developmental and transcriptional responses to host and ... · cuticles by the specific locust...
TRANSCRIPT

EUKARYOTIC CELL, May 2005, p. 937–947 Vol. 4, No. 51535-9778/05/$08.00�0 doi:10.1128/EC.4.5.937–947.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Developmental and Transcriptional Responses to Host and NonhostCuticles by the Specific Locust Pathogen Metarhizium
anisopliae var. acridum†Chengshu Wang and Raymond J. St. Leger*
Department of Entomology, University of Maryland, College Park, Maryland
Received 23 January 2005/Accepted 28 February 2005
Transcript patterns elicited in response to hosts can reveal how fungi recognize suitable hosts and themechanisms involved in pathogenicity. These patterns could be fashioned by recognition of host-specifictopographical features or by chemical components displayed or released by the host. We investigated this inthe specific locust pathogen Metarhizium anisopliae var. acridum. Only host (Schistocerca gregaria) cuticlestimulated the full developmental program of germination and differentiation of infection structures (appres-soria). Cuticle from beetles (Leptinotarsa decimlineata) repressed germination while cuticle from hemipteranbugs (Magicicada septendecim) allowed germination but only very low levels of differentiation, indicating thatthe ability to cause disease can be blocked at different stages. Using organic solvents to extract insects weidentified a polar fraction from locusts that allowed appressorial formation against a flat plastic (hydrophobic)surface. Microarrays comprising 1,730 expressed sequence tags were used to determine if this extract elicitsdifferent transcriptional programs than whole locust cuticle or nonhost extracts. Of 483 differentially regulatedgenes, 97% were upregulated. These included genes involved in metabolism, utilization of host cuticle compo-nents, cell survival and detoxification, and signal transduction. Surprisingly, given the complex nature of insectepicuticle components and the specific response of M. anisopliae var. acridum to locusts, very similar transcriptprofiles were observed on locust and beetle extracts. However, the beetle extract cluster was enriched in genesfor detoxification and redox processes, while the locust extract upregulated more genes for cell division andaccumulation of cell mass. In addition, several signal transduction genes previously implicated in pathoge-nicity in plant pathogens were only upregulated in response to locust extract, implying similarities in theregulatory circuitry of these pathogens with very different hosts.
Many fungal insect pathogens, such as Metarhizium aniso-pliae, infect host cuticles via spores that adhere and germinateto form a series of infection structures during penetration. Inthe presence of nutrients and water, conidia of M. anisopliaeform germ tubes. The germ tube continues undifferentiatedhyphal growth on a soft surface or if nutrient quality andquantity is not conducive to differentiation. On a host cuticle,however, apical elongation terminates and the germ tubeswells distally to form an appressorium, a major site of adhe-sion and for production of enzymes that help breach hostcuticle and establish a nutritional relationship with the host(33, 38). The formation of appressoria by strains of M. aniso-pliae var. anisopliae with a broad host range can also be in-duced efficiently by a hard hydrophobic surface (i.e., polysty-rene) in the presence of low levels of complex nitrogenousnutrients. However, pathogens with a narrow host range suchas M. anisopliae var. acridum (specific for acridids) germinatepoorly under these conditions and do not produce infectionstructures, suggesting they may be adapted to conditions per-taining to their hosts (36). The mechanisms for recognizinghost-related triggers and the processes responsible for appres-sorium initiation are likely involved in the expression of viru-
lence factors and parasitic specialization. The nature of theinductive triggers has not been determined, but while the pro-tein and chitin composition of insect procuticle appears similarin all insects, the epicuticular components are extremely het-erogeneous, even within the same genus, and therefore havethe potential to lead to different pathogen responses to partic-ular insects (7, 34).
A particular reason for focusing on M. anisopliae var. acri-dum is that it is being developed as a promising biologicalcontrol agent of desert locusts, Schistocerca gregaria, and otherorthopteran pests (30). These applied interests would obvi-ously benefit from an understanding of the molecular andbiochemical interactions between the pathogen and its hosts,particularly if genetic modifications are to be designed to en-hance pathogen attributes (30). In this study we investigatedthe conditions required for M. anisopliae var. acridum strainARSEF 324 to produce infection structures and developed amodel system that allowed control of environmental conditionsand inductive stimuli independent of host behavior. Havingstudied the biological significance of cuticular components fortriggering appressoria, we employed microarrays to initiate adetailed investigation of the molecular mechanisms involved.
MATERIALS AND METHODS
Organism and growth. M. anisopliae var. acridum ARSEF 324 originally iso-lated from Austracris guttulosa in Australia was obtained from the U.S. Depart-ment of Agriculture Entompathogenic Fungus Collection in Ithaca, N.Y. Cul-tures were maintained on potato dextrose agar.
Behavior of M. anisopliae var. acridum ARSEF 324 germinating on insect
* Corresponding author. Mailing address: 4112 Plant Science Build-ing, University of Maryland, College Park, MD 20742-4454. Phone:(301) 405-5402. Fax: (301) 314-9290. E-mail: [email protected].
† Supplemental material for this article may be found at http://ec.asm.org/.
937
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

wings. The membranous hind wings of locusts (S. gregaria), beetles (Leptinotarsadecimlineata), and 17-year periodical cicadas (Magicicada septendecim) weredissected away, surface sterilized in 5% sodium hypochlorite (5 min), and rinsedwith 4 changes (5 min each) of sterile distilled water. Wings were placed on wateragar (1.5%, wt/vol) plates, dried in a sterile-airflow cabinet and inoculated with200 �l distilled water containing about 10,000 conidia. Following incubation (upto 24 h) at 27.5°C, cuticles were examined microscopically.
Extraction of cuticular surface lipids. Groups of 5 or more adult locusts (S.gregaria), cicadas (M. septendecim), or an equivalent weight of beetles (L. decem-lineata) were killed by freezing, freeze-dried, and immersed in three- to fourfoldexcess of organic solvents (dichloromethane, hexane or methanol) and gentlyshaken for 10 min to extract cuticular lipids (17). All types of lipid tend todissolve in dichloromethane, which is of intermediate polarity. As it was desir-able to test the nonpolar lipids separately from the polar ones, hexane (a non-polar solvent) and methanol (a polar solvent) were also used to extract locusts.Hexane removes hydrocarbons, glycerides, sterol esters and other nonpolar lipidsfrom cuticles virtually free of nonlipid contaminants (16). Methanol does notextract these but extracts acylglycerols, free fatty acids, and phospholipids. How-ever, polar solvents may also extract amino acids, peptides, and carbohydrates(16). The extracts were collected in glass petri dishes, dried in a hood, andsuspended in absolute ethanol. Samples of the ethanol suspension were evapo-rated on preweighed aluminum dishes to determine the dry weight of extract andthe balance was stored at �20°C. To obtain the water-soluble fraction of theepicuticle, a methanol extract from locust was evaporated and the water-solublecomponents taken up with a small volume of water.
