difference in virulence of marine and freshwater isolates...
TRANSCRIPT

1
Difference in virulence of marine and freshwater isolates of viral
hemorrhagic septicemia virus in vivo correlates with in vitro ability to
infect gill epithelial cells and macrophages of rainbow trout
(Oncorhynchus mykiss)
1Bjørn E. Brudeseth,
2Helle F. Skall,
3*Øystein Evensen
1PHARMAQ AS, PO Box 267, N-0213 Oslo, Norway
2Technical University of Denmark, National Veterinary Institute, Hangøvej 2, DK-
8200 Aarhus N, Denmark
3Norwegian School of Veterinary Science, PO Box 8146 Dep, N-0033 Oslo Norway
* Corresponding author
Running title: Virulence traits of VHS virus ACCEPTED
Copyright © 2008, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Virol. doi:10.1128/JVI.01009-08 JVI Accepts, published online ahead of print on 27 August 2008
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

2
ABSTRACT
Two strains of viral hemorrhagic septicemia (VHS) virus with known different
virulence characteristics in vivo were studied (time-course) for their ability to infect
and translocate across a primary culture of gill epithelial cells (GEC) of rainbow trout
(RBT; Oncorhynchus mykiss). The strains included one low-virulent marine strain
(ma-VHSV; ma-1p8) and a highly pathogenic freshwater strain (fw-VHSV; fw-DK-
3592B). Infectivity to trout head kidney macrophages was also studied (time-course)
and difference in in vivo virulence was reconfirmed, the aim being to determine any
correlation between in vivo virulence with in vitro infectivity. The in vitro studies
showed that the fw-VHSV isolate infected and caused cytotoxic effect in monolayers
of GEC (virulence) at early time (2h) and the same virus strain translocated over a
confluent, polarized GEC-layer by 2h post inoculation. The marine isolate did not
infect monolayers of GEC and a delayed translocation across polarized GEC was seen
by 48h post inoculation. Primary cultures of head kidney macrophages were also
infected with fw-VHSV, with a maximum of 9.5% virus positive cells by 3 days post
infection, while for the ma-VHSV strain only 0.5% of the macrophages were positive
by 3 days of culture. In vivo studies showed that the fw-VHSV strain was highly
virulent for RBT fry and caused high mortality, with classical features of VHS. The
ma-VHSV showed very low virulence (only one pool of the sampled dead fish was
VHSV positive). This study has shown that the difference in virulence between ma-
and fw-strains of VHSV following in vivo infection in rainbow trout correlates with in
vitro ability to infect primary cultures of gill epithelial cells and head kidney
macrophages of the same species.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3
INTRODUCTION
Viral hemorrhagic septicemia (VHS) virus belongs to the Novirhabdovirus genus of
the family Rhabdoviridae. European VHS virus of freshwater origin causes disease
primarily in rainbow trout (RBT; Oncorhynchus mykiss) with high mortality.
Outbreaks of VHS have also been recorded in marine fish species like turbot
(Scophthalmus maximus; 17, 22, 23), and Japanese flounder (Paralichthys olivaceus;
9). VHSV has been implicated in mortality of Pacific herring (Clupea pallasi; 15) and
from later studies it was shown that this species is highly susceptible to experimental
challenge (11). Infection trials have shown that VHSV isolates originating from
marine fish (ma-VHSV) have low pathogenicity in RBT compared with freshwater
VHSV strains (fw-VHSV; 24). However, no studies have been carried to determine
the mechanisms for these differences at the level of host-pathogen interactions.
Fw-VHSV strains infect through waterborne exposure and the prime port of entry is
suggestively the skin and/or gills. This is based on observations where early virus
replication has been demonstrated in gill epithelial cells in situ of rainbow trout (1,
10; 18). Further, studies show that epithelial cells from skin and gills are capable of
supporting replication of VHSV (31) and the viral replication in excised fins
correlates with resistance to waterborne challenge (21). The progression of an IHNV
infection was suggested to occur from the gills into circulation, and/or from the oral
region to the gastrointestinal tract, with subsequent distribution to circulation (4).
Replication of IHNV at early stage in internal organs was proposed to take place in
the kidney (4). This is concordant with findings reported for both VHSV and IHNV in
RBT (1) and for ma-VHSV in turbot (2) using immunohistochemistry (2,6). In these
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

4
previous studies we showed that VHS and IHN viruses were detected in macrophages
and melanomacrophages at early time post infection, indicating that these cells
support virus replication in vivo. Replication of VHS and IHN viruses in macrophages
cultured in vitro has also been demonstrated (3, 5). In a recent study the fin base was
identified as a possible port of entry for IHNV (8).
In studies by Skall et al. (24) 139 ma-VHSV isolates from wild marine fish
and from farmed turbot did not cause mortality in RBT by immersion challenge, but
some isolates caused up to 60 % mortality by injection. In this study we have
investigated if the low virulence of ma-VHSV in RBT can be related to their ability to
translocate over a confluent gill epithelium. Pärt and colleagues (20) have developed a
method for in vitro culturing of gill epithelial cells on filters. The filter cultures of
GEC establish a monolayer firmly attached to glass and plastic supports. These cells
have the appearance of a differentiated epithelium and tight junctions are established
(20). This method has been used in studies of ion transport and acid-base regulations
in rainbow trout (30). In this experiment we used this in vitro model to study the
translocation of VHSV through a confluent gill epithelium and we compared a ma-
VHSV isolate (ma-1p8, genotype 1b) to virulent fw-VHSV reference strain (fw-DK-
3592B, genotype 1a). The marine strain was selected on the basis of being non-lethal
to RBT following immersion challenge, while causing around 40% mortality when
injected intraperitoneally in RBT. The same isolates were also tested for their ability
to infect isolated head kidney macrophages in vitro and the virulence of the isolates
used was again confirmed in experimental challenge of rainbow trout fry.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

