defining the host protective antigens secreted by the
TRANSCRIPT

Defining the host protective antigens secreted by
the murine whipworm, Trichuris muris
A thesis submitted to The University of Manchester for the degree of Doctor of Philosophy in the Faculty of Biology, Medicine and Health
2017
Rebecca K Shears
School of Biological Sciences
Division of Infection, Immunity and Respiratory Medicine

2
Table of contents
List of Figures ........................................................................................................ 5
List of Tables .......................................................................................................... 8
Abbreviations ....................................................................................................... 10
Abstract ................................................................................................................ 12
Acknowledgements ............................................................................................. 13
Declaration ........................................................................................................... 14
Copyright statement ............................................................................................ 14
Chapter 1: Introduction ....................................................................................... 15 1.1 Gastrointestinal nematodes: their prevalence, disease burden and the need for prophylactic vaccines ........................................................... 16 1.2 T. muris as a model for T. trichiura ................................................... 18
1.2.1 The life cycle of T. muris .................................................................. 18 1.2.2 Immune response during acute and chronic T. muris infection ....... 20 1.2.3 Mechanisms of T. muris expulsion .................................................. 23
1.2.3.1 Mucus production and other goblet cell secretions .................... 26 1.2.3.2 Increased rate of epithelial cell turnover ..................................... 28 1.2.3.3 Intestinal muscle hyper-contractility ........................................... 29 1.2.3.4 Mast cells and IgE production .................................................... 30 1.2.3.5 IgG antibody production and B cells ........................................... 30 1.2.3.6 Innate lymphoid cells and other early sources of Th2 cytokines 31 1.2.3.7 Regulation of the immune response during T. muris infection .... 32
1.3 Clinical and pre-clinical helminth vaccine candidates .................... 33 1.3.1 Hookworm vaccine candidates ........................................................ 33 1.3.2 Pre-clinical Ascaris vaccine candidates ........................................... 35 1.3.3 Experimental Trichuris vaccines ...................................................... 35 1.3.4 Schistosome vaccines ..................................................................... 39 1.3.5 Cestode vaccine candidates ............................................................ 40 1.3.6 The role of adjuvants in vaccines .................................................... 40
1.4 Extracellular vesicles as a source of antigenic material ................. 43 1.4.1 Exosome biogenesis and isolation from biological samples ............ 43 1.4.2 Exosome release by parasitic helminths ......................................... 45
1.5 Aims and objectives ........................................................................... 48
Chapter 2: Materials and methods ..................................................................... 49 2.1 Maintenance of animals ...................................................................... 50 2.2 Maintenance of parasites, ELV removal and preparation of adult ES 50 2.3 Preparation of larval ES ...................................................................... 51 2.4 Egg infectivity and dosage ................................................................. 52 2.5 Quantification of worm burdens ........................................................ 52 2.6 Collection of sera ................................................................................ 53 2.7 Anti-parasite IgG1 and IgG2a ELISAs ............................................... 53 2.8 Lymphocyte re-stimulation assay ..................................................... 53
2.8.1 Measuring cytokine production in cell supernatants ........................ 54 2.9 Fractionation of adult ES .................................................................... 56

3
2.9.1 Fractionation of adult ES by gel filtration chromatography .............. 56 2.9.2 Fractionation of adult ES and pool 3 by anion exchange and gel filtration chromatography ........................................................................... 57
2.10 SDS-PAGE ........................................................................................... 57 2.10.1 Coomassie blue staining ................................................................ 58 2.10.2 Silver staining of SDS-PAGE gels ................................................. 58
2.11 Assessing protein concentration of samples ................................... 58 2.12 Western blotting .................................................................................. 58
2.12.1 Western blotting using serum from T. muris infected mice ............ 59 2.12.2 Western blotting to detect His-tagged recombinant proteins ......... 59
2.13 Mass spectrometry and proteomic analysis of ES components .... 60 2.13.1 Mass spectrometry analysis of T. muris ELVs ............................... 61
2.14 Vaccination studies ............................................................................ 62 2.14.1 Proteinase K treatment of ES and subsequent vaccination ........... 63 2.14.2 Serum transfer from vaccinated to unvaccinated mice .................. 63 2.14.3 ELV vaccination studies................................................................. 64
2.15 DNA synthesis, transfections and collection of recombinant proteins ........................................................................................................ 65 2.16 Purification of recombinant proteins ................................................ 66 2.17 TEM analysis of ELV samples ............................................................ 67 2.18 DLS of ELVs ......................................................................................... 67 2.19 ELV fusion assay ................................................................................ 68 2.20 Graphing and statistical analysis ...................................................... 68
Chapter 3: Defining the host protective components within the soluble portion of T. muris ES ......................................................................................... 70
3.1 Introduction ......................................................................................... 71 3.2 Preparation of T. muris ES for fractionation using gel filtration chromatography .......................................................................................... 73
3.2.3 Investigating the suitability of gel filtration media to fractionate ES into smaller sub-groups.............................................................................. 74 3.2.2 Division of ES into four sub-groups using Superose 12 gel filtration media ......................................................................................................... 76 3.2.3 Investigating the cellular immune response to pools 2-4 ................. 78 3.2.4 Assessment of anti-parasite IgG serum antibody response during acute T. muris infection .............................................................................. 80
3.3 Vaccination with pools 2 to 4 stimulates protective immunity ....... 82 3.3.1 Assessment of antibody response following vaccination with pools 2 to 4 and subsequent infection .................................................................... 84
3.4 Vaccination with ES induces long-lasting protection against a subsequent low dose infection .................................................................. 89
3.4.1 Assessment of antibody response following long-term vaccination and subsequent infection .................................................................................. 90
3.5 Proteinase K degradation of ES abrogates its protective properties …………………………………………………………………………………94 3.6 Transfer of serum from ES vaccinated mice does not confer resistance to unvaccinated mice ............................................................... 95 3.7 Fractionation of ES by anion exchange chromatography ............... 97 3.8 Fractionation of pool 3 by anion exchange chromatography ......... 99 3.9 Vaccination with L2 ES induces protective immunity ................... 101

4
3.10 Evaluation of the AKR mouse model for vaccination studies ....... 104 3.11 Discussion ......................................................................................... 105
Chapter 4: Identification of immunogenic candidates within T. muris ES ………………………………………………………………………………………..112
4.1 Chapter introduction ......................................................................... 113 4.2 Identification of immunogenic candidates using a size exclusion chromatography and proteomics approach ........................................... 114
4.2.1 Fractionating ES using Superose 12 gel filtration media ............... 114 4.2.2 Fractionation of pool 3 using Superdex 75 gel filtration media ...... 122
4.3 Identification and synthesis of potential immunogenic candidates 128 4.4 In vitro assessment of the immunogenicity of candidate proteins 133 4.6 Discussion ......................................................................................... 140
Chapter 5: T. muris ELVs as a source of immunogenic material .................. 148 5.1 Chapter introduction ......................................................................... 149 5.2 ELVs can be isolated from T. muris ES by ultracentrifugation ..... 149 5.3 Mass spectrometry analysis shows that T. muris ELVs contain typical exosome markers and are enriched for proteins lacking a signal peptide ....................................................................................................... 151 5.4 Exosomes are able to fuse with colonic epithelial cells in vitro ... 154 5.5 Vaccination with T. muris ELVs can induce protective immunity and protection is dependent on intact vesicles ...................................... 156 5.6 Vaccination with ELVs boosts IgG1 serum antibody response to soluble ES components ............................................................................ 157 5.7 Identification of ELV components targeted by serum IgG antibodies following vaccination ................................................................................ 158 5.8 Discussion ......................................................................................... 160
6 Summary discussion ................................................................................... 166 6.1 Identification of immunogenic candidates within T. muris ES ...... 167 6.2 Identification of immunogenic candidates within T. muris ELVs . 171 6.3 Other considerations for Trichuris vaccine design ....................... 173 6.4 Conclusions and future perspectives ............................................. 175
References.......................................................................................................... 177
Appendix 1.......................................................................................................... 201
Appendix 2.......................................................................................................... 205
Appendix 3.......................................................................................................... 215
Word count: 40,277

5
List of Figures
Figure 1.1. Life cycle of T. muris…………………………………………………...19
Figure 1.2. Spectrum of immune responses to T. muris in different mouse
strains................................................................................................................20
Figure 1.3. Immune response during acute T. muris infection………………….25
Figure 1.4. Biogenesis of exosomes……………………………………………....44
Figure 3.1. Preparation of ES by removing P43 using nickel affinity chromatography………………………………………………………………………74 Figure 3.2. Investigating the suitability of Superdex 75, Superdex 200 and Superose 12 gel filtration media for fractionating ES by size exclusion chromatography………………………………………………………………………75 Figure 3.3. Fractionation of ES using the 24 ml Superose 12 column…………77 Figure 3.4. Cellular immune response to pools 2-4 and unfractionated ES…..79 Figure 3.5. Anti-parasite IgG1 serum antibody response to pools 2 to 4 and unfractionated ES…………………………………………………………………….81 Figure 3.6. Western blots showing anti-parasite IgG serum antibody response for T. muris infected (A) and naïve (B) C57BL/6 mice……………………………82 Figure 3.7. Vaccination of male C57BL/6 mice with pools 2 to 4 or unfractionated ES induces protective immunity…………………………………..83 Figure 3.8. Anti-parasite IgG1 serum antibody response for mice vaccinated with pools 2 to 4 or unfractionated ES……………………………………………..85 Figure 3.9. Anti-parasite IgG2a serum antibody levels for mice vaccinated with pools 2 to 4 or unfractionated ES…….…………………………………………….87 Figure 3.10. Western blots showing anti-ES serum antibody response for the sham, pool 2, 3, 4 or unfractionated ES vaccination groups…………………….88 Figure 3.11. Vaccination with pool 3 or unfractionated ES can induce long-lasting protective immunity…….……………………………………………………90 Figure 3.12. Anti-parasite IgG1 serum antibody levels following long-term vaccination and subsequent infection…….………………………………………..91 Figure 3.13. Anti-parasite IgG2a serum antibody levels following long-term vaccination and subsequent infection………..…………………………………….93

6
Figure 3.14. Proteinase K treatment of ES abrogates its protective properties……………………………………………………………………………...94 Figure 3.15. Worm burdens following transfer of serum from sham or ES vaccinated mice…….………………………………………………………………...94 Figure 3.16. Anti-parasite serum IgG1 and IgG2a antibody response for serum transfer groups…….………………………………………………………………….97 Figure 3.17. Fractionation of ES by anion exchange chromatography…....…..98 Figure 3.18. Worm burdens for male C57BL/6 mice vaccinated with ES sub-groups A to F…………………………………………………………………….99 Figure 3.19. Fractionation of pool 3 by anion exchange chromatography…...100 Figure 3.20. Worm burdens for male C57BL/6 mice vaccinated with pool 3 sub-groups A to D…….…………………………………………………………….101 Figure 3.21. Vaccination with L2 or adult ES can induce protective immunity……………………………………………………………………………..102 Figure 3.22. Western blots showing IgG antibody response to L2 and adult ES …….………………………………………………………………………………….103 Figure 3.23. SDS-PAGE separation of L2 and adult ES………………………104 Figure 3.24. Vaccination of male AKR mice with ES components protects against a subsequent high dose infection……………………………………….105 Figure 4.1. Cytokine production by infection-primed and naïve lymphocytes in response to stimulation with Superose 12 fractions 22 to 32………………….116 Figure 4.2. Cytokine production by infection-primed and naïve lymphocytes in response to Superdex 75 fractions 17 to 23……………………………………..123 Figure 4.3. Strategy for selecting immunogenic candidates for further investigation…….…………………………………………………………………...128 Figure 4.4. Purification of T. muris recombinant proteins……………………...131 Figure 4.5. Cytokine release by infection-primed and naïve lymphocytes following stimulation with recombinant proteins…………..…………………….134 Figure 4.6. Vaccination with recombinant T. muris proteins formulated with aluminum hydroxide does not induce protective immunity in male C57BL/6 mice …….………………………………………………………………………………….136

7
Figure 4.7. Vaccination with recombinant T. muris proteins formulated with Freund’s adjuvants or Montanide ISA 720 does not induce protective immunity in male C57BL/6 mice…….………………………………………………………..138 Figure 4.8. Vaccination with T. muris recombinant proteins induced production of IgG1 antibodies specific for these proteins…….…………………………..…139 Figure 5.1. Visualisation of ELVs isolated from T. muris ES………………….149 Figure 5.2. Size range of a typical T. muris ELV sample…….………………..150 Figure 5.3. Uptake of PKH26 labeled ELVs by colonic epithelial (HT-29) cells …….………………………………………………………………………………….155 Figure 5.4. Vaccination with T. muris ELVs induces protective immunity……156 Figure 5.5. IgG1 and IgG2a serum antibody response against ES following vaccination with ELVs..…….……………………………………………………....157 Figure 5.6. Western blots showing anti-ELV and anti-ES serum IgG response for sham, ELV and ES vaccination groups…….………………………………...158 Figure A1.1. Sequence for pCep-His vector used for expression of recombinant T. muris proteins.…….……………………………………………………………..201 Figure A2.1. BLAST search results for Ion trans 2 and Pfam-B 17708-domain containing protein (TMUE_s0066001200) …….………………………………...207 Figure A2.2. Anti-parasite IgG1 serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Montanide ISA 720…………..208 Figure A2.3. Anti-parasite IgG2a serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Montanide ISA 720…….209 Figure A2.4. Anti-parasite IgG1 serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with aluminum hydroxide…………210 Figure A2.5. Anti-parasite IgG2a serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with aluminum hydroxide……211 Figure A2.6. Anti-parasite IgG1 serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Freund’s adjuvants…………..212 Figure A2.7. Anti-parasite IgG2a serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Freund’s adjuvants……..213 Figure A2.8. Vaccination with T. muris recombinant proteins induced production of IgG2a antibodies specific for these proteins…………………….214