Behavior of M. anisopliae var. acridum ARSEF 324 germinating against plas-tic. To test the chemical requirements for strain differentiation against a smoothhydrophobic surface, conidia were induced to germinate by growing them in 5.5-cmpolystyrene petri dishes containing 2 ml of water supplemented with 0.0125% yeastextract (YE) and/or insect cuticle extracts. Various amounts of each extract werepipetted onto glass coverslips and evaporated, leaving a white greasy layer on oneside. The coverslips were then placed in the polystyrene petri dishes with the lipidlayer facing up. One hundred spores from each of four replicates were scoredmicroscopically to assess germination and differentiation frequency. The adherenceof conidia or germlings to polystyrene surfaces was determined by applying a jet ofwater from a pipette and counting the spores retained. The kinase inhibitors H89and PD-98059 (purchased from Biomol Research Laboratories) were added tocultures from stock solutions in dimethyl sulfoxide.
RNA extraction and cDNA microarray analysis. Mycelia from 36-h Sabourauddextrose broth (SDB) cultures were harvested and washed with sterile distilledwater. The mycelium was transferred to minimal medium (MM, containing 0.1%wt/vol KH2PO4; 0.05% wt/vol MgSO4 and 50% vol/vol tap water) or MM con-taining intact locust cuticle (10 mg/ml), dichloromethane extracts from locusts(locust extract), beetles (L. decimlineata) (beetle extract), or cicadas (cicadaextract), or the water-soluble fraction of methanolic extracts from locusts (i.e.,locust polar lipid) (all at 2 mg/ml). Cultures were shaken (100 rpm) for 6 h at27°C. Total RNA was extracted from mycelium using an RNeasy Plant minikitplus the treatment with DNase I (QIAGEN).
The RNA from SDB cultures was used as the reference sample for microarrayanalysis. Hybridizations were conducted using slides printed with 1,730 cDNAclones from M. anisopliae var. anisopliae ARSEF 2575 and a few genes from M.anisopliae var. acridum absent from the library of M. anisopliae var. anisopliae(15, 40). The M. anisopliae var. acridum genes sequenced to date are less than 5%divergent from their M. anisopliae var. anisopliae counterparts, even though theseare mostly secreted products encoding ecological traits that would be expected todiverge more than housekeeping genes (1, 14) so we did not envision anyproblems with genes from this strain being able to hybridize to the arrays.
Microarray data were analyzed as described before (40) with a TIGR TM4system (http://www.tigr.org/software/tm4/). Briefly, data normalization was per-formed based on the assumption that all spots within each block have the samestandard deviation for Log 2 ratios (29). The geometric mean of in-slide repli-cates was calculated with the MIDAS implemented process function after as-signing a unique identification number for each clone. One-class significanceanalysis of microarray (SAM) test (39) was conducted to identify the genes whoseexpressions were differentially regulated. For this analysis, the exchangeabilityfactor s0 was set at the fifth percentile with �, 1.0. Hybridizations were conductedin triplicate with RNA from different experiments.
RESULTS
Behavior of M. anisopliae var. acridum ARSEF 324 germi-nating on insect cuticles. At 24 h, 73% of conidia had germi-
nated on locust wings but production of appressoria waspatchy. In some regions almost 100% of germlings had termi-nal swellings at the end of short germ tubes (Fig. 1A), whileover other regions of the cuticle there was extensive hyphalgrowth with appressorial production usually being preceded bypronounced curling of germ tubes (Fig. 1B and C). Long hy-phae often grew together forming orderly aggregations withthe same orientation (Fig. 1C) but we saw no evidence thathyphae in close proximity attract each other. Hyphae oftencrossed over other hyphae that were at an angle to their di-rection of growth (Fig. 1D) but anastomosis was not observed.
M. anisopliae var. acridum, unlike M. anisopliae var. aniso-pliae (35), was never observed forming compound appressoria,i.e., multicellular appressoria comprising cells from multiplegermlings. Thus, even when hyphae were close together andwith the same directional growth each hyphae terminated in asimple appressorium (Fig. 1E). Subterminal appressoria (lat-eral to germ tubes) developed from 16 h. Appressorium pro-duction was not related to any discernible physical feature.Thus, hyphae were frequently deflected by hair sockets andother surface structures and changed direction of growth butthey did not produce appressoria against hair sockets as ob-served for M. anisopliae var. anisopliae (35).
At 24 h, 54% of conidia had germinated on cicada wings,often with extensive hyphal growth (Fig. 1F). Appressoria wererarely observed. In 45 fields of view containing several thou-sand germlings we observed only 36. Interestingly, these almostalways occurred in groups (Fig. 1G), suggesting that smallregions of this nonhost cuticle possessed conditions conduciveto differentiation. Only 4% of the conidia had germinated onbeetle cuticle by 24 h and we did not observe appressoria.
Behavior of M. anisopliae var. acridum germinating againstplastic. M. anisopliae var. acridum did not adhere or germinatein water alone, but yeast extract was sufficient to stimulate bothadhesion and germination of M. anisopliae var. acridum spores,although not differentiation of appressoria (Table 1). Sincemost analyses of locust S. gregaria lipids have used dichlo-romethane extracts (17), dichloromethane extracts from locust,cicada and beetle were tested for their effects on M. anisopliaevar. acridum with and without YE (Table 1). Extract frombeetle cuticle stimulated adhesion compared to water alonebut not germination. In fact, combined with YE, the beetleextract repressed germination compared to YE alone, suggest-ing the existence of an antifungal component in the beetlecuticle (Table 1). This inhibitory effect was not observed withM. anisopliae var. anisopliae strain ARSEF 2575 (unpublisheddata), indicating greater physiological plasticity by this broad-host-range strain.
The dichloromethane extracts from locusts and cicadas al-lowed germination without addition of YE, and when com-bined with YE both extracts stimulated higher levels (P � 0.01)of germination compared to YE alone. Unlike cicada extract,locust extract stimulated differentiation of small appressoriamorphologically very similar to appressoria formed on locustwings (Fig. 2A). Cicada extract combined with YE also in-duced formation of appressoria, but 8-fold fewer than whenlocust extract was combined with YE. However, whether grow-ing on locust extract or cicada extract, during the first 24 happressoria only formed on the petri dish surface overlaid bythe coverslip (some appressoria had formed outside the cov-
938 WANG AND ST. LEGER EUKARYOT. CELL
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

FIG
.1.
Micrographs
ofM
.anisopliaevar.acridum
germinating
onlocust(S.gregaria)
wings
(Ato
E)
orcicada
(M.septendecim
)w
ings(F
andG
)for
24h.
CO
,conidium;A
P,appressorium.B
ar,10�
m.
VOL. 4, 2005 TRANSCRIPTIONAL RESPONSES OF M. ANISOPLIAE 939
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

erslip by 48 h). The germ tubes and appressoria remainedattached to the hydrophobic petri dish even after removing thecoverslip.