5
MATERIALS AND METHODS
Preparation of GEC. RBT gill epithelial cells were isolated according to the method
described by Pärt et al. (19). RBT with a size of approximately 25-100g cultured in
fresh water were killed by a blow to the head followed by decapitation. The gill
arches were dissected and transferred to a Petri dish and rinsed twice with 10 ml of
PBS (Ca2+
and Mg2+
free) containing 200 µg/ml PEST (Penicillin/Streptomycin from
Gibco), 400 µg/ml gentamycin (Gibco) and 250 µg/ml Fungizone® (Amphotericin B,
Gibco 200 µg/ml.)
Coagulated blood was gently removed from the gills, and the filaments were excised
from the arches and washed twice in 10 ml of the PBS solution. The filaments were
thereafter transported to the lab on ice and transferred to 5 ml trypsin (2.5 % solution
from Gibco) and incubated on a shaker (120-200 rpm) for 20 minutes.
The cell suspension was aspirated from the tubes and filtrated through a 100 µm
nylon filter into a stopping solution; PBS containing 10 % Gibco Fetal bovine serum
(FBS). Remaining filaments were trypsinated in 5 ml 2.5 % trypsin solution for an
additional 20 minutes and filtered into the same stopping solution. The cell
suspension was then centrifuged at 200 x g for 10 minutes at 4ºC, and the cell pellet
was re-suspended in culture medium (Leibowitz L-15 medium supplemented with 2
mM L-glutamine, 5% FBS, 100µg/ml PEST and 200 µg/ml gentamycin). The cells
were then transferred to 75 cm2 Falcon culture flasks with GEC culture medium
(Leibowitz L-15 medium with 2 mM glutamine, 5 % FBS, 100 µg/ml PEST and 200
µg/ml gentamycin), and incubated at 20°C. After 24 hours incubation, the cells were
rinsed twice with PBS to remove non-attached cells. Culture medium was changed
every second day until the experiment started.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

6
Virus propagation. The virulent freshwater strain fw-DK-3592B (Genotype Ia) of
VHS virus isolated from RBT in Denmark (24), or the marine strain ma-1p8
(Genotype Ib), isolated from herring (Clupea harengus) in the Baltic Sea (16, 24)
were grown on the BF-2 cells in Eagle’s minimum essential medium with Earle’s
balanced salt solution (EBSS) supplemented with 16.4 mM Tris-buffer, 5.3 mM
NaHCO3, 10% fetal bovine serum (FBS), 4mM l-glutamine and 50 og ml-1
gentamicin. A stock of fw-DK-3592B with a dose of 8.66 log TCID50 ml-1
and a
stock of 1p8 with dose of 7.37 log TCID50 ml-1
were frozen in vials at -80 °C.
Infection assay GEC. Adherent gill epithelial cell were trypsinized 5-7 days after
isolation and transferred 400 000 - 500 000 cells/well to 24 well plates covered with
13mm diameter plastic coverslips. The GEC were cultured for 2-3 days. At time of
infection frozen vials with virus were thawed, and GEC were inoculated with 105
TCID50/ml of the fw-DK-3592B strain, or the ma-1p8 strain.
On each day from day 1 to 6 post inoculation subsets of wells were fixed by adding
1ml of 80% acetone to three wells of both inoculates and left for 2 min. Fixed cell
layers were then washed with Tris buffer saline (TBS) and incubated with IP5B11
(12) against VHSV-N protein for 1 hour at 37ºC. Coverslips were subsequently
washed with TBS and incubated for 30 min with rabbit anti mouse IgG (DAKO),
followed by APAAP (DAKO) mouse monoclonal for 30 min. After washing, Fast red
(1g/l; Sigma, St Louis, MO, USA) and Naphthol AS-MX-phosphate (0,2 g/l; Sigma)
with 1mM levamisole (Sigma) in 0.1 M TBS was added to develop for 20 min.
Counterstained with hematoxylin and mounted with Aquamount (BDH Laboratory).
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

7
The numbers of infected cells were estimated from 200 counted cells in each of three
parallel coverslips at 1, 2, 3 or 6 days post inoculation of virus.
Viability assay GEC. After 3-7 days of culturing the adherent GEC were trypsinized
and transferred to a 96 well plates at a concentration of 40.000 – 50.000 cells/well in
200 µl GEC culture medium. The GEC were sub-cultured for an additional 2-3 days
before removing media and adding 100 µl of a 106
TCID50/ml of fw-DK-3592B or
ma-1p8, respectively. After 0 to 7 days of inoculation, 20 µl of CellTiter AQueous One
solution reagent (Promega) was added to each well containing 100 µl of medium. The
cells were incubated for an additional 24 hours at 15ºC. The quantity of formazan
product in each well was measured at 490nm using a 96 well absorbance plate reader
and is directly proportional with the number of living cells in the culture well.
Transepithelial passage in GEC. GECs were grown in culture flasks for about 5-9
days, after which the cells were trypsinated and resuspended in medium. A sample
was counted by using KOVA Glasstic®
slide 10 with grids. 200 000 cells/insert was
added per filter (FALCON 0.4 µm pore size PET-track-etched membrane Cell culture
insert). The cells were cultured on cell culture chamber inserts with porous bottom
dishes providing virus access to pass through the membrane surface (Fig. 1). The
medium was frequently changed ensuring good cell growth. The transepithelial
resistance was measured daily by use of Millicell®
ERS meter with chopstick
electrodes (Millipore Co., Bedford, MA, USA). Result was expressed as K ohms x
cm2. Infection of cells was initiated when resistance exceeded 1 K ohms x cm
2
(usually after 24-48h of incubation). GEC inserts were inoculated with, 105
TCID50/ml of ma-1p8 or fw-DK-3592B. At 2, 4, 8, 24 and 48 hours, a medium vial of
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

