demonstration of constant upregulation of the telomerase rna component in human gastric carcinomas...
TRANSCRIPT

, . 185: 139–144 (1998)
DEMONSTRATION OF CONSTANT UPREGULATIONOF THE TELOMERASE RNA COMPONENT IN HUMAN
GASTRIC CARCINOMAS USING IN SITUHYBRIDIZATION
, *,
Institute for Pathology, Universitätsklinikum Benjamin Franklin, Free University Berlin, 12200 Berlin, Germany
SUMMARY
Upregulation of the ribonucleoprotein telomerase seems to be a prerequisite for immortality, a feature of malignant cells. Using apolymerase chain reaction (PCR)-based assay, it is possible to demonstrate telomerase activity (TA) in specimens of most humanmalignancies, whereas it is absent from most normal tissues. It remains unclear, however, why between 5 and 50 per cent of variousmalignant tumour samples give negative results when TA is measured by the telomeric repeat amplification protocol (TRAP). Theexpectation that reverse transcription (RT)-PCR for detection of the telomerase RNA component (hTR) would be able to complementor to replace the TRAP assay failed, since malignant as well as non-malignant tissue samples gave positive results in most instances. Inthe present study, in situ hybridization (ISH) was developed to demonstrate the RNA component of human telomerase at the single celllevel. With this method, 13 specimens of fresh frozen gastric carcinoma and four of normal, dysplastic, or inflamed gastric mucosa wereinvestigated and the results were compared with those obtained by RT-PCR and the TRAP assay. In addition, ISH was performed onformalin-fixed sections of the same cases. The TRAP assay revealed positive results in 8 out of 13 gastric carcinomas and was negativein all non-malignant tissues. RT-PCR led to amplification of the telomerase RNA component in all specimens tested, irrespective of thepresence or absence of malignant cells. By ISH, all gastric carcinomas showed strong telomerase RNA component-specific signals overmalignant cells, whereas only a few grains were detectable over some types of normal somatic cells, including activated lymphocytes. Inconclusion, high expression of the telomerase RNA component was restricted to the malignant cells of all the gastric carcinomasinvestigated, as shown by ISH. This indicates that the absence of TA in a proportion of carcinomas is due to methodological problemsof the TRAP assay and is not caused by biological factors. The detection of high levels of the telomerase RNA component by ISH isthus a useful technique for demonstrating malignant cells in frozen and formalin-fixed pathological specimens. ? 1998 John Wiley &Sons, Ltd.
J. Pathol. 185: 139–144, 1998.
KEY WORDS—telomerase; telomerase RNA component; polymerase chain reaction; TRAP assay; in situ hybridisation
*Correspondence to: Michael Hummel, PhD, Institute of Pathol-ogy, Klinikum Benjamin Franklin, Free University of Berlin, 12200Berlin, Germany. E-mail: [email protected]
Contract grant sponsor: Deutsche Forschungsgemeinschaft (DFG);Contract grant number: Hu 557/2-1.
INTRODUCTION
The ends of eukaryotic chromosomes are designatedas telomeres and in humans they consist of up tothousands of repetitions of the DNA sequence of5*-(TTAGGG)n-3*.1 Telomeres are essential for prevent-ing degradation and recombination of the chromo-somes. Due to the end replication problem, DNApolymerase cannot completely replicate the ends ofchromosomes.2,3 By each division, cells lose telomeresequences until the telomeres become unstable, leadingto cell senescence. It has been proposed that cells counttheir divisions through telomere shortening, a phenom-enon called the mitotic clock.4–6 This internal clock canbe stopped by de novo synthesis of telomeric repeats bythe ribonucleoprotein telomerase.7,8
Recently, a PCR-based method has been reported todetermine telomerase activity (TA) in cell supernatants
CCC 0123–3417/98/060212–07 $17.50? 1998 John Wiley & Sons, Ltd.