The nonpolar hexane extract from locusts induced very lowlevels of germination by 24 h (Table 2). However, when sup-plemented with YE, germination levels were similar to that inYE alone, indicating that it was not repressing germination.The hexane extract with or without YE did not induce produc-tion of appressoria, even when conidia were germinating di-rectly on the lipid layer of the coverslip. In contrast, the meth-anol fraction allowed germination without addition of YE andstimulated differentiation of small appressoria on the area ofthe petri dish overlaid by the coverslip (Fig. 2B). Supplement-ing the methanol extract with YE greatly increased both thesize of appressoria and the frequency of lobate appressoriaformed by short, swollen side branches from the original ap-pressorial cell (Fig. 2C). This suggests that YE does not con-tain inducers of appressorial differentiation but provides nu-tritional resources allowing the maturation of larger cells.
Since the inducer(s) appears to be confined to the polarfraction, we wished to determine whether the inducer was apolar lipid or a nonlipid contaminant. The methanol solventwas removed by evaporation and water-soluble componentswere taken up in a small volume of water. This water-solublefraction retained the ability to stimulate high levels of adhesionand germination (Table 2). However, the germlings showedunusual development. In particular, branching of germlingswas more numerous and compact than observed with methanolor dichloromethane extracts (Fig. 2D and E) or on locustcuticle and fewer appressoria were produced under coverslips.At high concentrations (4 mg/ml) the water-soluble fractionstimulated extensive hyphal growth under coverslips with evenfewer appressoria (Fig. 2F). In 3 out of 5 replicates we alsoobserved extensive directional growth by germ tubes towardsthe coverslip (Fig. 2G).
Effect of cAMP on appressoria formation. Because cyclicAMP (cAMP) is sufficient to induce appressorial formation inmany specific plant pathogens (2), we tested the effect of ex-ternal cAMP application on conidial development of M. aniso-
pliae var. acridum in YE and YE plus dichloromethane ex-tracts. Cyclic AMP added at 5, 10, or 20 mM produced nochange in germination frequencies and cAMP did not induceappressorial formation under conditions in which they do notnormally form, such as in YE culture, on a hydrophilic glasssurface, or not overlaid by a coverslip. Cyclic AMP exerts itseffects principally through the activation of cAMP-dependentprotein kinase A (PKA) (2). In spite of exogenous cAMPhaving no apparent effect, both germination and differentiationwere inhibited by the PKA inhibitor H89, suggesting thatcAMP/PKA is one of multiple pathways required to triggerdifferentiation. Consistent with this a highly selective mitogen-activated protein kinase kinase inhibitor PD-98059 was alsovery potent at inhibiting either germination or differentiation(Table 2).
Microarray analysis reveals distinctive gene transcriptionresponses to locust cuticle, to lipid extracts and to minimalmedium. Mycelium grown in shake cultures to prevent contactstimuli caused by settling was used as the source of RNA toprobe slides with 1,730 printed genes (40). These microarrayswere used for direct comparisons of gene expression levels asM. anisopliae var. acridum responded to nutrient deprivation(MM), intact locust cuticle, dichloromethane extracts fromlocusts, beetles, or cicadas, and the water-soluble fraction of amethanol extract from locusts (locust polar lipid, locust polarlipid) (Fig. 3).
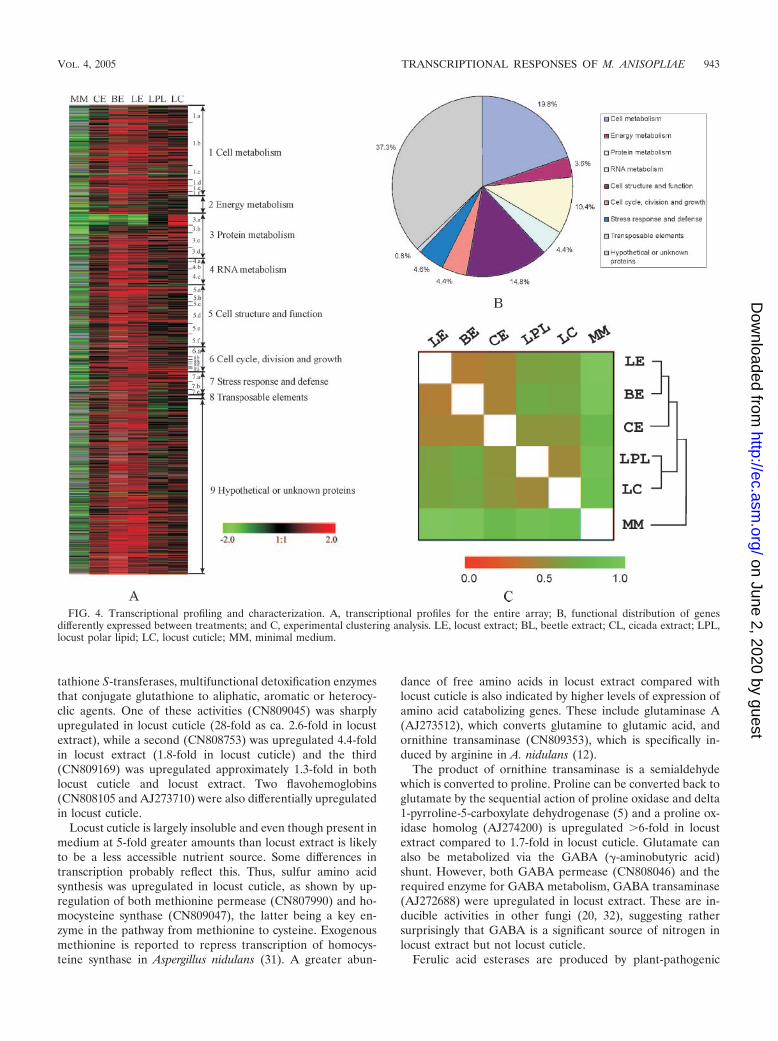
A SAM test identified 483 genes (28% of those arrayed) thatwere differentially up (471 genes) or down (12 genes) regulatedin one or more of the host related medium 6 h after transfer ofmycelium from nutrient rich SDB. In contrast, most geneswere downregulated during nutrient deprivation (Fig. 3B; Fig.4A). The printed genes were classified based on their potentialcellular function into 9 functional groups (40), as presented inFig. 4A. Viewed broadly, of the 483 differentially regulatedgenes, 19.8% were involved in cell metabolism, 14.8% in cellstructure and 10.4% in protein metabolism. Other significantlyvaried genes were evenly dispersed among the 6 remainingfunctional groups. In addition, 37.3% of genes with hypothet-ical or unknown function also showed differential expressionbetween experiments, sometimes being upregulated verysharply in one or other host related medium (Fig. 4A and B).
Based on the Euclidean distances estimated from the tran-scriptional profiles in different medium, MM (nutrient depri-vation) is divergent from the host-related medium while theseclustered into two distinct groups: expression patterns in thethree dichloromethane extracts (locust extract, beetle extractand cicada extract) cluster together, while locust polar lipidclusters with locust cuticle (Fig. 4C).
