development of a multivalent recombinant protein vaccine
TRANSCRIPT

Development of a flagellin-based multivalent vaccine against Pseudomonas aeruginosa
by
Eric T. Weimer
A Dissertation Submitted to the Graduate Faculty of Wake Forest University Graduate School of Arts and Sciences
In Partial Fulfillment of the Requirements
For the Degree of
DOCTOR OF PHILOSOPHY
Microbiology and Immunology
August 2009
Winston-Salem, NC
Approved by: Steven B. Mizel, Ph.D., Advisor __________________________ Examining Committee: Mark O. Lively, Ph.D., Chairman __________________________ Jason M. Grayson, Ph.D. __________________________ W. Edward Swords, Ph.D. __________________________ Rajender Deora, Ph.D. __________________________ Daniel J. Wozniak, Ph.D. __________________________

ACKNOWLEDGEMENTS
To my advisor, Dr. Mizel, I would like to thank you for all your time,
commitment, enthusiasm, and advice throughout my time in your lab. I would not be the
scientist I am today without your mentoring.
To my classmates: Kristen, Mary Jo, Cheraton, and Cynthia, I will miss all of you
and wish all of the success that you have worked so hard for. We have shared some of
the best moments. Kristen, you have been a terrific lab mate as well as a great friend.
My deepest thanks to all the members of the Department of Microbiology and
Immunology. It was an honor to be in a department that is so supportive of its students
and always looks out for their best interest. To Aaron Graff, James Phipps, and Dr. John
Bates, I want to thank you for all the help and advice. Lab would not have been the same
without you.
To Mom and Dad, I’m eternally gratefully for all your guidance and support. I
truly couldn’t have done this without you. To Shane, the best brother I could have asked
for, thank you for all your support and just being there when I needed you. Know there is
nothing you can’t achieve in this world if you want it bad enough.
To my love, Kristin, you have been the voice of reason through the storm. I can’t
say enough how much your love and support has meant to me. My weakness is your
strength and I appreciate your patience with me.
ii

Table of Contents
ACKNOWLEDGEMENTS................................................................................................ ii
LIST OF FIGURES ........................................................................................................... iv
LIST OF TABLES............................................................................................................. vi
ABBREVIATIONS .......................................................................................................... vii
ABSTRACT....................................................................................................................... ix
INTRODUCTION .............................................................................................................. 1
CHAPTER I: A Fusion Protein Containing OprF Epitope 8, OprI, and Type A and B Flagellins Promotes Enhanced Clearance of Nonmucoid Pseudomonas aeruginosa ...... 12
CHAPTER II: Immunization of young African green monkeys with OprF epitope 8-OprI-type A- and B-flagellin fusion proteins promotes the production of protective antibodies against nonmucoid Pseudomonas aeruginosa................................................. 49
DISCUSSION................................................................................................................... 69
REFERENCE LIST .......................................................................................................... 79
SCHOLASTIC VITAE................................................................................................... 101
iii

LIST OF FIGURES
Figure 1. Illustration of constructs used in this study …………………………...16
Figure 2. TLR5-specific signaling activity of P. aeruginosa type A and B flagellins and
OprF311-341–OprI Flagellins .…………………………………………..23
Figure 3. Immunization with OprF311-341–OprI–Flagellins promotes a potent antigen-
specific humoral response ...…………………………………………26
Figure 4. Generation of antigen-specific plasma cells and MBC by OprF311-341–OprI–
Flagellins immunization …………..……………………………………....29
Figure 5. OprF311-341–OprI–Flagellins immunization generates high affinity antigen-
specific IgG …………..……………………………………....32
Figure 6. Complement-activating activity of OprI, OprF, and type A and B flagellin-
specific IgG antibodies ….………………………………....…….35
Figure 7. Antibody-dependent complement (Comp)-mediated killing of P. aeruginosa by
OprF311-341–OprI–Flagellins-immunized mouse plasma ..........................................39
Figure 8. OprF311-341–OprI–Flagellins-immunized mice display enhanced rate of
clearance following pulmonary P. aeruginosa challenge ………...….…......44
Figure 9. OprF311-341–OprI–flagellin-immunized mice are protected against severe lung
pathology during pulmonary P. aeruginosa challenge ..........................................47
Figure 10. Intramuscular immunization of African green monkeys with OprF311-341-OprI-
Flagellins promotes a potent humoral immune response ..................55
Figure 11. OprF311-341-OprI-Flagellins elicit high-affinity antigen-specific IgG. ......58
iv

Figure 12. Complement activation and complement-mediated killing by OprI, OprF, and
type A and B-flagellin-specific IgG ......................................................61
Figure 13. Passive immunization of mice with immune monkey plasma enhances
clearance of nonmucoid P. aeruginosa ......................................................66
v

LIST OF TABLES
Table 1. Bacterial strains used in these studies ...........………………………..….14
Table 2. Complement-mediated killing of additional P. aeruginosa strains .….41
vi

ABBREVIATIONS CF cystic fibrosis
CFTR cystic fibrosis transmembrane conductance regulator
P. aeruginosa Pseudomonas aeruginosa
ABC ATP-Binding Cassette
Cl chloride
ENaC epithelium sodium channel
F phenylalanine
Na sodium
DNA deoxyribonucleic acid
ExoS, T, U, Y exotoxins S, T, U, and Y
AHL acyl homoserine lactones
C3 complement component 3
C5 complement component 5
LPS lipopolysaccharide
TLR toll-like receptor
TNF-α tumor necrosis factor alpha
IL-6 interleukin-6
NO nitric oxide
Opr outer membrane protein
IgG immunoglobulin G
PilA type IV pili protein
LD50 lethal dose 50
vii

DC dendritic cell
PRR pattern-recognition receptor
MHC major histocompatibility complex
ssRNA single-stranded ribonucleic acid
LB Luria-Bertaini broth
LBNS Luria-Bertaini broth lacking NaCl
PCR polymerase chain reaction
LAL limulus amebocyte lysate
ELISA enzyme-linked immunosorbent assay
NaSCN sodium thiocyanate
Avertin 2, 2, 2-tribromoethanol
ELISPOT enzyme-linked immunosorbent spot assay
FITC fluorescein isothiocyanate
H & E hematoxylin and eosin
MBC memory B cells
BM bone marrow
WT wild-type
Y. pestis Yersinia pestis
H. influenzae Haemophilus influenzae
S. aureus Staphylococcus aureus
S. pneumoniae Streptococcus pneumoniae
MMR Measles, mumbs, rubella vaccine
DTaP Diphtheria, tetanus, acellular pertussis toxin
viii

ABSTRACT
Although chronic Pseudomonas aeruginosa infection is the major cause of morbidity and
mortality in cystic fibrosis (CF) patients, there is no approved vaccine for human use
against P. aeruginosa. The goal of my research was to establish whether a multivalent
vaccine containing P. aeruginosa type A and B flagellins as well as the outer membrane
proteins OprF and OprI would promote enhanced clearance of P. aeruginosa. To test this
I utilized two animal models, mice and 4-6 month old African green monkeys.
Intramuscular immunization of mice with flagellins + OprI (separate), or OprI-Flagellins
fusion proteins generated significant anti-flagellin IgG responses. However, only OprI-
Flagellins fusion generated OprI-specific IgG. Although immunization of young African
green monkeys with OprF311-341-OprI-Flagellins promoted a high level of antigen-specific
IgG ten days post-boost, there was significant reduction in IgG three months later.
Immunization of mice and young African green monkeys with OprF311-341-OprI-
Flagellins elicited high affinity flagellins, OprI, and OprF-specific antibodies that
individually promoted extensive deposition of C3 on P. aeruginosa. Although these
antibodies exhibited potent antibody-dependent complement-mediated killing of
nonmucoid bacteria, they were significantly less effective with mucoid isolates. Mice
immunized with OprF311-341-OprI-Flagellins had a significantly lower bacterial burden
three days post-challenge and cleared the infection at a significantly faster rate than
OprF-OprI immunized mice. In addition, mice that were passively immunized with
OprF311-341-OprI-Flagellins monkey immune plasma had significantly less bacteria,
inflammation, and lung damage throughout the infection compared to control immunized
ix

mice. Based on my results, OprF311-341-OprI-A- and B-flagellin fusion proteins have
substantial potential as a vaccine against nonmucoid P. aeruginosa, which appears to be
the phenotype that initially colonizes CF patients.
x

INTRODUCTION Cystic fibrosis (CF) is a multisystem disease affecting the digestive system and
respiratory tract. The progressive lung disease is the major cause of morbidity and
mortality in CF patients. CF is an autosomal recessive disease caused by mutation in the
gene encoding the cystic fibrosis transmembrane conductance regulator (CFTR) protein
{1,2}. CFTR belongs to the ABC (ATP-Binding Cassette) family of proteins, a large
group of related proteins that share transmembrane transport functions. The protein
encodes six membrane-spanning regions, two intracellular nucleotide-binding folds, and
a highly charged “R” domain that contains phosphorylation motifs. Normally, CFTR
functions as a regulated chloride (Cl) ion transporter and to regulate epithelial sodium
channel (ENaC) {3}. More than 1,500 different mutations have been identified in the
CFTR protein and are divided into six different classes {4}. Class I mutations result in a
defect in protein production typically by nonsense, frameshift, or splice-site mutations.
Class II mutations are generally the result in defects in protein processing or transport.
The most common class II mutation is a deletion of phenylalanine (F) at position 508
referred to as ∆F508 {5,6}. The ∆F508 mutation causes the CFTR protein to misfold and
be retained within the Golgi {7,8}. Subsequent studies revealed that the ∆F508 CFTR is
found in extremely small quantities at the apical plasma membrane of epithelial cells and
transport very low or undetectable levels of Cl {8,9}. Thus, the ∆F508 mutation disrupts
CFTR function by two distinct mechanisms, by reducing surface protein levels and
reducing capacity to transport chloride ions. Class III mutations affect the regulation of
CFTR by preventing ATP hydrolysis at the nucleotide binding domains. Class IV
mutations result in a normal amount of CFTR at the membrane but reduced function and
1

class V mutations affect the synthesis of active CFTR. Unlike Class I, class VI mutations
produce functional CFTR but reduce the stability of the protein. Class I, II, III, and VI
mutations are generally associated with severe CF phenotypes. Class IV and V mutations
are typically associated with pancreatic exocrine sufficiency and milder disease {10}.
Defects in epithelial sodium (Na) and Cl transport, and accompanying
abnormalities in fluid secretion underlie many of the disease pathology observed in CF.
In CF patients, the lack of CFTR-mediated regulation of ENaC leads to abnormal sodium
transport while the defect in CFTR leads to abnormal chloride ion transport. The result
of this ion imbalance is the airway surface is depleted of water (i.e. thickened mucus) and
mucus plugging occurs in the upper respiratory tract {11,12}. Increased levels of
chloride ions in the respiratory tract of CF patients may inhibit the antimicrobial activity
of beta-defensins and neutrophils. In addition, increased fluid viscosity, impaired
mucociliary clearance, and dehydrated mucosal surfaces lead to the inability to clear
pathogens such as Haemophilus influenzae, Staphylococcus aureus, Burkholderia
cepacia, and eventually the Gram-negative, opportunistic pathogen Pseudomonas
aeruginosa {13}.
CF patients are particularly susceptible to P. aeruginosa infections due in part to
defects in normal respiratory tract function. The progressive mucus buildup within the
respiratory tract traps the bacteria in the hypoxic environment generated by increased
oxygen consumption by epithelial cells {14}. Within the low oxygen environment, P.
aeruginosa undergoes a distinct phenotypic change through accumulations of mutations
in mucA and the conversion to a mucoid phenotype {15-17}. The mucoid form of P.
aeruginosa is characterized by the over-production of the exopolysaccharide alginate and
2

loss of motility (i.e. loss of flagellin expression). There is a well-documented inverse
relationship between alginate and flagellin expression {18,19}. Alginate-overproducing
P. aeruginosa is rarely the initial infecting strain {20-25}. The appearance of mucoid P.
aeruginosa occurs over approximately a 10-year span and is a poor prognosis indicator in
CF patients {20,22,23,25}. Once infection has been established, neither the massive
infiltration of neutrophils nor the heavy use of antibiotics are capable of controlling the
bacteria. Recruited neutrophils consequently release elastase at concentrations that
overwhelm the protective capacity of lung and contribute to tissue destruction {26-28}.
As a result, large amounts of free DNA are released by degranulating neutrophils,
contributing to the increased viscosity of the airway mucus {15}. The accumulation of
free DNA acts as a scaffold for P. aeruginosa biofilm formation – a major virulence
factor of P. aeruginosa {29,30}. Over-production of alginate and eventual biofilm
development allows P. aeruginosa to evade the immune system and also increases
antibiotic resistance, making it extremely difficult to eradicate the infection {22,23}.
Chronic P. aeruginosa infection is the major contributor to respiratory failure in CF
patients.
Eventually, patients with CF succumb from respiratory failure because of a loss of
lung function. This results from the chronic inflammatory process triggered by persistent
P. aeruginosa infections {31-34}. Whole genome analysis of P. aeruginosa isolates
from CF patients revealed several virulence factors that are required to initiate acute
infections and are subsequently selected against during chronic infections. For example,
the exopolysaccharide Psl is important for attachment as well as biofilm initiation and
maintenance {35-37}. Many of these virulence factors allow P. aeruginosa to evade or
3

inhibit the immune response {38}. The type III secretion system is activated upon contact
with epithelial cells and is a major determinant of virulence {39}. P. aeruginosa secretes
four known effector proteins via type III secretion system: ExoS, ExoT, ExoU, and
ExoY. Secretion of the exotoxin S (ExoS) through the type III secretion system has been
shown to inhibit the activation of caspase-1 {40}. ExoT, possess an N-terminal GTPase
activating domain and a C-terminal adenosine diphosphate (ADP) ribosyltransferase
domain. ExoU is a phospholipase and ExoY is an adenylate cyclase {41,42}.
In contrast to invasive properties of type III secretion system, quorum sensing
regulates a complex circuit involving cell to cell signaling {43}. Quorum-sensing
molecules are typically acyl homoserine lactones (AHL), which are freely diffusible.
When a threshold AHL concentration is reached, AHL binds LasR/RhlR transcriptional
activators to induce expression of certain genes. P. aeruginosa predominately makes two
autoinducers: N-3-oxododecanoyl homoserine lactone and N-butyryl-L homoserine
lactone {44}. Activation of the quorum sensing signaling cascade promotes biofilm
formation and persistence in the lung {45}. In addition, quorum-sensing molecules can
directly modulate the immune response. Several investigators have shown that AHL
induces cyclooxygenase-2 and prostaglandin-E2, which inhibit the induction of adaptive
immune responses {44}. Outer membrane protein F (OprF) has also been shown to bind
interferon gamma and activate expression of quorum sensing virulence genes {46}.
Unlike quorum-sensing, the function of several virulence factors is inhibition of the
immune response.
Evasion of the complement cascade is mediated by binding and sequestering the
complement regulatory protein Factor H by bacterially expressed Tuf {47}. In addition,
4

P. aeruginosa secrete enzymes such as alkaline protease and elastase, which degrade
complement components and thus limit the role of complement in the clearance of early
pulmonary P. aeruginosa infections {48}. The critical role of complement in the
clearance of P. aeruginosa is evidenced by the observation that C3 and C5 KO mice were
unable to clear P. aeruginosa after challenge {49,50}. Also, P. aeruginosa express
lipopolysaccharide (LPS) variants that interfere with C3b deposition {51}.
The flagellum is an important virulence factor for P. aeruginosa, as non-
flagellated P. aeruginosa strains are less virulent {52}. P. aeruginosa strains that
initially colonize CF patients are generally flagella-positive, composed of ‘‘A’’ and/or
‘‘B’’ flagellin subtypes {53-55}. Both types of flagellin are crucial for establishing
infections in CF patients, as well as being involved in chemotaxis, motility, adhesion and
inflammation {56}. Type-A has a more variable molecular mass (45-52 kDa), while
type-B has an invariant molecular mass of approximately 50 kDa {57}. Both flagellins
are glycosylated {57,58}. Glycosylation of flagellin occurs in the domain outside the
region required for toll-like receptor-5 (TLR5) signaling {59}. Interaction with TLR5
induces the secretion of proinflammatory cytokines tumor necrosis factor alpha (TNF-α)
and interleukin-6 (IL-6) {60-62}. However, flagellin binding to TLR4/5 heteromeric
complexes is required for the induction of nitric oxide (NO) synthesis {62}. Also, P.
aeruginosa, Salmonella typhimurium, and Listeria monocytogenes flagellin has been
shown to interact with the intracellular cytoplasmic protein Ipaf {63-68}. Binding of
flagellin to Ipaf promotes activation of caspase-1, release of IL-1β, and subsequent cell
death in macrophages {69}. Delivery of flagellin to the cytoplasm and activation of Ipaf
of cells occurs via type III secretion system {40}. Flagellin, type III secretion system,
5

ExoS, LPS, alginate as well as other virulence factors play a key role in the establishing
and maintaining persistent P. aeruginosa infections in CF patients.
Initial efforts to develop a P. aeruginosa vaccine focused primarily on live
attenuated strains or components such as LPS, or alginate {70-79}. Several investigators
have utilized polysaccharide conjugate vaccines with carrier proteins keyhole limpet
hemocyanin, tetanus toxoid, or exotoxin A {70,74,79-81}. A live, attenuated vaccine
strain was employed by deletion of the aroA gene that is required for synthesis of
aromatic amino acids. This approach has been utilized to generate several other
attenuated strains, e.g. Salmonella species {82}. An attenuated aroA mutant of
Salmonella enterica serovar Typhimurium was used to express OprF-OprI as well as the
O-antigen from P. aeruginosa {72,83}. However, single aroA deletion mutants in S.
enterica retain significant virulence and thus make them unacceptable as human vaccines.
In contrast, P. aeruginosa aroA mutants are intrinsically less virulent and strains
expressing O-antigen have been evaluated in mice as a potential vaccine {84,85}. The
vaccine elicited high titers of O-antigen specific IgG that provide protection from
homologous but not heterologous challenge {84}.
Testing of LPS conjugate vaccines began in the late 1960s, when several
investigators evaluated a LPS vaccine in burn and CF patients. Although the vaccine
showed promise, it was ultimately stopped due to adverse side effects {86-92}.
Following modifications to the lipid A portion of LPS, an octavalent O-antigen
conjugated to exotoxin A was developed and tested in burn patients. In addition, the
vaccine was well tolerated and elicited high affinity antibodies that reduced the incidence
of infection in CF patients {74,93}. The results from this study were not able to obtain
6

regulatory approval due to lack of controls. A subsequent controlled study demonstrated
no significant difference between placebo and vaccine immunized CF patients {94}. As
indicated above, the major limitation of LPS vaccines has been toxicity and an inability
to provide protection against multiple P. aeruginosa serotypes {77,81,95,96}. In
contrast, the exopolysaccharide alginate is structurally less diverse than LPS. Alginate is
a linkage of β1-4 D-mannuronic acid and L-guluronic acid residues {97}. In humans and
mice, antibodies elicited by alginate immunization were mostly nonopsonic {78,98}.
Additional studies revealed the immunization with only the high molecular weight
fraction and at least 100µg of alginate elicited opsonic antibodies in a limited number of
healthy volunteers. However, the opsonic antibodies that were generated were not long-
lived, returning to background levels by 28 days post-boost {78,98}. Expression of
alginate is very low on nonmucoid strains of P. aeruginosa {99,100}. Given the low
expression level of alginate on initial infecting strains, investigators have focused on type
IV pili protein (PilA), type III secretion system protein, PcrV, ExoA, flagellin, and outer
membrane proteins I and F (OprI and OprF), as protective antigens. There is high level
of expression of flagellin, OprI, OprF, on nonmucoid P. aeruginosa as well as a high
degree of sequence conservation among CF isolates {72,101-117}. In deed, real-time
PCR using oprI-specific gene primers is often used to identify P. aeruginosa infection
{101,118,119}.
Immunization with the OprI antigen of P. aeruginosa and an appropriate adjuvant
elicited a protective response in mice that correlates with the titer of OprI-specific IgG
{103}. Importantly, Eckhardt et. al. {120} determined that the presence of OprI-specific
IgG that mediated complement activation as the best in vitro correlate of protection
7