(TRAP assay).9 With this approach, TA can be shown inmany human malignancies, but not in most normaltissues. This led to the concept that TA might serveas a marker to discriminate between malignant andnon-malignant human cell proliferation. However, asignificant proportion of human malignancies failed todemonstrate TA, casting doubt on the diagnostic valueof the TRAP assay.9–20 Alternative approaches aretherefore required to determine whether this is due totechnical problems or biological factors.
Since the sequence of the telomerase cDNA is notyet available, RT-PCR for the detection of thetelomerase RNA component (hTR) is employed todemonstrate indirectly the presence of telomerase. Sur-prisingly, the RNA component can be detected21,22 inmalignant as well as in non-malignant samples. It wastherefore concluded that the activity of telomerase isnot regulated through this RNA component.23,24 Totest this hypothesis and to demonstrate the cellulardistribution of the telomerase RNA component, weestablished an in situ hybridization and applied thistechnique to frozen as well as to formalin-fixed tissuesections.

140 B. HEINE ET AL.
MATERIALS AND METHODS
Cell lines and tissue samples
Cell lines (K562, Hela) were cultured in RPMImedium supplemented with 10 per cent fetal calf serum(FCS). Cell suspensions were centrifuged at 10 000 g for10 min and washed twice with phosphate-buffered saline(PBS). Tissue samples were obtained by gastrectomy fortherapeutic reasons. The tumours were classified accord-ing to Laurén.25 To investigate tissue samples for thepresence of TA, 15 ìm thick tissue sections were pro-duced from frozen tissue blocks obtained in the courseof the routine diagnostic procedure. The tissues werecontrolled for the presence of neoplastic cells by histo-logical examination of sections directly above andbeneath the extracted portion.
TRAP assay
Telomerase activity was determined as previouslydescribed, with minor modifications.9 Briefly, aliquots of6 ìg, 0·6 ìg, and 0·06 ìg of total protein from eachsample were incubated. Two TRAP assays were per-formed from each sample. The supernatants of TRAP-negative samples were mixed with TRAP-positive cell
? 1998 John Wiley & Sons, Ltd.
line supernatants (5:1; 1:5) in order to detect telomeraseinhibitors.
Analysis and quantification of the TRAP assay
TRAP-positive samples were semi-quantified employ-ing the reliable peak areas between 37 and 150 bp of theTRAP amplification products. The sum of the corre-sponding peak areas of a positive control (K562; 6 ìg)obtained from the same GeneScan run were set to 100per cent and the peaks of the negative control (lysisbuffer) were set to 0 per cent. The relative TA (rTA) ofa given sample X was calculated using the followingformula:
rTAX=((X1"nc)/(pc"nc)+(X2"nc)/(pc"nc))/2#100%
where X1 and X2 are the peak areas of sample Xobtained from two different TRAP assays; pc is the peakarea, positive control; and nc is the peak area, negativecontrol.
Fig. 1—Non-radioactive TRAP assay for the detection of telomerase activity. Fluorescence-labelled amplificates were separated on a DNAsequencing gel and analysed by the GeneScan software. (A) Representative TRAP-positive sample (Hela cells; 0·6 ìg protein). The appearanceof TRAP-positive carcinomas is identical to that seen with cell line samples. Note the 6 bp distance between each peak, representing the repetitivenature of the telomeric repeats. (B) TRAP-negative adenocarcinoma (case 13; 6 ìg protein). (C) Mixture of the supernatants from Hela cells(1 ìg protein) and case 13 (5 ìg protein). Note the disappearance of the specific signals. (D) Negative control (PBS)
RT-PCR for the telomerase RNA component (hTR)
RT-PCR for the detection of the telomerase RNA(hTR) component was performed using a GeneAmp?