8
Figure A3.1. BLAST search results for T. muris TSP-1 domain containing protein (TMUE_s0070003500) …….……………………………………………..215 Figure A3.2. Worm burden and IgG1/IgG2a serum antibody response for ES titration experiment…….…………………………………………………………...216
List of Tables
Table 2.1. Description of mouse strains and infection dose used for experiments described in this thesis……………………………………………….50 Table 2.2. Mascot search criteria…….…………………………………………….55 Table 4.1. List of identified proteins with peak abundance around Superose 12 fractions 24 to 27…….…………………………………………………………..…118 Table 4.2. List of identified proteins with peak abundance around Superdex 75 fractions 20 to 22. …….……………………………………………………………125 Table 4.3. List of potential immunogenic candidates…….……………………..128 Table 5.1. List of exosome markers identified in T. muris ELV samples……..151
Table 5.2. List of shared ELV and ES proteins …...….…...……………………152
Table 5.3. Possible identities of ELV components targeted by IgG antibodies following vaccination…….……………………………………………………….…159 Table A1.1. List of proteins identified within adult T. muris ES………………...CD Table A1.2. List of proteins identified within pool 1……………………………..CD Table A1.3. List of proteins identified within pool 2………………………...…...CD Table A1.4. List of proteins identified within pool 3……………………………..CD Table A1.5. List of proteins identified within pool 4………………………...…...CD
Table A1.6. List of L2 larval ES protein……………………………………...…...CD
Table A2.1. Mass spectrometry analysis of recombinant Serpin…….……….205
Table A2.2. Mass spectrometry analysis of recombinant Lactoglutathione lyase ………………………………………………………………………………………..205
Table A2.3. Mass spectrometry analysis of recombinant Translationally controlled tumour protein…….………….…………………………………………205

9
Table A2.4. Mass spectrometry analysis of recombinant TPD52 domain containing protein…….………………………………………………………….....206
Table A2.5. Mass spectrometry analysis of recombinant Hypothetical protein ………………………………………………………………………………………..206
Table A2.6. Comparison of protein sequences for T. muris immunogenic candidates with T. trichiura homologues…………………………………………206
Table A2.7. BLAST search results for T. muris Ion trans 2 and Pfam-B 17708-domain containing protein (TMUE_s0066001200). ………….…………………207
Table A3.1. List of T. muris ELV proteins………………………………………...CD
Table A3.2. BLAST search results show that there is significant homology between the T. muris TSP-1 domain containing protein (TMUE_s0070003500) and S. mansoni proteins…….…………………………………………………….215 Table A3.3. BLAST search results for T. muris Vacuolar protein sorting associated protein (TMUE_s0093001800) …….………………………………..217

10
Abbreviations
ABTS 2, 2'-azino-bis (3-ethylbenzthiazoline)-6-sulphonic acid
Alix Apoptosis linked gene 2 interacting protein X 1
ANOVA Analysis of variance
BCA Bicinchoninic acid
BCIP 5-bromo-4-chloro-3-indolyl phosphate
BSA Bovine serum albumin
CaCl2 Calcium chloride
cDNA Complementary DNA
CFA Complete Freund’s adjuvant
CO2 Carbon dioxide
DC Dentritic cell
dH2O Distilled water
ddH2O Double-distilled water
DIR Drug induced resistance
DLS Dynamic light scattering
DMEM Dulbecco’s modified Eagle’s medium
DTT Dithiothreitol
DNA Deoxyribonucleic acid
ELISA Enzyme-linked immunosorbent assay
EM Electron microscopy
ES Excretory/secretory product without P43 and ELVs
ES+P43 Native excretory/secretory product
EV Extracellular vesicle
ELV Exosome-like vesicle
FBS Foetal bovine serum
HCl Hydrochloric acid
HEK293 Human embryonic kidney 293
His Histidine
IFA Incomplete Freund’s adjuvant
IFN-γ Interferon gamma
Ig Immunoglobulin
IL Interleukin
ILV Intraluminal vesicle
LC-MS Liquid chromatography-tandem mass spectrometry
MACS Magnetic affinity cell sorting
MHC Major histocompatibility complex
mRNA Messenger ribonucleic acid
miRNA Micro ribonucleic acid
MLN Mesenteric lymph node
MS/MS Tandem mass spectrometry
MVB Multi-vesicular body
Mw Molecular weight

11
MWCO Molecular weight cut off
NaCl Sodium chloride
Na2CO3 Sodium carbonate
NBT Nitro blue tetrazolium
P43 Poly-cysteine and histidine tailed protein isoform 2
PBS Phosphate buffered saline
PBST 0.05% v/v Tween 20 in PBS
PCR Polymerase chain reaction
p.i. Post infection
Ripa Radioimmunoprecipitation assay
RNA Ribonucleic acid
RPMI Roswell park memorial institute medium
SA-POD Streptavidin peroxidase
SCID Severe combined immunodeficiency
SDS Sodium dodecyl sulphate
SIV Simian immunodeficiency virus
SDS-PAGE Sodium dodecyl sulphate-polyacrylamide gel electrophoresis
SEM Standard error of mean
STH Soil-transmitted helminth
TBST Tris-buffered saline-Tween
TFE Trifluoroethylene
TGF Transforming growth factor
Th T helper cell
TLR Toll-like receptor
TNF-α Tumour necrosis factor α
Treg T regulatory cell
TSP Tetraspanin
Tween 20 Polyoxyethylene(20)sorbitan monolaurate

12
Abstract
Soil-transmitted helminths are a major cause of morbidity for humans and their livestock. A
combination of better sanitation, anthelminthic drugs and vaccines are predicted to reduce
the morbidity of these parasites in humans. The drugs currently used to treat these
infections, albendazole and mebendazole, are fairly ineffective against Trichuris trichiura
(human whipworm), and there are reports of drug resistance arising within parasite
populations in Vietnam and Zanzibar. There are also no commercially available vaccines
against human STH species, and very few against their veterinary counterparts. The
murine whipworm, T. muris, has been used for over 50 years as a model for T. trichiura.
These parasites share homology at the genomic and transcriptomic levels, and the immune
responses associated with both acute and chronic infection have been well studied using
the T. muris mouse model.
T. muris excretory/secretory products have been studied in the context of vaccination for
over four decades, however relatively little progress has been made towards identifying the
molecular components that stimulate protective immunity following vaccination or during
acute infection. Here, a stringent selection protocol was developed using chromatography
and mass spectrometry methods combined with a measurement of T cell cytokine
production. The work presented in this thesis provides a novel framework for identifying
potential immunogenic candidates within adult T. muris excretory/secretory products.
Exosome-like vesicles isolated from adult T. muris ES were also explored as a source of
host protective material. Vaccination with exosome-like vesicles protected male C57BL/6
mice from a subsequent low dose infection, which would ordinarily progress to chronicity,
and a number of potential immunogenic candidates were identified.
Over the course of this thesis, several important observations were made relating to
characteristics of the immune response induced by vaccination with ES. Firstly,
proteinaceous material is likely to be responsible for the host protective properties of ES.
Secondly, vaccination with ES products stimulates long-lasting immunity. Thirdly,
vaccination with ES collected from both larval and adult stages stimulates protective
immunity. The number of potential immunogenic candidates has also been narrowed down
from over four hundred to just eleven. Given the homology between T. muris and
T. trichiura at both the genomic and transcriptomic levels, this work has the potential to
advance vaccine design for T. trichiura and other Trichuris parasites.

13
Acknowledgements
First and foremost I would like to thank my supervisors, Dave and Richard for
the continued help and guidance throughout my PhD. Thank you for being such
supportive supervisors and for making the PhD so enjoyable. I am also grateful
to the Wellcome Trust for funding this project.
I want to thank Dr Allison Bancroft for the excellent training and invaluable
discussions over the years and Dr Caroline Ridley for teaching me everything I
know about chromatography and protein purification.
Thank you to all members of the Thornton and Grencis labs for the useful
advice I have received in lab meetings and for keeping me smiling throughout
the PhD.
I want to thank my friends – old and new – particularly the Come Dine With Me
girls, who have helped make so many fun memories.
Lastly, I’d like to thank my parents and Rory for their continued love, support
and motivation.

14
Declaration
I declare that that no portion of the work referred to in this thesis has been submitted in
support of an application for another degree or qualification at this or any other
university or institute of learning.
Copyright statement
The author of this thesis (including any appendices and/or schedules to this thesis)
owns certain copyright or related rights in it (the “Copyright”), and she has given The
University of Manchester certain rights to use such Copyright, including for
administrative purposes.
Copies of this thesis, either in full or in extracts and whether in hard or electronic copy,
may be made only in accordance with the Copyright, Designs and Patents Act 1988 (as
amended) and regulations issued under it or, where appropriate, in accordance with
licensing agreements which the University has from time to time. This page must form
part of any such copies made.
The ownership of certain Copyright, patents, designs, trademarks and other intellectual
property (the “Intellectual Property”) and any reproductions of copyright works in the
thesis, for example graphs and tables (“Reproductions”), which may be described in
this thesis, may not be owned by the author and may be owned by third parties. Such
Intellectual Property and Reproductions cannot and must not be made available for use
without the prior written permission of the owner(s) of the relevant Intellectual Property
and/or Reproductions.
Further information on the conditions under which disclosure, publication and
commercialisation of this thesis, the Copyright and any Intellectual Property and/or
reproductions described in it may take place is available in the University IP Policy (see
http://documents.manchester.ac.uk/display.aspx?DocID=24420), in any relevant
Thesis restriction declarations deposited in the University Library, The University
Library’s regulations (http://www.library.manchester.ac.uk/about/regulations/) and in
The University’s policy on Presentation of Theses.

15
Chapter 1: Introduction

16
1.1 Gastrointestinal nematodes: their prevalence, disease burden and
the need for prophylactic vaccines
Gastrointestinal nematodes are a major source of morbidity in humans and their
livestock (1). The four main species of clinical relevance are the hookworms
Necator americanus and Ancylostoma duodenale, the roundworm Ascaris
lumbricoides, and whipworm Trichuris trichiura (2, 3). Infections occur following
the ingestion of embryonated eggs (A. lumbricoides and T. trichiura) or after
contact with larvae (hookworms) in the soil, hence why these parasites are also
known as soil-transmitted helminths (STHs) (2, 3). These parasites are endemic
in many parts of tropical and subtropical Africa, Asia and Central America (2, 4).
Over a billion people are infected with one or more STH, and estimates for the
disease burden of these parasites range from 4.5 to 39 million disability
adjusted life years (5, 6). STH infections have a profound effect on school
attendance and economic productivity in endemic areas, and the combined
morbidity of these infections is equal to that of malaria, tuberculosis or HIV, yet
they receive comparatively little research attention (2, 7).
For T. trichiura, heavy worm burdens are associated with Trichuris dysentery
syndrome, symptoms of which include stomach pain, chronic, bloody diarrhea,
and in extreme cases, rectal prolapse (8). Population studies demonstrate that
the distribution of worm burdens is highly overdispersed, meaning that the
majority of people harbour low-level infections, while relatively fewer people
have heavy worm burdens (9, 10). In endemic areas, T. trichiura infections are
acquired from a young age, with 90% of children under 5 infected (9). Infection
rates remain high across each age group, with 85% of 40 year olds
infected (9, 10). Studies have also been carried out to investigate re-infection
rates following anthelminthic treatment (9, 10). These reports demonstrate that
people with low-level infections tend to acquire low-level infections following
anthelminthic treatment, whereas individuals with high worm burdens tend to
acquire high worm burdens (9, 10). These data suggest a direct relationship
between initial worm burden and worm burden upon re-infection, which may

17
suggest that some people are naturally more resistant to T. trichiura than
others (9, 10).
The major anthelminthics used to treat STH infections are the benzamidizole
drugs, albendazole and mebendazole, which bind to and inhibit nematode
β-tubulin, preventing microtubule depolymerisation and killing the worm over a
number of days (2). Some studies have shown that treating children regularly
with anthelminthics as part of mass drug administration programs has a positive
effect on children’s iron levels and physical development (11-13), however, a
more recent meta-analysis found that a single dose of albendazole or
mebendazole has poor efficacy against T. trichiura, and that treatment with
benzamidazoles alone had little impact on hookworm-associated anaemia
(7, 14). Combining albendazole with praziquantel did however improve
haemaglobin levels in hookworm patients with moderate anaemia (11).
Recently a new class of anthelminthics, the dihydrobenz[e][1,4]oxazepin-2(3H)-
ones have been shown to have in vitro and in vivo efficacy against T. muris
(14, 15). This suggests that new anthelminthic drug treatments may be
available in the near future, however studies show that post-treatment rates of
re-infection are high, and that drug treatment may prevent the development of
protective immunity (16-19). There is also evidence of benzamidazole
resistance arising, based on field studies carried out in Vietnam and
Zanzibar (20, 21).
A combination of anthelminthic drugs, vaccines and improvements to sanitation
are predicted to reduce the morbidity of STHs (22). There are currently no
anthelminthic vaccines licensed for use in humans, however there are two
hookworm vaccine candidates undergoing clinical trials, and several pre-clinical
vaccine candidates for schistosome species and A. lumbricoides (3, 22-27).
Comparatively little progress has been made towards identifying vaccine
candidates for T. trichiura, although vaccination with material excreted/secreted
by the parasite (known as ES) has been shown to stimulate protective immunity
in a number of mouse models (19, 28, 29). In addition, a recent paper showed
that vaccination of mice with recombinant serine/threonine phosphatase 2A

