detection of poliovirus, hepatitis avirus, sewage …aem.asm.org/content/60/7/2400.full.pdf ·...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, JUly 1994, p. 2400-2407 Vol. 60, No. 70099-2240/94/$04.00+0Copyright © 1994, American Society for Microbiology
Detection of Poliovirus, Hepatitis A Virus, and Rotavirusfrom Sewage and Ocean Water by Triplex Reverse
Transcriptase PCRYU-LI TSAI,* BICH TRAN, LOUIS R. SANGERMANO, AND CAROL J. PALMER
Environmental Sciences Laboratory, County Sanitation Districts of Orange County, Fountain Valley, Califomia 92728
Received 31 January 1994/Accepted 6 May 1994
A triplex reverse transcriptase PCR (RT-PCR) was developed to simultaneously detect poliovirus, hepatitisA virus (HAV), and rotavirus in sewage and ocean water. Sewage and ocean water samples seeded with thethree different viruses were concentrated by ultrafiltration. The unseeded ocean water and sewage samples wereconcentrated by vortex flow filtration and/or ultrafiltration. Random hexamers and a rotavirus downstreamprimer were used to initiate reverse transcription. Three different sets of primers specific for poliovirus, HAV,and rotavirus cDNAs were mixed in the PCR mixture to amplify the target DNA. Three distinct amplified DNAproducts representing poliovirus, HAV, and rotavirus were identified by gel electrophoresis as 394-, 192-, and278-bp sequences, respectively. Dot blot and Southern analyses were used to confirm the amplified products foreach virus present in the environmental samples. Except for poliovirus, the sensitivity of triplex RT-PCR forthe detection of rotavirus and HAV was found to be similar to that of monoplex RT-PCR, which uses only oneset of primers to amplify a single type of virus. The triplex RT-PCR has greater advantages over monoplexRT-PCR for virus detection, namely, the rapid turnaround time and cost effectiveness.
Enteric viruses have been associated with many outbreaks ofwaterborne nonbacterial gastroenteritis (16, 18-20, 28) and arean important public health concern. It has been reported thatat least 37 different human viruses have been isolated fromdrinking water all around the world (11). The traditionaldetection of viruses in water, including monitoring treatmentremoval efficiency, is done by labor-intensive tissue culturemethodology (2). The problems with tissue culture methodsare well-known and include the lack of susceptible cell lines formany important waterborne viruses (such as Norwalk virus),lack of sensitivity, lengthy analysis time (up to 6 weeks), andproblems in detecting low virus numbers, which is the typicalsituation in environmental water samples.
Faster and more sensitive technology is now available for thedetection of viruses in both environmental and clinical sam-ples. The use of molecular techniques, such as PCR andnucleic acid hybridization, for the detection of enteric virusesin environmental samples has been well documented in recentyears (1, 3, 6, 8, 14, 15, 17, 26, 27, 30). Another rapid method,developed by Graff et al. (13), uses an antigen capture PCRtechnique to detect hepatitis A virus (HAV) from treated anduntreated sewage sludge samples. These new methods offerseveral advantages over traditional tissue culture. These ad-vantages include rapid turnaround time and a high degree ofsensitivity. Molecular techniques do not require tissue cultureor cell line maintenance. Through enzymatic amplification, thePCR is capable of detecting specific genes from one viablebacterial cell containing 10-15 g of DNA in 100 ml of water (5)and the viral genomic RNA in diluents containing less than 1PFU (9, 26). Because the PCR does not require cell cultiva-tion, it has been effectively used to detect viruses which aredifficult to cultivate or are nonculturable (8, 14, 26, 27), such asHAV and Norwalk virus from sewage or human stools. The
* Corresponding author. Mailing address: Environmental SciencesLaboratory, County Sanitation Districts of Orange County, 10844 EllisAve., Fountain Valley, CA 92728. Phone: (714) 962-2411. Fax: (714)962-2591.
PCR has been used to detect pathogenic microorganisms indifferent environments and it could be used as an analyticaltool to indicate the potential of disease outbreaks.
Conventional reverse transcriptase PCR (RT-PCR) ismonoplex and includes only one specific set of primers in thereaction mixture and can detect only one virus or one targetRNA sequence in a sample (1, 3, 9, 12, 15, 22, 26). MultiplexPCR is the simultaneous PCR amplification of gene sequencesassociated with different organisms or different genes withinthe same organism. This technique has been used to detectdual genes from Escherichia coli or Legionella pneumophila (4)in water and to detect different Salmonella spp. in soil andwater (29). In this study, we have developed a triplex RT-PCRmethod, which uses three different sets of primers in onereaction mixture, to detect poliovirus, HAV, and rotavirussimultaneously in sewage and ocean water. Because of the timesavings in time and cost-effectiveness, the triplex RT-PCRmethod provides a more efficient way to detect various viralRNAs in environmental samples than the monoplex RT-PCRmethod.
MATERUILS AND METHODS
Samples. Ocean water samples were collected from bothcoastal and offshore waters in sterile 20-liter polypropylenecontainers. Sewage samples from primary influent and second-ary effluent were collected in 250-ml sterile polypropylenecontainers. All samples were stored on ice. For virus detection,the ocean water samples (15 liters) and sewage samples (100ml) were concentrated within 4 h after collection by vortex flowfiltration and ultrafiltration as described previously (26).
Virus strains. Poliovirus type 1 strain LSc was maintained inBuffalo green monkey kidney cells, and HAV strain HM175was maintained on BS-C-1 cells (an African green monkeykidney-derived cell line) and/or FRhK-4 cells (fetal rhesuskidney-derived cell line). Two human rotaviruses (VR-2018strain Wa and VR-970 strain D) were purchased from Amer-ican Type Culture Collection, Rockville, Md. One human
2400
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

TRIPLEX RT-PCR ON SEWAGE AND OCEAN WATER 2401
rotavirus provided by National Institutes of Health and oneanimal strain rotavirus SA-1 1 were used for the determinationof primer specificity. All rotaviruses were propagated on theMA-104 cell lines.
