diets of free-ranging mexican gray wolves janet e. …
TRANSCRIPT

DIETS OF FREE-RANGING MEXICAN GRAY WOLVES
IN ARIZONA AND NEW MEXICO
by
JANET E. REED. B.S.
A THESIS
IN
WILDLIFE SCIENCE
Submitted to the Graduate Faculty
of Texas Tech University in Partial Fulfillment of the Requirements for
the Degree of
MASTER OF SCIENCE
Approved
Chairperson of the Committee
Accepjpd
Dean of the Graduate School
May, 2004

ACKNOWLEDGEMENTS
I thank the United States Fish and Wildlife Service (USFWS) for fiinding this
project I appreciated the direction of Mexican wolf recovery leader and committee
member, Brian T Kelly, the assistance of Mexican wolf Interagency Field Team
personnel (i e , USFWS, Arizona Game and Fish Department, New Mexico Department
of Game and Fish, USD A APHIS Wildlife Services, and US Forest Service [USFS])
and all the volunteers who coUected scats Thank you, David R Parsons and Wendy
Brown, for mentoring and believing in me
I thank Dr Warren B Ballard, my major advisor, for providing encouragement
and guidance throughout this study Special thanks go to committee member Dr Robert
J Baker for welcoming me into his genetics lab and to Celine C Perchellet, Adam
Brown, and Drs Jeff Wickliffe and Federico Hoffman for their dedication, assistance,
and humor in the DNA lab I also thank committee members Drs Philip S Gipson, Paul
R. Krausman, and Mark C Wallace for their ad\ice and support A special thank you to
Dr David B Wester for facilitating the statistical analyses
I thank the USFS Alpine and Clifton Ranger Districts for their assistance on the
forest A special thank you to Myron C Burnett, USFS Wilderness Ranger, for rising
above the opposition to teach me to pack burros and believing that I could handle the job
I also thank the residents of Alpine for welcoming me into their community and all the
forest visitors who assisted with scat collecting Thank you Waypoint Enterprises, Show
Low, Arizona, for your donation of UTM and Multi Waypointers

My parents, Roger C and Betty A Stone, my sister, Anita D. Moore, and my
fnend, Alan B Alley, provided unwavering support and encouragement throughout my
education. Thanks to my two burros, Pancho and Lefty, who carried my gear and the
stinky carnivore scats tirelessly over many miles. Finally, my dog. Kaiser von
Stockwerk, provided faithfiil companionship for MVJ years His unconditional love was
greatiy appreciated
111

TABLE OF CONTENTS
ACKNOWLEDGEMENTS ii
ABSTRACT vi
UST OF TABLES viii
USTOFnOURES x
CHAPTER
I INTRODUCTION 1
Literature Cited 2
n fflSTORICAL OVERVIEW OF THE MEXICAN GRAY WOLF
AND ITS RECOVERY 3
Literature Cited 7
in DIFFERENTIATING MEXICAN WOLF AND COYOTE SCATS
USING DNA ANALYSIS 10
Abstract 10
Introduction 11
Study Area 12
Materials and Methods 14
Results 22
Discussion 24
Acknowledgements 31
Literature Cited 32
IV

IV. DIETS OF FREE-RANGING MEXICAN GRAY WOLVES IN
ARIZONA AND NEW MEXICO 45
Abstract 45
Introduction 47
Study Area 48
Methods 51
Resuhs 55
Discussion 57
Acknowledgements 64
Literamre Cited 65
V SUMMARY 82

ABSTRACT
The Mexican gray wolf (Catiis lupus baileyi) was extirpated from the
southwestern United States before any systematic smdies could be conducted, therefore
littie is known about the subspecies' natural history From April 1998 through October
2001, we coUected carnivore scats {n = 1,682) from the Blue Range Wolf Recovery Area
(BRWRA) in .\rizona and New Mexico to study the diets of free-ranging Mexican
wolves We identified the scats to species using traditional field methods (i e , diameter,
location, and sign) and odor, but expected that it would be difficult to separate Mexican
wolf and sympatric coyote {Cams latrans) scats We tested this hypothesis whh fecal
DNA analysis (molecular scatology) to verify the accuracy of identifying Mexican wolf
and sympatric coyote scats Our DNA results showed scats > 28 mm diameter could be
identified as deposited by Mexican wolves, a high overlap in scat diameters for Mexican
wolves and sympatric coyotes, and a difference in the 2 species' scat diameter means
To determine the diets of Mexican wolves, we used our DNA results to refine our
scat identification criteria Scats for diet analysis were identified as deposited by
Mexican wolves by DNA analysis, diameters ^ 28 mm, 2 den site locations, and ttacks.
Diet analysis of 55 scats with diameters ^ 28 mm collected from areas where 6 Mexican
wolf packs received supplemental food items indicated Cienega and Hawks Nest packs
consumed non-supplemental food items during supplemental feeding and Campbell Blue,
Lupine, Mule, and Pipestem packs did not Diet analysis of 251 scats revealed non-
supplememal fed Mexican wolves consumed large-sized food items, primarily elk
VI

{Cerws elaphus caiKhJensis [nelsoni]) adults and calves We compared this diet
composition to that found in 26 scats identified with DNA analysis and found more large-
sized food items appeared in Mexican wolf scats identified by our refined traditional
method than in scats identified with DNA analysis There was no difference between
diameter means or number of food items per scat between the 2 scat identification
methods There was a difference between diet composition for 26 Mexican wolf scats
and that found in 21 sympatric coyote scats identified with DNA analysis, with Mexican
wolves consuming more large-sized food items than sympatric coyotes. There was also a
difference between diameter means and number of food items per scat for the 2 species,
vsith sympatric coyote scats containing more food items per scat than Mexican wolf scats.
Lastly, we found a difference in diet composition of Vtexican wolves when compared to
diet composition reported in 7 previous diet studies of other North American gray
wolves Mexican wolf diet analysis revealed more large-sized food items than the
subspecies' larger, northern counterparts
vn

UST OF TABLES
3.1 Comparison of accuracy of predicting which species (Mexican wolf or coyote) deposited a scat using discriminant analysis based on combination of 3 measurements takoi from scats identified to species with DNA analysis 43
3.2 Locations of scats (n = 47) identified as Mexican wolf or coyote with DNA analysis 44
4.1 Food items found in scats (n = SS) ^ 28 mm diameter collected from areas where Mexican gray wolf packs (n = 6) were fed supplemental food items (i.e., carnivore logs and road-killed elk, deer and jackrabbit) 71
4.2 ConqMurison of diet composition found in scats (n = 55) ^ 28 mm diameter from areas where Mexican gray wolf packs (n = 6) were fed supplemental food items (i.e., carnivore logs and road-killed elk, deer, and jackrabbit) to determine presence of non-supplemental food items (e.g., elk aduhs and calves, deo* adults and £Eiwns, domestic bovine, and insects) 72
4.3 Food items (n = 265) found in free-ranging Mexican gray wolf scats (/i = 251) 73
4.4 Comparison among years (n = 4,1998-2001) of diet composition found in scats {n = 251) of free-ranging Mexican gray wolves collected from April 1998 to October 2001 in Arizona and New Mexico 74
4.5 Comparison between seasons (n = 2, fiill-winter versus s|Hing-summer) of diet composition found in scats (/i = 251) of free-ranging Mexican gray wolves in Arizona and New Mexico (April 1998 - October 2001) 75
4.6 Comparison of diet conq)08ition among packs (n = 4) found in free-ranging Mexican wolf scats (n = 251) collected from AJptii 1998 to October 2001 in Arizona and New Mexico 76
4.7 Food items (n = 33) found in free-ranging Mexican gray wolf scats (n = 26) collected from April 1998 to October 2001 in Arizona and New Mexico 77
4.8 Comparison of diet c<Hiq)Osition of Mexican wolf scats (n = 277) collected from April 1998 to Oct(A>er 2001 in Arizona and New Mexico and identified u«ng 2 methods: DNA analysis (n = 26) and refined traditional (/t = 251; i.e., ^ 28 mm diameter, 2 den sites, and tracks) 78
4.9 Coiiq>ari8(ni of diet composition found in free-ranging Mexican wolf scats (/f = 26) and sympatric coyote scats (/i = 21) collected from hipdX 1998 to October 2001 in Arizona and New Mexico 79
viu

4.10 Food items found in free-ranging Mexican gray wolf scats (n = 26) and sympatric coyote scats (n = 21) coUected from April 1998 to October 2001 in Arizona and New Mexico 80
4. U ConqpariscHi of diet composition found in free-ranging Mexican wolf scats (n = 277) and diet oompositi(» reported in other North American gray wolf diet studies (n = 7) 81
IX

LIST OF FIGURES
2.1 The Bhie Range Wolf Recovery Area (BRWRA) in Arizona and New Mexico where Mexican gray wolves were released beginning April 1998 9
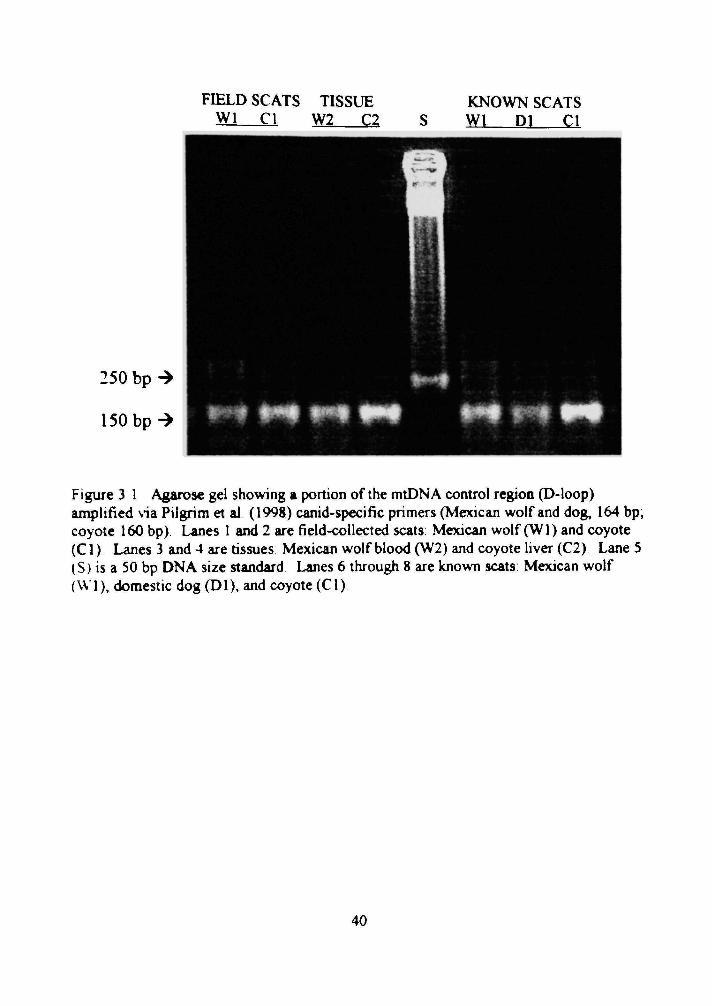
3 1 Agarose gd showing a portion of the mtDN A control region (D-loop) amplified via Pilgrim et al. (1998) canid-specific primers (Mexican wolf and dog, 164 bp; coyote 160 bp) 40
3 2 Agarose gel showing restriction fragment digestion with BstN I of mtDNA control r^on (D-loop) purified polym«-ase chain reaction (PPCR) isolated from Mexican wolf and coyote scats 41
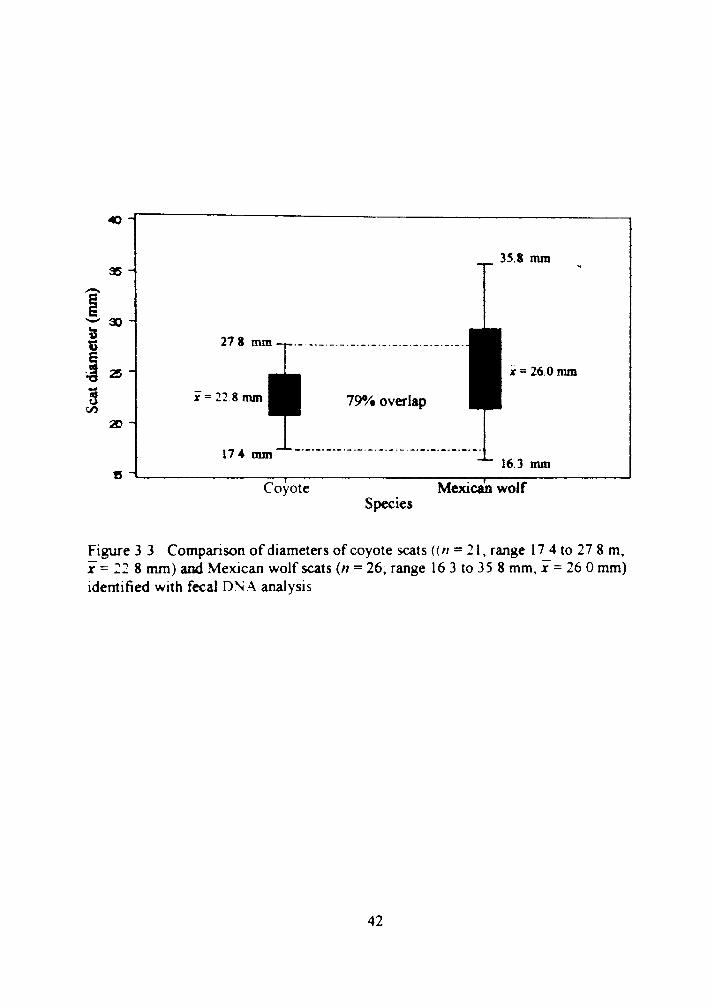
3.3 Comparison of diameters of coyote scats (n = 21, range 17.4 to 27.8 m; X = 22.8 nun) and Mexican wolf scats (n = 26; range 16.3 to 35.8 nun, x = 26.0 nun) identified with fecal DNA analysis 42

CHAPTER I
INTRODUCTION
The following chapters constitute partial fulfillment of the requirements for the
degree of Master of Science in Wildlife Science for the Graduate School at Texas Tech
University These chapters are the resuh of research conducted on Mexican gray wolves
(Cojus lupus baileyi) m Arizona and New Mexico from April 1998 through October
2001 Chapter II is an historical overview of the Mexican gray wolf and its recovery
Chapters ID and I\' are 2 manuscripts intended for submission to peer-reviewed journals.
Chapter III discusses differentiating Mexican wolf and coyote scats using fecal DNA
analysis (molecular scatology) Chapter IV reports the diets of free-ranging Mexican
wolves in .\rizona and New Mexico from April 1998 through October 2001, and
compares the diets of Mexican wolves to sympatric coyotes and to other North American
gray wolves Chapter V is a summary of all chapters ,\11 chapters represent my ideas,
analyses, and writing ability Each chapter has several coauthors, which were determined
using guidelines provided by Dickson and Conner (1978) and the CBE Style Manual
Committee (1994) .Authorships for chapters are as follows
Chapter III Janet E Reed, Robert J Baker, Warren B Ballard, and Brian T Kelly
Chapter IV; Janet E Reed, Warren B Ballard, Philip S Gipson, Brian T Kelly, Paul
R Krausman, Mark C Wallace, and David B Wester

LiteraUire Cited
CBE Style Manual Committee 1994 Scientific style and format: the CBE manual for authors, editors, and publishers Sixth Edition Council of Biology Editors, Cambridge University Press, New York, New York, USA.
Dickson, J. G., and R N Conner 1978 Guidelines for authorship of scientific articles. Wildlife Society Bulletin 6: 260-261

