demonstration of a hormonal inhibitor of proximal tubular
TRANSCRIPT

Demonstration of a Hormonal Inhibitor of
Proximal Tubular Reabsorption during Expansion of
Extracellular Volume with Isotonic Saline
FLOYD C. RECTOR, JR., MANUEL MARTINEZ-MALDONADO, NEIL A. KURTZMAN,JACK C. SELLMAN, FRED OEirlfm, and DONALD W. SELDINwith the technical assistance of ALLEN C. NUNN
From the Department of Internal Medicine, The University of Texas SouthwesternMedical School, Dallas, Texas and Veterans Administration Hospital,Dallas, Texas 75235
A B S T R A C T Evidence for the elaboration of ahormonal inhibitor of renal tubular reabsorptionin response to expansion of extracellular fluid vol-ume was obtained by examining the effects ofplasma from rats and dogs undergoing saline diu-resis on the rate of proximal tubular reabsorptionmeasured both directly by micropuncture tech-niques and indirectly by clearance techniques.
Intravenous infusion of plasma from saline-loaded rats and dogs, but not plasma from con-trol animals, inhibited the intrinsic reabsorptivecapacity of the proximal tubule (as estimated fromthe shrinking-drop technique) by 35%, and re-duced fractional reabsorption (as estimated fromthe tubular fluid-to-plasma ratio) by 20%o. In ad-dition the natriuretic plasma increased urine flow,solute-free water clearance, and potassium excre-tion in rats with hereditary diabetes insipidus, in-dicating an increase in the delivery of filtrate outof the proximal tubule to the more distal dilutingsegments of the nephron.The hormonal inhibition of proximal tubular
reabsorption had an extremely rapid onset of action(within seconds after instillation into the tubularlumen) and a short duration of action (less than
Dr. Kurtzman is a Special Fellow, Office of the Sur-geon General, U. S. Army.Address requests for reprints to Dr. Floyd C. Rector,
Jr., Department of Internal Medicine, University of TexasSouthwestern Medical School, Dallas, Tex. 75235.Received for publication 11 September 1967 and in re-
vised form 27 November 1967
30 min after cessation of an intravenous infusion).Inhibitory activity was lost from natriuretic plasmaupon dialysis and could be recovered in the dialy-sate. Dialysates of natriuretic plasma, when in-jected directly into the tubular lumen, also in-hibited proximal reabsorption, indicating an actionon the luminal side of the cell.
INTRODUCTION
It is now abundantly clear that the augmentedexcretion of sodium which results from expansionof extracellular fluid (ECF) volume with isotonicsaline is mediated, neither by an increase in glo-merular filtration rate (GFR) nor by suppressionof aldosterone secretion, but rather by some yetunidentified mechanism which inhibits the tubularreabsorption of sodium (1-4). The fact that sa-line infusion during water diuresis increases bothsolute-free water clearance (CH2O) and sodium ex-cretion suggests that a major site of inhibition isthe proximal tubule (3), an inference amply con-firmed by micropuncture studies (5-8).The mechanism whereby expansion of ECF
volume reduces proximal tubular reabsorption hasnot yet been identified. Several investigators (1,9-12) have suggested that the inhibiting influencemay be mediated by the elaboration of a natriu-retic hormone, whereas others (13-16) have in-voked alterations in various physical factors, suchas colloid osmotic pressure, blood pressure, renal
The Journal of Clinical Investigation Volume 47 1968 761

vascular resistance, and distribution of renal bloodflow.
Cross-circulation studies, designed to demon-strate the presence of a natriuretic hormone, haveyielded equivocal results (1, 4, 11, 12). The cross-circulation technique, however, is not ideallysuited to examine for the presence of a natriuretichormone. Micropuncture studies have shown thatsaline infusions suppress reabsorption primarily inthe proximal tubule (5-8), and that much of thefiltrate delivered out of this segment is reabsorbeddistally, with only a portion of the increment ap-pearing in the final urine. Under conditions ofcross-circulation, the small increase in sodium ex-cretion commonly observed might not accuratelyreflect the true magnitude of proximal tubularinhibition. On the other hand, such small increasesin sodium excretion that do occur may well bethe consequence, not of the cross-circulation of ahumoral inhibitor, but rather of hemodynamicchanges associated with increased renal blood flowand compositional changes associated with dilu-tion of the blood, both of which are known to re-sult from the cross-circulation procedure and arethemselves capable of inducing natriuresis (15, 17,18).The present experiments were designed to ob-
viate these difficulties by testing for the presenceof a humoral inhibitor in plasma from animalsundergoing saline diuresis (natriuretic plasma),not only by examining its effect on urinary so-dium excretion, but rather by directly assessing itscapacity to inhibit proximal tubular reabsorption.It was shown that an infusion of natriuretic plasmainhibited proximal tubular reabsorption, as judgedby three different techniques: prolongation of therate of reabsorption of a shrinking drop of salineduring stopped flow; depression of the inulin tubu-lar fluid to plasma ratio (TF/P) during freeflow; and increased urinary excretion of free wa-ter and potassium in rats with hereditary diabetesinsipidus. On the basis of these results the conclu-sion was drawn that expansion of extracellularvolume causes the release of a humoral agentwhich inhibits sodium reabsorption in the proximalportions of the nephron.
METHODSThe effect of plasma from antidiuretic animals and fromanimals undergoing saline diuresis (natriuretic plasma)on proximal tubular reabsorption was assayed in three
groups of rats. Groups I and II consisted of Sprague-Dawley rats, whereas group III contained Brattlebororats with hereditary diabetes insipidus.1 Rats in groupsI and II were anesthetized with sodium pentobaribitaland prepared for micropuncture as previously described(19). In order to measure GFR by a constant infusiontechnique the assay rats in groups I and II were sub-jected to right unilateral nephrectomy 1-3 wk before theacute experiment. In groups I and III GFR was mea-sured with radioactive inulin as previously described (19).In group III GFR was calculated on the basis of boththe inulin infusion and the urinary excretion, with goodagreement between the two methods. In group II inulinwas given at a rate of 2 mg/min to maintain a plasmaconcentration of approximately 100 mg/100 ml. To re-place surgical loss of ECF volume occurring during thesurgical procedure each rat was given 1 ml of bicarbonate-saline solution (25 mm NaHCOs + 110 mm NaCl). Thisamount of saline replacement is less than previously used(19) in order to avoid any possibility of overexpandingthe assay animal and thus obscuring possible effects ofnatriuretic plasma on proximal reabsorption. Transittime was measured with lissamine green as previouslydescribed (19).Group I. In the first group of 18 rats the intrinsic re-
absorptive capacity of the proximal tubule was measuredwith the shrinking-drop technique of Gertz (20, 21).Five rats were infused with plasma from rats, while theremaining 13 received plasma from dogs. Four of the ratswere given 5 mg of deoxycorticosterone acetate (DOCA)in oil intramuscularly 4 hr before the experiment.