, . 185: 139–144 (1998)

141CONSTANT UPREGULATION OF THE TELOMERASE RNA COMPONENT IN HUMAN GASTRIC CARCINOMAS
RNA-PCR kit (Perkin-Elmer/Applied Biosystems,Weiterstadt, Germany). Random hexamer primedreverse transcription was performed with 2 ìl of lysateprepared for the TRAP assay in a 20 ìl reaction volumeaccording to the manufacturer’s recommendation. PCRfor amplification of hTR was carried out in 100 ìlemploying the following primers (0·15 ì each): HTR-UP: 5*-ACCCTAACTGAGAAGGGCGT-3*; HTR-LOW: 5*-GCCAGCAGCTGACATTTTTT-3*. Fortycycles (96)C for 15 s, 55)C for 30 s, and 72)C for 30 s)were used, employing 2 m MgCl2. The sequence of theresulting 142 bp amplificate was determined byautomated DNA sequencing and was confirmed tocomprise the expected proportion of the telomeraseRNA component.
In situ hybridization (ISH)
The probe for detection of the telomerase RNAcomponent (hTR) by ISH was generated by RT-PCRemploying primers deduced from published sequences:TEL-UP: 5*-GGTGGCCATTTTTTGTCTAAC-3*;TEL-LOW: 5*-TGCATGTGTGAGCCGAGT-3*. TotalRNA (1 ìg) from PHA-activated peripheral blood lym-phocytes was used for reverse transcription and ampli-fication as described above. The resulting 417 bpamplificate was subcloned in pAMP1 (GibcoBRL,Gaithersburg, U.S.A.) and sequenced (373A, Perkin-Elmer/Applied Biosystems, Weiterstadt, Germany).Hybridization was carried out with 35S-labelled run-offtranscripts as described elsewhere.26 In brief, dewaxedand rehydrated paraffin sections were exposed to 0·2
? 1998 John Wiley & Sons, Ltd.
HCl and 0·125 mg/ml pronase (Boehringer-Mannheim,Germany), followed by acetylation with 0·1 trieth-anolamine, pH 8·0/0·25 per cent (v/v) acetic anhydrideand dehydration through graded ethanol. Slideswere hybridized to 2–4#105 cpm of labelled probesovernight at 50)C and exposed for 8–12 weeks.
RESULTS
Frozen tissue from 13 gastric carcinomas and fromone dysplastic, one inflamed, and two normal samples ofgastric mucosa was investigated using the TRAP assayto determine telomerase activity, as well as RT-PCR andISH to demonstrate the telomerase RNA component. Inaddition, we performed ISH on paraffin sections of thesame cases.
Table I—Detection of telomerase activity and the telomerase RNA component in malignant and non-malignant gastric tissuesamples
CaseNo. Diagnosis Typing Staging Grading
Age(years) Sex rTa
Telomerase RNAcomponent (hTR)
RT-PCR ISH
1 Adenocarcinoma Intestinal type pT2pN0 GII 73 M 85% Pos. Pos.*2 Adenocarcinoma Intermediate type pT3pN2 GIII 58 F 50% Pos. Pos.*3 Adenocarcinoma Poorly differentiated pT2pN1 GIII 77 M 45% Pos. Pos.*4 Adenocarcinoma Poorly differentiated pT3pN2 GIII 79 F 40% Pos. Pos.*5 Adenocarcinoma Diffuse type pT2pN1 GIII 66 M 15% Pos. Pos.*6 Adenocarcinoma Intestinal type pT1pN1 GII 56 M 15% Pos. Pos.*7 Adenocarcinoma Poorly differentiated pT3pN1 GIII 58 M 10% Pos. Pos.*8 Adenocarcinoma Intestinal type pT2pN0 GII 42 M 5% Pos. Pos.*9 Adenocarcinoma Diffuse type pT3pN0 GIII 54 F Neg. Pos. Pos.*
10 Adenocarcinoma Intestinal type pT3pN1 GIII 50 M Neg. Pos. Pos.*11 Adenocarcinoma Intestinal type pT4pN1 GIII 82 F Neg. Pos. Pos.*12 Adenocarcinoma Intestinal type pT1pN1 GII 77 F Neg. Pos. Pos.*13 Adenocarcinoma Diffuse type pT2pN1 GIII 72 F Inhib. Pos. Pos.*14 Dysplastic mucosa — High grade 81 F Neg. Pos. —†15 Gastritis Chronic active, II — — 50 M Neg. Pos. —‡16 Normal mucosa — — 85 F Neg. Pos. —‡17 Normal mucosa — — 51 M Neg. Pos. —‡
*Strong expression in neoplastic cells.†Weak to moderate accumulation of grains over dysplastic cells.‡Weak accumulation of grains over normal mucosa cells and activated lymphocytes.rTA=relative telomerase activity deduced from the TRAP assay; inhib.=telomerase inhibition; RT-PCR=reverse transcription polymerase chain
reaction; ISH=in situ hybridization.