Virus seeding. A virus mixture (30,ul) containing 1.0 x 104PFU of HAV strain HM175, 6.0x 104 PFU of poliovirus type1 strain LSc, and 1.6 x 104 PFU of rotavirus strain Wa (ATCCVR-2018) was seeded into 100-ml portions of filtered andnonfiltered samples. The filtered samples, including bothocean water and secondary sewage effluent, were filter steril-ized with 0.2-,um-pore-size Nalgene disposable filterware(Nalge Co., Rochester, N.Y.). The seeded and unseededsamples (100-ml portions) were concentrated by ultrafiltrationwith Centriprep-100 and Centricon-100 (Amicon, Inc., Bev-erly, Mass.) as described previously (26). The final concen-trates (2,ul) were used as templates for the triplex RT-PCR.The remaining concentrates were stored at -20°C for futureanalysis.
Triplex RT-PCR. The triplex RT-PCR was performed with aGeneAmp PCR System 9600 (Perkin-Elmer, Norwalk, Conn.).All the reagents required for RT-PCR were included in aGeneAmp RNA PCR kit (Perkin-Elmer). Each RT reactionmixture (16,ul) contained the following: 4 RI of 25mM MgCl2;2 RI of 1Ox PCR buffer 11 (500 mM KCl, 100 mM Tris-HCl,pH 8.3); 2 RI each of 10 mM dGTP, 10 mM dATP, 10 mMdTTP, and 10 mM dCTP; 1 ,ul of 50,uM random hexamers;and 1 RI of 10,uM End9 primer (12), 5'-GGTCACATCATACAATTCTAATC TAAG-3'. Two microliters of concentratedsample was added to the RT reaction mixture and subse-quently heated at 99°C for 5 min and cooled at 4°C for 5 minbefore the addition of 1 [I of RT (50 U/pA) and 1 pA of RNaseinhibitor (20 U/,ul). The reverse transcription was completedafter the following steps: 25°C for 10 min, 42°C for 30 min and4°C for 5 min. The PCR mixture (80 pA) containing AmpliTaqDNA polymerase and three sets of oligonucleotide primerswas added to the finished RT mixture (20 pI). Except for theprimers, all other components used in the PCR process werekept at optimal concentrations as suggested by the manufac-turer (Perkin-Elmer). Primers for poliovirus 5' noncodingregion (3, 10, 24) (Polio-R, 5'-ACGGACACCCAAAGTA-3';Polio-L, 5'-AGCACTTCTGTTTCCC-3'), for HAV capsidregion (21) (HAVC-R, 5'-CTCCAGAATCATCTCCAAC-3';HAVC-L, 5'-CAGCACATCAGAAAGGTGAG-3') and for asegment of rotavirus gene encoding the major outer capsidglycoprotein vp7 (End9 and Rota785 [5'-TTCGAAATTG-TAAGAAATTAG-3']) were used to amplify 394-, 192-, and278-bp sequences, respectively. The final concentrations foreach set of primers in the reaction mixture were 50 nM. ThePCR was carried out by the following protocol: initial dena-turation step at 95°C for 2 min; 40 cycles, with 1 cycleconsisting of 1 min at 95°C, 1 min at 55°C, and 1 min at 72°C;and final extension step at 72°C for 7 min. The PCR productswere stored at 4°C before analysis. The internal oligonucleo-tide probes for poliovirus, HAV, and rotavirus PCR productswere POLIO-IN (5'-ACATAAGAATCCTCCGGCCCCTGA-3'), HAVC-IN (5'-7TfGCITCCFC-I-lTlATCATGCTAT-3') (9),and RTPB858 (5'-CATAACAGCAGATCCAACAAC-3'), re-spectively. All oligonucleotides were synthesized by a DNA/RNA synthesizer (model 392; Perkin-Elmer, Applied Biosys-tems Division, Foster City, Calif.). The RT-PCR internalprobes, POLIO-IN and RTPB858, and the PCR primer,Rota785, were designed by using OLIGO 4.1 computer soft-ware (National Biosciences, Plymouth, Minn.).DNA hybridization on PCR products. The amplified PCR
products were identified under UV irradiation by electro-phoresis on a 2% SeaKem GTG agarose gel (FMC BioProd-
ucts, Rockland, Maine) stained with ethidium bromide (0.5,ug/ml). The amplified DNA was transferred onto Hybond-N+positively charged nylon membranes (Amersham, ArlingtonHeights, Ill.) by Southern blotting (23) or dot blotting (7). APosiBlot pressure blotter (Stratagene, La Jolla, Calif.) and aMinifold I dot blotter (Schleicher & Schuell, Keene, N.H.)were used for Southern blotting and dot blotting, respectively.All three PCR internal probes were nonradioactively labeledby using a Genius 5 DNA labeling kit (Boehringer Mannheim,Indianapolis, Ind.) by the protocols suggested by the manufac-turer. The DNA hybridization on amplified target fragmentsand chemiluminescence detection were performed as de-scribed previously (25). The triplex PCR products on themembrane were first probed with HAVC-IN. The membranewas then boiled in deionized water for 30 min to strip off theold probe and reprobed with POLIO-IN. Last, the membranewas stripped off again and reprobed with RTPB858. Theprobes were tested for cross-reactivity with the triplex PCRproducts by dot blot analysis.
Application of triplex RT-PCR on sewage and ocean waterconcentrates. The undiluted and serially diluted sewage andocean water concentrates were used as templates in the triplexRT-PCR. Positive controls, including a virus mixture (poliovi-rus, HAV, and rotavirus), each type of virus, and a RNAfragment (a plasmid pAW109 transcript) from the RT-PCR kit(Perkin Elmer), were employed to confirm the amplificationresults. A negative control containing no template was used toensure that carryover contamination did not occur in thereaction.
In a separate experiment, serially diluted virus mixturescontaining poliovirus, rotavirus, and HAV ranging from 2 x102 to 2 x 105 PFU/ml were seeded into 100-ml portions oftarget virus-free primary influent and were concentrated to200-pul for triplex RT-PCR and monoplex RT-PCR. Thesensitivity of detection for these viruses present in primaryinfluent was determined by dot blot analysis.