18
(from Angiostrongylus costaricensis) linked to a synthetic lipid, oleic-vinyl
sulphone, lead to expulsion of an established T. muris infection in AKR
mice (30). These advances will be discussed in section 1.3.
1.2 T. muris as a model for T. trichiura
The approach used in this thesis to identify vaccine candidates for Trichuris
parasites will focus on identifying T. muris antigens that induce protective
immunity in mice. T. muris is a naturally-occurring murine parasite that has
been used for decades as a model for T. trichiura (31, 32). The T. muris model
has enabled researchers to dissect the immune responses associated with
acute (resolving) and chronic infection (reviewed in section 1.2.2) (31, 33).
There is extensive homology between the genome and transcriptome of
T. muris and T. trichiura (33), and therefore the work presented in this thesis
has the potential to advance vaccine design for T. trichiura and other Trichuris
parasites.
1.2.1 The life cycle of T. muris
Trichuris species are transmitted via the faecal-oral route, and the host is
infected following the ingestion of embryonated eggs in contaminated food,
water or soil (31). The eggs hatch upon reaching the caecum in response to
specific microbial cues, releasing the first stage larvae, L1, which burrow into
the caecal epithelium using the stichosome at the anterior end of the
worm (31, 34). The burrowing of larvae into the epithelium leads to the
formation of structures resembling syncytial tunnels, and the parasite appears
to be in direct contact with the cytoplasm of host cells throughout the
infection (35). The larvae grow outwards into the lumen and undergo four
moults to become adults. Adult worms have a characteristic whip-like shape,
with a thin anterior that is embedded in the caecal epithelium, and a thick

19
posterior that protrudes out into the lumen to facilitate mating (31). The parasite
is dioecious, and females release eggs into the caecal lumen following mating.
These eggs are expelled in the faeces and must undergo a period of
embryonation outside the host before they are infective (Figure 1.1) (31).
Figure 1.1. Life cycle of T. muris. Infection occurs via the faecal-oral route. Eggs hatch in the host caecum releasing L1 larvae, which burrow into the caecal epithelial crypts. Larvae undergo four moults to become adults at the time points specified on the diagram. Male and female worms mate and eggs are released into the caecal lumen, where they exit the host in the faeces. Eggs must undergo a period of embryonation before they are infective. P.i. = post-infection.
There are over 50 species within the Trichuris genus, and each species has a
specific mammalian host (34). Egg hatching is triggered by host body
temperature and is reliant on the host microflora, which may explain why these
parasites reside in the caecum and colon, where the largest number of bacteria
are found within the body (34). Hayes and co-workers showed that treating mice
with antibiotics prior to and during T. muris infection lead to a reduction in worm
burden, while incubating embryonated eggs with faecal explants or with various
laboratory strains of bacteria and yeast triggered egg hatching over a period of
30 minutes to 18 hours. The authors found that bacteria cluster at the opercula,
the sticky plugs at either end of the egg, which is also where the worm
emerge (34).
Ingestion of embryonated eggs
Lumen
Eggs hatch in caecum, L1 larvae burrow into epithelium
Lamina propria
L2 larvae
L3 larvae
L4 larvae Adult
Mating produces eggs, which are expelled in the faeces
Day 9-11 p.i.
90 mins
Day 32 p.i.

20
1.2.2 Immune response during acute and chronic T. muris infection
Studies with inbred mice and gene knock out animals have been key for
dissecting the immune responses associated with acute and chronic T. muris
infection (31, 36). These studies have highlighted the role of host genetics,
gender, infection dose, and parasite strain in the relative
resistance/susceptibility to infection (Figure 1.2) (31, 36).
Figure 1.2. Spectrum of immune responses to T. muris in different mouse strains. Diagram was constructed based on published data (references in brackets) and observations from our laboratory.
The importance of T cells during T. muris expulsion was recognised as early as
1974 by Wakelin and Selby, who demonstrated that thymectomised and
irradiated NIH mice were unable to expel worms, but that transferring MLN cells
or thymocytes from untreated animals could reverse this (37). Lee and
colleagues also showed that transferring T cell (but not B cell) enriched
lymphocyte populations from T. muris infected CBA/Ca mice protected naïve
mice of the same strain from a subsequent T. muris infection (38). Ito and
co-workers reported similar findings, showing that transferring splenocytes or
thymocytes from wild type BALB/c mice led to T. muris expulsion in normally
susceptible athymic Nude mice (BALB/c background) (39). Researchers
recognised a specific role for CD4 T cells in worm expulsion by demonstrating
that depleting these cells using neutralising antibodies rendered wild type
BALB/c (39, 42, 48) C57BL/6 (48)
Unable to expel low dose
infection.
Able to expel high dose
infection.
AKR (19, 29)
SCID (42)
NUDE (39)
Unable to expel both high
(200 eggs) and low (20 eggs)
dose infections.
Resistance
Th2 Susceptibility
Unable to expel high and low dose infections.
Possible to induce resistance
by vaccination with ES.

21
BALB/c mice susceptible to T. muris infection, while depleting CD8 T cells had
no effect (40). Depleting natural killer T cells also had no effect on the
susceptibility of B10.BR mice to T. muris (40, 41). Similarly, researchers found
that transferring CD4 T cells from wild type BALB/c to SCID mice, which lack
both B and T cells, resulted in worm expulsion (42).
It is well known that different CD4 T helper (Th) cell subsets are associated with
acute and chronic T. muris infection. Naturally susceptible strains such as AKR
mice, and resistant strains such as BALB/c have been key in dissecting these
responses (31, 36). Worm expulsion is brought about by Th2 cells, which
secrete IL-4, 5, 9, 10 and 13, whereas Th1 cells, which secrete IFN-γ, are
associated with chronic infection (31, 36). The interplay between these Th1/Th2
cytokines is emphasised by studies whereby depleting IFN-γ or administering
recombinant IL-4 enabled worm expulsion in AKR mice, while blocking IL-4
function rendered BALB/k mice susceptible to T. muris infection (43). These
studies highlight the influence of host genetics on the relative
resistance/susceptibility to T. muris infection. The effector mechanisms that
bring about worm expulsion in response to Th2 cytokine release will be
discussed in section 1.2.3.
In addition to host genetics, gender is known to influence worm expulsion. For
example, male and female IL-4 deficient mice (BALB/c background) respond
differently to T. muris infection. Females have delayed worm expulsion relative
to wild type mice, whereas males are unable to expel worms, leading to chronic
infection (44). Bancroft and colleagues showed that this gender difference was
due to IL-13, as depleting IL-13 in female IL-4 deficient mice lead to chronic
infection, whereas administering recombinant IL-13 allowed male IL-4 deficient
mice to expel worms (44). Another example of gender differences in T. muris
expulsion is evident in TNF-α (p55/p75) deficient mice. As with IL-4 deficient
mice, female p55/p75 deficient mice (C57BL/6 background) are resistant to
infection, while males are susceptible. This gender difference can be reversed
by neutralising IL-13 in female mice or by IL-13 treatment of males (45). These
studies also emphasise the importance of the Th2 cytokine, IL-13, in driving

22
worm expulsion and suggest that male mice naturally produce less IL-13 than
females (44, 45).
Further studies showed that sex hormones are responsible for the gender
differences in worm expulsion reported for IL-4 deficient mice. Hepworth and
co-workers showed that the androgen dihydrotestosterone is associated with a
reduction in T cell activation by dendritic cells (DCs) and a diminished immune
response in male IL-4 deficient mice (46). This effect was reversed when males
were castrated, enabling mice to expel worms. Lower levels of IL-18 mRNA
were reported in castrated mice, suggesting that androgens may promote a Th1
environment (47). The authors showed that IL-18 neutralisation allowed male
IL-4 deficient mice to expel worms (46) and administering recombinant IL-18 to
C57BL/6 mice resulted in decreased IL-4 and IL-13 production (47). The
female-related hormone 17-β-estradiol has also been shown to enhance Th2
responses in vitro (46).
Infection dose is also known to influence resistance/susceptibility to T. muris
infection. Most laboratory strains, including C57BL/6 and BALB/c mice, are
resistant to a high dose infection (200-400 eggs), typically expelling worms
before they reach patency, however when given a low dose infection (10-40
eggs), these mice are unable expel worms, leading to long-lasting, chronic
infection (48). The exceptions are AKR mice and immuno-compromised strains,
such as SCID mice, which are unable to expel both high and low dose
infections (49, 50). Interestingly, Bancroft and colleagues demonstrated that
BALB/c and C57BL/6 mice are protected from a low dose T. muris infection if it
is preceded by a high dose infection, while mice are susceptible to a high dose
if a low dose is administered first (48). The authors also showed that it is
possible to induce resistance by giving multiple low dose infections (known as a
trickle infection regime) once a critical worm threshold is reached. These data
suggest that antigen dose can affect T helper cell polarisation and ultimately the
outcome of T. muris infection (48).
Similar findings were reported for wild house mice and field mice (51). Behnke
and Wakelin demonstrated that laboratory-bred wild mice expelled high dose

23
T. muris infections, while low dose infections progressed to chronicity (51). They
also sampled a population of wild house mice, and showed that the majority had
low-level infections, while 6 female mice harboured larger worm burdens (51).
The authors suggest that the larger worm burdens in female mice may be
attributed to an altered immune response due to pregnancy and/or
lactation (51). Human population studies also demonstrate that the majority of
individuals have low-level infections (9, 10), and this is reflected in the cytokine
response following re-stimulation of blood leukocytes with T. trichiura antigens,
since IL-4, IL-5 and IL-9 production was restricted to a small proportion (7 to
17%) of the study group, while a larger proportion (32 to 96%) produced IL-10,
IFN-γ and TNF-a (52). These studies demonstrate that the observations made
with laboratory strains of mice reflect natural infection in wild populations (51),
and mirror the infection dynamics described for T. trichiura within human
populations (9, 10).
Different T. muris isolates can also affect the host immune response. Most
inbred mouse strains are able to expel a high dose of the Edinburgh (E) and
Japan (J) isolates, but may be susceptible to the Sobreda (S) isolate (31). The
S isolate appears to abrogate Th2 immunity, by increasing IFN-γ production,
leading to higher titres of anti-parasite serum IgG2a (53, 54). A more recent
study found that mice infected with the S isolate had higher numbers of
regulatory T cells (Tregs) compared to those infected with the E isolate (55).
However, the E isolate has been used in the experiments reported in this thesis.
1.2.3 Mechanisms of T. muris expulsion
The IL-4 and TNF-α knock out studies described in the previous section
emphasised the importance of the Th2 cytokine, IL-13, for T. muris expulsion
(44, 45). IL-13 is known to stimulate a number of effector mechanisms that drive
worm expulsion, including de novo production of the intestinal mucin, Muc5ac,
and increasing the rate of epithelial cell turnover (56, 57). Another important
effector mechanism is intestinal muscle hyper-contractility, which is brought

24
about by the Th2 cytokine, IL-9 (58). This cytokine is also responsible for the
mastocytosis and IgE production observed during gastrointestinal nematode
infections, while IL-5 and CCL11 induce eosinophilia (59-62). Although the
intestinal influx of these cells is considered a hallmark of gastrointestinal
nematode infections, these responses appear to be dispensable for T. muris
worm expulsion (59-62). This section will examine the effector mechanisms
induced by Th2 cytokines during T. muris infection and will discuss the relative
contribution of these to worm expulsion. These are summarised in Figure 1.3.

25
Figure 1.3. Immune response during acute T. muris infection. Damage caused by T. muris burrowing into the host epithelium may stimulate release of alarmins, such as IL-25, IL-33 and TSLP. ILC2s release Th2 cytokines in response to these alarmins, along with basophils, eosinophils and NK cells. Antigen presenting cells (APCs) such as DCs phagocytose antigens (such as ES components). Antigens are processed and presented to naïve T cells on MHC class II (MHCII). T cell activation occurs if the T cell receptor recognises antigen presented on MHCII, and T cells become polarised towards the Th2 phenotype due to high levels of IL-4 in the local environment. IL-9 and IL-13 trigger effector mechanisms (purple boxes), leading to worm expulsion. In addition, activated T cells can provide a second signal for activation of B cells. A proportion of B cells become antibody-secreting plasma cells. Antibody class switching (towards IgG1) occurs in response to Th2 cytokines. Anti-parasite IgG1 antibodies recognise native antigens in ES (top of figure).
IL-33
TSLP
IL-25
ILC2
IL-4
IL-9 IL-13
Baso IL-4
NK cells
IL-13
Thn
IL-4
IL-4
IL-4 IL-4
IL-4
IL-4
Th2
IL-13
IL-4
IL-9
IL-4
IL-13
IL-9
Intestinal hypercontractility
Mucus production
Epithelial escalator
B cell
IgM
IgG1
Eosino
IL-13
IL-4
IL-9
IL-5
IL-5
Early sources of Th2 cytokines
Damage caused by worm burrowing into epithelium may
stimulate release of IL-25, IL-33 and TSLP
Uptake and processing of
antigen by APCs
Antigen presentation and
activation of T cells
Polarisation towards Th2
phenotype due to IL-4 in local
environment Activation of B cells by T cells
A proportion of B cells become
antibody-secreting plasma cells
Antibody class switching and
production of anti-parasite IgG1
in response to Th2 cytokine
environment
B cell
IgG1
IgG1
IgG1
APC
RELM-β
Setd7
Mastocytosis, IgE
IL-5
Eosinophilia