Telomerase activity (TA)
The TRAP assay revealed positive results in 8 out of13 cases (62 per cent) of gastric carcinoma. One of thenegative samples displayed no TA due to inhibitoryeffects (Figs 1B and 1C). There was no significantcorrelation between the different tumour subtypes, thegrading, the staging, and the age or sex of the patients.Non-malignant samples were negative in all instances(n=4), including one high-grade dysplasia of the mucosa(Table I). Telomerase-positive samples showed thecharacteristic 6 bp ladder of the TRAP amplificates(Fig. 1A), whereas this ladder was absent in RNasepretreated samples and in the negative control (Fig. 1D).
, . 185: 139–144 (1998)
142 B. HEINE ET AL.
RT-PCR for the telomerase RNA component (hTR)
Using RT-PCR for amplification of the RNA compo-nent of telomerase, all specimens, both malignantand non-malignant, gave positive results (Table I).Moreover, the intensity of the amplificates showedno convincing differences between malignant andnon-malignant samples. Pretreatment of the sampleswith RNase led to the disappearance of the PCRproducts, indicating the specificity of this approach.
In situ hybridization for the telomerase RNA component(hTR)
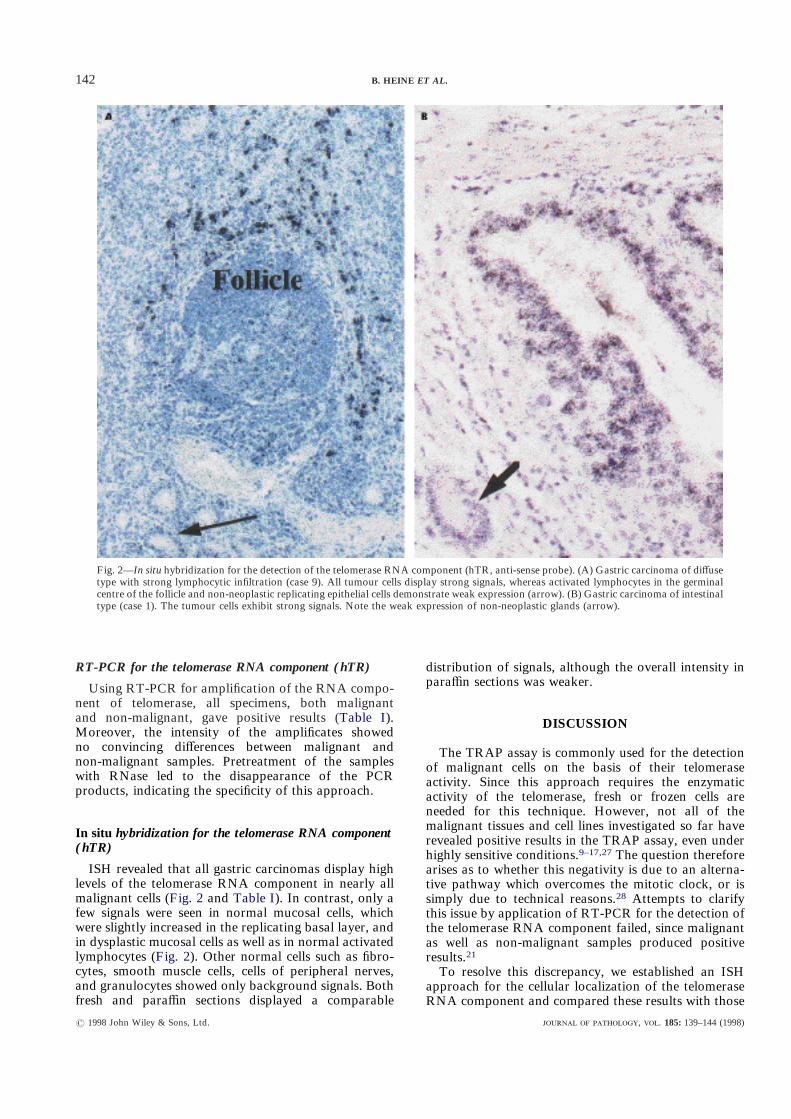
ISH revealed that all gastric carcinomas display highlevels of the telomerase RNA component in nearly allmalignant cells (Fig. 2 and Table I). In contrast, only afew signals were seen in normal mucosal cells, whichwere slightly increased in the replicating basal layer, andin dysplastic mucosal cells as well as in normal activatedlymphocytes (Fig. 2). Other normal cells such as fibro-cytes, smooth muscle cells, cells of peripheral nerves,and granulocytes showed only background signals. Bothfresh and paraffin sections displayed a comparable
? 1998 John Wiley & Sons, Ltd.
distribution of signals, although the overall intensity inparaffin sections was weaker.
Fig. 2—In situ hybridization for the detection of the telomerase RNA component (hTR, anti-sense probe). (A) Gastric carcinoma of diffusetype with strong lymphocytic infiltration (case 9). All tumour cells display strong signals, whereas activated lymphocytes in the germinalcentre of the follicle and non-neoplastic replicating epithelial cells demonstrate weak expression (arrow). (B) Gastric carcinoma of intestinaltype (case 1). The tumour cells exhibit strong signals. Note the weak expression of non-neoplastic glands (arrow).