RESULTS
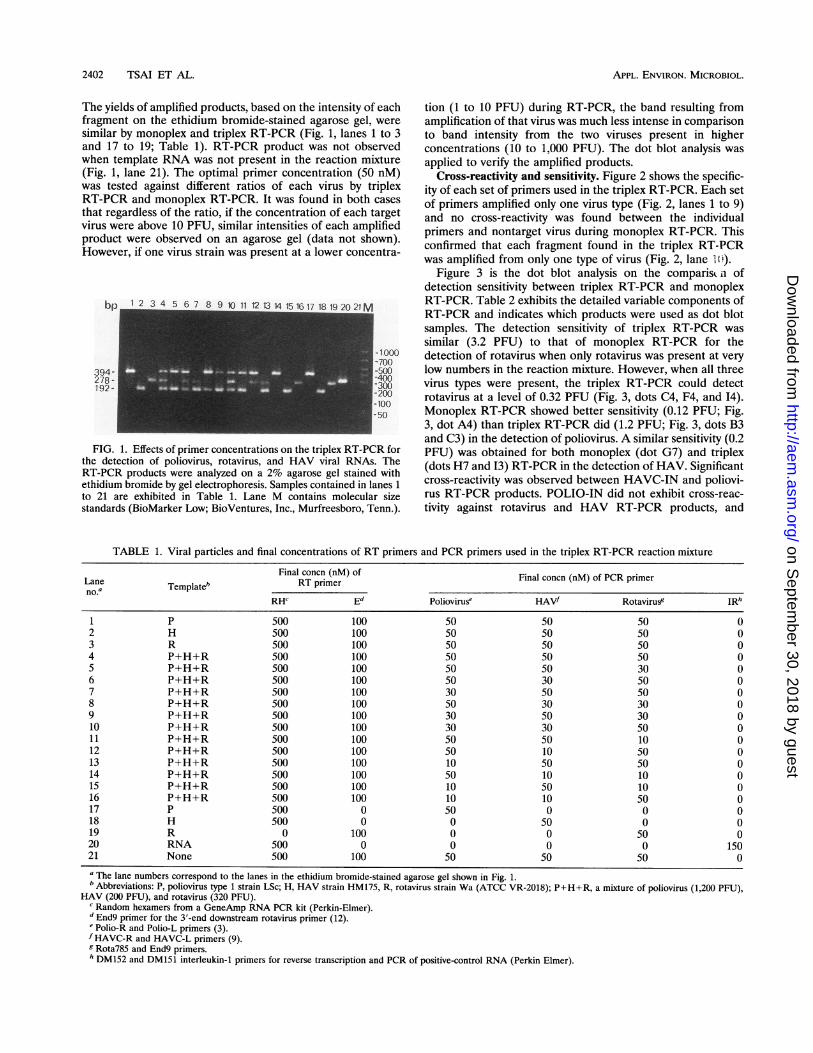
Triplex RT-PCR with various primer concentrations. Figure1 shows the effects of multiple primer concentrations on theamplification efficiencies of different viral RNAs during thetriplex RT-PCR. The amplified DNA products were 394, 278,and 192 bp long, which correspond to the sizes of the cDNAtarget fragments transcribed from viral RNAs of poliovirus,rotavirus, and HAV, respectively. The estimated initial virusnumbers in each reaction mixture were 1,200, 200, and 320PFU for poliovirus, HAV, and rotavirus, respectively. Thecomposite primer concentrations for three different virusesused in the triplex RT-PCR are listed in Table 1. When eachset of primers was initially maintained at a final concentrationof 50 nM in the reaction mixture, the best amplification resultfor all three viruses was obtained (Fig. 1, lane 4; Table 1).Although the reduction of poliovirus and HAV primer con-centrations from 50 nM to 30 nM also gave a similar result(Fig. 1, lane 10; Table 1), primer concentrations of 50 nM eachwere chosen for triplex RT-PCR throughout this study becauseof the simplicity of preparation. In addition, when the rotavirusprimer concentration was decreased to 30 nM and the othertwo sets of primers were maintained at either 30 or 50 nM,significant reduction of rotavirus RT-PCR products was found(Fig. 1, lanes 5, 8, and 9; Table 1). Moreover, when theconcentrations of one or two primer pairs were reduced to 10nM, no amplified products from corresponding primers wereobserved (Fig. 1, lanes 11 to 16; Table 1). No cross-reactivitywas found among the three primer sets for the target DNA.
VOL. 60, 1994
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
APPL. ENVIRON. MICROBIOL.
The yields of amplified products, based on the intensity of eachfragment on the ethidium bromide-stained agarose gel, weresimilar by monoplex and triplex RT-PCR (Fig. 1, lanes 1 to 3and 17 to 19; Table 1). RT-PCR product was not observedwhen template RNA was not present in the reaction mixture(Fig. 1, lane 21). The optimal primer concentration (50 nM)was tested against different ratios of each virus by triplexRT-PCR and monoplex RT-PCR. It was found in both casesthat regardless of the ratio, if the concentration of each targetvirus were above 10 PFU, similar intensities of each amplifiedproduct were observed on an agarose gel (data not shown).However, if one virus strain was present at a lower concentra-
bp 1 2 3 4 5 6 7 8 9 1112131415161718 19 20 21M
I ~~~~~~~~~~~~~~-700
2978 _ -4001 92- 2300J ~~~~~~~~~~~~~~-100
-50
FIG. 1. Effects of primer concentrations on the triplex RT-PCR forthe detection of poliovirus, rotavirus, and HAV viral RNAs. TheRT-PCR products were analyzed on a 2% agarose gel stained withethidium bromide by gel electrophoresis. Samples contained in lanes 1
to 21 are exhibited in Table 1. Lane M contains molecular sizestandards (BioMarker Low; BioVentures, Inc., Murfreesboro, Tenn.).
tion (1 to 10 PFU) during RT-PCR, the band resulting fromamplification of that virus was much less intense in comparisonto band intensity from the two viruses present in higherconcentrations (10 to 1,000 PFU). The dot blot analysis wasapplied to verify the amplified products.
Cross-reactivity and sensitivity. Figure 2 shows the specific-ity of each set of primers used in the triplex RT-PCR. Each setof primers amplified only one virus type (Fig. 2, lanes 1 to 9)and no cross-reactivity was found between the individualprimers and nontarget virus during monoplex RT-PCR. Thisconfirmed that each fragment found in the triplex RT-PCRwas amplified from only one type of virus (Fig. 2, lane 1 ).