CHAPTER II
HISTORICAL OVERVIEW
OF THE MEXICAN GRAY WOLF AND ITS RECOVERY
Historically, the Mexican gray wolf (Ca7>/5 lupus baileyi), or lobo, lived the
farthest south of all gray wolves (C lupus) on North America and in the most arid
environment at elevations from 1,200 to 3,300 m ranging throughout central and
southeastern Arizona, western Texas, southern New Mexico, and most of Old Mexico
(Young and Goldman 1944, Brown 1983, Parsons 1996) h is believed that the Mexican
wolf comributed to the overall historical biological diversity and ecological functioning
of both the southwestern ecosystems and the continued evolution of species the Mexican
wolf preyed upon (Parsons 1998)
It is uncertain if the Mexican wolf was sympatric with the coyote {Canis latrans)
in the Southwest before humans altered its natural habitat (Brown 1983) Gier (1975)
and Bekoff and Wells (1986) reported that historical coyote distnbutions were confined
primarily to plains and deserts until the spread of civilization and the reduction of gray
wolf ranges
By the early 1880s, humans and associated livestock had moved into the
Southwest in large numbers (Brown 1983, Shumway 1998) Due to overgrazing and
unregulated subsistence and market hunting, native prey species were in decline and
some predators turned to the more abundant and easily caught livestock (United States
Fish and Wildlife Service 1987) Government and private predator comrol programs

designed to protect livestock in the United States and Mexico accelerated the extirpation
of the Mexican wolf from the Southwest by the late 1960s (Gipson and Ballard 1998).
In 1975, the Arizona-Sonora Desert Museum proposed to capture Mexican
wolves from the remaining population in Mexico and to place them in a captive-breeding
program for future re-estabhshment of the subspecies in its historic range (Siminski
1997) The United States government listed the Mexican wolf as endangered in 1976 and
Mexico began protecting this subspecies, as well (United States Fish and Wildlife Service
1996) The last 4 Mexican wolves known to exist were captured in Durango and
Chihuahua, Mexico from 1977 to 1980 (McBride 1978) and 3 of them founded an official
captive-breeding program (Garcia-Moreno et al 1996, Hedrick et al 1997) To increase
genetic diversity in the official Certified (renamed McBride) lineage, 2 other captive
Mexican wolf populations, Arag6n and Ghost Ranch lineages, were certified genetically
pure in July 1995 (Hedrick et al 1997) and incorporated into the official captive-breeding
program The first offspring from cross-lineage pairings were produced in 1997 (Parsons
1998) .\s of October 2001, the global population of Mexican wolves consisted of 227
individuals, the majority of which were held in 40 zoos and wildlife sanctuaries
throughout the United Stales and Mexico (P Suninski, Arizona-Sonora Desert Museum,
personal communication)
Recovery efforts for the Mexican wolf were outlined in the Mexican Wolf
Recovery Plan (United States Fish and Wildlife Service 1982) and research flinded by the
Lmted States Fish and Wildlife Service (USFWS) supported experimental releases of
Mexican wolves imo isolated Southwest habitat where close monitoring could be

conducted (Bednarz 1988) The Final Envu-onmental Impact Statement (FEIS) for the
Mexican wolf proposed reintroduction of the subspecies into a portion of its historical
range in Arizona, New Mexico, and Texas (United States Fish and Wildlife Service
1996) Three areas were considered the Blue Range Wolf Recovery Area (BRWRA) in
.\rizona and New Mexico, White Sands Missile Range Wolf Recovery Area in New
Mexico, and Big Bend National Park in Texas This proposal was formally approved by
Secretar, of the Interior Bmce Babbitt's March 1997 Record of Decision (United States
Fish and Wildlife Service 1997). In April 1998, the first 11 captive-reared Mexican
wol\ es-5 generations removed from the wild-were released within the primary recovery
zone of the BRWRA on the Apache National Forest in east-central Arizona (Figure 2 1).
In the spring 2000, Mexican wolves previously released in Arizona were translocated to
the secondary recovery zone on the Gila National Forest and Wilderness in New Mexico
As of October 2001, at least 37 Mexican wolves were resident within the BRWTIA and
31 of those were radiocollared (United States Fish and Wildlife Service, unpublished
data)
In June 2000, we began a study to determine the diets of free-ranging Mexican
gray wolves in Arizona and New Mexico We collected 1,682 carnivore scats from April
1998 through October 2001 and identified them to species using traditional identification
methods and odor We verified scat identification accuracy with fecal DNA analysis
(molecular scatology) and refined our identification of Mexican wolf scats We then
analyzed Mexican wolf scats identified with DNA analysis and our refined traditional

identification method to determine the diet composition of free-ranging Mexican wolves
in Arizona and New Mexico

Literature Cited
Bednarz,! C 1988 The Mexican wolf biology, history, and prospects for reestabhshment in New Mexico Endangered species report No. 18 United States Fish and Wildlife Service, Albuquerque, New Mexico, USA.
Bekoff, M . and M C Wells 1986 Social ecology and behavior of coyotes Advances in tiie Stiidy of Behavior 16: 251-338
Brown, D E 1983 The wolf in the Southwest: the making of an endangered species. The University of .\rizona Press, Tucson, Arizona, USA
Garcia-Moreno, J , M D Matocq, M S Roy, E Geffen, and R K Wayne 1996 Relationships and genetic purity of the endangered Mexican wolf based on analysis of microsatellite loci Conservation Biology 10 376-389
Gier. H T 1975 Ecology and behavior of the coyotes (Co^iw/a/rans), pages 247-262/>/ M W Fox, editor The Wild Canids their systematics, behavioral ecology and evolution Van Nostrand Reinhold, New York, USA
Gipson, P S , and W B Ballard 1998 Accountsof famous North American wolves Canadian Field-Naturalist 112 724-739
Hednck, P W , P S Miller, E Geffen, and R K Wayne 1997 Genetic evaluation of the three captive Mexican wolf lineages Zoo Biology 16 47-69
McBride, R T 1978 Statusof the gray wolf (ran/5/M^.vZ)a//ev/) in Mexico United States Fish and Wildlife Service Report, Albuquerque, New Mexico, USA.
Parsons, D R 1996 Case Smdy the Mexican wolf Pages 101-123 inE A. Herrera and L F Huenneke, editors New Mexico's natural heritage biological diversity in the Land of Enchantment New Mexico Journal of Science 36 101-123
Parsons, D R. 1998 "Green fire" returns to the Southwest reintroduaion of the Mexican wolf Wildlife Society Bulletin 26: 799-807
Shumway, E W 1998 Alpine, Arizona a stroll through history, Americopy, Mesa, Arizona, USA
Simmski, D P 1997 A history of cooperation in Mexican wolf conservation Proceedings American Zoo and Aquarium Association Regional Conference 1997 384-385

United States Fish and Wildlife Service. 1982. Mexicanwolf recovery plan. United States Fish and Wildlife Service, Albuquerque, New Mexico, USA
United States Fish and Wildlife Service 1987. Restoring America's wildlife 1937-1987: the first 50 years of the Federal Aid in Wildlife Restoration (Pitman-Robertson .\ct) Unhed States Department of the Interior, United States Fish and Wildlife Service, Washington, D C , USA.
United States Fish and Wildlife Service 1996 Reintroduction of the Mexican wolf within its historic range in the southwestern United States Final Environmental Impact Statement. United States Fish and Wildlife Service, Albuquerque, New Mexico, US.\
United States Fish and Wildlife Service 1997. Record of decision and statement of fmdings on the environmental impact statement on reintroduction of the Mexican gray wolf to rts historic range in the southwestern United States United States Fish and Wildlife Service, Albuquerque, New Mexico, USA
Young, S P . andE A Goldman 1944 The wolves of North America The American Wildlife Institute, Washington, D C , USA


CHAPTER III
DIFFERENT1.\TING MEXICAN GRAY WOLF AND
COYOTE SCATS USING DNA ANALYSIS
Abstract
We used fecal DNA analysis (molecular scatology) to test a subsample of scats
(// = 203) identified as deposited by Mexican gray wolves (Ca j/5 lupus baileyi) or
coyotes {Cams latrans) and compared the resuhs to the identification of scats using
traditional methods (i e , diameter, location, and sign) and odor We then used the scats
identified with DN.A. analysis to evaluate discriminant analysis for classifying scats using
3 measurements (i e , diameter, mass, and length) Forty-nine (24° o) of the field-
collected scats {n = 203) tested provided amplifiable DN.A., which identified 28 scats
deposhed by Mexican wolves and 21 deposited by coyotes Scats identified with DNA
analysis for the 2 species had a 79% diameter overiap (Mexican wolf, 16 3 to 35 8 mm,
coyote, 17 4 to 27 8 mm) and scats ^ 28 mm diameter were Mexican wolf scats There
was a difference (/ = -2 428. Z' = 0 019) between diameter means for the 2 species
(Mexican wolf ir = 26 0 mm, coyotex = 22 8 mm) Of 45 scats that would have been
field-identified as deposited by Mexican wolves based on location and odor criteria,
DN.A analysis showed that 19 (42°o) were deposited by coyote, and of 41 scats that
would have been field-identified as deposhed by coyotes based on diameter < 30 mm
criterion, 20 (49%) were deposited by Mexican wolves We then used Halfpenny's
(1986) suggested diameter criterion for field-identification of scats, which identified 3 of
10

the scats as gray {Urocyon cmereoargenleus) or red (Vulpes vulpes) fox (0% correct), 24
as coyote (62o o correct), and 20 as Mexican wolf (75% correct). Discriminant analysis
indicated that diameter and mass offered the best resuhs for accurately classifying coyote
scats (86° o), but provided relatively low accuracy for classifying Mexican wolf scats
(65° o) Molecular scatology appears to be a more definitive scat identification technique
than methods previously utilized (i e , traditional and odor)
Introduction
No scientific smdies were conducted on the Mexican gray wolf (Canw lupus
baileyi) before it was extirpated from the wild, therefore little is known of the subspecies'
natural history (Brown 1983) Previous studies indicate that the Mexican wolf is the
smallest (Hall and Kelson 1959, Bogan and Mehlhop 1983). the most genetically distinct
(Wayne et al 1992, Nowak 1995, Garcia-Moreno et al 1996), and the most endangered
(McBride 1980. Brown 1983, Bednarz 1988, Ginsberg and Macdonald 1990) subspecies
of gray wolf (C lupus) on North America In April 1998, the United States Fish and
Wildlife Service (USFWS) began releasing captive Mexican wolves into the Blue Range
Wolf Recovery .\rea (BRWRA) on the Apache and Gila National Forests and the Gila
Wilderness in Arizona and New Mexico to restore the subspecies into a portion of its
historical range
In June 2000, we began a study to determine the diets of captive-bom and wild-
bom .Mexican gray wolves free-ranging in the southwestern United States From April
1998 through October 2001, we collected carnivore scats {n = 1,682) from the BRWRA
I I

and identified them to species using traditional field methods (i e , diameter, location, and
sign) and odor Mexican wolves are the smallest subspecies of gray wolves found on
North America, therefore we hypothesized that traditional scat identification techniques
might have a high error rate in separating Mexican wolf and coyote (Canis latrans) scats
collected where the 2 species were sympatric
There are no published reports using fecal DNA analysis (molecular scatology) to
differentiate between gray wolf and coyote scats for diet analysis The purpose of this
research was to accurately differentiate Mexican wolf and coyote scats in order to study
the diets of Mexican wolves Accurate identification of scats would be beneficial for
determining the level of dietary overlap of gray wolves and coyotes in areas where they
are s>Tnpatric, managing gray wolf and prey populations, and investigating livestock
depredation incidences in areas where gray wolves have been reintroduced or are
recovering
Study .Area
We conducted research in the BRW'R.A (Figure 2 1), a 17,820 km^ area that
included the Apache National Forest (NF) in east-central Arizona and the Gila NF and
Wilderness in west-central New Mexico Our focus was the 2,600 km^ primary recovery
zone in .Arizona where captive Mexican wolves were released and wild-bom since April
1998 The secondary recovery zone (Gila National Forest and Wilderness, New Mexico)
was included beginning spring 2000 due to translocations of Mexican wolves previously
released in Arizona The BRWRA was bordered by the White Mountain (Fort Apache)
12

and San Carlos Apache Reservations to the west, while private lands were scattered
within and bordered the east, north and south Almost all areas were grazed by domestic
livestock
Elevations within the BRWRA ranged from < 1,220 m along the San Francisco
River to 3,350 m on Mount Baldy, Escudilla Mountain, and the Mogollon Mountains
(United States Fish and Wildlife Service 1996) Rolling hills with moderately steep-
walled canyons and sandy washes characterized the lower elevations, while mgged
slopes, deep canyons, elevated mesas, and rock cliffs typified the higher elevations.
Major vegetation included ponderosa pine (Pittusponderosa), aspen (Populus
iremuloides), fir (Abies spp ), juniper (Juniperus spp ), piflon (Pirrus cembroides),
mesquhe (Prosopis spp ), evergreen oaks {Quercus spp ), and a variety of grasses and
forbs Annual temperatures reported for the BRWHA averaged 16 4-C maximum and
-3 \°C minimum, annual precipitation averaged 52 1 cm, and annual snowfall averaged
139 3 cm (Desert Research Institute, Western Region Climate Center, Reno, Nevada,
impublished data)
From April 1998 through October 2(X)1, > 87 Mexican wolves were either
released from captivity or bom in the wild within the BRWTIA As of October 2001,
> 37 Mexican wolves resided in the BRWRA and 31 of those were fitted with radio
collars (United States Fish and Wildlife Service, unpublished data) Other predators
residem in the BRWRA included humans (Homo sapiens), coyote, gray (Urocyon
cmereoargenleus) and red (Vulpes vulpes) fox, bobcat (Felis rufus), mountain lion (Puma
concolor), and black bear (Ursus americanus) (United States Fish and Wildlife Service
13

1993, Arizona Game and Fish Department 1994) Density estimates for these predators
were unavailable, but the United States Forest Service (USFS) and Arizona Game and
Fish Department (AGFD) reported coyote densities to be relatively high (personal
communication) These sources and the USFWS also reported that feral and stray dogs
or wolf-dog hybrids were rare residents, if at all, within the BRWRA, therefore we
assumed a low likelihood of collecting dog or wolf-dog hybrid scats.
Materials and Methods
Sample collection
Carnivore scats (/; = 1,682) were collected opportunistically from the BRWRA by
Mexican wolf Interagency Field Team (IFT, i e , U S Fish and Wildlife Service, Arizona
Cjame and Fish Department, New Mexico Department of Game and Fish, USDA APHIS
Wildlife Services, and U S Forest Service) personnel from April 1998 through October
2001 We actively collected scats from June through August 2000 and March through
October 2(X)1 in areas where captive-released, translocated, and wild-bom Mexican
wolves were known to frequent The scat collection process employed an opportunistic
sampling strategy (Frenzel 1974) in which the sampling areas followed Mexican wolves
as they moved within the study area as reported by the USFWS Our search methods
included driving forest roads, hiking or horseback riding forest trails, ridgelines, and
npanan areas, and investigating forest campsites, opened release pens, den sites and kill
or carcass sites
14

Scats were collected using disposable mbber or food preparation gloves and
placed in brown paper bags, which were labeled with date, location, and scat number.
We aged scats as old, recent, or fresh according to appearance, exposure of deposition
site, and weather conditions (Ciucci et al 1996) Scats were identified initially to species
using traditional identification techniques (i e , location, diameter, and sign) and odor as
described below We did not autoclave the scats, as there were no reports of
Echinoci\:cus granulosus in the Southwest The scats were allowed to ah dry in the
brown paper bags before storing in large plastic containers at room temperature until
analyzed
The location criterion was established as areas frequented by Mexican wolves,
such as opened release pens, den sites, and kill or carcass sites We measured the
maximum diameter of each air-dried scat as described by Scott (1943). Weaver and Fritts
(1979), Green and Flinders (1981), and Danner and Dodd (1982). however, we took 2
measurements at the maximum diameter to the nearest 0 1 mm with a 152 mm dial
caliper (General Tools Manufacturing Co , LLC, New York, New York) and used the
average for diameter size Thompson (1952) established the minimum diameter for
identifying northern gray wolf scats in the field as > 24 mm and has been the accepted
critenon for several smdies (Mech 1970, Stephenson and Johnson 1972, Peterson 1974,
Van Ballenberghe et al 1975) However, Weaver and Fritts (1979) suggested > 30 mm
minimum diameter be used for identifying gray wolf scats, which was the diameter
critenon used by Arjo et al (2002) Halfpenny (1986) reviewed 3 stiidies and suggested
> 25 mm diameter for identifying gray wolf scats Initially, we used the more
15

conservative > 30 mm diameter criterion for classifying scats as deposited by Mexican
wolves Sign criterion was tracks of Mexican wolves Mexican wolf IFT personnel
provided instmction for identifying Mexican wolf scats by odor We were provided with
scats identified as Mexican wolf scats by the IFT personnel and were instmcted that the
odor (i e , sweet, musky) of those scats was that of wolf No odor description was
available for identify ing coyote scats.
Genetic identification
We used fecal DN.A analysis to identify scats deposited by either Mexican wolves
or coyotes Our design was to isolate and analyze Mexican wolf and coyote DNA from
shed epithelial cells from the intestinal lining found on scats (Kohn et al 1995, Foran et
al 1997, Kohn and Wayne 1997, Reed et al 1997, Frantzen et al 1998) We examined a
portion of the mhochondrial DNA (mtDNA) control region (D-loop, Pilgrim et al. 1998)
to ditTerentiate Mexican wolf scats from s\mpatric coyote scats collected from the field
We subsampled 203 of the field-collected scats by taking 2 to 6 fecal surface
samples from each scat for DNA analysis The surface of the scats were scraped to
decrease the possibility of removing undigested prey parts from within the scats and to
increase the probability of obtaining sloughed epithelial cells (Reed et al. 1997) Each of
the subsamples, comprising ^ 1 g of fecal material taken from each scat, was placed in a
1 5 mL micro centrifuge tube and stored at room temperature (Taberlet et al 1997) The
remainder of each scat was used for diet analysis To optimize laboratory efforts, scats
were not subsampled randomly but were selected primarily from scats field-identified as
16

deposited by Mexican wolves (/»= 169) We sorted the master database for scats
identified as probably deposited by Mexican wolf scats (/»= 1.111) based on traditional
field metiiods and odor, tiien selected every 5* scat to be subsampled for DNA analysis.
DN.A isolation
Fecal material is likely to contain relatively low concentrations of DNA much of
which is degraded (Gerloff et al 1995, Kohn et al 1995), yet mtDNA is likely to be
present on a scat m greater quantity than single-copy nuclear DNA (Reed et al 1997,
Woods etal 1999) Polymerase chain reaction (PCR) amplification of short mtDNA
fragments results in more consistem amplifications than longer fragments (Kohn et al
1995) Therefore, it was necessary to identify and establish a simple, rapid and reliable
protocol that extracted as much DNA as possible from degraded samples and removed
any potential PCR inhibitors (Boom et al 1990, Deuter et al 1995. Kohn et al 1995.
Reed et al 1997) We tried 4 DNA isolation methods Proteinase K method (Sambrook
etal 1989). salt extraction method (Miller et al 1988), Qiagen DNeasy* Tissue Kit
(Qiagen, Valencia, California, USA), and guanidinium thiocyanate (GuSCN) and silica
method (Boom et al 1990, Hoss et al 1992, Hbss & Piabo 1993) Our resuhs were
similar to those of Hoss and Paabo (1993) and Reed et al (1997), with the GuSCN and
silica method having the highest isolation efficiency and providing the most consistent
resuhs We used this method as our DNA isolation protocol and optunized it for our
application, as described below
17