compared to total IgG response and antibody affinity. In related studies, administration
of OprI-specific monoclonal antibodies mediated protection {104}. Four intramuscular
immunizations of healthy volunteers with 50µg of OprI absorbed on alum (aluminum
hydroxide) elicited elevated antibodies levels that remained high for 4 months post-
immunization {121,122}. In addition, an adenovirus expressing epitope 8 (amino acids
311-341) (Epi8) of OprF provided protection against acute P. aeruginosa infection
{116,117}. DNA vaccination with oprF elicited antibodies that were opsonic and
reduced the appearance of lesion during P. aeruginosa infection {113,123}. From the
success of these studies, several investigators began focusing on a fusion peptide
containing OprF and OprI as a potential vaccine candidate. Although large amounts of
this protein were required for an optimal response, immunization with an OprF-OprI
fusion protein resulted in a 95-fold increase in the lethal dose-50 (LD50) for mice. A
subsequent studies in burn, chronic obstructive pulmonary disease, and CF patients
revealed that 1mg of a OprF-OprI fusion protein was required to elicit a 3-fold increase in
IgG titers {110,124,125}. However, it is not known whether antibodies to OprF-OprI
mediate opsonization and killing of P. aeruginosa strains. In addition, investigators have
used type IV pili coupled to either tetanus toxoid or exotoxin A. Immunization of rabbits
with either conjugate vaccine elicited antibodies that reduced adherence and enhanced
clearance of P. aeruginosa following pulmonary infection {126,127}. In light of any
vaccine that leads to a substantial reduction in the incidence of infection, investigators to
use multivalent vaccines. For example, Saha et. al. {106} used DNA vaccination with
PcrV, PilA, OprF, and OprI to demonstrate the effectiveness of multivalent immunization
at enhancing the clearance of P. aeruginosa. A phase III clinical trial of P. aeruginosa
8

flagellins in CF patients demonstrated that the vaccine was well tolerated and caused a
30% reduction in the incidence of infection {128}. Several investigators have shown that
passive immunization with anti-flagellin IgG antibodies mediates protection from P.
aeruginosa infection {56,129,130}.
The use of adjuvants has been shown to enhance adaptive immune responses.
Currently, the only FDA-approved adjuvant is alum. Until recently, the adjuvant effect
of alum was not understood. Kool and colleagues {131,132} demonstrated that alum
particles activate inflammatory dendritic cell (DC) and promote release of uric acid that
elicits an inflammatory response. Alum has been shown to activate the cytoplasmic
NOD-like receptor called NLRP3 or Nalp3, which associates with ASC and caspase-1 to
form a protein complex called inflammasome. The activation of the inflammasome leads
to the secretion of several proinflammatory cytokines such as IL-1β and IL-18
{132,133}. NOD receptors are cytoplasmic form of pattern-recognition receptors (PRR).
TLR are membrane bound PRR. PRR on DC function to alert the immune system to
“danger”. PRR are a collection of receptors that recognize pathogen-associated
molecular patterns (PAMPs) on a diverse group of pathogens. Surface and endosomal
recognition of bacterial, viral, and fungal pathogens occurs through TLR. Upon TLR
stimulation, DC undergo a maturation process leading to increased expression of co-
stimulatory molecules CD40, CD80, and CD86 as well as the secretion of cytokines such
as IL-2, IL-12, and type I interferon. DC downregulate phagocytosis and increase
antigen processing and presentation of antigen peptides in the context of MHC class II or
I. DC are potent activators of CD8+ and CD4+ T cells, providing a key link between
innate and adaptive immunity. TLR agonists function not only as adjuvants but also as
9

antigens. TLR agonists fused to proteins are more effective than the separate proteins
{115,134-138}. CpG (TLR9), ssRNA (TLR7 & TLR8), LPS (TLR4), poly I:C (TLR3)
and flagellin (TLR5) have all been shown to function as adjuvants {61,134,136,138-
140,140-148}.
The conserved regions of flagellin are responsible for binding directly to a leucine
rich repeat on the extracellular domain of TLR5 {149}. Schirm et al. {59} demonstrated
that amino acids 88-97 were required for type A flagellin signaling via TLR5. Signaling
via TLR5 is dependent on IRAK-1 {150} and MyD88 {151}. Honko and Mizel {60}
demonstrated the in vivo stimulatory effects of flagellins from P. aeruginosa and
Salmonella. Intratracheal (i.t.) administration of flagellin induced maximal neutrophilic
infiltration within 12 hours. Consistent with the neutrophilic infiltration, cytokines TNF-
α and IL-6 as well as chemokines macrophage inflammatory protein-1 alpha and
keratinocyte-derived chemokine were upregulated four-fold in flagellin treated mice
{60}.
Flagellin has been proven to be an effective adjuvant in mice {60,61,134-
136,143,152,153} as well as cynomolgus and African green monkeys {61,154}. Arnon
and colleagues first demonstrated that flagellin could function as an adjuvant by
enhancing the influenza response by intranasal immunization with recombinant flagellin
expressing influenza peptides. Importantly, prior exposure to flagellin does not inhibit
the effectiveness of flagellin {143,152,155,156}. A recent study, demonstrated the
adjuvant effect of flagellin is mediated by tlr5+CD11c+ cells {157}. Flagellin is an
effective adjuvant for CD4+ T cells in vivo and in vitro {135,140,144}. The efficacy of
flagellin to promote a protective response has been demonstrated for bacteria and viral
10

pathogens when used with recombinant proteins as antigens {134-136,145,155}. Several
investigators have reported protection against P. aeruginosa using flagellin as an antigen
and adjuvant {115,128,145}.
Although experimental P. aeruginosa vaccines have shown promise in initial
preclinical and clinical trials, none have achieved the level of response required for
protection against P. aeruginosa in CF patients. A limitation of the previous vaccines
was the age of the CF patients when they were immunized. Given the pathophysiologic
events associated with CF in combination with the pathogenic mechanisms associated
with P. aeruginosa infection, I hypothesize that vaccine efficacy will be greatly enhanced
if administered to CF patients at a very young age. In addition, although the magnitude
of the humoral response was determined in previous studies, the functional quality of the
induced antibodies was not addressed, e.g., antibody affinity and the ability to activate
complement or promote opsonization {115,120,121,158,159}.
After critical review of the literature, I have identified several features that are
important for an effective P. aeruginosa vaccine: the presence of a potent adjuvant, the
ability to induce high titer antigen-specific IgG that exhibits a high degree of functional
activity (for example, complement activation), multivalency, and the ability to induce a
robust memory response. To that end, I generated a multivalent vaccine containing type
A and B-flagellin, OprF, and OprI and have evaluated its immunogenicity and protective
potential in mice as well as 4-6 month old African green monkeys.
11

Chapter I
A Fusion Protein Containing OprF Epitope 8, OprI, and Type A and B Flagellins
Promotes Enhanced Clearance of Nonmucoid Pseudomonas aeruginosa
E.T. Weimer, H. Lu, N. D. Kock, D.J. Wozniak, S.B. Mizel
The following manuscript was published in Infection and Immunity, volume 77, issue 6, pages 2355-2366, 2009 and is reprinted with permission. Stylistic variations are due to the requirement of the journal. E.T. Weimer performed the experiments and prepared the manuscript. Dr. Steven B. Mizel acted in an advisory and editorial capacity. Copyright © American Society for Microbiology 2009.
12

MATERIALS AND METHODS
Strains and plasmids. Bacterial strains and plasmids used in this study are
described in Table 1. Escherichia coli cultures were maintained at 37ºC in Luria-Bertaini
(LB; 10 g/L tryptone, 5 g/L yeast extract, 5 g/L NaCl) broth, while P. aeruginosa was
cultured in LB broth lacking NaCl (LBNS; 10 g/L tryptone, 5 g/L yeast extract). Solid
media were prepared by adding 1.0-1.5% select agar (Gibco-BRL). Plasmids in E. coli
were selected using media supplemented with antibiotics at the following concentrations:
carbenicillin (Cb 100 µg ml-1), gentamicin (Gm 10 µg ml-1). Plasmids in P. aeruginosa
were selected on media containing Cb (300 µg ml-1), Gm (100 µg ml-1), and irgasan (Irg
25 µg ml-1). E. coli strain JM109 was used for all cloning procedures while E. coli SM10
was used to transfer plasmids into P. aeruginosa by bi-parental mating {160}. The P.
aeruginosa strains used were PAO1 and its derivatives WFPA850, WFPA852,
WFPA854, WFPA860, WFPA862, WFPA864, and WFPA866. Vectors pEX18Gm,
pEX18Ap or derivatives were used for cloning and gene replacements (Table 1).
Construction of non-polar deletion mutations in fliC, oprF, and oprI. To
engineer unmarked, non-polar deletion mutations in fliC, oprF, and oprI, we utilized a
previously described method {161}. Internal fragments of coding sequences within each
gene were deleted using a modified PCR technique termed splicing by overlap extension
{162}. In this assay, four gene-specific primers were employed in three separate PCR
reactions to generate DNA fragments with a defined in-frame deletion of coding
sequences within the fliC,
13

Table 1. Bacterial Strains used in this study Strain Description Source PAK WT {19} PAO1 WT {19}
WFPA850 In-frame fliC deletion in PAO1
This study
WFPA852 In-frame oprF deletion in PAO1
This study
WFPA854 In-frame oprI deletion in PAO1
This study
WFPA860 In-frame fliC and oprI deletions in PAO1
This study
WFPA862 In-frame fliC and oprF deletions in PAO1
This study
WFPA864 In-frame fliC, oprF, and oprI deletions in PAO1
This study
WFPA866 In-frame oprF and oprI deletions in PAO1
This study
T69833 Mucoid CF isolate Wozniak unpublished 1286 Nonmucoid CF isolate Wozniak unpublished
PDO300M Mucoid PAO1 {163} PDO300NM Nonmucoid PDO300
deficient in alginate production
{163}
14

oprF, or oprI genes. The primers were also designed such that the final amplicon,
harboring the specified deletion allele harbored restriction sites to allow direct cloning
into pEX18Ap or pEX18Gm resulting in plasmid pHL150 (∆fliC), pHL153 (∆oprF), or
pHL155 (∆oprI). The mutant alleles were introduced into the PAO1 chromosome as
outlined {164}. The merodiploids were resolved by growing on sucrose-containing
media and introduction of the deletion allele, which was verified by PCR.
Recombinant proteins. DNA encoding full-length type A-flagellin of P.
aeruginosa strain PAK and DNA encoding full-length type B-flagellin of strain PAO1
were each amplified by PCR and ligated into pET29a. DNA encoding the mature OprI
antigen of P. aeruginosa strain PAO1 (amino acids 21-83) was amplified by PCR and
ligated into pET29a or to the 5’ end of type A and B-flagellin genes in pET29a
generating constructs that encode OprI-type A Flagellin and OprI-type B flagellin. DNA
encoding the OprF epitope 8 (amino acids 311-341) of P. aeruginosa strain PAO1 was
amplified by PCR and ligated into pET29a or to the 5’ end of oprI-A-flagellin and oprI-
B-flagellin. The structure of each of the final proteins are presented in diagrammatic
form in Figure 1.
All expressed proteins were purified by metal ion affinity chromatography as
previously described {61,165}. Acrodisc Q membranes were used to deplete endotoxin
and nucleic acids. Endotoxin levels were <10pg/µg for all of the proteins (as detected by
QCL-1000 chromogenic Limulus amebocyte lysate (LAL) test kit, Cambrex Corporation
(East Rutherfrod, NJ)).
15

Figure 1. Illustration of the constructs used in this study.
16

17

ELISA for TNF-α and antigen-specific IgG. TNF-α levels in cultures of RAW 424
(TLR5+) or RAW 264.7 (TLR5-) cells were measured using a commercial ELISA kit
(OptiEIA ELISA, Becton Dickinson) according to the manufacturer's instructions. Data
represent three independent experiments with triplicate samples in each experiment.
Titers of antigen-specific IgG were measured using MaxiSorb plates coated with
100µl of antigen (A-flagellin, B-flagellin, OprI, or OprF) at 10 µg/ml in sterile PBS. The
plates were incubated overnight at 4°C and then blocked with 10% newborn calf serum in
PBS. Plasma samples (in triplicate) were added, and the plates incubated overnight at
4°C, followed by secondary anti-Ig antibodies (Roche Diagnostics) for 2h at room
temperature. Peroxidase activity was detected with 3,3’,5,5’-tetramethylbenzidine
(TMB) liquid substrate system (Sigma-Aldrich) and stopped with 2 N H2SO4. Endpoint
dilution titers were defined as the inverse of the lowest dilution that resulted in an
absorbance value (at 450 nm) of 0.1 over that of naive plasma. Groups of at least 7 mice
were used. To determine relative antibody affinities, the ELISA assay was conducted as
described above with the addition of a 15 min incubation with sodium thiocyanate
(NaSCN) (Sigma) solution as described previously {165,166}.
Mice. 6-8 week old BALB/c and DBA/2 mice were purchased from Charles
River Laboratories. All animals were maintained under pathogen-free conditions. All
research performed on mice in this study complied with federal and institutional
guidelines set forth by the Wake Forest University Animal Care and Use Committee.
Intramuscular (i.m.) immunization of mice. Groups of 7 mice were anesthetized
with Avertin (2, 2, 2-tribromoethanol [Sigma]; tert-amyl alcohol [Fisher]) by
intraperitoneal injection. Small volumes (20µL total) containing antigen and adjuvant in
18

PBS were injected using a 29 ½ G needle into the right calf of mice. Mice were boosted
at 4 weeks via the same route, and bled two weeks later. Plasma was prepared and stored
at -70°C until analysis.
ELISPOT assay. The frequency of antigen-specific plasma cells was determined
using limiting dilution analysis as previously described {167}. Briefly, Immobulin-P
high-affinity protein binding ELISPOT plates (Millipore) were coated with 100µL of A-
flagellin, B-flagellin, OprI, or OprF (10µg/mL) in sterile PBS. Bone marrow and spleen
were collected 45 days post boost, single cell suspensions were prepared, and dilutions of
the cells (5x105/well) were added to the antigen-coated wells. Plates were then incubated
at 37°C for 5 hours, washed, and probed with goat anti-mouse (4°C overnight). Plates
were developed using HRP-Avidin D diluted 1:1000 (Southern Biotechnology) and 3-
amino-9-ethylcardbazole (AEC) and dried overnight. Spots were enumerated using a
dissecting microscope. Only wells that contained ≥ 4 spots were counted for analysis.
Total spleen cell plasma cell numbers were calculated by multiplying the number of cells
in the spleen by the number of spots per million spleen cells. Total bone marrow plasma
cell numbers were calculated in the same manner with an additional multiplication by 7.9
to compensate for total bone marrow {168}.
To determine the frequency of antigen-specific memory B cells, the bone marrow
and spleen cells were incubated in vitro for 5 days in the presence of 1µg/mL OprF311-341-
OprI-Flagellins and then plated as described above. The number of memory B cells was
determined by subtracting the number of plasma cells from the 5 hr incubation from the
total number of plasma cells after the 5-day culture. Results are shown for two
independent experiments.
19

Antigen-specific IgG binding to P. aeruginosa. P. aeruginosa strains were
incubated with heat-inactivated control or immune mouse plasma for 1 h at 4°C prior to
staining with AlexaFlour647-conjugated anti-mouse IgG (Invitrogen) for 1 hr at 4°C.
Data are representative of two experiments with triplicate samples in each experiment.
Antigen-specific IgG-mediated complement activation. Control and immune
mouse plasma were diluted 1:10 and heat-inactivated at 56°C for 1 hour prior to use. P.
aeruginosa strains were grown in LANS broth to an OD600 of 0.5 (~108 cfu/mL), washed
2 times with sterile PBS, and then incubated with mouse plasma for 1 hr. The bacteria
were then washed and incubated for 1 hr at 37°C with 5% rabbit serum (Innovative
Research) as a source of complement. Finally, the bacteria were stained with goat anti-
rabbit C3-FITC (MP Biomedical). Flow cytometric analysis was performed using a BD
FACSCaliber and data analyzed with FloJo software (Tree Star, Inc., Ashland, OR).
Representative histograms of three experiments are shown. Complement-mediated
killing was performed as described above with the exception that the bacteria were
incubated for 4 hr with rabbit serum. A time-course experiment revealed minimal killing
at 1 hour with rabbit serum (data not shown). Percent bacteria killed was quantitated by
(number of input bacteria – number of recovered bacteria) / (number of input bacteria) X
100.
Respiratory challenge with agar-embedded P. aeruginosa. P. aeruginosa strains
were grown in LBNS to 108 cfu/mL. One-part bacteria were added to 9 parts warm
(52°C) 1.5% trypticase soy agar. After five minutes, the agar:bacteria mixture was
injected into rapidly spinning warm heavy mineral oil using a 22 gauge needle. The
20

suspension was then mixed for 6 min. The agar beads were then cooled on ice for 20 min
and washed 3 times with sterile PBS. The final volume was adjusted to approximately
5mL. To determine the number of cfu/mL, the agar:bacteria beads were homogenized
and bead size determined by comparison to 100-150µm chromatography beads. Mice (6-
7 per group) were anesthetized with Avertin by intraperitoneal injection and then 50µL of
agar-embedded bacteria were instilled intra-tracheally using a sterile gel-loading tip.
Histology. Lungs were harvested and transferred to 10% formalin for 24 h. The
tissue then was trimmed, embedded in paraffin, cut at 4 µm, and stained with
hematoxylin and eosin by routine methods. For histological examination, groups of four
mice were used for each condition. Slides were scored as blind groups on an increasing
severity index that incorporates values for consolidation, bronchiolar and vascular
degenerative changes, and edema (range for each factor: 0 to 4). Total inflammation
score was calculated by the sum of all categories. Representative images are shown from
4 animals/group with 3 sections by animal.
Statistical analyses. Statistical analysis was performed using SigmaStat 3.10
(Systat Software, Inc., Point Richmond, CA). For normally distributed data sets,
significance was determined using the Student's t-test. The significance of data sets,
which were not normally distributed, or were of unequal variance were determined using
the Mann–Whitney rank sum test. Where applicable a two-way ANOVA test was
applied. P values of less than 0.05 were considered significant.
21

RESULTS
TLR5-specific signaling activity of P. aeruginosa A- and B-flagellin and OprF311-341-
OprI-Flagellins. In order to generate antigens with flagellin as the adjuvants, we
generated several constructs as shown in Figure 1. In view of the insertion of the OprF
and OprI sequences at the N-terminus of flagellin, it was important to determine if this
addition would have a negative impact on the ability of each flagellin, i.e., type A or B, to
signal via TLR5. To test the ability of P. aeruginosa flagellins either alone or as part of a
tri-fusion with OprF and OprI (see Fig 1), to signal via TLR5, RAW 424 (TLR5+) or
RAW 264.7 (TLR5-) cells were incubated with 1pM-1nM of each protein and production
of TNF-α was assessed. Stimulation of RAW 424 cells with P. aeruginosa type A or B-
flagellin, OprI-type A or B-flagellin, or OprF311-341-OprI-type A or B-flagellin resulted in
a concentration-dependent increase in TNF-α (Fig 2A-C). In contrast, none of these
proteins induced TNF-α production in cultures of TLR5- RAW 264.7 cells. Consistent
with previous results with the P. aeruginosa flagellins, the half-maximal stimulation
occurred at 16 pM for A-flagellin and 40 pM for B-flagellin {169}. There was no
significant difference in the half-maximal stimulation between type A or B flagellin,
OprI-type A or B-flagellins, or OprF311-341-OprI-type A or B-flagellins. Thus, the
presence of OprF-OprI at the N-terminus of type A or B-flagellin does not alter
recognition and signaling via TLR5.
Immunization with OprF311-341-OprI-Flagellins promotes a potent antigen-specific
humoral response. To assess the ability of OprF311-341-OprI-A and B-
22

Figure 2. TLR5-specific signaling activity of P. aeruginosa A- and B-flagellin and
OprF311-341-OprI-Flagellins. RAW 424 (TLR5+) and RAW 264.7 (TLR5-) were
stimulated with 10-9 to 10-12 M of protein. At 4 hours post stimulation, supernatants were
harvested and the amount of TNF-α was determined by ELISA. A) A-flagellin and B-
flagellin B) OprI-Flagellins fusion C) OprF311-341-OprI-Flagellins fusion. Data represent
the results of three independent experiments done in triplicate.
23

24

flagellin to promote an antigen-specific humoral response, groups of 7 BALB/c or
DBA/2 mice were immunized with 5µg of each type A and B-flagellin + 10µg OprI, 5µg
OprI-type A and B-flagellin fusion proteins, or 5µg OprF311-341-OprI-type A and B-
flagellin fusion proteins. Prior experiments established that immunization of BALB/c
mice with 5µg OprI-Flagellins generated a maximal IgG response to flagellin and OprI
(data not shown). Control mice received either OprI or OprF311-341-OprI at equivalent
molar doses. DBA/2 mice were used because previous studies identified DBA/2 mice as
more susceptible to P. aeruginosa infection than BALB/c and C57BL/6 mice {32,34}.
Four weeks later, mice were boosted in an identical manner. Two-weeks after the boost
the mice were bled and plasma was prepared for analysis of circulating antigen-specific
IgG. Mice immunized with OprI-Flagellins or OprF311-341-OprI-Flagellins exhibited a
robust OprI-specific IgG response (Fig 3). In contrast, there was no significant OprI-
specific IgG in mice given only OprI or type A and B-flagellin + OprI. In all cases,
flagellin-specific responses were extremely robust. Mice immunized with OprF311-341-
OprI-A- and B-flagellin exhibited a high-level of OprF-specific IgG as well as flagellin
and OprI-specific IgG.
In addition to determining the titers of antigen-specific IgG following
immunization with OprF311-341-OprI-A- and B-flagellin, we also evaluated IgG isotypes
and IgE. Plasma was prepared from immune mice as described above and antigen-
specific IgG subclasses and IgE were determined by ELISA. Immunization of mice with
OprF311-341-OprI-Flagellins did not elicit any detectable antigen-specific IgE (data not
shown). This finding is consistent with our prior
25