Figure 3 is the dot blot analysis on the comparisL n ofdetection sensitivity between triplex RT-PCR and monoplexRT-PCR. Table 2 exhibits the detailed variable components ofRT-PCR and indicates which products were used as dot blotsamples. The detection sensitivity of triplex RT-PCR wassimilar (3.2 PFU) to that of monoplex RT-PCR for thedetection of rotavirus when only rotavirus was present at verylow numbers in the reaction mixture. However, when all threevirus types were present, the triplex RT-PCR could detectrotavirus at a level of 0.32 PFU (Fig. 3, dots C4, F4, and 14).Monoplex RT-PCR showed better sensitivity (0.12 PFU; Fig.3, dot A4) than triplex RT-PCR did (1.2 PFU; Fig. 3, dots B3and C3) in the detection of poliovirus. A similar sensitivity (0.2PFU) was obtained for both monoplex (dot G7) and triplex(dots H7 and 13) RT-PCR in the detection of HAV. Significantcross-reactivity was observed between HAVC-IN and poliovi-rus RT-PCR products. POLIO-IN did not exhibit cross-reac-tivity against rotavirus and HAV RT-PCR products, and
TABLE 1. Viral particles and final concentrations of RT primers and PCR primers used in the triplex RT-PCR reaction mixture
Final concn (nM) of Final concn (nM) of PCR primerLane Templateb RT primerno.'
RH" Ed Poliovirus" HAVf Rotavirue IRh
1 P 500 100 50 50 50 02 H 500 100 50 50 50 03 R 500 100 50 50 50 04 P+H+R 500 100 50 50 50 05 P+H+R 500 100 50 50 30 06 P+H+R 500 100 50 30 50 07 P+H+R 500 100 30 50 50 08 P+H+R 500 100 50 30 30 09 P+H+R 500 100 30 50 30 010 P+H+R 500 100 30 30 50 011 P+H+R 500 100 50 50 10 012 P+H+R 500 100 50 10 50 013 P+H+R 500 100 10 50 50 014 P+H+R 500 100 50 10 10 015 P+H+R 500 100 10 50 10 016 P+H+R 500 100 10 10 50 017 P 500 0 50 0 0 018 H 500 0 0 50 0 019 R 0 100 0 0 50 020 RNA 500 0 0 0 0 15021 None 500 100 50 50 50 0" The lane numbers correspond to the lanes in the ethidium bromide-stained agarose gel shown in Fig. 1.b Abbreviations: P, poliovirus type 1 strain LSc; H, HAV strain HM175, R, rotavirus strain Wa (ATCC VR-2018); P+H+R, a mixture of poliovirus (1,200 PFU),
HAV (200 PFU), and rotavirus (320 PFU).C Random hexamers from a GeneAmp RNA PCR kit (Perkin-Elmer).d End9 primer for the 3'-end downstream rotavirus primer (12).' Polio-R and Polio-L primers (3).fHAVC-R and HAVC-L primers (9).g Rota785 and End9 primers.h DM152 and DM151 interleukin-1 primers for reverse transcription and PCR of positive-control RNA (Perkin Elmer).
2402 TSAI ET AL.
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

TRIPLEX RT-PCR ON SEWAGE AND OCEAN WATER 2403
FIG. 2. Analysis of primer specificity on target viruses in RT-PCRby gel electrophoresis. Primers included in each reaction mixture arerotavirus primers (Rota785 and End9) (lanes I to 3), HAV primers(HAVC-L and HAVC-R) (lanes 4 to 6), poliovirus primers (Polio-Land Polio-R) (lanes 7 to 9), and all three sets of primers (lane 10). Thevirus templates contained in each reaction are rotavirus (300 PFU[lanes 3, 5, and 7]), HAV (200 PFU [lanes 1, 6, and 8]), poliovirus(1,200 PFU [lanes 2, 4, and 9]), all three viruses (lane 10), RNART-PCR control from a RNA PCR kit (Perkin Elmer) (lane 11), andnegative control containing no template (lane 12). Lane M containsmolecular size standards (BioMarker Low; BioVentures, Inc.). Thenumbers to the left of the gel indicate the number of base pairs.
RTPB858 did not exhibit cross-reactivity against poliovirus andHAV RT-PCR products.
Triplex RT-PCR on sewage and ocean samples. Figure 4illustrates the application of triplex RT-PCR on ocean waterand sewage. Poliovirus, HAV, and rotavirus were not detectedin coastal water (Fig. 4, lanes 1 to 4). An amplified DNAfragment similar in size to the positive HAV product (lane 6)was found in ocean water 30 m deep around a sewage outfallarea, but both rotavirus and poliovirus were not found (Fig. 4,lanes 5 to 8). Poliovirus, HAV, and rotavirus were not detectedin filtered ocean water (lane 9), but suspicious HAV RNA wasobserved in unfiltered secondary effluent (lane 10). Both of thesuspicious HAV PCR fragments found in outfall deep-oceanwater and secondary effluent were probed with HAV internalprobe and they were all negative (Fig. 4B). However, whenunfiltered secondary effluent and filtered ocean water wereseeded with poliovirus, HAV, and rotavirus, the expectedamplified product was observed, as shown in lanes 11 and 12(Fig. 4A). They were all positive when hybridized with therespective internal probes (Fig. 4B). The amplified rotavirustarget fragments in the seeded samples were not clearlydemonstrated in the agarose gel, but they hybridized with theRTPB858 internal probe after Southern blotting (Fig. 4B).Except for positive controls, none of the three viruses weredetected in filtered ocean water and secondary effluent bymonoplex RT-PCR (lanes 14 to 22). However, more back-ground RNA or DNA from microflora in the ocean water andsecondary sewage was amplified by triplex RT-PCR than bymonoplex RT-PCR. This result suggested that the method (26)used in concentrating viruses from environmental water sam-ples was successful in removing substances in the samples thatcould cause inhibition of the PCR. Figure 4B is a compositeautoradiogram, showing three images of the Southern analysiswhen the same membrane was probed with three different
probes separately. Each internal probe positively identified itstriplex RT-PCR products amplified from seeded water samplesand respective positive control. Unlike dot blot analysis (Fig. 3,dots GI to G4 and HI to H4) no cross-reactivity betweenHAVC-IN and poliovirus RT-PCR products was observed onthe Southern analysis (Fig. 4B). The rotavirus amplified prod-ucts were dual-sized fragments on Southern blots but only asingle-sized fragment was observed on agarose gel (Fig. 4,lanes 11 to 13 and 16).
