!edfats
Dietfats
Enzymes Gallbladder
Colipase TrypsinLipase
Hydrophilic
Lipophilic
BileSalt
Fatty acids andmonoglyceridesand cholesteroland bile salts
MicelleEEC
Inactivepro-colipase
Fatty acids andmonoglyceridesand cholesterol
TriglycerideCholesterol
Figure 44.12 Luminal phase of fat digestion. Churning of the stomach emulsifies dietary lipids. As these fats enter the duodenum they stimulate enteroendocrine cells (EEC) to secrete cholecystokinin (CCK). CCK stimulates the pancreas to release digestive enzymes including lipase and pro‐colipase needed for fat digestion. CCK also stimulates the gallbladder to contract causing secretion of bile salts into the lumen. Pro‐colipase is cleaved by trypsin to form active colipase, which is a cofactor needed for full activity of lipase. Lipase, colipase, and the bile salts work together on the emulsified fat to convert the triglycerides to monoglycerides and free fatty acids. The liberated fatty acids and monoglycerides, as well as cholesterol and fat‐soluble vitamins, are surrounded by bile salts to form micelles. Micelles are several hundred fold smaller than the emulsified fat droplet.
Estómago
Vesícula biliar
Hidro`lica
Lipo`lica
Sales Biliares
Triglicéridos Colesterol
Triglicéridos Colesterol Vit A,E,D,K
CCK
Lipasa Colipasa Tripsina
pro-‐Colipasa inac7va
Enzimas
Micela
Estómago
Ácidos grasos Monoglicéridos Colesterol Vit A, E, D, K
CCK: Colecistoquinina; CEE: Célula del sistema endocrino entérico
CEE
Otras enzimas pancreá&cas necesarias para la diges&ón de los lípidos de la dieta: colesterol esterasa y fosfolipasa
Chapter 44: Digestion and Absorption of Nutrients 515
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Moving lipids across the apical membrane of villous enterocytesFatty acids and monoglycerides alone are not able to access the apical membrane of the villous enterocytes for absorption as they cannot cross the unstirred water layer and glycocalyx due to their hydrophobic nature. However, the micelle, carrying its load of lipids, is surrounded by bile salts with their hydrophilic end protruding. This allows them to traverse the unstirred water layer and glycocalyx to reach the apical surface of the enterocyte. The micelle comes in direct contact with the apical membrane and the lipophilic contents of the micelle diffuse through the apical cell membrane into the cytosol down their concentration gradient (Figure 44.13). The bile salts themselves are too water‐soluble to enter the the cell. The bile salts reenter the lumen and collect another load of lipids to be carried to another villous enterocyte. They can do this numerous times before they are finally swept to the ileum. In the ileum, specific bile salt transporters absorb the bile salts via receptor‐mediated endocytosis and carry them in the portal circulation back to the liver where they can be absorbed by hepatocytes and be again excreted into the bile. In humans, it is estimated that each bile salt is recycled twice to absorb the fat in one meal.
At the brush border of the apical membrane any diglycerides or triglycerides that managed to be incorporated into the micelle are broken down by a brush border lipase to monoglyceride and fatty acids, which quickly diffuse into the cell. Once the mono-glycerides and fatty acids enter the cell, they are quickly taken
to the smooth endoplasmic reticulum and converted back into triglycerides. This serves two purposes: it reduces the osmolarity of the cytosol and it removes fatty acids and mono-glycerides from the cytosol. This maintains the fatty acid and monoglyceride gradient forcing the fatty acids and monoglycer-ides across the apical membrane so that the entire lipid load of the micelle can diffuse into the cytosol compartment.
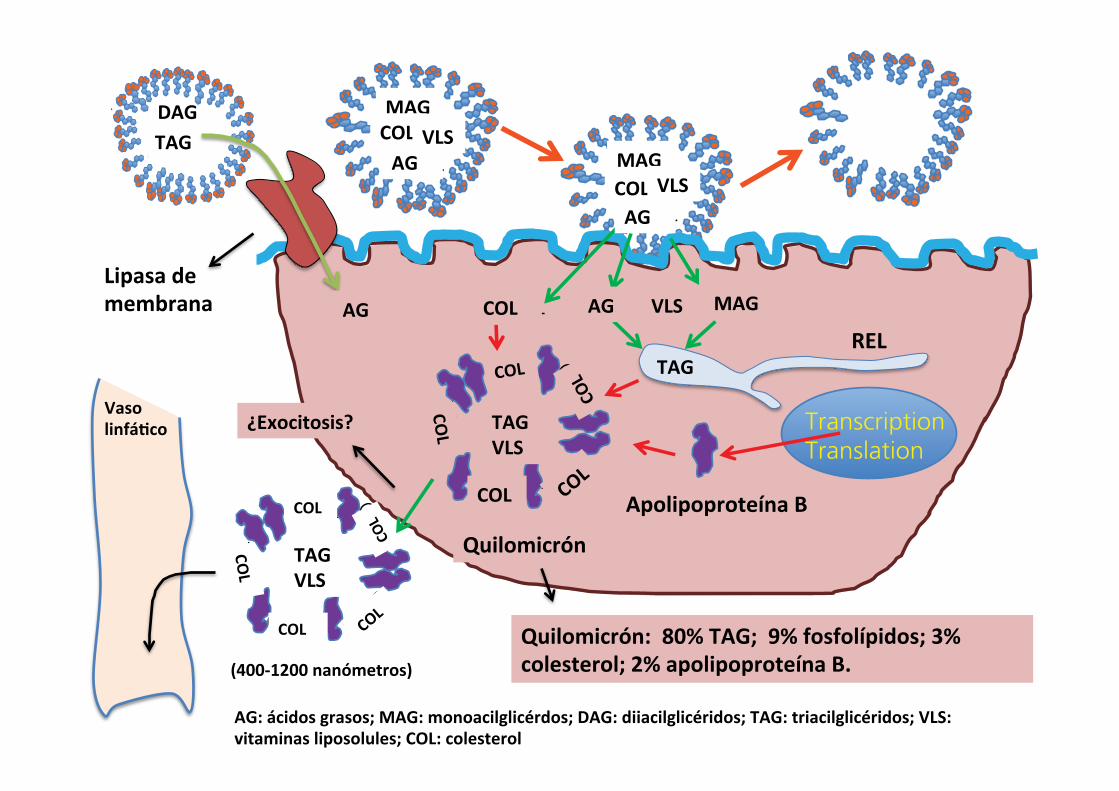
Moving lipids across the basolateral membrane of villous enterocytesThe newly formed triglycerides and cholesterol (and fat‐soluble vitamins) in the cytosol are too water‐insoluble to move out of the cell across the basolateral membrane and too insoluble to circulate in the blood (Figure 44.13). The solution that has evolved is to package these lipid materials with lipopro-teins (apolipoproteins). An apolipoprotein has hydrophilic and hydrophobic ends and forms a special structure that has many of the properties of cell membranes, i.e., contains phospholipids and cholesterol. The apolipoproteins surrounding a collection of triglyceride molecules form a structure called a chylomicron. The chylomicron comprises about 80% triglyceride, which forms the interior of the chylomicron. The outer surface is composed of phosphoglycerides (9%), cholesterol (3%), and apolipoprotein B (2%). It is believed that chylomicrons cross the basolateral membrane of the villous enterocytes by exocytosis into the extracellular fluid. They are too large (400–1200 nm) to enter the portal circulation so instead they enter the lacteal vessels within the lamina propria and then the lymphatic circulation to join the blood circulation via the thoracic duct.