Figure 3. Immunization with OprF311-341-OprI-Flagellins promotes a potent
antigen-specific humoral response. BALB/c or DBA/2 mice were immunized
intramuscularly with 5µg of A-flagellin and B-flagellin + 10µg OprI (white bars), 5µg
OprI-A-flagellin + 5µg OprI-B-flagellin (grey bars), or 5µg OprF311-341-OprI-A-flagellin
+ 5µg OprF311-341-OprI-B-flagellin (black bars). At 4 wks post-immunization, animals
were boosted and 2 wks post-boost blood was collected and antigen-specific total IgG
was determined by ELISA. Data represent at least 7 mice per group in triplicate. * =
P<0.05, using a Mann–Whitney rank sum test.
26

27

work demonstrating that flagellin does not promote antigen-specific IgE responses {61}.
Although high titers of antigen-specific IgG2a were induced, the overall response to
OprI-A- and B-flagellins or OprF311-341-OprI-A- and B-flagellin was biased towards IgG1
(data not shown). This finding is consistent with our prior work on flagellin as an
adjuvant in a Y. pestis vaccine {61}.
Generation of antigen-specific plasma and memory B cells (MBC) in response to
OprF311-341-OprI-A- and B-flagellin. In view of the robust antigen-specific IgG
response, we evaluated the frequency of antigen-specific plasma and memory B cells
generated in response to OprF311-341-OprI-A- and B-flagellin. Mice were immunized with
5µg of OprF311-341-OprI-A- and B-flagellin as described above and 45 days post-boost,
bone marrow (BM) and spleen were harvested and the frequency of antigen-specific
plasma and memory B cells were determined by ELISPOT. Antigen-specific plasma
cells were determined following 5 hr incubation. Eighty-five percent of antigen-specific
plasma cells were found in the BM and 15% in the spleen. Consistent with the IgG titer
data (Fig 3), there were more plasma cells for type A- and B-flagellin (~200/106 BM
cells) than OprI (42) and OprF (30) (Fig 4A). No plasma cells were detected in wells that
contained cells from non-immune mice. Although significantly more antigen-specific
plasma cells were found in the bone marrow, a substantial number of plasma cells
remained in the spleen (Fig 4B). The retention of antigen-specific plasma cells in the
spleen correlated with the immunogenicity of each antigen.
28

Figure 4. Generation of antigen-specific plasma and memory B cells by OprF311-341-
OprI-Flagellins immunization. DBA/2 mice were immunized intramuscularly with 5µg
of OprF311-341-OprI-Flagellins. Bone marrow and spleens were harvested 40-days post-
boost and analyzed for antigen-specific plasma and memory B cells by ELISPOT. (A)
Frequency of antigen-specific plasma and MBC cells. (B) Total number of plasma and
MBC cells. Results are the average of 2 independent experiments on 5 mice.
29

30

In contrast to plasma cells, the generation of MBC was equivalent across flagellins and
OprI (Fig 4A). The fewer number of OprF-specific memory B cells (108 MBC/106 cells)
was not unexpected given the presence of only a single epitope. Nonetheless, our results
clearly establish that OprF311-341-OprI-A- and B-flagellin elicits not only a significant
numbers of plasma cells, but also a substantial pool of memory B cells.
OprF311-341-OprI-Flagellin immunization generates high-affinity antigen-specific IgG.
Since antigen affinity plays a critical role in the functional activity of an antibody, we
evaluated the relative affinity of the IgG generated following immunization with OprF311-
341-OprI-A- and B-flagellin. The relative affinity of antibodies can be assessed in an
ELISA by determining the concentration of sodium thiocyanate required to reduce
antibody binding by 50%. As shown in Fig 5, immunization with OprI-Flagellins or
OprF311-341-OprI-Flagellins generated IgG with equivalent relative affinities for the three
antigens. For comparative purposes, a flagellin + Y. pestis F1 antigen vaccine generated
F1-specific IgG requiring 3M sodium thiocyanate for 50% reduction in binding {165}.
Given the observation that these antibodies provide complete protection against
respiratory challenge with Y. pestis {61,154}, we have defined high affinity IgG as those
antibodies requiring 2-3 M sodium thiocyanate for 50% reduction in antigen binding.
OprF311-341-OprI-A-and B-flagellin immune plasma had an average relative IgG affinity
approaching 3 M for flagellin, OprI, and OprF (Fig 5). Thus,
31

Figure 5. OprF311-341-OprI-Flagellins immunization generates high-affinity antigen-
specific IgG. Plasma samples from mice that received OprI-Flagellins or OprF-OprI-
Flagellins were used to determine relative antibody affinity for A-flagellin, B-flagellin,
OprI, and OprF. Antigen-specific IgG affinity was determined by ELISA using dilutions
of sodium thiocyanate (NaSCN). Data are presented as molar concentration of NaSCN
required to reduce absorbance 50%. Samples from the same mice were used in the
experiments presented in Figures 2 and 3. There were 7 mice per group with each sample
done in triplicate.
32

33

the data are consistent with the conclusion that OprF311-341-OprI-A- and B-flagellin elicits
high-affinity antigen-specific IgG.
Complement activating activity of antibodies specific for OprI, OprF, and type A and
B-flagellins. To assess the functional activity of each of the antigen-specific IgG, it was
first necessary to generate P. aeruginosa mutants lacking one or more of these antigens
(see Table 1 and Material and Methods). P. aeruginosa type-B-flagellin expressing strain
PAO1 was used as the genetic background for the mutants. Each mutant exhibited
growth kinetics that were similar to that of the wild-type strain (data not shown). P.
aeruginosa strains were incubated with immune or control mouse plasma at 4°C for 1
hour and then stained for the presence of IgG. As shown in Fig 6A, wild-type P.
aeruginosa bound significant amounts of IgG specific for flagellin, OprI, and OprF.
Furthermore, mutants positive for only flagellin, OprF, or OprI also bound high levels of
IgG. Experiments using a type-A-flagellin expressing strain, PAK, demonstrated similar
results (data not shown). These results demonstrate that the antibodies generated against
the recombinant fusion protein recognize these antigens in their cell-associated forms.
This is particularly important in the case of OprF, since only a single epitope was present
in the OprF311-341-OprI-flagellin fusion proteins.
Having established the ability of the individual populations of IgG to recognize
the cell-associated antigens, we next evaluated the potential of these antibodies to
activate complement. Previous work has clearly established the
34

Figure 6. Complement activating activity of OprI, OprF, and type A and B-
flagellin-specific IgG. Plasma samples from OprF311-341-OprI-Flagellins were incubated
with P. aeruginosa and IgG binding and C3 deposition was determined by flow
cytometry. The antigens expressed by each P. aeruginosa strain are shown. (A)
Antigen-specific IgG binding to P. aeruginosa. Left column – Plasma samples from
OprI-Flagellins immunized mice. Filled regions refer to control strain WFPA860
(∆fliC∆oprI) which lacks flagellin and OprI. Right column - Plasma samples from OprF-
OprI-Flagellins immunized mice. WT, wild-type; B-Flagellin+, fliC+∆oprI∆oprF;
OprI+, oprI+∆fliC∆oprF; OprF+, oprF+∆fliC∆oprI. Filled regions refer to control
WFPA864 (∆fliC∆oprI∆oprF) strain lacking all three antigens. (B) C3 deposition on P.
aeruginosa strains. (C) Percent C3 positive of the data shown in B. *=P<0.05, ** =
P<0.001, compared to WFPA860 (∆fliC∆oprI) (top) or WFPA864 (∆fliC∆oprI∆oprF)
(bottom). Statistics were performed using Student’s t-test. Data are from 2 independent
experiments performed in triplicate.
35

36

importance of the complement system in the clearance of P. aeruginosa {49-51,120}. To
assess the ability of antibodies specific for OprF, OprI, type A and B-flagellin IgG to
activate complement, we measured the extent of IgG-dependent C3 deposition on P.
aeruginosa. The various P. aeruginosa strains were incubated with a 1:10 dilution of
heat-inactivated immune mouse plasma for 1 hour, and then 5% rabbit complement was
added for an additional hour. The bacteria were then stained with FITC-labelled C3-
specific antibody and the extent of C3-deposition was determined by flow cytometry. A
time course revealed that 1-hour incubation with serum was optimal for C3 deposition
and yielded minimal cell death (data not shown). As a control, we used a P. aeruginosa
strain lacking flagellin, OprI, and OprF. OprI-A- and B-flagellin or OprF311-341-OprI-A-
and B-flagellin immune plasma promoted significant C3 deposition on the surface of
wild-type P. aeruginosa (Fig 6A). By using mutants that lack one or more of the eliciting
antigens, we found that IgG with specificity for each of the eliciting antigens promoted
robust C3 deposition (Fig 6B-C). When all three antigens were present, there was a
synergistic increase in the level of C3 deposition.
These results indicate that OprF311-341-OprI-A- and B-flagellin immunization
generated antigen-specific IgG that exhibited a high-degree of functional activity and that
the combination of flagellins, OprI, and OprF-specific IgG triggered the highest level of
C3 deposition.
Antibody-dependent, complement-mediated killing of P. aeruginosa by OprF311-341-
OprI-A- and B-flagellin immune plasma. In view of the robust ability of OprF, OprI,
and type A and B-flagellin-specific IgG to promote C3 deposition, we next examined the
37

ability of those antibodies to promote complement-mediated killing of P. aeruginosa.
Bacteria were incubated with heat-inactivated immune plasma for 1 hr and then 5%
rabbit complement was added for an additional 4 hr at 37°C. It is important to note that
like wild-type bacteria, the P. aeruginosa mutants were not susceptible to significant non-
specific killing by normal serum (data not shown). Approximately 90% of wild-type,
nonmuciod P. aeruginosa (PAO1, PAK, and 1286) as well as strains expressing B-
flagellin, OprI, or OprF were susceptible to antibody-dependent complement mediated
killing (Fig 7 and Table 2). In contrast, only 18% of mucoid P. aeruginosa (T68933 and
PD0300M) were susceptible to killing (Table 2). This result is not unexpected, given the
presence of a large amount of alginate exopolysaccharide in the mucoid strains that
would likely mask OprI and OprF. In support of this conclusion, we found that a strain
of PAO1 (PDO300NM) deficient in alginate production (and thus nonmucoid) was quite
sensitive to killing (Table 2). In addition, the generally applicable inverse relationship
between flagella and alginate expression {19} would also limit the effectiveness of the
flagellin-specific IgG. The antigen-dependence of the killing was evidenced by the very
low level of killing with bacteria lacking all three of the eliciting antigens. When the
source of complement was heat-inactivated, only background levels of killing were
observed. Taken together, these findings clearly demonstrate that the antibodies
generated in response to OprF311-341-OprI-A- and B-flagellin exhibit potent antigen
binding, complement activating activity, and killing of nonmucoid, but not mucoid P.
aeruginosa.
38

Figure 7. Antibody-dependent complement-mediated killing P. aeruginosa by
OprF311-341-OprI-Flagellins immunized mouse plasma. Plasma samples from OprF311-
341-OprI-Flagellins immunized mice were diluted 1:10 and heat-inactivated at 56°C for 1
hr. Samples were then supplemented with 5% rabbit complement for 4 hr at 37°C. Data
are from 4 samples over two experiments, each sample done in duplicate. * P= <0.05,
using Student’s t-test.
39

40

Table 2. Complement-mediated killing of additional P. aeruginosa strains
Strain Percent Killed
PAK (WT) 86.5 ±0.2%
1286 (nonmucoid CF isolate) 84.3 ±0.8%
PD0300M (mucoid PAO1) 15.2 ±1%
PD0300NM (alginate deficient PDO300) 83.4±0.4%
T68933 (mucoid CF isolate) 24.8 ±0.3%
41

Enhanced clearance of P. aeruginosa in OprF311-341-OprI-Flagellins immunized mice.
Nonmucoid P. aeruginosa does not cause a chronic infection in healthy mice as it does in
CF patients. If large doses of bacteria are used, the mice quickly succumb to bacteremia
(Weimer, Wozniak, and Mizel, unpublished observations). With small doses, the mice
rapidly clear the bacteria. In view of the lack of a suitable animal model that closely
mimics the situation in CF patients, i.e., chronic infection, investigators have evaluated
agar-embedded mucoid P. aeruginosa as a way to infect mice such that rapid septic
shock is avoided and the time of infection is lengthened {34,170}. For example,
Stevenson and colleagues {32,34} used the agar bead model to demonstrate that DBA/2
mice were more susceptible to mucoid P. aeruginosa than were BALB/c or C57BL/6
mice. However, since the initial P. aeruginosa infection in CF patients is mediated by
nonmucoid strains {22,23,25}, we felt it was more appropriate to use nonmucoid bacteria
in the agar bead model. Preliminary results revealed that unimmunized mice did not
succumb when infected intra-tracheally with up to 3.5 x 106 cfu of nonmucoid P.
aeruginosa embedded in agar beads, but the mice did exhibit substantial morbidity. In
view of the finding that DBA/2 mice are more susceptible to P. aeruginosa, we used this
strain to evaluate the ability of OprF311-341-OprI-Flagellins immunization to promote
enhanced clearance of P. aeruginosa embedded in agar. DBA/2 mice were immunized as
described above and infected intratracheally with 3.5x106 cfu agar-embedded PAO1.
Lungs were harvested 1, 3, and 5 days post-infection and bacteria enumerated by serial
dilutions on LANS plates. One day after challenge, immunized mice displayed a marked
decrease in bacterial burden compared to control mice (Fig 8). After 3 days, 5 of 6 mice
immunized with OprF311-341-OprI-Flagellins had cleared the infection. In contrast, the
42

control mice had large numbers of bacteria in the lungs. Although the control mice
cleared the infection by day 5, our results clearly demonstrate that immunization with
OprF311-341-OprI-Flagellins had a dramatic effect on the rate of bacterial clearance. It is
important to emphasize that the ability of mice immunized with OprF-OprI to clear the
infection by day 5 reflects a limitation of this model and not the efficacy of the OprF311-
341-OprI-Flagellins vaccine.
Reduced lung pathology following pulmonary P. aeruginosa challenge in OprF311-341-
OprI-Flagellin immunized mice. In addition to determining bacterial burden following
challenge, we also evaluated the histopathology of lungs from mice immunized with
OprF311-341-OprI-Flagellin or OprF-OprI. Lungs were harvested 1, 3, and 5 days after P.
aeruginosa challenge. One day after P. aeruginosa challenge, alveolar walls from
OprF311-341-OprI-Flagellin immunized mice displayed slight thickening owing to
congestion and increased numbers of inflammatory cells. In contrast, lungs from mice
immunized with OprF-OprI developed bronchopneumonia, with airway-oriented
neutrophils, edema, and abundant visible bacteria (Fig 9A). After 3 days, immune mice
exhibited only minor inflammatory changes in the lung, whereas more severe pneumonia
with diffuse consolidation was present in the control animals. After 5 days, the lungs of
immune mice were normal, while those of the controls had thickened alveolar walls, a
result of congestion and inflammatory cells (Fig 9B). In summary, mice immunized with
OprF311-341-OprI-Flagellin displayed minimal lung pathology which completely resolved
by day 5 post challenge. The absence of lung pathology in the immune mice not only
43

Figure 8. OprF311-341-OprI-Flagellins immunized mice display enhanced rate of
clearance following pulmonary P. aeruginosa challenge. DBA/2 mice were
immunized twice with 5µg of OprF311-341-OprI-Flagellins and challenged intratracheally
with 3.5x106 cfu of agar-embedded PAO1. The right lungs were harvested 1, 3, and 5
days post-infection and bacterial burden was assessed by counting. Data are the average
of 6-7 mice per group. Dotted line indicates the limit of detection. #P=0.053,
**P=0.002, using a Mann–Whitney rank sum test.
44

45

demonstrates the efficacy of the vaccine in promoting bacterial clearance, but also the
ability of the vaccine to promote clearance without inducing secondary tissue damage. In
striking contrast, mice immunized with OprF-OprI demonstrated severe pneumonia
which only partially resolved by day 5 (Fig 9A). In conjunction with the results of in
vitro experiments (Figs 5 and 6), it is clear that OprF311-341-OprI-Flagellin immunization
promotes an adaptive immune response that promotes the generation of antigen-specific
IgG that exhibits robust functional activity, facilitates rapid clearance, and prevents the
development of severe pneumonia following P. aeruginosa infection.
46

Figure 9. OprF311-341-OprI-Flagellins immunized mice are protected against severe
lung pathology during pulmonary P. aeruginosa challenge. The left lung of identical
mice used in Figure 8 was evaluated for histology. Lungs were fixed in 10% formalin for
24 hours, paraffin embedded, and 4µm sections were cut. Slides were stained with H&E.
Representative images are shown from 6 sections/animal. (A) OprF-OprI immunized
mice. A1-3 magnified 4x. A4 magnified 40x to show bacteria. A5-6 magnified 20x (B)
OprF311-341-OprI-Flagellins immunized mice. B1-3 magnified 4x. B4-6 magnified 20x.
(C) Slides were blindly scored for consolidation, bronchiolar and vascular degenerative
changes, alveolar wall thickness, and edema. The score for inflammation was determined
by the sum for each category. Dotted line indicates lowest score possible. *P<0.05,
using Student’s t-test.
47

48

Chapter II
Immunization of young African green monkeys with OprF epitope 8-OprI-type A- and B-
flagellin fusion proteins promotes the production of protective antibodies against
nonmucoid Pseudomonas aeruginosa
E.T. Weimer, S.E. Ervin, D.J. Wozniak, S.B. Mizel
The following manuscript was submitted to Vaccine, May 27, 2009. Stylistic variations
are due to the requirement of the journal. E.T. Weimer performed the experiments and
prepared the manuscript. Dr. Steven B. Mizel acted in an advisory and editorial capacity.
49

MATERIALS AND METHODS
Strains and plasmids. Escherichia coli was cultured in Luria-Bertaini (LB; 10
g/L tryptone, 5 g/L yeast extract, 5 g/L NaCl) broth at 37oC and P. aeruginosa in LB
broth lacking NaCl (LBNS; 10 g/L tryptone, 5 g/L yeast extract). Solid media was
prepared by adding 1.0-1.5% select agar (Gibco-BRL). Plasmid selection in E. coli was
done using media supplemented with antibiotics at the following concentrations:
carbenicillin (Cb; 100 µg/ml), gentamicin (Gm; 10 µg/ml) and in P. aeruginosa using
media containing Cb (300 µg/ml), Gm (100 µg/ml), and irgasan (Irg; 25 µg/ml). E. coli
strain JM109 was used for all cloning procedures. E. coli SM10 was used to transfer
plasmids into P. aeruginosa by bi-parental mating {160}. The P. aeruginosa strains used
were PAO1 and its derivatives: WFPA850, WFPA852, WFPA854, WFPA860,
WFPA862, WFPA864, and WFPA866. These non-polar deletion mutants were
constructed by overlap extension as previously described {115}.
Recombinant proteins. OprF311-341-OprI-Flagellins were purified by metal ion
affinity chromatography as previously described {61,115,165}. Acrodisc Q membranes
were used to remove endotoxin and nucleic acids. Endotoxin levels were <10pg/µg for
all of the proteins (as detected by QCL-1000 chromogenic LAL test kit, Cambrex
Corporation (East Rutherfrod, NJ)).
ELISA for antigen-specific IgG. Titers of antigen-specific IgG were measured
using MaxiSorb plates coated with 100µl of purified recombinant antigen (A-flagellin, B-
flagellin, OprI, or OprF) at 10 µg/ml in sterile PBS. The plates were incubated overnight
at 4°C and then blocked with 10% newborn calf serum in PBS. Plasma samples (in
triplicate) were added and the plates incubated overnight at 4°C. Secondary anti-Ig
50

antibodies (Roche Diagnostics) were then added for 2h at room temperature. Peroxidase
activity was detected with 3,3’,5,5’-tetramethylbenzidine (TMB) liquid substrate system
(Sigma-Aldrich). Endpoint dilution titers were defined as the inverse of the lowest
dilution that resulted in an absorbance value (at 450 nm) of 0.1 over that of pre-
immunization plasma. To determine relative antibody affinities, the ELISA assay was
conducted as described above with the addition of a 15 min incubation with sodium
thiocyanate (NaSCN) (Sigma) solution as described previously {115,165,166}.
Monkeys. Immunization studies with 4-6 month old African green monkeys
(Chlorocebus aethiops SK) (approximately 1 kg) were conducted at the Behavioral
Science Foundation, St. Kitts, using a protocol approved by the foundation-associated
Animal Care and Use Committee. The animals were bled several days prior to
immunization to obtain control plasma. Groups of 7 monkeys were immunized with 1, 3,
10, or 30µg of OprF311-341-OprI-Flagellins (in a volume of approximately 160 µl) in the
gluteus maximus muscle on two occasions separated by 28 days. Ten days and three
months after the boost, blood was collected and plasma was prepared for analysis of
antigen-specific IgG titers.
Antigen-specific IgG-mediated complement activation. Antibody-specific
complement activation was determined using flow cytometry as previously described
{115}. Complement-mediated killing was determined by incubating bacteria with serial
dilutions of plasma samples for 4 hr and then plating aliquots {115}. Percent bacteria
killed was quantified by [(number of input bacteria – number of recovered bacteria) /
(number of input bacteria)] X 100.
51