Figure 5 shows additional application of the triplex RT-PCRprocedure to two primary influent and two secondary effluentsewage samples. In this experiment, 15 liters (instead of 100ml) of sewage sample was collected and further concentratedby vortex flow filtration and ultrafiltration. Several distinctPCR products amplified from background organisms wereobserved on an agarose gel. Poliovirus, rotavirus, and HAVRNAs were detected in one undiluted and 10-fold-dilutedprimary influent concentrate (Fig. 5, lanes 5 and 6), but theywere not detected in the 100- or 1,000-fold dilutions. OnlyHAV RNA was detected in the second primary influent sample(Fig. 5, lane 1). The secondary effluent concentrates werenegative (Fig. 5, lanes 9 to 16). These results were confirmedwith subsequent Southern blot analysis (data not shown).
Table 3 shows the dot blot hybridization results using bothtriplex and monoplex RT-PCR to amplify various concentra-tions of poliovirus, rotavirus, and HAV from seeded primaryinfluent samples. Triplex RT-PCR detected viral RNAs equiv-alent to 10 PFU of poliovirus, rotavirus, and HAV seeded into100 ml of the primary influent. However, except for rotavirusand HAV, monoplex RT-PCR was able to detect polioviruslevels as low as 1 PFU in the seeded primary influent.
DISCUSSION
This study has developed a rapid and efficient method tosimultaneously detect three medically important viruses com-monly transmitted via water and/or shellfish. The efficiency oftriplex RT-PCR was carefully controlled by the concentrationof each set of primers involved in the reaction. An optimalprimer concentration of 50 nM for each primer was deter-mined as yielding the best results for the triplex RT-PCRprotocol for the detection of poliovirus, HAV, and rotavirus.Even when the viruses were present at the levels detectable byRT-PCR (200 to 1,200 PFU), no PCR products of targetviruses could be found if primer concentrations were de-creased to 10 nM. This result indicates that the lower limit ofthe primer concentration was 10 nM in the triplex RT-PCR.Conversely, when 10-fold-concentrated primers (500 nM) wereused in the reaction mixture, primers-dimers appeared andonly rotavirus RNA was amplified in the triplex reaction (datanot shown). Therefore, these results suggest that the optimi-zation of primer concentration is critical to determine theefficacy of triplex or multiplex PCR on the detection of thetarget organisms in a mixed sample. Under optimized primerconcentration and fixed virus number (10' PFU), no differencein amplification efficiency was found by triplex or monoplexRT-PCR for detection of each virus type. Moreover, theamount of each virus did not affect the amplification of theother two viruses at the optimal primer concentration. Inaddition to random hexamers, the End9 primer was required inreverse transcription to obtain successfully triplex RT-PCRresults. If the End9 oligonucleotide (downstream primer) wasnot present in the RT reaction mixture, insufficient amplifica-tion of rotavirus RNA was observed (data not shown). There-fore, the End9 primer was critical to the success of the RTreaction for the triplex PCR.
VOL. 60, 1994
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
FIG. 3. Dot blot analysis of sensitivity test on triplex RT-PCR andmonoplex RT-PCR. The variable components used in both triplex andmonoplex RT-PCR are listed in Table 2. The same Southern blot-transferred membrane was probed with each of three different RT-PCR internal probes in separate hybridizations. The hybridizationsignals shown in each row are POLIO-IN for rows A, B, and C,RTPB858 for rows D, E, and F, and HAVC-IN for rows G, H, and I.
The primer sets used in this study have proved to be veryspecific in the triplex PCR reaction. No cross-reactivity be-tween the virus primers was observed. The published primers(HAVC-R, HAVC-L, End9, Polio-R, and Polio-L), whichwere used in the triplex RT-PCR, have been screened againstother virus types and were determined to be type specific byother investigators (3, 8, 12). Therefore, it was feasible to usethese primers in the triplex RT-PCR to specifically detectpoliovirus, HAV, or rotavirus in environmental samples. Therotavirus primers used in this study were specific to rotaviruses
TABLE 2. Major components used in the triplex and monoplex RT-PCR for the dot blot analysis. Dot number shown in this tablecorresponds to the dot blot analysis in Fig. 3
RT RT-PCRDot no." TemplateO PFU primee PCR primer" TyPee
Al, Dl, Gl P 120 RH+E Ppr MA2, D2, G2 P 12 RH+E Ppr MA3, D3, G3 P 1.2 RH+E Ppr MA4, D4, G4 P 0.12 RH+E Ppr MA5, D5, G5 H 20 RH+E Hpr MA6, D6, G6 H 2 RH+E Hpr MA7, D7, G7 H 0.2 RH+E Hpr MA8, D8, G8 H 0.02 RH+E Hpr MA9, D9, G9 R 3,200 RH+E Rpr MAIO, D10, G10 R 320 RH+E Rpr MAll, Dll, Gll R 32 RH+E Rpr MA12, D12, G12 R 3.2 RH+E Rpr MBi, El, HI P 120 RH+E Ppr+Hpr+Rpr TB2, E2, H2 P 12 RH+E Ppr+Hpr+Rpr TB3, E3, H3 P 1.2 RH+E Ppr+Hpr+Rpr TB4, E4, H4 P 0.12 RH+E Ppr+Hpr+Rpr TB5, E5, H5 H 20 RH+E Ppr+Hpr+Rpr TB6, E6, H6 H 2 RH+E Ppr+Hpr+Rpr TB7, E7, H7 H 0.2 RH+E Ppr+Hpr+Rpr TB8, E8, H8 H 0.02 RH+E Ppr+Hpr+Rpr TB9, E9, H9 R 3,200 RH+E Ppr+Hpr+Rpr TB10, EIO, H10 R 320 RH+E Ppr+Hpr+Rpr TBl, Ell, H1l R 32 RH+E Ppr+Hpr+Rpr TB12, E12, H12 R 3.2 RH+E Ppr+Hpr+Rpr TCl, Fl, II P 120 RH+E Ppr+Hpr+Rpr T
H 20R 320
C2, F2, 12 P 12 RH+E Ppr+Hpr+Rpr TH 2R 32
C3, F3, 13 P 1.2 RH+E Ppr+Hpr+Rpr TH 0.2R 3.2
C4, F4, 14 P 0.12 RH+E Ppr+Hpr+Rpr TH 0.02R 0.32
C5, F5, I5 RNA RH DM151+DM152 MC6, F6, I6 NONE RH+E Ppr+Hpr+Rpr T
a The dot numbers correspond to the dot blot analysis shown in Fig. 3.b p, poliovirus type 1 strain LSc; H, HAV strain HM175; R, rotavirus strain Wa (ATCC VR-2018).' RH, random hexamers from a GeneAmp RNA PCR kit (Perkin-Elmer); E, End9 primer for the 3'-end downstream rotavirus primer (12).dPpr, Polio-R and Polio-L primers; Hpr, HAVC-R and HAVC-L primers; Rpr, Rota785 and End primers.eM, monoplex; T, triplex.