DISCUSSION
The TRAP assay is commonly used for the detectionof malignant cells on the basis of their telomeraseactivity. Since this approach requires the enzymaticactivity of the telomerase, fresh or frozen cells areneeded for this technique. However, not all of themalignant tissues and cell lines investigated so far haverevealed positive results in the TRAP assay, even underhighly sensitive conditions.9–17,27 The question thereforearises as to whether this negativity is due to an alterna-tive pathway which overcomes the mitotic clock, or issimply due to technical reasons.28 Attempts to clarifythis issue by application of RT-PCR for the detection ofthe telomerase RNA component failed, since malignantas well as non-malignant samples produced positiveresults.21
To resolve this discrepancy, we established an ISHapproach for the cellular localization of the telomeraseRNA component and compared these results with those
, . 185: 139–144 (1998)

143CONSTANT UPREGULATION OF THE TELOMERASE RNA COMPONENT IN HUMAN GASTRIC CARCINOMAS
obtained by the TRAP assay and RT-PCR. Our ISHresults clearly demonstrate that the telomerase RNAcomponent is expressed in high amounts only in themalignant cells of all 13 gastric carcinomas investigated.In contrast, the TRAP assay produced positive results inonly 8 out of 13 cases. This relatively low TRAPefficiency is most likely due to the fact that the tissuesamples were not collected under conditions optimizedfor the detection of telomerase activity, but wereobtained in the course of the routine diagnostic pro-cedure. The positivity of the RT-PCR in all malignant aswell as non-malignant tissue samples confirms thelimited value of this approach.24 As shown by our ISH,this positivity of non-malignant samples in the RT-PCRis most likely due to low rates of expression of thetelomerase RNA component detectable in some non-malignant cell types, such as cells of the replicating basallayer of the gastric mucosa and activated lymphocytes.