The large-scale upregulation of gene expression produced bycuticular components implies regulation by several modula-tors, consistent with which insect cuticle components activateda series of genes involved in signal transduction and transcrip-tional regulation. Of particular interest, putative signal trans-duction genes previously implicated in pathogenicity in M.anisopliae and/or other fungi (37) were either upregulated inall host-related media, all three dicloromethane extracts, orjust in locust extract (Fig. 5A). The exceptions were CN807989,which is similar to a pathogenicity transmembrane proteinfrom Magnaporphe grisea that is upregulated in all media, in-cluding MM, and adenylate cyclase (AJ251971), which showed
TABLE 1. Effects of water, yeast extract, and different insectdichloromethane extracts on spore adhesion (A), germination (G),
and appressorium differentiation (D)
Treatmenta
% Change
15 h 24 h
A G D A G D
Water 0 0 0 0 0 0YE 95 36 0 100 82 0Beetle extract 21 0 0 27 �1 0Beetle extract � YE 100 3 0 100 57 0Cicada extract 37 11 0 50 48 0Cicada extract � YE 100 73 0 100 �99 19Locust extract 42 23 1 98 78 7Locust extract � YE 100 83 3 100 �99 63
a Ethanol containing 1 mg of beetle, locust, or cicada dichloromethane extractwas evaporated on coverslips that were placed in 5.5-cm petri dishes containing2 ml of water or 0.0125% yeast extract (YE) and 10,000 conidia of M. ansiopliaevar. acridum ARSEF 324. Percent germination and differentiation frequencieswere determined from counts on the petri dish surface overlaid by the coverslip.Adhesion was determined on separate petri dishes not containing coverslips.
940 WANG AND ST. LEGER EUKARYOT. CELL
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

minimal changes in regulation. Otherwise, PKA (AJ273657)and CN809265, similar to a virulence factor in Gibberella moni-formis that functions as a two component signaling responseregulator for transmitting extracellular signals (6), were up-regulated in all media except MM, while mitogen-activatedprotein kinase kinase (AJ272796) was upregulated (�2-fold)in all three dicloromethane extracts but not in locust polarlipid, locust cuticle, or MM.
Activation of adenylate cyclase is mediated by heterotrimic
GTP-binding proteins (G�, G�, and G). A G protein � sub-unit was only upregulated in locust extract (ca. 6-fold), whilethe � subunit, also involved in regulating infection-relatedmorphogenesis in Magnaporphe grisea (26), was downregulated�4-fold in the three dichloromethane extracts and MM, butwas upregulated in locust cuticle and locust polar lipid by 3.3-and 1.6-fold, respectively. Most significantly, several homologsfor regulators of infection structure formation in other fungiwere only upregulated in locust extract. This resulted in signal
FIG. 2. Micrographs of M. anisopliae var. acridum germinated on polystyrene for 24 h with insect extracts. Germlings grown with dichlo-romethane (A) or methanol (B) extracts from locusts showing terminal hyphal swellings. C, production of lobate appressoria when the methanolextract was combined with 0.0125% yeast extract. D and E, branching of germlings in the water-soluble fraction (1 mg/ml) of the methanol extract;F, extensive hyphal growth at 4 mg/ml that may demonstrate directional growth towards the coverslip (G). Panels A to F show growth on the areaof the petri dish overlaid by the coverslip. CO, conidium; AP, appressorium; CS, coverslip. Bar, 10 �m.
VOL. 4, 2005 TRANSCRIPTIONAL RESPONSES OF M. ANISOPLIAE 941
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

transduction components upregulated in beetle extract andcicada extract clustering separately from locust extract (Fig.5A). Thus, CN808063, which is similar to MAS1 protein, re-quired for appressorial formation in M. grisea (19), is upregu-lated 2.7-fold in locust extract but downregulated 4.4-fold inbeetle extract and 1.6-fold in cicada extract. Likewise,CN808339 that is similar to a CAP20-like protein involved invirulence, penetration, and appressorium formation in Blume-
ria graminis (18) was upregulated in locust extract and down-regulated in other media.
The downstream target genes of many signaling pathwaysare still poorly understood. However, the transcriptome inbeetle extract possessed some unique features as shown in Fig.5b, which lists genes preferentially responding to locust extractbut not to beetle extract and vice versa. The beetle extractcluster does not represent a defined functional category but isenriched in genes associated with detoxification and redoxprocesses, including the antioxidant defense protein thiore-doxin AJ273832 (upregulated 2.3-fold in beetle extract butdownregulated in locust extract), aldehyde dehydrogenaseCN809673 (upregulated 6.5-fold), and formate dehydrogenaseCN809022 (upregulated 20-fold). It also contains genes forreallocation of resources such as a sugar transport protein(AJ273888), a sterol carrier protein (CN809167), and bothhigh- and low-affinity copper transporters (AJ273995 andCN808791) that were differentially upregulated 25- and 40-fold, respectively. Besides signal transduction genes, those dif-ferentially upregulated in locust extract included some for cellwall synthesis (e.g., CN808518) and cell cycle regulation (e.g.,AJ274064), implying more rapid cell division and accumulationof cell mass (Fig. 5B).
We also identified clusters of genes specifically induced byeither growth on intact locust cuticle or its dichloromethaneextract (Fig. 5C). Genes differentially upregulated duringgrowth on intact locust cuticle included the M. anisopliae ho-molog of protein disulfide isomerase (CN808269) along withcalnexin (AJ273132) and several chaperones in the heat shockclass (AJ272997, AJ273036, AJ273534, AJ273662, andCN809938).
Some of the enzymes upregulated in locust cuticle have toxicsubstrates known to be present in procuticle. Thus, a putativedioxygenase (AJ272846) with homologs that cleave aromatichydrocarbon components of phenolics was upregulated. Likeother fungi, M. anisopliae var. acridum contains multiple glu-
FIG. 3. Raw intensity (RI) graph of overall gene expression profiles from microarray analysis of locust-related extracts (A) and the others (B).I(A) and I(B) represent the intensity data from channel A (Cy3) and channel B (Cy5), respectively. LE, locust dichloromethane extracts; LC, locustcuticle; LPL, locust polar lipid; BE, beetle extract; CE, cicada extracts; MM, minimal medium.