2404 TSAI ET AL.
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

TRIPLEX RT-PCR ON SEWAGE AND OCEAN WATER 2405
A.
394-278'192-
B.2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 M
FIG. 4. Detection of poliovirus, rotavirus, and HAV on sewage andocean water samples by triplex RT-PCR and monoplex RT-PCR.Lanes: 1, coastal water concentrate (CWC), undiluted; 2, 10-fold-diluted CWC; 3, 100-fold-diluted CWC; 4, 1,000-fold-diluted CWC; 5,outfall-depth ocean water concentrate (ODWC), undiluted; 6, 10-fold-diluted ODWC; 7, 100-fold-diluted ODWC; 8, 1,000-fold-dilutedODWC; 9, filtered ocean water concentrate, undiluted; 10, secondaryeffluent concentrate, undiluted; 11, concentrate from seeded (poliovi-rus [1,200 PFU], rotavirus [320 PFU1, HAV [200 PFU]) filtered oceanwater, undiluted; 12, concentrate from seeded (poliovirus [1,200 PFU],rotavirus [320 PFU], HAV [200 PFU]) secondary effluent, undiluted;13, positive control containing poliovirus (1,200 PFU), rotavirus (320PFU), and HAV (200 PFU); 14, 17, and 20, 10-fold-diluted CWC; 15,18, and 21, 10-fold-diluted ODWC; 16, positive rotavirus control (320PFU); 19, positive HAV control (200 PFU); 22, positive polioviruscontrol (1,200 PFU); 23, RNA control from a RNA PCR kit (PerkinElmer); 24, negative control containing no template; M, molecular sizestandards (BioMarker Low, BioVentures, Inc.). Lanes 1 to 13 areresults from triplex RT-PCR, and lanes 14 to 24 are the results frommonoplex RT-PCR (lanes 14 to 16, rotavirus; lanes 17 to 19, HAV;lanes 20 to 22, poliovirus). (A) Triplex and monoplex RT-PCRproducts on an ethidium bromide-stained 2% Seakem agarose gel. (B)Autoradiogram of the panel A gel produced by Southern analysisagainst poliovirus, rotavirus, or HAV internal probe and subsequentchemiluminescence detection.
of human origin. Another rotavirus primer (Pan-VG9) (21) incombination with End9 was able to amplify the viral RNA fromboth human and animal rotavirus strains, such as strain SA-11.However, because the amplified product (208 bp) was too closeto the size of HAV product (192 bp), it became indistinguish-able from the HAV products on the agarose gel. Therefore,the Rota785 primer was designed and used in the triplexRT-PCR to produce a distinct rotavirus-specific fragment (278bp).The dot blot analysis indicated that monoplex RT-PCR was
10 times more sensitive than triplex RT-PCR in detection ofpoliovirus but monoplex and triplex RT-PCR have similarsensitivities when only rotavirus or HAV was present in the
FIG. 5. PCR analysis of triplex RT-PCR on primary influent andsecondary effluent sewage samples for detection of poliovirus, rotavi-rus, and HAV on an ethidium bromide-stained agarose gel. Lanes: 1,primary influent concentrate 1 (PIC1), undiluted; 2, 10-fold-dilutedPIC1; 3, 100-fold-diluted PIC1; 4, 1,000-fold-diluted PIC1; 5, primaryinfluent concentrate 2 (PIC2), undiluted; 6, 10-fold-diluted PIC2; 7,100-fold-diluted PIC2; 8, 1,000-fold-diluted PIC2; 9, secondary effluentconcentrate 1 (SEC1), undiluted; 10, 10-fold-diluted SEC1; 11, 100-fold-diluted SEC1; 12, 1,000-fold-diluted SEC1; 13, secondary effluentconcentrate 2 (SEC2), undiluted; 14, 10-fold-diluted SEC2; 15, 100-fold-diluted SEC2; 16, 1,000-fold-diluted SEC2; 17, positive controlscontaining 200 PFU each of poliovirus, rotavirus, and HAV; 18, RNAcontrol from a RNA PCR kit (Perkin Elmer); 19, negative controlcontaining no template; M, molecular size standards (BioMarker Low;BioVentures, Inc.). The numbers to the right of the gel are in basepairs.