8
100 µl of 3 replicate cell culture chambers was sampled from the basal side of the
filter (Fig. 1). The samples were transferred to a sub-confluent layer of BF-2 cells
cultured on 24 wells plates (20-48 hours old) at 1:100 dilution. The inoculated cells
were incubated at 15flC and inspected for up to 7 days after inoculation for the
occurrence of CPE following primary incubation. After 7 days of culture, cell culture
medium (supernatant) was passed onto fresh (20-48 hours old) BF-2 cells cultured on
24 well plates (2nd
passage). Inoculated cells were inspected for the occurrence of
CPE during the secondary incubation period lasting up to 7 days after inoculation.
Cytopathic effects (CPE) consisted of cell rounding of focal appearance, followed by
detachment of cells from the plastic surface. Identification of virus was performed by
use of a standard ELISA technique using mab IP5B11 as primary antibody (19).
In vitro infection of RBT macrophages. Disease-free RBT (no history of disease
during the life-cycle of the fish) of a size of 30-80g, cultured in fresh water were
anaesthetized with chlorbutanol (1g/L) and killed by decapitation. The head kidney
was dissected out and transferred to incubation buffer (HEPES-buffered (20mM, pH
7.4) saline solution). The head kidney was torn into pieces with two sterile needles
and gently pressed through a nylon mesh (pores 0.3 mm) with a glass rod. The cell
fraction was concentrated by centrifugation and resuspended in incubation buffer.
Homogenous cell fractions were obtained by discontinuous Percoll (Pharmacia)
gradient density centrifugation (G/rpm). Cells concentrated in the bands between the
fraction densities 37% and 54% Percoll were diluted in L-15 medium containing
penicillin 100 U/ml, streptomycin 100 たg/ml and kanamycin 100 たg/ml (all from
Gibco) to give a final cell concentration of 106 cells/ml. One ml of this solution was
seeded onto 12 mm Ø coverslips in culture wells, and incubated at 14ºC in air. The
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

9
cell medium was changed and the wells washed twice with PBS to remove the non-
adherent cell fraction. The following day (after 24 hours of incubation) the wells were
inoculated with identical concentrations, 3.9x107
infective particles, of the fw-DK-
3592B strain or the ma-1p8 strain and the number of infected cells was estimated
from 200 counted cells in two parallel coverslips incubated for 1, 2, 3 or 6 days post
infection. The cell layers were fixed in 80% acetone, washed with Tris buffer saline
(TBS) and incubated with mab IP5B11 for 1 hour at 37ºC. Coverslips were then
washed with TBS and incubated for 30 min with rabbit anti mouse IgG (DAKO),
followed by APAAP (DAKO) mouse monoclonal for 30 min. After washing, Fast red
(1g/l; Sigma, St Louis, MO, USA) and Naphthol AS-MX-phosphate (0,2 g/l; Sigma)
with 1mM levamisole (Sigma) in 0.1 M TBS was added to develop for 20 min.
Counterstained with hematoxylin and mounted with an aqueous mounting medium
(Aquamount, BDH Laboratory).
Viability assay M . The wells were added 200 ol of 8 oM ethidium homodimer (red
fluorescent visualizes dead cells) and 2 oM calcein AM (green fluorescent visualizes
live cells; Molecular Probes) and diluted in phosphate buffered saline (PBS) in each
well. Incubation was for 30-120 minutes before the coverslips were evaluated in an
inverted fluorescence microscope (Leica). At 6 days post inoculation the number of
live and dead cells was estimated from 100 counted cells in eight parallel coverslips.
In vivo virulence study in RBT. The fw-DK-3592B and ma-1p8 strains were tested
for their in vivo virulence in rainbow trout fry. Both isolates were propagated and
titrated on the bluegill fry caudal trunk cell line (BF-2; 28) according to standard
procedures (12). Both isolates used were of low passage numbers (maximum 5
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

10
passages). RBT, SPF fish reared at the laboratory, of an average weight of
approximately 0.2g (average of 15 fish) were used in the trial. Approximately 200 fish
were transferred to each of the small tanks (no. 1-4) containing 8 liter of softened tap
water, with temperature maintained at 9-11ºC throughout the experiment. The
challenge dose was 1.6 x 105 TCID50/ml for ma-1p8 (tank 4) and of 2.9 x 10
3
TCID50/ml for fw-DK-3592B (tank 3) and virus free medium for the negative control
(tank 1). The fish were challenged for 2 hours in aerated water by immersion, after
which water flow was resumed. Mortality was recorded daily up to 42 days post
infection. Fish that died during the trial and the survivors were sampled for virus
isolation including the negative control group. From the negative control group and
the group infected with ma-1p8, fish dying on the same day were all pooled (up to 7
fish/pool). The pooled sample was frozen at ÷25flC or ÷80flC until examination. At
termination 25 fish from each group were examined for virus in 3 pools and examined
as described (16).
Virus distribution and quantification. From tank 1-4 sequential sampling of 6 fish
from each group/tank, were collected at day 0 (before onset), 1, 2, 3, 5, 7, 10, 14, and
21 post infection (p.i.). Fish were culled for immunohistochemistry (3 fish) and virus
quantification (3 fish). Immunohistochemistry was performed on slides of paraffin
embedded specimens (14) and examined as described previously (7). Fish were
thawed in cold water for virus quantification, weighed and EMEM was added to give
a dilution of 1:10 (w/v). The fish were grounded with mortar, pestle and sterile sand.
Hexamicin was added and the samples were kept at 4ºC overnight and titrated on BF-
2 cells in 96-well plates (NUNC) with calculation of titer after 7 days. Samples that
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