Fate of chylomicrons in the circulationWhen the chylomicron is released from the enterocyte the main lipoprotein on its surface is apolipoprotein B48. As the chylomicron circulates in the blood it will encounter high‐density lipoproteins (HDLs) released by the liver. The HDLs are covered with apolipoproteins C and E on their surface. When the chylomicron encounters HDL, it unloads some of its triglyceride to the HDL and in turn receives some apolipopro-tein C and E, which is incorporated on the surface of the chylo-micron. Apolipoprotein C receptors exist on adipose, mammary, and skeletal and cardiac muscle. As the chylomicrons pass through these tissues, the apolipoprotein C on the surface can bind the chylomicron to its receptor on these tissues. The tri-glycerides are transported into the tissue and the now empty chylomicron remnant is released back into the circulation. As it passes through the liver sinusoids, the apolipoprotein E on the surface of the chylomicron remnant becomes bound to apolipo-protein E receptors on the hepatocytes and the chylomicron remnant is taken into the hepatocyte by endocytosis. The liver can use the remaining triglycerides for energy and can store vitamins A, D, and E that were in the chylomicrons or the liver can repackage the triglyceride with the cholesterol and phospholipids into other lipoprotein particles such as very low density lipoproteins (VLDL) and HDL. VLDL can deliver triglyceride to peripheral tissues such as adipose, muscle, and
FAMGLYCCHOL FA
MGLYCCHOL
FA MGLYC
TG
CHOL
CHOL
CHOL
CHOL
CHOL
CHOL
CHOLCHOL
CHOL
CH
OL
CH
OL
TranscriptionTranslation
Apolipoprotein
TG
TG
Figure 44.13 Brush border phase of lipid digestion and absorption. Lipids packaged into the micelle are able to cross the unstirred water layer above the enterocytes. On making contact the fatty acids (FA), monoglycerides (MGLYC), cholesterol (CHOL), and fat‐soluble vitamins diffuse across the apical membrane to the cytosol. The FA and MGLYC are combined to reform triglycerides (TG), encouraging further diffusion of FA and MGLYC from the micelle. The empty micelle bile salts return to the lumen to collect another load of FA and MGLYC. The enterocyte produces apolipoproteins that are combined with cholesterol to form a chylomicron structure which surrounds cytosol TG and fat‐soluble vitamins. The chylomicron is then exocytosed across the basolateral membrane. It is too large to enter venules so it enters the lacteal and lymphatics to reach the thoracic duct.
514 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII: D
igest
ion
, A
bso
rpti
on
, an
d M
eta
bo
lism
Digestion and absorption of fat
Dietary fats are generally in the form of triglycerides. Their diges-tion may begin in the mouth as lingual glands produce pharyn-geal lipase that converts triglycerides to fatty acids, monoglycerides, and diglycerides. This enzyme is relatively stable in acid and is thought to play a role in digestion of milk fat by neonates that may not be producing the full complement of pancreatic enzymes and liver bile. In general, the amount of fat that is digested by pharyn-geal lipase has a negligible effect in normal fat digestion. The first step in fat digestion takes place in the stomach where the dietary fats are subjected to the churning action of stomach contractions. This causes dietary fats to form an emulsion with water – a suspension of fine droplets of fat in water. This is often aided by incorporation of dietary phospholipids into the emulsion. The emulsified fat droplets then enter the duodenum (Figure 44.12). The presence of fats in the duodenum elicits secretion of CCK by crypt enteroendocrine cells. CCK causes the pancreas to secrete
enzymes and also causes contraction of the gallbladder (not pre-sent in the horse or rat). Several critical enzymes can be found in pancreatic secretions: pancreatic lipase, secreted in an active form; colipase, secreted in an inactive pro‐colipase form; phos-pholipases, secreted in an inactive form (cleave phospholipids of cell membranes); and cholesterol esterase, secreted in the active form. Just as with proteolytic enzymes, the inactive lipid‐digest-ing enzymes are activated in the lumen of the duodenum on cleavage by the enzyme trypsin. Bile contains the bile salts that play several roles in fat digestion.