Passive immunization and respiratory challenge with agar-embedded P.
aeruginosa. Groups of 5-7 week old female DBA/2 mice (6 mice per group) were given
100µL of pre-immunization or post-immunization monkey plasma intra-peritoneally and
then two days later were challenged intratracheally with 2x106 colony forming units (cfu)
of agar-embedded PAO1 {34,115-117,171}. The agar-embedded PA01 were prepared
and given intratracheally as previously described {115}.
Histology. Lungs were harvested and placed in 10% formalin for 24 h. The
tissue then was trimmed, embedded in paraffin, cut at 4 µm, and stained with
hematoxylin and eosin by routine methods. Three sections per lung from each of 4 mice
in control and immune groups were evaluated. Representative images are presented.
Statistical analyses. Statistical analysis was performed using GraphPad Prism
5.02 for Windows (GraphPad Software, San Diego California USA). The significance of
data sets was determined using the Mann–Whitney rank sum test. Where applicable a
one-way ANOVA test was applied. P values of less than 0.05 were considered
significant.
52

RESULTS
OprF311-341-OprI-Flagellins promote a robust humoral immune response in young
African green monkeys. In view of our previous study indicating that OprF311-341-OprI-
Flagellins promote the production of protective IgG in mice {115}, we evaluated the
ability of OprF311-341-OprI-Flagellins to elicit a humoral immune response in 4-6 month
old African green monkeys. We chose monkeys at this age because we favor the view
that protection against P. aeruginosa in CF patients will be maximal if immunity is
established at an early age prior to P. aeruginosa infection. To that end, groups of seven
4-6 month old African green monkeys (~ 1 kg) were immunized intramuscularly with 1,
3, 10, or 30µg of total protein (equal amounts of each fusion protein). Four weeks later,
the monkeys were boosted via the same route, plasma was prepared ten days post-boost,
and evaluated by ELISA for titers of anti-OprF, OprI, and type A and B flagellin IgG.
Pre-immunization plasma from each animal was used as a control. The results using the
30µg dose are shown in Figure 10. As might be expected, the flagellin-specific IgG
responses were extremely strong. However, the magnitude of the OprI and OprF-specific
IgG titers were also quite high, generally exceeding 105. Enhanced antigen-specific IgG
production was also observed in monkeys immunized with the 1, 3, and 10 µg doses (data
not shown). Although some of the monkeys had low pre-immunization titers against
each antigen, immunization with OprF311-341-OprI-Flagellins induced a dramatic increase
in the plasma IgG titers against all of the eliciting antigens (Fig 10). Importantly, the
induced IgG was not due to reactivation of memory B cells, as immunization with OprF-
OprI fusion protein, without flagellin, did not elicit any increase in plasma IgG (data not
shown).
53

To determine the stability of antigen-specific IgG following immunization with
OprF311-341-OprI-Flagellins, 4-6 month old African green monkeys were immunized as
described above. Three months post-boost, plasma was prepared and antigen-specific
IgG was determined by ELISA. Pre-immunization plasma was used as a control.
Surprisingly, there was a substantial reduction in the level of IgG specific for flagellin,
OprI and OprF (Fig 10). However, the decrease in antigen-specific IgG was not from
reduced stability of the plasma IgG as the calculated half-life was 26.9 days (data not
shown). Reduced antigen-specific IgG production was also observed in monkeys
immunized with the 1, 3, and 10 µg doses (data not shown). Based on nonlinear
regression analysis, the animals possessed protective levels of IgG out to 2.5 months after
immunization (Fig 10B). Thus, ten days post-immunization plasma was used for all
subsequent experiments.
OprF311-341-OprI-Flagellins elicit high affinity IgG against flagellin, OprI, and OprF.
Given that antigen affinity plays a critical role in the overall functional activity of an
antibody, we evaluated the relative affinity of the IgG generated following immunization
with OprF311-341-OprI-A- and B-flagellin. The relative affinity of antibodies was assessed
by determining the concentration of sodium thiocyanate required to reduce antibody
binding by 50% in an ELISA. Samples were pooled from the 30µg pre- and ten days
post-immunizations.
54

Figure 10. Intramuscular immunization of African green monkeys with OprF311-341-
OprI-Flagellins promotes a potent humoral immune response. African green
monkeys were immunized twice intramuscularly with 30µg of OprF311-341-OprI-
Flagellins. Ten-days and three months post-boost, plasma was prepared and ELISA used
to determine antigen-specific IgG titers. (A) Open circles represent the pre-immunization
titers for each animal. Filled squares represent individual monkeys ten days post-
immunization IgG titer. Filled triangles represent individual monkeys three months post-
immunization (7 monkeys per group in triplicate). (B) Average nonlinear regression
analysis on IgG titers from individual moneys immunized with 30µg of OprF311-341-OprI-
Flagellins. Using one-way ANOVA analysis, post-immunization IgG titers were
significantly higher than pre-immunization IgG titers (P<0.05).
55

56

For comparative purposes, in mice immunized with OprF311-341-OprI-Flagellins 3M
NaSCN was required to reduce immune complex formation by 50% for type A and B
flagellins and 2M for OprI and OprF {115}. Given the observation that these antibodies
provide enhanced clearance of P. aeruginosa, we have classified high affinity antibodies
as requiring 2-3 M sodium thiocyanate for 50% reduction in antigen binding. As shown
in Fig 11, immunization with OprF311-341-OprI-Flagellins generated IgG with relatively
high-affinity for flagellin, OprI, and OprF. OprF311-341-OprI-A-and B-flagellin immune
plasma had an average IgG affinity approaching 3M sodium thiocyanate for A- and B-
flagellin and OprI, and 2M for OprF (Fig 11). In contrast, pre-immunization IgG
exhibited significantly lower affinities than post-immunization IgG. For example, pre-
immunization anti-OprI IgG had a relative affinity of 600 mM as opposed to 3M for
immunization-induced anti-OprI IgG (Fig. 11A). The presence of high affinity anti-A-
flagellin IgG prior to immunization is most likely a result of exposure to environmental
P. aeruginosa. However, immunization with OprF311-341-OprI-Flagellins significantly
enhanced the affinity of A-flagellin-specific IgG (Fig 11). These results are consistent
with the conclusion that OprF311-341-OprI-A- and B-flagellin elicits high-affinity, antigen-
specific IgG.
57

Figure 11. OprF311-341-OprI-Flagellins elicit high-affinity antigen-specific IgG.
Pooled plasma samples from monkeys that received 30µg of OprF311-341-OprI-Flagellins
were used to determine relative antibody affinity for A-flagellin, B-flagellin, OprI, and
OprF. Antigen-specific IgG affinity was determined by ELISA using dilutions of sodium
thiocyanate (NaSCN). Data are presented as percent maximal absorbance relative to
PBS. Dotted line indicates the 50% maximal absorbance, which is an indicator of
antibody affinity. Samples were obtained from the monkeys used in Fig 10.
58

59

IgG specific for flagellin, OprI, and OprF mediate complement activation and killing
of nonmucoid P. aeruginosa. To assess the functional activity of each of the antigen-
specific IgG, we utilized several P. aeruginosa mutants lacking one or more of the
eliciting antigens {115}. P. aeruginosa type-B-flagellin expressing strain PAO1 was
used as the genetic background for the mutants. Using these mutant strains, we evaluated
the ability of flagellin, OprI, and OprF-specific IgG antibodies to individually mediate C3
deposition on P. aeruginosa. The various P. aeruginosa strains were incubated with
dilutions of heat-inactivated pooled immune monkey plasma for 1 hour, and then 5%
rabbit complement was added for an additional hour. The bacteria were then stained with
FITC-labelled C3-specific antibody and the extent of C3-deposition was determined by
flow cytometry. A mutant P. aeruginosa strain that was unable to synthesize flagellin,
OprI, and OprF was used as a control. Consistent with previous observations in mice
{115}, antibodies specific for flagellin, OprI, and OprF mediated high levels of
complement deposition on wild-type P. aeruginosa strain PAO1 (Fig. 12A). The
magnitude of the effect was dependent on the concentration of immune plasma and was
detectable at a dilution as low as 1:1,000 with wild-type PAO1 as the target. The antigen
specificity of the effect was evidenced by the ability of immune plasma to promote
significantly greater C3 deposition on mutant PAO1 strains expressing only one of the
eliciting antigens than on bacteria lacking flagellin, OprI, and OprF expression (shaded
peaks in Fig. 12A). The benefit of having a multivalent vaccine containing flagellin,
OprI, and OprF is evidenced by the observation that C3 deposition on
60

Figure 12. Complement activation and complement-mediated killing by OprI,
OprF, and type A and B-flagellin-specific IgG. Pooled plasma samples from the
monkeys immunized with 30 μg OprF311-341-OprI-Flagellins were incubated with wild
type and mutant P. aeruginosa PAO1. (A) C3-depostion was determined by flow
cytometry after bacteria had been incubated with plasma for 1 hr and rabbit serum for an
additional hr. WT, wild-type; B-Flagellin+, fliC+∆oprI∆oprF; OprI+, oprI+∆fliC∆oprF;
OprF+, oprF+∆fliC∆oprI. The shaded peak was obtained using the WFPA864 strain
(∆fliC∆oprI∆oprF) lacking all three antigens. (B) 1:10, 1:100, 1:1000, and 1:10,000
dilutions of plasma samples were incubated with PAO1 (WT) for 4 hrs and complement-
dependent killing was determined by plating and enumerating bacteria. (C) Mutant
strains of P. aeruginosa were used to determine the ability of flagellin, OprI, and OprF-
specific IgG to mediate complement-dependent killing of the bacteria. In each case, a
1:100 dilution of plasma was used. (D) Effect of immune plasma on PAO1, as well as
nonmucoid (1286) and mucoid (T68933) P. aeruginosa clinical isolates. Data represents
3 independent experiments performed in triplicate. * P<0.05, compared to WFPA864
(∆fliC∆oprI∆oprF) strain lacking all three antigens.
61
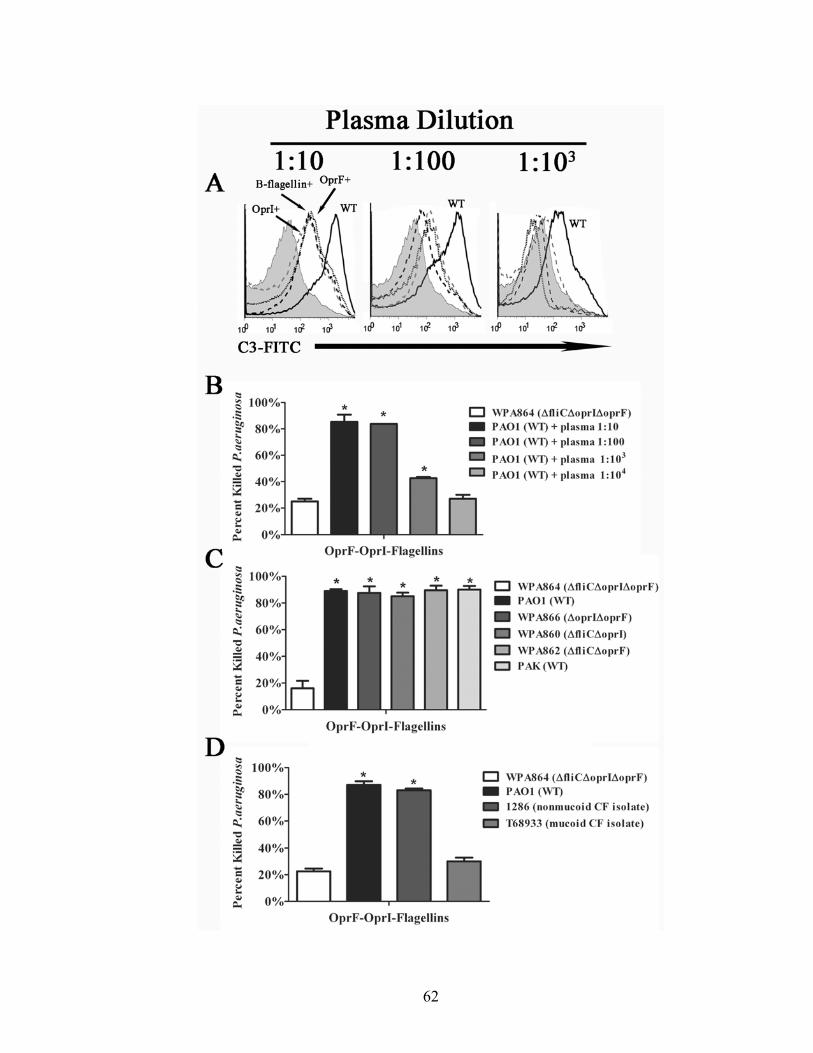
62

PAO1 expressing all three antigens was still measurable at a 1:1,000 dilution, whereas C3
deposition was only minimally evident at this dilution when individual PAO1 mutants
were used—a finding that is consistent with the notion that the individual populations of
anti-type A and B flagellin, OprI, and OprF synergize in promoting C3 deposition.
In view of the robust complement activation mediated by immune monkey
plasma, we assessed whether the plasma was equally effective in promoting complement-
mediated killing of P. aeruginosa. Aliquots of wild-type PAO1 were incubated for 4 hr
with varying dilutions of heat-inactivated pooled immune plasma and then supplemented
with rabbit serum as a source of complement prior to plating {115}. To control for non-
specific effects, WPA864, a mutant strain lacking type B flagellin, OprF, and OprI
expression was also evaluated. As shown in Fig 12B, pooled immune plasma promoted
high level, concentration-dependent killing of wild-type P. aeruginosa PAO1. As
expected, the low level of killing observed with WPA864 was similar to that observed in
the absence of complement (Table 2) or pre-immunization plasma (data not shown) and
thus demonstrates that the observed effect of the pooled immune monkey plasma is
antigen-specific. Consistent with the ability of anti-flagellin, OprI and OprF IgG to
individually induce C3 deposition, high level complement-mediated killing was observed
when the PAO1 mutants expressing only one of the eliciting antigens were used as targets
(Fig. 12C).
In a prior study {115}, we found that plasma from mice immunized with OprF311-
341-OprI-Flagellins promoted robust complement-mediated killing of nonmucoid, but not
mucoid strains of P. aeruginosa. The diminished ability of immune plasma to kill
mucoid strains is most likely due to the absence of flagella on mucoid P. aeruginosa
63

{172} and interference of antibody and/or complement deposition by alginate
exopolysaccharide. To determine if pooled immune plasma from monkeys also exhibits
activity against non-mucoid, but not mucoid strains of P. aeruginosa, we incubated wild-
type PAO1, strain 1286, a nonmucoid CF isolate, strain T68933, a mucoid CF isolate,
and PAO1 WPA864 (∆fliC∆oprI∆oprF) with a 1:100 dilution of pooled immune monkey
plasma for 4 hr prior to addition of rabbit serum as a source of complement. Aliquots of
the bacteria were then plated and surviving bacteria were enumerated. As shown in Fig
12D, the pooled immune plasma promoted high level killing of nonmucoid wild-type
PAO1 and 1286 clinical isolate, but had little if any effect on the mucoid isolate T68933.
Passive immunization of mice with plasma from OprF311-341-OprI-Flagellins
immunized monkeys enhances clearance of P. aeruginosa following pulmonary
challenge. To test the ability of antibodies from immunized monkeys to mediate
clearance of nonmucoid P. aeruginosa, we passively immunized mice with pooled pre-
immunization or immune monkey plasma. Due to the development of a lethal systemic
Arthus –like reaction in DBA/2 mice given a volume of monkey plasma in excess of 100
µl (unpublished observations), we were forced to limit the passive immunization volume
to 100 µl. Using this volume of monkey plasma and intra-peritoneal injection, there was
no observable morbidity or mortality in any of the mice. Therefore, groups of 5-6 female
DBA/2 mice were given 100 µl of pooled pre-immunization or post-immunization
plasma intra-peritoneally and then after two days the mice were challenged
intratracheally with 2x106 cfu of agar-embedded PAO1 {115}. Animals were sacrificed
three days post-infection and one lung was harvested for bacterial enumeration and the
64

other for hematoxylin and eosin (H & E) staining. As shown in Figure 13A, mice that
received immune plasma had approximately two-log lower numbers of bacteria than mice
that received pre-immunization plasma. Indeed, two of the mice that received immune
plasma had no detectable bacteria.
Since some of the mice that received immune monkey plasma had no detectable
bacteria in their lungs, we considered the possibility that these mice may have had higher
circulating IgG of the transferred immune monkey plasma as a result of differences in the
kinetics of distribution of the monkey plasma during the 48 hr after intraperitoneal
injection. Thus, some mice were better able to clear the bacteria following challenge. To
evaluate this possibility, we prepared plasma at the time of sacrifice (day 3 after
challenge) from the same mice shown in Fig 13A determined the titers of circulating
antigen-specific monkey IgG. The results of this analysis are presented in Figure 13B.
Recipient mice had type A and B flagellin-specific IgG titers approaching 105, whereas
OprI and OprF-specific IgG titers were in the range of 1-4 x 104. The IgG titers represent
a 30-fold dilution of the transferred plasma and are consistent with a blood volume of
approximately 3 ml. In contrast, mice that received pre- immunization plasma had
antigen-specific IgG titers below the limit of detection (data not shown). Importantly, the
level of total antigen-specific IgG (sum of flagellin, OprI, and OprF-specific IgG) three
days post-infection inversely correlated (r2 = 0.89) with the bacterial load in a given
animal. Based on these results, it is quite likely that all of the mice receiving immune
plasma would have had undetectable bacterial loads if their antigen-specific monkey IgG
titers had been above 4-5 x 105—the equivalent of undiluted immune monkey plasma
(Fig. 13B, right panel).
65

Figure 13. Passive immunization of mice with immune monkey plasma enhances
clearance of nonmucoid P. aeruginosa. Mice were passively immunized
intraperitoneally with 100µL of pre- or post-immunization monkey plasma. Two days
post-injection mice were intratracheally challenged with agar-embedded PAO1 [16].
Three days post-infection, the mice were sacrificed and lungs harvested for bacterial
counts and H+E staining. (A) Lung bacterial burden three days post-infection. * P<0.05.
(B) Flagellin, OprI, and OprF-specific IgG titers three days post-infection were
determined by ELISA. The sum of flagellin, OprI, and OprF-specific IgG (Total antigen-
specific IgG) inversely correlates with bacterial burden r2=0.89. (C) 4µm thick lung
sections were stained with H+E. Left panel is representative of mice that received pre-
immunization plasma. Center panel is representative of mice that received post-
immunization plasma that had >102 bacteria. Right panel is representative of mice that
received post-immunization plasma with undetectable (<102) bacteria.
66

67

These findings are consistent with the conclusion that the titers of anti-flagellins, OprI,
and OprF in the immune monkeys may be of sufficient magnitude to promote enhanced
clearance of P. aeruginosa in these animals.
Mice immunized with OprF311-341-OprI-Flagellins are spared from the serious
inflammation generated in response to respiratory infection with P. aeruginosa {115}.
To determine if mice receiving pooled immune monkey plasma are also spared from
serious lung inflammation following challenge with agar-embedded P. aeruginosa
PAO1, H & E lung sections from mice receiving pre-immunization or post-immunization
monkey plasma were examined (Fig. 13C). As might be expected from the variability in
the bacterial load in lungs (Fig. 13A) and circulating titers of antigen-specific IgG (Fig.
13B) mice that received immune monkey plasma displayed variable levels of lung
inflammation 3 days post-challenge. The mice that received immune monkey plasma and
had sufficient circulating titers of monkey IgG to rapidly and completely clear the
infection had no signs of lung pathology (Fig. 13C right panel), whereas mice that had
detectable bacteria had signs of lung inflammation (Fig. 13C middle panel). However, it
is important to emphasize that the extent of lung inflammation in these mice was
significantly less than observed in mice that received pre-immunization plasma (Fig. 13C
left panel). The latter group of mice clearly had severe bronchial pneumonia that was
characterized by consolidation, alveolar wall thickening, and edema.
68