TABLE 2. Effects of locust organic extracts on spore adhesion (A),germination (G), and appressorium differentiation (D)
Treatmenta
% Change
15 h 24 h
A G D A G D
YE 95 39 0 100 79 0Dichloromethane 40 28 3 97 81 9Dichloromethane � YE 100 87 9 100 �99 66Dichloromethane � YE
� H89 (5 �M)84 61 0 100 87 0
Dichloromethane � YE� PD98059 (10 nM)
80 22 0 95 65 0
Hexane 0 0 0 50 10 0Hexane � YE 95 55 0 100 80 0Methanol 100 91 3 98 �99 47Methanol � YE 100 93 53 100 �99 63LPL (1 mg/ml) 95 67 0 100 �99 13LPL (2 mg/ml) 100 94 4 100 �99 17LPL (4 mg/ml) 100 95 0 100 �99 0LPL (1 mg/ml) � YE 95 85 3 100 �99 8
a Ethanol containing 1 mg of locust dichloromethane, hexane, or methanolextract was evaporated on coverslips that were placed in 5.5-cm petri dishescontaining 2 ml of water or 0.0125% yeast extract (YE) and 10,000 conidia of M.anisopliae var. acridum ARSEF 324. The effects of YE alone and the water-soluble fraction of the methanol extract (locust polar lipid [LPL]) were testedusing ethanol-washed cover slips. Percent germination and differentiation fre-quencies were determined from counts on the petri dish surface overlaid by thecoverslip. Adhesion was determined on separate petri dishes not containingcoverslips.
942 WANG AND ST. LEGER EUKARYOT. CELL
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from
tathione S-transferases, multifunctional detoxification enzymesthat conjugate glutathione to aliphatic, aromatic or heterocy-clic agents. One of these activities (CN809045) was sharplyupregulated in locust cuticle (28-fold as ca. 2.6-fold in locustextract), while a second (CN808753) was upregulated 4.4-foldin locust extract (1.8-fold in locust cuticle) and the third(CN809169) was upregulated approximately 1.3-fold in bothlocust cuticle and locust extract. Two flavohemoglobins(CN808105 and AJ273710) were also differentially upregulatedin locust cuticle.
Locust cuticle is largely insoluble and even though present inmedium at 5-fold greater amounts than locust extract is likelyto be a less accessible nutrient source. Some differences intranscription probably reflect this. Thus, sulfur amino acidsynthesis was upregulated in locust cuticle, as shown by up-regulation of both methionine permease (CN807990) and ho-mocysteine synthase (CN809047), the latter being a key en-zyme in the pathway from methionine to cysteine. Exogenousmethionine is reported to repress transcription of homocys-teine synthase in Aspergillus nidulans (31). A greater abun-
dance of free amino acids in locust extract compared withlocust cuticle is also indicated by higher levels of expression ofamino acid catabolizing genes. These include glutaminase A(AJ273512), which converts glutamine to glutamic acid, andornithine transaminase (CN809353), which is specifically in-duced by arginine in A. nidulans (12).
The product of ornithine transaminase is a semialdehydewhich is converted to proline. Proline can be converted back toglutamate by the sequential action of proline oxidase and delta1-pyrroline-5-carboxylate dehydrogenase (5) and a proline ox-idase homolog (AJ274200) is upregulated �6-fold in locustextract compared to 1.7-fold in locust cuticle. Glutamate canalso be metabolized via the GABA (-aminobutyric acid)shunt. However, both GABA permease (CN808046) and therequired enzyme for GABA metabolism, GABA transaminase(AJ272688) were upregulated in locust extract. These are in-ducible activities in other fungi (20, 32), suggesting rathersurprisingly that GABA is a significant source of nitrogen inlocust extract but not locust cuticle.
Ferulic acid esterases are produced by plant-pathogenic
FIG. 4. Transcriptional profiling and characterization. A, transcriptional profiles for the entire array; B, functional distribution of genesdifferently expressed between treatments; and C, experimental clustering analysis. LE, locust extract; BL, beetle extract; CL, cicada extract; LPL,locust polar lipid; LC, locust cuticle; MM, minimal medium.
VOL. 4, 2005 TRANSCRIPTIONAL RESPONSES OF M. ANISOPLIAE 943
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

fungi and are involved in the degradation of pectin and xylan,from which they release aromatic acids on cereal-derived sub-strates. The expression of a putative homolog (AJ273114) inlocust extract implies the presence of a hydrophobic substratewith bulky substituents of the benzene ring (9). The ability ofstrain 324 to use at least some lipoidal components of thedichloromethane extract was suggested by upregulation of sev-eral classes of lipases. Most of these were similarly regulated inlocust extract and locust cuticle. The exceptions included alipase (AJ274323) and a nitroalkane dioxygenase (CN809138)that converts nitroalkanes to the corresponding aldehyde andnitrite. They are �5-fold upregulated in locust extract. Nitrite
reductase (CN808073) is also sharply upregulated in locustextract (Fig. 5C).
DISCUSSION
One of the outstanding features of M. anisopliae is its abilityto parasitize a wide range of different insect species. Manyworkers have characterized the host range of different isolatesin attempts to characterize strains on the basis of pathogenicity(28). Most of the information on why isolates attack somehosts and not others is concerned with the effects of differenthost components on adhesion and germination (35). This focus
FIG. 5. Transcriptional profiling and characterization. Differentially expressed genes involved in signaling pathways (A; *, indicates whosegenes transcriptional expression varied significantly between the treatments); genes differentially expressed between locust extract and beetleextract (B) and between locust extract and locust cuticle (C). The blocks in gray show the intensity values (Cy3 and Cy5) have been rescaled aszero after background filtering and normalization.
944 WANG AND ST. LEGER EUKARYOT. CELL
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

was reasonable given that these are the earliest stages of in-fection. Consistent with it, beetle cuticle and extracts repressgermination of M. anisopliae var. acridum, although allowinglow levels of adhesion. However, there are many stages in theprocess of infection where the ability of an isolate to causedisease may be influenced, and so a widening of our knowledgein this area is needed.
The inability of M. anisopliae var. acridum to infect cicadas isapparently determined at the stage of appressorial formation.M. anisopliae var. acridum adheres and germinates on cicadacuticle but forms few small appressoria suggesting differencesin stimulation on the locust and cicada cuticles. The patchydistribution of infection structures produced by M. anisopliaevar. acridum on cicada cuticles also suggests that the fungusrecognizes specific signals from the structures or chemistry ofthe cuticle surface that vary with site, and that it behavesdifferently as a reaction to these signals. These stimuli arelikely to be chemical rather than physical as the ability of anextract from locust cuticles to stimulate abundant appressorialformation against a flat surface excludes an important role fortopographical features on the locust cuticle in initiating ap-pressoria formation.
Spores of most plant pathogens will germinate in wateralone. Like many other entomopathogenic fungi (4), M. aniso-pliae var. acridum apparently requires water-soluble nutrientsfrom the host surface to germinate and develop. The extent towhich parasitic specificity of entomopathogenic fungi is deter-mined by cuticular lipids is uncertain, with apparent contradic-tions in the literature. In part this probably reflects lack ofprecision in the chemical nomenclature or incomplete meth-odology description (21) as well as strain differences. Strainswith multiple hosts are likely to respond to multiple nonspe-cific signals. It is highly improbable that diverse insects secretea single universal signaling chemical to which all fungi respond.Individual strains may sense one signal or a set of specificsignals within a complex cocktail of insect chemicals and wouldrespond according to what is secreted by any given host insect.