11
did not produce cytopathogen effect (CPE) upon 1st passage were sub-cultured by 1
additional passage.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

12
RESULTS
Virus infectivity and cytopathic effects in GEC. The infectivity of strain ma-1p8
was assessed in GEC by use of immunohistochemistry and any cytopathic effects
were assessed on the basis of morphological changes (CPE) as defined previously.
GECs were infected with a dose of 105 TCID50/ml per well and over a period of 7
days cells were negative by immunocytochemistry and no cytopathic effects were
observed. For fw-DK-3592B, 23% of the GECs were found infected by
immunocytochemical staining by day 1 post infection (Fig. 2), and 64% were infected
by day 2. At day 3 and 4, counting was not possible as no cells were found attached to
the coverslips because of virus-induced cytopathic effect.
Viability assay GEC. The virulence of the ma-1p8 isolate was also evaluated by
assessing the viability of ma-1p8-infected GEC using CellTiter 96® AQueous One
Solution Cell Proliferation Assay, used for determining the number of viable cells in
cytotoxicity assays. As this is considered a more sensitive method than morphological
assessment it was included to document cytotoxicity (CPE) induced by ma-1p8
infection. GECs inoculated with isolate ma-1p8 was not different from non-infected
cells while GECs infected with fw-DK-3592B showed a rapid decline in viability and
at three days post infection viability was close to zero, indicating full cytolysis (Fig.
3).
GEC transepithelial passage of VHSV. In an attempt to mimic the in vivo infection
of virus across the gill epithelial barrier (Fig. 1), GECs were grown on filters where a
polarization of the cells is established (19, 29) and the filter inserts were subsequently
inoculated with ma-1p8 on the apical side (Fig. 1). Media vials collected from 3
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

13
replicate chambers at each time point from the basal side of GECs inoculated with
ma-1p8 were virus negative by culture on BF-2 cells and by use of ELISA at 2, 4, 8
and 24 hours p.i.. By 48 hours p.i. 2 out of 3 replicate media vials were virus positive
by 48 hours p.i. by culture on BF-2 (CPE) and confirmed by ELISA (Table 1).
Conversely, GECs infected with fw-DK-3592B were virus positive (1 of 3 replicates)
in the basal medium at 2 hours after virus was added to the apical medium. Two of
three replicates were positive by 4, 8, and 24 and by 48 hours p.i. all three replicates
were virus positive in the basal medium (Table 1). These findings indicate a rapid
translocation of the fw-DK-3592B strain across the polarized epithelial cells.
In vitro infection of RBT macrophages. As previous studies (5, 28) have implicated
that macrophages possibly play role in primary replication of VHS virus after the
virus bypass the primary barriers (skin, mucus), head kidney macrophages isolated by
gradient centrifugation were infected with the ma-VHSV and fw-VHSV isolates.
Interestingly, few macrophages were found infected with the fw-DK-3592B strain,
averaging from 4% by day 1 in two parallel wells and peaking at 8.75% positive cells
by culture day 3 (Table 2). By 6 day of culture, the percentage positive cells had
declined to 3.5%. Positive staining for virus in infected macrophages was seen in
confined structures in the cell cytoplasm (Fig. 4). By this time and later, a few
macrophages (viability assessment described below) were found with a condensed
nucleus or in lysis, possibly as a result of virus replication. In macrophage cultures
infected with the ma-1p8 strain, an average of 0.5% virus-positive cells were found by
day 1 and there was no increase in the number of infected macrophages over the
culture period (Table 2). By day 6, no cells were found positive.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

14
Viability assays of M . Since detection of VHS virus by immunoenzyme methods
has limited sensitivity (7) and there is a possibility of underestimating the number of
virus-infected cells using this method, evaluation of cell viability of macrophages kept
in culture was also included, both in infected and non-infected parallels. The viability
of macrophages was assessed by staining the cultures using a combination of ethidium
homodimer (red fluorescence; dead cells) and calcein AM (green fluorescence; live
cells) to visualize live versus dead macrophages in culture.
The viability assay (counting of 100 cells in 8 parallels) showed that 78% of non-
infected macrophages were viable by day 6 in culture. This was not different from
what was found for cultures infected with strain ma-1p8, while for strain fw-DK-
3592B, only 55% of the cells were viable by day 6 post infection (Fig. 5). By visual
examination in a fluorescence microscope the condensed cells were identified as dead
cells (Fig. 6).
In vivo challenge study in RBT
The in vivo virulence characteristics of isolates fw-DK-3592B and ma-1p8 have been
tested previously (24), but since they had been subject to additional passages in cell
culture, a challenge trial in rainbow trout fry was performed to confirm previous
results. For isolate fw-DK-3592B, the cumulative mortality was 100% by day 13 and
fish died with classical VHS symptoms. Dead fish sampled at day 7 and 12 were all
found VHSV positive by culture and ELISA. For isolate ma-1p8, the cumulative
mortality by day 24 post challenge was 26%, higher than for the non-infected controls
(15% by day 24). No fish died between day 24 and 42 in the ma-1p8 challenged group
or the controls.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