We isolated genomic DNA from: whole blood (n = 38) previously collected from
released Mexican wolves, museum specimen coyote tissue (liver, n = 4), and Mexican
wolf scats (n = 13) collected from captive individuals, coyote scats (n = 5) collected
where no wx)l\ es occurred along the New Mexico and Texas border, and domestic dog
scats (/; = 5) The DNA isolated from tiiese sources provided the reference collection for
genetically identifying target species' (Foran et al 1997) Addhionally, we attempted to
isolate DN.A from museum specimen rodent tissue (liver, /; = 8), and hair (n = 4) and
grass {n ^ 2) from field-collected scats to test for possible amplication of food items
ingested
We modified an extraction buffer (5 M GuSCN, 0 1 M Tns-HCl pH 6 4, 0 2 M
EDT.A pH 8 0, 13% Triton X-100), bringing the volume to 50 mL with ddHjO (Boom et
al 1990, Hoss and Paabo 1993, Hoss 1994) One millimeter of extraction buffer was
added to each <. 1 g of dry fecal material in a 1 5 mL micro centrifiige tube The products
were vortexed, then incubated overnight at 55°C with constant agitation Samples were
then centnfiiged for 10 min at 13.000 rpm in a Microfuge* 18 Centrifuge (Beckman
Couher^, Inc , Palo Alto, California, USA) Approximately 7(X) [iL of the supernatant
was transferred to a sterile 1 5 mL micro centrifuge tube and 300 (iL of fresh extraction
buffer and 50 nL of silica matrix were added (Boom et al. 1990) The mixture was
vortexed before incubated at 55°C for 10 min with constant agitation After
centrifligation (1 min, 5,000 rpm), the supernatant was decanted and discarded. The
remaining silica pellet was washed twice with 1 mL of a modified wash buffer (5 M
GuSCN, 0 1 M Tns-HCl pH 6 4, Boom et al 1990) and once with 1 mL of 70% ethanol,
18

centrifiiging each time for 1 min at 5,000 rpm The pellet was dried at 55°C and then the
extracted DNA was eluted at 55°C with constant agitation for 30 min in 100 ^L elution
buff'er (10 mM Tris-CI, pH 8 5) The product was centrifliged (5 min, 13,000 rpm) and
75 [iL of the supernatant was removed carefully to avoid transferring silica particles that
could inhibit the PCR (Hoss and PMbo 1993) and placed in a sterile 15 mL micro
centrifuge tube These samples were stored at -20°C until PCR amplification
DN A isolation procedures were performed under a fume hood in a laboratory
exclusiN ely dedicated to DNA isolation and PCR amplification Care was taken
throughout all procedures to ensure no cross-contamination To monhor contamination
during DN.A isolations, negative controls (no fecal material added to the extraction
buffer) were treated identically through both the isolation procedure and the subsequent
PCR amplifications (Handt et al 1994)
PCR amplification
We used the PCR to amplify < 200 base pairs (bp) of the mtDNA control region
(D-loop) using canid-specific mtDNA primers (F-4479679, R-4479680, Integrated DNA
Technologies, Inc , Coralville, Iowa, USA) designed by Pilgrim et al (1998) to produce
species-specific bands of 164 bp for wolves and dogs and 160 bp for coyotes
Four microliters of excremental DNA extract were placed in a sterile 0 5 mL
micro centrifiige tube with a 46 nL volume containing 0 2 mM each dNTP, 3 25 mM
.MgCIj, IX buff'er, 0 5 piM each pnmer, 0 5 U Tag polymerase (Ml865, Promega
Corporation, .Madison, Wisconsin, USA), and 25 0 nL ddH20, then topped with 2 drops
19

of autoclaved mineral oil to prevent evaporation. The DNA and reagent mixtiires were
amplified in a programmable Perkin Elmer CeUis 480 DNA thermal cycler (Perkin
Elmer, Norwalk, Connecticut, USA) for 32 cycles, each cycle consisting of a
denaturation step for 20 sec at 95°C, an annealing step for 40 sec at 50°C, and an
extension step of 30 sec at 72°C Three microliters of tiie amplification reaction were
mixed with 3 |iL standard tracking dye and were separated at 60 milliamps on a 2%
agarose gel containing etiiidium bromide in TAE buff'er (Ausubel et al 1989) Gels were
visualized under ultraviolet (LTV) excitation on an Eagle Eye n (Stratagene*, La Jolla,
California, US.A) Amplicoms were sized using a 50 bp DNA ladder (Fisher Scientific
BP 2550-100)
Negative extraction and PCR controls were used alongside the excremental DNA
samples to monitor contamination of the PCR reagents As positive controls, we used the
DN.A extracted from blood samples collected from previously released Mexican wolves.
DNA analysis
Pilgrim et al (1998) identified a control region length difference between coyotes
and wolves and dogs, which also results in a restriction-site polymorphism present in
wolves and dogs but lacking in coyotes Using the restriction enzyme Mva I (BstN I,
New England BioLabs Inc , Beverly, Massachusetts, USA), Pilgrim et al (1998)
distinguished coyote amplicons from those of wolf and dog Twenty-five PCR products
were digested with 0 35 ^L restriction enzyme BstN I, 3 fiL lOX buff'er, 3 ^L BSA and
15 95 ^L ddH20 at 37°C for 3 hr Digested amplicons were analyzed on a 2 5% agarose
20

gel containing ethidium bromide in TAE buff'er, then visualized under UV excitation on
an Eagle Eye n Ehgested products were sized using a 50 bp ladder Products that
remained undigested at 160 bp were identified as coyote and fragments that digested into
2 fi^gments of 49 and 113 bp were identified as wolf
To confirm the results of the mtDNA restriction fragment analysis, we sequenced
tiie same fragment used for the digestions of 31 scats PCR products were purified using
the QLAquick PCR Purification Kit (Qiagen) following the supplier's instmctions
Punfied PCR (PPCR) products were cycle-sequenced in both directions (2 reactions)
using a programmable PTC-200 Pehier Thermal Cycler (MJ Research, Inc , Watertown,
Massachusetts, USA). 1 5 ^L PPCR, 1 0 nL 2 mM forward or reverse primer. 4 ^L Big
D\e Terminator v3 0 (4390244, Applied Biosystems, Foster City, California, USA), and
3 5 (iL ddHjO Each cycle-sequenced product was purified with 3 0 ^L 3 M NaOAc pH
4 6, b2 5 ^L 100% EtOH, and 14 5 nL ddHjO for 15 min at room temperaUire, then
centrifliged for 20 min at 13,(XX) rpm The supernatant was decanted and discarded, then
the pellet was washed with 250 |iL 70% EtOH followed by centrifligation for 5 min at
13,(XX) rpm The EtOH was decanted and discarded, then the pellet was air-dried at room
temperature overnight in the dark Twenty microliters of Template Suppression Reagent
(Applied Biosystems) were added to the dried pellet, then vortexed The pellet was
shaken down by hand and incubated at 95°C for 3 min, then quick spun Raw sequence
data were collected using a 310 Genetic Analyzer (Applied Biosystems) Resuhing
sequences were aligned and analyzed using Sequencher v3 1 (Gene Codes Corporation,
Inc , Arm Arbor, Michigan, USA) Aligned, proofed sequences were compared to

homologous wolf and coyote sequences ((jenbank No AF487754, AF020700) and
domestic dog mtDNA sequences (Genbank No AF487751, Slade et al. 1994) The
difference between scat diameter means for Mexican wolf and coyote was calculated
using a standard /-test (Ott 1998) and deemed significant if P <. 0 05
Discriminant analysis
Based on the measurement values (i e , diameter, mass, and length) of the scats,
we used discriminant analysis (Williams 1982) to classify scats as either Mexican wolf or
coyote Forty-seven of the scats identified with DNA analysis as deposited by Mexican
wolf or coyote were weighed to the nearest 0 1 g on a OHAUS Precision Plus TP4000
scale (OHAUS Corporation, Florham Park, New Jersey, USA), and the total length of
each scat was measured with a metric straight edge to the nearest 0 1 cm We used
discriminant analysis to classify scats based on (1) diameter, (2) diameter and mass,
(3) diameter and length, and (4) diameter, mass, and length Since coyote density was
unknown within the smdy area, we accepted prior probability (q,) = 0 50
Resuhs
Genetic identification
Forty-nine (24°o) of 203 scats provided amplifiable DNA (Figure 3 1) Twenty-
eight were identified as Mexican wolf scats and 21 were identified as coyote scats with
restriction fragment analysis (n = IS, Figure 3 2), sequence analysis (n = 24), or both
(/J = 7) Two of the scats identified as deposited by Mexican wolves were uncollectibles,
which are loose, liquid feces containing little idemifiable prey material (Floyd et al
22

1978) Uncollectible scats cannot be measured for diameter or length and traditionally
have not been included in diet stiidies Therefore, the 2 uncollectible scats were
discarded, leaving 26 Mexican wolf scats for further analysis
Mexican wolf scats (n = 26) ranged in diameter from 16 3 to 35.8 mm, while
coyote scats (n = 21) ranged in diameter from 17 4 to 27 8 mm (Figure 3.3). We found a
79«/o o%erlap in scat diameter for Mexican wolf (AJ = 16) and coyote (n = 2l) There was
a difference (t = -2 428, P = 0.019) between scat diameter means for the 2 species
(Mexican wolf x = :6 0 mm, coyote r = 22 8 mm. Figure 3 3) Scat diameters > 28 mm
appeared adequate for identifying Mexican wolf scats without other identification criteria
(i e , location and sign)
Of the 45 scats that would have been identified as deposited by Mexican wolves
based on location and odor criteria, 19 (42° o) were acmally deposited by coyotes, and of
the 41 scats that would have been identified as deposited by coyotes based on diameter
< 30 mm criterion, 20 (49%) were actually deposited by Mexican wolves Using
Halfijenny's (1986) suggested diameter criterion for identifying the scats of 3 carnivore
species, scat diameters < 18 mm would have been identified as fox (n = 3, 0% correct),
diameters 18 mm to 25 mm would have been identified as coyote (n = 24, 62% correct),
and diameters > 25 mm would have been identified as Mexican wolf (/? = 20, 75%
correct)
Foran et al (1997) reported that ingested prey tissue did not produce erroneous
resuhs when targeting species-specific DNA and our resuhs supported this claim Chir
attempts to isolate DNA from rodent tissue (liver) and ingested grass and prey hair taken
23

from scats provided no amplifiable PCR products using the canid-specific primers
designed by Pilgrim et al. (1998)
Discriminant analysis
For scats identified as Mexican wolf or coyote wrth DNA analysis, we used
discriminant analysis for 4 classifications (Table 3 1) Scat diameter (Classification 1)
accuratdy identified 81% of coyote scats, but only 50% of Mexican wolf scats Scat
diameter and mass (Classification 2) accurately identified 86% of coyote scats and 65%
of Mexican wolf scats Scat diameter and length (Classification 3) accurately identified
68% of coyote scats and 59% of Mexican wolf scats Lastly, a combination of scat
diameter, mass, and length (Classification 4) accurately identified 79% of coyote scats
and 55% of Mexican wolf scats
Discussion
Fecal material is often the most common sign and easily collected source of
information for rare, secretive carnivores (Putman 1984), and scat analysis appears to
provide the best noninvasive sampling method for determining diets of free-ranging
carnivore species However, which species actually deposited a scat must be accurately
determined for the method to be vahd Traditional scat identification criteria have been
based primarily on morphology (Halfpenny 1986, Foran et al 1997), which can be
subjective and confounded by sympatric species that are comparably sized and share
similar diets (Weaver and Fritts 1979, Green and Flinders 1981, Danner and Dodd 1982,
24

Foran et al 1997). Halfpenny (1986) reported that visual identification of scat to species
by experienced naturaUsts had error rates approaching 50 to 66%. Scott (1941, 1943)
reported tiiat fecal passage sizes varied approximately in proportion to the amount of
food consumed Weaver and Fritts (1979) reported tiiat scat diameter might be
influenced by diet composition and suggested that canid scats could not be identified to
species based on diameter alone Previous dietary analysis based on scat identification
using diameter alone may have biased results in favor of prey items that produce larger
scat diameters (Danner and Dodd 1982)
Halfjpenny (1986) reported that scat diameter and length values do not provide
positive identification of a species, because both values are too variable to be used as
adequate diagnostic criteria Green and Flinders (1981) reported that the dry mass of
scats \aried considerably for coyotes and red foxes and suggested that diameter used in
conjunction with mass could be used for identifying scats of the 2 species Discriminant
analysis of diameter and mass values of scats (Classification 2) provided the most
accurate identification for both species (coyote, 86%, Mexican wolf, 65%) However,
14°'o of the scats deposited by coyotes were classified incorrectly as Mexican wolf scats
and 35° 0 of the Mexican wolf scats were classified incorrectly as coyote scats Although
diameter and mass values provided a relatively high percentage of accuracy for
identifying coyote scats, we considered the resuhs unsatisfactory for identifying Mexican
wolf scats
Uncollectible scats, as defined by Floyd et al (1978), have traditionally been
discarded because it is difficult to identify them to species and uncollectibles usually
25

contain little identifiable prey material We were able to isolate DNA and identify the
species of 2 uncollectibles subsampled When using the appropriate primers, it is also
possible to amplify any prey DNA found in uncollectibles so that they can now be
included in cami\ore diet studies, if desired
Location of where scats have been deposited has also been used to identify wolf
scats for diet analysis smdies Scats collected from wolf den and rendezvous sites
undoubtedK have provided accurate diet information because coyotes rarely attend these
areas (Ballard et al 2003) However, the information only provides wolf diet data for
late spring through summer (Murie 1944, Mech 1966, Ballard et al 1987, Fuller 1989,
Spaulding et al 1997) Identification of scats collected from kill and carcass sites
(Thompson 1952, .Arjo et al 2(X)2) may be more problematic in areas where wolves and
coyotes are svmpatric because coyotes often scavenge from wolf kills (Paquet 1992,
Phillips and Smith 1996) The scats (n = 47, Table 3 2) from which we were able to
isolate DN.A were collected from forest trails (21 scats), forest highways, roads, and
2-tracks (16 scats), an elk carcass (1 scat), a wolf den (1 scat), an opened release pen
(2 scats), and other locations (i e , stream bed, cabin area, crater, canyon, mesa, and ridge,
6 scats)
Odor has also been reported as a scat identification technique, but it is subjective
(Stokes and Stokes 1986, Bang 2001) and currently cannot be quantified The only
mention of odor in the literature as an identification technique was for fox (Scott 1943,
Murie 1954, Wilcomb 1956, Korschgen 1980, Turkowski 1980), not for wolves or
coyotes Halfpenny (1986) suggested that odor resuhed from the camivore's diet
26