Evidence for a mixture of signals in the locust extract wasprovided by the response of M. anisopliae var. acridum todifferent fractions. Total extraction with dichloromethane wasvery effective at inducing infection structure formation undercoverslips but M. anisopliae var. acridum was largely unrespon-sive to the nonpolar lipid fraction that comprises the bulk ofthe locust epicuticle (17). The polar components extracted withmethanol elicited a response similar to that obtained withdichloromethane but its water-soluble fraction (locust polarlipid) produced not only appressoria but a bushy type of hyphalbranching pattern. At high levels (4 mg/ml) locust polar lipidproduced extensive hyphal growth at the expense of appresso-rial production. This is consistent with observations on M.anisopliae var. anisopliae (33) that suggest that a primary roleof the appressoria is to establish a nutritional relationship withthe host and is not necessary in the presence of a sufficiency ofnutrients. The directional growth that occurred towards thecoverslip was reminiscent of directional growth shown on lo-cust cuticle except that appressoria were not produced. Direc-tional growth could be adaptive if it increased the probabilitythat hyphae will contact other host related signals.
In spite of different, presumably selectable responses to in-sects, the ultimate physiological and morphological responses
of most M. anisopliae strains in terms of appressorial formationare similar. However, several differences were noted betweenthe behavior of M. anisopliae var. acridum and M. anisopliaevar. anisopliae on cuticle surfaces (33). Failure of M. anisopliaevar. acridum to produce compound appressoria may not be ofgreat functional significance as the simple M. anisopliae var.acridum appressoria have developed at a similar time they willprobably also penetrate at a similar time, resulting in an effec-tive multiple invasion. However, they do imply genetic differ-ences that are employed during penetration events. Potentiallyrelated to our observation that M. anisopliae var. acridum doesnot anastomatize and form compound appressoria, CN808914related (E, 10�101) to Het-C (heterokaryon incompatibility)from Neurospora crassa was sharply upregulated (�5-fold) indichloromethane extracts. Podospora anerina uses common sig-nal transduction pathways in regulating development and veg-etative incompatibility, including the G protein � subunit (23,24). Thus, potential relationships can be anticipated betweenfungal appressorium differentiation and vegetative incompati-bility controls.
Aside from the G protein � subunit, all the signal transduc-tion genes previously implicated in pathogenicity in plantpathogens that we had homologs for in the M. anisopliae var.anisopliae arrays were upregulated in locust extract. Thus, al-though the signals that induce germination and differentiationare different, similar signal transduction pathways may mediatethese signals in plant and insect pathogens. Fluctuations in therelative levels of both Ca2� and cAMP effect changes in thecytoskeleton and cell wall of M. anisopliae var. anisopliae dur-ing formation of appressoria (34, 37). However, while cAMP isalso sufficient to induce appressorium formation in many spe-cific plant pathogens (2) this was not the case for M. anisopliaevar. acridum. Evidence for commonalities was provided by theinhibitor studies with H89 and PD-98059 as PKA and mitogen-activated protein kinases have been implicated in developmen-tal processes in many other fungal pathogens (10).
The mitogen-activated protein kinase cascade is a highlyconserved module that mediates new gene expression in re-sponse to external signals in a series of developmental pro-cesses in fungi including germination, appressorium formation,and pathogenicity (41). Branching and the curvilinear mode ofgrowth that often preceded differentiation both in vivo and invitro imply an effect on the Spitzenkorfer (the vesicle-gener-ating apparatus at the hyphal tip) that by its presence andposition orients cell wall component disposition of the hyphalapex and controls the direction of growth (3). Previously, wesuggested that the transition from polarized hyphal growth togerm tube expansion (appressoria) might be linked to a re-duced displacement rate or dispersal of the Spitzenkorfer, re-directing cell wall synthesis from the apical tip to the entire cellsurface (34). Vesicles are transported along microtubules andphosphorylation of mitogen-activated protein kinase is in-volved in regulating the organization of microtubules (27). TheCa2�-dependent phosphorylation of three Metarhizium pro-teins recognized by antiserum to mitogen-activated proteinkinase (34) suggests a possible mechanism for both Ca2�- andmitogen-activated protein kinase on differentiation.
Our microarray data indicate that induction of transcriptionis an overriding feature of the response of M. anisopliae var.acridum to insect-related medium with relatively little tran-
VOL. 4, 2005 TRANSCRIPTIONAL RESPONSES OF M. ANISOPLIAE 945
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

scriptional repression observed. Before this study we had sus-pected that host recognition is determined by regulatory con-trols that allow expression of pathogenicity genes that are notexpressed on nonhosts. The similar transcriptional responsesof M. anisopliae var. acridum grown on inductive (for appres-soria) locust extract and noninductive beetle extract is evidencethat specialization to locust hosts has not eliminated a substan-tial stereotypical response to epicuticular components. As bee-tle extract represses germination and growth of M. anisopliaevar. acridum, it contains components that inhibit developmentand the differentially regulated transcripts equivalently ex-pressed in both locust extract and beetle extract cannot besufficient to overcome these inhibitors. It is possible that genesspecifically upregulated by locust extract are essential for de-velopment.
Based on the overall transcriptional profiles for the totalarray of 1,730 cDNAs, locust extract and beetle extract clustermore closely with each other than with cicada extract (Fig. 4C)but locust extract forms an outgroup when cluster analysis islimited to coregulated genes involved in signal transduction(Fig. 5A). This correlates with differences between locust ex-tract and beetle extract or cicada extract in their ability toinduce appressorial formation, consistent with perception andregulatory factors in M. anisopliae var. acridum controllinginfection related differentiation. Among these regulatory ele-ments, some of the biggest expression differences were ob-served for a Mas1-like protein (CN808063) and a CAP20-likeprotein (CN808339) that were both upregulated in locust ex-tract while being downregulated in beetle extract and cicadaextract (Fig. 4A). Deletion of their homologs in Magnaporthegrisea and Colletotrichum gloeosporiodies, respectively, allowsappressorial formation but greatly reduces penetration andvirulence (18, 42), and they may have similar effects in otherplant pathogens (19).
Other differences between the transcriptional profiles in-duced by locust extract and beetle extract occurred principallyamong genes involved in detoxification, oxidant defense andtransport. These differential shifts in gene expression occurredin the context of equivalent expression of other genes in thesame functional categories in both locust extract and beetleextract. Thus, many genes involved in detoxification, oxidantdefense and transport were comparably induced by incubatingin locust extract and in beetle extract suggesting that bothextracts contain a cocktail of inductive and repressive compo-nents. The overall favorable chemistry of the locust extract wasshown by differential upregulation of genes for cell wall syn-thesis (CN808518) and cell cycle regulation (AJ274064) imply-ing more rapid cell division and accumulation of cell mass.