15
VHS was confirmed by histology and immunohistochemistry with
classical histological changes in the fw-DK-3592-B group by day 3 post
infection in target organs such as kidney (not shown). For ma-1p8
infected fish, histopathological changes were not observed at any
collection times and no virus was detected by immunohistochemistry at
any time points post challenge. By culture fish were positive by day 2
post challenge for fw-DK-3592B with a low titer (7.1 x 102 TCID50/g
tissue) reaching a titer peak by day 7 (1.3 x 108 TCID50/g tissue). In the
ma-1p8 group 10 pools of dead fish (pools of 1-7 fish, 31 fish total), were all negative
for VHSV by culture. At day 42, 3 pools of 8, 8 and 9 fish, respectively were
examined and one pool (of 8 fish) was virus positive. In the controls, all pools
examined were negative for VHSV. Together, this indicates a high non-specific
mortality in the ma-1p8 and the control groups.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

16
DISCUSSION
In previous studies it has been well documented that marine isolates of VHSV
are non-pathogenic to rainbow trout following bath challenge while variable mortality
(up to 60%) has been found after injection challenge (24). The underlying factors
have not been elucidated in any detail. In this study we confirmed published results
that a marine isolate of VHSV (ma-1p8) is very inefficient at infecting fry of rainbow
trout following bath challenge. These findings correlate with a low ability of the virus
to infect and translocate over polarized, primary gill epithelial cell (GEC) cultures and
the low in vitro infectivity of isolate ma-1p8 in primary cultures of head kidney
macrophages. These results contrast the findings for the highly virulent freshwater
isolate of VHSV fw-DK-3592B.
The portal of entry for VHSV infection of rainbow trout has not been
conclusively proven. We have previously shown that fw-VHSV can be detected in gill
epithelial cells by immunohistochemistry at early time following an experimental
infection, however limited to a few epithelial cells (1). Similar observations have been
made by others (3, 10, 18) but these studies can only serve as indication as to how the
virus gains access to systemic circulation in trout. An interesting observation is the
finding that a low virulent strain of VHSV (Makah) for rainbow trout showed low
replication in fin tissue and not at all in gill tissue ex vivo (31). Our findings are in
concert with this observation.
Several studies have suggested that kidney and spleen macrophages have an
important role as target cells for the initial replication of VHSV (10) and previous
studies have shown that kidney and spleen macrophages are infected in vivo (1,6,7).
In this study we found that the strain fw-DK-3592B infected head kidney
macrophages in culture (Fig. 4) also leading to reduced cell viability, i.e. the virus
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

17
strain was virulent (able to induce cell damage) to isolated head kidney macrophages.
In contrast a very low percentage of the cultured macrophages were infected with ma-
1p8 and the number of viable cells was not different from the non-infected control
(Fig. 5) indicating lack of virulence towards macrophages. Also, the number of cells
that was infected and supported viral replication was limited, below 10% for fw-DK-
3592B and 0.5% for ma-1p8. There is also a possibility that we have underestimated
the number of virus positive cells since infected cells are more easily washed off from
the slide during incubation for immunohistochemical staining and viability staining.
However, our findings are concordant with a previous study (28) where it was shown
that in primary macrophage cultures from rainbow trout 8% of the cells supported
VHSV replication and for turbot (Scophthalmus maximus) 1.7% of the cells were
found positive (a strain of rainbow trout origin was studied). No significant cytopathic
effect was observed in experiments performed by Tafalla et al. (28) in contrast to our
findings while Estepa et al. (5) showed that VHSV lysed macrophages from RBT in
vitro which is more in line with what we observed. It thus seems that macrophages
can be infected with virulent strains of VHSV (of rainbow trout origin). However, the
initial VHSV replication occur in endothelial cells and to a lesser degree in
macrophages. Further there is also a relatively low infectivity in macrophages in
vitro, and taken together it therefore remains to document that macrophages are the
most important target cells for VHS virus at early time of infection.
Concordant with previous findings ma-1p8 infects fry of rainbow trout at a
very low prevalence following in vivo challenge, while the freshwater strain was
highly virulent (10). The mortality developed rapidly in the fw-DK-3592B challenged
fish from day 4 to 10 and with 100% mortality by day 12 p.i. The background
mortality in the controls was higher than normal possibly associated with fry being
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

18
used for challenge. These sized fish are more susceptible to handling and sorting
which can account for some of the background mortality. Fry were used since they are
considered more susceptible to challenge than larger fish which would provide a
sensitive biological test system. Overall from histological examination,
immunohistochemistry and virus re-isolation there is good documentation provided to
state that isolate ma-1p8 was avirulent with no VHS related mortality occurring over
the course of the experiment. At termination three pools were examined and one of
the pools was found positive for VHSV by culture, showing that isolate ma-1p8 can
infect rainbow trout but without clinical signs or pathology in internal organs,
corroborating previous findings by Skall et al. (24).
Over the last 20 years VHSV have been isolated from several marine fish
species (25) and this has created a concern that marine strains of VHSV can be a
potential source of infection for farmed rainbow trout (25). Studies performed by
Skall et al. (24) demonstrate very clearly that rainbow trout has a very low
susceptibility to several VHSV originating from marine fish when tested by
immersion challenge. 139 ma-VHSV isolates from wild marine fish and from farmed
turbot did not cause mortality in RBT by immersion, but some isolates caused
mortality by injection (24). The low virulence of ma-VHSV in RBT can be related to
their limited ability to pass a confluent gill epithelium or the external barriers to
infection. However, one should not rule out the possibility that ma-VHSV can cause
disease in RBT in a commercial setting and studies (26,27) show that the ma-VHSV
isolates are genetically closely related to fw-VHSV isolates. Recently the first known
disease outbreak in sea-farmed rainbow trout caused by genotype 3 of VHSV was
documented in Norway (13). Bath challenge trials in rainbow trout with this isolate
showed high mortality and is the first registration of a marine VHS virus of genotype
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