However, some wolf biologists have reported through personal communications that they
can identify wolf scats by odor, but tiiis claim has not been scientifically substantiated
and our resuhs suggest that odor was not a reliable method for this stiidy
Another attempt to identify carnivore scats has employed bile acids (Eneroth and
Sjovall 1969, .Alfred 1980, Major et al 1980, Clinite 1981, Johnson et al 1984, Quinn
and Jackman 1994) or pH (Green and Fhnders 1981) with httle or no success According
to Halfpenny (1986), these methods have proven more successful for herbivores than
carnivores
Recent noninvasive sampling studies of free-ranging mammals (Foran et al 1997,
Kohn and W ayne 1997, Reed et al 1997, Ernest et al 2000, Lucchini et al 2002) have
confirmed that fecal DNA analysis provides a more accurate assignment of the species
that deposited a scat rather than morphology of scats With noninvasive sampling and
fecal DN.A analysis, biologists can collect individual fecal samples at any time, from any
location, to study free-ranging species without having to disturb them (Hoss et al 1992,
Taberlet and Bouvet 1992. Morin et al 1993, Kohn and Wayne 1997, Taberlet et al
1999) We were able to collect scats from the field and isolate DNA for species
identification without ever seeing or dismpting the individuals
DNA from sloughed intestinal epithelial cells found on fecal material provides an
objective marker for species identification, and potentially for individuals, that remains
invariable during the life of an animal (Foran et al 1997) Mitochondrial DNA is
inherited maternally and is non-recombining (Brown 1985), making it well suited for
species identification of fecal samples (Moritz 1994, Kohn and Wayne 1997) and
27

mammalian population genetic stiidies (Randi et al. 1994, Slade et al. 1994, Taberlet et
al 1995) Mitochondrial DNA's extia-nuclear genome provides several hundred copies
per cell (Kohn et al 1995) and is more amenable to genetic manipulation than single-
copy nuclear genes (Kohn and Wayne 1997, Frantten et al 1998). The control region
(D-loop) is usually more polymorphic and informative for differentiating species (Pilgrim
et al 1998) because it evolves faster in mammals than either the rest of the mtDNA
molecule or most single-copy nuclear DNA (Avise 1994) Beginning with small
quantities of relatively short target sequences of DNA (Walsh et al. 1991. Mullis et al
1994. Reed et al 1997), the PCR is an enzymatic process that can experimentally
synthesize a large number of copies of specific DNA sequences from degraded or impure
samples (Saiki et al 1985, Mullis and Faloona 1987, Saiki et al 1988, Amheim et al
1990, Walsh et al 1991) When applied to mtDNA sequences useful for estimating
genetic differences between closely related species can be amplified (Kocher et al 1989,
.A\ise 1994) Species identification of scats using DN.A-based assays provides an
accurate method that is rapid (3-4 days), repeatable, and relatively inexpensive (The cost
is approximately $5 00 [USJ/sample for the disposable items, enzymes, and chemicals
required for sequencing Other costs include the sequencer, non-disposable lab
equipmem [e g , pipetors, cameras, and computers] and salaries of students and
technicians ) Furthermore, the resuhs are definitive and not subject to confidence
intervals or probabilistic estimation (Foran et al 1997) However, our low DNA isolation
success (24%) may be cause for concem
28

DNA isolated from fecal material is often of low quantity and quality (Taberlet et
al 1996) In addition, epitiielial cells usually are distributed unevenly (Kohn et al 1995)
and our fecal subsamples may not have included tiie cells required for DNA isolation.
Fecal material may also contain Taq DNA polymerase or PCR inhibitors, however, tiiis
problem can be reduced by using tiie silica-based extraction method (Boom et al 1990,
H6SS and Paabo 1993, Kohn et al 1995) Our modest DNA isolation success was
attnbuted to low-quality and low-quamity DNA found on scats Furthermore, the scats
we tested were dry-stored for up to 5 years at the time we conducted DNA analysis.
Reed et al (1997) and Lucchini et al (2002) reported that fecal sample age affected DNA
isolation success and suggested that fresh scats would be more suitable for DNA analysis
-Although our DNA isolation success (24%) was low, we found, as did Foran et al
(1997), that a scat's physical appearance was not a definitive guide to the DNA quality
a\ailable Another possibility influencing our low DNA isolation success could have
been that some of the scats were deposited by non-target species whose DNA could not
be amplified with the canid-specific primers designed for wolf and coyote that we used
We found, however, that the targeted species could be identified if a PCR product could
be obtained, since DNA too degraded to amplify produced no results as opposed to
incorrect results (Foran et al 1997) Our results were consistent with the findings of
Pilgrim et al (1998) that wolf and coyote mtDNAs were distinct and could be
differentiated by a single restriction site and length polymorphism We recognize that
there is a possibility that a low percentage of the scats identified as wolf could have been
deposited by feral dogs or wolf-dog hybrids if they existed within the BRWRA.
29

Therefore, we recommend that ftiture wolf DNA research utilize primers (Foran et al.
1997) that differentiate between wolf and dog DNA,
With a sample size of 47 scats identified to species using DNA analysis, we
suggest that our results be interpreted with caution Our resuhs demonstrated, however,
that identification of Mexican wolf and coyote scats using DNA analysis was more
accurate than identification methods previously available Molecular scatology can
facilrtate the identification of species, individuals, their gender, food habhs, and
pathology This would require an experimental design with extended systematic transects
from which fresh fecal samples are obtained, coupled with an appropriate preservation of
fecal material and DN.A isolation method (Reed et al 1997, Wasser et al 1997, Frantzen
et al 1998, Taberlet et al 1999, Lucchini et al 2(X)2), and using appropriate species-
sp>ecific primers (Foran et al 1997). These data could then be used further to estimate
home range (Adams et al 2(K)3), reproductive patterns, kinship stmcture and population
size (Kohn and W ayne 1997) Molecular scatology also has potential to detect
hybndization (Lehman et al 1991, Wayne et al 1992, Gondii et al 1994, Pilgrim et al
1998. Vila and Wayne 1999, Adams et al 2003) Finally, these data could be used for
validating the presence of wolves in livestock depredation incidences
Our resuhs suggest that previous diet studies using traditional scat identification
methods may have misrepresented the diets of both the North American gray wolf and
coyote where the 2 species were sympatric Fecal DNA analysis provides an accurate
method for assessing the visual identification of scat samples collected from the field and
improves diet analysis (Reed et al 1997) Molecular scatology appears to have
30

significant potential as a noninvasive sampling technique to momtor and manage free-
ranging Mexican wolves where the subspecies is sympatric with other carnivore species
Acknowledgements
The United States Fish and Wildlife Service funded this research and provided
scats from captive Mexican wolves The University of New Mexico and Arizona
Veterinary Diagnostic Lab provided blood samples from free-ranging Mexican wolves
The Museum of Texas Tech University provided coyote and rodent tissue samples.
Kaiser von Stockwerk generously provided domestic dog scats A Brown, C C.
Perchellet, and Drs F Hoffinan and J Wickliffe contributed invaluable assistance and
dedication in the DN.A lab Dr D B Wester facilitated the statistical analysis This is a
Texas Tech University. College of Agriculmral Sciences and Natural Resources technical
fwjblication T-X-.\XX
31

Literatiire Cited
Adams, J R , B T KeUy, and L P Waits 2003 Using faecal DNA sampling and GIS to monitor hybridization between red wolves (Canis rufus) and coyotes (Canis latrans) Molecular Ecology 12: 2175-2186
Alfred, D R 1980 Biochemical identification of carnivore scats Thesis, Mississippi State University, Mississippi, USA.
.Amheim, N , T WTiite, and W E Rainey 1990 Application of PCR: organismal and population biology BioScience 40: 174-182
Arizona Game and Fish Department 1994 Predator population estimates Unpublished report, Arizona Game and Fish Department, Phoenix, Arizona, USA
Arjo, W M , D R Pletscher, and R R Ream 2002 Dietary overlap between wolves and coyotes in northwestern Montana Journal of Mammalogy 83: 754-766.
Ausubel, F M . R Brent, R E. Kingston, D D Moore, J G Seigdman, J A Smith, and K Struhl 1989 Current protocols in molecular biology John Wiley & Sons, New York, New York, USA
.A\ise, J C 1994. Molecular markers, namral history, and evolution Chapman & Hall, New York, New York, USA
Ballard, W B , J S Whitman, and C L Gardner 1987 Ecology of an exploited wolf population in south-central Alaska Wildlife Monograph 98
Ballard, W B , L N Carbyn, and D W Smitii 2003 Pages 259-271 m L D Mech and L Boitani, editors Wolves behavior, ecology and conservation University of Chicago Press, Chicago, Illinois, USA
Bang, P 2001 Animal tracks and signs Oxford University Press, New York, USA
Bednarz, J C 1988 The Mexican Wolf: biology, history, and prospects for reestabhshment in New Mexico Endangered Species Report No 18 United States Fish and Wildlife Service, Albuquerque, New Mexico, USA
Bogan, M A, and P Mehlhop 1983 Systematic relationships of gray wolves (Coww lupus) in southwestern North America Occasional papers of the Museum of Southwestern Biology 1
32

Boom, R , C J A. Sol, M M M Salimans. C L Jansen, P M E Wertheim-van Dillen, and J van der Noordaa. 1990 Rapid and simple method for purification of nucleic acids Journal of Clinical Microbiology 28 495-503.
Brown, D E 1983 The wolf in the Southwest the making of an endangered species. The Univwsity of Arizona Press, Tucson, Arizona, USA.
BrowTvW M 1985 The mitochondrial genome of animals Pages 95-130//iR J Maclnryre, editor Molecular Evolutionary Genetics Plenum, New York, USA.
Ciucci, P , L Boitani, E Raganella Pelliccioni, M Rocco, and I. Guy 1996 A comparison of scat-analysis methods to assess the diet of the wo\f Canis lupus Wildlife Biology 2 37-48
Chnrte, E W 1981 Biochemical analysis of mountain lion and bobcat scats: difference between species and sex Thesis, San Jose State University, San Jose, California, USA
Danner, D .A , and N Dodd. 1982 Comparison of coyote and gray fox scat diameters Journal of Wildlife Management 46 240-241
Deuter, R , S Peitsch, S Hertel, and O Muller 1995 A method for preparation of faecal DN.A suitable for PCR Nuclei Acids Research 23 3800-3801
Eneroth, P , and J Sjovall 1969 Methodsof analysis in the biochemistry of bile acids Methods of Enzymology XV Academic Press, New ^'ork. New York, USA
Ernest, R B , M C T Penedo, B P May, M Syvanen, and W M Boyce 2000 Molecular tracking of mountain lions in the Yosemite Valley region in California: genetic analysis using microsatellites and faecal DNA Molecular Ecology 9: 433-441
Floyd, T J , L D Mech, and P A. Jordan 1978 Relating wolf scat content to prey consumed Journal of Wildlife Management 42: 528-532
Foran, D R, K R Crooks, and S C Mima 1997 Species identification from scat: an unambiguous genetic method Wildlife Society Bulletin 25 835-839.
Frantzen, M A J , J B Silk, J W H Ferguson, R K Wayne, and M H Kohn 1998 Empirical evaluation of preservation methods for faecal DNA. Molecular Ecology 7 1423-1428
33

Frenzel, L D 1974 Occurrence of moose in food of wolves as revealed by scat analyses: a review of North American stiidies Le Natiiraliste Canadien 101: 467-479
Fuller, T.K 1989 Population dynamics of wolves in north-central Minnesota. Wildlife Monograph 105
Garcia-Moreno. J . M D Matocq, M S Roy, E GefiFen, and R K Wayne 1996 Relationships and genetic purity of the endangered Mexican wolf based on analysisof microsatellite loci Conservation Biology 10: 376-389
Gerloff, U . C Schldtterer. K Rassmann, I. Rambold, G Hohmann, B Fmth, and D. Tautz 1995 Amplification of hypervariable simple sequence repeats (microsatellites) from excrememal DNA of wild living Bonobos (Pan paniscus) Molecular Ecology 4 515-518
Ginsberg, J R , and D W Macdonald 1990 Foxes, wolves, jackals and dogs An action plan for the conservation of canids World Conservation Union, Gland, Switzerland
Gottelli, D , C Sillero-Zubiri, G D Applebaum, M S Roy. D J Girman, J. Garcia-Moreno, E A. Ostrander, and R K Wayne 1994 Molecular genetics of the most endangered canid the Ethiopian wolf Ca?»5 5/ 1 15/5 Molecular Ecology 3 301-312
Green, J S , and J T Flinders 1981 Diameter and pH comparisons of coyote and red fox scats Journal of Wildhfe Management 45 765-767
Halfpenny, J 1986 .A field guide to mammal tracking in North America Johnson Printing Company, Boulder, Colorado, USA
Hall, R E , and K R Kelson 1959 The mammals of North .America The Ronald Press Company, New York, New York, USA
Handt, O , M Hoss, M Krings, and S Paabo 1994 Ancient DNA - methodological challenges Experientia 50 524-529
Hoss, -M , M Kohn, and S Pdlbo 1992 Excrement analysis by PCR. Nature 359: 199
Hdss, M , and S Piabo 1993 DNA extraction from Pleistocene bones by a silica-based purification method Nucleic Acids Research 21 3913-3914
Hoss, M 1994 More about the sihca method Ancient DNA Newsletter 2: 10-12.
34

Johnson, M K, R C Belden, and D R Aked. 1984 Differentiating mountain lion and bobcat scats. Journal of Wildlife Management 48: 239-244.
Kocher, T. D , W K Thomas, A Meyer, S V. Edwards, S Paabo, F. X Villablanca, and A. C Wilson 1989 Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers Proceedings of the National Academy of Sciences of tiie United States of America 86: 6196-6200.
Kohn, M H . F Knauer, A StoflFella, W Schroder, and S Piiabo 1995 Conservation genetics of the European brown bear - a study using excremental PCR of nuclear and mitochondrial sequences Molecular Ecology 4: 95-103
Kohn, M H . and R K Wayne 1997 Facts from feces revisited Trends in Ecology and Evohition 12 223-227
Korschgen, L J 1980 Procedures for food-habits analyses Pages 113-127 mH S Mosby, R. H Giles, Jr, and S D Schemnitz, editors Wildlife management techniques manual, 4*** edition: revised The Wildlife Society, Washington, D C . USA
Lehman, N , A Eisenhawer, K Hansen, L D Mech, R O Peterson, P J P Gogan, and R K Wayne 1991 Introgression of coyote mitochondrial DNA into sympatric North .American gray wolf populations Evolution 45 104-119
Lucchini, \'., E Fabbri, F Mamcco, S Ricci, L Boitani, and E Randi 2002 Noninvasive molecular tracking of colonizing wolf (Con/i lupus) packs in the western Italian Alps Molecular Ecology 11 857-868
Major, M . M K. Johnson, W S Davis, and T F Kellogg 1980 Identifying scats by recovery of bile acids Journal of Wildlife Management 44 290-293
.McBride, R T 1980 The Mexican wolf (Caw/5 lupus baileyi) a historical review and observations on its stams and distribution Endangered species report 8 Unrted States Fish and Wildlife Service, Albuquerque, New Mexico, USA
.Mech, L D 1966 The wolves of Isle Royale, United States National Park Service Fauna Series Number 7
Mech, L D 1970 The wolf ecology and behavior of an endangered species University of MinnesoU Press, Minneapolis, Minnesota, USA.
Miller, S A , D D Dykes, and H F Polesky 1988 A simple salting out procedure for extracting DNA from human nucleated cells Nucleic Acids Research 16: 1215.
35

Morin, P A . J Wallis, J J Moore. R Chakraborty, and D S Woodmff" 1993 Noninvasive sampling and DNA amplification for paternity exclusion, community stmcttire, and phylogeography m wild chimpanzees Primates 34 347-356.
Moritz, C 1994 Applications of mitochondrial DNA in conservation: a critical review. Molecular Ecology 3 401 -411.
Mullis, K B , andP A Faloona 1987 Specific synthesis of DNA/n v//ro via a polyraerase-catalysed chain reaction Methods Enzymol 155: 335-350
Mullis. K B . F Ferre. and R .A Gibbs, edhors 1994 The polymerase chain reaction Birkhauser, Boston, Massachusetts, USA
Murie, A 1944 The wolves of Mount McKinley United States National Park Service Fauna Series, Number 5
Murie. O J 1954 Animal tracks, 2™* edhion Peterson Field Guides Houghton Mifflin Company. Boston, Massachusetts, USA
Nowak, R. M 1995 Another look at wolf taxonomy Pages 375-397 in L N Carbyn, S H Fritts. and D R Seip, editors Ecology and conservation of wolves in a changing world Occasional Publication 35 Canadian Circumpyolar Instimte, Edmonton, Alberta, Canada
Ott, L 1988 An introduction to statistical methods and data analysis, 3" edition PWS-KENT Publishing Company. Boston, Massachusetts, US.A
Paquet, P C 1992 Prey use strategies of sympatric wolves and coyotes in Riding Mountain National Park, Manitoba Journal of Mammalogy 73 337-343
Peterson, R O 1974 Wolf ecology and prey relationships on Isle Royale Dissertation Purdue L niversity, Lafayette, Louisiana, USA
Phillips, M K.andD W Smith 1996 The wolves of Yellowstone Voyageur Press. Stillwater, .Minnesota, USA
Pilgnm, K L , D K Boyd, and S H Forbes 1998 Testing for wolf-coyote hybridization in the Rocky Mountains using mitochondrial DNA Journal of W ildlife Management 62 683-689
Putman, R J 1984 Facts from faeces Mammal Review 14 79-97
Quinn, T , and W R Jackman 1994 Infiuenceof diet on detection of fecal bile acids by thin-layer chromatography Journal of Wildlife Management 58 295-299
36