reaction mixture. The triplex RT-PCR was able to detect threetypes of viruses when each virus was present at levels less than1 PFU. The multiple primers present in the PCR mixture couldcause competition between primers for the target cDNA andthus affect the detection sensitivity, especially when the cDNAis present at very low concentrations. This could also impactthe detection sensitivity on the environmental water samples,because only low numbers of viruses are present in thesesamples. Therefore, steps involving the concentration of alarge volume of water are required to concentrate virusesbefore the application of triplex RT-PCR. The HAVC-INprobe slightly cross-reacted with the poliovirus product in thedot blot analysis, but this cross-reactivity was not found in theSouthern analysis. The HAVC-IN probe did confirm thecorrect HAV fragment in the triplex RT-PCR. Experimentalresults shown in Fig. 2 indicated that each set of primers usedin this study was very specific to one virus type, and hence thecross-reactivity found in the dot blot analysis was not due tononspecific primer annealing during the RT-PCR. Because theamplified poliovirus PCR product was blotted onto a verylimited area (13 mm2) in the dot blot analysis, this couldincrease the chances of a cross-reaction between HAVC-INand poliovirus PCR product. In order to solve this problem, amore stringent wash condition, raising the low-salt wash tem-perature to 55°C, was performed for the HAVC-IN, but itreduced the detection sensitivity for HAV. Therefore, thecross-reaction might be due to nonspecific hybridization be-tween probe and nontarget DNA, which was commonly foundin the nucleic acid hybridization. The viruses used for seedingwere stored at -20°C. Viruses were propagated on each cellline and counted before they were stored. The die-off rate afterstorage was not determined. Therefore, the PFU on the
VOL. 60, 1994
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 3. Triplex and Monoplex RT-PCR and dot blot analyses on a primary influent sewage seeded with different densities of poliovirus,rotavirus, and HAV'
Result' of dot blot hybridization and RT-PCRSeeded PFU of Triplex Monoplex
each virus' * _Poliovirus Rotavirus HAV Poliovirus Rotavirus HAV
2x105(1,000) + + + + + +2x104(100) + + + + + +2 x 103 (10) + + + + + +2 x 102 (1) - - + -0
a The primary influent sewage was tested for poliovirus, rotavirus, and HAV and found to be free of these virus before the seeding.b Only 1/100th of the aliquot (2 ,ul) of the final concentrate (200 ,ul) was used for RT-PCR. The value shown in parentheses are equivalent to the number of actual
viral particles of each virus being amplified.c +, dot plot probed positive with RT-PCR internal probe; -, dot plot probed negative with RT-PCR internal probe.
sensitivity experiment was estimated from the initial PFU. Theactual PFU used in the sensitivity test could be less.The application of triplex RT-PCR on some concentrated
environmental samples showed positive results for the targetviruses. Although amplified background DNA was observed inboth deep-sea water and secondary effluent sewage samples, itwas negative for poliovirus, HAV, or rotavirus after Southernanalysis. This result underscores the importance of additionaltesting by Southern blotting to confirm the correct amplifica-tion products. The amplified rotavirus target fragments showedtwo bands on the Southern analysis but only one band (278 bp)on an ethidium-stained agarose gel. This observation indicatedthat a smaller fragment homologous to the rotavirus internalprobe was produced during the triplex reaction. However, thesmaller fragment, which could have resulted from incompleteprimer extension, did not affect the interpretation of theresults. Because there are many unidentified substancespresent in the environmental samples, these impurities couldinterfere with the RT-PCR during the amplification process.The detection sensitivity of both triplex and monoplex RT-PCR on seeded primary influent was lower than that on seededvirus samples (Table 3, Fig. 3). In a previous report (27), wehave detected enteroviruses and HAV in primary and second-ary effluent samples using monoplex RT-PCR. In addition,there are other reports in the literature that document thedetection of viral RNA in groundwater (1), sewage sludge (13),surface waters (15), and river water (9a) by monoplex RT-PCR. In this current study, the application of a triplex RT-PCRamplification method was clearly an improvement over previ-ously reported monoplex RT-PCR methods, since analysistime was greatly reduced and three types of virus could bedetected with one amplification step.
In conclusion, this study focused primarily on the develop-ment and application of triplex RT-PCR for the detection ofpoliovirus, HAV, and rotavirus from environmental watersamples. The success of this method is an improvement overcurrent monoplex RT-PCR because it provides a more rapidand efficient way to detect these three medically importantviruses. This sensitive technology could help researchers toinvestigate the presence of virus in environmental samples andcould also be of benefit in clinical diagnoses.
ACKNOWLEDGMENTSThis study is based upon work supported by the National Water
Research Institute under awards WQ-91-01 and HR-92-06. Matchingfund supports were received from County Sanitation Districts ofOrange County.We thank Mark Sobsey for providing HAV strain HM175 and Moy
Yahya and Marylin Koval for cultivating poliovirus type 1 and rotavirus
SA-11. We are grateful to G. Fred Bonilla for sample collection andconcentration and to Patrick McNelly for editing the manuscript.
REFERENCES1. Abbaszadegan, M., M. S. Huber, C. P. Gerba, and I. L. Pepper.
1993. Detection of enteroviruses in groundwater with the poly-merase chain reaction. Appl. Environ. Microbiol. 59:1318-1324.
2. American Public Health Association. 1992. Standard methods forthe examination of water and wastewater, 18th ed. AmericanPublic Health Association, Washington, D.C.
3. Atmar, R. L., T. G. Metcalf, F. H. Neill, and M. K. Estes. 1993.Detection of enteric viruses in oysters by using the polymerasechain reaction. Appl. Environ. Microbiol. 59:631-635.
4. Bej, A. K., M. H. Mahbubani, R. Miller, J. L. DiCesare, L. Hal,and R. M. Atlas. 1990. Multiplex PCR amplification and immobi-lized capture probes for detection of bacterial pathogens andindicators in water. Mol. Cell. Probes 4:353-365.
5. Bej, A. K., R. J. Steffan, J. DiCesare, L. Haff, and R. M. Atlas.1990. Detection of coliform bacteria in water by polymerase chainreaction and gene probes. Appl. Environ. Microbiol. 56:307-314.
6. Chapman, N. M., S. Tracy, C. J. Gauntt, and U. Fortmueller. 1990.Molecular detection and identification of enteroviruses usingenzymatic amplification and nucleic acid hybridization. J. Clin.Microbiol. 28:843-850.
7. Davis, L. G., M. D. Dibner, and J. F. Battey. 1986. Basic methodsin molecular biology, p. 147-149. Elsevier Science Publishing Co.,New York.
8. De Leon, R., S. M. Matsui, R. S. Baric, J. E. Herrmann, N. R.Blacklow, H. B. Greenberg, and M. D. Sobsey. 1992. Detection ofNorwalk virus in stool specimens by reverse transcriptase-poly-merase chain reaction and nonradioactive oligoprobes. J. Clin.Microbiol. 30:3151-3157.