In each assay rat 6-10 measurements were made dur-ing each of three periods (control, experimental, and re-covery; see below for experimental protocol).Group II. In the second group of 12 rats fractional
reabsorption in the proximal tubule was calculated fromthe ratio of inulin concentration in tubular fluid andplasma [(TF/P)In] at the end of the proximal convolu-tion. The end of the proximal convolution was identifiedby the passage of lissamine green (7, 22). In each ratfour samples of tubular fluid were collected in each ofthree periods (control, experimental, recovery).Group III. In order to estimate the effect of natriuretic
plasma on the delivery of filtrate out of the proximal tu-bule, experiments were carried out in six rats with heredi-tary diabetes insipidus (Brattleboro strain). In the ab-sence of antidiuretic hormone (ADH) urine flow can beused as a rough approximation of the amount of fluiddelivered out of the proximal portions of the nephron tothe distal diluting segments. Urine volume (V), osmolarclearance (Co.m), solute-free water clearance (Cimo), andsodium and potassium excretion were measured duringcontrol, experimental, and recovery periods. Four to five10-min urine collections from an indwelling bladdercatheter were obtained in each period; arterial blood wasobtained at the midpoint of each urine collection.Experimental protocol. The same experimental proto-
col was used in all three groups of assay rats. The ex-periment was divided into three periods: control, experi-
1 Courtesy of Dr. Heinz Valtin.
762 Rector, Martinez-Maldonado, Kurtzman, SeUman, Oerther, and Seldin

mental, and recovery. Each period consisted of an initial30 min for equilibration and 30-45 min for shrinking-dropmeasurements or collection of tubular fluid and urinesamples. Plasma was infused intravenously through theentire experiment at a rate of 0.02 ml/min. (Preliminarystudies demonstrated that this amount of plasma by it-self would not change proximal reabsorption.) Duringthe first or control period plasma from control animalswas infused. During the second or experimental periodthe control plasma was discontinued and natriureticplasma was infused. During the third or recovery periodnatriuretic plasma was discontinued and the originalcontrol plsma reinfused. In an occasional rat the sequencewas varied.
In two experiments 5 ml of natriuretic plasma was di-alyzed through Visking cellophane membrane against 50ml of isotonic bicarbonate-saline solution for 18-24 hr at4VC. The dialyzed plasma was then infused into group Iassay rats according to the standard experimental pro-tocol. In an additional experiment, 5 ml of natriureticplasma was dialyzed against an equal volume of distilledwater for 18-24 hr at 40C and the dialysate then infusedinto a group I rat at 0.04 ml/min during the experimentalperiod.
In five other experiments 3 ml of antidiuretic plasmaand 3 ml of natriuretic plasma were each dialyzed against3 ml of isotonic bicarbonate-saline solution for 18-24 hrat 4VC. The plasma was discarded and the dialysate wasthen injected into the lumen of a proximal tubule and itsreabsorptive rate measured with the shrinking-drop tech-nique. In these experiments measurements were first madewith isotonic bicarbonate-saline solution, then with dialy-sate of control plasma, next with dialysate of natriureticplasma, and finally again with dialysate of control plasma.
Collection of plasma. For each experiment hepa-rized arterial blood from three to four control rats waspooled. In another group of three to four rats, salinediuresis was induced by infusing bicarbonate-saline solu-tion intravenously at 0.4 ml/min. When urinary sodiumexcretion rose to approximately 10%o of the filtered so-dium, the rats were bled directly from the aorta. Theblood was centrifuged and the plasma stored at 4VC untilthe time of the experiment. Each batch of plasma couldbe utilized for two or three experiments. There was nodifference in the results if a fraction of pooled plasmaor plasma from individual rats was infused into the re-
cipient rats.In experiments utilizing dog plasma both control and
natriuretic plasmas were obtained from the same donoranimal. In the first three experiments (Table II, experi-ments 6, 7, 9), both control and natriuretic plasma wereobtained from the femoral artery. In subsequent experi-ments all samples were obtained through a catheter im-planted into a jugular vein. Heparinized saline filledthe entire length of the catheter until the blood sampleswere obtained. After collection of control plasma, bi-carbonate-saline solution was infused through a leg veinat 15 ml/min. When sodium excretion increased to ap-proximately 10-15% of the filtered sodium, blood wasobtained for the experimental period.
Analytic methods. The concentration of inulin in tu-bular fluid was measured by the method of Vurek andPegram (23). Inulin in plasma filtrate and urine wasmeasured by the anthrone method (24). The concentra-tion of inulin-14C was measured by adding samples ofinfusion solution, plasma, urine, and tubular fluids to 10ml of Bray's solution and counting in a Packard Tri-Carb liquid scintillation counter (Packard InstrumentsCo., Inc., Downers Grove, Ill.). The details of this tech-nique have already been described (19). Sodium and po-tassium in plasma and urine were measured by flamephotometry with lithium as an internal standard. Urineand plasma osmolalities were measured by the tech-nique of Ramsay and Brown (25).
Calculations. Fractional reabsorption was calculatedfrom the (TF/P) In ratio with the expression,
fractional reabsorption = [1 - (P/TF)ln]. (1)
The intrinsic reabsorptive capacity, K (expressed as mil-liliter reabsorbed per second per milliliter of tubular vol-ume), was either measured directly with the split-droptechnique and calculated from the expression (20, 21),
0.693K= (2)
where tj is the time required to reabsorb 50% of the in-jected column of saline from the tubular lumen, or esti-mated indirectly from the (TF/P) I, and transit time(T) by the expression (7, 19, 22),
log ~~~~~~~~(3)( )In 2.3- 3
The ratio of tubular volume (rrTd) to GFR/per nephron(Vo) was estimated from the expression (7),
fractional reabsorption = K-(4
where r is the tubular radius and d is the tubular length.
RESULTS
Effects on intrinsic reabsorptive capacity (groupI). The effects of plasma from control animalsand from animals undergoing saline diuresis areshown in Tables I and II; experiments usingplasma from rats are listed in Table I and thoseusing plasma from dogs are listed in Table II.The administration of natriuretic plasma increasedsodium excretion modestly in five experiments,considerably in one (Table I, experiment 1), andnot at all in one (Table I, experiment 5). Urineflow rose only slightly. GFR was not significantlyaltered by the natriuretic plasma in any experi-ment.The effects of control and natriuretic plasmas
on the intrinsic reabsorptive capacity of the proxi-mal tubule are also shown in Tables I and II.