The emulsified fat droplet entering the duodenum is too hydrophobic and too large for pancreatic lipase to access the droplet and begin to break down the triglycerides. Bile salts are essentially detergents produced in the liver by combining cholesterol with an amino acid. One end of the molecule, com-posed of the cholesterol moiety, is hydrophobic which allows it to form ionic bonds with hydrophobic fatty acids. The other end is very hydrophilic due to the amino acid component. Bile salts surround the emulsified fat droplet and break it into smaller fat droplets suspended in the water in the lumen. This increases the surface area available for enzymatic degradation of the triglycerides to occur. Colipase must bind to pancreatic lipase to allow it to be fully active and this complex then begins to digest triglycerides to monoglycerides and two fatty acids at the surface of the small fat droplet created by the bile salts. Cholesterol esterase may free cholesterol from the droplet and phospholipase liberates fatty acids and monoglyceride from phospholipids. As the lipolytic action of these enzymes prog-resses, more bile salts surround the liberated fatty acids, mono-glycerides, cholesterol, and fat‐soluble vitamins (e.g., vitamins A, D, and E) that were in the diet to form small bile salt‐covered structures known as micelles.
1 What happens to fat in the stomach?
2 Colipase is secreted from the pancreas as pro‐colipase. What is its function and why is it not secreted in an active form?
3 What are the end products of lipase digestion of fat?
4 What is the function of bile salts in the process of fat digestion? How do they function in fat absorption?
5 What happens after the micelle contacts the apical surface of the villous cell?
6 Why are monoglycerides and fatty acids converted back to triglycerides inside enterocytes?
7 What is an apolipoprotein and what does it do?
8 What is HDL?
CCK
Stomach
Emulsi!edfats
Dietfats
Enzym
es Gallbladder
Colipase TrypsinLipase
Hydrophilic
Lipophilic
BileSalt
Fatty acids andmonoglyceridesand cholesteroland bile salts
MicelleEEC
Inactivepro-colipase
Fatty acids andmonoglyceridesand cholesterol
TriglycerideCholesterol
Figure 44.12 Luminal phase of fat digestion. Churning of the stomach emulsifies dietary lipids. As these fats enter the duodenum they stimulate enteroendocrine cells (EEC) to secrete cholecystokinin (CCK). CCK stimulates the pancreas to release digestive enzymes including lipase and pro‐colipase needed for fat digestion. CCK also stimulates the gallbladder to contract causing secretion of bile salts into the lumen. Pro‐colipase is cleaved by trypsin to form active colipase, which is a cofactor needed for full activity of lipase. Lipase, colipase, and the bile salts work together on the emulsified fat to convert the triglycerides to monoglycerides and free fatty acids. The liberated fatty acids and monoglycerides, as well as cholesterol and fat‐soluble vitamins, are surrounded by bile salts to form micelles. Micelles are several hundred fold smaller than the emulsified fat droplet.
DAG TAG
MAG COL AG
COL AG
MAG
MAG COL AG
TAG COL
COL
COL
TAG VLS
COL
COL
COL
TAG VLS
Apolipoproteína B
REL
Quilomicrón
Quilomicrón: 80% TAG; 9% fosfolípidos; 3% colesterol; 2% apolipoproteína B.
Vaso linfá&co
(400-‐1200 nanómetros)
¿Exocitosis?
Lipasa de membrana AG
VLS
VLS
VLS
AG: ácidos grasos; MAG: monoacilglicérdos; DAG: diiacilglicéridos; TAG: triacilglicéridos; VLS: vitaminas liposolules; COL: colesterol

Chapter 44: Digestion and Absorption of Nutrients 505
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Active transport across the cell membraneActive transport across membranes implies that energy will have to be provided, usually in the form of ATP, to move ions or molecules. Active transport is usually required when moving a substance against its electrochemical gradient. The cell mem-brane proteins that carry out this function are often referred to as pumps since they are generally moving ions from an area of low concentration to an area of higher concentration. The active transport pumps are generally highly specific in the substance they will pump, but can develop very high concentration gradients across membranes.
An example of such a pump is the electrogenic Na+/K+‐ATPase pump. This pumping protein utilizes the energy in one ATP molecule to move three sodium atoms from inside the cell to the outside of the cell against sodium’s electrical and concentration gradient in exchange for two potassium atoms moving into the cell down their electrical gradient but against the concentration gradient. This pump keeps extracellular sodium concentration high and intracellular potassium concentration high. It also gen-erates an electrical potential difference across the cell membrane, keeping the inside negative relative to the outside of the cell.
Nonionic diffusionWeak acids and weak bases are compounds that exist in both a dissociated and a nondissociated state. In the nondissociated state they are both water‐ and lipid‐soluble. In this state they have no charge and being lipid‐soluble they can freely cross the lipid bilayer of cell membranes. In the dissociated state their charge makes them unable to cross the lipid bilayer and that charge also makes them soluble in water only. The nondissoci-ated and dissociated forms of weak acids and weak bases are in equilibrium and the concentration of nondissociated and disso-ciated forms is dependent on the pH of the solution they occupy. The pH at which 50% of the weak acid or base is in the dissoci-ated form and 50% is in the nondissociated form is called the pKa for the compound. How this affects the equilibrium and concentrations of nondissociated and dissociated forms is best illustrated using acetic acid as an example. Acetic acid in water exists in the nondissociated state, HAc, and in the dissociated state, Ac–, as described by the following equation:
HAc H Ac
The pKa for acetic acid is 4.76. In a pH 4.76 solution, 50% of the acetic acid will exist in the nondissociated state, designated HAc, and 50% will exist in the dissociated state as Ac–. If the pH of the solution is 5.76 (and remembering that pH is a logarithmic scale), the reduction in H+ ions shifts the equilibrium further to the right and now just 10% of the acetic acid is in the HAc form while 90% is in the Ac– dissociated form. If the solution has a pH of 6.76, not unlike the pH in the colon of the horse, just 1% of the acetic acid is in the nondissociated form, while 99% is in the dissociated form. Using the horse colon cell membrane as an example, the small amount of HAc in the nondissociated form will freely cross the apical membrane down its concentration
gradient to the interior of the cell (Figure 44.4). By removing HAc from the lumen the equilibrium of acetic acid dissociation will be shifted toward the left to replace the lost HAc, allowing another HAc to cross the cell membrane. Once on the other side of the membrane the HAc quickly dissociates to form H+ and Ac–. The Ac– is now trapped inside the cell. However, as long as HAc is being produced in the lumen and is crossing the apical membrane, there will be a second equilibrium set up for acetic acid at the opposite side of the cell near the basolateral mem-brane. Here Ac– and H+ will again be in equilibrium with HAc. As HAc is formed it will move out to the extracellular fluids down its concentration gradient as it is lipid‐soluble and able to freely cross the basolateral cell membrane.