Randi, E , L Gentile, G Boscagli, D Huber, and H. U Roth 1994 Mitochondrial DNA sequence divergence among some west European brown bear (Ursus arctos L.) populations Lessons for conservation Heredity 73 480-489.
Reed, J Z , D J Tollit, P M Thompson, and W Amos 1997 Molecular scatology: the use of molecular genetic analysis to assign species, sex and individual identity to seal fifieces Molecular Ecology 6: 225-234
Saiki, R K . S Scharf, F Faloona, K B Mulhs, G T. Hom, H A Eriich, and N. Amheim 1985 Enzymatic amplification of B-globin genomic sequences and restriction she analysis for diagnosis of sickle cell anemia Science 230: 1350-1354
Saiki, R K , D R Gelfand, S Stoffel, S J Scharf, R Higuchi, G T Horn, K B Mullis, andH .A Eriich 1988 Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase Science 239 487-491
Sambrook, J . E F Fritsch, and T Maniatis 1989 Molecular cloning a laboratory manual. 2° edition Cold Spring Harbour Laboratory Press. New York, USA
Scott, T G 1941 Methods and computation in fecal analysis with reference to the red fox United States Department of the Imerior, Fish and Wildlife Service Iowa State College Journal of Science 15: 279-285
Scott, T G 1943 Some food coactions of the northern plains red fox Ecological Monographs 13 427-479
Slade, R W, C Moritz, and A Heideman 1994 Muhiple nuclear-gene phytogenies: apphcation to pinnipeds and comparison with a mitochondrial DNA gene phylogeny Molecular Biology and Evolution 11 341-356
Spaulding, R L , P R Krausman, and W B Ballard 1997 Summer diet of gray wolves, Canis lupus, in northwestern Alaska The Canadian Field-Naturalist 112: 262-266
Stephenson, R O , and L Johnson 1972 Wolf report, Alaska Federal Aid m Wildlife Research Report Project W-17-3, Juneau, Alaska, USA
Stokes, D W, andL Q Stokes 1986 A guide to animal tracking and behavior Little, Brown and Company, Boston, Massachusetts, USA
Taberlet, P , and J Bouvet 1992 Bear conservation genetics Nature 358 197
37

Taberlet, P , J. J E Swenson, F Sandegren, and A. Bjarvall. 1995. Localization of a contact zone between two highly divergent mitochondrial DNA lineages of the brown bear Ursus arctos in Scandinavia Conservation Biology 5: 1255-1261
Taberlet, P , S Griffin, B Goossens, S Questiau, V Manceau, N, Escaravage, L. P Waits, and J Bouvet 1996 Reliable genotyping of samples with very low DNA quantities using PCR Nucleic Acids Research 24 3189-3194.
Taberlet. P , J -J Camarra, S Griffin, E Uhres, O Hanotte, L P Waits, C Dubois-Paganon, T Burke, and J Bouvet 1997. Noninvasive genetic tiacking of the endangered Pyrenean brown bear population. Molecular Ecology 6: 869-876.
Taberlet, P . L P Waits, and G Luikart 1999 Noninvasive genetic sampling: look before you leap Trends in Ecology and Evolution 14: 323-327.
Thompson, D Q 1952 Travel, range, and food habits of timber wolves in Wisconsin Journal of Mammalogy 33: 429-442
Turkowski, F J 1980 Camivora food habits and habitat use in ponderosa pine forests United States Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station, Research Paper RM-215
United States Fish and Wildlife Service 1993 Biological evaluation for listed, proposed, and sensitive species of the forest-wide animal damage management plan on the Apache-Sitgreaves National Forests United States Fish and Wildlife Service. Phoenix, Arizona, USA
United States Fish and W ildlife Service 1996 Rdntroduction of the Mexican wolf within its historic range in the southwestem United States Final environmental impact statement. United States Fish and Wildlife Service, Albuquerque, New Mexico, USA
Van Ballenberghe, V , A W Erickson, and D Byman 1975 Ecology of the timber wolf in northeastern Minnesota Wildlife Monograph 43
\ ila, C , and R K Wayne 1999 Hybridization between wolves and dogs Conservation Biology 13: 195-198
Walsh, P S , D A Metzger, and R Riguchi 1991 Chelex* 100 as a medium for simple extt-action of DNA for PCR-based typing from forensic material BioTechniques 10: 506-513
38

Wasser, S K., C S Houston, G M Koehler, G G Cadd, and S R. Fain 1997 Techniques for application of faecal DNA methods to field studies in Ursids Molecular Ecology 6 1091-1097
W ayne, R K , N Uhman, M W Allard, and R. L Honeycutt 1992. Mitochondrial DNA variability of the gray wolf genetic consequences of population decline and habitat fiiigmentation Conservation Biology 6: 559-569.
Weaver, J L , and S H Fritts 1979 Comparison of coyote and wolf scat diameters. Journal of Wildlife Management 43 786-788
W ilcomb, M S 1956 Fox populations and food habits in relation to game bird survival, W illiamette Valley, Oregon. Oregon State College Agricultursd Experiment Station Technical Bulletin 38
Williams, B K 1982. Some observations on the use of discriminant analysis in ecology Ecology 64 1283-1291.
Woods, J G, D Paetkau, D Lewis, B N McLellan, M Proctor, and C Strobeck 1999 Genetic tagging of free-ranging black and brown bears Wildlife Society Bulletin 27 616-627
39
FIELD SCATS TISSUE Wl _Ci W2 C2
KNOWN SCATS Wl DJ £ 1
250 bp ^
150 b p ^
Figure 3 1 Agarose gel showing a pxDrtion of the mtDNA control region (D-loop) amplified via Pilgrim et al (1998) canid-specific primers (Mexican wolf and dog, 164 bp; coyote 160 bp) Lanes 1 and 2 are field-collected scats Mexican wolf (WI) and coyote (CI) Lanes 3 and 4 are tissues Mexican wolf blood (W2) and coyote hver (C2) Lane 5 (S) is a 50 bp DNA size standard Lanes 6 through 8 are known scats: Mexican wolf (W I), domestic dog (Dl), and coyote (CI)
40

DIGESTED Wl W2
UNDIGESTED CI C2 C3
250 bp ^
150bp->
100bp->
Figure 3 2 Agarose gel showing restriction fragment digestion with BstN I of mtDNA control region (D-loop) purified polymerase chain reaction (PPCR) isolated from Mexican wolf and coyote scats The first lane is a 50 bp DN.A size standard Lanes 2 and 3 show digested Mexican wolf mtDNA (W1-W2, 113 bp), and the last 3 lanes show undigested coyote (C1-C3, 160 bp)
41
V5
« -
35 -
X -
25 -
2D -
35 S mm
27 8 ^u^_r-
r =22 8mm
174 mm
x = 26.0 mm
16.3 nun
Coyote Species
Mexican wolf
Figure 3 3 Comparison of diameters of coyote scats ((// = 21. range 17 4 to 27 8 m, X = 22 8 mm) and Mexican wolf scats {n = 26, range 16 3 to 35 8 mm, x = 26 0 mm) idemified with fecal DN.A analysis
42

Table 3 1 Comparison of accuracy of predicting which species (Mexican wolf or coyote) deposited a scat using discriminant analysis based on combination of 3 measurements taken from scats identified to species with DNA analysis Scats were collected in southeastern Arizona and southwestem New Mexico from April 1998 to October 2001
I 2 3 4
Classification Diameter (mm) Diameter and mass (g) Diameter and length Diameter, mass and length (cm)
Coyote (%) 81 86 68 79
Mexican wolf (%) 50 65 59 55
43

Table 3.2. Locations of scats (n = 47) identified as Mexican wolf or coyote with DNA analysis Scats were collected in southeastern Arizona and southwestem New Mexico from April 1998 to October 2001
Location Forest trails Forest roads and highways
Forest roads Forest 2-tracks Highway
Elk carcass Mexican wolf den Opened release pen Otiier Total
Mexican wolf 10 10
1 1 0 4 26
6 3 1
11 6
0 0 2 2
21
Coyote
4 2 0
Total 21 16
1 1 2 6 47
44

CHAPTER IV
DIETS OF FREE-RANGING MEXICAN GRAY WOLVES
IN ARIZONA AND NEW MEXICO
Abstract
There were no systematic diet smdies of Mexican gray wolves (Canis lupus
baileyi) before their extupation by the late 1960s from the southwestem United States.
We collected carnivore scats (n = 1,682) from the Apache and Gila National Forests of
Arizona and New Mexico from April 1998 through October 2001 and identified the scats
to species using traditional field methods (i e , diameter, location, and sign) and odor
We verified the accuracy of scat identification with fecal DNA analysis (molecular
scatology, n = 26, Reed 2004) to determine the diets of free-ranging Mexican wolves in
Arizona and New Mexico We analyzed scats > 28 mm diameter (n = 55) collected from
areas where Mexican wolf packs (n = 6) were fed supplemental food items and
determined if the packs consumed non-supplemental food items to detect a shift in diet as
they acclimated to a wild existence We foimd a difference (Go, = 12 995, P = 0 023) m
diet composition among packs, with the Cienega and Hawks Nest packs consuming more
non-supplememal food hems than the Lupine, Campbell Blue, Mule and Pipestem packs.
We analyzed Mexican wolf scats (w = 251) identified with our refined traditional field
methods established with DNA analysis (Reed 2004) and found the diet composition of
tiiose scats consisted of large-sized food items (92 8% percent frequency of occurrence
[PFO]), primarily elk (Cervus elaphus canadensis [nelsonij) adults (36 6% PFO) and
45

calves (36 2% PFO). We found no difference in Mexican wolf diets among years (n = 4,
God, = 2 588, P = 0 460). between seasons (n - 2. G^j, = 0 490, P = 0 484), or among
packs (n = 4. G.^= 7.719, P = 0.052) We analyzed Mexican wolf scats (n = 26)
identified witii DNA analysis and compared the resuhs to tiie diet composition of
Mexican wolf scats (n = 251) identified witii our refined ti-aditional methods. There was
a difference (G*^ = 9 761, P = 0 002) in diet composition between the 2 methods, with
tiie scats idemified witii DNA analysis composed of fewer large-sized food items (72.7%
PFO) than found in the scats identified with our refined tiaditional methods (92.8%
PFO) However, we found no difference (/ = - I 95, f = 0 001) between diameter means
(refined tt-aditional method x = 29 3 mm, DNA analysis r = 26 0 mm) and no difference
(/ = 1 50,P = 0 005) between number of food items per scat for the 2 identification
methods (refined traditional method = 1 06 food items per scat, DNA analysis 1 27 food
items per scat) We then compared diet composition found in scats identified with DNA
analysis as Mexican wolf scats (n = 26) and sympatric coyote scats (C. latrans: n = 2l)
and found a difference (Gajj = 9 647, P = 0002) Mexican wolf diets consisted primarily
of large-sized food items (72 7% PFO), while coyote diets were more variable and
composed of more medium- and small-sized food items (63 4% PFO) There was a
difference (/ = -2 428, P = 0 019) between diameter means for the 2 species (Mexican
wolf X = 26 0 mm, coyote x = 22 8 mm. Reed 2004) There was also a difference (/ =
2 849, P = 0 007) between number of food items per scat for the 2 species (Mexican wolf
X = 1 27 food hems per scat, coyote x = I 95 food items per scat) We compared the diet
composition of Mexican wolf scats (n = 277) identified with our refined traditional field
46

methods (n = 251) and DNA analysis (n = 26) to tiie diets reported in previous North
American gray wolf (C. lupus) stiidies (n = 7) and found a difference (Gadj = 462,492,
P = < 0 0001) Mexican wolf diet consisted primarily of large-sized food items (90.6%
PFO), which differed from late spring and summer- and carcass-based diets previously
reported for North American gray wolves Our results suggest that free-ranging Mexican
wolves consumed more large-sized food items than either sympatric coyotes or other
North .American gray wolves
Introduction
Little is known of the Mexican gray wolfs (Ccaiis lupus baileyi) natural history
because systematic studies were not conducted on the subspecies before it was extirpated
from the wild (Brown 1983) The Mexican wolf is the smallest and southern-most
occurring gray wolf (C lupus) of North America (Young and (joldman 1944, Hall and
Kelson 1959, Bogan and Mehlhop 1983, Nowak 1995) The subspecies is also the most
genetically distinct (Wayne et al 1992, Garcia-Moreno et al 1996) and the most
endangered (McBride 1980, Brown 1983, Bednarz 1988, Ginsberg and Macdonald 1990)
North .American gray wolf The United States Fish and Wildlife Service (USFWS) began
releasing captive-reared Mexican wolves in April 1998 into the Blue Range Wolf
Recovery Area (BRWRA) in Arizona and New Mexico
The purpose of our smdy was to determine the diets of free-ranging Mexican gray
wolves in Arizona and New Mexico and to determine if there were any differences
among years, between seasons, or among packs We also determined if non-
47

supplemental food items were consumed by Mexican wolves while they were fed
supplemental food items From the resuhs of our fecal DNA analysis (molecular
scatology. Reed 2004). we were able to compare the diet composhion of Mexican wolves
with that of sympatric coyotes (Ca/iis latrans) Lastiy we compared the diet composition
of Mexican wohes to that reported in 7 previous diet studies of northern gray wolves
This information could prove beneficial for managing Mexican wolf and prey
populations and investigating depredation incidences in areas where Mexican wolves are
recovering in the southwestem United States
Study Area
We conducted our research within the BRWTIA, which encompassed 17,700 km
and included all of the .Apache and Gila National Forests in east-central Arizona and
west-central New Mexico The primary recovery zone (2,600 km') in Arizona, in which
capti\e-reared Mexican wolves were released since April 1998. was our principal focus
In Spring 2000, our research efforts extended to the secondary recovery zone (Gila
National Forest and Wilderness, New Mexico) due to translocations of Mexican wolves
previously released in .Arizona The White Mountain (Fort Apache) and San Carlos
Indian Reservations bordered the BRWRA on the west and private lands bordered the
area to the east, north and south, and were scattered within public lands Domestic cattle
grazed almost all areas (USFS, unpublished data)
The BRWRA s elevations ranged from about 1,200 m along the San Francisco
River to the 3,350 m mountain ranges of Mount Baldy. Escudilla, and Mogollon (United
48

States Fish and Wildlife Service 1996) Lower elevations were characterized by rolling
hills with moderately steep-walled canyons and sandy washes, while higher elevations
were typified by mgged slopes, deep canyons, elevated mesas, and rock cliffs (Umted
States Fish and Wildlife Service 1996) Dominant vegetation included ponderosa pine
(Pittus ponderosa), aspen (Popuhs tremuloides), fir (Pseudotsuga menziesii and Abies
spp ), juniper (Juniperus spp ), piflon (Pinus cembroides), mesquhe (Prosopis spp ),
evergreen oaks (Quercus spp ), and a variety of grasses and forbs (United States Fish and
Wildlife Service 1996) Annual temperatures averaged 16 4°C maximum and-3.1°C
minimum and most precipitation fell during thunderstorms (annual average 52.1 cm)
from July through September and snow (annual average 139 3 cm) from December
through March (Desert Research Institute, Western Region Climate Center, Reno,
Nevada, unpublished data)
From April 1998 through October 2001, USFWS reported > 87 Mexican wolves
either released from captivity or bom in the wild within the BRWRA Approximately 37
Mexican wolves were free-ranging as of October 2(X)1 and 31 of those individuals were
fitted with radio collars Other predators within the study area included humans (Homo
sapiens), mountain lion (Puma concolor), black bear (Ursus americanus), coyote, bobcat
(Felis rufus), and gray (Urocyon cmereoargenleus) and red (Vulpes vulpes) fox (United
States Fish and Wildlife Service 1993, Arizona Game and Fish Department 1994)
Density estimates were unavailable for these species
Potential large-sized prey within the BRWRA included Rocky Mountain elk
(Cerxvs elaphus canadensis [nelsoni]), Coues white-tailed (Odocoileus virginianus
49

couesi) and desert mule (O. hemionus eremicus) deer, pronghom (Antilocapra
americana\ and Rocky Mountain bighorn sheep (Ovis canadensis. United States Fish
and Wildlife Service 1996). It was estimated that tiie BRWRA supported 57.000 deer
and 16,000 elk (Parsons 1998); however, the densities of these ungulates were unknown
during our study. Potential medium- and small-sized prey included javelina (Dicotyles
kyacu. United States Fish and Wildlife Service 1996), beaver (Castor canadensis),
jackrabbit (Lepus spp ), cottontail rabbit (Syhnlagus spp ), skunk (Mephitis spp), various
tree (Schmts and Tamiasciurus spp ) and ground squirrels (Spermophilus spp), chipmunk
(Eutamiasspp),rat(Neotomaspp),mice(Per<miyscusspp), vole(Microtusspp), and
other small mammals (Hofifmeister 1986, Southwest Region US Forest Service 1992),
porcupines (Erethizon dorsatum), and Merriam's turkey (Meleagris gaJlopavo, Groebner
etal. 1995).
Elk calving occurred during May and June, mule deer fawning occurred June
through August, and white-tailed deer fiiwning occurred during August (AGFD,
unpublished data. New Mexico Department of Game and Fish 1998—2001). Native
ungulate harvesting occurred intermittently from late August through January for white-
tailed and mule deer and from September through mid-December for elk (Arizona Game
and Fish Departmem 1998—2001, New Mexico Department of Game and Fish 1998—
2001), which could have resulted in carrion for Mexican wolves.
50