Hormonal Inhibitor of Proximal Tubular Reabsorption
(4)
763

Preliminary experiments revealed that plasmafrom control animals had no effect on intrinsicreabsorptive capacity. The average t4 of 11.4 secand K of 0.062 sec'1 obtained during control peri-ods (Tables I and II) are similar to those obtainedin assay rats not infused with donor plasma. Theinfusion of plasma from animals undergoing salinediuresis uniformly prolonged the tj and decreasedK. In preliminary experiments changes in proxi-mal reabsorption were noted within 10-15 minafter starting the infusion of natriuretic plasma,and maximal effects were attained within 20-30min; upon discontinuing the infusion of natri-uretic plasma the inhibitory effects on proximalreabsorption were completely dissipated within20-30 min. For this reason the equilibrationperiod of 30 min was utilized in all of the experi-ments in Tables I and II. Plasma from natriureticrats increased tj to an average value of 20.1 secand inhibited K by an average of 40% (Table I).Plasma from natriuretic dogs inhibited K by anaverage of 33% (Table II). Upon discontinuingthe infusion of natriuretic plasma and reinfusing
the original control plasma, tj and K returned toor towards the control values in every experiment.In experiment 17 (Table II) in an additionalexperimental period natriuretic plasma inhibitedK to the same degree as in the first experimentalperiod, and in the final recovery period K returnedto the control level. In experiment 18, despite re-versal of the sequence in which natriuretic andcontrol plasmas were infused, K was still 43%lower during the infusion of natriuretic plasmathan during the administration of control plasma.Pretreatment of the assay animals with DOCA(Table I, experiments 4, 5; Table II, experiments13, 14) did not alter the response to either controlor natriuretic plasma.
Average transit time (T) through the proximaltubule fell from a control value of 12.5 sec to 10.6sec during the administration of natriuretic plasmaand then rose to 12.2 sec during the recoveryperiod. Per cent of reabsorption, calculated fromthe K and T in each experiment (equation 3), fellfrom a control level of 53 to 35% during theinfusion of natriuretic plasma and rose again to
TABLE I
Effect of Natriuretic Rat Plasma on Intrinsic Reabsorptive Capacity of the Proximal Tubule
Intrinsic reabsorptivecapacity
Reabsorptive Urine Sodium TransitRat No. half-time (ti/2)* K Inhibition GFR flow excretion time (T)
ml/minsec sec-1 % per g I'/min pEqimin sec
1. Control 11.1 4 0.28 0.062 4.73 6 0.27 12.5Experimental 16.7 :1 0.18 0.042 33 4.81 59 10.89 8.2Recovery 10.6 4 0.16 0.065 4.94 14 5.36 11.6
2. Control 14.9 1 0.15 0.046 5.28 5 0.69 10.3Experimental 19.8 41 0.14 0.035 25 5.36 11 1.46 9.9Recovery 15.1 ± 0.15 0.046 5.12 3 0.64 11.5
3. Control 12.4 4 0.28 0.058 7.12 3 0.07 12.5Experimental 20.8 + 0.16 0.033 42 7.68 5 0.36 10.6Recovery 10.7 i 0.29 0.065 7.91 2 0.07 13.0
4.t Control 12.2 1 0.24 0.057 5.98 4 0.10 11.9Experimental 25.3 1 0.53 0.027 52 5.80 4 0.21 9.0Recovery 10.4 i 0.11 0.067 5.17 3 0.10 11.5
5.t Control 12.3 ± 0.23 0.056 6.45 5 0.63 14.0Experimental 22.8 :1 0.35 0.030 49 6.89 6 0.65 14.0Recovery 11.0 1 0.24 0.063 6.97 7 0.64 14.0
K, intrinsic readsorptive capacity; GFR, glomerular filtration rate.* Mean 4 standard error.$ Assay animal pretreated with DOCA.
764 Rector, Martinez-Maldonado, Kurtzman, Sellman, Oerther, and Seldin
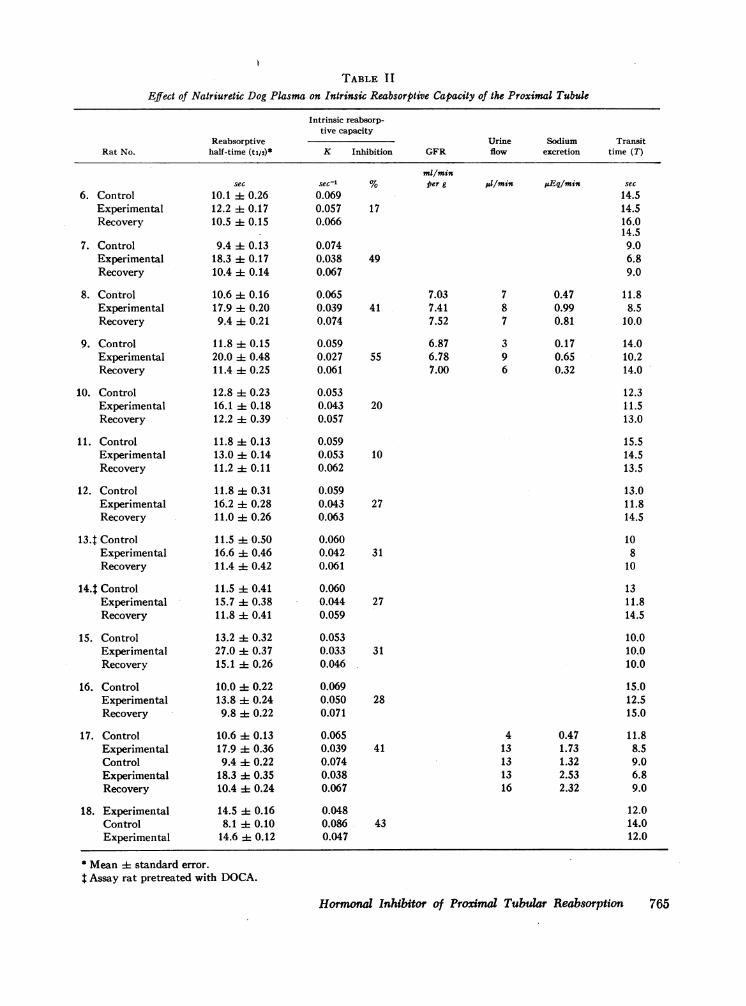
TABLE I IEffect of Natriuretic Dog Plasma on Intrinsic Reabsorptive Capacity of the Proximal Tubule
Intrinsic reabsorp-tive capacity
Reabsorptive Urine Sodium TransitRat No. half-time (tl/2)* K Inhibition GFR flow excretion time (T)
6. ControlExperimentalRecovery
7. ControlExperimentalRecovery
8. ControlExperimentalRecovery
9. ControlExperimentalRecovery
10. ControlExperimentalRecovery
11. ControlExperimentalRecovery
12. ControlExperimentalRecovery
13.t ControlExperimentalRecovery
14.t ControlExperimentalRecovery
15. ControlExperimentalRecovery
16. ControlExperimentalRecovery
17. ControlExperimentalControlExperimentalRecovery
18. ExperimentalControlExperimental
sec10.1 4 0.2612.2 4 0.1710.5 4 0.15
9.4 i 0.1318.3 4 0.1710.4 4 0.14
10.6 ± 0.1617.9 i 0.209.4 4 0.21
11.8 i 0.1520.0 t 0.4811.4 + 0.25
12.8 ± 0.2316.1 0.1812.2 i 0.39
11.8 0.1313.0 ± 0.1411.2 + 0.11
11.8 + 0.3116.2 0.2811.0 : 0.26
11.5 41 0.5016.6 + 0.4611.4 0.42
11.5 :1 0.4115.7 4 0.3811.8 0.41
13.2 + 0.3227.0 : 0.3715.1 + 0.26
10.0 0.2213.8 i 0.249.8 : 0.22
10.6 :1 0.1317.9 1 0.369.4 -4 0.2218.3 i 0.3510.4 0.24
14.5 i 0.168.1 -± 0.1014.6 i 0.12
sealI0.0690.0570.066
0.0740.0380.067
0.0650.0390.074
0.0590.0270.061
0.0530.0430.057
0.0590.0530.062
0.0590.0430.063
0.0600.0420.061
0.0600.0440.059
0.0530.0330.046
0.0690.0500.071
0.0650.0390.0740.0380.067
0.0480.0860.047
ml/minper g 1J/min juEq/min
17
49
7.0341 7.41
7.52
6.8755 6.78
7.00
7 0.478 0.997 0.81
3 0.179 0.656 0.32
20
10
27
31
27
31
28
41
43
4 0.4713 1.7313 1.3213 2.5316 2.32
* Mean standard error.