The locust polar lipid and intact cuticle (locust cuticle) in-duced a similar transcriptional response that may reflect sim-ilarities in composition between the water-soluble fraction ofthe epicuticle and the water-soluble fraction released by wholeinsect cuticle in liquid medium. The larger number of geneswhose expression is altered by the dichloromethane extractscompared to locust cuticle or locust polar lipid indicates thattheir direct and indirect effects on the cell are broader. How-ever, several clusters of genes were specifically induced bygrowth on intact locust cuticle including chaperones and fol-dases (Fig. 5C). A number of studies have shown that these areupregulated in response to stress or secretion of foreign pro-
teins (8). Thus, in Trichoderma reesei the levels of proteindisulfide isomerase is a function of the quantity of total se-creted protein being produced (13). The M. anisopliae var.acridum homolog (AJ273116) was upregulated in locust cuticle(�3-fold).
One of the major groups of secreted proteins is the pro-teases. Consistent with the proteinaceous nature of the procu-ticle 6 out of 8 genes encoding secreted subtilisin proteinaseswere expressed at higher levels in locust cuticle (Table S1 inthe supplemental material). An exception was subtilisin Pr1B,which was upregulated 13-fold in beetle extract compared with3.6- and 1.5-fold on locust extract and locust cuticle, respec-tively. This unique profile implies that this subtilisin has adistinctive role. Some of the enzymes upregulated in locustcuticle such as dioxygenase and glutathione S-transferaseshave toxic substrates known to be present in procuticle.
Two flavohemoglobins (CN808105 and AJ273710) were alsodifferentially upregulated in locust cuticle. Fungal flavohemo-globins are produced to counteract nitrosative stress (22) andas such promote fungal virulence to mammals (11). Their up-regulation in locust cuticle suggests that if insects also employnitrosative challenge, their pathogens have the necessary en-zymatic defenses. Several of the enzymes differentially ex-pressed in locust polar lipid, locust extract, and locust cuticlewould affect intracellular levels of proline. Proline acts as asignaling/regulatory molecule inversely controlling cell division(25), and it would be interesting to determine the linkage, ifany, between intracellular levels of proline and the higherpercentage of appressorium differentiation in locust extract(Table 2).
In conclusion, we have demonstrated that at least four dif-ferent signals affect appressorial formation in a specific insectpathogen: a polar cuticle fraction from an appropriate host,nutrient levels, a hydrophobic surface, and the unknown signalimparted by the coverslip. The cuticle fraction elicits an activetranscriptional response, and we have identified a list of induc-ible genes that respond to this fraction and intact locust cuticle.The gene set defined in this study will now permit rigorouscomparison of gene expression programs by M. anisopliae var.anisopliae strains with widely different hosts. Such studiescould address the origin of intraspecies differences and corre-late these differences with the underlying metabolic and bio-synthetic differences that define different host ranges, identifythe mechanisms by which novel pathogens emerge with differ-ent host ranges and identify targets for restricting or broaden-ing host ranges.
ACKNOWLEDGMENT
This work was supported by USDA grants (2003-351-07-13658 and2003-353-02-13588).
REFERENCES
1. Bagga, S., G. Hu, S. E. Screen, and R. J. St. Leger. 2004. Reconstructing thediversification of subtilisins in the pathogenic fungus Metarhizium anisopliae.Gene 324:159–169.
2. Barhoom, S., and A. Sharon. 2004. cAMP regulation of “pathogenic” and“saprophytic” fungal spore germination. Fungal Genet. Biol. 41:317–326.
3. Bartnicki-Garcia, S., F. Hergert, and G. Gierz. 1989. Computer simulationof morphogenesis: mathematical basis for the hyphal tip growth. Proto-plasma 153:46–57.
4. Boucias, D. G., and J. C. Pendland. 1984. Nutritional requirements forconidial germination of several host range pathotypes of the entomopatho-genic fungus Nomuraea rileyi. J. Invertebr. Pathol. 43:288–292.
946 WANG AND ST. LEGER EUKARYOT. CELL
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from

5. Brandriss, M. C., and B. Magasanik. 1997. Genetics and physiology ofproline utilization in Saccharomyces cerevisiae: enzyme induction by proline.J. Bacteriol. 140:498–503.
6. Catlett, N. L., O. C. Yoder, and B. G. Turgeon. 2003. Whole-genome analysisof two-component signal transduction genes in fungal pathogens. Eukaryot.Cell. 2:1151–1161.
7. Charnley, A. K. 1984. Physiological aspects of destructive pathogenesis ininsects by fungi: A speculative review, p. 229–270. In J. M. Anderson,A. D. M. Rayner and D. W. H. Walton (ed.), Invertebrate-microbial inter-actions. British Mycological Society Symposium 6. Cambridge UniversityPress, London, U.K.
8. Conesa, A., P. J. Punt, N. van Luijk, and C. A. van den Hondel. 2001. Thesecretion pathway in filamentous fungi: a biotechnological view. FungalGenet. Biol. 33:155–171.
9. Crepin, V. F., C. B. Faulds, and I. F. Connerton. 2004. Functional classifi-cation of the microbial feruloyl esterases. Appl. Microbiol. Biotechnol. 63:647–652.
10. Dean, R. A. 1997. Signal pathways and appressorium morphogenesis. Annu.Rev. Phytopathol. 35:211–234.
11. De Jesus-Berrios, M., L. Liu, J. C. Nussbaum, G. M. Cox, J. S. Stamler, andJ. Heitman. 2003. Enzymes that counteract nitrosative stress promote fungalvirulence. Curr. Biol. 13:1963–1968.
12. Dzikowska, A., M. Kacprzak, R. Tomecki, M. Koper, C. Scazzocchio, and P.Weglenski. 2003. Specific induction and carbon/nitrogen repression of argi-nine catabolism gene of Aspergillus nidulans–functional in vivo analysis of theotaA promoter. Fungal Genet. Biol. 38:175–186.
13. Foreman, P. K., D. Brown, L. Dankmeyer, R. Dean, S. Diener, N. S. Dunn-Coleman, F. Goedegebuur, T. D. Houfek, G. J. England, A. S. Kelley, H. J.Meerman, T. Mitchell, C. Mitchinson, H. A. Olivares, P. J. Teunissen, J.Yao, and M. Ward. 2003. Transcriptional regulation of biomass-degradingenzymes in the filamentous fungus Trichoderma reesei. J. Biol. Chem. 278:31988–31997.
14. Freimoser, F. M., S. Screen, S. Bagga, G. Hu, and R. J. St. Leger. 2003.Expressed sequence tag (EST) analysis of two subspecies of Metarhiziumanisopliae reveals a plethora of secreted proteins with potential activity ininsect hosts. Microbiology 149:239–247.
15. Freimoser, F. M., G. Hu, and R. J. St. Leger. 2005. Variation in geneexpression patterns as the insect pathogen Metarhizium anisopliae adapts todifferent host cuticle or nutrient deprivation in vitro. Microbiology 151:361–371.