26
1.2.3.1 Mucus production and other goblet cell secretions
The intestinal mucus barrier is the first line of defense against gastrointestinal
nematodes (36). Mucins are the main components of mucus and are
responsible for its gel-like properties. Muc2 is the major intestinal mucin, and is
produced by goblet cells (36). Goblet cell hyperplasia has been reported during
T. muris, Nippostrongylus brasiliensis, Heligosomoides polygyrus and
Trichinella spiralis infections (63-66). The importance of intestinal mucus for
worm expulsion is clear from Muc2 deficient mice, which lack an effective
mucus barrier and exhibit delayed T. muris expulsion compared to their wild
type counterparts (67). During acute T. muris infection, the mucus layer
thickens and production of Muc2 as well as the transmembrane mucins, Muc4
and Muc13 is increased (68). Resistant strains of mice (BALB/c and C57BL/6)
also produce Muc5ac in the intestinal tract in response to T. muris infection.
Muc5ac expression is normally restricted to the eyes, lungs and stomach and is
absent from the intestine during homeostasis (56). Hasnain and colleagues
demonstrated that Muc5ac deficient mice (C57BL/6 background) are unable to
expel T. muris, despite generating a strong Th2 response, indicating that
intestinal Muc5ac production is critical for worm expulsion (56).
Hasnain and co-workers demonstrated that intestinal Muc5ac production during
T. muris infection was dependent on the cytokine IL-13 (Figure 1.3). The
authors showed that IL-4 deficient mice (BALB/c background) produce Muc5ac
in response to a high dose infection, enabling them to expel worms, whereas
(IL-4R) deficient mice, which are unable to respond to both IL-4 and IL-13, fail to
produce Muc5ac, and infection progresses to chronicity (56). The authors also
used an ATP assay to measure the viability of worms following incubation with
human MUC2 and MUC5AC produced by HT29 and LS174T cells in vitro. They
reported a dose-dependent decrease in worm viability following incubation with
MUC5AC, while MUC2 had no effect on worm viability. This suggests that
MUC5AC/Muc5ac is a pharmacological agent that directly damages
worms (56). De novo Muc5ac synthesis was also observed in wild type
C57BL/6 mice following infection with N. brasiliensis and T. spiralis, while

27
Muc5ac deficient mice showed delayed expulsion of these parasites. However,
the authors could not detect intestinal Muc5ac in C57BL/6 mice given a primary
infection of H. polygyrus, a parasite that persists for months in most strains of
mice (56). These studies suggest that Muc5ac plays a crucial role in the
resolution of gastrointestinal nematodes infections (56).
Muc5ac also appears to contribute to the network properties and viscosity of the
intestinal mucus. Hasnain and colleagues reported that the intestinal mucus of
Muc5ac deficient mice is more porous than that of wild type mice, and proposed
that the increased viscosity may be important for retaining anti-parasitic
molecules, such as serine proteases and resistin-like molecule beta (RELM-β),
which may also contribute to worm damage (68). The authors also found that
during acute infection, a large proportion of goblet cell-associated mucins were
sulphated, whereas during chronic infection the majority were sialylated (69).
Hasnain and colleagues demonstrated that mucin sulphation is driven by IL-13,
and that sulphate anion transporter-1 (Sat-1) deficient mice (C57BL/6
background) had significantly fewer sulphated goblet cell-associated mucins.
The authors found that high dose infections progressed to chronicity in Sat-1
deficient mice, despite a strong Th2 response (69). These studies demonstrate
an important role for mucins in influencing mucus barrier properties and driving
worm expulsion.
IL-13 also induces RELM-β secretion by goblet cells during T. muris infection.
As with Muc5ac, Artis and co-workers demonstrated that IL-4 deficient mice
produce RELM-β during acute infection, whereas IL-4R deficient mice do
not (63). The authors demonstrated that RELM-β binds to the bacillary band of
T. muris, specifically binding to pore structures that may contain chemosensory
apparatus (63). The authors also demonstrate that RELM-β impairs
chemoattractant-driven migration of Strongyloides stercoralis L3 larvae in vitro.
Taken together, this suggests that RELM-β may impair the chemosensory
functions of T. muris such that the worm is less equipped to navigate the
intestinal environment, which may in turn contribute to worm expulsion (63).
However, Nair and co-workers demonstrated that T. muris is expelled from

28
RELM-β deficient mice and reported an alternative role for RELM-β in
promoting Th1 immunity during infection through activating inflammatory
macrophages (70, 71). These experiments suggest a less important (or even a
negative) role for RELM-β in T. muris expulsion (70, 71). Herbert and
colleagues found that RELM-β inhibited the feeding of H. polygyrus on host
tissue in vivo, and that this molecule plays an important role in H. polygyrus and
N. brasiliensis expulsion, but had no effect on T. spiralis viability (71). These
studies suggest that RELM-β may contribute to the expulsion of luminal, but not
tissue-dwelling, gastrointestinal nematodes.
1.2.3.2 Increased rate of epithelial cell turnover
The rate of epithelial cell turnover in the caecum is also increased in response
to IL-13 during acute T. muris infection (Figure 1.3) (57). For BALB/c mice, the
rate of turnover peaks at day 14 post infection (p.i.), which coincides with
parasite expulsion, whereas the turnover rate in AKR mice, where infection
progresses to chronicity, is half that of BALB/c mice. Cliffe and colleagues
showed that the epithelial turnover rate for IL-13 deficient mice (BALB/c
background) was lower than for wild type BALB/c mice, suggesting that IL-13 is
responsible for the increased turnover (57). They found that the chemokine
CXCL10 has a negative affect on the rate of turnover in AKR mice, and that
blocking CXCL10 with antibodies allows AKR mice to expel worms effectively
through an increase in cell turnover (57). This suggests that during acute
infection, IL-13 counteracts the regulatory effect of CXCL10 on epithelial cell
turnover, which contributes to worm expulsion (57). Cliffe and colleagues also
showed that blocking CXCL10 significantly increased the rate of turnover in
SCID mice, enabling worm expulsion despite a lack of T cells and lower levels
of IL-13. The authors propose a model whereby increasing the rate of turnover
acts as an ‘epithelial escalator’, moving worm-embedded epithelial cells from
the crypts towards the lumen, where the cells are shed and worms are then
expelled from the body (57).

29
Oudhoff and colleagues recently reported that Setd7 deficient mice (C57BL/6
background), which lack the lysine methlytransferase, Setd7, have increased
resistance to T. muris (72). The authors found that Sedt7 deficient mice were
able to expel a high dose T. muris infection at an earlier time point compared to
their wild type counterparts, with 75% fewer worms at day 14 p.i. compared to
wild type mice. They provide evidence to suggest that Sedt7 controls intestinal
epithelial cell turnover through methylation of members of the Hippo and Wnt
signalling pathways, and that the rate of turnover is increased in Setd7 deficient
mice, enabling rapid worm expulsion (72). The authors showed that worm
expulsion in Setd7 deficient mice was independent of adaptive immunity, as
Setd7/Rag-1-/- mice were able to expel worms despite a lack of B and T cells
(albeit not to the same degree as Setd7-/- mice). Oudhoff and co-workers found
that specifically deleting Setd7 in intestinal epithelial cells rendered mice more
resistant to high and low dose T. muris infections, but had no effect on
H. polygyrus expulsion (72). These experiments are strongly supportive of
epithelial cell turnover as an important effector mechanism for expulsion of gut
epithelial-dwelling nematodes and suggest that manipulation of the Sedt7
pathway can increase resistance to Trichuris parasites (72).
1.2.3.3 Intestinal muscle hyper-contractility
Another effector mechanism that contributes to T. muris expulsion is intestinal
hyper-contractility, mediated by intestinal smooth muscle cells (Figure 1.3) (58).
Khan and co-workers demonstrated that treating C57BL/6 mice with IL-9 during
the early stages of infection (day 7 or 14 p.i.) increased intestinal contractility
and promoted earlier expulsion of both T. spiralis and T. muris (58). The authors
also showed that stimulating splenocytes from T. spiralis infected animals with
IL-9 led to increased IL-4 and IL-13 production in vitro, and that IL-9 increased
mucosal mast cell protease-1 levels and goblet cell hyperplasia in T. spiralis
infected animals (58). Furthermore, they showed that blocking IL-9 function,
either by treating mice with anti-IL-9 antibodies or by vaccinating against IL-9,
significantly impaired muscle contractility and nematode expulsion in T. muris

30
infected mice, but had no significant effect on T. spiralis infected mice. T. muris
and T. spiralis reside in different niches within the intestine (caecum and upper
small intestine respectively), which might explain these differences (58).
1.2.3.4 Mast cells and IgE production
IL-9 is also responsible for the mastocytosis and IgE response that accompany
gastrointestinal nematode infections, however these do not appear to play
major roles in T. muris worm expulsion (62). Lee and colleagues showed that
NIH mice (a resistant strain) expel worms ten days before mastocytosis
develops (73), while Betts and co-workers demonstrated that blocking mast cell
development with anti-c-kit antibodies (c-kit is the major transcription factor
required for mast cell development) did not impair worm expulsion (59).
Koyama and colleagues also concluded that mucosal mast cells are not
required for T. muris expulsion from studies with mast cell deficient W/Wv
mice (74).
1.2.3.5 IgG antibody production and B cells
In addition to IgE, acute T. muris infections are associated with anti-parasite
IgG1 class switching, while IFN-γ promotes IgG2a production, particularly
during chronic infection (75). Early studies suggested a role for antibodies in
generating immunity to T. muris (76, 77), however a more recent study has
suggested a role for B cells in Th2 cytokine production (78). Blackwell and
colleagues described a susceptible phenotype for B cell deficient μMT mice
(C57BL/6 background), reporting that MLN-derived lymphocytes harvested from
these mice produced very low levels of Th2 cytokines in response to antigen
re-stimulation (78). The authors demonstrated that administration of anti-IL-12
neutralising antibodies enabled μMT mice to expel worms, suggesting that the
susceptible phenotype of these mice is a result of an inability to block a Th1
immune response, presumably through diminished Th2 cytokine release (78).

31
The authors show that transferring anti-parasite IgG1 antibodies from naturally
resistant NIH mice enabled μMT mice to expel worms (78). However, given that
resistance can be induced in SCID mice (deficient in B and T cells) by
transferring CD4 T cells alone, it would seem that B cells and antibodies play a
minor role in generating immunity to T. muris (50).
1.2.3.6 Innate lymphoid cells and other early sources of Th2 cytokines
Recently there has been great interest within the field of immunology in
understanding the molecular events that bridge innate and adaptive immunity,
particularly concerning the cell populations that are responsible for the initial
increase in cytokines that drive T cell polarisation (36). Innate lymphoid cells
(ILCs) are likely to be important in this process. ILCs are divided into three
groups: group 1 includes type 1 ILCs and natural killer cells, which secrete
IFN-γ, group two refers to type 2 ILCs (also known as ILC2s), which secrete
Th2 cytokines, and group 3 includes Rorγt+ type 3 ILCs and lymphoid tissue
inducers, which both secrete IL-17 and IL-22 (79). ILC2s are the most relevant
in terms of gastrointestinal nematode expulsion as these secrete Th2 cytokines
(particularly IL-5 and IL-13) in response to epithelial derived IL-33, IL-25 and
thymic stromal lymphopoietin (TSLP) (36). Basophils and NK cells are also
thought to be an early source of Th2 cytokines during gastrointestinal nematode
infection (80-83).
IL-33, IL-25 and TSLP are all upregulated in the intestine during early
gastrointestinal nematode infection (84). IL-33 and IL-25 behave similarly during
gastrointestinal infection – mice deficient in either cytokine have impaired
(85-87) or delayed (88, 89) worm expulsion, and treating susceptible AKR mice
with recombinant IL-33 or IL-25 during early infection enhances Th2 immunity
and worm expulsion (84, 87). However, neither cytokine could prevent a chronic
T. muris infection developing in SCID mice, suggesting that these cytokines are
dependent on adaptive immunity to exert their anti-nematode functions (84, 87).
IL-25 is known to induce multi-potent progenitor type 2 cells, which secrete Th2

32
cytokines during early infection (90), while TSLP appears to neutralise the
effects of IFN-γ and IL-12/23, which indirectly enhances Th2 immunity (91).
1.2.3.7 Regulation of the immune response during T. muris infection
Chronic T. muris infection can lead to severe intestinal inflammation, resulting in
changes to gut architecture and physiology similar to those observed during
inflammatory bowel disease (92). IL-10, TGF- and Tregs are thought to
regulate the immune response in order to minimise intestinal damage (31). The
importance of IL-10 in regulating inflammation during T. muris infection is
emphasised by the severe pathology seen in IL-10 deficient mice (C57BL/6
background), with mice eventually succumbing to infection (93). An IL-35-
dependent subset of Tregs, iTR35, has also been implicated in regulating
inflammation during T. muris infection (94). These cells lack the typical Treg
transcription factor, Foxp3, and are thought to suppress intestinal inflammation
through IL-35 secretion in the absence of IL-10 and TGF- (94).
There is also some evidence to suggest that TGF- regulates the development
of Th2 immunity. Veldhoen and colleagues show that TGF- can re-program
Th2 cells so that they no longer express GATA-3 (the transcription factor
associated with Th2 cells) or express the signature Th2 cytokines IL-4, IL-5 and
IL-13, but instead secrete high levels of IL-9 (95). This re-programming fails to
occur in CD4dnTGFβRII mice (C57BL/6 background), which have a truncated
form of the TGF- receptor II, rendering them unresponsive to this cytokine.
These mice have delayed worm expulsion, suggesting that TGF- mediated
re-programming is involved in the resolution of infection (95). In contrast,
Worthington and colleagues showed that TGF- signalling interferes with the
development of Th2 immunity during the early stages of a low dose infection,
leading to production of an inappropriate Th1 response and chronic
infection (96). These studies show that the role of IL-10, TGF- and Tregs
during T. muris infection is complex, however, these factors do not appear to
play a major role in worm expulsion.