DISCUSSION
The goal of my thesis studies was two-fold: to establish a set of criteria for a
vaccine against P. aeruginosa and then to develop and test a vaccine based on these
criteria in adult mice as well as very young African green monkeys. Based on my results,
I conclude that the OprF311-341-OprI-A- and B-flagellins vaccine meets all of the proposed
criteria: the vaccine contains flagellin, a potent adjuvant, is multivalent, generates high-
titer antigen-specific IgG that exhibits a high degree of functional activity, generates a
robust memory response, and enhances clearance of nonmucoid P. aeruginosa without
secondary tissue damage. In addition, OprF311-341-OprI-A- and B-flagellins vaccine
promotes a humoral immune response in 4-6 month old monkeys that induces not only a
high level of antigen-specific IgG but also high affinity IgG that mediate complement-
dependent killing of nonmucoid P. aeruginosa. Although the antigen-specific IgG
induced by this vaccine did not promote complement-mediated killing of mucoid P.
aeruginosa (Table 2), it is important to emphasize that longitudinal studies of CF patients
have clearly demonstrated that the initial P. aeruginosa infection is mediated by
nonmucoid bacteria {22,23,25}. The overexpression of alginate on mucoid P.
aeruginosa may prevent antibodies from binding to antigen or inhibit the deposition on
C3 on the surface of the bacteria. The latter hypothesis is consistent with the
observations that S. pneumoniae capsule polysaccharide prevents the Fc region of
antibodies from binding to Fc receptors on phagocytic cells as well as reduces the amount
of C3 deposited on the surface of the bacteria {173,174}. I wish to emphasize that
although the OprF311-341-OprI-Flagellins has many of the features of an efficacious
69

vaccine, the evaluation of its potential for use in CF patients is limited at this time by the
lack of a suitable animal model that closely mirrors the situation in CF patients.
In addition to promoting high-level antigen-specific IgG production (Fig 3 and
10), flagellin facilitated the generation of significant numbers of antigen-specific memory
B cells (Fig 4). In view of these potent actions of flagellin, it is likely that the very
limited reduction in the incidence of P. aeruginosa infections in CF patients immunized
with sheared flagella may be due to a low level of functional adjuvant activity of the
material {128}. Although the adjuvants Pam3Cys and Pam2Ser {175} , alum {81,176},
adenovirus {116,117}, activated DC {171}, and B-cell activating factor {177} were
found to enhance the response to P. aeruginosa antigens, it appears that flagellin is far
more effective as evidenced by the dramatic difference in the amount of antigen required
and the resultant titers of antigen-specific IgG.
Multivalency not only promotes synergistic activity of individual antibodies in
activating complement but also enhances vaccine coverage against nonmucoid P.
aeruginosa strains (Fig 6). The value of multivalency was also demonstrated by Saha et.
al. {106} who used multivalent DNA vaccination with OprF-OprI, PilA, and PcrV
antigens.
The titers of flagellins, OprI, and OprF-specific IgG following immunization with
OprF311-341-OprI-Flagellins are in most cases two logs higher than those reported in other
studies {81,103,106,116,117,128,145,178}. For example, von Specht et. al. {179}
required three immunizations with 70-fold more antigen to achieve equivalent antibody
responses. In addition, Sorichter et. al. {124} reported that intranasal immunization of
CF patients with 1mg of OprF-OprI fusion protein promoted a 3-fold increase in IgG
70

titers. The difference may be due to the extraordinary potency of flagellin as an adjuvant,
as well as the use of fusion proteins that enhance the efficiency of antigen delivery to
dendritic cells via the binding of the associated flagellin to TLR5 on these cells {157}.
The finding that immunization with OprF311-341-OprI-Flagellins promotes the generation
of large numbers of plasma cells is consistent with the very high titers of induced IgG.
Furthermore, the generation of immunologic memory is evidenced by the relatively high
frequency of antigen-specific MBC.
In line with prior work on flagellin, OprF, and OprI antigens, I have shown that
all three antigens significantly affect the clearance of P. aeruginosa in mice. In addition,
antibodies specific for each antigen activate complement and mediate killing of P.
aeruginosa (Figs 6, 7). In this regard, Von Specht and colleagues {120} found that the
level of complement-activating OprI-specific IgG correlated with the level of protection
against P. aeruginosa. In addition, several investigators have demonstrated that high-
affinity IgG that exhibits a high degree of functional activity can mediate protection
against P. aeruginosa {80,180}. My results clearly support these conclusions. The
finding that intramuscular immunization with OprF311-341-OprI-Flagellins promotes
clearance is consistent with the conclusion that protection is not dependent on the
availability of antigen-specific IgA. In this regard, Pier et al. {181} demonstrated using
LPS-based vaccination that intraperitoneal immunization could prevent mucosal
colonization by P. aeruginosa.
A number of studies {182-185} have presented evidence in support of the
conclusion that heightened production of Th2 cell-derived cytokines such as IL-4, IL-10,
and IL-13 is associated with a poor prognosis in CF patients. In view of the notion that
71

isotype switching to IgG1 is Th2 cell-driven, the question arises as to whether the
observed IgG1 bias (relative to IgG2a) of the humoral response to OprF311-341-OprI-
Flagellins might exacerbate the pathology in CF patients. First, the linkage between Th
bias and prognosis in CF patients is based on cytokine production and not IgG isotype.
Indeed, two studies revealed that elevated levels of alginate-specific IgG2 and IgG3, and
not IgG1, are associated with a poor prognosis in CF patients {186,187}. It is also
important to note that class switching to IgE is also promoted by Th2 cells. Yet we have
never observed any increase in IgE in response to OprF311-341-OprI-Flagellins or any
flagellin-based vaccine {61}. Thus, we believe that the humoral response driven by the
adjuvant activity of flagellin does not fit the paradigm of a classic Th2 response and thus
the possibility that a flagellin-based P. aeruginosa vaccine might cause substantial
secondary tissue damage is unlikely. This conclusion is clearly supported by the
observation (Fig 9) that the lungs of mice immunized with OprF311-341-OprI-Flagellins did
not exhibit any evidence of residual tissue damage following clearance of the bacteria.
In contrast to the protective, inflammation-sparing effect of immunization with
OprF311-341-OprI-Flagellins, P. aeruginosa infection in CF patients generates a humoral
immune response against P. aeruginosa antigens such as LPS, exotoxin A, and alginate
that is ineffective in promoting clearance of the bacteria. High levels of P. aeruginosa-
specific IgG have been associated with increased inflammation and lung damage most
likely from increased immune complex formation {23,188,189}. The P. aeruginosa-
specific antibodies exhibit mixed affinity or low functional activity {25,190}, in part to
the fragmentation of these antibodies by released elastase{191}. Importantly, Tosi et. al.
{25} found that the opsonic activity declined as chronic P. aeruginosa infection was
72

established. The generation of low affinity antibodies during chronic P. aeruginosa
infection reduces the ability of CF patients to clear immune complexes and results in
increased inflammation {192}. In addition, IgG isolated from CF patients airway is
fragmented, explaining why the high levels of P. aeruginosa IgG exhibit low functional
activity. The fragmentation of the IgG was directly correlated with the elastase levels
{191}. Since CF patients have a progressive increase in elastase concentrations,
vaccination at an early age would circumvent fragmentation of the antibodies by elastase.
In contrast to infection elicited IgG, vaccine-induced antibodies generally display
a high affinity. For example, Bruderer et. al. {193} demonstrated that vaccination
promoted a 100-fold increase in antibody affinity compared to naturally acquired
antibodies of the same isotype. Given the chronic nature of P. aeruginosa infections, it is
quite likely that the B and T cells undergo “exhaustion”. Constant antigen stimulation in
T cells leads to reduced proliferation, differentiation, and function {194,195}. Thus, the
possible lack of sufficient CD4+ T cell help may explain the reduced antibody affinity
seen in CF patients. Alternatively, the inability of CF patients to generate a high quality
IgG response following P. aeruginosa infection may be due to a combination of several
factors, most importantly virulence factors expressed by P. aeruginosa and the abnormal
respiratory environment.
Although OprF311-341-OprI-A- and B-flagellin elicits a humoral immune response
that promotes rapid clearance of bacteria, it remains to be determined if this vaccine will
be efficacious in CF patients. The pathophysiologic events that occur in CF patients as
well as the virulence factors expressed by P. aeruginosa {196} clearly represent a
significant challenge for any vaccine. The presence of a defective CFTR leads to
73

abnormal ion transport and water depletion in the airway of CF patients that ultimately
results in an increase in fluid viscosity, impaired mucociliary clearance, increased
bacterial trapping in the mucus layer facilitating chronic infection, and a reduction in
antibody penetration {33}. In addition, infection by S. aureus and H. influenzae, and
later P. aeruginosa contribute to the development of a hyper-inflammatory environment
in the respiratory tract {13}. All of these events contribute to a vicious cycle of infection
and inflammation that ultimately causes severe pathology. Chronic inflammation can
adversely affect vaccine efficacy due to the high level production of prostaglandins that
exert an inhibitory effect on lymphocytes {197}. In addition, chronic inflammation
promotes the continual recruitment of neutrophils to the lung that in turn release elastase
that induces tissue damage and release of DNA—a substance that provides a scaffold for
P. aeruginosa biofilms {29,30}.
The major cause of chronic inflammation in CF patients is persistent P.
aeruginosa infection. Over-production of alginate and subsequent, biofilm development
allows P. aeruginosa to evade the immune system and also increases antibiotic
resistance, making it extremely difficult to eradicate the infection {22,23}. Mucoid
conversion is also associated with a significant decrease in lung function in CF patients
{198}. Our findings indicate the best time to vaccinate CF patients using OprF311-341-
OprI-A- and B-flagellins would be prior to mucoid conversion, since complement-
mediated killing occurred only in nonmucoid bacteria (Fig 7, 12, and Table 2).
The pathophysiologic events associated with CF in combination with the
pathogenic mechanisms associated with P. aeruginosa represent a significant challenge
to the elements of protective immunity. Thus, it is important to evaluate the efficacy of
74

OprF311-341-OprI-Flagellins in a hyper-inflammatory environment that closely mimics CF.
To that end, studies have been initiated in the laboratory of Dr. Steven Mizel using CFTR
KO mice (on a C57BL/6 background) to assess the ability of the vaccine to elicited a
protective antibody response in the lungs of mice undergoing progressive damage caused
by excessive inflammation. C57BL/6 CFTR KO mice display increased susceptibility to
P. aeruginosa as well as a progressive lung disease that resembles CF {13,199}.
However, no P. aeruginosa vaccine has been evaluated in these animals. I hypothesize
that there is an inverse correlation between vaccine efficacy and age in the C57BL/6
CFTR KO mice. It is quite likely that vaccine efficacy will be most effective at a very
early stage in the disease process. Perhaps the limited success of previous vaccine
studies was not solely due to the limited inherent efficacy of the vaccines, but rather to
the timing of immunization. In addition, previous clinical trials focused primarily on the
level of antigen-specific IgG induced, instead of the quality or functional activity of the
IgG {115,120,121,158,159}.
Given that the timing of immunization and the quality of the immune response
play important roles in determining vaccine efficacy, I tested OprF311-341-OprI-Flagellins
in 4-6 month old African green monkeys. In line with my observations in mice, I found
that the vaccine elicited a high level of antigen-specific IgG at low antigen doses (Fig
10). Consistent with a previous report {61}, the presence of pre-existing flagellin IgG
does not affect vaccines efficacy (Fig 10). Individual antigen-specific IgG was capable
of mediating complement activation and complement-dependent killing of nonmucoid P.
aeruginosa strains (Fig 12). Importantly, passive immunization of mice with immune
monkey plasma enhanced the clearance of nonmucoid P. aeruginosa (Fig 13). Thus, I
75

believe that this study provides a strong foundation for future studies in infant CF
patients.
Children receive numerous vaccines including diphtheria, tetanus, acellular
pertussis toxin (DTaP), inactivated poliovirus, H. influenzae type B, measles, mumps,
rubella (MMR), and Streptococcus pneumoniae. Each of these vaccines is clearly
effective at reducing the incidence of infection. Children receive the vaccines over a
period of 18-months. Most childhood vaccines are given at least 3 times to achieve
protective antibody levels {200}. Importantly, lowering the number of immunizations
required to generate high levels of antibodies will reduce the cost of immunization and
number of doctor visits. My results with very young African green monkeys, OprF311-341-
OprI-Flagellins demonstrate that this vaccine promotes high levels of antigen-specific
IgG with only two immunizations using small amounts of antigen (Fig 10). The finding
that OprF311-341-OprI-Flagellins induces a high level of antibody production with only
two immunizations once again demonstrates the extraordinary potency of flagellin as an
adjuvant. However, I did observe a substantial decrease in the antigen-specific IgG three
months post-boost (Fig 10B). This result is consistent with what has been reported for
early life vaccination against H. influenzae type B, S. pneumoniae, and MMR {201-204}.
For example, infants immunized with a conjugate H. influenzae polysaccharide vaccine
had high antigen-specific IgG one month post-immunization but the IgG level fell 84%
by seven month post-immunization {205,206}. This is in contrast to what has been
observed for adult animals, in which the antigen-specific IgG titer remains high for at
least 3-6 months {207-209}. However, recent evidence suggests that the infant immune
system doesn’t support development of long-lived plasma cells {210}. At a young age,
76

the bone marrow lacks sufficient secretion of key plasma cells survival factors such as a
proliferation-inducing ligand, IL-6, and stromal cell-derived factor 1 to promote long-
lived plasma cells {211-216}. Thus, I hypothesis that the lack of sustainable IgG titers in
the young monkeys is due to the inability of the young immune system to support the
generation of long-lived plasma cells.
Although OprF311-341-OprI-Flagellins immunized monkeys have reduced antigen-
specific IgG titers three months after immunization, I hypothesize that the vaccine
induces the production of a large population of memory B cells. This hypothesis is based
on my results with mice in which large numbers of antigen-specific memory B cells were
generated after immunization with OprF311-341-OprI-Flagellins (Fig 4). Previous studies
have demonstrated that infants are biased towards generation of memory B cells
{217,218}. In addition, antigen-specific memory B cells can survive for extended
periods and upon reactivation rapidly generate a high titer of protective antibodies
{209,219-223}. Thus, it will be important to determine how the antigen-specific memory
B cells levels change over time in African green monkeys immunized with OprF311-341-
OprI-Flagellins.
The development of vaccines that are effective at generating a high quality long-
lasting immune response in very young CF patients may not only prevent the initial P.
aeruginosa infection, but may also prevent much of the inflammation induced by chronic
mucoid P. aeruginosa infection. However, it is quite likely that pathogens such as S.
aureus and H. influenzae would continue to be a significant concern in these patients. In
view of the flexibility of flagellin for use as an adjuvant and carrier, it may be that a
cocktail of flagellin fusion proteins targeting a range of CF pathogens may afford broad
77

immunity and thus spare the respiratory system from significant pathogen-induced
inflammatory damage.
The use of cocktail immunizations is an emerging concept in the field of
vaccinology. Several clinical trials have focused on reducing the number of
immunizations children receive by combining vaccines {224-226}. Many of the current
childhood vaccines are polysaccharide conjugate vaccines. The use of flagellin as a
carrier for polysaccharides has not been extensively studied. In view of the protective
efficacy of polysaccharides from S. pneumoniae, H. influenzae, and P. aeruginosa, it
would be beneficial to determine how conjugation of polysaccharides to flagellin
modulates the humoral immune response. Psl, a polysaccharide that is important for
attachment and biofilm formation and maintenance {35-37,227,228}, is an excellent
candidate for evaluation as a component of a flagellin conjugate vaccine. Conjugation of
Psl to flagellin would demonstrate a “proof of concept” that flagellin linked
polysaccharide vaccines are effective in promoting long-lived plasma and memory B
cells.
My studies represent a significant step forward for P. aeruginosa vaccine
development. Importantly, I have established a set of criteria for an effective vaccine and
have demonstrated the validity of these criteria. In addition, I have demonstrated the
efficacy of OprF311-341-OprI-Flagellins in adult mice as well as very young African green
monkeys.
78

REFERENCE LIST
1. Bear, C. E., C. Li, N. Kartner, R. J. Bridges, T. J. Jensen, M. Ramjeesingh, and J. R. Riordan. 1992. Purification and functional reconstitution of the cystic fibrosis transmembrane conductance regulator (CFTR). Cell 68:809-818.
2. Rommens, J. M., M. C. Iannuzzi, B. Kerem, M. L. Drumm, G. Melmer, M. Dean, R. Rozmahel, J. L. Cole, D. Kennedy, and N. Hidaka. 1989. Identification of the cystic fibrosis gene: chromosome walking and jumping. Science 245:1059-1065.
3. Lu, C., C. Jiang, S. Pribanic, and D. Rotin. 2007. CFTR stabilizes ENaC at the plasma membrane. Journal of Cystic Fibrosis 6:419-422.
4. Cystic Fibrosis Mutation Database. 3-2-2007.
5. Gan, K. H., H. J. Veeze, A. van den Ouweland, D. Halley, H. Scheffer, A. van der Hout, S. E. Overbeek, J. C. de Jongste, W. Bakker, and H. Heijerman. 1995. A Cystic Fibrosis Mutation Associated with Mild Lung Disease. N Engl J Med 333:95-99.
6. Riordan, J. R., J. M. Rommens, B. Kerem, N. Alon, R. Rozmahel, Z. Grzelczak, J. Zielenski, S. Lok, N. Plavsic, J. L. Chou, and . 1989. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245:1066-1073.
7. Kerem, E. 2005. Pharmacological induction of CFTR function in patients with cystic fibrosis: mutation-specific therapy. Pediatr. Pulmonol. 40:183-196.
8. Welsh, M. J., G. M. Denning, L. S. Ostedgaard, and M. P. Anderson. 1993. Dysfunction of CFTR bearing the delta F508 mutation. J Cell Sci. Suppl 17:235-239.
9. Widdicombe, J. H., M. J. Welsh, and W. E. Finkbeiner. 1985. Cystic fibrosis decreases the apical membrane chloride permeability of monolayers cultured from cells of tracheal epithelium. Proc. Natl. Acad. Sci. U. S. A 82:6167-6171.
10. Zielenski, J. 2000. Genotype and phenotype in cystic fibrosis. Respiration 67:117-133.
11. Mall, M., B. R. Grubb, J. R. Harkema, W. K. O'Neal, and R. C. Boucher. 2004. Increased airway epithelial Na+ absorption produces cystic fibrosis-like lung disease in mice. Nat Med 10:487-493.
12. Mall, M. A., J. R. Harkema, J. B. Trojanek, D. Treis, A. Livraghi, S. Schubert, Z. Zhou, S. M. Kreda, S. L. Tilley, E. J. Hudson, W. K. O'Neal, and R. C. Boucher.
79

2008. Development of Chronic Bronchitis and Emphysema in {beta}-Epithelial Na+ Channel-Overexpressing Mice. Am. J. Respir. Crit. Care Med. 177:730-742.
13. Chmiel, J., M. Berger, and M. Konstan. 2002. The role of inflammation in the pathophysiology of CF lung disease. Clinical Reviews in Allergy and Immunology 23:5-27.
14. Worlitzsch, D., R. Tarran, M. Ulrich, U. Schwab, A. Cekici, K. C. Meyer, P. Birrer, G. Bellon, J. Berger, T. Weiss, K. Botzenhart, J. R. Yankaskas, S. Randell, R. C. Boucher, and G. Doring. 2002. Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients. J Clin Invest 109:317-325.
15. Brown, R. K., H. Wyatt, J. F. Price, and F. J. Kelly. 1996. Pulmonary dysfunction in cystic fibrosis is associated with oxidative stress. Eur Respir J 9:334-339.
16. Mathee, K., O. Ciofu, C. Sternberg, P. W. Lindum, J. I. Campbell, P. Jensen, A. H. Johnsen, M. Givskov, D. E. Ohman, S. Molin, N. Hoiby, and A. Kharazmi. 1999. Mucoid conversion of Pseudomonas aeruginosa by hydrogen peroxide: a mechanism for virulence activation in the cystic fibrosis lung. Microbiology 145 ( Pt 6):1349-1357.
17. Oliver, A., R. Canton, P. Campo, F. Baquero, and J. Blazquez. 2000. High Frequency of Hypermutable Pseudomonas aeruginosa in Cystic Fibrosis Lung Infection. Science 288:1251-1253.
18. Garrett, E. S., D. Perlegas, and D. J. Wozniak. 1999. Negative control of flagellum synthesis in Pseudomonas aeruginosa is modulated by the alternative sigma factor AlgT (AlgU). J Bacteriol. 181:7401-7404.
19. Tart, A. H., M. C. Wolfgang, and D. J. Wozniak. 2005. The alternative sigma factor AlgT represses Pseudomonas aeruginosa flagellum biosynthesis by inhibiting expression of fleQ. J Bacteriol. 187:7955-7962.
20. Abman, S. H., J. W. Ogle, R. J. Harbeck, N. Butler-Simon, K. B. Hammond, and F. J. Accurso. 1991. Early bacteriologic, immunologic, and clinical courses of young infants with cystic fibrosis identified by neonatal screening. J Pediatr. 119:211-217.
21. Govan, J. R. and V. Deretic. 1996. Microbial pathogenesis in cystic fibrosis: mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol Rev 60:539-574.
22. Burns, J. L., R. L. Gibson, S. McNamara, D. Yim, J. Emerson, M. Rosenfeld, P. Hiatt, K. McCoy, R. Castile, A. L. Smith, and B. W. Ramsey. 2001. Longitudinal assessment of Pseudomonas aeruginosa in young children with cystic fibrosis. J Infect Dis 183:444-452.
80