Weak bases also set up similar equilibria when placed in solution:
BaseOH Base OH
Their pKa will generally be in excess of 8.0 and their dissociation is promoted by placing them in more acid solutions. This system is surprisingly efficient and can operate even when the nondisso-ciated form would be expected to comprise less than 0.01% of the total amount of weak acid or weak base present. Nonionic diffusion is the main method used by ruminants and hindgut fermenters to absorb the volatile fatty acids (VFAs) produced by bacterial fermentation of cellulosic plant materials. Most of the drugs utilized in veterinary medicine are weak acids or weak bases and this is the method they utilize to cross cell membranes.
H+ + Ac–
H+ + Ac–
H+ + Ac–
Apical
Basolateral
HAc
HAc
HAc
HAc
Figure 44.4 Nonionic diffusion. Assume a high concentration of acetic acid in the lumen of the colon. It establishes an equilibrium, with a portion existing in the uncharged nondissociated state (HAc) and a portion in the charged dissociated state (Ac–). The lipid bilayer cell membrane is freely permeable to weak acids such as acetate when they are in the nondissociated uncharged state (HAc). Once HAc has crossed the apical membrane it again establishes an equilibrium, with a portion of the acetic acid in the HAc and Ac– states. This equilibrium is also established at the basolateral membrane. The HAc can then cross the basolateral membrane. Once in the extracellular fluid the HAc reestablishes an equilibrium, with some in the HAc and some in the Ac– form. The acetic acid in the charged Ac– form is trapped in the extracellular fluid as it cannot recross the lipid bilayer.
Difusión no iónica: Absorción de ácidos débiles (acé&co, propiónico y buhrico) a través del epitelio de las cámaras de fermetación

508 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Basolateral membrane transportChloride concentration in the extracellular fluid is about 102–108 mmol/L. Inside the enterocyte the Cl– concentration might get as high as 30 mmol/L. Cl– will move to the extracel-lular fluid against its concentration gradient but with its electrical gradient.1 Cl–/K+ cotransporter: K+ will be moving into the extracellular
fluid down its concentration gradient but against its electrical gradient. The combined force of K+ moving down its concentration gradient and Cl– moving down its electrical gradient permits both molecules to overcome the forces resisting this movement (Figure 44.7).
2 Cl– pump: chloride can be actively pumped across the baso-lateral membrane at the expense of an ATP. This mechanism is used primarily in the lower intestine (Figure 44.7).
3 Na+/Cl– pump: sodium and chloride can be actively pumped across the basolateral membrane at the expense of an ATP. This mechanism is used primarily in the lower intestine (Figure 44.7).
Paracellular absorption into the extracellular fluidWhen Cl– concentration is very high, as in the uppermost duo-denum (110–120 mmol/L), the concentration gradient between the lumen and the extracellular fluid (102–108 mmol/L) will
allow Cl– to pass through the tight junctions between adjoining enterocytes directly into the extracellular fluid. This mechanism is important in the first few centimeters of the duodenum and quickly removes a large amount of chloride from the chyme; this action also increases the pH of the digesta in the duodenum (Figure 44.7).
Potassium
Paracellular K+ transportThe bulk of K+ absorption occurs across the tight junctions and K+ moves between cells directly into the extracellular fluid, particularly in the lower small intestine. K+ concentration in extracellular fluid is low (4–6 mmol/L) while the concentration of K+ in the lumen can be many fold higher. The concentration of K+ in the lumen actually increases as the digesta moves down
Table 44.1 Major mechanisms utilized for absorption of electrolytes, hexoses, amino acids, and water in various segments of the intestinal tract.
Mechanism Duodenum Upper jejunum Middle jejunum Lower jejunum Ileum Colon
Na+/Cl– cotransporter + + + + ++ +++Na+/hexose cotransporter ++++++ ++++ + + – –Na+/amino acid cotransporter ++++++ ++++ + + – –Cl–/HCO3
– exchange – – – – ++ +++Paracellular Cl– absorption ++++ – – – – –Paracellular K+ absorption – – – – + +++Water +++ +++ + + ++ +
Venule
Na+Na+Cl–
Na+
Na+
Na+
Na+
Na+ Na+Na+ Na+
Cl–
Cl–
Cl–
Cl–
Cl–Cl–Cl– Cl–
Cl–Cl–
Cl–
Cl–
Cl–
Cl–
Cl–Cl–
Cl–
2Cl–
2Cl–
HCO3–
Cl– HCO3–
ATP
3Na+
3Na+
K+
K+
K+ K+ K+
K+
K+
K+
K+
K+ 2K+
2K+
K+ K+
ATP
ATP
Figure 44.7 Major electrolyte absorption mechanisms. The apical membrane has a Na+/Cl– cotransporter, a Na+/K+/2Cl– cotransporter, and a Cl–/HCO3
– ATPase pump to bring electrolytes from the lumen into the cytosol. The basolateral membrane uses a Na+/Cl– ATPase pump, a K+/Cl– cotransporter, and the 3Na+/2K+ exchange ATPase pump (the electrogenic pump) to move electrolytes from the cytosol into the extracellular fluid. In the lower small intestine and colon most of the K+ is absorbed paracellularly. In the uppermost duodenum Cl– can also be absorbed paracellularly.