33
1.3 Clinical and pre-clinical helminth vaccine candidates
As outlined at the start of this thesis, a combination of better sanitation,
anthelminthic drugs and prophylactic vaccines is predicted to reduce the
morbidity caused by STHs in humans (22). This section will review the progress
made towards identifying vaccine candidates for medically important helminths,
including N. americanus, A. lumbricoides, T. trichiura and Schistosoma species.
Highly effective recombinant vaccines have been developed for some cestode
species, including Taenia solium, a porcine cestode that causes
neurocysticercosis in humans (97, 98).
1.3.1 Hookworm vaccine candidates
The identification and development of vaccine candidates for N. americanus,
which is thought to cause the majority of hookworm infections worldwide, is led
by the human hookworm vaccine initiative (HHVI). This group was set up with
the aim of identifying vaccine candidates from both the L3 larval and adult stage
of the parasite (99, 100). The rationale for targeting the L3 larval stage is based
on the success of a radiation-attenuated L3 larval vaccine for the canine
hookworm, A. caninum in the 1970s (100). The first vaccine candidate to reach
clinical trials was N. americanus Activation Secreted Protein-2 (Na-ASP-2),
which is one of the most abundant antigens secreted by L3 larvae (101).
Recombinant Na-ASP-2 was expressed by yeast and insect cells, and proof-of-
principal animal studies showed that vaccinating hamsters and dogs with the
recombinant protein prior to hookworm infection resulted in reduced worm
burdens (101, 102). There were no noticeable side effects when a cohort of
young healthy adults was vaccinated with recombinant Na-ASP-2 in the United
States, however when humans were vaccinated in endemic areas in Brazil,
three of the seven adults developed urticaria, an allergic reaction in the skin,
within two hours of vaccination (103). This appeared to be due to high levels of
pre-existing anti-Na-ASP-2 IgE antibodies resulting from prior infection, which
led to an immediate hypersensitivity type reaction. The adverse reaction to the

34
vaccine led to the clinical trial being terminated and the antigen was
abandoned (103).
An alternative recombinant vaccine is now being developed, consisting of two
proteins, N. americanus aspartic protease-1 (Na-APR-1) and N. americanus
glutathione-S-transferase-1 (Na-GST-1), formulated with an aluminum
hydroxide adjuvant (100). The candidates are being tested separately in clinical
trials, with the ultimate aim of administering these proteins together as a single
vaccine (100). Both candidates were selected based on their putative role in the
breakdown of host haemoglobin by adult stages in the gut to provide the
parasite with iron. Na-APR-1 is a 24 kDa aspartic protease that digests
haemoglobin, whereas Na-GST-1 is a 45 kDa glutathione-S-transferase that
detoxifies haem (100). These candidate proteins are being expressed in yeast
and tobacco plants respectively, however, for safety reasons the Na-APR-1 has
been modified so that its proteolytic activity is removed (100, 104-106).
Vaccination of laboratory animals with the modified recombinant Na-APR-1 has
been shown to protect against subsequent infection by inducing neutralising
antibodies that target the native hookworm enzyme, meaning that the parasite
is unable to digest host haemoglobin (106). This also led to a reduction in iron
loss during infection (106). Similar results have been reported for
Na-GST-1 (107).
An effective hookworm vaccine should reduce the number of gut-dwelling
parasites such that symptoms (namely anaemia) are reduced, however the
HHVI team argue that this may be possible without achieving sterilising
immunity (99, 100). These pre-clinical results are therefore very encouraging,
and the two recombinant proteins are now being tested in combination in a
phase II clinical trial in Gabon, having successfully completed phase I trials in
the US and Brazil (107). Clinical studies are also being carried out to assess
whether using an additional adjuvant, such as the synthetic Toll-like receptor
(TLR) 4 agonist, glucopyranosyl, or the TLR9 agonist, CpG
oligodeoxynucleotide, can induce better protection (107).

35
1.3.2 Pre-clinical Ascaris vaccine candidates
The porcine roundworm, A. suum, is often used as a model for A. lumbricoides.
These species are antigenically identical, and the life cycle of these parasites
involves the migration of larvae from the small intestine to the lungs via the
portal and systemic circulation. A. suum can complete this part of its life cycle in
rodents such as mice and rabbits, which are often used as experimental models
of A. suum infection in place of pigs (22). A number of Ascaris vaccine
candidates have been identified by antibody-based screening methods using
sera from immunised rabbits or pigs. Some of these candidates have been
tested in Ascaris challenge models, including As14 and As16, which are 14 and
16 kDa-sized proteins found in Ascaris ES material and worm homogenate.
Vaccinating mice intranasally with recombinant forms of these proteins
formulated with cholera toxin B subunit resulted in a 60% reduction in migrating
larvae (108, 109). Two other candidates of unknown function, As24 and As37
were also identified by antibody-based screening methods, and vaccinating
mice with recombinant forms of these proteins formulated with Freund’s
complete adjuvant (CFA) reduced migrating larvae by 58 and 69% respectively
(22, 110-112). Finally, a DNA vaccine coding for a glycolytic enzyme,
As-Enol-1, has been demonstrated to reduce the lung larval burden by 61% in
vaccinated mice (113, 114).
1.3.3 Experimental Trichuris vaccines
In the 1980s, Jenkins and colleagues demonstrated that the ES material
released by the mouse whipworm, T. muris, could stimulate protective immunity
in mice (28). They showed that vaccinating NIH mice subcutaneously with
100 μg of ES formulated with either aluminum hydroxide or CFA promoted
earlier expulsion of a high dose infection (around 70-75% reduction in worm
burden at day 9 p.i. compared to sham vaccinated mice) (28). The authors also
demonstrated that subcutaneous vaccination was more effective than peritoneal
vaccination – mean worm burden was reduced by 70% compared to sham

36
vaccinated controls when mice were vaccinated subcutaneously with 100 μg of
ES (formulated without adjuvant), compared to 33% for intraperitoneal
vaccination. Jenkins and others have shown that subcutaneous vaccination with
worm homogenate (formulated with CFA) induced similar levels of protection as
ES, and that oral vaccination with worm homogenate formulated with cholera
toxin could also induce protective immunity in BALB/c and C57BL/6
mice (115, 116).
Jenkins and colleagues demonstrated that ES can be fractionated into a
number of smaller sub-groups using ammonium sulphate precipitation and
isoelectric focusing. They vaccinated NIH mice with the resulting sub-fractions,
and assessed worm burdens at day 9 p.i., to determine which fractions
contained protective material. They found that vaccinating mice with material
precipitated with 85-100% ammonium sulphate or at pH 6.9 to 7.3 lead to a
significant reduction in worm burden compared to sham vaccinated mice at day
9 p.i. (28). Similar approaches were used in this thesis to fractionate ES into a
smaller sub-fractions based on the size and charge of its components. The
immunogenicity of these sub-fractions has been tested using in vitro assays and
vaccination experiments, and mass spectrometry has enabled the identification
of proteins within the immunogenic sub-fractions.
More recent studies show that subcutaneous vaccination of AKR mice with
100 μg of T. muris ES formulated with either aluminum hydroxide or incomplete
Freund’s adjuvant (IFA) induced expulsion of a high dose infection in this
naturally susceptible strain (19, 29). Dixon and colleagues demonstrated that
subcutaneous vaccination of AKR mice with ES formulated with CFA increased
Th2 cytokine production by MLN lymphocytes following infection, compared to
sham vaccinated controls (29). The authors also report increased goblet cell
hyperplasia and an influx of alternatively activated macrophages, offering clues
about the protective immune response following vaccination (29).

37
Sequencing of the T. trichiura and T. muris transcriptomes identified a number
of functional groups that are upregulated in the anterior end of the worm, which
is postulated to be the site of ES release (33). These include proteases,
particularly chymotrypsin-like serine proteases, protease inhibitors/WAP domain
containing proteins, and DNases. As such, these proteins may be of interest to
vaccine research. T. muris has 75 genes encoding chymotrypsin-like serine
proteases, which is more than the other nematodes studied to date (33).
Three-quarters of these are upregulated in the anterior portion of the worm and
two-thirds are secreted (33). Hasnain and colleagues have demonstrated that
serine proteases released in T. muris ES are able to degrade Muc2, the major
component of the mucus barrier, which may alter the barrier properties, allowing
the worms to persist to chronic infection (117). Foth and colleagues suggest
that by altering the environment in such a way, Trichuris proteases may be
targeted by the host immune system, as part of the evolutionary arms race
between pathogen and host (33).
Foth and colleagues also report that 72% of the 111 T. muris protease inhibitors
are serine protease inhibitors (serpins) and many of these are WAP domain-
containing proteins (33). Mammalian secretory leukocyte protease inhibitor
protein (SLPI) is a WAP protein and has several additional functions, including
anti-inflammatory, anti-microbial and immunomodulatory roles, as well as roles
in innate immunity and wound healing (33). As such, Foth and colleagues
suggest that the T. muris WAP proteins could have similar additional
functions (33). The archetypal WAP domain is composed of eight cysteine
residues arranged in a 4-disulphide core, however only one of the T. muris
WAP proteins contains this typical structure and this is also true for
T. trichiura (33). Interestingly, this protein shares some homology to
Caenorhabditis elegans mesocentin, a protein that gives an RNA interference
phenotype (33), suggesting that this Trichuris protein could perhaps play an
immunomodulatory role. The remaining 19 T. muris WAP proteins have a novel
structure consisting of a six cysteine core, which raises the possibility that these
proteins carry out Trichuris specific functions, and as such may be key targets
for the host immune system (11).

38
Transcripts for several DNases were upregulated at the anterior end of T. muris,
however, aside from one exception, these share little homology with
mammalian DNases and are only distantly related (33). This was also the case
for T. trichiura and the closely related tricephalid parasite, Trichinella spiralis,
which suggests that these DNases could carry out nematode-specific
functions (33). An interesting suggestion put forward by Foth and colleagues is
that these DNases degrade host DNA which is released when the epithelium is
damaged as a result of worm burrowing, a process which would normally
stimulate an inflammatory response (33). It is of course beneficial for Trichuris
species to minimise inflammation and immunopathology to allow prolonged
infection (118). These DNases also warrant further investigation as vaccine
candidates due to their possible role in promoting chronicity.
Gomez-Samblas and colleagues have recently identified a vaccine candidate
with immunoprotective properties against a number of helminth parasites,
including T. muris (30). The candidate is a recombinant serine-threonine
phosphatase 2 enzyme from the nematode A. costaricensis, and is linked to a
synthetic self-adjuvant oleic-vinyl sulphone group. The authors showed that
vaccinating AKR mice with this protein induced expulsion of established
T. muris infection. The group also showed that egg output was reduced as early
as 7 days following vaccination (30). The ability of this recombinant subunit
vaccine to drive expulsion of a chronic infection is interesting, as it would
suggest that the vaccine is potent enough to reverse an ongoing Th1 response
(30). This is an important consideration for vaccinating people in endemic
areas, as they are likely to be infected from an early age and could be infected
prior to vaccination (2). This candidate protein has also been shown to induce
protective immunity against ovine helminth species, namely Haemonchus
contortus and Teladorsagia circumcincta (119).

39
1.3.4 Schistosome vaccines
There are several schistosome vaccine candidates in clinical or pre-clinical
trials. Although schistosomes are trematodes (flatworms) rather than
nematodes (roundworms), these vaccine candidates could still offer some
useful insights for helminth vaccine design. The most advanced of these
vaccine candidates is the S. haematobium glutathione-S-transferase,
Sh28GST, which was deemed safe and effective in Phase I clinical trials (23).
The other vaccine candidates target S. mansoni, the major cause of intestinal
schistosomiasis. These candidates were identified using a protein array
containing S. mansoni and S. japonicum proteins (27). IgG antibody responses
to these proteins were measured for individuals with drug induced resistance
(DIR) and rhesus macaques, which, unlike humans, are naturally resistant to
schistosome infections (27). The term DIR refers to individuals who develop
resistance to schistosome infection following praziquantel treatment (27).
Praziquantel destroys flukes, which exposes the host immune system to a
range of schistosome proteins that it may not normally come into contact with,
and some individuals develop at least some level of resistance to subsequent
schistosome infections (termed DIR). This resistance can last a year or more,
and appears to correlate with anti-parasite IgG levels (27). Rhesus macaques
naturally expel schistosome infections once the parasite reaches patency if
worm burdens reach a certain threshold, and this expulsion mechanism is also
thought to be antibody-mediated (27).
Pearson and co-workers have taken advantage of these phenomena to guide
candidate selection (27). Serum IgG antibodies from DIR individuals and
macaques recognised several proteins, including Smp80 (calpain), tetraspanins
(such as Sm-TSP-1 and 2), glutathione-S-transferases, and glucose
transporters (27). Out of these vaccine candidates, Sm-TSP-2 is currently
undergoing Phase I clinical trials in the USA (23), while Sm80, Sm-TSP-1 and
Sm29, all S. mansoni tegument proteins, are being evaluated in pre-clinical
studies (23-26). The latter has been tested in non-human primates, and the
authors showed that three vaccinations resulted in a 10-36% decrease in
parasite burdens compared to sham vaccinated controls (26).