I Assay rat pretreated with DOCA.
Hormonal Inhibitor of Proximal Tubular Reabsorption 765
see14.514.516.014.59.06.89.0
11.88.5
10.0
14.010.214.0
12.311.513.0
15.514.513.5
13.011.814.5
10810
1311.814.5
10.010.010.0
15.012.515.0
11.88.59.06.89.0
12.014.012.0

TABLE II IEffect of Natriuretic Plasma on Per cent Reabsorption in the Proximal Tubule
Sodium ProximalRat No. GFR excretion Transit time (TF/P)In* reabsorption
ml/min per kg AEqmmis sec % % changeExperiments using rat plasma1. Control 4.70 0.28 12.0 196. -i 0.26 49
Experimental 4.57 0.58 7.9 1.18 1 0.09 15 -70Recovery 5.52 0.99 8.3 1.58 + 0.16 37
2. Control 3.38 0.25 11.2 2.42 1 0.09 59Experimental 3.29 0.30 8.5 2.09 d 0.08 52 -12Recovery 3.29 0.08 11.7 2.88 d 0.14 65
Experiments using dog plasma3. Control 4.84 0.17 11.8 3.33 i 0.42 70
Experimental 5.89 0.13 10.0 2.32 i 0.30 57 -19Recovery 4.45 0.10 11.0 2.78 1 0.09 64
4. Control 4.55 0.76 13.0 2.41 i 0.08 59Experimental 4.56 0.74 7.9 1.72 i 0.05 42 -29Recovery 4.83 0.30 10.8 2.41 ± 0.23 59
5. Control 4.87 0.44 9.5 2.16 + 0.02 54Experimental 4.99 0.31 8.5 1.84 0.10 46 -15Recovery 5.02 0.13 9.5 2.17 d 0.16 54
6. Control 4.98 1.01 15.0 2.13 4 0.18 52Experimental 5.00 1.44 10.1 1.68 d 0.06 41 -22Recovery 5.85 1.03 9.2 1.88 4 0.13 47
7. Control 5.96 0.94 9.8 2.00 4 0.15 50Experimental 4.90 1.14 9.1 1.64 4 0.06 39 -22Recovery 5.33 1.71 9.7 2.08 0.10 52
8. Control 9.76 0.31 11.3 2.10 4 0.08 52Experimental 7.44 2.40 8.8 2.21 i: 0.36 55 +6
9. Control 7.00 0.09 12.0 2.00 4 0.15 50Experimental 7.00 0.34 11.0 1.84 0.26 46 -8Recovery 6.90 1.57 12.3 1.88 1 0.02 47
10. Control 6.23 0.42 13.0 2.71 4 0.39 67Experimental 5.98 1.10 11.7 2.31 4 0.11 57 -15Recovery, 6.05 0.84 10.9 2.54 0.37 61
11. Control 7.64 0.09 16.6 2.61 i 0.21 62Experimental 7.13 0.28 10.3 2.38 d 0.17 58 -6Recovery 6.57 0.38 11.0 2.50 ± 0.16 60
12. Experimental 4.66 11.6 2.36 ± 0.08 58Control 5.55 13.0 3.21 L 0.06 69 -57Experimental 3.70 12.1 2.26 0.07 56
(TF/P)I, ratio of inulin concentration in tubular fluid and plasma.* Mean + standard error.
766 Rector, Martinez-Maldonado, Kurtzman, Sellman, Oerther, and Seldin
v

52%o during the recovery period; the averagedepression in per cent of reabsorption was 34%o.The ratio of tubular volume to GFR per nephron,7rr2d/Vo, also calculated from K and T (equations3 and 4), was not altered by the natriureticplasma.
Effect of per cent reabsorption (group II). Inthe second group of experiments (Table III) theadministration of natriuretic plasma did not pro-duce any significant increase in sodium excretionthat could be attributed to the effects of the plasma.In those experiments in which sodium excretiondid increase during the experimental period, itdid not return to the control level during therecovery period. GFR was not changed by thenatriuretic plasma.
During control periods the (TF/P)n,, averaged2.42 and per cent of reabsorption averaged 58%o.The administration of natriuretic plasma reducedthe (TF/P)I,, in 11 out of 12 experiments; in
one of these (experiment 9) the (TF/P)In didnot rise during the recovery period. During theexperimental period (TF/P) In averaged 1.96 andper cent of reabsorption averaged 47%; per centof reabsorption was depressed an average of 19%.The average transit time (T) fell from 12.3 sec
during the control period to 9.6 sec during theexperimental period and rose to 10.4 sec duringthe recovery period. The intrinsic reabsorptivecapacity, K, calculated from the (TF/P)lIn andtransit time (equation 3), was depressed an aver-age of 22% by the natriuretic plasma. The ratioof tubular volume to GFR per nephron, also cal-culated from the (TF/P)ln and T (equations 3and 4), was not altered during the experimentalperiods.
Effect on water and solute excretion in ratswith diabetes insipidus. To determine the effectof natriuretic plasma on the delivery of filtrate outof the proximal tubule, six studies were performed
ILE IVEffect of Natriuretic Rat Plasma on Water and Solute Excretion in Rats with
Hereditary Diabetes Insipidus*
Urine Urine Sodium PotassiumRat No. osmolality flow Cosm CH20 excretion excretion GFR
mil/minmOsm/kg ;JI/min ;I/min MI/min gEq/min jsEq/min per kg
1. Control 254 8.0 7.0 1.0Experimental 123 42.0 17.8 24.2Recovery 195 12.0 8.0 4.0
2. Control 165 13.0 8.3 4.7 0.24 0.31Experimental 85 60.7 20.5 40.2 1.31 0.56Recovery 196 24.0 17.0 7.0 0.36 0.12
3. Control 231 25.0 19.8 5.2 0.40 0.11 5.36Experimental 189 84.0 51.6 32.4 2.92 1.84 5.16Recovery 235 45.0 35.5 9.5 1.11 0.60 5.20
4. Control 215 4.2 3.2 1.0 0.33 0.19Experimental 126 38.8 15.8 23.0 1.99 0.50Recovery 171 21.1 12.3 8.8 0.21 0.06
5. Control+ 228 11.7 8.3 3.4 0.16 0.14Experimental 162 87.6 45.8 41.8 1.08 1.16Recovery 239 33.3 24.2 9.1 0.45 0.54
6. Control: 195 38.5 26.3 12.2 0.66 0.05 4.98Experimental 145 143.6 66.4 77.2 4.21 3.09 5.19Recovery 195 42.5 30.3 12.3 0.17 0.40 5.00
CosM, osmolar clearance; CH20, solute-free water clearance.* All values are the average of three to four collection periods.t Control plasma was obtained from rats with hereditary diabetes insipidus.