1 How do potassium atoms cross from the lumen of the gut to the interior of the villous cell?
2 How do potassium atoms cross from the interior of the cell to the extracellular !uid?
3 Where is paracellular absorption of potassium most likely to occur and why?
Cl-‐
Cl-‐
ATP
Na+ 12-‐15 mM K+ 140 mM Cl-‐ 30 mM (duodeno) Cl-‐ 4mM (íIeon)
Na+ 140 mM K+ 4 mM Cl-‐ 102-‐108 mM
Na+ variable: > LIC; < LEC mM; disminuye hacia el íleon. K+ variable (30-‐40 mM) > LEC; < LIC. Cl-‐ 120 mM en duodeno; disminuye a casi 0 mM en íleon
LIC
LEC
Digesta (Luz del intes&no) Absorción de Na+, K+ y Cl-‐
LIC: líquido intracelular; LEC: líquido extracelular

516 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
mammary gland. The remnant of a VLDL is called a low‐density lipoprotein (LDL). It still contains a large amount of cholesterol and has the unhealthy habit in humans of delivering cholesterol to endothelial cells lining arteries. This is why LDL is often referred to as “bad cholesterol.” The HDL particles released from the liver can actually remove triglyceride and cholesterol from arteries and other peripheral tissues and take these materials to the liver for processing. This property has earned them the moniker of “good cholesterol.”
Absorption of water
Two forces are at work to move water (Figure 44.14): osmotic pressure is generated by the movement of solute from one compartment to another creating an osmotic gradient; hydro-static pressure is a physical force created when water is moved into a confined area. Aquaporin channels exist in the apical and basolateral membranes of enterocytes that allow water to move across the lipid bilayer of the cell membranes to follow the osmotic gradient. These aquaporin channels do not allow any charged ions through them. The key to absorption of water is to absorb solute. Solute is typically absorbed across the apical membrane and crosses the basolateral membrane to enter the lateral space between epithelial cells. This draws water into the lateral space between cells through the aquaporins. As more solute and more water enters this confined space it increases hydrostatic pressure. Distension of the elastic cell membranes of the lateral space also allows the hydrostatic pressure to increase. Under normal circumstances the path of least resistance for this water will be to cross the basement membrane of the fenestrated capillary endothelium to enter the circulation. A critical fact is that the tight junction offers more resistance to flow of water than the fenestrated endothelial basement membrane of the capillary bed. The tight junctions are not impermeable to water, just less permeable than the capillary endothelium. Water chan-nels formed by claudin proteins (not the same as aquaporins) in the tight junction generally resist the flow of water, but this
1 What are aquaporins? Do they allow water to move down or against its osmotic gradient?
2 Why is movement of solute critical for absorption of water?
3 Can water cross the tight junctions? Under what circumstances?
4 How does the !ow of blood at the villous tip affect water absorption?
5 Where is the bulk of water absorbed in the digestive tract?
6 Can the colon compensate for malabsorption in the upper intestine? What can it absorb that might help with water absorption?
SoluteNa, Cl, K,sugars,aminoacids
300mosmol
500mosmol
400mosmol
ArterioleVenule
H2OH2O
H2O
H2OH2O
H2O
H2O
H2O
H2O H2O
H2OH2O AA
AA
AA
AA
AAAA
AANa+
Na+
Na+Na+
Na+
Na+ Na+
Na+
Na+
Na+
Na+
Na+Na+ Na+
Na+
AAAA
AA
AA
AA
AA
600mosmol
Capillary bed
1
3
2
H2OH2OH2O
H2OH2OH2O
H2OH2O
H2O
H2O
Figure 44.14 Water absorption. Water is pulled into areas high in solute. (1) In the villous tip the countercurrent flow of arteriole and venule blood causes solute in the descending venule to diffuse into the ascending arteriole to be carried back to the villous tip. The osmolarity at the villous tip can approach 600 mosmol. (2) At the level of individual enterocytes, water utilizes aquaporin to follow its osmotic gradient, created by absorption of solutes such as Na+ and amino acids (AA) into the extracellular fluid through the enterocytes. Aquaporins are specialized water channels that allow water but not charged ions through their channels. (3) As water moves into the lateral spaces, hydrostatic pressure builds up between the cells causing distension and bulging of the cell walls below the tight junction. The hydrostatic pressure then drives the water and solute into the fenestrated capillaries, which have lower resistance to the flow of water than do the tight junctions.
Absorción de agua
1) Para absorber agua hay que absorber solutos
2) el agua atraviesa las membranas apical y asolateral por canales específicos (acuaporinas). Efecto osmó&co
3) el agua se acumula en el espacio lateral entre 2 enterocitos y aumenta la presión hidrostá&ca.
4) La unión estrecha es más resistente al aumento de la presión en este espacio que la membrana basal y el agua atraviesa la membrana basal hacia el LIC.

Chapter 44: Digestion and Absorption of Nutrients 517
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
resistance can be overcome if osmotic forces are great enough on one or another side of the membrane.