40
Another S. mansoni protein, the fatty acid binding protein, Sm14 was declared
safe and effective in Phase I clinical trials in Brazil (120). Recombinant Sm14
was expressed in yeast, and healthy volunteers were vaccinated
intramuscularly with 50 μg of protein plus a glucopyranosyl lipid adjuvant a total
of three times. Vaccination induced anti-Sm14 IgG antibody production and a
Th1 polarised CD4 T cell response (120). Interestingly, cross-reactivity between
Sm14 and the Fasciola hepatica (an ovine liver fluke) F15 protein has been
reported (121). Recombinant Sm14 was shown to induce protective immunity
against F. hepatica in a murine model, as well as the natural host,
sheep (121, 122). Other parasites of great medical and agricultural importance
have been shown to express fatty acid binding proteins with homology to Sm14,
including the canine tapeworm, E. granulosus, and the Chinese liver fluke,
Clonorchis sinensis, suggesting that this vaccine candidate could be effective
against several parasites of clinical and veterinary importance (123, 124).
1.3.5 Cestode vaccine candidates
Highly effective recombinant vaccines have been developed for a number of
cestodes, including T. ovis, T. solium, T. saginata and E. granulosus (98).
Antibody-based screening methods were employed to identify immunogenic
candidates within the ES collected from Taenia oncospheres (the infective larval
stage found within Taenia eggs) (92). The rationale for this approach was based
on studies carried out in the 1930s, showing that vaccination with Taenia
oncospheres induces protective immunity in a range of veterinary species, and
the transfer of serum or colostrum from infected or vaccinated animals can
protect naïve individuals from a subsequent infection (125-127). Several
immunogenic candidates were identified by combining SDS-PAGE fractionation
of oncosphere ES and immunoblot analysis, including To16, To18 and
To45W (128, 129).
Previous studies also showed that there was significant cross-reactivity
between the oncosphere antigens of different Taenia species (98). This was

41
reflected in vaccination experiments whereby antigens from one taeniid species
were able to induce protective immunity against heterologous species, and
passive transfer of immunity was achieved using immune sera raised against
heterologous species (125-127). Homologues of To16, To18 and To45W were
identified in T. solium (porcine tapeworm) and T. saginata (bovine tapeworm)
oncospheres. Vaccination with recombinant Tsol18, the T. solium homologue of
To18, was highly effective at protecting pigs against a subsequent T. solium
infection (close to 100% protection), in both experimental settings and field
studies (130, 131). However, unlike T. solium, vaccination with TSA-18, the
T. saginata homologue of To18, was not sufficient to protect cattle against a
subsequent T. saginata infection. However, vaccination with TSA-18 in
combination with TSA-9 (the T. saginata homologue of To45W), resulted in 94
to 99% protection (132). These studies demonstrate that is possible to achieve
close to 100% protection against a metazoan parasite by vaccinating with one
or two recombinant antigens.
Antibody screening and fractionation methods were also used to identify a
suitable vaccine candidate for E. granulosus, a 25 kDa protein called
EG95 (98). Experimental vaccine studies showed that vaccinating sheep with
EG95 resulted in 96-100% protection against E. granulosus (133, 134). A
homologue of EG95 has been identified in E. multicularis, suggesting that it may
be possible to develop an effective vaccine for this parasite, as well as other
zoonotic Echinococcus species, as has been possible for Taenia species (98).
1.4 The role of adjuvants in vaccines
Adjuvants are used to enhance the magnitude and breadth of
vaccinations (135). Often, adjuvant usage means that less material is required
in order to stimulate protective immunity (this phenomena is known as antigen
sparing) and fewer vaccinations may be required to achieve an appropriate
level of protection (135). A range of adjuvants are available, and these can be
broadly divided into three classes: emulsions, immunostimulatory complexes
(ISCOMs), and mineral salts (135). Freund’s adjuvants (CFA and IFA) are

42
amongst the most widely used adjuvants in experimental vaccines (135). These
fall under the emulsions category and are very effective at inducing both B and
T cell responses (135). However, CFA and IFA are not used in a clinical setting
due to safety reasons (135). Since the advent of Freund’s in 1930s, safer
alternatives have been developed, namely water-in-oil adjuvants such as
Montanide ISA 51 and ISA 720, which have been licenced for use in humans
(135). These adjuvants have been used with great success in pre-clinical and
phase I clinical trials for vaccines against malaria, Epstein-Barr virus, hepatitis C
virus and simian immunodeficiency virus (SIV) (136-141). Montanide ISA 720
was also used in pre-clinical studies with the schistosome vaccine candidate,
cathepsin B, where vaccination reduced worm burden by 60% compared to
sham vaccinated control mice (142). Other emulsion adjuvants include MF59,
which is an oil-in-water adjuvant (like Freund’s) licenced for use in humans
(135). This adjuvant forms part of a licenced influenza vaccine, and has been
used in clinical trials for hepatitis B virus, herpes simplex virus and HIV (135).
Aluminum hydroxide is the most common mineral salt adjuvant (135). Like the
majority of adjuvants, the exact mechanism by which aluminum hydroxide
exerts its adjuvanticity is unknown (135). It was proposed that aluminum
hydroxide acts as an antigen ‘depot’, allowing slow release of antigen over time
(143). However, Hutchinson and colleagues showed that removal of the antigen
‘depot’ 2 hours after vaccination had no appreciable effect on antigen-specific T
and B cell activation (144). Another study suggests that the damage caused by
aluminum hydroxide causes cell death, leading to release of host DNA, which
can act as a damage associated molecular pattern (DAMP) and drive immune
cell activation through various Toll-like receptors (145). There is also some
evidence that activation of the NLRP3 inflammasome by aluminum hydroxide
mediates adjuvanticity (146-148).
Lastly, ISCOM adjuvants, such as QuilA, which is composed of a mixture of
saponins from Quillaia saponaria, encapsulate antigens in 40 nm cages (135).
These adjuvants stimulate potent CD8 T cell responses in mice and
humans (149-151).

43
1.5 Extracellular vesicles as a source of antigenic material
Helminth vaccination studies have traditionally focussed on identifying antigenic
proteins within parasite secretions or homogenates. However, there is
increasing recognition of vesicular components within parasite secretions, and
these have been somewhat overlooked in terms of identifying host protective
components (152-154). Two recently published papers describe a protective
role for parasite-derived extracellular vesicle (EV) vaccinations. Coakley and
colleagues report a significant reduction in H. polygyrus worm burden and egg
output when mice were vaccinated intraperitoneally with H. polygyrus exosome-
like vesicles (ELVs) prior to infection, while Trelis and co-workers showed that
subcutaneous vaccination of mice with Echinostoma caproni ELVs prior to
infection lead to significant improvement of clinical symptoms and reduced
mortality (155, 156). A sub-objective of this thesis was to investigate the
potential of T. muris vesicular components as a source of protective antigens,
and as such the following sections will provide an overview of exosome
biogenesis, and will discuss the potential for helminth-derived EVs as vaccine
candidates, immunomodulators and mediators of host-parasite communication.
1.5.1 Exosome biogenesis and isolation from biological samples
The three key types of EVs are exosomes, microvesicles and apoptotic bodies.
These vesicles are mostly spherical in shape and are composed of a
phospholipid bilayer with a protein/RNA core (157). Exosomes are the smallest
type of EVs, ranging from 30 to 100 nm in size, and are formed via the
endosomal pathway (157). Exosomes have been shown to play an important
role in the transfer of proteins, messenger RNAs (mRNAs) and micro RNAs
(miRNAs) between cells (158-161). This transfer can alter the behaviour of
recipient cells, which may be of particular importance in the context of host-
parasite interactions, where parasite exosomes have been shown to stimulate
and/or modulate host immunity through ELV release (162-165). The first step in
exosome biogenesis is the invagination of the plasma membrane to form

44
endosomes. The second step is the inward budding of the late endosomal
membrane to form intraluminal vesicles (ILVs) inside a multivesicular body
(MVB). These ILVs are released upon fusion with the plasma membrane, and
once extracellular, they are referred to as exosomes (Figure 1.4) (132).
Figure 1.4. Biogenesis of exosomes. Exosomes are formed by the invagination of the endosomal membrane to form intraluminal vesicles (ILVs, yellow) inside a multivesicular body (MVB). These vesicles are released following fusion of the MVB with the plasma membrane. Once released into the extracellular environment, vesicles are referred to as exosomes. PM is plasma membrane.
Isolation of microvesicles and exosomes from biological samples is often
carried out using differential ultracentrifugation. This involves centrifuging the
material at progressively higher speeds, starting at around 200 g to pellet cells,
followed by 10,000 g to pellet microvesicles and 100,000 g to pellet
exosomes (166). This is considered the ‘gold standard’ for exosome isolation,
however other methods include using density gradient centrifugation, size
exclusion chromatography or commercially available kits (157, 163, 166).
Isolation of exosomes and microvesicles is confirmed by electron microscopy,
and where possible, by the presence of certain markers. Typical exosome
markers include Alix, and TSG101, which are both involved in MVB biogenesis,

45
and tetraspanins such as CD9, CD63 and CD81, while CD40 is a key marker of
microvesicles (157, 167).
1.5.2 Exosome release by parasitic helminths
Helminths are parasitic worms belonging to several phyla, including nematodes
(roundworms), trematodes (flukes) and platyhelminths (flatworms) (168). There
are examples in the literature of members of all three of these phyla secreting
ELVs (152, 165, 169-171). The first examples of ELVs isolated from parasitic
helminth species were E. caproni and F. hepatica (152). These were isolated
from parasite ES by differential ultracentrifugation and contained roughly half of
the proteins identified in the soluble portion of the ES produced by these
parasites, suggesting that vesicle trafficking may be an important mechanism
for releasing these proteins into the extracellular environment, especially those
lacking a signal peptide (152). Host proteins were also detected in both
E. caproni and F. hepatica ELVs, suggesting that these vesicles may enable
two-way communication between the parasite and host. The authors found that
E. caproni ELVs are internalised by rat intestinal epithelial cells (IEC-18), further
suggesting that the parasite uses these vesicles to communicate with the host,
delivering antigens and/or immunomodulatory components to host cells (152).
More recently, Trelis and colleagues demonstrated that E. caproni ELVs have
potential as vaccines, showing that vaccinating mice with these ELVs prior to
infection reduced symptom severity and increased survival (156). Although
there was no reduction in parasite burden compared to non-immunised mice,
the authors show that ELV vaccination promoted IL-10 and TGF-ß production
during infection, suggesting that the decrease in symptom severity may be as a
result of the anti-inflammatory environment that ensues (156).
ELVs have also been isolated from Dicrocoelium dendriticum, S. japonicum and
S. mansoni (169, 172, 173). S. japonicum ELVs have been shown to modulate
the phenotype of RAW 264.7 macrophages in vitro, skewing the population

46
towards a classical M1 phenotype, with a significant increase in surface
CD16/32 expression (a marker of classical macrophage activation), increased
TNF-α production and elevated NO activity (169). Four of the top five most
abundant proteins in S. mansoni ELVs are previously identified vaccine
candidates – Sm-TSP-1, Sm29, saposin B domain-containing protein and
cytoplasmic dynein light chain, suggesting that ELVs may be an important
source of protective antigens (174-177). Triose phosphate isomerase and
glyceraldehyde-6-phosphate dehydrogenase were also identified, both of which
induce a protective response when recombinant forms were administered by
vaccination (173). ELVs have also been isolated from S. mansoni
schistosomula (the tissue migrating life cycle stage) ES products, and have
been identified on the surface of cercariae, the free-swimming form that
penetrates the mammalian host’s skin to initiate infection (178).
The first parasitic nematode known to release ELVs is the rodent intestinal
parasite, H. polygyrus (165). The vesicles released by this parasite fit the
definition of exosomes, as they are 50-100 nm in size, and the authors identified
several known exosome markers associated with the vesicles, including Alix,
enolase and HSP70. The authors report the presence of an Argonaute protein
as well as several small RNAs, suggesting that the parasite may be able to alter
host protein expression using RNA silencing, although the exact mechanism by
which this occurs is yet to be determined (165). The authors demonstrate that
H. polygyrus ELVs are internalised by murine epithelial cells in vitro, which
would allow these small RNAs and Argonaut to be delivered to host cells (165).
Buck and co-workers also show that H. polygyrus ELVs can modulate the
inflammatory airway response to the fungal allergen, Alternaria alternata in vivo,
by preventing the activation of ILC2s, thereby blocking eosinophil recruitment
(165). More recently, Coakley and colleagues demonstrated that intraperitoneal
vaccination with H. polygyrus ELVs prior to infection resulted in a significant
reduction in worm burden compared to sham vaccinated controls (153).
Brugia malayi, which causes lymphatic filariasis, was the first human parasitic
helminth shown to release ELVs (179). L3 larvae are transmitted by mosquitos

47
and migrate to the lymphatics, where they undergo two moults to become adults
(179). The adults live in the lymphatic vasculature and are responsible for the
morbidity caused by this parasite (179). Interestingly, ELV release appears to
be primarily a feature of L3 larvae, with adult worms releasing far fewer ELVs.
The larval ELVs were shown to contain a number of typical exosome markers
including HSP70, enolase and elongation factor-1α (179). The authors report
significant overlap with proteins identified in E. caproni and F. hepatica
exosomes, and found several miRNAs with identical sequences to human
miRNAs with known immunomodulatory functions, suggesting that this could be
a mechanism used by the parasite to promote its own survival (179). The
authors demonstrated that B. malayi ELVs are internalised by J774A.1
macrophages and stimulate an increase in granulocyte colony-stimulating
factor, monocyte chemoattractant protein-1, IL-6, and macrophage inflammatory
protein-2 production by these cells. This suggests the ELVs promote classical
M1 macrophage activation, rather than the alternative M2 phenotype (179).
ELVs have been isolated from Trichuris spcies, as well as from the ovine
parasitic nematode T. circumcinta and canine heartworm Dirofilaria immitis
(154, 180-182). Hansen and colleagues report the presence of both ELVs and
microvesicle-like structures in the secretory products of T. suis L1 larvae, and
also observed vesicles budding from the surface of larvae (154). Tritten and
colleagues isolated ELVs from adult T. muris ES, showing that these vesicles
contain at least 14 parasite-derived miRNAs and 73 proteins (180), while
Eichenberger and co-workers demonstrated that T. muris ELVs can fuse with
murine colonic organoids in vitro (183).
These studies show that helminth ELVs may be a source of both
immunomodulatory and antigenic material. The potential for T. muris ELVs as
vaccine candidates will be investigated in this thesis.