Our results clearly show that the telomerase RNAcomponent is not ubiquitously expressed in all cells atthe same level, as suggested by the results of our ownand other RT-PCR results.21,23,24 Instead, only a pro-portion of lymphocytes, most likely activated, as well asproliferating basal layer cells, displayed weak expressionof hTR. This points to their potential to overcome themitotic clock. However, since their proliferation rate islow, it is tempting to speculate that the TA escapeddetection by the TRAP due to the small amount oftelomerase present in these cells. The negative outcomeof the TRAP assay in a significant number of malignan-cies in this and other studies might therefore representtechnical artefacts due to degradation, inhibition, orother unknown reasons. An alternative telomerase-independent mechanism for the blockage of the mitoticclock, as postulated for TRAP-negative cases,28 isunlikely. Our results are in line with very recent datafrom Yashima et al.,29 who reported telomerase positiv-ity in nearly all malignancies of the lung after applica-tion of the TRAP assay and hTR ISH. Theirobservation of focally increased expression of the telom-erase RNA component in carcinomas in situ as anindicator of early invasive tumour growth requires,however, further investigation.
In conclusion, malignant cells harbour elevated levelsof the telomerase RNA component in all gastric carci-nomas investigated. In contrast, only a few types ofnon-malignant cells display expression of hTR, at lowrates. Whereas the detection of telomerase activity isrestricted to fresh or frozen materials, ISH for thecellular demonstration of the telomerase RNA compo-nent is applicable on frozen as well as on formol-fixedtissue sections. This ISH approach might therefore serveas a more reliable tool than the TRAP assay andRT-PCR to determine the presence of malignant cells,even in routinely processed pathological specimens.
NOTES ADDED IN PROOF
During the review process the cDNA of one of thetelomerase subunits was published (Meyerson M. et al.,Cell, 1997 Aug 22; 90(4): 785–795). Preliminary exper-
? 1998 John Wiley & Sons, Ltd.
imental data (Nakayama J. et al., Nature Genetics, 1998Jan; 18(1): 65–68) indicate that this part of the telomer-ase is more strictly regulated than its RNA component.
ACKNOWLEDGEMENTS
We are grateful to Elisabeth Oker for her excellenttechnical assistance and Janet Yates for editorial helpwith the preparation of the manuscript. This work wassupported by the Deutsche Forschungsgemeinschaft(DFG; Hu 557/2-1).
REFERENCES1. Moyzis RK, Buckingham JM, Cram IS, et al. A highly conserved repetitive
DNA sequence, (TTAGGG)n, present at the telomeres of human chromo-somes. Proc Natl Acad Sci USA 1988; 85: 6622–6626.
2. Watson JD. Origin of concatemeric T7 DNA. Nature New Biol 1972; 239:197–201.
3. Olovnikov A. A theory of marginotomy. The incomplete copying oftemplate margin in enzymic synthesis of polynucleotides and biologicalsignificance of the phenomenon. Theor Biol 1973; 41: 181–190.
4. Harley CB, Futcher AB, Greider CW. Telomeres shorten during aging ofhuman fibroblasts. Nature 1990; 345: 458–460.
5. Lindsey J, McGill NI, Lindsey LA, Green DK, Kooke HJ. In vivo loss oftelomeric repeats with age in humans. Mut Res 1991; 256: 45–48.
6. Allsopp RC, Vaziri H, Patterson C, et al. Telomere length predictsreplicative capacity of human fibroblasts. Proc Natl Acad Sci USA 1992; 89:10114–10118.
7. Greider CW, Blackburn EH. Identification of a specific telomere terminaltransferase activity in tetrahymena extracts. Cell 1985; 43: 405–413.
8. Morin G. The human telomere terminal transferase enzyme is a ribonucleo-protein that synthesizes TTAGGG repeats. Cell 1989; 59: 521–529.
9. Kim NW, Piatyszek MA, Prowse KR, et al. Specific association of humantelomerase activity with immortal cells and cancer. Science 1994; 266:2011–2015.
10. Hiyama E, Yokoyama T, Tatsumoto N, et al. Telomerase activity in gastriccancer. Cancer Res 1995; 55: 3258–3262.
11. Hiyama K, Hiyama E, Ishioka S, et al. Telomerase activity in small-cell andnon-small-cell lung cancers.. J Natl Cancer Inst 1995; 87: 895–902.