A phenomena that aids absorption of water at the villous tip is the countercurrent movement of solute from vein to arteriole in the lamina propria of a villus. Recall that an arteriole carries blood to the tip of the villus and this arteriole is in close proximity to a venule carrying blood away from the tip of the villus. During digestion and absorption of a meal, the postcapil-lary venules bring large amounts of absorbed solute into the venules. As the venule passes down the villus, solute can diffuse from the venule into the arteriole down its concentration gra-dient (see Figure 44.14). The arteriole brings this solute to the villous tip and eventually the osmolarity of the lamina propria at the villous tip can reach 500–600 mosmol, about twice the osmolarity of plasma. This draws water from the lumen of the intestine into the villous tip and on to the extracellular fluid.
Water balance in the gastrointestinal tractNearly all the water that enters the gastrointestinal tract is removed, so that feces generally contain only small amounts of water. A 20‐kg dog might drink (or ingest with its diet) 600 mL water each day. Salivary glands add 300 mL, gastric juices 600 mL, bile 300 mL, pancreatic secretions 600 mL, small intestine (Brunner’s glands and crypt cell secretions) add another 600 mL, and the colonic mucus adds 50 mL water to the lumen of the gastrointestinal tract for a total of 3050 mL water entering the lumen (Table 44.2). Only about 35 mL, about 1%, of this water exits the tract with the feces. The bulk of the water, about 2650 mL, is absorbed by the small intestine. The duo-denum and jejunum absorb about 1600 mL or 52%. The ileum absorbs about 1060 mL or about 35%, and the colon absorbs 365 mL or 12% of the water entering the gastrointestinal tract. Water is absorbed with close to 99% efficiency. The water balance for the dog is therefore ingested 600 mL, retained 565 mL (which was likely used for respiration humidity and urine production).
Water balance in the horse (and other hindgut fermenters) is somewhat different. A 500‐kg horse would be expected to ingest about 40 L of water each day (Table 44.3). Salivary and gastric juices add 41 L of water, and pancreatic, bile, and small and large intestine secretions add another 63 L for a total of 144 L of water entering the gastrointestinal tract. The small intestine absorbs 79 L of water daily. The cecum absorbs 18 L, the ventral colon absorbs 13 L, the dorsal colon absorbs 18.5 L, and the transverse and descending colon absorb another 8 L of water, for a total of 57.5 L for the large intestine. The horse colon absorbs a much greater amount of water than the monogastric colon. It uses the osmotic force of VFA absorption to absorb much of this water. The net loss of water to the feces is about 7.5 L/day. On balance the horse retains 32.5 L of the 40 L ingested.
Malabsorptive diarrheaBacteria and viruses can damage the tight junctions and/or villous absorptive cells and can interfere with absorption of solute, both the solute that was in the diet and the solute secreted
from crypt cells and in pancreatic, and salivary secretions. The inability to absorb solute will also cause some loss in efficiency of water absorption. If water absorption efficiency in the 20‐kg dog drops from nearly 99% to 90% due to enteritis in a section of the intestine, fecal water content will increase from 35 to 275 mL, which will create a very watery feces. If damage to the gastrointestinal tract is restricted to the colon and no water is absorbed by the colon, fecal water content will only increase by the 365 mL the colon is expected to absorb. However, damage to large segments of the small intestine can cause loss of up to 2650 mL water.
Bacterial and viral infections can not only destroy villous absorptive cells, they can initiate inflammation reactions that generally cause hypersecretion by the crypt and goblet cells in an attempt to flush away the offending pathogens. This can greatly increase the amount of fluid lost with the feces. Bacterial infections tend to be fairly localized and cause local damage to either the small or large intestine. Rotaviruses attack the cells at the tips of villi causing some malabsorption. Coronaviruses, such as the causative agents of transmissible gastroenteritis and porcine epidemic diarrhea, kill villous entrocytes the full length of the villus and are therefore much more severe infections. Parvoviruses causing canine parvo and feline panleukopenia attack rapidly dividing cells – in the intestine these are the crypt
Table 44.2 Water secretion and absorption in a 20‐kg dog.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 600 mL —Salivary glands 300 mL —Gastric juice 600 mL —Bile 300 mL —Pancreatic secretions
600 mL —
Small intestine 600 mL 2650 mLColon 50 mL 365 mLTotals 3050 mL 3015 mL = 35 mL
in feces
Table 44.3 Water secretion and absorption in a 500‐kg horse.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 40 L —Salivary glands 30 L —Gastric juice 11 L —Bile 5 L —Pancreatic secretions
10 L —
Small intestine 47 L 79 LColon 1 L 57.5 LTotals 144 L 136.5 L = 7.5 L
in feces
Alimento y agua
Saliva Jugo gástrico
Bilis Secreciones pancreá7cas
Intes7no delgado
Colon Total
en heces
Agua Ingerida y secretada hacia la luz
Agua absorbida de la luz
Balance de agua en el tubo diges7vo de un perro de 20 kg de peso
Chapter 44: Digestion and Absorption of Nutrients 517
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
resistance can be overcome if osmotic forces are great enough on one or another side of the membrane.
A phenomena that aids absorption of water at the villous tip is the countercurrent movement of solute from vein to arteriole in the lamina propria of a villus. Recall that an arteriole carries blood to the tip of the villus and this arteriole is in close proximity to a venule carrying blood away from the tip of the villus. During digestion and absorption of a meal, the postcapil-lary venules bring large amounts of absorbed solute into the venules. As the venule passes down the villus, solute can diffuse from the venule into the arteriole down its concentration gra-dient (see Figure 44.14). The arteriole brings this solute to the villous tip and eventually the osmolarity of the lamina propria at the villous tip can reach 500–600 mosmol, about twice the osmolarity of plasma. This draws water from the lumen of the intestine into the villous tip and on to the extracellular fluid.