19
3 being virulent for rainbow trout (13). It is an indication of a possible adaption of a
marine isolates of VHSV to a new host. Future studies including infectivity and
virulence towards RBT gill epithelial cells of this new variant of a marine VHSV
isolate would be an interesting endeavor and should be pursued.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

20
REFERENCES
1. Brudeseth, B.E., Castric, J., Evensen, Ø. 2002. Studies on pathogenesis
following single and double infection with viral hemorrhagic septicamia virus
and infectious hematopoietic necrosis virus in rainbow trout (Oncorhynchus
mykiss). Vet. Pathol. 39:180-9.
2. Brudeseth, B.E., Raynard, R.S., King, J.A., Evensen, Ø. 2005. Sequential
pathology after experimental infection with marine viral hemorrhagic
septicemia virus isolates of low and high virulence in turbot (Scophthalmus
maximus L). Vet. Pathol. 42:9-18.
3. Chilmonczyk, S., Voccia, I., Monge, D. 1995. Pathogenesis of viral
haemorrhagic septicaemia virus: cellular aspects. Vet. Res. 26:505-11.
4. Drolet, B.S., Rohovec, J.S., Leong, J.A. 2002. The route of entry and
progression of infectious haematopoietic necrosis virus in Oncorhynchus
mykiss (Walbaum): a sequential immunohistochemical study. J Fish Dis
17:337-47.
5. Estepa, A., Frias, D., Coll, J.M. 1992. Susceptibility of trout kidney
macrophages to viral hemorrhagic septicemia virus. Viral Immunol. 5:283-92.
6. Evensen, Ø., Meier, W., Wahli, T., Olesen, N.J., Jørgensen, P.E.V., Håstein,
T. 1994. Comparison of immunohistochemistry and virus cultivation for
identification of viral haemorrhagic septicaemia virus in experimentally
infected rainbow trout (Oncorhynchus mykiss). Dis. Aquat. Organ. 20:101-9.
7. Evensen, Ø., Olesen, N.J. 1997. Immunohistochemical detection of VHS virus
in paraffin-embedded specimens of rainbow trout (Oncorhynchus mykiss): the
influence of primary antibody, fixative and antigen unmasking on method
sensitivity. Vet. Pathol. 34:253-61.
8. Harmache, A., LeBerre, M., Droineau, S., Giovannini, M., Brémont, M. 2006.
Bioluminescence imaging of live infected salmonids reveals that the fin bases
are the major portal of entry for Novirhabdovirus. J. Virol. 80(7):3655-9.
9. Isshiki, T., Nishizawa, T., Kobayashi, T., Nagano, T., Miyazaki, T. 2001. An
outbreak of VHSV (viral haemorrhagic septicaemia virus) infection in farmed
Japanese flounder Paralichthys olivaceus in Japan. Dis. Aquat. Organ. 47:87-
99.
10. Kinkelin de, P., Chilmonczyk, S., Dorson, M., Le Berre, M., Baudaouy, A.M.
1979. Some pathogenic facets of rhabdoviral infection of salmonid fish. In:
Bachmann , P.A. (Ed.) Munich symposia on microbiology: Mechanisms of
viral pathogenesis and virulence. WHO Publ., Munich p. 357-75.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