12. Counter C, Hirte H, Bacchetti S, Harley C. Telomerase activity in humanovarian carcinoma. Proc Natl Acad Sci USA 1994; 91: 2900–2904.
13. Chadeneau C, Hay K, Hirte H, Gallinger S, Bacchetti S. Telomerase activityassociated with acquisition of malignancy in human colorectal cancer.Cancer Res 1995; 55: 2533–2536.
14. Tahara H, Nakanishi T, Kitamoto M, et al. Telomerase activity in humanliver tissues: comparison between chronic liver disease and hepatocellularcarcinomas. Cancer Res 1995; 55: 2734–2736.
15. Nilsson P, Mehle C, Remes K, Roos G. Telomerase activity in vivo inhuman malignant hematopoietic cells. Oncogene 1994; 9: 3043–3048.
16. Hiyama E, Hiyama K, Yokohama T, Matsuura Y, Piatyszek M, Shay JW.Correlating telomerase activity levels with human neuroblastoma outcomes.Nature Med 1995; 1: 249–255.
17. Brousset P, Al Saati T, Chaouche N, Schlaifer D, Chittal S, Delsol G.Telomerase activity in reactive and neoplastic lymphoid tissues: infrequentdetection of activity in Hodgkin’s disease. Blood 1997; 89: 26–31.
18. Mehle C, Piatyszek M, Ljungsberg B, Shay J, Roos G. Telomerase activityin human renal cell carcinoma.. Oncogene 1996; 13: 161–166.
19. Nouso K, Urabe Y, Higashi T, et al. Telomerase as a tool for the differentialdiagnosis of human hepatocellular carcinoma. Cancer 1996; 78: 232–236.
20. Counter CM, Gupta J, Harley CB, Leber B, Bacchetti S. Telomeraseactivity in normal leukocytes and in hematologic malignancies. Blood 1995;85: 2315–2320.
21. Feng J, Funk W, Wang S, et al. The RNA component of human telomerase.Science 1995; 269: 1236–1241.
22. Broccoli D, Godley L, Donehower L, Varmus H, de Lange T. Telomeraseactivation in mouse mammary tumors: lack of detectable telomere shorten-ing and evidence for regulation of telomerase RNA with cell proliferation.Mol Cell Biol 1996; 16: 3765–3772.
23. Blasco M, Rizen M, Greider C, Hanahan D. Differential regulation oftelomerase activity and telomerase RNA during multi-stage tumorigenesis.Nature Genet 1996; 12: 200–204.
24. Avilion A, Piatyszek M, Gupta J, Shay J, Bacchetti S, Greider C. Humantelomerase RNA and telomerase activity in immortal cell lines and tumortissues. Cancer Res 1996; 56: 645–650.
25. Laurén P. The two histological main types of gastric carcinoma: diffuse andso-called intestinal type. Acta Pathol Microbiol Immunol Scand 1965; 64 (A):31–49.
, . 185: 139–144 (1998)

144 B. HEINE ET AL.
26. Foss HD, Herbst H, Oelmann E, et al. Lymphotoxin, tumour necrosisfactor and interleukin-6 gene transcripts are present in Hodgkin andReed–Sternberg cells of most Hodgkin’s disease cases. Br J Haematol 1993;84: 627–635.
27. Heine B, Hummel M, Müller M, Heicappell R, Miller K, Stein H.Non-radioactive measurement of telomerase activity in human bladdercancer, bladder washings, and in urine. J Pathol 1998; 184: 71–76.
? 1998 John Wiley & Sons, Ltd.
28. Bryan T, Englezou A, Gupta J, Bacchetti S, Reddel R. Telomeraseelongation in immortal human cells without detectable telomerase activity.EMBO J 1995; 14: 4240–4248.
29. Yashima K, Litzky L, Kaiser L, et al. Telomerase expression in respiratoryepithelium during multistage pathogenesis of lung carcinomas. Cancer Res1997; 57: 2373–2377.
, . 185: 139–144 (1998)