Water balance in the gastrointestinal tractNearly all the water that enters the gastrointestinal tract is removed, so that feces generally contain only small amounts of water. A 20‐kg dog might drink (or ingest with its diet) 600 mL water each day. Salivary glands add 300 mL, gastric juices 600 mL, bile 300 mL, pancreatic secretions 600 mL, small intestine (Brunner’s glands and crypt cell secretions) add another 600 mL, and the colonic mucus adds 50 mL water to the lumen of the gastrointestinal tract for a total of 3050 mL water entering the lumen (Table 44.2). Only about 35 mL, about 1%, of this water exits the tract with the feces. The bulk of the water, about 2650 mL, is absorbed by the small intestine. The duo-denum and jejunum absorb about 1600 mL or 52%. The ileum absorbs about 1060 mL or about 35%, and the colon absorbs 365 mL or 12% of the water entering the gastrointestinal tract. Water is absorbed with close to 99% efficiency. The water balance for the dog is therefore ingested 600 mL, retained 565 mL (which was likely used for respiration humidity and urine production).
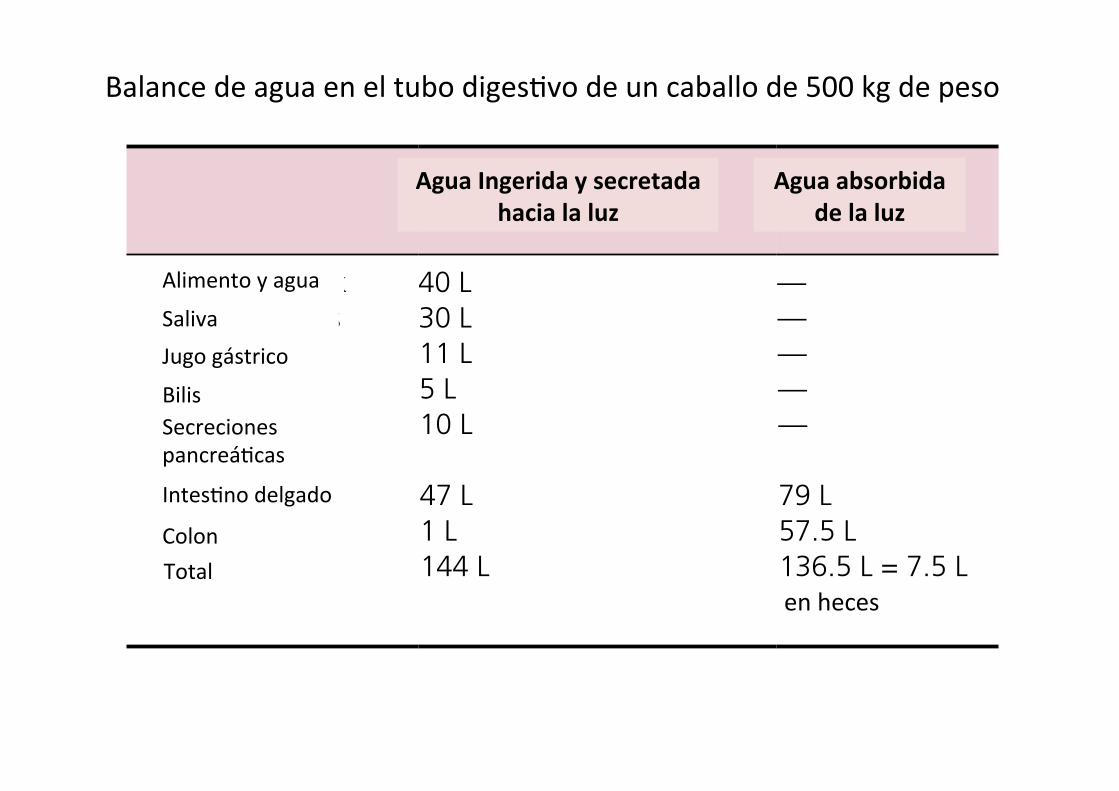
Water balance in the horse (and other hindgut fermenters) is somewhat different. A 500‐kg horse would be expected to ingest about 40 L of water each day (Table 44.3). Salivary and gastric juices add 41 L of water, and pancreatic, bile, and small and large intestine secretions add another 63 L for a total of 144 L of water entering the gastrointestinal tract. The small intestine absorbs 79 L of water daily. The cecum absorbs 18 L, the ventral colon absorbs 13 L, the dorsal colon absorbs 18.5 L, and the transverse and descending colon absorb another 8 L of water, for a total of 57.5 L for the large intestine. The horse colon absorbs a much greater amount of water than the monogastric colon. It uses the osmotic force of VFA absorption to absorb much of this water. The net loss of water to the feces is about 7.5 L/day. On balance the horse retains 32.5 L of the 40 L ingested.
Malabsorptive diarrheaBacteria and viruses can damage the tight junctions and/or villous absorptive cells and can interfere with absorption of solute, both the solute that was in the diet and the solute secreted
from crypt cells and in pancreatic, and salivary secretions. The inability to absorb solute will also cause some loss in efficiency of water absorption. If water absorption efficiency in the 20‐kg dog drops from nearly 99% to 90% due to enteritis in a section of the intestine, fecal water content will increase from 35 to 275 mL, which will create a very watery feces. If damage to the gastrointestinal tract is restricted to the colon and no water is absorbed by the colon, fecal water content will only increase by the 365 mL the colon is expected to absorb. However, damage to large segments of the small intestine can cause loss of up to 2650 mL water.