Hormonal Inhibitor of Proximal Tubular Reabsorption 767

in rats with hereditary diabetes insipidus. Asshown in Table IV urine flow rose sharply inevery, experiment during the infusion of natri-uretic plasma, increasing from an average flowof- 16.7 fil/min during control periods to 76.1,ul/min during the experimental periods. Althoughthere was a significant drop in urine flow duringthe recovery period, the average value remainedat levels almost twice the control values. Since theurine flow of 31.3 4ul/min during recovery is closeto the flow before the rats were anesthetized, it ispossible that the relatively low flow at the begin-ning was the consequence of anesthesia.The marked rise in urine volume in all experi-
ments was principally due to an increase in CH20,which rose from a control value of 4.6 Ml/min toa value of 43.2 ul/min during the experimentalperiod. As in the case of urine flow, although therewas a sharp drop in CH20 during recovery, thevalue was about twice that observed during con-trol periods. To exclude the possibility that thecontrol plasma might have had much higher levelsof ADH than the natriuretic plasma, thus causingthe lower urine flow and CH2o during control and''recovery periods than during the experimentalperiods, two experiments (Table IV, experiments5 and 6) were performed with control plasmataken from rats with hereditary diabetes insipidus.The differences between control and natriureticplasmas in these two experiments were the sameas-those in the other four experiments.
Natriuretic plasma also increased solute excre-tion in these rats. Cosm rose from a control level
of 12.1 Ml/min to an experimental value of 33.1dul/min and fell to 20.1 Mil/min during the recoveryperiod. There was a modest increase in sodiumexcretion in all experiments, with a fall towardscontrol levels during the recovery periods. Similarchanges in potassium excretion were also noted.In the two experiments in which it was measuredGFR was not changed.
Characteristics of the hormonal inhibitor ofproximal reabsorption. Natriuretic plasma re-tained its inhibitory activity after standing at roomtemperature for 3-4 hr. As seen in Fig. 1 it alsoretained its activity when stored at 4VC for aslong as 30 days. Inhibitory activity did not appearin control plasma after standing at room tempera-ture for 3-4 hr or after storage at 4VC for severaldays. Thus the substance appears to be relativelystable. The full extent of its heat stability, how-ever, has not yet been ascertained.As shown in Table V the inhibitory activity was
lost when natriuretic plasma was dialyzed againstlarge volumes of Ringer's bicarbonate solution.When natriuretic plasma was dialyzed againstequal volumes of Ringer's bicarbonate solution,inhibitory activity was observed when the dialysatewas infused intravenously. Dialysate of controlplasma had no effect. These studies, showing lossof the inhibitor from natriuretic plasma duringdialysis and its recovery in the dialysate, indicatethat the substance is probably a small molecularweight, water-soluble compound.
Site of action of the proximal inhibitor. Theability to recover the inhibitor in dialysates of
I; I
O 10 20 30
I Days of Aging
CONTROL PLASMA
I,
NATRIURET~~~~~~~~~~~~~~IC0
~~I'01
NATRIURETIC PLASMA
FIGURE 1 Effect of storage at 4VC on inhibitory activity of control and natriuretic plasma.
768 Rector, Martinez-Maldonado, Kurtzman, Sellman, Oerther, and Seldin
50 r
40 V
Per centInhibition
ofReabsorptive
Rate
30 H
20 F
10 F
0
50
40
30
20
10
0FRESH
CON TROLPLASMA
i

TABLE V
Effect of Dialysis on Inhibitory Activityof Natriuretic Plasma
Intrinsic reabsorp-tive capacity
Reabsorp-tive half- Inhi-
Expt. No. time (ti/t) K bition
sec sect %Dialysis against large volumes
of fluid1. Control plasma 11.0 0.063
Natriuretic plasma 15.2 0.045 28Dialyzed natriuretic plasma 11.3 0.061 3Control plasma 11.0 0.063
2. Control plasma 10.8 0.064Natriuretic plasma 16.5 0.042 34Dialyzed natriuretic plasma 11.1 0.062Control plasma 11.0 0.063
Dialysis against equal volumesof fluid*
3. Control plasma 11.3 0.61Natriuretic plasma 17.0 0.41 35Dialyzed natriuretic plasma 10.8 0.64 0Dialysate 16.6 0.42 34Control plasma 10.8 0.64
* Dialysate infused intravenously at 0.04 ml/min. Plasmainfused intravenously at 0.02 ml/min.
natriuretic plasma made it possible to determinewhether the substance could exert an effect whenplaced in contact with the luminal membrane ofthe tubule cell. As shown in Table VI dialysatesof control plasma, when injected directly into thetubular lumen, were reabsorbed at the same rateas Ringer's bicarbonate solution. In contrast,dialysates of natriuretic plasma were reabsorbedat a much slower rate. The dialysates of natri-uretic plasma inhibited K by an average of 35%,a degree of inhibition similar to that observed wheneither natriuretic plasma or dialysates were infusedintravenously. These studies, therefore, clearly in-dicate that the substance can act from the luminalside of the cell.
It was surprising that the intratubular injectionof dialysate, which should produce very high localconcentrations, gave only the same degree of inhi-bition as the intravenous infusions. To pursue thequantitative aspects of this problem further theeffect of serially diluting jugular venous plasmaand dialysates was examined. As shown in Fig. 2
TABLE VIIntratubular Action of Dialysates of Natriuretic Plasma
Intrinsic reab-sorptive capacity
Reabsorptivehalf-time Inhi-
Expt. No. (t1/2) K bition
sec sec'1 %1. Ringer's solution 10.8 4 0.32 0.064
Dialysate-control plasma 9.6 i 0.20 0.072Dialysate-natriuretic plasma 17.7 + 0.24 0.039 45Ringer's solution 9.1 : 0.40 0.076
2. Ringer's solution 15.0 ± 0.42 0.046Dialysate-control plasma 15.5 i 0.54 0.045Dialysate-natriuretic plasma 20.3 i 0.61 0.034 24Ringer's solution 15.0 ± 0.57 0.046
3. Ringer's solution 11.3 + 0.38 0.061Dialysate-control plasma 12.2 d 0.24 0.057Dialysate-natriuretic plasma 20.2 h 0.58 0.034 43Ringer's solution 11.5 ± 0.21 0.061
4. Dialysate-control plasma 11.5 i 0.16 0.060Dialysate-natriuretic plasma 16.2 i 0.09 0.043 29Dialysate-control plasma 11.5 ± 0.08 0.060
5. Dialysate-control plasma 11.0 ± 0.89 0.063Dialysate-natriuretic plasma 16.3 ± 0.31 0.043 33Dialysate-control plasma 11.0 h 0.46 0.063
the inhibitory effect of natriuretic plasma infusedintravenously was almost completely lost whenthe plasma was diluted 1:4 with Ringer's bicar-bonate. In three other experiments natriureticplasma infused intravenously lost all activity ata 1: 2 dilution. In one of these experiments inhibi-tory activity was obtained when the diluted plasmawas infused at twice the rate (0.04 ml/min). Incontrast, dialysate injected into the tubular lumenproduced the same inhibitory effect (approxi-
40 - tntrotubulorDiolyste
Per CentInhibition of 30 -
ProximalReobsorptive 20us
Rate 20 -
10
1:32 1:16 1:8 1:4 1:2 1:1 UndilutedDilution Factor
FIGURE 2 Effect of concentration of hormonal inhibi-tor on intrinsic reabsorptive capacity of the proximaltubule. The dialysis of plasma against an equal volumeof Ringer's bicarbonate results in a concentration of in-hibitor in the dialysate of exactly half that of undialyzedplasma. For this reason the horizontal axis is adjustedso that undiluted dialysate has a dilution factor of 1:1.