48
1.6 Aims and objectives
Acquired immunity to T. muris is mediated by IL-13, derived mainly from Th2
cells (44, 184). Previous work shows that vaccination with material excreted by
T. muris can induce protective immunity in a range of mouse models (19, 28).
An ideal vaccine might include ES antigens that stimulate IL-13 secretion by
Th2 cells. The aim of this thesis is to identify such antigens as a pipeline to
develop anti-Trichuris vaccines. A stringent vaccination protocol involving
subcutaneous vaccination of male C57BL/6 mice followed by administration of a
low dose infection was developed in order to assess the immunogenicity of
various native T. muris proteins and recombinant immunogenic candidates
in vivo. A sub-objective was to explore the potential of T. muris ELVs as a
source of anti-Trichuris vaccine candidates.
The specific objectives of this thesis are as follows:
1. Identify methods for fractionating T. muris ES into smaller sub-groups
and assess the immunogenicity of these using in vitro assays and
vaccination experiments.
2. Develop a stringent vaccination protocol to assess the efficacy of various
ES vaccines.
3. Identify potential vaccine candidates using a proteomics approach and
assess the immunogenicity of these proteins (in recombinant form) both
in vitro and in vivo.
4. Investigate T. muris ELVs as a potential source of immunogenic
components.

49
Chapter 2: Materials and methods

50
2.1 Maintenance of animals
C57BL/6 (Envigo), SCID and AKR mice (bred at the University of Manchester)
were used for the experiments described in this thesis (Table 1). All mice were
maintained in individually ventilated cages at 22oC ± 1°C and 65% humidity with
a 12 hour light-dark cycle. Mice had free access to food and water, and all
procedures were carried out on mice 6-8 weeks of age or older, under the
Home Office Scientific Procedures Act (1986). Animals were humanely killed by
CO2 inhalation followed by terminal exsanguination or cervical dislocation.
Strain Sex Infection
dose Use
SCID Male/female High Adult ES collection
C57BL/6 Male High L2 ES collection
C57BL/6 Female High Studies involving acute infection (eg re-stimulation assays)
C57BL/6 Male Low Vaccine studies
AKR Male High Vaccine studies
Table 2.1. Description of mouse strains and infection dose used for experiments described in this thesis.
2.2 Maintenance of parasites, ELV removal and preparation of adult ES
The Edinburgh (E) strain of T. muris was used for all experiments. The parasite
was maintained in SCID mice, which were infected with 400 T. muris
embryonated eggs by oral gavage. Adult worms were harvested between 42
and 45 days p.i. Upon killing, the caecum of each animal was removed, cut
open longitudinally, and the caecal content was removed by shaking. The
worms were carefully removed from the caeca using forceps, and were
transferred to clean petri dishes of RPMI media supplemented with 500 U/ml
penicillin and 500 µg/ml streptomycin (all Sigma). Once all worms were
removed, they were transferred to 6-well plates (Corning) with 4 ml media (plus
antibiotics) per well, and were incubated at 37oC and 100% humidity. Media

51
was collected after 4 hours, and worms were transferred to fresh 6-well plates
for overnight incubation in 6 ml media per well.
The next day, worms were removed from the overnight culture using sterile
forceps. The 4-hour and overnight worm cultures were centrifuged at 720 g for
15 minutes to separate the eggs (pellet) from the ES (supernatant). The pellet
was re-suspended in ultrapure water (ddH2O) and eggs were collected after
passing through a 100 μm sterile cell strainer (Fisher Scientific). Eggs were
allowed to embryonate in tissue culture flasks (Helena Biosciences) at room
temperature in the dark for three to four months, and were stored under these
conditions until required.
The ES was passed through a 0.22 μm filter (Millipore), and ELVs were
removed by ultracentrifugation at 100,000 g for 2 hours in polyallomer tubes
(Beckman Coulter, UK). The ELV pellet was washed by ultracentrifugation at
100,000 g for 2 hours in PBS. The poly-cysteine and histidine tailed protein
isoform 2 (P43) was removed from ES samples using NTA-Nickel affinity beads
(Qiagen). The ES was centrifuged at 2000 g to pellet the NTA-Nickel affinity
beads and attached P43. Unless otherwise stated, ‘ES’ refers to samples that
have undergone both ultracentrifugation and the P43 removal step, while
‘ES+P43’ refers to whole ES (with P43) that has not undergone these steps.
These samples were stored at -20oC until required.
2.3 Preparation of larval ES
In order to collect ES from T. muris L2 larvae, male C57BL/6 mice were given a
high dose T. muris infection and were killed at day 15 p.i. The caeca were
removed, cut into small sections (approximately 5 mm in length) and incubated
in PBS with 0.9% (w/v) NaCl for 2 hours at 37oC, with regular vigorous shaking
to remove L2 larvae from the epithelium. Worms were individually isolated using
a pipette, and were transferred to RPMI media supplemented with 500 U/ml

52
penicillin and 500 µg/ml streptomycin. Worms were transferred to fresh media
for overnight culture at 37oC, 5% CO2. L2 ES was collected after 72 hours, and
was concentrated using a pre-washed Amicon Ultra 0.5 ml centrifugal filter with
a 10 kDa molecular weight cut off (MWCO, Merck Millipore). PBS was added to
dilute out the media, and this was then concentrated to approximately 200 µl.
2.4 Egg infectivity and dosage
The infectivity of each batch of eggs was tested prior to usage in experiments.
SCID mice were infected with 400 eggs by oral gavage and worm burdens were
assessed after 35-42 days. Infectivity was calculated as a percentage of
embryonated eggs gavaged (typically between 45 and 60%).
Eggs were washed before each infection by pelleting eggs at 720 g for
15 minutes, pouring away the supernatant, and then re-suspending the eggs in
ddH2O. To prepare eggs for a high dose infection, the washed eggs were
diluted to 90-100 eggs per 50 μl, and mice were infected with 200 μl of the
suspension by oral gavage, so that each mouse received approximately 200
infective eggs (48). For low dose infections, eggs were individually isolated
using a pipette and mice were infected with 25 eggs in 200 μl ddH2O by oral
gavage (48).
2.5 Quantification of worm burdens
Upon sacrifice, the caecum of each mouse was removed and worms were
counted using a binocular dissection microscope (Leica).

53
2.6 Collection of sera
Blood was collected post mortem by cardiac puncture. Serum was collected
following centrifugation at 13,000 g for 15 minutes and stored at -20oC until
required.
2.7 Anti-parasite IgG1 and IgG2a ELISAs
Unless otherwise stated, 96-well immunograde plates (Brand GmbH) were
coated with ES+P43 diluted to 5 μg/ml in 0.05 M carbonate bicarbonate buffer
(pH 7.6, 50 μl/well) (185). Plates were incubated at 4oC overnight in the dark.
The following day, plates were washed three times with PBST (0.05% v/v
Tween 20 in PBS) using a Skatron Scan washer 400 (Molecular Devices) and
blocked with 3% (w/v) BSA (Melford) in PBST for 60 minutes. Serum was
serially diluted in PBST from 1/20 to 1/2560, and this was added to the plates
following another wash step (50 μl per well, 90 minutes incubation). Plates were
washed in PBST, following which bound IgG1 and IgG2a were detected using
biotinylated rat anti-mouse IgG1 (BioRad, 1:2000 dilution) and anti-IgG2a (BD
Biosciences, 1:1000 dilution) respectively (50 μl per well, 60 minutes
incubation). Following another wash step, streptavidin-peroxidase conjugate
(Roche, 1:1000 dilution) was added (75 μl per well). Plates were washed for a
final time before ABTS substrate (10% 2,2’azino 3-ethyl benzthiazoline in
0.045 M citrate buffer) was added (100 μl per well). Plates were read at 405 nm
with a reference of 490 nm on a VersaMax microplate reader (Molecular
devices, UK).
2.8 Lymphocyte re-stimulation assay
Female C57BL/6 mice were used for re-stimulation assays. These mice were
given a high dose infection and sacrificed at day 20 p.i., which corresponds to
peak Th2 cytokine response during acute infection (Bancroft, personal

54
communication). Age-matched naïve controls were also sacrificed. The
mesenteric lymph nodes (MLNs) were removed and were transported back to
the laboratory on ice in RPMI media supplemented with 2% FBS, 2 mM
L-glutamine (GIBCO/Invitrogen), 100 U/ml penicillin, 100 µg/ml streptomycin
(wash media). Each MLN was disrupted using the piston of a 2.5 ml syringe,
and cells were passed through a 100 μm sterile cell strainer (Fisher Scientific)
to make a cell suspension in wash media. The cell suspension was centrifuged
at 430 g for 5 minutes to pellet the cells, the supernatant was removed and cells
were re-suspended in 10 ml wash media. This wash step was repeated twice,
and after the second time, the cells were re-suspended in 1 ml (for cells taken
from naïve mice) or 2 ml (for cells from infected mice) RPMI supplemented with
20% FBS, 100 U/ml penicillin, 100 µg/ml streptomycin and 2 mM L-glutamine.
The number of cells in each sample was measured using a CASYcounter®
(Scharfe systems) and the volume was adjusted to give a concentration of
1x107 cells/ml. For re-stimulation assays, the cells from three infected mice
were pooled and the cells from two naïve mice were pooled in order to allow
multiple antigen fractions to be tested. Cells were re-stimulated with various ES
proteins diluted to 50 μg/ml in RPMI media supplemented with 100 U/ml
penicillin, 100 μg/ml streptomycin in a 1:1 (v/v) ratio of cell suspension to
protein. Cells were incubated for 42 hours at 37oC and 5% CO2 in sterile 96-well
flat bottom tissue culture plates (Corning). Plates were centrifuged at 430 g for 5
minutes to pellet cells and the supernatants were transferred to sterile 96-well
round bottom tissue culture plates (Corning). Supernatants were stored at -20oC
until required.
2.8.1 Measuring cytokine production in cell supernatants
Cytokine levels in the cell supernatants (see above) were measured by
cytometric bead array, using a Mouse/Rat Soluble Protein assay kit (BD
Biosciences). The kit contained wash buffer, assay diluent, capture bead diluent
and detection reagent diluent. Cytokine-specific Mouse/Rat Soluble Protein Flex

55
Kits, each containing capture beads, detection reagent, and cytokine standards
were purchased separately from BD Biosciences. Levels of the following
cytokines were assessed: IL-6, IL-9, IL-10, IL-13, IL-17A, IFN-γ and TNF-α. To
assess IFN-γ production, supernatants were diluted 1:10 with RPMI plus
antibiotics.
Cytokine standards were prepared according to the manufacturer’s instructions
and were serially diluted 2-fold from 2500 to 10 pg/ml in assay diluent. The
serially diluted cytokine standards and cell supernatants were pipetted into
individual wells of a pre-washed (with wash buffer) 96-well round bottom tissue
culture plate (16.6 μl per well). Next, the capture beads were prepared in
capture bead diluent. The volume to be prepared was determined by multiplying
the number of test wells by 16.6 μl. Each test well received 0.33 μl of each
capture bead in 16.6 μl capture bead diluent, so the volume of each bead to
add to the capture bead diluent was determined by multiplying the number of
test wells by 0.33 μl. Beads were vortexed for 10 seconds before they were
added to the capture bead diluent and this was vortexed regularly as it was
added to the plate (16.6 μl per well). Plates were incubated on a digital shaker
(500 rpm) for 60 minutes at room temperature to allow capture beads to bind
the samples.
The detection reagent was prepared in detection reagent diluent. As with the
preparation of the capture bead reagent, the volume to be prepared was
determined by multiplying the number of test wells by 16.6 μl. Each test well
received 0.33 μl of each detection reagent in 16.6 μl detection reagent diluent,
so the volume of each detection reagent to add to the detection reagent diluent
was determined by multiplying the number of test wells by 0.33 μl. The
detection reagent was mixed by gentle pipetting and 16.6 μl were added to each
test well. Plates were incubated on a digital shaker (500 rpm) in the dark for 60
minutes at room temperature to allow detection reagents to bind.
Wash buffer was added to the test wells (150 μl per well), following which,
plates were centrifuged at 430 g to pellet beads. Plates were tipped upside

56
down to remove wash buffer, following which 70 μl wash buffer were added to
each well. The assay plates were read using a MACSQuant Analyzer 10
(Miltenyi Biotech) and data was extracted using FCAP array software, Version
3.01 (BD Bioscience).
2.9 Fractionation of adult ES
2.9.1 Fractionation of adult ES by gel filtration chromatography
Prior to fractionation, adult ES was dialysed in 25 mM Tris-HCl, 10 mM NaCl
(pH 7.4) and concentrated by approximately 50-fold. All 2.4 ml columns
(Superdex 75, Superdex 200 and Superose 12) were attached to an ӒKTA
Ettan purification system (all GE Healthcare). Samples were injected and
separated on the column at a flow rate of 0.05 ml/min. Protein was eluted in
0.05 ml fractions in 25 mM Tris, 10 mM NaCl (pH 7.4) and the column effluent
was monitored for UV absorbance at 280 nm.
The 24 ml Superose 12 and Superdex 75 columns were attached to an
ӒKTAprime plus purification system (all GE Healthcare). Samples were injected
and separated on the column at a flow rate of 0.5 ml/min. Protein was eluted in
0.5 ml fractions in 25 mM Tris, 10 mM NaCl (pH 7.4) and the column effluent
was monitored for UV absorbance at 280 nm. Unicorn software was used to
control these fractionation processes, and elution profiles were viewed using the
PrimeView software (GE Life Sciences). The protein content of the resulting
fractions was separated by SDS-PAGE and gels were visualised using either
Coomassie blue or silver nitrate staining (see section 2.10).