23. Li, Z., M. R. Kosorok, P. M. Farrell, A. Laxova, S. E. H. West, C. G. Green, J. Collins, M. J. Rock, and M. L. Splaingard. 2005. Longitudinal Development of Mucoid Pseudomonas aeruginosa Infection and Lung Disease Progression in Children With Cystic Fibrosis. JAMA 293:581-588.
24. Sibley, C. D., M. D. Parkins, H. R. Rabin, K. Duan, J. C. Norgaard, and M. G. Surette. 2008. A polymicrobial perspective of pulmonary infections exposes an enigmatic pathogen in cystic fibrosis patients. PNAS.
25. Tosi, M. F., H. Zakem-Cloud, C. A. Demko, J. R. Schreiber, R. C. Stern, M. W. Konstan, and M. Berger. 1995. Cross-sectional and longitudinal studies of naturally occurring antibodies to Pseudomonas aeruginosa in cystic fibrosis indicate absence of antibody-mediated protection and decline in opsonic quality after infection. J Infect Dis 172:453-461.
26. Sonawane, A., J. Jyot, R. During, and R. Ramphal. 2006. Neutrophil Elastase, an Innate Immunity Effector Molecule, Represses Flagellin Transcription in Pseudomonas aeruginosa. Infect. Immun. 74:6682-6689.
27. Tamura, Y., S. Suzuki, and T. Sawada. 1992. Role of elastase as a virulence factor in experimental Pseudomonas aeruginosa infection in mice. Microb. Pathog. 12:237-244.
28. Voynow, J. A., B. M. Fischer, D. E. Malarkey, L. H. Burch, T. Wong, M. Longphre, S. B. Ho, and W. M. Foster. 2004. Neutrophil elastase induces mucus cell metaplasia in mouse lung. Am J Physiol Lung Cell Mol Physiol 287:L1293-L1302.
29. Hirche, T. O., R. Benabid, G. Deslee, S. Gangloff, S. Achilefu, M. Guenounou, F. Lebargy, R. E. Hancock, and A. Belaaouaj. 2008. Neutrophil Elastase Mediates Innate Host Protection against Pseudomonas aeruginosa. J Immunol 181:4945-4954.
30. Walker, T. S., K. L. Tomlin, G. S. Worthen, K. R. Poch, J. G. Lieber, M. T. Saavedra, M. B. Fessler, K. C. Malcolm, M. L. Vasil, and J. A. Nick. 2005. Enhanced Pseudomonas aeruginosa Biofilm Development Mediated by Human Neutrophils. Infect. Immun. 73:3693-3701.
31. Fick, R. B., Jr., F. Sonoda, and D. B. Hornick. 1992. Emergence and persistence of Pseudomonas aeruginosa in the cystic fibrosis airway. Semin. Respir Infect 7:168-178.
32. Stotland, P. K., D. Radzioch, and M. M. Stevenson. 2000. Mouse models of chronic lung infection with Pseudomonas aeruginosa: models for the study of cystic fibrosis. Pediatr. Pulmonol. 30:413-424.
33. Heijerman, H. 2005. Infection and inflammation in cystic fibrosis: A short review. Journal of Cystic Fibrosis 4:3-5.
81

34. Tam, M., G. Jackson Snipes, and M. M. Stevenson. 1999. Characterization of Chronic Bronchopulmonary Pseudomonas aeruginosa Infection in Resistant and Susceptible Inbred Mouse Strains. Am. J. Respir. Cell Mol. Biol. 20:710-719.
35. Ma, L., H. Lu, A. Sprinkle, M. R. Parsek, and D. Wozniak. 2007. Pseudomonas aeruginosa Psl is a galactose- and mannose-rich exopolysaccharide. J. Bacteriol.JB.
36. Ma, L., K. D. Jackson, R. M. Landry, M. R. Parsek, and D. J. Wozniak. 2006. Analysis of Pseudomonas aeruginosa Conditional Psl Variants Reveals Roles for the Psl Polysaccharide in Adhesion and Maintaining Biofilm Structure Postattachment. J. Bacteriol. 188:8213-8221.
37. Jackson, K. D., M. Starkey, S. Kremer, M. R. Parsek, and D. J. Wozniak. 2004. Identification of psl, a Locus Encoding a Potential Exopolysaccharide That Is Essential for Pseudomonas aeruginosa PAO1 Biofilm Formation. J. Bacteriol. 186:4466-4475.
38. Leid, J. G., C. J. Willson, M. E. Shirtliff, D. J. Hassett, M. R. Parsek, and A. K. Jeffers. 2005. The Exopolysaccharide Alginate Protects Pseudomonas aeruginosa Biofilm Bacteria from IFN-{gamma}-Mediated Macrophage Killing. J Immunol 175:7512-7518.
39. Hauser, A. R., S. Fleiszig, P. J. Kang, K. Mostov, and J. N. Engel. 1998. Defects in Type III Secretion Correlate with Internalization of Pseudomonas aeruginosa by Epithelial Cells. Infect. Immun. 66:1413-1420.
40. Galle, M., P. Schotte, M. Haegman, A. Wullaert, H. J. Yang, S. Jin, and R. Beyaert. 2008. The Pseudomonas aeruginosa Type III secretion system plays a dual role in the regulation of caspase-1 mediated IL-1beta maturation. J Cell Mol Med 12:1767-1776.
41. Sato, H., D. W. Frank, C. J. Hillard, J. B. Feix, R. R. Pankhaniya, K. Moriyama, V. Finck-Barbancon, A. Buchaklian, M. Lei, R. M. Long, J. Wiener-Kronish, and T. Sawa. 2003. The mechanism of action of the Pseudomonas aeruginosa-encoded type III cytotoxin, ExoU. EMBO J 22:2959-2969.
42. Vance, R. E., A. Rietsch, and J. J. Mekalanos. 2005. Role of the Type III Secreted Exoenzymes S, T, and Y in Systemic Spread of Pseudomonas aeruginosa PAO1 In Vivo. Infect. Immun. 73:1706-1713.
43. Davies, D. G., M. R. Parsek, J. P. Pearson, B. H. Iglewski, J. W. Costerton, and E. P. Greenberg. 1998. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280:295-298.
44. Smith, R. S., S. G. Harris, R. Phipps, and B. Iglewski. 2002. The Pseudomonas aeruginosa Quorum-Sensing Molecule N-(3-Oxododecanoyl)Homoserine
82

Lactone Contributes to Virulence and Induces Inflammation In Vivo. J. Bacteriol. 184:1132-1139.
45. Drenkard, E. and F. M. Ausubel. 2002. Pseudomonas biofilm formation and antibiotic resistance are linked to phenotypic variation. Nature 416:740-743.
46. Wu, L., O. Estrada, O. Zaborina, M. Bains, L. Shen, J. E. Kohler, N. Patel, M. W. Musch, E. B. Chang, Y. X. Fu, M. A. Jacobs, M. I. Nishimura, R. E. W. Hancock, J. R. Turner, and J. C. Alverdy. 2005. Recognition of Host Immune Activation by Pseudomonas aeruginosa. Science 309:774-777.
47. Kunert, A., J. Losse, C. Gruszin, M. Huhn, K. Kaendler, S. Mikkat, D. Volke, R. Hoffmann, T. S. Jokiranta, H. Seeberger, U. Moellmann, J. Hellwage, and P. F. Zipfel. 2007. Immune Evasion of the Human Pathogen Pseudomonas aeruginosa: Elongation Factor Tuf Is a Factor H and Plasminogen Binding Protein. J Immunol 179:2979-2988.
48. Gross, G. N., S. R. Rehm, and A. K. Pierce. 1978. The effect of complement depletion on lung clearance of bacteria. J Clin Invest 62:373-378.
49. Mueller-Ortiz, S. L., S. M. Drouin, and R. A. Wetsel. 2004. The Alternative Activation Pathway and Complement Component C3 Are Critical for a Protective Immune Response against Pseudomonas aeruginosa in a Murine Model of Pneumonia. Infect. Immun. 72:2899-2906.
50. Younger, J. G., S. Shankar-Sinha, M. Mickiewicz, A. S. Brinkman, G. A. Valencia, J. V. Sarma, E. M. Younkin, T. J. Standiford, F. S. Zetoune, and P. A. Ward. 2003. Murine Complement Interactions with Pseudomonas aeruginosa and Their Consequences During Pneumonia. Am. J. Respir. Cell Mol. Biol. 29:432-438.
51. Schiller, N. L. 1988. Characterization of the susceptibility of Pseudomonas aeruginosa to complement-mediated killing: role of antibodies to the rough lipopolysaccharide on serum-sensitive strains. Infect. Immun. 56:632-639.
52. Feuillet, V., S. Medjane, I. Mondor, O. Demaria, P. P. Pagni, J. E. Galan, R. A. Flavell, and L. Alexopoulou. 2006. Involvement of Toll-like receptor 5 in the recognition of flagellated bacteria. PNAS 103:12487-12492.
53. Anderson, T. R., T. C. Montie, M. D. Murphy, and V. P. McCarthy. 1989. Pseudomonas aeruginosa flagellar antibodies in patients with cystic fibrosis. J Clin Microbiol 27:2789-2793.
54. Ansorg, R. 1978. [Permanence and parellelism of serogroups of Pseudomonas aeruginosa in hospitalized patients (author's transl)]. Zentralbl. Bakteriol. [Orig. A] 240:191-196.
83

55. Anderson, T. R. and T. C. Montie. 1989. Flagellar antibody stimulated opsonophagocytosis of Pseudomonas aeruginosa associated with response to either a- or b-type flagellar antigen. Can. J Microbiol 35:890-894.
56. Nilsson, E., A. Amini, B. Wretlind, and A. Larsson. 2007. Pseudomonas aeruginosa infections are prevented in cystic fibrosis patients by avian antibodies binding Pseudomonas aeruginosa flagellin. Journal of Chromatography B 856:75-80.
57. Verma, A., M. Schirm, S. K. Arora, P. Thibault, S. M. Logan, and R. Ramphal. 2006. Glycosylation of b-Type Flagellin of Pseudomonas aeruginosa: Structural and Genetic Basis. J. Bacteriol. 188:4395-4403.
58. Verma, A., S. K. Arora, S. K. Kuravi, and R. Ramphal. 2005. Roles of Specific Amino Acids in the N Terminus of Pseudomonas aeruginosa Flagellin and of Flagellin Glycosylation in the Innate Immune Response. Infect. Immun. 73:8237-8246.
59. Schirm, M., S. K. Arora, A. Verma, E. Vinogradov, P. Thibault, R. Ramphal, and S. M. Logan. 2004. Structural and Genetic Characterization of Glycosylation of Type a Flagellin in Pseudomonas aeruginosa. J. Bacteriol. 186:2523-2531.
60. Honko, A. N. and S. B. Mizel. 2004. Mucosal Administration of Flagellin Induces Innate Immunity in the Mouse Lung. Infect. Immun. 72:6676-6679.
61. Honko, A. N., N. Sriranganathan, C. J. Lees, and S. B. Mizel. 2006. Flagellin Is an Effective Adjuvant for Immunization against Lethal Respiratory Challenge with Yersinia pestis. Infect. Immun. 74:1113-1120.
62. Mizel, S. B., A. N. Honko, M. A. Moors, P. S. Smith, and A. P. West. 2003. Induction of Macrophage Nitric Oxide Production by Gram-Negative Flagellin Involves Signaling Via Heteromeric Toll-Like Receptor 5/Toll-Like Receptor 4 Complexes. J Immunol 170:6217-6223.
63. Amer, A., L. Franchi, T. D. Kanneganti, M. Body-Malapel, N. Ozoren, G. Brady, S. Meshinchi, R. Jagirdar, A. Gewirtz, S. Akira, and G. Nunez. 2006. Regulation of Legionella phagosome maturation and infection through flagellin and host IPAF. J. Biol. Chem.M604933200.
64. Miao, E. A., R. K. Ernst, M. Dors, D. P. Mao, and A. Aderem. 2008. Pseudomonas aeruginosa activates caspase 1 through Ipaf. PNAS 105:2562-2567.
65. Franchi, L., J. Stoolman, T. D. Kanneganti, A. Verma, R. Ramphal, and G. Nunez. 2007. Critical role for Ipaf in Pseudomonas aeruginosa-induced caspase-1 activation. Eur. J Immunol 37:3030-3039.
66. Franchi, L., A. Amer, M. Body-Malapel, T. D. Kanneganti, N. Ozoren, R. Jagirdar, N. Inohara, P. Vandenabeele, J. Bertin, A. Coyle, E. P. Grant, and G.
84

Nunez. 2006. Cytosolic flagellin requires Ipaf for activation of caspase-1 and interleukin 1[beta] in salmonella-infected macrophages. Nat Immunol advanced online publication.
67. Miao, E., E. Andersen-Nissen, S. Warren, and A. Aderem. 2007. TLR5 and Ipaf: dual sensors of bacterial flagellin in the innate immune system. Seminars in Immunopathology 29:275-288.
68. Miao, E. A., C. M. Alpuche-Aranda, M. Dors, A. E. Clark, M. W. Bader, S. I. Miller, and A. Aderem. 2006. Cytoplasmic flagellin activates caspase-1 and secretion of interleukin 1[beta] via Ipaf. Nat Immunol advanced online publication.
69. Sutterwala, F. S., L. A. Mijares, L. Li, Y. Ogura, B. I. Kazmierczak, and R. A. Flavell. 2007. Immune recognition of Pseudomonas aeruginosa mediated by the IPAF/NLRC4 inflammasome. J. Exp. Med.jem.
70. Kashef, N., Q. Behzadian-Nejad, S. Najar-Peerayeh, K. Mousavi-Hosseini, M. Moazzeni, and G. E. Djavid. 2006. Synthesis and characterization of Pseudomonas aeruginosa alginate-tetanus toxoid conjugate. J Med Microbiol 55:1441-1446.
71. DiGiandomenico, A., J. Rao, K. Harcher, T. S. Zaidi, J. Gardner, A. N. Neely, G. B. Pier, and J. B. Goldberg. 2007. Intranasal immunization with heterologously expressed polysaccharide protects against multiple Pseudomonas aeruginosa infections. PNAS 104:4624-4629.
72. Arnold, H., D. Bumann, M. Felies, B. Gewecke, M. Sorensen, J. E. Gessner, J. Freihorst, B. U. von Specht, and U. Baumann. 2004. Enhanced Immunogenicity in the Murine Airway Mucosa with an Attenuated Salmonella Live Vaccine Expressing OprF-OprI from Pseudomonas aeruginosa. Infect. Immun. 72:6546-6553.
73. Scarff, J. M. and J. B. Goldberg. 2008. Vaccination against Pseudomonas aeruginosa Pneumonia in Immunocompromised Mice. Clin. Vaccine Immunol. 15:367-375.
74. Cryz, S. J., Jr., A. Lang, A. Rudeberg, J. Wedgwood, J. U. Que, E. Furer, and U. Schaad. 1997. Immunization of cystic fibrosis patients with a Pseudomonas aeruginosa O-polysaccharide-toxin A conjugate vaccine. Behring Inst. Mitt.345-349.
75. Lai, Z., R. Kimmel, S. Petersen, S. Thomas, G. Pier, B. Bezabeh, R. Luo, and J. R. Schreiber. 2005. Multi-valent human monoclonal antibody preparation against Pseudomonas aeruginosa derived from transgenic mice containing human immunoglobulin loci is protective against fatal pseudomonas sepsis caused by multiple serotypes. Vaccine 23:3264-3271.
85

76. Preston, M. J., A. A. Gerceker, M. E. Reff, and G. B. Pier. 1998. Production and Characterization of a Set of Mouse-Human Chimeric Immunoglobulin G (IgG) Subclass and IgA Monoclonal Antibodies with Identical Variable Regions Specific for Pseudomonas aeruginosa Serogroup O6 Lipopolysaccharide. Infect. Immun. 66:4137-4142.
77. Stanislavsky, E. S. and J. S. Lam. 1997. Pseudomonas aeruginosa antigens as potential vaccines. FEMS Microbiology Reviews 21:243-277.
78. Pier, G. B., D. DesJardin, M. Grout, C. Garner, S. E. Bennett, G. Pekoe, S. A. Fuller, M. O. Thornton, W. S. Harkonen, and H. C. Miller. 1994. Human immune response to Pseudomonas aeruginosa mucoid exopolysaccharide (alginate) vaccine. Infect. Immun. 62:3972-3979.
79. Theilacker, C., F. T. Coleman, S. Mueschenborn, N. Llosa, M. Grout, and G. B. Pier. 2003. Construction and Characterization of a Pseudomonas aeruginosa Mucoid Exopolysaccharide-Alginate Conjugate Vaccine. Infect. Immun. 71:3875-3884.
80. Zuercher, A. W., M. P. Horn, J. U. Que, A. Ruedeberg, M. H. Schoeni, U. B. Schaad, P. Marcus, and A. B. Lang. 2006. Antibody responses induced by long-term vaccination with an octovalent conjugate Pseudomonas aeruginosa vaccine in children with cystic fibrosis. FEMS Immunol Med Microbiol 47:302-308.
81. Zuercher, A. W., M. P. Horn, H. Wu, Z. Song, C. J. Bundgaard, H. K. Johansen, N. Hoiby, P. Marcus, and A. B. Lang. 2006. Intranasal immunisation with conjugate vaccine protects mice from systemic and respiratory tract infection with Pseudomonas aeruginosa. Vaccine 24:4333-4342.
82. Stocker, B. A. 1988. Auxotrophic Salmonella typhi as live vaccine. Vaccine 6:141-145.
83. DiGiandomenico, A., J. Rao, and J. B. Goldberg. 2004. Oral Vaccination of BALB/c Mice with Salmonella enterica Serovar Typhimurium Expressing Pseudomonas aeruginosa O Antigen Promotes Increased Survival in an Acute Fatal Pneumonia Model. Infect. Immun. 72:7012-7021.
84. Priebe, G. P., G. J. Meluleni, F. T. Coleman, J. B. Goldberg, and G. B. Pier. 2003. Protection against Fatal Pseudomonas aeruginosa Pneumonia in Mice after Nasal Immunization with a Live, Attenuated aroA Deletion Mutant. Infect. Immun. 71:1453-1461.
85. Priebe, G. P., M. M. Brinig, K. Hatano, M. Grout, F. T. Coleman, G. B. Pier, and J. B. Goldberg. 2002. Construction and Characterization of a Live, Attenuated aroA Deletion Mutant of Pseudomonas aeruginosa as a Candidate Intranasal Vaccine. Infect. Immun. 70:1507-1517.
86