Hormonal Inhibitor of Proximal Tubular Reabsorption 769

mately 35%) when diluted 1:2 and 1:4 as withthe undiluted dialysate; activity was less at a 1:8dilution and was completely gone'at a 1: 16 dilu-tion. Since the inhibitor is diluted by a factor of2 in the process of dialysis, the undiluted dialysateis equivalent to a 1 : 1 dilution of plasma and the1:2, 1: 4, 1 :8, and 1: 16 dilutions of dialysateare equivalent respectively to 1: 4, 1: 8, 1: 16, and1:32 dilutions of plasma. The intratubular injec-tion, therefore, appears to result in the deliveryto the reabsorptive site of about eight times moreinhibitor than does the intravenous infusion. Thestudies with the dilutions of dialysate (Fig. 2)indicate that the process of inhibition exhibitssaturation characteristics with a maximum inhibi-tion of approximately 30-40o. For this reason,both the intravenous infusion of plasma and theintratubular injection of dialysate produce thesame inhibitory effect despite the much higherlocal concentrations attained with the intratubularinjections.
DISCUSSIONPreviously' we have suggested that fractionalreabsorption in the proximal tubule is regulatedby'the interaction of two variables: the intrinsicreabsorptive capacity, K, and the ratio of tubularvolume of GFR per nephron [rrr2d/Vo (7, 19,22)]. Recent studies in our laboratory (7) haveshown that expansion of ECF volume with isotonicsaline in rats depresses fractional reabsorption inthe proximal tubule by reducing K 339 andI-rrd/Vo 25go. We proposed that'the decrease inK might be due to some hormonal inhibitor,whereas the decrease in irr2d/Vo might be medi-ated by changes in renal interstitial volume sec-ondary to alterations in the hydrostatic and oncoticpressures in the peritubular capillary bed (7).Indeed, Earley, Martino, and Frielder (15) haveshown that experimental maneuvers which modifyhydrostatic and oncotic pressures in the post-glomerular capillaries markedly influence sodiumexcretion.The present experiments clearly demonstrate
that a hormonal inhibitor of proximal reabsorptionis elaborated in response to expansion of ECFvolunie with isotonic saline. Three lines of evi-dence'support this conclusion. First, infusions ofplasma. from animals undergoing saline diuresisinhibited the. intrinsic reabsorptive capacity of the
proximal tubule, as measured by the shrinking-drop technique. The fact that dialysates of natri-uretic plasma placed directly into the tubularlumen also inhibited intrinsic reabsorptive capacityunequivocally establishes a direct inhibitory actionon the tubular epithelium rather than any indirecteffects secondary to either compositional changesin blood or alterations in renal hemodynamics.Second, infusions of natriuretic plasma reduced(TF/P),1, ratios, indicating a net reduction ofproximal reabsorption. Third, infusions of natri-uretic plasma increased both urine flow and cationexcretion in rats with hereditary diabetes insipidus.Since minimal amounts of water are reabsorbedin the distal diluting segments of the nephron inthe absence of ADH, the rise in urine flow canbe used as evidence of accelerated delivery offiltrate out of the proximal portions of the nephron.Although there was a significant increase in so-dium excretion in all six experiments (Table IV),most of the increased sodium delivered out of theproximal tubule was reabsorbed distally, resultingin augmented CH2O. In addition, potassium excre-tion rose also, indicating that a portion of thesodium delivered out of the proximal tubule wasreabsorbed distally in exchange for potassium. Allthree types of studies, therefore, indicate thatplasma from animals undergoing saline diuresiscontains a humoral agent capable of inhibiting theintrinsic reabsorptive capacity of the proximaltubule, thereby producing a net suppression ofproximal reabsorption and increasing the deliveryof filtrate to more distal portions of the nephron.The exact mechanism by which this hormone
inhibits proximal reabsorption cannot be ascer-tained from the present studies. However, certaincharacteristics of its action can be described. First,the material acts both when infused intravenouslyand when injected directly into the tubule. Thisfinding does not necessarily mean that the hormonereaches the transport site through both surfaces ofthe tubular cell. Substances such as digitalis, vaso-pressin, and aldosterone, which are known to influ-ence transport across epithelial structures (frogskin, toad bladder, and isolated collecting ducts),have been demonstrated to gain access to theirsite of action through only one surface of theepithelial cell, even though the final site of actionmay be on the opposite surface. If the renal tubularcell behaves like these isolated epithelial structures,
770 Rector, Martinez-Maldonado, Kurtzman, Se-lman, Oerther, and Seldin

it is probable that the tubular surface is the portalof entry, since the material injected intratubularlycould have had contact with only the luminal sur-face of the cell. The material injected intravenouslywould not be limited to the blood surface of therenal tubular cell. Since it is dialyzable, it candoubtless pass through the glomerulus and thuswould have access to the luminal surface of thetubule.The second important characteristic of the in-
hibitory action on the proximal tubule is that thehumoral agent exhibits saturation kinetics. Thestudies utilizing serial dilutions of dialysate (Fig.2) indicate that a maximal inhibition of 35-40%is maintained over about an eightfold concentrationrange. As the dialysate is diluted by greater than1:8 the inhibiting effect is rapidly lost. The satura-tion kinetics explain the finding that the infusionof natriuretic plasma at a very low rate (0.02ml/min) inhibited the intrinsic reabsorptive capac-ity to the same degree (Tables I and II) as thatobserved in rats undergoing massive saline diure-sis (7, 8, 26), where the circulating level of theinhibitor has been shown, in unpublished studies,to be very much higher. Indeed, preliminary stud-ies in our laboratory suggest that there is a roughcorrelation between the level of circulating hor-mone and the magnitude of ECF volume expan-sion. The significance of such a variation in plasmalevels is not clear, however, since the presentstudies would indicate that maximal inhibition isachieved at very low plasma concentrations andthat any further rise would have little additionaleffect. It is possible, on the other hand, that if thehormone acts only from the luminal side of thecell and is either reabsorbed or inactivated by thetubular epithelium, variations in plasma concen-tration might be of physiologic importance bydetermining the length of the tubule inhibited.