Methods
We collected 1.682 carnivore scats from April 1998 through October 2001 from
areas where captive-released, translocated, and wild-bom Mexican wolves were known to
frequent within the BRWRA on the Apache and Gila National Forests of Arizona and
New Mexico Personnel of the Mexican wolf Interagency Field Team (IFT, i e , US
Fish and W ildlife Service, .Arizona Game and Fish Department, New Mexico Department
of Game and Fish, USDA APHIS Wildlife Services, and U S Forest Service) collected
scats opportumstically from April 1998 through October 2001 and we collected scats
actively from June through August 20(X) and March through October 2001 An
opportunistic sampling strategy (Frenzel 1974) was employed as the sampling areas
followed Mexican wolves as they moved within the study area We collected scats along
forest roads, trails, ridgelines, and riparian areas, and from opened release pens,
campsites, den sites, and kill and carcass sues Scats collected by 4 Drag Ranch
personnel in areas where Mexican wolves were repyortedly depredating cattle were not
available for this smdy
Scat collectors wore disposable mbber or food preparation gloves and placed the
scats in brown paper bags, which were labeled with date, location, and scat number
Scats were aged as old, recent, or fresh based on appearance, exposure of deposition site,
and weather conditions (Ciucci et al 1996) We identified scats to carnivore species
using traditional field methods (i e , diameter, location, and sign) reported in the hterature
and odor There were no reports of Echinococcus granulosus in Arizona or New Mexico,
therefore we did not autoclave the scats The bagged scats were ah dried before storing
51

at room temperatiire in large plastic comainers until fiirther analysis We used DNA
analysis to verify traditional scat idemification methods for a subsample of scats (Reed
2004). From our DNA analysis resuhs, we refined the tiaditional scat identification
techniques as described below
The location criterion was defined as den sites (n = 2) only We measured the
maximum diameter of each dried scat (Scott 1943, Weaver and Fritts 1979, Green and
Flinders 1981. Danner and Dodd 1982) using 152 mm dial calipers (General Tools
Manufacturing Co . LLC. New ^ ork. New York) However, we took 2 measurements to
the nearest 0 I mm and used the average for diameter size The diameter criterion for
identifying gray wolf scats was > 24 mm diameter established by Thompson (1952) and
has been accepted for several smdies (Mech 1970. Stephenson and Johnson 1972.
Peterson 1974, Nan Ballenberghe et al 1975) Weaver and Fritts (1979) suggested scat
diameters > 30 mm be used to identify gray wolf scats Halfpenny (1986) proposed > 25
mm diameters for identifying gray wolf scats after reviewing 3 studies For identifying
Mexican wolf scats in the field, we chose the more conservative > 30 mm diameter
criterion, then redefined it as > 28 mm based on results from DNA analysis (Reed 2004).
We used wolf tracks and visual observations for sign criterion Some wolf biologists
believe that they can identify wolf scats by odor (Halfpenny 1986) We attempted to
idemify Mexican wolf scats based on odor (eg , a sweet, musky odor) as instmcted by
.Mexican wolf IFT personnel No distinguishing odor for coyote scats was reported.
We established a reference collection of potemial prey items from the BRWRA
We trapped and euthanized small prey and collected road-killed small prey, which were
52

prepared as museum specimens Hair samples were taken from various body areas (eg.,
mmp, hind legs, belly, back, and neck) of road-killed elk and deer Elk calf hair samples
were taken from the carcass remains of a known Mexican wolf kill. Fawn hair samples
were taken from a 2-month-old mule deer orphan residing at Sipes White Mountain
Wildlife Refijge, Springerville, Arizona.
We estimated the diets of Mexican wolves from 332 of the 1,682 scats. Twenty-
one additional scats were positively identified as coyote scats with DNA analysis (Reed
2004) The remaining 1,329 scats were < 28 mm diameter and could not be positively
identified to carnivore species These scats were excluded from this diet study and were
analyzed for another
We analyzed 55 scats with diameters > 28 mm collected from areas where
Mexican wolves were fed supplemental food items (i e , 11 1 kg carnivore logs [zoo diet
formulation for wild canids. Central Nebraska Packing. Inc . North Platte, Nebraska,
USA] and road-killed elk, deer, and jackrabbit) By comparing food items found in these
scats with supplemental food records (United States Fish and Wildlife Service [USFWS],
unpublished data), we were able to determine if any non-supplemental food (i e , elk
aduhs and calves, deer adults and fawns, domestic bovine, and insects) were consumed.
We determined overall diet of non-supplemental fed Mexican wolves by
analyzing scats (n = 25\) identified as deposhed by the subspecies based on > 28 mm
diameter (Reed 2004), den site location, and sign criteria, plus the scats (n = 26)
identified as deposhed by Mexican wolves with DNA analysis (Reed 2004). The 251
scats identified by our refined traditional methods were used to determine differences
53

among years, between seasons, and among packs. We then compared the diet
composition of the Mexican wolf scats identified with our refined fradhional methods to
scats identified with DNA analysis We also compared the diet composition found in
Mexican wolf scats (n = 26) to coyote scats (n = 21) identified with DNA analysis (Reed
2004) to determine a difference between the 2 species Mexican wolf scats identified
with our refined tt^ditional methods (n = 251) and DNA analysis (n = 26) were combined
for comparison with 7 North American gray wolf diet studies to determme any
differences in diet composition
Scats were broken apart by hand and undigested food items (i e , hak, bone, teeth,
claw s, and hooves) were separated We identified the undigested food items
macroscopically by comparing to our reference collection
We calculated frequency of occurrence (FO) of prey items (i e . number of
occurrences of a prey item divided by the total number of scats) and percent frequency of
occurrence (PFO) of prey items (i e , number of occurrences of a prey item divided by the
total number of occurrences of all prey items) We used PFO for all statistical analyses.
To fiacilitate statistical comparisons, we pooled ungulate prey (i e , adult and neonate elk
and deer, and domestic bovine) and considered them large-sized food hems We then
pooled smaller food items (i e , medium- and small-sized mammals, birds, reptiles,
insects, and vegetation) and considered them medium- and small-sized food hems. We
did not include non-food hems (e g , rocks, sticks, pine needles, string, plastic, mbber,
paper, and ahiminum foil) We used /-tests to examine differences in diameter means and
number of food items per scat between Mexican wolf scats identified whh our refined
54

tt^ditional identification methods and DNA analysis, and between Mexican wolf and
sympatric coyote scats identified with DNA analysis We used chi-square likelihood
ratio contingency-table analysis (G-test. Ott 1988) corrected for continuity (Williams
1976) to determine the presence of non-supplemental food items found in scats identified
with our refined traditional techniques collected where Mexican wolves were fed
supplemental food items and to determine differences in diet composition among years
(1998—2001), between 2 seasons (spring-summer March—August, fall-winter:
September—Febmary), between packs (n = 4). between scat identification methods
(refined traditional and DN.A analysis), between species (Mexican wolves and sympatric
coyotes), and between Mexican wolves and northern gray wolves All tests were
considered significant at the probability level of/* ^ 0 05
Results
We found a difference {Godj - 12 995. P = 0 023. Table 4 1) in diet composhion
of scats (n = 55) > 28 mm diameter collected from areas where Mexican wolf packs
(n = 6) were fed supplemental food items Two packs, Cienega and Hawks Nest,
consumed non-supplemental food items while being fed supplemental food items, and
Lupine, Campbell, Mule and Pipestem packs did not (Table 4 2)
Diet composition of Mexican wolf scats (n = 251) identified with our refined
traditional field methods consisted mainly of native ungulates (88 6% PFO, Table 4 3).
Smaller mammalian prey (5 3% PFO), birds (0 4% PFO), insects (0,8% PFO), and
vegetation (0 8% PFO) were of lesser importance Cattle comprised 4 2% PFO We
55

found no differences in diet among years (n = A,G^ = 2 588, P = 0 460, Table 4.4),
between seasons (n = 2. G«* = 0 490, P = 0 484. Table 4 5), or among packs (// = 4,
C/a4=7 719, P =0 052. Table 4 6) All 3 comparisons indicated Mexican wolf diets
were composed mainly of large-sized food hems
Diet composhion of Mexican wolf scats (n = 26) identified with DNA analysis
consisted of a variable diet composed of large-sized food items (72 7% PFO; Table 4 7)
and medium- and small-sized food hems (27 3°'o PFO) We compared these results with
the diet composition of the scats identified with our refined traditional methods and found
a difference (Ga^ = 9 761. P = 0 002, Table 4 8) in diets based on scat identification
techniques Diet composhion of scats identified with our refined tradhional methods
consisted of more large-sized food items (92 8°o PFO) and fewer medium- and small-
sized food Items (7 2°o PFO) than found in the scats identified by DN.A analysis We
found no difference between diameter means (I = -\ 95, P =0 052) for the 2 scat
idemification methods (refined traditional method x = 29 3 mm diameter, DNA analysis
X = 26 0 mm diameter) We also found no difference between number of food hems per
Mexican wolf scat (t = I 50, P = 0 135) for the 2 scat identification methods (1 06 food
hems per scat identified with our refined traditional methods. Table 4 3, 1 27 food hems
per scat identified with DN.A analysis. Table 4 7)
We found a difference (G^d, = 9 647, P = 0 002, Table 4 9) in diet composhion
found in Mexican wolf (n = 26) and sympatric coyote (/»= 21) scats when the 2 species
were poshively identified with DNA analysis (Reed 2004). Mexican wolf diets consisted
mainly of large-sized food rtems (72 7% PFO), while coyote diet had a more variable diet
56

consisting of medium- and small-sized food items (63.4% PFO). We also found a
difference (t = 2.849, P = 0.007) in number of food hems per scat for the 2 species (1.27
food items per Mexican wolf scat; 1 95 food items per coyote scat. Table 4.9).
We pooled scats identified with DNA analysis (n = 26) and those identified with
our refined traditional field techniques (n = 251) to compare witii previous North
American gray wolf diet studies (n = 7). We found a difference (Gad/ = 462.492,
P = < 0.0001, Table 4.11) between the diets of Mexican wolves and other North
American gray wolves, with Mexican wolves consuming a higher percentage of large-
sized fixKl items (90.6% PFO) than reported for North American gray wolves (range
58 5% to 82 9% PFO).
Discussion
Most early researchers of the Mexican wolf based their assessment of the
subspecies' diet on field observations and stomach analysis (Young and Goldman 1944).
Bailey (1931) reported the subspecies fed primarily on cattie, as determined from
undigested hair found in scats. Leopold (19S9) hypothesized that the Mexican wolfs diet
consisted mainly of deer, but included bighorn sheep, pronghom, javelina, rabbits,
rodents, and some plam foods. McBride (1980) reported that deer were the principal
native prey of Mexican wolves, but they also consumed pronghom, rabbhs, and mice.
Brown (1983) speculated that the Mexican wolf historically preyed almost entu-ely on the
diminutive Coues white-tailed deer Parsons (1996) acknowledged that the Mexican
wolfs historic diets were not well documented, but suggested that the subspecies preyed
57

primarily on whrte-tailed and mule deer, and ahematively on elk, pronghom, javelina,
beaver, rabbits, hares, and other small mammals
Introduced domestic livestock increased in Arizona and New Mexico from 1880
to 1890 and native prey populations decreased due to unregulated subsistence and market
hunting (Unhed States Fish and Wildlife Service 1987) Historically, the BRWRA had a
plentiftil supply of elk, deer, and otiier wild game (Shumway 1998). By 1890 the native
elk (Cer\tis merriami) were extinct and current populations of Rocky Mountain elk were
tt^nslocated to tiie area around 1925 (Nelson 1902, Shumway 1998) With the reduction
of the large native ungulate prey base, the Mexican wolf reportedly turned to the more
abundam and more-easily caught livestock (Young and Goldman 1944, Brown 1983,
Parsons 1996) Leopold (1959) indicated that not all wolves were livestock killers and
that such behavior was that of particular packs or individuals Historical records of the
Mexican wolfs diets during the extirpation campaign focused primarily on wolves that
depredated domestic cattle and the reports may have been exaggerated (Gipson and
Ballard 1998)
To smdy the diets of free-ranging Mexican wolves, it was necessary to select a
sampling technique that would not interfere with recovery efforts Scat analysis is a
noninvasive field technique widely used in determining carnivore diets, and scats are
readily available and easily collected (Scott 1941, Putman 1984) However, carnivore
scats must be identified accurately to species for scat analysis to be a valid method of
determining the diet of the species under smdy (Reed 2004)
58

Frequency of occurrence of food hems per scat is the most commonly used and
easily applied method of diet analysis (Leopold and Krausman 1986, Spaulding et al
1997). although this method may overemphasize tiie frequency of small prey (Corbett
1989) We used the PFO method for this study because our objective was to determine
the diet composhion and relative amounts of food items consumed by Mexican wolves to
compare with previous North American gray wolf diet studies If PFO did over
emphasize smaller food rtems in the diet composhion of Mexican wolves, h was not
apparent in our resuhs
Diet analysis based upon scats identified with DNA analyses as deposhed by
Mexican wolves and sympatric coyotes revealed a difference between the 2 species'
diets, which was consistent with Thurber et al (1992) and Arjo et al (2002) who found
that coyote diets contained more small-sized food items than sympatric gray wolf diets
This could be contributed to morphological and social differences between the 2 species.
Coyotes are medium-sized carnivores that usually prey on smaller animals (Mech 1970,
Pilgrim et al 1998), and the presence of ungulate remains in their scats may resuh from
feeding on carrion (Ozoga and Harger 1966, Berg and Chesness 1978, Weaver 1979)
Wolves, on the other hand, are 3 to 4X the mass of coyotes and are more likely to hunt in
cooperative packs than coyotes, which allows preying on larger animals (Mech 1970,
Ballard et ai 1987, .Moehlman 1989, Thurt)er and Peterson 1993, Dale et al 1995) In
our smdy area, Mexican wolves weighed 22 7 to 40 9 kg (Unhed States Fish and Wildlife
Service 1996) and southwestem coyotes averaged 9 5 to 10 6 kg (Woolsey 1985)
59

Although our sample sizes were small, DNA analysis of scats may provide a more
comprehensive representation of wolf and coyote diets than analysis of scats identified to
species by ttadhional methods used by previous diet studies. Early northern gray wolf
diet smdies (Murie 1944. Cowan 1947. Thompson 1952, Mech 1966, Pimlott et al. 1969)
supported the conclusion that the wolfs diet consisted mainly of ungulate prey either
killed or scavenged (acmal predation and carrion feedmg cannot be distinguished by scat
analysis [Pimlott 1967]), as did more recent studies (Ballard et al 1987, Arjo et al. 2002).
Although Mech (1970) reported that wolves prey on sick, weak, and unfit prey.
Brown (1983) reported that there was no evidence in southwestem historic records that
indicated such hunting behavior by Mexican wolves However, during the first year of
Mexican wolf recovery efforts, all confirmed Mexican wolf prey carcasses were elk and
most remains were young of the year and old or injured individuals (Parsons 1998), and
the first documented native prey kill was by the Hawks Nest pack whhin 2 weeks after
tiieir release (USFWS, unpublished data) From May 1998 tiirough October 2001,
Defenders of Wildlife-Bailey Wildlife Foundation Wolf Compensation Tmst
(unpublished data) paid $16,612 (US) for confirmed and unconfirmed Mexican wolf-
related injuries to 2 horses, 1 calf, and 1 guard dog, and deaths of 1 bull, 2 cows, 1 heifer,
11 calves, and 1 herding dog within Arizona and New Mexico Our resuhs combined
with USFWS kill reports and Defenders of Wildlife-Bailey Wildlife Foundation Wolf
Compensation Trust reports indicated that Mexican wolves consumed more large-sized
native ungulates than domestic livestock
60