86. Alexander, J. W., M. W. Fisher, and B. G. MacMillan. 1971. Immunological control of Pseudomonas infection in burn patients: a clinical evaluation. Arch Surg. 102:31-35.
87. Pennington, J. E., H. Y. Reynolds, R. E. Wood, R. A. Robinson, and A. S. Levine. 1975. Use of a Pseudomonas aeruginosa vaccine in pateints with acute leukemia and cystic fibrosis. Am J Med 58:629-636.
88. Wood, R. E., J. E. Pennington, and H. Y. Reynolds. 1983. Intranasal administration of a Pseudomonas lipopolysaccharide vaccine in cystic fibrosis patients. Pediatr. Infect Dis 2:367-369.
89. Alexander, J. W. and M. W. Fisher. 1970. Vaccination for Pseudomonas aeruginosa. Am J Surg. 120:512.
90. Alexander, J. W., M. W. Fisher, B. G. MacMillan, and W. A. Altemeier. 1969. Prevention of invasive pseudomonas infection in burns with a new vaccine. Arch Surg. 99:249-256.
91. Jones, C. E., Jr., J. W. Alexander, and M. W. Fisher. 1970. Clinical evaluation of Pseudomonas hyperimmune globulin and plasma. Surg. Forum 21:238-240.
92. Schemmer, K. E., J. W. Alexander, and M. W. Fisher. 1969. Immunologic response of patients with severe burns to pseudomonas vaccination. Surg. Forum 20:69-71.
93. Donta, S. T., P. Peduzzi, A. S. Cross, J. Sadoff, C. Haakenson, S. J. Cryz, Jr., C. Kauffman, S. Bradley, G. Gafford, D. Elliston, T. R. Beam, Jr., J. F. John, Jr., B. Ribner, R. Cantey, C. H. Welsh, R. T. Ellison, III, E. J. Young, R. J. Hamill, H. Leaf, R. M. Schein, M. Mulligan, C. Johnson, E. Abrutyn, J. M. Griffiss, D. Slagle, and . 1996. Immunoprophylaxis against klebsiella and pseudomonas aeruginosa infections. The Federal Hyperimmune Immunoglobulin Trial Study Group. J Infect Dis 174:537-543.
94. Doring, G. and G. B. Pier. 2008. Vaccines and immunotherapy against Pseudomonas aeruginosa. Vaccine 26:1011-1024.
95. Hatano, K., J. B. Goldberg, and G. B. Pier. 1995. Biologic activities of antibodies to the neutral-polysaccharide component of the Pseudomonas aeruginosa lipopolysaccharide are blocked by O side chains and mucoid exopolysaccharide (alginate). Infect. Immun. 63:21-26.
96. Liu, P. V. and S. Wang. 1990. Three new major somatic antigens of Pseudomonas aeruginosa. Journal Of Clinical Microbiology 28:922-925.
97. Skjak-Braek, G., H. Grasdalen, and B. Larsen. 1986. Monomer sequence and acetylation pattern in some bacterial alginates. Carbohydr. Res 154:239-250.
87

98. Garner, C. V., D. DesJardins, and G. B. Pier. 1990. Immunogenic properties of Pseudomonas aeruginosa mucoid exopolysaccharide. Infect. Immun. 58:1835-1842.
99. Bragonzi, A., D. Worlitzsch, G. Pier, P. Timpert, M. Ulrich, M. Hentzer, J. Andersen, M. Givskov, M. Conese, and G. Doring. 2005. Nonmucoid Pseudomonas aeruginosa Expresses Alginate in the Lungs of Patients with Cystic Fibrosis and in a Mouse Model. J Infect Dis 192:410-419.
100. Wozniak, D. J., T. J. Wyckoff, M. Starkey, R. Keyser, P. Azadi, G. A. O'Toole, and M. R. Parsek. 2003. Alginate is not a significant component of the extracellular polysaccharide matrix of PA14 and PAO1 Pseudomonas aeruginosa biofilms. Proc. Natl. Acad. Sci. U. S. A 100:7907-7912.
101. De Vos, D., C. Bouton, A. Sarniguet, P. De Vos, M. Vauterin, and P. Cornelis. 1998. Sequence Diversity of the oprI Gene, Coding for Major Outer Membrane Lipoprotein I, among rRNA Group I Pseudomonads. J. Bacteriol. 180:6551-6556.
102. Duchene, M., C. Barron, A. Schweizer, B. U. von Specht, and H. Domdey. 1989. Pseudomonas aeruginosa outer membrane lipoprotein I gene: molecular cloning, sequence, and expression in Escherichia coli. J. Bacteriol. 171:4130-4137.
103. Finke, M., M. Duchene, A. Eckhardt, H. Domdey, and B. U. von Specht. 1990. Protection against experimental Pseudomonas aeruginosa infection by recombinant P. aeruginosa lipoprotein I expressed in Escherichia coli. Infect. Immun. 58:2241-2244.
104. Finke, M., G. Muth, T. Reichhelm, M. Thoma, M. Duchene, K. D. Hungerer, H. Domdey, and B. U. von Specht. 1991. Protection of immunosuppressed mice against infection with Pseudomonas aeruginosa by recombinant P. aeruginosa lipoprotein I and lipoprotein I-specific monoclonal antibodies. Infect. Immun. 59:1251-1254.
105. Gerke, J. R. and J. S. Nelson. 1977. Oral vaccination and multivalent vaccine against Pseudomonas aeruginosa keratitis. Invest. Ophthalmol. Vis. Sci. 16:76-80.
106. Saha, S., F. Takeshita, S. Sasaki, T. Matsuda, T. Tanaka, M. Tozuka, K. Takase, T. Matsumoto, K. Okuda, N. Ishii, K. Yamaguchi, D. M. Klinman, K. Q. Xin, and K. Okuda. 2006. Multivalent DNA vaccine protects mice against pulmonary infection caused by Pseudomonas aeruginosa. Vaccine 24:6240-6249.
107. Baumann, U., E. Mansouri, and B.-U. von Specht. 2004. Recombinant OprF-OprI as a vaccine against Pseudomonas aeruginosa infections. Vaccine 22:840-847.
108. Finnen, R. L., N. L. Martin, R. J. Siehnel, W. A. Woodruff, M. Rosok, and R. E. Hancock. 1992. Analysis of the Pseudomonas aeruginosa major outer membrane protein OprF by use of truncated OprF derivatives and monoclonal antibodies. J. Bacteriol. 174:4977-4985.
88

109. Gocke, K., U. Baumann, H. Hagemann, J. Gabelsberger, H. Hahn, J. Freihorst, and B. U. Specht. 2003. Mucosal vaccination with a recombinant OprF-I vaccine of Pseudomonas aeruginosa in healthy volunteers: comparison of a systemic vs. a mucosal booster schedule. FEMS Immunology & Medical Microbiology 37:167-171.
110. Mansouri, E., S. Blome-Eberwein, J. Gabelsberger, G. Germann, and B. U. von Specht. 2003. Clinical study to assess the immunogenicity and safety of a recombinant Pseudomonas aeruginosa OprF-OprI vaccine in burn patients. FEMS Immunol Med Microbiol 37:161-166.
111. Mansouri, E., J. Gabelsberger, B. Knapp, E. Hundt, U. Lenz, K. D. Hungerer, H. E. Gilleland, Jr., J. Staczek, H. Domdey, and B. U. von Specht. 1999. Safety and Immunogenicity of a Pseudomonas aeruginosa Hybrid Outer Membrane Protein F-I Vaccine in Human Volunteers. Infect. Immun. 67:1461-1470.
112. Martin, N. L., E. G. Rawling, R. S. Wong, M. Rosok, and R. E. Hancock. 1993. Conservation of surface epitopes in Pseudomonas aeruginosa outer membrane porin protein OprF. FEMS Microbiol Lett. 113:261-266.
113. Price, B. M., L. J. Barten, D. R. Galloway, B. U. von Specht, L. B. Gilleland, H. E. Gilleland, Jr., and J. Staczek. 2002. Enhancement of the protective efficacy of an oprF DNA vaccine against Pseudomonas aeruginosa. FEMS Immunol Med Microbiol 33:89-99.
114. Rawling, E. G., N. L. Martin, and R. E. Hancock. 1995. Epitope mapping of the Pseudomonas aeruginosa major outer membrane porin protein OprF. Infect. Immun. 63:38-42.
115. Weimer, E. T., H. Lu, N. D. Kock, D. J. Wozniak, and S. B. Mizel. 2009. A fusion protein vaccine containing OprF epitope 8, OprI, and Type A and B flagellin promotes enhanced clearance of nonmucoid Pseudomonas aeruginosa. Infect. Immun. 77:2356-2366.
116. Worgall, S., A. Krause, M. Rivara, K. K. Hee, E. V. Vintayen, N. R. Hackett, P. W. Roelvink, J. T. Bruder, T. J. Wickham, I. Kovesdi, and R. G. Crystal. 2005. Protection against P. aeruginosa with an adenovirus vector containing an OprF epitope in the capsid. J. Clin. Invest. 115:1281-1289.
117. Worgall, S., A. Krause, J. Qiu, J. Joh, N. R. Hackett, and R. G. Crystal. 2007. Protective Immunity to Pseudomonas aeruginosa Induced with a Capsid-Modified Adenovirus Expressing P. aeruginosa OprF. J. Virol. 81:13801-13808.
118. Qin, X., J. Emerson, J. Stapp, L. Stapp, P. Abe, and J. L. Burns. 2003. Use of Real-Time PCR with Multiple Targets To Identify Pseudomonas aeruginosa and Other Nonfermenting Gram-Negative Bacilli from Patients with Cystic Fibrosis. Journal Of Clinical Microbiology 41:4312-4317.
89

119. De Vos, D., A. Lim, Jr., J. P. Pirnay, M. Struelens, C. Vandenvelde, L. Duinslaeger, A. Vanderkelen, and P. Cornelis. 1997. Direct detection and identification of Pseudomonas aeruginosa in clinical samples such as skin biopsy specimens and expectorations by multiplex PCR based on two outer membrane lipoprotein genes, oprI and oprL. Journal Of Clinical Microbiology 35:1295-1299.
120. Eckhardt, A., M. M. Heiss, W. Ehret, W. Permanetter, M. Duchene, H. Domdey, and B. U. von Specht. 1991. Evaluation of protective mAbs against Pseudomonas aeruginosa outer membrane protein I by C1q binding assay. Zentralbl. Bakteriol. 275:100-111.
121. von Specht, B. U., H. Christian Lucking, B. Blum, A. Schmitt, K. Dieter Hungerer, and H. Domdey. 1996. Safety and immunogenicity of a Pseudomonas aeruginosa outer membrane protein I vaccine in human volunteers. Vaccine 14:1111-1117.
122. von Specht, B. U., H. C. Lucking, B. Blum, A. Schmitt, K. D. Hungerer, and H. Domdey. 1997. The Pseudomonas aeruginosa outer membrane protein I vaccine: immunogenicity and safe administration in man. Behring Inst. Mitt.326-337.
123. Price, B. M., D. R. Galloway, N. R. Baker, L. B. Gilleland, J. Staczek, and H. E. Gilleland, Jr. 2001. Protection against Pseudomonas aeruginosa Chronic Lung Infection in Mice by Genetic Immunization against Outer Membrane Protein F (OprF) of P. aeruginosa. Infect. Immun. 69:3510-3515.
124. Sorichter, S., U. Baumann, A. Baumgart, S. Walterspacher, and B. U. von Specht. 9 A.D. Immune responses in the airways by nasal vaccination with systemic boosting against Pseudomonas aeruginosa in chronic lung disease. Vaccine 27:2755-2759.
125. Knapp, B., E. Hundt, U. Lenz, K. D. Hungerer, J. Gabelsberger, H. Domdey, E. Mansouri, Y. Li, and B. U. von Specht. 1999. A recombinant hybrid outer membrane protein for vaccination against Pseudomonas aeruginosa. Vaccine 17:1663-1666.
126. Hertle, R., R. Mrsny, and D. J. Fitzgerald. 2001. Dual-function vaccine for Pseudomonas aeruginosa: characterization of chimeric exotoxin A-pilin protein. Infect Immun. 69:6962-6969.
127. Hsieh, J. C., D. M. Tham, W. Feng, F. Huang, S. Embaie, K. Liu, D. Dean, R. Hertle, D. J. Fitzgerald, and R. J. Mrsny. 2005. Intranasal immunization strategy to impede pilin-mediated binding of Pseudomonas aeruginosa to airway epithelial cells. Infect Immun. 73:7705-7717.
128. Doring, G., C. Meisner, M. Stern, and for the Flagella Vaccine Trial Study Group. 2007. A double-blind randomized placebo-controlled phase III study of a
90

Pseudomonas aeruginosa flagella vaccine in cystic fibrosis patients. PNAS 104:11020-11025.
129. Neville, L. F., Y. Barnea, O. Hammer-Munz, E. Gur, B. Kuzmenko, H. Kahel-Raifer, R. Eren, A. Elkeles, K. G. Murthy, C. Szabo, A. L. Salzman, S. Dagan, Y. Carmeli, and S. Navon-Venezia. 2005. Antibodies raised against N'-terminal Pseudomonas aeruginosa flagellin prevent mortality in lethal murine models of infection. Int. J Mol. Med 16:165-171.
130. Barnea, Y., Y. Carmeli, L. F. Neville, H. Kahel-Reifer, R. Eren, S. Dagan, and S. Navon-Venezia. 2009. Therapy with anti-flagellin A monoclonal antibody limits Pseudomonas aeruginosa invasiveness in a mouse burn wound sepsis model. Burns In Press, Corrected Proof.
131. Kool, M., T. Soullie, M. van Nimwegen, M. A. M. Willart, F. Muskens, S. Jung, H. C. Hoogsteden, H. Hammad, and B. N. Lambrecht. 2008. Alum adjuvant boosts adaptive immunity by inducing uric acid and activating inflammatory dendritic cells. J. Exp. Med.jem.
132. Kool, M., V. Petrilli, T. De Smedt, A. Rolaz, H. Hammad, M. van Nimwegen, I. M. Bergen, R. Castillo, B. N. Lambrecht, and J. Tschopp. 2008. Cutting Edge: Alum Adjuvant Stimulates Inflammatory Dendritic Cells through Activation of the NALP3 Inflammasome. J Immunol 181:3755-3759.
133. Eisenbarth, S. C., O. R. Colegio, W. 'Connor, F. S. Sutterwala, and R. A. Flavell. 2008. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature 453:1122-1126.
134. Cuadros, C., F. J. Lopez-Hernandez, A. L. Dominguez, M. McClelland, and J. Lustgarten. 2004. Flagellin Fusion Proteins as Adjuvants or Vaccines Induce Specific Immune Responses. Infect. Immun. 72:2810-2816.
135. Huleatt, J. W., A. R. Jacobs, J. Tang, P. Desai, E. B. Kopp, Y. Huang, L. Song, V. Nakaar, and T. J. Powell. 2006. Vaccination with recombinant fusion proteins incorporating Toll-like receptor ligands induces rapid cellular and humoral immunity. Vaccine In Press, Corrected Proof.
136. McDonald, W. F., J. W. Huleatt, H. G. Foellmer, D. Hewitt, J. Tang, P. Desai, A. Price, A. Jacobs, V. N. Takahashi, Y. Huang, V. Nakaar, L. Alexopoulou, E. Fikrig, and T. J. Powell. 2007. A West Nile virus recombinant protein vaccine that coactivates innate and adaptive immunity. J Infect Dis 195:1607-1617.
137. Wille-Reece, U., B. J. Flynn, K. Lore, R. A. Koup, R. M. Kedl, J. J. Mattapallil, W. R. Weiss, M. Roederer, and R. A. Seder. 2005. HIV Gag protein conjugated to a Toll-like receptor 7/8 agonist improves the magnitude and quality of Th1 and CD8+ T cell responses in nonhuman primates. Proc. Natl. Acad. Sci. U. S. A 102:15190-15194.
91

138. Wille-Reece, U., C. y. Wu, B. J. Flynn, R. M. Kedl, and R. A. Seder. 2005. Immunization with HIV-1 Gag Protein Conjugated to a TLR7/8 Agonist Results in the Generation of HIV-1 Gag-Specific Th1 and CD8+ T Cell Responses. J. Immunol. 174:7676-7683.
139. Davila, E., R. Kennedy, and E. Celis. 2003. Generation of Antitumor Immunity by Cytotoxic T Lymphocyte Epitope Peptide Vaccination, CpG-oligodeoxynucleotide Adjuvant, and CTLA-4 Blockade. Cancer Res 63:3281-3288.
140. Schwarz, K., T. Storni, V. Manolova, A. Didierlaurent, J. C. Sirard, P. Rothlisberger, and M. F. Bachmann. 2003. Role of Toll-like receptors in costimulating cytotoxic T cell responses. Eur. J. Immunol. 33:1465-1470.
141. Ben Yedidia, T., R. Tarrab-Hazdai, D. Schechtman, and R. Arnon. 1999. Intranasal Administration of Synthetic Recombinant Peptide-Based Vaccine Protects Mice from Infection by Schistosoma mansoni. Infect. Immun. 67:4360-4366.
142. Cunningham, A. F., M. Khan, J. Ball, K. M. Toellner, K. Serre, E. Mohr, and I. C. MacLennan. 2004. Responses to the soluble flagellar protein FliC are Th2, while those to FliC on Salmonella are Th1. Eur. J Immunol 34:2986-2995.
143. McEwen, J., R. Levi, R. J. Horwitz, and R. Arnon. 1992. Synthetic recombinant vaccine expressing influenza haemagglutinin epitope in Salmonella flagellin leads to partial protection in mice. Vaccine 10:405-411.
144. McSorley, S. J., B. D. Ehst, Y. Yu, and A. T. Gewirtz. 2002. Bacterial Flagellin Is an Effective Adjuvant for CD4+ T Cells In Vivo. J Immunol 169:3914-3919.
145. Saha, S., F. Takeshita, T. Matsuda, N. Jounai, K. Kobiyama, T. Matsumoto, S. Sasaki, A. Yoshida, K. Q. Xin, D. M. Klinman, S. Uematsu, K. J. Ishii, S. Akira, and K. Okuda. 2007. Blocking of the TLR5 Activation Domain Hampers Protective Potential of Flagellin DNA Vaccine. Journal of Immunology 179:1147-1154.
146. Strindelius, L., M. Filler, and I. Sjoholm. 2004. Mucosal immunization with purified flagellin from Salmonella induces systemic and mucosal immune responses in C3H/HeJ mice. Vaccine 22:3797-3808.
147. Asahi-Ozaki, Y., S. Itamura, T. Ichinohe, P. Strong, S. i. Tamura, H. Takahashi, H. Sawa, M. Moriyama, M. Tashiro, T. Sata, T. Kurata, and H. Hasegawa. 2006. Intranasal administration of adjuvant-combined recombinant influenza virus HA vaccine protects mice from the lethal H5N1 virus infection. Microbes and Infection 8:2706-2714.
148. Llopiz, D., J. Dotor, A. Zabaleta, J. Lasarte, J. s. Prieto, F. Borr+ís-Cuesta, and P. Sarobe. Combined immunization with adjuvant molecules poly(I:C) and anti-
92

CD40 plus a tumor antigen has potent prophylactic and therapeutic antitumor effects. Cancer Immunology, Immunotherapy.
149. Mizel, S. B., A. P. West, and R. R. Hantgan. 2003. Identification of a sequence in human toll-like receptor 5 required for the binding of Gram-negative flagellin. J Biol. Chem. 278:23624-23629.
150. Moors, M. A., L. Li, and S. B. Mizel. 2001. Activation of interleukin-1 receptor-associated kinase by gram-negative flagellin. Infect. Immun. 69:4424-4429.
151. Hayashi, F., K. D. Smith, A. Ozinsky, T. R. Hawn, E. C. Yi, D. R. Goodlett, J. K. Eng, S. Akira, D. M. Underhill, and A. Aderem. 2001. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 410:1099-1103.
152. Ben Yedidia, T., H. Marcus, Y. Reisner, and R. Arnon. 1999. Intranasal administration of peptide vaccine protects human/mouse radiation chimera from influenza infection. Int. Immunol. 11:1043-1051.
153. Sfondrini, L., A. Rossini, D. Besusso, A. Merlo, E. Tagliabue, S. Menard, and A. Balsari. 2006. Antitumor Activity of the TLR-5 Ligand Flagellin in Mouse Models of Cancer. J Immunol 176:6624-6630.
154. Mizel, S. B., A. H. Graff, N. Sriranganathan, S. Ervin, C. J. Lees, M. O. Lively, R. R. Hantgan, M. J. Thomas, J. Wood, and B. Bell. 2008. A Fusion Protein, Flagellin/F1/V, is an Effective Plague Vaccine in Mice and Two Species of Nonhuman Primates. Clin. Vaccine Immunol.CVI.
155. Levi, R. and R. Arnon. 1996. Synthetic recombinant influenza vaccine induces efficient long-term immunity and cross-strain protection. Vaccine 14:85-92.
156. Ben Yedidia, T. and R. Arnon. 2005. Towards an epitope-based human vaccine for influenza. Hum. Vaccin. 1:95-101.
157. Bates, J. T., S. Uematsu, S. Akira, and S. B. Mizel. 2009. Direct Stimulation of tlr5+/+ CD11c+ Cells Is Necessary for the Adjuvant Activity of Flagellin. J Immunol 182:7539-7547.
158. Pier, G. B., J. M. Saunders, P. Ames, M. S. Edwards, H. Auerbach, J. Goldfarb, D. P. Speert, and S. Hurwitch. 1987. Opsonophagocytic killing antibody to Pseudomonas aeruginosa mucoid exopolysaccharide in older noncolonized patients with cystic fibrosis. N Engl J Med 317:793-798.
159. Pollack, M., N. L. Koles, M. J. Preston, B. J. Brown, and G. B. Pier. 1995. Functional properties of isotype-switched immunoglobulin M (IgM) and IgG monoclonal antibodies to Pseudomonas aeruginosa lipopolysaccharide. Infect. Immun. 63:4481-4488.
93