Finally, the luminal inhibition has a very rapidonset of action, within seconds after its instillationinto the tubular lumen. In this respect it differsfrom aldosterone, where a lag of at least 0.5 hroccurs before transport is stimulated, presumablybecause aldosterone stimulates the synthesis of anintermediate agent, perhaps a specific protein,which directly influences transport (27, 28). Therapid onset of action of the hormonal inhibitorsuggests that it has a more direct effect and doesnot act by stimulating the synthesis of an active
intermediary. Its duration of action is compara-tively short, lasting no more than 20-30 min afterthe discontinuation of the intravenous infusion ofnatriuretic plasma. Sustained natriuresis, therefore,to the extent that it is mediated by the humoralinhibitor, would seem to require persistent expan-sion of ECF volume to insure continued pro-duction.The elaboration of a potent hormonal inhibitor
of proximal reabsorption in response to ECFvolume expansion would explain some, but notall, of the changes observed during saline diuresis.The infusion of natriuretic plasma inhibited theintrinsic reabsorptive capacity to the same degreeas did massive saline infusions (7), yet fractionalreabsorption during free flow was reduced only20% in contrast to the 50% depression observedduring saline diuresis. This discrepancy is probablythe consequence of two factors. First, because ofhigher plasma concentrations of hormone the entirelength of the proximal tubule might be maximallyinhibited during saline diuresis, but not duringinfusion of natriuretic plasma. Since for technicalreasons most shrinking-drop measurements aremade in the earlier portions of the proximal con-volution, this discrepancy might not be detected.Second, saline infusions reduced the ratio of tubu-lar volume to GFR per nephron by approximately25 o, whereas the infusion of natriuretic plasmahad no effect on tubular geometry. Since the abso-lute rate or proximal reabsorption has been shownto vary directly with tubular volume ( 19-22);,2
2 The influence of tubular geometry also affords anattractive explanation for the different effects of the hor-mone and another potent inhibitor of proximal reabsorp-tion, furosemide. As judged by the shrinking-drop tech-nique, furosemide inhibits the intrinsic reabsorptive ca-pacity of the proximal tubule to the same extent as doesthe hormone (29). Yet furosemide does not depress netproximal reabsorption, whereas the hormone does. Theexplanation, doubtless, revolves about the potent inhibi-tion of reabsorption in the ascending limb by furosemide(30, 31). In consequence, large amounts of fluid issue intothe collecting duct, generating internal hydronephrosis,which, in turn, causes proximal tubular dilatation, therebyovercoming the inhibitory effect of the drug (29). Thehormone, at least in the concentrations resulting from theinfusion of natriuretic plasma, exerts little, if any, distaleffect, so that internal hydronephrosis does not occur.Therefore, the inhibition of intrinsic reabsorptive ca-pacity is not masked by the enhancing effects of tubulardilatation, and net inhibition of proximal reabsorptionresults.
Hormonal Inhibitor of Proximal Tubular Reabsorption 7771

the reduction of 7rr2d/Vo during saline diuresiscould account for the greater suppression of frac-tional reabsorption. The fall in 7rr2d/Vo is proba-bly the consequence of an increased interstitialvolume secondary to the combined effects of de-creased plasma protein, reduced renal vascularresistance, and increased blood pressure (7, 14,15) which occur during massive saline infusionsbut not during the infusion of small amounts ofnatriuretic plasma.
Another major difference between the effects ofthe hormone and saline diuresis is that, despitesignificant inhibition of proximal reabsorption, theinfusion of natriuretic plasma failed to increasesodium excretion appreciably. This finding clearlyindicates the importance of inhibition of reabsorp-tion beyond the proximal tubule in promotingsaline diuresis. Evidence from both CH20 studies(32, 33) and micropuncture experiments (34)indicate that the expansion of ECF volume sup-presses sodium reabsorption in the distal nephron.There are several possible explanations as to whythe infusion of saline, but not the infusion of smallamounts of natriuretic plasma containing the hor-mone, inhibits distal reabsorption. It is possiblethat the hormone inhibits sodium reabsorptionalong the entire length of the nephron, but that,owing to the low circulating levels achieved by theplasma infusions, the hormone is reabsorbed fromthe tubular fluid and does not reach sufficientlyhigh boncentrations distally to inhibit the tubule.In contrast, in animals infused with saline farhigher concentrations of hormone circulate in theblood, much of which escapes proximal reabsorp-tion and achieves sufficiently high luminal concen-trations distally to produce maximal inhibition.Even in the presence of high circulating levels,however, the hormone would probably account foronly a fraction of the inhibition of distal reabsorp-tion. Hemodynamic factors, such as increased med-ullary blood flow with washout of the hypertonicmedullary interstitium (35) and redistribution offiltrate from deep nephrons with high capacityloops to superficial nephrons with low capacityloops (16, 36) probably account for most of thereduction of the total effective reabsorptive capac-ity of the distal nephrons during saline diuresis.
It would appear, therefore, that although apotent inhibitor of proximal reabsorption is re-leased during ECF volume expansion, its full
physiologic effect is not expressed unless it isaccompanied by the associated renal hemodynamicchanges that occur during saline diuresis. Thesechanges might potentiate the effect of the hormoneby altering the geometry of the proximal tubule,by increasing medullary blood flow, and by redis-tributing filtrate from deep nephrons with highcapacity loops to superficial nephrons with lowcapacity loops.
ACKNOWLEDGMENTSThis work was supported in part by a grant from theAmerican Heart Association and -in part by traininggrant 5 TI HE 5469 from the National Institutes ofHealth, U. S. Public Health Service. Dr. Martinez-Maldonado performed his work as a trainee of the U. S.Public Health Service, grant 5 TI HE 5469.
REFERENCES
1. DeWardener, H. E., I. H. Mills, W. F. Clapham,and C. J. Hayter. 1961. Studies on the efferentmechanism of sodium diuresis which follows the ad-ministration of intravenous saline in the dog. Clin.Sci. 21: 249.
2. Levinsky, N. G., and R. C. Lalone. 1963. The mecha-nism of sodium diuresis after saline infusion in thedog. J. Clin. Invest. 42: 1261.
3. Rector, F. C., Jr., G. Van Giesen, F. Kiil, and D. W.Seldin. 1964. Influence of expansion of extracellularvolume on tubular reabsorption of sodium independentof changes in glomerular filtration rate and aldos-terone activity. J. Clin. Invest. 43: 341.
4. Levinsky, N. G. 1966. Nonaldosterone influences onrenal sodium transport. Ann. N. Y. Acad. Sci. 139:295.
5. Dirks, J. H., W. J. Cirksena, and R. W. Berliner.1965. The effect of saline infusion on sodium reab-sorption by the proximal tubule of the dog. J. Clin.Invest. 44: 1160.