Mech (1970) and Messier and Crete (1985) indicated wolves concentrated on the
smallest or easiest to catch large prey species in areas where > 2 large prey species
inhabrt the same area Our resuhs suggest that Mexican wolves were consuming
primarily adult elk (males ^ 454 5 kg, females ^ 227 3 kg, AGFD, unpublished data) and
calves, the larger of tiie native ungulate species available within the BRWRA, and not the
smaller white-tailed deer (males < 56 8 kg, females <. 36 4 kg; AGFD, unpublished data)
or mule deer (males < 102 3 kg, females ^ 56 8 kg, AGFD, unpublished dau). When
compared to previous diet studies for North American gray wolves, our resuhs suggest
that Mexican gray wolves consumed a higher proportion of large-sized native ungulates
than their northern counterparts This may have been the result of our conservative
identification of Mexican wolf scats using diameters ^ 28 mm, which may have biased
our diet analysis results towards the larger prey commonly found in large diameter scats
(Daimer and Dodd 1982) Additionally, previous gray wolf diet smdies included scats
collected from den and rendezvous sites and kill or carcass shes, while our Mexican wolf
scats were collected from all locations frequented by the subspecies, during all seasons,
and 2 den srtes
Our results indicate that elk aduhs and calves were the primary food source for
Nfexican wolves in Arizona and New Mexico from April 1998 through October 2001
Gray wolves in differem areas rely on different prey, and usually the wolfs diet is
compnsed of 1 or 2 species (Mech 1970) Elk were the most abundant and largest prey
available whhin the BRWRA (AGFD, unpublished data, USFS, unpubhshed data)
Although we did not study prey selection, we were curious why Mexican wolves
61

consumed mostiy elk aduhs and calves We can only hypothesize based on previous
lueramre of prey selection studies of gray wolves First, the density of elk within
Arizona and New Mexico was unknown, however, elk densrty was reported to be high
(Ballard et al 1998. AGFD, unpublished data) Therefore, h is unknown if the elk herd
within the BRWRA had neared or reached ecological carrying capacity. Furthermore, we
did not know if elk were subjected to winter severity (Mech et al 2001), malnutrition,
disease, or injunes or death from human activity ( e g , hunting and vehicles) that possibly
played a role in the Mexican wolfs consumption of elk One reason that a large
proportion of elk remains were found in Mexican wolf scats may have been because of
the low-densrty, early colonizing stage of wolves (Fritts and Mech 1981. Boyd et al
1994) These authors, as did Carbyn (1974. 1983) and Huggard (1992), found that
wolves primarily preyed upon the most vulnerable ungulates juvenile, old, or post-mt
males These data and ours suggest low-density, colonizing Mexican wolves may have
preyed upon vulnerable elk at a higher rate than other wolves in established northern
populations
Another reason why Mexican wolves consumed primarily elk may be because of
wolf-naive elk Although the resident elk population had been exposed to other predators
(i e , mountain lions, black bears, and coyotes), the elk had not been exposed to wolves.
This may have increased the vulnerability of resident elk that had never seen wolves until
confronted with them in 1998 Naive prey confronted with new predators have been less
wary than prey previously exposed to such danger (Byers 1998, Berger 1999, 2001)
62

To answer why Mexican wolves consumed primarily elk, ftuther study would be
required Mexican wolves are 1 of the 4 major predator species in Arizona and New
Mexico, and little research has been conducted on the other wild carnivores (i.e.,
mountain lion, black bear, and coyote) that now share the same prey base A muhi-
carmvore prey selection study would provide the information necessary for optimal
management of predators and then prey within Arizona and New Mexico
Our estimates of the diets of free-ranging Mexican gray wolves in Arizona and
New Mexico were based on a small sample of scats This is a result of our conservative
approach to identif>ing Mexican wolf scats based on DNA analysis and refined
traditional identification methods (i e , > 28 mm diameter, den site locations, and tracks)
Our results suggest that our refined traditional scat identification methods may have
biased estimates of the diets of Mexican wolves toward large ungulate food items WTien
researching the diets of colonizing and established wolf populations, h is important to
provide wildlife managers and the public whh accurate information We recommend that
future gray wolf diet studies incorporate DNA analysis of scats to ensure that only gray
wolf scats are included in diet analyses for the species This approach could prove
beneficial in resolving human-wolf conflicts in areas where gray wolves are reintroduced
or recovering
63

Acknowledgements
The Unhed States Fish and Wildlife Service funded this research The Mexican
wolf Interagency Field Team (IFT, i e , US. Fish and Wildlife Service, Arizona Game
and Fish Department, New Mexico Department of Game and Fish, USDA APHIS
Wildlife Services, and U S Forest Service) p)ersonnel assisted with the collection of scats
and provided Mexican wolf location data The United States Forest Service Alpine and
Clifton Ranger District personnel assisted whh logistics This is a Texas Tech
University, College of Agricultural Sciences and Natural Resources technical publication
T-X-XXX
64

Literatiire Ched
Arizona Game and Fish Departmem 1994 Predator population estimates Unpublished report. Arizona Game and Fish I>epartment, Phoenix, Arizona, USA.
.\rizona Game and Fish Departmem 1998-2001. Arizona hunting regulations. Arizona Game and Fish Department, Phoenix, Arizona, USA.
-Arjo, W M . D H Pletscher, and R R Ream 2002 Dietary overlap between wolves and coyotes in Northwestern Montana Journal of Mammalogy 83: 754-766.
Bailey. V 1931. Mammals of New Mexico United States Department of Agriculttire, Bureau of Biological Survey of North American Fauna No 53
Ballard, W B . H A Whitlaw, B F Wakeling, R L Brown, J C deVos, Jr, and M C. Wallace 1998 Survival of female elk in northern Arizona Journal of Wildlife Management 64 500-504
Ballard, W B . J S Whitman, and C L Gardner 1987 Ecology of an exploited wolf population in south-central Alaska Wildlife Monograph 98
Bednarz, J C 1988 The Mexican wolf biology, history, and prospects for reestabhshment in New Mexico Endangered Species Report No 18 United Slates Fish and Wildlife Service, Albuquerque, New Mexico. USA
Berg, W E , and R A Chesness 1978 Ecology of the coyote in northern Minnesota Pages 229-247 m M Bekoff, editor Coyotes biology, behavior, and management Academic Press, New York, USA
Berger, J 1999 .Anthropegenic extinction of top carnivores and interspecific animal behaviour implications of the rapid decoupling of a web involving wolves, bears, moose and ravens Proceedings of Royal Society of London, Series B, 266: 2261-2267.
Berger, J , J E Swenson, and I Per-Illson 2001 Recolonizing carnivores and naive prey conservation lessons from Pleistocene extinctions Science 291 1036-1039.
Bogan, M A . and P Mehlhop 1983 Systematic relationships of gray wolves (Caw/5 lupus) in southwestem North America Occasional Papers of the Museum of Southwestem Biology 1
Boyd, D K , R R Ream, D H Pletscher, and M W Fairchild 1994 Prey taken by colonizing wolves and hunters in the Glacier National Park area Journal of Wildlife Management 58 289-295
65

Brown, D E 1983 The wolf in tiie Southwest: tiie making of an endangered species. The Universrty of Arizona Press, Tucson, Arizona, USA.
Byers, J A 1998. American pronghom: social adaptations and ghosts of predators past. University of Chicago Press, Chicago, Dlinois, USA
CarbvTi, L N 1974 Wolf predation and behavioral interactions with elk and otiier ungulates in an area of high prey diversity Dissertation, Universrty of Toronto, Ontario, Canada
Carbyn, L N 1983 Wolf predation on elk in Riding Mountain National Park, Manitoba Journal of Wildlife Management 47: 963-976
Ciucci, P , L Bortani, E Raganella Pelliccioni, M Rocco, and I Guy 1996. A comparison of scat analysis methods to assess the diet of the wolf Can/5 lupus Wildlife Biology 2 37-48
Corbett, L. K 1989 Assessing the diet of dingoes from feces a comparison of 3 methods Journal of Wildhfe Management 53 343-346
Cowan, I M 1947 The timber wolf in the Rocky Mountain national parks of Canada. Canadian Journal of Research 25 139-174
Dale. B W , L G Adams, and R T Bowyer 1995 Wimer wolf predation in a muhiple ungulate prey system. Gates of the Arctic National Park, Alaska Pages 223-230 m L N Carbyn, S H Fritts, and D R Seip. edhors Ecology and conservation of wolves in a changing world Canadian Circumpolar Instimte Occasional Publication 35
Danner. D A, and N Dodd 1982 Comparison of coyote and gray fox scat diameters. Journal of Wildlife Management 46 240-241
Frenzel, L D 1974 Occurrence of moose in food of wolves as revealed by scat analyses a review of North American smdies Le Naturaliste Canadien 101: 467-479
Fritts, S H, andL D Mech 1981 Dynamics, movements, and feedmg ecology of a newly protected wolf population in northwestern Minnesota Wildlife Monograph 80
Garcia-Moreno, J , M D Matocq, M S Roy, E Geflfen, and R K Wayne 1996 Relationships and genetic purity of the endangered Mexican wolf based on analysisof microsatellhe loci Conservation Biology 10 376-389,
66

Gmsberg, J R . and D W Macdonald 1990 Foxes, wolves, jackals, and dogs. An action plan for the conservation of canids Worid Conservation Union, Gland, Switzerland
Gipson, P S . and W B Ballard 1998 Accounts of famous North American wolves. The Canadian Field-Nattiralist 112: 724-739
Green, J S , and J T Flinders 1981 EMameter and pH comparisons of coyote and red fox scats Journal of Wildhfe Management 45: 765-767
Groebner, D J, A L Girmendonk, and T B Johnson 1995 A proposed cooperative reintroduction plan for the Mexican wolf in Arizona Arizona Game and Fish Departmem, Phoenix, Technical Report 56
Halfipenny, J. 1986 A field guide to mammal tracking in North America. Johnson Printing Company, Boulder, Colorado, USA
Hall, R E , and K R Kelson 1959 The mammals of North America The Ronald Press, New York, New York, USA
Hoffmeister, D F 1986 Mammals of Arizona University of Arizona Press and Arizona Game and Fish Department, Tucson, .Arizona, USA
Hoss, M , M Kohn, and S Paabo 1992 Excrement analysis by PCR Namre 359: 199.
Huggard, D J 1992 Prey selectivity of wolves in Banff National Park II Age, sex, and condhion of elk Canadian Journal of Zoology 71:140-147
Leopold, A S 1959 Wildlife of Mexico the game birds and mammals University of California Press, Berkeley and Los Angeles, California, USA.
Leopold, B D , and P R Krausman 1986 Diets of 3 predators in Big Bend National Park, Texas Journal of W ildlife Management 50 290-295
McBride, R T 1980 The Mexican wolf ((awi5/«pM5 ^a/Vey;) Unhed States I>epartment of the Interior, Fish and Wildlife Service Endangered Species Report No 8
Mech, L. D 1966 The wolves of Isle Royale United States Department of the Interior, National Park Service Fauna Series No 7
Mech, L. D 1970 The wolf the ecology and behavior of an endangered species Universrty of Minnesota Press, Minneapolis, Minnesota, USA
67

Mech, L D . D W Smith, K M Murphy, and D R. MacNuhy 2001. Winter severity and wolf predation on a formerly wolf-free elk herd Journal of Wildlife Management 65 998-1003
Messier, F , and M Crete 1985. Moose-wolfpopulationdynamicsand the natural regulation of moose populations Oecologia(Berhn) 65: 503-512.
Moehlman, P D 1989 Intt-aspecific variation in canid social units Pages 143-163 in J. L Gittleman, edrtor Carnivore behavior, ecology, and evolution. Cornell Umversrty Press, Ithaca, New York, USA
Murie. A 1944 The wolves of Mount McKinley United States National Park Service Fauna Series No 5
Nelson, E W 1902 A new species of elk from Arizona Bulletin of the American Museum of Natural History 16 1-12
New Mexico Department of Game and Fish 1998—2(X)1 Big Game and Furbearer Rules and Information New Mexico Department of Game and Fish, Santa Fe, New Mexico, US.A
Nowak, R M 1995 .Another look at wolf taxonomy Pages 375-397 m L N Carbyn, S H Fritts. and D R Seip, editors Ecology and conservation of wolves in a changing world Occasional Publication No 35 Canadian Chcumpolar Institute, Edmonton, Alberta, Canada
Ozoga, J L , and E M Harger 1966 Winter activhies and feeding habrts of northern Michigan coyotes Journal of Wildlife Management 30 809-818
Ott, L 1988 An introduction to statistical methods and data analysis, 3"* edhion PWS-KENT Publishing Company, Boston, Massachusetts, USA
Parsons, D R 1996 Case stiidy the Mexican wolf Pages 101-123 /n E A. Herrera and L F Huermeke, editors New Mexico's nattiral heritage biological diversity in the land of enchantment New .Mexico Journal of Science, Volume 36
Parsons, D R 1998 "Green fire" returns to the Southwest reintroduction of the Mexican wolf Wildlife Society Bulletin 26 799-807
Peterson, R O 1974 Wolf ecology and prey relationships on Isle Royale Dissertation Purdue University, Lafayette, Louisiana, USA
68

Pilgrim, K L , D K Boyd, and S H Forties 1998 Testing for wolf-coyote hybndization in tiie Rocky Mountains using mhochondrial DNA. Journal of Wildlife Management 62 683-689
Phnlott, D H 1967 Wolf predation and ungulate populations American Zoolocv 7 2t)7-278
Pimlott. D H, J A Shannon, and G B Kolenosky 1969 The ecology of tiie timber wolf in Algonquin Provincial Park Ontario Department of Lands and Forest Research Report No 87
Putman, R J 1984 Facts from feces Mammal Review 14 79-97.
Reed, J E , R J Baker, W B Ballard, and B T Kelly In review Differentiating Mexican wolf and coyote scats using DNA analysis Wildlife Society Bulletin
Scott T G 1 541 Methods and computation in fecal analysis with reference to the red fox Iowa State College Journal of Science 15: 279-285
Scott, T. G. 1943 Some food coactions of the northern plains red fox Joumal Paper No J-1131 Iowa Agricultural Experiment Station, .Ames, Iowa, USA
Southwest Region, United States Forest Service 1992 Fish, amphibians, reptiles and mammals a species checklist for the Gila National Forest Pamphlet, United States Department of Agriculture, Forest Service, Silver Citv, New Mexico, USA.
Spaulding, R L , P R Krausman, and W B Ballard 1997 Calculation of prey biomass consumed by wolves in northwest Alaska Joumal of Wildlife Research 2 128-132
Stephenson, R O , and L Johnson 1972 Wolf report, Alaska Federal Aid in Wildlife Restoration Report, Project W-17-3, Juneau, Alaska, USA
Thompson, D Q 1952 Travel, range, and food habhs of timber wolves in Wisconsin Joumal of Mammalogy 33 429-442
Thurber, J M, R O Peterson, J D Woolington, and J A Vucetich 1992 Coyote coexistence with wolves on the Kenai Peninsula, Alaska Canadian Joumal of Zoology 70 2494-2498
Thurber, J M , and R O Peterson 1993 Effects of population density and pack size on the foraging ecology of gray wolves Joumal of Mammalogy 74: 879-889
69

United States Fish and Wildlife Service 1993 Biological evaluation for listed, proposed, and sensrtive species of the forest-wide animal damage management plan on the Apache-Srtgreaves National Forests Unhed States Fish and Wildlife Service, Phoenix, .Arizona, USA
United States Fish and Wildlife Service. 1996 Reintroduction of the Mexican wolf within rts historic range in the southwestem United States: final environmental impact statement United States Fish and Wildlife Service, Albuquerque, New Mexico. USA
\'an Ballenberghe, V , .A W Erickson, and D. Byman 1975 Ecology of the timber wolf in northeastern Minnesota Wildlife Monograph 43
Wayne, R K , N Lehman, M W Allard, and R L Honeycutt 1992 Mitochondrial DN.A variabihty of the gray wolf genetic consequences of population decline and habitat fragmentation. Conservation Biology 6: 559-569
Weaver, J. L 1979 Influence of elk carrion upon coyote populations in Jackson Hole, W yoming Pages 152-157 m M S Boyce and L D Hayden, editors. North American elk ecology, behavior, and management University of Wyoming Press, Laramie, Wyoming, USA
Weaver, J L . and S H Fritts 1979 Comparison of coyote and wolf scat diameters Joumal of Wildlife Management 43 786-788
Williams, D A 1976 Improved hkelihood ratio tests for complete contingency tables. Biometrika 63 33-37
Woolsey, N G 1985 Coyote field guide Special report number 15, Arizona Game and Fish Commission, Phoenix, Arizona, USA
Young, S P , andE A Goldman 1944 The wolves of North America The American Wildlife Instittite, Washington, D C , USA
70

(/)
S g -gH
ilSl "•lis-
en 2
PL,
z
- I I I I I
I fS I <s
I
%
Z
Z
Z
(A
- J ! I
I -
r^
- I I I !
— — Tf 00
M ON • * rS
3
<S
S. I 3 i. S I 5 (J S £ C S
o
s 4-*
i a •a
men
t
4>
"B. o. S II
bu V3
siu
V _*rf
food
-3 c E
JU
supp
^ o c II
( L I C/}
Z
71