21
11. Kocan, R., Bradley, M., Elder, N., Meyers, T.R., Batts, W.N. Winton, J.R.
1997. North American strain of viral hemorrhagic septicemia virus is highly
pathogenic for laboratory-reared Pacific herring. J. Aquatic Animal Health 9:
279-90.
12. Lorenzen, N., Olesen, N.J., Jørgensen, P.E.V. 1988. Production and
characterization of monoclonal antibodies to four Egtved virus structural
proteins. Dis. Aquat. Organ. 4:35-42.
13. Lyngstad, T.M., Høgåsen, H.R., Ørpetveit, I., Hellberg, H., Dale, O.B.,
Lillehaug, A. Scientific evaluation of the eradication of viral hemorrhagic
septicemia in Storfjorden (in Norwegian). National Veterinary Institute’s
Report Series no. 3, 2008, ISSN 1890-3290 electronic version.
14. Meier, W., Pfister, K. 1981. Viral hemorrhagic septicemia (VHS virus) in pike
(Esox lucius L.). Clinical, macroscopic, histological and electron-
microscopical findings; direct visualisation of Egtved virus. Schweiz. Arch.
Tierheilkd. 123:37-49.
15. Meyers, T. R. Winton, J. R. 1995. Viral hemorrhagic septicemia virus in North
America. Ann. Rev. Fish Dis. 5:3-24.
16. Mortensen, H.F., Heuer, O.E., Lorenzen, N., Otte, L., Olesen, N.J. 1999.
Isolation of viral haemorrhagic septicaemia virus (VHSV) from wild marine
fish species in the Baltic Sea, Kattegat, Skagerrak and the North Sea. Vir. Res.
63:95-106.
17. Munro, A.S. 1996. Report on the first recorded outbreak of viral haemorrhagic
septicaemia (VHS) in Great Britain and subsequent actions to contain,
eradicate and investigate the origins of the infection. Scottish Aquaculture
Research Report Number 3. The Scottish Office Agriculture, Environment and
Fisheries Department, Aberdeen
18. Neukirch, M., Glass, B. 1984. Some aspects of virus shedding by rainbow
trout (Salmo gairdneri Rich.) after waterborne infection with viral
haemorrhagic septicaemia (VHS) virus. Zent. Bakt. Mikro. Hyg. 257:433-38.
19. Olesen, N.J., Jørgensen, P.E.V. 1991. Rapid detection of viral haemorrhagic
septicaemia virus in fish by ELISA. J. Appl. Ichtyol. 7:183-86.
20. Pärt, P., Norrgran, L., Bergström, E., Sjöberg, P. 1993. Primary cultrures of
epithelial cells from rainbow trout gills. J. Exp. Biol. 175:219-32.
21. Quillet, E., Dorson, M., Aubard, G., Torhy, C. 2001. In vitro viral
haemorrhagic septicaemia virus replication in excised fins of rainbow trout:
correlation with resistance to waterborne challenge and genetic variation. Dis.
Aquat. Organ. 45(3):171-82.
22. Ross, K., McCarthy, U., Huntly, P., Wood, B., Stuart, D., Rough, E., Smail, D.
& Bruno, D. 1994. An outbreak of viral heamorrhagic septicaemia (VHS) in
turbot (Scophthalmus maximus L.) in Scotland. Bull. European Ass.Fish Path.
14:213-14.
23. Schlotfeldt, H., Ahne, W., Vestergård Jørgensen, P., Glende, W. 1991.
Occurrence of viral haemorrhagic septicaemia in turbot (Schopthalmus
maximus L.) -a natural outbreak. Bull. European Ass.Fish Path. 11:105-7.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