6. Cortney, M. A., M. Mylle, W. E. Lassiter, and C. W.Gottschalk. 1965. Renal tubular transport of water,solute, and PAH in rats loaded with isotonic saline.Am. J. Physiol. 209: 1199.
7. Rector, F. C., Jr., J. C. Sellman, M. Martinez-Maldonado, and D. W. Seldin. 1967. The mechanismof suppression of proximal tubular reabsorption bysaline infusions. J. Clin. Invest. 46: 47.
8. Landwehr, D. M., R. M. Klose, and G. Giebisch. 1967.Renal tubular sodium and water reabsorption in theisotonic sodium chloride-loaded rat. Am. J. Physiol.212: 1327.
9. Cort, J. H. 1965. Electrolytes, Fluid Dynamics andthe Central Nervous System. Academic Press Inc.,New York. 139.
10. Lichardus, B., and J. W. Pearse. 1966. Evidence fora humoral natriuretic factor released by blood volumeexpansion. Nature. 209: 407.
772 Rector, Martinez-Maldonado, Kurtzman, Sellman, Oerther, and Seldin

11. Johnston, C. I., and J. 0. Davis, 1966. Evidence fromcross-circulation studies for a humoral mechanism inthe natriuresis of saline loading. Proc. Soc. Exptl.Biol. Med. 121: 1058.
12. Johnston, C. I., J. 0. Davis, S. S. Howards, andF. S. Wright. 1967. Cross-circulation experiments onthe mechanism of the natriuresis during saline loadingin the dog. Circulation Res. 20: 1.
13. Earley, L. E., and R. M. Friedler. 1965. Studies on themechanism of natriuresis accompanying increasedrenal blood flow and its role in the renal responseto extracellular volume expansion. J. Clin. Invest. 44:1857.
14. Earley, L. E., and R. M. Friedler. 1966. The effectsof combined renal vasodilatation and pressor agentson renal hemodynamics and the tubular reabsorptionof sodium. L. Clin. Invest. 45: 542.
15. Earley, L. E., J. A Martino, and R M. Friedler 1966.Factors affecting sodium reabsorption by the proxi-mal tubule as determined during blockade of distalsodium reabsorption. J. Clin. Invest. 45: 1668.
16. Barger, A. C. 1966. Renal hemodynamic factors incongestive heart failure. Ann. N. Y. Acad. Sci. 139:276.
17. Starling, E. H., and E. B. Verney. 1925. The secretionof urine as studied on the isolated kidney. Proc. Roy.Soc. (London) Ser. B. 97: 321.
18. Mills, I. H., G. N. Osbeldiston, G. M. Grain. andB. L. Wise. 1966. The effect of change in perfusionpressure and hematocrit in the perfused isolated dogkidney. Proc. Phys. Soc. (London) 44.
19. Rector, F. C., Jr., F. P. Brunner, and D. W. Seldin.1966. Mechanism of glomerulotubular balance. I.Effect of aortic constriction and elevated uretero-pelvic pressure on glomerular filtration rate, frac-tional reabsorption, transit time, and tubular size inthe proximal tubule of the rat. J. Clin. Invest. 45: 590.
20. Gertz, K. H. 1963. Transtubulare Natriumchloridfluisseund Permeabilitiit fur Nichtelektrolyte in proximalenund distalen Konvolut der Rattenniere. PfleugersArch. Ges. Physiol. 276: 336.
21. Brunner, F. P., F. C. Rector, Jr., and D. W. Seldin.1966. Mechanism of glomerulotubular balance II.Regulation of proximal tubular reabsorption by tubu-lar volume, as studied by stopped-flow microperfusion.J. Clin. Invest. 45: 603.
22. Gertz, K. H., J. A. Mangos, G. Braun, and H. D.Pagel. 1965. On the glomerular tubular balance inthe rat kidney. Pfleugers Arch. Ges. Physiol. 285: 360.
23. Vurek, G. G., and S. E. Pegram. 1966. Fluorometricmethod for the determination fo nanogram qualitiesof inulin. Anal. Biochem. 16: 409.
24. Davidson, W. D., and M. A. Sackner. 1963. Simpli-fication of the anthrone method for the determinationsof inulin in clearance studies. J. Lab. Clin. Med. 62:351.
25. Ramsay, J. A., and R. H. J. Brown. 1955. Simplifiedapparatus and procedure for freezing point determi-nation upon small volumes of fluid. J. Sci. Instr. 32:372.
26. Hayslett, J. P., M. Kashgarian, and F. H. Epstein.1967. Changes in proximal and distal reabsorptionproduced by rapid expansion of extracellular fluid.J. Clin. Invest. 46: 1254.
27. Edelman, I. S., R. Borgoroch, and G. A. Porter.1963. On the mechanism of action of aldosterone onsodium transport. The role of protein synthesis.Proc. Natl. Acad. Sci. U. S. 50: 1169.
28. Williamson, H. E. 1963. Mechanism of the anti-natriuretic action of aldosterone. Biochem. Pharmacol.12: 1449.
29. Rector, F. C., Jr., F. P. Brunner, J. C. Sellman, andD. W. Seldin. 1966. Pitfalls in the use of micropunc-ture for the localization of diuretic action. Ann. N. Y.Acad. Sci. 139: 400.
30. Suki, W., F. C. Rector, Jr., and D. W. Seldin. 1965.The site of action of furosemide and other sulfona-mide diuretics in the dog. J. Clin. Invest. 44: 1458.
31. Seldin, D. W., G. Eknoyan, W. N. Suki, and F. C.Rector, Jr. 1966. Localization of diuretic action fromthe pattern of water and electrolyte excretion Ann.N. Y. Acad. Sci. 139: 328.
32. Eknoyan, G., W. N. Suki, F. C. Rector, Jr, andD. W. Seldin. 1967. Functional characteristics of thediluting segment of the dog nephron and the effectof extracellular volume expansion on its reabsorptivecapacity. J. Clin. Invest. 46: 1178.
33. Stein, R. M., R. G. Abramson, T. Kahn, and M. F.Levitt. 1967. Effects of hypotonic saline loading inhydrated dog: evidence for a saline-induced limit ondistal tubular sodium transport. J. Clin. Invest. 46:1205.
34. Sellman, J. C., F. J. Oerther, F. C. Rector, Jr., andD. W. Seldin. 1967. Evidence for inhibition of distaltubular reabsorption during saline diuresis. Clin. Res.15: 370. (Abstr.)
35. Earley, L. E., and R. M. Friedler. 1964. Observationson the mechanism of decreased tubular reabsorptionof sodium and water during saline loading. J. Clin.Invest. 43: 1928.
36. Sellman, J. C., M. Martinez-Maldonado, F. C. Rector,Jr., and D. W. Seldin. 1967. Influence of saline infu-sion on renal cortical hemodynamics and proximaltubular reabsorption. Clin. Res. 15: 78. (Abstr.)
Hormonal Inhibitor of Proximal Tubular Reabsorption 773