Table 4 2 Comparison of diet composhion found in scats (n = 55) ^ 28 mm diameter from areas where Mexican gray wolf packs (n = 6) were fed supplemental food hems (i e , carnivore logs and road-killed elk, deer, and jackrabbrt) to determine presence of non-supplemental food items ( eg , elk adults and calves, deer adults and fawns, domestic bovine, and insects) Scats were collected from April 1998 to October 2001 in Arizona and New Mexico and compared to Mexican wolf Interagency Field Team dated supplemental food records Comparison values are express as percent frequency of occurrence (PFO)
Pack Lupine Campbell Blue Mule Pack Pipestem Cienega Hawks Nest
Total
No food rtems 3
- I T
8 5 5 12 55
SF' 3 16 5 2 1 2 29
Food hems NSF^
1 6 3 3 4 10 27
NSF PFO (%)' 25 0 a 27.3 a 37.5 a 60 0ab 80 0ab 833 b 49 1
'SF = supplemental food items * NSF = non-supplemental food hems *PFO (%) = percent frequency of occurrence Percentages followed by the same lower
case letter were not significantly different (P>0 05, G-test. adjusted for continuity)
72
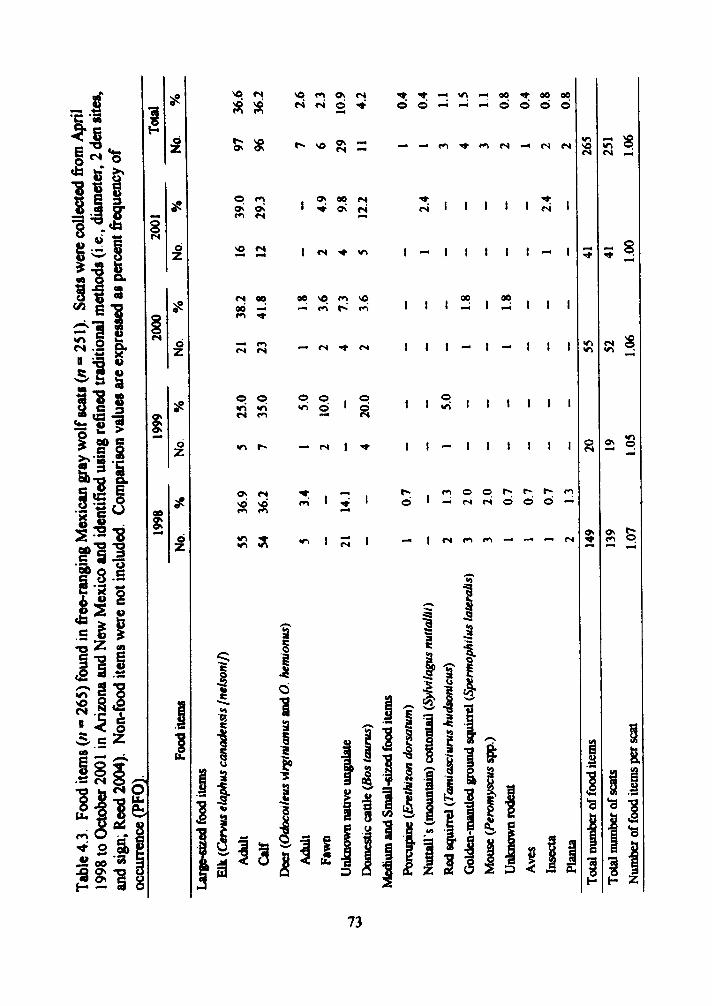
gb-
^ ^ «
O O
vO «N
r 4 00 00 —'
vO W 0 \ <S «s P4 d •^
t ^ vO Ov —
o\ ao
I «s • • «r\
— <N ' t <N
o o CO
O wS
O d tH4
, p ' d cs
- IN I
9- f>l >d vd
«n I
S "a •5
d d « <ri - - 00 ^ ^ ^M ^H O
' « 00
d d
-^ — r 0 ^ f < ^ « S - H j s ( M
l i l t !
I - I I I I I - I
I t 00 00
1 * 1 " I I I
I I » - I - I I I
i I ^ 1 1 1 I t I
J I - I I I I I I
f*> o o t * —i r< «s d
1^ r» d d
5!
.<:! S
OS O
I <s r^ n^ —• — —< M
1
?
I I:
!•
!13 I £
111 I § I I 5 I
a I .8 .<«
I 1 £
111 ( "8 !> il I .1 I . 13 2 3 •< a o.
i
•s
t= z
73

Table 4 4 Comparison among years (n = 4, 1998-2001) of diet composhion found in scats (n = 251) of free-ranging Mexican gray wolves collected from April 1998 to October 2001 in Arizona and New Mexico Scats were identified to species by refined tiaditional field methods (i e, diameter, location, and sign; Reed 2004). Food rtems were combined as large-sized (i e . aduh and young ungulates) food items and medium- and small-sized food items (i e . medium- and small-sized mammals, birds, insects, and vegetation). Non-food rtems were not included. Comparison values are expressed as percent frequency of occurrence (PFO)
Year 1998 1999 20OO 2001 Total
Large-sized food hems
135 19 53 39
246
Medium - and small-sized food rtems
14 1 2 2 19
Large-sized food hems PFO (%)'
90.6 95.0 964 95 1
'PFO ("*/o) = percent frequency of occurrence, none differed significantly (P>0 05, G-test, adjusted for cominuity)
74

Table 4 5 Comparison between seasons (;i = 2, fall-winter versus spring-summer) of diet composhion friund in scats (/i = 251) of free-ranging Mexican gray wolves in Arizona and New Mexico (April 1998 - October 2001) Scats were identified to species by refined traditional field methods (i e , diameter, location, and sign. Reed 2004) Food items were combined as large-sized (aduh and young ungulates) food hems and medium-and small-sized food rtems (i e , medium- and small-sized mammals, birds, msects, and vegetation) Non-food rtems were not included Comparison values are expressed as percent frequency of occurrence (PFO)
Season fall-winter spring-summer Total
Large-sized food items
36 210 246
Medium- and small-sized food hems
4 15 19
Large-sized food rtems PFO (%)'
900 933
'PFO (%) = percent frequency of occurrence, none differed significantly (P > 0 05, G-test. adjusted for continuity)
75

Table 4 6 Comparison of diet composhion among packs (n = 4) found in free-ranging Mexican wolf scats (n = 251) collected from April 1998 to October 2001 in Arizona and New Mexico Scats were identified by refined traditional field methods (i.e., diameter, location, and sign. Reed 2004) Food items were combined as large-sized (i.e., adult and young ungulates) food rtems and medium- and small-sized food hems (i e., medium- and small-sized mammals, bhds, insects, and vegetation) Non-food items were not included. Comparison values are expressed as percent frequency of occurrence (PFO).
Pack Hawks Nest Campbell Blue Francisco Cienega Total
Large-sized food rtems
66 98 20 26
210
Medium- and small-sized food rtems
8 9 0 0 17
Large-sized food hems PFO (%)'
892 91 6
100 0 1000
'PFO ("o) = percent frequency of occurrence, none differed significantly (P > 0 05, G-test, adjusted for continuity)
76

Table 4.7. Food items (n = 33) found in free-ranging Mexican gray wolf scats (n = 26) collected from April 1998 to October 2001 in Arizona and New Mexico. Scats were identified by DNA analysis (Reed 2004). Non-food items were not included. Comparison values are expressed as percent frequency of occurrence (PFO).
Food rtems Large-sized food items
Elk (Cervus elaphus canadensis fnels<mij) Aduh Calf
Deer (Odocoileus virginianus and 0. hemionus) aduh Unknown native ungulate Domestic cattle (Bos taunts)
Medium- and Small-sized food rtems Javelina (Dicotyles tajacu) Red squirrel (Tamiasciurus hudsonicus) Mouse (Peromyscus spp.) Unknown rodent Birds Insects
Total number of food items Total number of scats Number food items per scat
No.
5 14 2 I 2
1 I 2 2 2 1
33 26
I 27
PFO (%)'
15.2 42.4 6.1 3.0 6.1
3.0 3.0 6 1 6.1 6 1 30
'PFO (%) = percent frequency of occurrence
77

Table 4 8 Comparison of diet composhion of Mexican wolf scats (n = 277) collected from .April 1998 to October 2001 in Arizona and New Mexico and identified using 2 methods DN.A analysis (n = 2b) and refined traditional (n = 251, i e , > 28 mm diameter, 2 den srtes, and ttacks) Food items were combmed as large-sized (i e , aduh and young ungulates) food items and medium- and small-sized food hems (i e., medium- and small-sized mammals, bhds, insects, and vegetation) Non-food hems were not mcluded. Comparison \^ues are expressed as percent frequency of occurrence (PFO)
Method DN.A analysis Refined traditional
identification Total
Large-sized food hems
24 246
270
Medium- and small-sized food hems
9 19
28
Large-sized food hems PFO (%)'
72.7 a 92 8 b
'PFO (°'o) = percent frequency of occurrence Percentages followed by the same lower case letter are not significantly different (P > 0 05, G-te^, adjusted for continuity)
78

Table 4 9, Comparison of diet composhion found in free-ranging Mexican wolf scats (n = 26) and sympatric coyote scats (/»= 21) collected from April 1998 to October 2001 in Arizona and New Mexico Scats were identified by DNA analysis (Reed 2004). Food hems were combmed as large-sized (i e . aduh and young ungulates) food hems and medium- and small-sized food hems (i e , medium- and small-sized mammals, birds, insects, and vegetation) Non-food hems were not included Comparison values are expressed as percent frequency of occurrence (PFO)
Species Mexican gray wolf Coyote Total
Large-sized food hems
24 15 39
Medium- and small-sized food hems
9 26 35
Large-sized food rtems PFO (%)'
72.7 a 36 6 b
'PFO l" o) = percem frequency of occurrence Percertages followed by the same lower case letter are not significantly different (P > 0 05, G-test. adjusted for continuity)
79

Table 4.10. Food items found in free-rangmg Mexican gray wolf scats (n = 26) and sympatric coyote scats (n = 21) collected from April 1998 to October 2001 in Arizona and New Mexico. Scats were identified by DNA analysis (Reed 2004). Non-food hems were not mcluded. Comparison values are expressed as percent frequency of occiurence (PFO).
Food items Large-sized food items
Elk (Cervus ekphia cxmadensis [nelsoni]) Adult Calf
Deer (Odocoileus virginiamis and O. hemionus) Adult Fawn
Unknown nati\<e nngiiiat^ Domesbc cattle (Bos Uwrns)
Medium- and SnuO-sized food items Ja 'dina {Dicotyles tajacu) Nunall's (mountain) cottomail (Sytvilagus nuttallii) Red squind {Taniasdums hudsonicus) Golden-ffiantled ground squiirel (Spermophilus lauratis) Gny-coUared chipmunk Mouse {Peromyscus spp.) Unknown rodeot Shrew Bink Reptiles Insects VegEtatioa
Total nomter of food items Total mmter of scats Noraber food items per scat
Mexicanwolf No. PFO(%)'
5 15.2 14 42.4
2 6.1 — — 1 3.0 2 61
1 30 — — 1 30 — — — — 2 61 2 6 1 — — 2 6 1
1 3.0 _ _ 33 26
1.27
No.
2 7
1 2 -3
1 4 1 4 1 1 — 1 — 1 7 5
41 21
1.95
Coyote HfO(%)
4.9 17.1
2.4 4.9 — 7.3
2.4 9.8 2.4 9.8 2.4 2.4 — 2.4 — 2.4
17.1 12.2
'PFO (•/•) = percem frequency of occurrence
80

Table 4 11 Comparison of diet composhion found in free-ranging Mexican wolf scats (n = 277) and diet composition reported in other North American gray wolf diet studies (n = 7) Mexican wolf scats (/; = 270) were identified by refined traditional field methods (" = 251) and DNA analysis (n = 26. Reed 2004) Scats were collected from April 1998 to October 2001 in Arizona and New Mexico Food items were combined as large-sized (i e , adult and young ungulates) food items and medium- and small-sized food hems. Non-food items were not included Comparison values are expressed as percent frequency of occurrence (PFO)
Source Ballard etal (1987) Thompson (1952) Spaulding et al (1997) Murie (1944) Mech (1966) Cowan (1947) .Arjo etal (2002) This study
Large-sized food items
3,263 421
2,402 935 392 353 753 270
Medium- and small-sized food hems
2,316' 292'
1,082 406 124 94
155 28
Large-sized food hems PFO (%)'
58 5 a 59 0 a 68 9 b 697 b 76 0 c 79 0 cd 829 d 90 6 e
'PFO (" o) = percent frequency of occurrence Percentages followed by the same lower case letter are not significantly different (P > 0 05. G-test, adjusted for continuity)
^Data included unidentified ungulates and undefined "unidentified" food hems 'Data mcluded unidentified non-food items
81

CHAPTER V
SUMMARY
This was the first systematic stiidy to determine the diets of free-ranging Mexican
gray wolves (Carus hipus baileyi) in tiie soutiiwestem United States. We collected
carnivore scats (n = 1,682) from tiie Bhie Range Wolf Recovery Area (BRWRA) in
Arizona and New Mexico from April 1998 tiirough October 2001. We identified tiie
scats to species using ti^dhional field methods (i.e., diameter, location, and sign) and
odor. We then used fecal DNA analysis (molecular scatology) to verify the accuracy of
identifying Mexican wolf scats with tradhional methods and odor. DNA analysis of
Mexican wolf (n = 26) and sympatric coyote (» = 21) scats showed a 79% overlap in
diameto- size (Mexican wolf; 16 3 to 35 8 mm, coyote, 17 4 to 27 8 mm) and scats ^ 28
mm diameter were deposhed by Mexican wolves We found a difference (/ = -2.428,
P = 0 019) between diameter means for the 2 species (Mexican wolf F= 26 0 mm, coyote
X = 22.8 mm). From our DNA analysis resuhs, we refined the traditional methods for
identifying Mexican wolf scats as diameters ^ 28 mm, den site locations, and tracks, as
well as scats identified whh DNA analysis Diet analysis of Mexican wolf scats (n = 55)
with diameters > 28 mm collected from areas where 6 packs received supplemental
carnivore logs and road-killed elk, deer, and jackrabbit showed that non-supplemental
food items were consumed by Hawks Nest and Cienega packs (G«^ = 12.995, P = 0.023),
but not by Lupine, Campbell Blue, Mule or Pipestem packs. Diet analysis of Mexican
wolf scats (n = 251) identified according to > 28 mm diameter, den she location, and
82

tracks revealed a diet composhion consisting mainly of large-sized food hems (92.8%
PFO). primarily elk aduhs (36 6% PFO) and calves (36,2% PFO). There was no
difference in diets among years (n = 4. Gad/ = 2 588, P = 0.460), between seasons (n = 2,
Gad, = 0 490. P = 0 484), or among packs (n = 4, Gad, = 7 719, P = 0.052). We found a
difference (G^^ = 9 761, P = 0 002) in diet composhion of Mexican wolf scats identified
with our refined ttaditional methods (n = 251) when compared to that of DNA analysis
identified scats (n = 26) There were more large-sized food items in the scats identified
with our refined ttaditional method than found in scats identified with DNA analysis.
There was no difference (/ = -1 95, P = 0 001) between diameter means (refined
traditional method x = 29 3 mm, DNA method x = 26 0 mm) There was also no
difference (t = \ 50, P = 0 005) in number of food items per scat when comparing the 2
methods (refined tradhional method x 1 06 food items per scat, DNA method x = 1 27
food hems per scat) We found a difference ((}ad, = 9 647, Z' = 0 002) in diet composhion
of Mexican wolves and sympatnc coyotes, with Mexican wolf diets consisting primarily
of large-sized food hems (72 7°o PFO) and sympatnc coyote diets consisting primarily of
medium- and small-sized food items (63 4% PFO) There found a difference between the
2 species' scat diameter means {t = -2 428. P = 0 019, Mexican wolf x = 26 0 mm,
coyote X = 22 8 mm) There was also a difference (/ = 2 849, P = 0 007) between number
of food items per scat for the 2 species (Mexican wolf F = 1 27 food items per scat,
coyote x = 1 95 food items per scat) Finally, we found a difference (Gad, = 462 492,
/> = < 0 0001) in diet composhion of Mexican wolves when compared to diets reported
previously for other North American gray wolves Mexican wolf diet analysis revealed
83

more large-sized food hems (90 6% PFO) than that reported in 7 northern gray wolf diet
smdies (range 58 5% to 82 90/0 PFO) Our results imply that free-ranging Mexican gray
wolves in .\nzona and New Mexico consumed more large-sized food hems, primarily elk
aduhs and calves, than sympatric coyotes and the larger, northern gray wolves. This
information could prove useftil in managing both Mexican wolf and prey populations, as
well as determining the presence of Mexican wolves in depredation incidences in Arizona
and New Mexico
84

PERMISSION TO COPY
In presenting this diesis m partial fiilfilhnent of tiie requirements for a master's
degree at Texas Tech University or Texas Tech University Health Sciences Center, I
agree that the Library and my major department shall make it freely available for
research purposes. Permission to copy this thesis for scholarly purposes may be
granted by the Director of the Library or my major professor. It is understood that any
copying or publication of this thesis for financial gain shall not be allowed without my
fiuther written permission and that any user may be liable for copyright infringement.
A|gree (Permission is granted.)
^tudem Signature Date
Disagree (Permission is not granted.)
Student Signature Date