9. De Leon, R., C. Shieh, R. S. Baric, and M. D. Sobsey. 1990.Detection of enteroviruses and hepatitis A virus in environmentalsamples by gene probes and polymerase chain reaction, p. 833-853. In Advances in water analysis and treatment. Proceedings ofthe Water Quality Technology Conference, San Diego, Calif.American Water Works Association, Denver.
9a.Divizia, M., G. Morace, R. Gabrieli, G. Pisani, and A. Pana. 1993.Application of the PCR technique to the detection of hepatitis-Avirus in the environment. Water Sci. Technol. 27:223-225.
10. Gama, R. E., P. R. Horsnell, P. J. Hughes, C. North, C. B. Bruce,W. Al-Nakib, and G. Stanway. 1989. Amplification of rhinovirusspecific nucleic acids from clinical samples using the polymerasechain reaction. J. Med. Virol. 28:73-77.
11. Gerba, C. P., and J. B. Rose. 1990. Viruses in source and drinkingwater, p. 380-395. In Drinking water microbiology. Springer-Verlag, New York.
12. Gouvea, V., R. I. Glass, P. Woods, K. Taniguchi, H. F. Clark, B.Forrester, and Z.-Y. Fang. 1990. Polymerase chain reaction am-plification and typing of rotavirus nucleic acid from stool speci-mens. J. Clin. Microbiol. 28:276-282.
13. Graff, J., J. Ticehurst, and B. Flehmig. 1993. Detection of hepatitisA virus in sewage sludge by antigen capture polymerase chain
2406 TSAI ET AL.
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

TRIPLEX RT-PCR ON SEWAGE AND OCEAN WATER 2407
reaction. Appl. Environ. Microbiol. 59:3165-3170.14. Jiang, X., J. Wang, D. Y. Graham, and M. K. Estes. 1992.
Detection of Norwalk virus in stool by polymerase chain reaction.J. Clin. Microbiol. 30:2529-2534.
15. Kopecka, H., S. Dubrou, J. Prevot, J. Marechal, and J. M.L6pez-Pila. 1993. Detection of naturally occurring enteroviruses inwaters by reverse transcription, polymerase chain reaction, andhybridization. Appl. Environ. Microbiol. 59:1213-1219.
16. Morse, D. L., J. J. Guzewich, J. P. Hanrahan, R. Stricof, M.Shayegani, R. Deibel, J. C. Grabau, N. A. Nowak, J. E. Herrmann,G. Cukor, and N. R. Blacklow. 1986. Widespread outbreaks ofclam- and oyster-associated gastroenteritis. Role of Norwalk virus.N. Engi. J. Med. 314:678-681.
17. Muir, P., F. Nicholson, M. Jhetam, S. Neogi, and J. E. Banatvala.1993. Rapid diagnosis of enterovirus infection by magnetic beadextraction and polymerase chain reaction detection of enterovirusRNA in clinical specimens. J. Clin. Microbiol. 31:31-38.
18. Murphy, A. M., G. S. Grohmann, P. J. Christopher, W. A. Lopez,G. R. Davey, and R. H. Millsom. 1979. An Australia-wide outbreakof gastroenteritis from oysters caused by Norwalk virus. Med. J.Aust. 2:329-333.
19. Paul, M. O., and E. A. Erinle. 1982. Influence of humidity onrotavirus prevalence among Nigerian infants and young childrenwith gastroenteritis. J. Clin. Microbiol. 15:212-215.
20. Richards, G. P. 1985. Outbreaks of shellfish-associated entericvirus illness in the United States: requisite for development of viralguidelines. J. Food Prot. 48:815-823.
21. Schwab, K. J., R. De Leon, R. S. Baric, and M. D. Sobsey. 1991.Detection of rotaviruses, enteroviruses and hepatitis A virus byreverse transcriptase-polymerase chain reaction, p. 475-491. InAdvances in water analysis and treatment. Proceedings of theWater Quality Technology Conference, Orlando, Fla. AmericanWater Works Association, Denver.
22. Schwab, K. J., R. De Leon, and M. D. Sobsey. 1993. Development
of PCR methods for enteric virus detection in water. Water Sci.Technol. 27:211-218.
23. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
24. Toyoda, H., M. Kohara, Y. Kataoka, T. Omata, N. Imura, and A.Nomoto. 1984. Complete nucleotide sequences of all three polio-virus serotype genomes. Implication for genetic relationship, genefunction and antigenic determinants. J. Mol. Biol. 174:561-585.
25. Tsai, Y.-L., C. J. Palmer, and L. R. Sangermano. 1993. Detectionof Escherichia coli in sewage and sludge by polymerase chainreaction. Appl. Environ. Microbiol. 59:353-357.
26. Tsai, Y.-L., M. D. Sobsey, L. R. Sangermano, and C. J. Palmer.1993. Simple method of concentrating enteroviruses and hepatitisA virus from sewage and ocean water for rapid detection byreverse transcriptase-polymerase chain reaction. Appl. Environ.Microbiol. 59:3488-3491.
27. Tsai, Y.-L., M. T. Yahya, C. D. McGee, L. R. Sangermano, and C. J.Palmer. 1993. Detection of enteroviruses and hepatitis A virus insewage and ocean water by reverse transcriptase-polymerase chainreaction, p. 115-122. In Proceedings of the Water EnvironmentFederation 66th Annual Conference & Exposition, vol. 7, Anaheim,Calif. Water Environment Federation, Alexandria, Va.
28. Wanke, C. A., and R. L. Guerrant. 1987. Viral hepatitis andgastroenteritis transmitted by shellfish and water. Infect. Dis. Clin.N. Am. 1:649-664.
29. Way, J. S., K. L. Josephson, S. D. Pillai, M. Abbaszadegan, C. P.Gerba, and I. L. Pepper. 1993. Specific detection of Salmonellaspp. by multiplex polymerase chain reaction. Appl. Environ.Microbiol. 59:1473-1479.
30. Wilde, J., R. Van, L Pickering, J. Eiden, and R. Yolken. 1992.Detection of rotaviruses in the day care environment by reversetranscriptase polymerase chain reaction. J. Infect. Dis. 166:507-511.
VOL. 60, 1994
on Septem
ber 30, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from