Bacterial and viral infections can not only destroy villous absorptive cells, they can initiate inflammation reactions that generally cause hypersecretion by the crypt and goblet cells in an attempt to flush away the offending pathogens. This can greatly increase the amount of fluid lost with the feces. Bacterial infections tend to be fairly localized and cause local damage to either the small or large intestine. Rotaviruses attack the cells at the tips of villi causing some malabsorption. Coronaviruses, such as the causative agents of transmissible gastroenteritis and porcine epidemic diarrhea, kill villous entrocytes the full length of the villus and are therefore much more severe infections. Parvoviruses causing canine parvo and feline panleukopenia attack rapidly dividing cells – in the intestine these are the crypt
Table 44.2 Water secretion and absorption in a 20‐kg dog.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 600 mL —Salivary glands 300 mL —Gastric juice 600 mL —Bile 300 mL —Pancreatic secretions
600 mL —
Small intestine 600 mL 2650 mLColon 50 mL 365 mLTotals 3050 mL 3015 mL = 35 mL
in feces
Table 44.3 Water secretion and absorption in a 500‐kg horse.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 40 L —Salivary glands 30 L —Gastric juice 11 L —Bile 5 L —Pancreatic secretions
10 L —
Small intestine 47 L 79 LColon 1 L 57.5 LTotals 144 L 136.5 L = 7.5 L
in feces
Chapter 44: Digestion and Absorption of Nutrients 517
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
resistance can be overcome if osmotic forces are great enough on one or another side of the membrane.
A phenomena that aids absorption of water at the villous tip is the countercurrent movement of solute from vein to arteriole in the lamina propria of a villus. Recall that an arteriole carries blood to the tip of the villus and this arteriole is in close proximity to a venule carrying blood away from the tip of the villus. During digestion and absorption of a meal, the postcapil-lary venules bring large amounts of absorbed solute into the venules. As the venule passes down the villus, solute can diffuse from the venule into the arteriole down its concentration gra-dient (see Figure 44.14). The arteriole brings this solute to the villous tip and eventually the osmolarity of the lamina propria at the villous tip can reach 500–600 mosmol, about twice the osmolarity of plasma. This draws water from the lumen of the intestine into the villous tip and on to the extracellular fluid.
Water balance in the gastrointestinal tractNearly all the water that enters the gastrointestinal tract is removed, so that feces generally contain only small amounts of water. A 20‐kg dog might drink (or ingest with its diet) 600 mL water each day. Salivary glands add 300 mL, gastric juices 600 mL, bile 300 mL, pancreatic secretions 600 mL, small intestine (Brunner’s glands and crypt cell secretions) add another 600 mL, and the colonic mucus adds 50 mL water to the lumen of the gastrointestinal tract for a total of 3050 mL water entering the lumen (Table 44.2). Only about 35 mL, about 1%, of this water exits the tract with the feces. The bulk of the water, about 2650 mL, is absorbed by the small intestine. The duo-denum and jejunum absorb about 1600 mL or 52%. The ileum absorbs about 1060 mL or about 35%, and the colon absorbs 365 mL or 12% of the water entering the gastrointestinal tract. Water is absorbed with close to 99% efficiency. The water balance for the dog is therefore ingested 600 mL, retained 565 mL (which was likely used for respiration humidity and urine production).
Water balance in the horse (and other hindgut fermenters) is somewhat different. A 500‐kg horse would be expected to ingest about 40 L of water each day (Table 44.3). Salivary and gastric juices add 41 L of water, and pancreatic, bile, and small and large intestine secretions add another 63 L for a total of 144 L of water entering the gastrointestinal tract. The small intestine absorbs 79 L of water daily. The cecum absorbs 18 L, the ventral colon absorbs 13 L, the dorsal colon absorbs 18.5 L, and the transverse and descending colon absorb another 8 L of water, for a total of 57.5 L for the large intestine. The horse colon absorbs a much greater amount of water than the monogastric colon. It uses the osmotic force of VFA absorption to absorb much of this water. The net loss of water to the feces is about 7.5 L/day. On balance the horse retains 32.5 L of the 40 L ingested.
Malabsorptive diarrheaBacteria and viruses can damage the tight junctions and/or villous absorptive cells and can interfere with absorption of solute, both the solute that was in the diet and the solute secreted
from crypt cells and in pancreatic, and salivary secretions. The inability to absorb solute will also cause some loss in efficiency of water absorption. If water absorption efficiency in the 20‐kg dog drops from nearly 99% to 90% due to enteritis in a section of the intestine, fecal water content will increase from 35 to 275 mL, which will create a very watery feces. If damage to the gastrointestinal tract is restricted to the colon and no water is absorbed by the colon, fecal water content will only increase by the 365 mL the colon is expected to absorb. However, damage to large segments of the small intestine can cause loss of up to 2650 mL water.
Bacterial and viral infections can not only destroy villous absorptive cells, they can initiate inflammation reactions that generally cause hypersecretion by the crypt and goblet cells in an attempt to flush away the offending pathogens. This can greatly increase the amount of fluid lost with the feces. Bacterial infections tend to be fairly localized and cause local damage to either the small or large intestine. Rotaviruses attack the cells at the tips of villi causing some malabsorption. Coronaviruses, such as the causative agents of transmissible gastroenteritis and porcine epidemic diarrhea, kill villous entrocytes the full length of the villus and are therefore much more severe infections. Parvoviruses causing canine parvo and feline panleukopenia attack rapidly dividing cells – in the intestine these are the crypt
Table 44.2 Water secretion and absorption in a 20‐kg dog.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 600 mL —Salivary glands 300 mL —Gastric juice 600 mL —Bile 300 mL —Pancreatic secretions
600 mL —
Small intestine 600 mL 2650 mLColon 50 mL 365 mLTotals 3050 mL 3015 mL = 35 mL
in feces
Table 44.3 Water secretion and absorption in a 500‐kg horse.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 40 L —Salivary glands 30 L —Gastric juice 11 L —Bile 5 L —Pancreatic secretions
10 L —
Small intestine 47 L 79 LColon 1 L 57.5 LTotals 144 L 136.5 L = 7.5 L
in feces
Alimento y agua
Saliva Jugo gástrico
Bilis Secreciones pancreá7cas
Intes7no delgado
Colon Total
en heces
Balance de agua en el tubo diges7vo de un caballo de 500 kg de peso
Agua Ingerida y secretada hacia la luz
Agua absorbida de la luz