160. Toder, D. S. 1994. Gene replacement in Pseudomonas aeruginosa. Methods Enzymol. 235:466-474.
161. Sundin, C., M. C. Wolfgang, S. Lory, A. Forsberg, and E. Frithz-Lindsten. 2002. Type IV pili are not specifically required for contact dependent translocation of exoenzymes by Pseudomonas aeruginosa. Microbial Pathogenesis 33:265-277.
162. Warrens, A. N., M. D. Jones, and R. I. Lechler. 1997. Splicing by overlap extension by PCR using asymmetric amplification: an improved technique for the generation of hybrid proteins of immunological interest. Gene 186:29-35.
163. Malhotra, S., L. A. Silo-Suh, K. Mathee, and D. E. Ohman. 2000. Proteome Analysis of the Effect of Mucoid Conversion on Global Protein Expression in Pseudomonas aeruginosa Strain PAO1 Shows Induction of the Disulfide Bond Isomerase, DsbA. J. Bacteriol. 182:6999-7006.
164. Hoang, T. T., R. R. Karkhoff-Schweizer, A. J. Kutchma, and H. P. Schweizer. 1998. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 212:77-86.
165. Bates, J. T., A. N. Honko, A. H. Graff, N. D. Kock, and S. B. Mizel. 2008. Mucosal adjuvant activity of flagellin in aged mice. Mechanisms of Ageing and Development 129:271-281.
166. Macdonald, R. A., C. S. Hosking, and C. L. Jones. 1988. The measurement of relative antibody affinity by ELISA using thiocyanate elution. J Immunol Methods 106:191-194.
167. Slifka, M. K. and R. Ahmed. 1996. Limiting dilution analysis of virus-specific memory B cells by an ELISPOT assay. Journal of Immunological Methods 199:37-46.
168. Benner, R., A.van Oudenaren, and G.Koch. Induction of antibodyformation in mouse bone marrow., pp. 247-262.
169. Ciacci-Woolwine, F., P. F. McDermott, and S. B. Mizel. 1999. Induction of Cytokine Synthesis by Flagella from Gram-Negative Bacteria May Be Dependent on the Activation or Differentiation State of Human Monocytes. Infect. Immun. 67:5176-5185.
170. Kikuchi, T. and R. G. Crystal. 2001. Antigen-pulsed dendritic cells expressing macrophage-derived chemokine elicit Th2 responses and promote specific humoral immunity. J. Clin. Invest. 108:917-927.
171. Worgall, S., T. Kikuchi, R. Singh, K. Martushova, L. Lande, and R. G. Crystal. 2001. Protection against Pulmonary Infection with Pseudomonas aeruginosa
94

following Immunization with P. aeruginosa-Pulsed Dendritic Cells. Infect. Immun. 69:4521-4527.
172. Garrett, E. S., D. Perlegas, and D. J. Wozniak. 1999. Negative Control of Flagellum Synthesis in Pseudomonas aeruginosa Is Modulated by the Alternative Sigma Factor AlgT (AlgU). J. Bacteriol. 181:7401-7404.
173. Musher, D. M. 1992. Infections caused by Streptococcus pneumoniae: clinical spectrum, pathogenesis, immunity, and treatment. Clin Infect Dis 14:801-807.
174. Abeyta, M., G. G. Hardy, and J. Yother. 2003. Genetic Alteration of Capsule Type but Not PspA Type Affects Accessibility of Surface-Bound Complement and Surface Antigens of Streptococcus pneumoniae. Infect. Immun. 71:218-225.
175. Heurtault, B. +., P. Gentine, J. S. b. Thomann, C. Baehr, B. t. Frisch, and F. Pons. Design of a Liposomal Candidate Vaccine Against Pseudomonas aeruginosa and its Evaluation in Triggering Systemic and Lung Mucosal Immunity. Pharmaceutical Research.
176. von Specht, B. U., B. Knapp, G. Muth, M. Broker, K. D. Hungerer, K. D. Diehl, K. Massarrat, A. Seemann, and H. Domdey. 1995. Protection of immunocompromised mice against lethal infection with Pseudomonas aeruginosa by active or passive immunization with recombinant P. aeruginosa outer membrane protein F and outer membrane protein I fusion proteins. Infect. Immun. 63:1855-1862.
177. Tertilt, C., J. Joh, A. Krause, P. Chou, K. Schneeweiss, R. G. Crystal, and S. Worgall. 2009. Expression of B Cell Activating Factor Enhances Protective Immunity of a Vaccine Against P. aeruginosa. Infect. Immun.IAI.
178. Holder, I. A. and J. G. Naglich. 1986. Experimental studies of the pathogenesis of infections due to Pseudomonas aeruginosa: immunization using divalent flagella preparations. J Trauma 26:118-122.
179. von Specht, B. U., J. Gabelsberger, B. Knapp, E. Hundt, H. Schmidt-Pilger, S. Bauernsachs, U. Lenz, and H. Domdey. 2000. Immunogenic efficacy of differently produced recombinant vaccines candidates against Pseudomonas aeruginosa infections. Journal of Biotechnology 83:3-12.
180. Lee, N. G., S. B. Jung, B. Y. Ahn, Y. G. Kim, Y. Lee, Y. J. Jeon, and W. J. Park. 1999. Protection of mice against P. aeruginosa infections by large-scale affinity-purified human IgG specific to P. aeruginosa outer membrane proteins. Vaccine 18:665-674.
181. Pier, G. B., G. Meluleni, and J. B. Goldberg. 1995. Clearance of Pseudomonas aeruginosa from the murine gastrointestinal tract is effectively mediated by O-antigen-specific circulating antibodies. Infect. Immun. 63:2818-2825.
95

182. Moser, C., P. O. Jensen, T. Pressler, B. Frederiksen, S. Lanng, A. Kharazmi, C. Koch, and N. Hoiby. 2005. Serum concentrations of GM-CSF and G-CSF correlate with the Th1/Th2 cytokine response in cystic fibrosis patients with chronic Pseudomonas aeruginosa lung infection. APMIS 113:400-409.
183. Moser, C., S. Kjaergaard, T. Pressler, A. Kharazmi, C. Koch, and N. Hoiby. 2000. The immune response to chronic Pseudomonas aeruginosa lung infection in cystic fibrosis patients is predominantly of the Th2 type. APMIS 108:329-335.
184. Hartl, D., M. Griese, M. Kappler, G. Zissel, D. Reinhardt, C. Rebhan, D. J. Schendel, and S. Krauss-Etschmann. 2006. Pulmonary TH2 response in Pseudomonas aeruginosa-infected patients with cystic fibrosis. Journal of Allergy and Clinical Immunology 117:204-211.
185. Brazova, J., A. Sediva, D. Pospisilova, V. Vavrova, P. Pohunek, J. Macek, J. Bartunkova, and H. Lauschmann. 2005. Differential cytokine profile in children with cystic fibrosis. Clinical Immunology 115:210-215.
186. Pressler, T., S. S. Pedersen, F. Espersen, N. Hoiby, and C. Koch. 1992. IgG subclass antibody responses to alginate from Pseudomonas aeruginosa in patients with cystic fibrosis and chronic P. aeruginosa infection. Pediatr. Pulmonol. 14:44-51.
187. Pressler, T., S. S. Pedersen, F. Espersen, N. Hoiby, and C. Koch. 1990. IgG subclass antibodies to Pseudomonas aeruginosa in sera from patients with chronic Ps. aeruginosa infection investigated by ELISA. Clin Exp Immunol 81:428-434.
188. West, S. E., L. Zeng, B. L. Lee, M. R. Kosorok, A. Laxova, M. J. Rock, M. J. Splaingard, and P. M. Farrell. 2002. Respiratory infections with Pseudomonas aeruginosa in children with cystic fibrosis: early detection by serology and assessment of risk factors. JAMA 287:2958-2967.
189. Kosorok, M. R., L. Zeng, S. E. West, M. J. Rock, M. L. Splaingard, A. Laxova, C. G. Green, J. Collins, and P. M. Farrell. 2001. Acceleration of lung disease in children with cystic fibrosis after Pseudomonas aeruginosa acquisition. Pediatr. Pulmonol. 32:277-287.
190. Polanec, J., J. Patzer, J. Grzybowski, M. Strukelj, and Z. P. Pavelic. 1997. Amount and Avidity of IgG Antibodies to Pseudomonas aeruginosa Exotoxin A Antigen in Cystic Fibrosis Patients. Pathol Oncol Res 3:26-29.
191. Fick, R. B., Jr., G. P. Naegel, S. U. Squier, R. E. Wood, J. B. Gee, and H. Y. Reynolds. 1984. Proteins of the cystic fibrosis respiratory tract. Fragmented immunoglobulin G opsonic antibody causing defective opsonophagocytosis. J Clin Invest 74:236-248.
96

192. Ciofu, O., T. D. Petersen, P. Jensen, and N. Hoiby. 1999. Avidity of anti-P. aeruginosa antibodies during chronic infection in patients with cystic fibrosis. Thorax 54:141-144.
193. Bruderer, U., S. J. Cryz, Jr., U. B. Schaad, M. Deusinger, J. U. Que, and A. B. Lang. 1992. Affinity constants of naturally acquired and vaccine-induced anti-Pseudomonas aeruginosa antibodies in healthy adults and cystic fibrosis patients. J Infect Dis 166:344-349.
194. Bucks, C. M., J. A. Norton, A. C. Boesteanu, Y. M. Mueller, and P. D. Katsikis. 2009. Chronic Antigen Stimulation Alone Is Sufficient to Drive CD8+ T Cell Exhaustion. J Immunol 182:6697-6708.
195. Mueller, S. N. and R. Ahmed. 2009. High antigen levels are the cause of T cell exhaustion during chronic viral infection. PNAS.
196. Wat, D., C. Gelder, S. Hibbitts, I. Bowler, M. Pierrepoint, R. Evans, and I. Doull. 2007. Is there a role for influenza vaccination in cystic fibrosis? J Cyst. Fibros.
197. Chemnitz, J. M., J. Driesen, S. Classen, J. L. Riley, S. Debey, M. Beyer, A. Popov, T. Zander, and J. L. Schultze. 2006. Prostaglandin E2 Impairs CD4+ T Cell Activation by Inhibition of lck: Implications in Hodgkin's Lymphoma. Cancer Res 66:1114-1122.
198. Muhlebach, M. S., P. W. Stewart, M. W. Leigh and T. L. Noah. 1999. Quantitation of Inflammatory Responses to Bacteria in Young Cystic Fibrosis and Control Patients. Am. J. Respir. Crit. Care Med. 160:186-191.
199. Guilbault, C., J. P. Novak, P. Martin, M. L. Boghdady, Z. Saeed, M. C. Guiot, T. J. Hudson, and D. Radzioch. 2006. Distinct pattern of lung gene expression in the Cftr-KO mice developing spontaneous lung disease compared with their littermate controls. Physiol. Genomics 25:179-193.
200. CDC. 2009 Child & Adolescent Immunization Schedules. 2-26-2009.
201. Heath, P. T., J. Bowen-Morris, D. Griffiths, H. Griffiths, D. W. M. Crook, and E. R. Moxon. 1997. Antibody persistence and Haemophilus influenzae type b carriage after infant immunisation with PRP-T. Arch Dis Child 77:488-492.
202. Sell, S. H., P. F. Wright, W. K. Vaughn, J. Thompson, and G. Schiffman. 1981. Clinical studies of pneumococcal vaccines in infants. I. Reactogenicity and immunogenicity of two polyvalent polysaccharide vaccines. Rev Infect Dis 3 Suppl:S97-107.
203. Gans, H. A., A. M. Arvin, J. Galinus, L. Logan, R. DeHovitz, and Y. Maldonado. 1998. Deficiency of the Humoral Immune Response to Measles Vaccine in Infants Immunized at Age 6 Months. JAMA 280:527-532.
97

204. Davidkin, I., S. Jokinen, M. Broman, P. Leinikki, and H. Peltola. 2008. Persistence of measles, mumps, and rubella antibodies in an MMR-vaccinated cohort: a 20-year follow-up. J Infect Dis 197:950-956.
205. Booy, R., S. Hodgson, H. Griffiths, H. M. Chapel, and E. R. Moxon. 1993. Antibody persistence after accelerated immunisation against Haemophilus influenzae type b. BMJ 306:971-972.
206. Booy, R., S. A. Taylor, S. R. Dobson, D. Isaacs, G. Sleight, S. Aitken, H. Griffiths, H. Chapel, R. T. Mayon-White, and J. A. Macfarlane. 1992. Immunogenicity and safety of PRP-T conjugate vaccine given according to the British accelerated immunisation schedule. Arch Dis Child 67:475-478.
207. Slifka, M. K., R. Antia, J. K. Whitmire, and R. Ahmed. 1998. Humoral Immunity Due to Long-Lived Plasma Cells. Immunity 8:363-372.
208. Slifka, M. K. and R. Ahmed. 1996. Long-term antibody production is sustained by antibody-secreting cells in the bone marrow following acute viral infection. Ann. N Y. Acad. Sci. 797:166-176.
209. Amanna, I. J., N. E. Carlson, and M. K. Slifka. 2007. Duration of Humoral Immunity to Common Viral and Vaccine Antigens. N Engl J Med 357:1903-1915.
210. Pollard, A. J., K. P. Perrett, and P. C. Beverley. 2009. Maintaining protection against invasive bacteria with protein-polysaccharide conjugate vaccines. Nat Rev Immunol advanced online publication.
211. Wols, H. A. M., G. H. Underhill, G. S. Kansas, and P. L. Witte. 2002. The Role of Bone Marrow-Derived Stromal Cells in the Maintenance of Plasma Cell Longevity. J Immunol 169:4213-4221.
212. Pihlgren, M., N. Schallert, C. Tougne, P. Bozzotti, J. Kovarik, A. Fulurija, M. Kosco-Vilbois, P. H. Lambert, and C. A. Siegrist. 2001. Delayed and deficient establishment of the long-term bone marrow plasma cell pool during early life. Eur J Immunol 31:939-946.
213. Pihlgren, M., C. Tougne, P. Bozzotti, A. Fulurija, M. A. Duchosal, P. H. Lambert, and C. A. Siegrist. 2003. Unresponsiveness to Lymphoid-Mediated Signals at the Neonatal Follicular Dendritic Cell Precursor Level Contributes to Delayed Germinal Center Induction and Limitations of Neonatal Antibody Responses to T-Dependent Antigens. J Immunol 170:2824-2832.
214. Pihlgren, M., M. Friedli, C. Tougne, A. F. Rochat, P. H. Lambert, and C. A. Siegrist. 2006. Reduced ability of neonatal and early-life bone marrow stromal cells to support plasmablast survival. J Immunol 176:165-172.
215. Belnoue, E., M. Pihlgren, T. L. McGaha, C. Tougne, A. F. Rochat, C. Bossen, P. Schneider, B. Huard, P. H. Lambert, and C. A. Siegrist. 2008. APRIL is critical
98

for plasmablast survival in the bone marrow and poorly expressed by early-life bone marrow stromal cells. Blood 111:2755-2764.
216. O'Connor, B. P., V. S. Raman, L. D. Erickson, W. J. Cook, L. K. Weaver, C. Ahonen, L. L. Lin, G. T. Mantchev, R. J. Bram, and R. J. Noelle. 2004. BCMA Is Essential for the Survival of Long-lived Bone Marrow Plasma Cells. J. Exp. Med. 199:91-98.
217. Smith, K. G., A. Light, G. J. Nossal, and D. M. Tarlinton. 1997. The extent of affinity maturation differs between the memory and antibody-forming cell compartments in the primary immune response. EMBO J 16:2996-3006.
218. Gatto, D., T. Pfister, A. Jegerlehner, S. W. Martin, M. Kopf, and M. F. Bachmann. 2005. Complement receptors regulate differentiation of bone marrow plasma cell precursors expressing transcription factors Blimp-1 and XBP-1. J. Exp. Med. 201:993-1005.
219. Kelly, D. F., A. J. Pollard, and E. R. Moxon. 2005. Immunological Memory: The Role of B Cells in Long-term Protection Against Invasive Bacterial Pathogens. JAMA 294:3019-3023.
220. Amanna, I. J., M. K. Slifka, and S. Crotty. 2006. Immunity and immunological memory following smallpox vaccination. Immunol Rev 211:320-337.
221. Rohner, G. B., M. D. Snape, D. F. Kelly, T. John, A. Morant, L. M. Yu, A. Borkowski, F. Ceddia, R. Borrow, C. A. Siegrist, and A. J. Pollard. 2008. The Magnitude of the Antibody and Memory B Cell Responses during Priming with a Protein-Polysaccharide Conjugate Vaccine in Human Infants Is Associated with the Persistence of Antibody and the Intensity of Booster Response. J Immunol 180:2165-2173.
222. Gatto, D., S. W. Martin, J. Bessa, E. Pellicioli, P. Saudan, H. J. Hinton, and M. F. Bachmann. 2007. Regulation of Memory Antibody Levels: The Role of Persisting Antigen versus Plasma Cell Life Span. J. Immunol. 178:67-76.
223. Bernasconi, N. L., E. Traggiai, and A. Lanzavecchia. 2002. Maintenance of Serological Memory by Polyclonal Activation of Human Memory B Cells. Science 298:2199-2202.
224. Lee, C. Y., J. Thipphawong, L. M. Huang, P. I. Lee, H. H. Chiu, W. Lin, H. Debois, D. Harrison, F. Xie, and L. Barreto. 1999. An evaluation of the safety and immunogenicity of a five-component acellular pertussis, diphtheria, and tetanus toxoid vaccine (DTaP) when combined with a Haemophilus influenzae type b-tetanus toxoid conjugate vaccine (PRP-T) in Taiwanese infants. Pediatrics 103:25-30.
225. Lin, T. Y., Y. H. Wang, L. Y. Chang, Y. C. Huang, H. T. Kao, P. Y. Lin, H. K. Lu, P. Chavand, and E. Ortiz. 2007. A fully liquid diphtheria-tetanus-five
99

component acellular pertussis-inactivated poliomyelitis-Haemophilus influenzae type b conjugate vaccine: immunogenicity and safety of primary vaccination in Taiwanese infants. International Journal of Infectious Diseases 11:129-136.
226. Thisyakorn, U., C. Pancharoen, S. Chuenkitmongkol, and E. Ortiz. 2009. Immunogenicity and safety of a DTaP-IPV//PRP-T vaccine (Pentaxim) booster during the second year of life in Thai children primed with an acellular pertussis combined vaccine. Southeast Asian J Trop. Med Public Health 40:282-294.
227. Ma, L., M. Conover, H. Lu, M. R. Parsek, K. Bayles, and D. J. Wozniak. 2009. Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog 5:e1000354.
228. Overhage, J., M. Schemionek, J. S. Webb, and B. H. A. Rehm. 2005. Expression of the psl Operon in Pseudomonas aeruginosa PAO1 Biofilms: PslA Performs an Essential Function in Biofilm Formation. Appl. Environ. Microbiol. 71:4407-4413.
100

SCHOLASTIC VITAE NAME: Eric T. Weimer ADDRESS: Department of Microbiology and Immunology Wake Forest University Health Sciences Medical Center Boulevard Winston-Salem, North Carolina 27157-1064 Telephone - (336) 716-4138 Email – [email protected] EDUCATION: 2001-2005 East Carolina University Greenville, NC B.S. (Biology) Magna Cum Laude 2005-2009 Wake Forest University School of Medicine Winston Salem, NC Ph.D. (Microbiology & Immunology)
"Development of a multivalent flagellin-based Pseudomonas aeruginosa vaccine"
Steven B. Mizel, Ph.D., Advisor PROFESSIONAL MEMBERSHIPS: American Association of Immunologists BIBLIOGRAPHY: E.T. Weimer, S. Ervin, D. Wozniak, S. Mizel. Immunization of young African green
monkeys with OprF epitope 8-OprI-type A- and B-flagellin fusion proteins promotes the production of protective antibodies against nonmucoid Pseudomonas aeruginosa. Submitted to Vaccine, May 2009.
E.T. Weimer, H. Lu, N. Kock, D. Wozniak, S. Mizel. A fusion protein vaccine
containing OprF epitope 8-OprI-type A- and B-flagellin promotes enhanced
101

clearance of nonmucoid Pseudomonas aeruginosa. Infect Immun. June 2009 77:2356-2366.
R.D. Yammani, S. Pejawar-Gaddy, T.G. Gurley, E.T. Weimer, E.M. Hiltbold, M.A.
Alexander-Miller. Regulation of maturation and activating potential in CD8+ versus CD8- dendritic cells following in vivo infection with vaccinia virus. Virology. 2008 Aug 15;378(1):142-50.
102