16. Gilby, A. R. 1980. Chemical methods (lipids), p. 217–252. In T. A. Miller(ed.), Cuticle techniques in arthropods. Springer-Verlag, New York, N.Y.
17. Heifetz, Y., I. Boekhoff, H. Breer, and S. W. Applebaum. 1997. Cuticularhydrocarbons control behavioural phase transition in Schistocerca gregarianymphs and biochemical responses in antennae. Insect Biochem. Mol. Biol.27:563–568.
18. Hwang, C. S., M. A. Flaishman, and P. E. Kolattukudy. 1995. Cloning of agene expressed during appressorium formation by Colletotrichum gloeospo-rioides and a marked decrease in virulence by disruption of this gene. PlantCell 7:183–193.
19. Irie, T., H. Matsumura, R. Terauchi, and H. Saitoh. 2003. Serial Analysis ofGene Expression (SAGE) of Magnaporthe grisea: genes involved in appres-sorium formation. Mol. Genet. Genomics 270:181–189.
20. Kumar, S., N. S. Punekar, V. SatyaNarayan, and K. V. Venkatesh. 2000.Metabolic fate of glutamate and evaluation of flux through the 4-aminobu-tyrate (GABA) shunt in Aspergillus niger. Biotechnol. Bioeng. 67:575–584.
21. Lecuona, R., J.-L. Clement, G. Riba, C. Joulie, and P. Juarez. 1997. Sporegermination and hyphal growth of Beauveria sp. on insect lipids. J. Econ.Entomol. 90:118–123.
22. Liu, L., M. Zeng, A. Hausladen, J. Heitman, and J. S. Stamler. 2000.Protection from nitrosative stress by yeast flavohemoglobin. Proc. Natl.Acad. Sci. USA 97:4672–4676.
23. Loubradou, G., J. Begueret, and B. Turcq. 1997. A mutation in an HSP90gene affects the sexual cycle and suppresses vegetative incompatibility in thefungus Podospora anserina. Genetics 147:581–588.
24. Loubradou, G., J. Begueret, and B. Turcq. 1999. MOD-D, a G alpha subunitof the fungus Podospora anserina, is involved in both regulation of develop-ment and vegetative incompatibility. Genetics 152:519–528.
25. Maggio, A., S. Miyazaki, P. Veronese, T. Fujita, J. I. Ibeas, B. Damsz, M. L.Narasimhan, P. M. Hasegawa, R. J. Joly, and R. A. Bressan. 2002. Doesproline accumulation play an active role in stress-induced growth reduction?Plant J. 31:699–712.
26. Nishimura, M., G. Park, and J. R. Xu. 2003. The G-beta subunit MGB1 isinvolved in regulating multiple steps of infection-related morphogenesis inMagnaporthe grisea. Mol. Microbiol. 50:231–243.
27. Nixon, R. A., I. Fischer, and S. E. Lewis. 1990. Synthesis, axonal transport,and turnover of the high molecular weight microtubule-associated proteinMAP 1A in mouse retinal ganglion cells: tubulin and MAP 1A displaydistinct transport kinetics. J. Cell Biol. 110:437–448.
28. Nowierski, R. M., Z. Zeng, S. Jaronski, F. Delgado, and W. Swearingen.1996. Analysis and Modeling of Time-Dose-Mortality of Melanoplus sangui-nipes, Locusta migratoria migratorioides, and Schistocerca gregaria (Or-thoptera: Acrididae) from Beauveria, Metarhizium, and Paecilomyces isolatesfrom Madagascar. J. Invertebr. Pathol. 67:236–252.
29. Quackenbush, J. 2002. Microarray data normalization and transformation.Nat. Genet. 32(Suppl.):496–501.
30. Roberts, D. W., and R. J. St. Leger. 2004. Metarhizium spp., cosmopolitaninsect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 54:1–70.
31. Sienko, M., J. Topczewski, and A. Paszewski. 1998. Structure and regulationof cysD, the homocysteine synthase gene of Aspergillus nidulans. Curr. Genet.33:136–144.
32. Solomon, P. S., and R. P. Oliver. 2002. Evidence that gamma-aminobutyricacid is a major nitrogen source during Cladosporium fulvum infection oftomato. Planta 214:414–420.
33. St. Leger, R. J., T. M. Butt, M. S. Goettel, R. S. Staples, and D. W. Roberts.1989. Production in vitro of appressoria by the entomopathogenic fungusMetarhizium anisopliae. Exp. Mycol. 13:274–288.
34. St. Leger, R. J., R. M. Cooper, and A. K. Charnley. 1991. Characterization ofchitinase and chitobiase produced by the entomopathogenic fungus Meta-rhizium anisopliae J. Invertebr. Pathol. 58:15–426.
35. St. Leger, R. J. 1991. Integument as a barrier to microbial infections, p.286–308. In A. Retnakaran and K. Binnington (ed.), The physiology of insectepidermis. CIRO, Canberra, Australia.
36. St. Leger, R. J., B. May, L. L. Allee, D. C. Frank, R. C. Staples, and D. W.Roberts. 1992. Genetic differences in allozymes and in formation of infectionstructures among isolates of the entomopathogenic fungus Metarhiziumanisopliae. J. Invertebr. Pathol. 60:89–101.
37. St. Leger, R. J. 1993. Biology and mechanisms of invasion of deuteromycetefungal pathogens, p. 211–229. In N. C. Beckage, S. N. Thompson and B. A.Federici (ed.), Parasites and pathogens of insects, vol. 2. Academic Press,New York, N.Y.
38. St. Leger, R. J., L. Joshi, M. J. Bidochka, N. W. Rizzo, and D. W. Roberts.1996. Characterization and ultrastructural localization of chitinases fromMetarhizium anisopliae, M. flavoviride, and Beauveria bassiana during fungalinvasion of host (Manduca sexta) cuticle. Appl. Environ. Microbiol. 62:907–912.
39. Tusher, V. G., R. Tibshirani, and G. Chu. 2001. Significance analysis ofmicroarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci.USA 98:5116–5121.
40. Wang, C.-S., G. Hu, and R. J. St. Leger. Differential gene expression byMetarhizium anisopliae growing in root exudate and host (Manduca sexta)cuticle or hemolymph reveals mechanisms of physiological adaptation. Fun-gal Genet. Biol., in press.
41. Xu, J.-R. 2000. MAP kinases in fungal pathogens. Fungal Genet. Biol.31:137–152.
42. Xue, C., G. Park, W. Choi, L. Zheng, R. A. Dean, and J. R. Xu. 2002. Twonovel fungal virulence genes specifically expressed in appressoria of the riceblast fungus. Plant Cell 14:2107–2119.
VOL. 4, 2005 TRANSCRIPTIONAL RESPONSES OF M. ANISOPLIAE 947
on June 2, 2020 by guesthttp://ec.asm
.org/D
ownloaded from