22
24. Skall, H.F., Slierendrecht, W.J., King, J.A., Olesen, N.J. 2004. Experimental
infection of rainbow trout Oncorhynchus mykiss with viral haemorrhagic
septicaemia virus isolates from European marine and farmed fishes. Dis.
Aquat. Organ. 58:99-110.
25. Skall, H.F., Olesen, N.J. Mellergaard, S. 2005. Viral haemorrhagic
septicaemia virus in marine fish and its implications for fish farming – A
review. J. Fish Dis. 28:509-29.
26. Snow, M., Cunningham, C.O., Melvin, W.T., Kurath, G. 1999. Analysis of the
nucleoprotein gene, identifies distinct lineages of viral haemorrhagic
septicaemia virus (VHSV) within the European marine environment. Vir. Res.
63:35-44.
27. Snow, M., Bain, N., Black, J., Taupin, V., Cunningham, C.O., King, J.A.,
Skall, H.F., Raynard, R.S. 2004. Genetic population structure of marine viral
haemorrhagic septicaemia virus (VHSV). Dis. Aquatic Organ. 61:11-21.
28. Tafalla, C., Figueras, B., Novoa, B. 1998. In vitro interaction of viral
haemorrhagic septicaemia virus and leukocytes from trout (Oncorhynchus
mykiss) and turbot (scophthalmus maximus). Vet. Immunol.Immunopathol.
62:359-66.
29. Wolf, K., Gravell, M., Malsberger, R. G. 1966. Lymphocystis virus: Isolation
and propagation in centrarchid fish cell lines. Science 151:1004-5.
30. Wood, C.M., Pärt, P. 2000. Intracellular pH regulation and buffer capacity in
CO2/HCO3-buffered media in cultured epithelial cells from rainbow trout
gills. J. Comp. Physiol. [B] 170(3):175-84.
31. Yamamoto, T., Batts, W.N., Winton, J.R. 1992. In vitro infection of salmonid
epidermal tissues by infectious hematopoietic necrosis virus and viral
hemorrhagic septicemia virus. J. Aquat. Animal Health 4:231-39. ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Fig. 1 depicts the layout of the filters on which the GEC was grown. The setup
mirrors the water-to-blood barrier of the gills of rainbow trout.
Fig. 2. Gill epithelial cells (GECs) infected with fw-DK-3592B at 1 day p.i. Single
cells are detected as virus positive in the cytoplasm by alkaline phosphatase (red
color) using mab IP5B11 and APAAP/Fast Red. The slide is counterstained with
Mayer’s hematoxylin (blue nuclei). Bar = 10 たm.
“GILLS” Basolateral
medium
”WATER”
Apical
medium
Samples transferred to BF2.
”BLOOD” Filter
If CPE:
VHSV verified by ELISA
Well
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Fig. 3. GEC cultures were infected with strains fw-DK-3592B or ma-1p8,
respectively and incubated for 7 days. 1 parallel was left non-infected. Mean
absorbance value of GEC incubated with CellTiter AQueous One solution reagent (at
490 nm) is given and each value is based on mean value of 11 wells. Background
absorbance is set to zero.
Fig. 4. Macrophages, infected with fw-DK-3592B at 3 days p.i. cells with cytoplasmic
staining (red coloration). Virus antigen is detected by alkaline phosphatase coupled
antibodies visualized by Fast Red. Counterstained with Mayer’s hematoxylin. Bar =
10 たm.
GEC viability
0
0.05
0.1
0.15
0.2
0.25
1 3 5 7
Days post infection
Absorb
ance 6
00 n
m
fw-DK3592B
ma-1p8
Non-infected
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
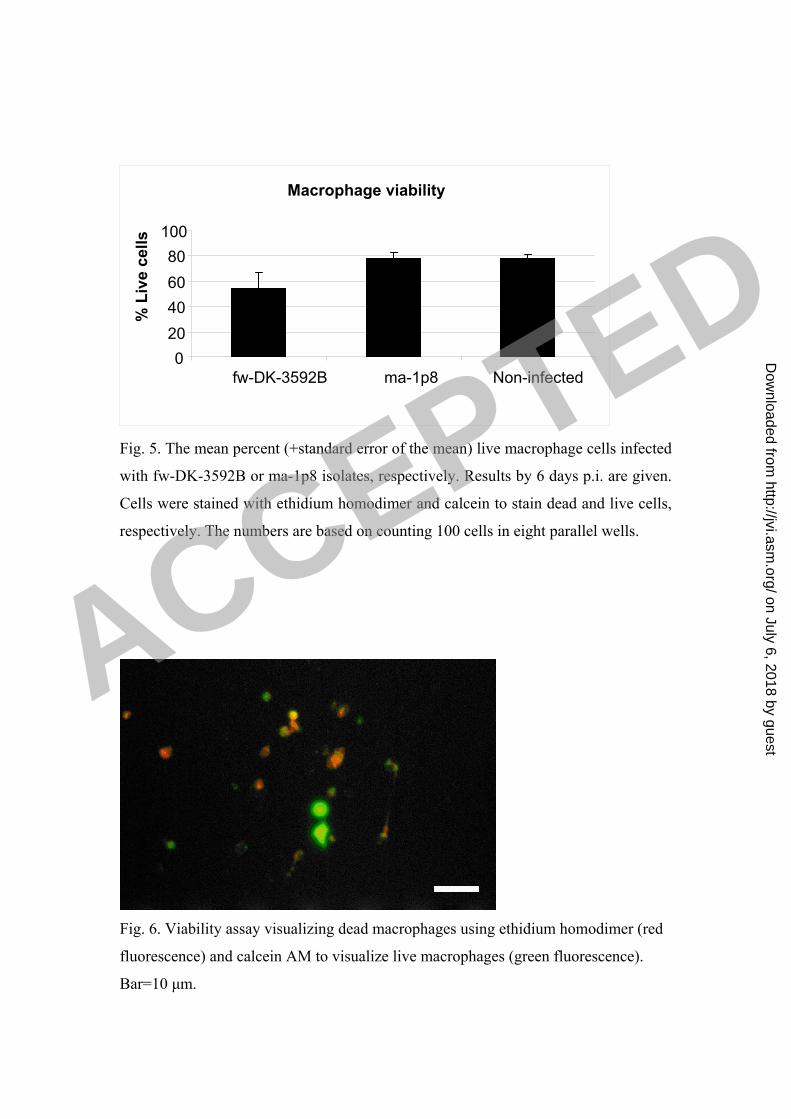
Fig. 5. The mean percent (+standard error of the mean) live macrophage cells infected
with fw-DK-3592B or ma-1p8 isolates, respectively. Results by 6 days p.i. are given.
Cells were stained with ethidium homodimer and calcein to stain dead and live cells,
respectively. The numbers are based on counting 100 cells in eight parallel wells.
Fig. 6. Viability assay visualizing dead macrophages using ethidium homodimer (red
fluorescence) and calcein AM to visualize live macrophages (green fluorescence).
Bar=10 たm.
Macrophage viability
0
20
40
60
80
100
fw-DK-3592B ma-1p8 Non-infected
% L
ive c
ells
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

0 %
10 %
20 %
30 %
40 %
50 %
60 %
70 %
80 %
90 %
100 %
1 4 7 10 13 16 19 22 25 28 31 34 37 40
Days after challenge
% c
um
ula
tive m
ort
ality
fw-DK-3592B
ma-1p8
Control
Fig. 7. The percentage cumulative mortality following VHSV challenge with fw-DK-
3592B and ma-1p8 strains is shown. Each group was challenged in a separate tank. A
separate control tank with non-challenged fish was also included. Fish sampled for
studies of viral distribution are not included in the figure.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

Hours p.i. fw-DK-3592B ma-1p8
2 1 0
4 2 0
8 2 0
24 2 0
48 3 2
Table 1. GEC filter inserts infected with fw-DK-3592B and ma-1p8 on the apical side and
samples collected at the basal side (see Fig. 1 for details) were examined for presence of virus
in BF-2 cells. The number of positive wells (of 3 per time point) is indicated over an
incubation period of 48h after virus was added to the apical side.
% +ve cells infected with fw-DK-3592B % +ve infected with ma-1p8 Days p.i.
C1 C2 C1 C2
1 3,0 5,0 0 1,0
2 4,0 8,0 0 0,25
3 8,0 9,5 0,5 0,5
6 3,5 3,5 0 0
Table 2. Detection of VHSV in cultured macrophages isolated from head kidney of RBT and
subsequently infected with isolate fw-DK-3592B or isolate ma-1p8. The number of infected
cells was estimated from 200 counted cells in each of two parallel coverslips (C1 and C2)
after staining of virus positive cells using mab IP5B11 and developed with APAAP.
Percentage positive cells are indicated.
ACCEPTED
on July 6, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from