57
2.9.2 Fractionation of adult ES and pool 3 by anion exchange and gel
filtration chromatography
Adult ES was also fractionated using a 2.4 ml MiniQ anion exchange column.
This column was attached to an ӒKTA Ettan plus purification system, and
fractionation was controlled by Unicorn software. Samples were injected onto
the column and proteins were eluted by increasing the salt concentration of the
buffer (25 mM Tris, pH 7.4) from 10 to 300 mM NaCl over 15 column volumes,
followed by a gradient of 300 to 1000 mM NaCl over 8 column volumes.
Pool 3 (the product of pooling Superose 12 fractions 24-27) was further
fractionated using a Resource Q anion exchange column. This column was
attached to an ӒKTAprime plus purification system and proteins were also
eluted by increasing the salt concentration of the buffer (25 mM Tris, pH 7.4)
from 10 to 300 mM NaCl over 15 column volumes, followed by 1000 mM NaCl
for 5 column volumes. The protein content of the resulting fractions was
separated by SDS-PAGE and gels were stained using either Coomassie blue or
silver nitrate staining (see section 2.10).
2.10 SDS-PAGE
SDS-PAGE was carried out using NuPage Novex 4-12% BisTris Mini-gels in 1x
MOPs running buffer (both Invitrogen, Life Technologies). Samples were
reduced in 10 mM dithiothreitol (DTT), 1 x NuPAGE LDS sample buffer
(Invitrogen) and heated at 100oC for 5 minutes. Precision Plus Protein All Blue
Standards (Bio-Rad Laboratories) were run along side samples, to enable
molecular weight assessment of protein bands. Electrophoresis was carried out
using XCell II Mini-cell electrophoresis tanks (Novex) at 200 V for 48 minutes.
Protein bands were visualised using either Coomassie blue or silver nitrate
staining.

58
2.10.1 Coomassie blue staining
Gels were washed briefly in dH2O, and were stained with InstantBlue
(Expedeon Ltd) for at least 1 hour. Gels were washed extensively with dH2O,
and were imaged using a ChemiDoc MP Imaging system (Bio-Rad).
2.10.2 Silver staining of SDS-PAGE gels
Gels were washed briefly in dH2O, fixed for 20 minutes in 50% (v/v) methanol,
5% (v/v) acetic acid, and then rinsed in 50% (v/v) methanol for 10 minutes. Gels
were washed for 10 minutes in dH2O, sensitised in 0.02% sodium thiosulphate
for 1 minute, and were then washed in dH2O for 5 minutes. Gels were
submerged in 0.1% (w/v) silver nitrate in dH2O for 20 minutes, followed by
another wash step. Gels were developed in 0.04% (v/v) formalin, 2% (w/v)
Na2CO3 in dH2O. Once bands were visualised, development of gels was
terminated using 5% (v/v) acetic acid. Gels were rinsed in dH2O and were
imaged using a ChemiDoc MP Imaging system.
2.11 Assessing protein concentration of samples
Protein concentration was assessed using either a Nanodrop 1000 (Labtech
International) or a bicinchoninic acid (BCA) assay kit (Fisher Scientific),
according to the manufacturer’s instructions. For ELV samples, vesicles were
lysed with 0.1% (v/v) SDS prior to assessment of protein concentration.
2.12 Western blotting
Samples were loaded onto NuPAGE Novex 4-12% BisTris mini gels alongside a
protein ladder (Precision Plus Protein All Blue Standards if assessing serum

59
antibody binding, or Odyssey® One-Color Protein Molecular Weight Marker,
Li-cor, if assessing His-probe binding) and electrophoresis was carried out as
described in section 2.10. Electrotransfer of proteins from NuPAGE gels to
nitrocellulose membrane was carried out using an XCell IITM semi-wet Blot
Module (Invitrogen) run at 35 V for one hour with 20% (v/v) methanol,
1x NuPAGE® transfer buffer. Membranes were blocked with 5% skimmed milk
(Marvel) in Tris-buffered saline-Tween (TBST; 10 mM Tris-base / 150 mM NaCl
/ 0.1 % (v/v) Tween-20, pH 8.0) for 15 to 30 minutes, and were then rinsed 4
times in TBST for a total of 20 minutes.
2.12.1 Western blotting using serum from T. muris infected mice
Following the block and washing steps, membranes were probed with serum
from ES vaccinated and/or T. muris infected mice (1:300 dilution) overnight. The
following day, membranes were washed in TBST for 20 minutes and bound
antibody was detected using an anti-mouse IgG (whole molecule) alkaline
phosphatase conjugated antibody (1:10,000 dilution in TBST, Sigma). Following
a wash step in TBST for 20 minutes, immunoblots were revealed using
chromogenic substrates, BCIP (5-bromo-4-chloro-3-indolyl-phosphate, 100%
v/v in dimethylformamide) and nitro blue tetrazolium (NBT, 70% v/v in
dimethylformamide) in a 1:2 ratio in TBST. Membranes were washed for 20
minutes in TBST and were imaged using a ChemiDoc MP Imaging system.
2.12.2 Western blotting to detect His-tagged recombinant proteins
Following the block and washing steps, membranes were probed with a
His-probe (H-3 mouse monoclonal IgG1, Santa Cruz Biotechnology).
Membranes were washed in TBST for 20 minutes, and bound His-probe was
detected using an AlexaFluor 680 goat anti-mouse IgG (H+L) antibody (Life
Technologies). Following a wash step in TBST for 20 minutes, His blots were

60
visualised at 700 and 800 nm using the Odyssey CLx infrared imaging system
(LI-COR Biosciences).
2.13 Mass spectrometry and proteomic analysis of ES components
Samples were reduced with 20 mM DTT for 2 hours and alkylated with 50 mM
iodoacetemide for 20 minutes in the dark. Ammonium bicarbonate was added to
make a final concentration of 0.1 M, following which sequencing grade trypsin
(Promega) was added at a ratio of 1:50 (trypsin to protein by weight), and
samples were incubated at 37oC overnight. The resultant peptides were
acidified to pH 2 with formic acid and purified using ZipTip desalting pipette tips
(Millipore). ZipTips were pre-washed four times in acetonitrile, then six times in
0.1% (v/v) formic acid. The ZipTips were loaded with sample, washed eight
times in 0.1% (v/v) formic acid, and then peptides were eluted using 50% (v/v)
acetonitrile, 0.1% (v/v) formic acid. A centrifugal vacuum evaporator was used
to dry the samples, and these were then re-suspended in 10 μl 5% (v/v)
acetonitrile, 0.1% (v/v) formic acid.
The BioMS core facility team at the University of Manchester performed liquid
chromatography-tandem mass spectrometry (LC-MS) on each sample using a
NanoAcquity LC coupled to a LTQ Velos mass spectrometer, and the results
were analysed using Mascot MS/MS ion search (Matrix Science, see Table 2.2
for search criteria) and searched against the T. muris proteome, version 2.2
(Sanger Centre). Scaffold Proteome Software (Scaffold, USA) was used to
calculate the exclusive unique peptide count for each protein (criteria set to 95%
protein threshold, 50% peptide threshold, minimum 2 peptides identified). The
Sanger Centre FTP site
(ftp://ftp.sanger.ac.uk/pub/project/pathogens/Trichuris/muris/genes/) was used
to search for T. muris protein and cDNA sequences. The SignalP Server
version 4.1 (http://www.cbs.dtu.dk/services/SignalP/, Technical University of
Denmark) was used to predict whether proteins had signal peptides. The

61
homology between T. muris candidate proteins and those of other species was
analysed using the BLAST protein search tool
(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=Blast
Search&LINK_LOC=blasthome, National Institute of Health).
Enzyme Trypsin
Maximum Missed Cleavages 1
Fixed modifications
Carbamidomethyl
(C)
Quantitation method None
Variable modifications Oxidation (M)
Peptide Mass Tolerance 1.2
Peptide Mass Tolerance Units Da
Fragment Mass Tolerance 0.6
Fragment Mass Tolerance
Units Da
Mass values Monoisotopic
Instrument type Default
Isotope error mode 0
Table 2.2. Mascot search criteria.
2.13.1 Mass spectrometry analysis of T. muris ELVs
Mass spectrometry analysis of ELV samples was performed as described by
Marcilla et al. (2012). ELVs were precipitated with 100 μl chloroform/methanol
(1:1 ratio) and then dissolved in 50 μl 50% (v/v) trifluoroethylene (TFE), 50 mM
ammonium bicarbonate. Samples were treated with 2 mM DTT for 30 minutes
at 60oC, followed by 5 mM iodoacetamide for 30 minutes at room temperature
(in the dark), and finally 10 mM DTT at room temperature for 30 minutes. The
samples were diluted with 50 mM ABC such that the final concentration of TFE
was <5%. Samples were incubated with 500 ng trypsin overnight at 37oC,
following which samples were acidified using formic acid and peptides were

62
purified using a ZipTip step, as described in section 2.13. Samples were dried in
a vacuum centrifuge, reconstituted in 10 μl 5% (v/v) acetonitrile, 0.1% (v/v)
formic acid and mass spectrometry analysis was carried out as described
above.
2.14 Vaccination studies
All vaccination studies were carried out using male C57BL/6 or AKR mice.
Unless otherwise stated, mice were vaccinated subcutaneously with 30 μg of
protein (either native ES proteins, or recombinant proteins) diluted in Tris buffer
(pH 7.4), followed by a second vaccination 14 days later with 15 μg antigen.
Vaccinations were formulated with aluminum hydroxide (1:1 v/v ratio of protein
to adjuvant), and 100 μl was administered using a 25-gauge needle
(BD Microlance). Vaccinations were prepared by adding aluminum hydroxide
drop-wise to the antigen preparation on a digital shaker. The antigen/adjuvant
solution was then mixed on a 360o rotator for approximately 40 minutes. For the
recombinant proteins vaccination study, an additional group was included,
whereby mice were vaccinated with each of the 5 recombinant proteins in a
single vaccination, formulated with aluminum hydroxide (30 μg of each protein,
150 μg protein in total), followed by a second vaccination 14 days later (15 μg of
each protein, 75 μg in total).
Vaccinations with Freund’s adjuvants and Montanide ISA 720 were prepared
using two luer lock syringes and one I-connector. Material was pushed back
and forth between the two syringes to produce a thick emulsion. Freund’s
adjuvants were prepared in 1:1 dilution (v/v) of antigen to adjuvant; the first of
the three vaccinations was formulated with CFA, while the second and third
were formulated with IFA (both Sigma). Montanide ISA 720 vaccinations were
prepared in 70:30 ratio (v/v) of adjuvant to antigen. All sham vaccinations were
carried out by diluting adjuvant with Tris buffer (pH 7.4), according to the ratios
stated above.

63
Mice were infected with 25 T. muris eggs 28 to 30 days after the first
vaccination and were sacrificed at day 32 p.i., as this is when the worms reach
adulthood and the infection is considered to have reached patency (31). Worm
burdens were assessed, along with anti-parasite IgG1 and IgG2a serum
antibody levels.
2.14.1 Proteinase K treatment of ES and subsequent vaccination
ES was denatured by heating at 95oC in 6M guanidinium chloride, 50 mM Tris
(pH8), 5 mM DTT for 5 minutes. The denatured material was buffer exchanged
to 50 mM Tris, 5 mM CaCl2 (pH 8.0) using a pre-washed Amicon Ultra 0.5 ml
centrifugal filter (10 kDa MWCO) and 100 μg/ml proteinase K was added. The
reaction was incubated overnight at 40oC and the following day, peptides were
collected using a pre-washed Amicon Ultra 0.5 ml centrifugal filter
(10 kDa MWCO). A small sample of the treated ES was separated by
SDS-PAGE (gel stained with Coomassie blue) to confirm degradation. C57BL/6
mice were vaccinated subcutaneously with 30 μg of the digested material
(protein concentration measured before proteinase K addition), followed by
vaccination with 15 μg of material 14 days later (antigen diluted in Tris buffer,
pH 8.0). As a negative control, a group of 3 mice were vaccinated with
proteinase K in Tris buffer (pH 8.0) prepared as described above. All
vaccinations were formulated with aluminum hydroxide as described in
section 2.14.
2.14.2 Serum transfer from vaccinated to unvaccinated mice
C57BL/6 mice were vaccinated with ES formulated with aluminum hydroxide as
described in section 2.14. These mice were sacrificed 14 days after the second
vaccination and serum was collected as described in section 2.6. The serum
was heated at 54oC for 30 minutes to inactivate cytokines and other serum
proteins. Naïve C57BL/6 mice were injected intraperitoneally with 500 μl of

64
serum from ES or sham-vaccinated mice. These mice were infected with 25 T.
muris eggs by oral gavage immediately following the serum transfer and were
sacrificed at day 32 p.i. to assess worm burdens.
2.14.3 ELV vaccination studies
C57BL/6 mice were vaccinated subcutaneously with 3 µg of ELV material
(formulated without adjuvant), followed by a second vaccination with 1.5 µg of
material 14 days later. The sham group was vaccinated subcutaneously with
100 μl of PBS only. A positive control group was included, whereby mice were
vaccinated subcutaneously with 30 μg ES, followed by 15 μg of ELV material
14 days later (vaccinations formulated with aluminum hydroxide as described in
section 2.14). Mice were infected with 25 T. muris eggs 14 days after the
second vaccination and were sacrificed at day 32 p.i. as described above.
Experiments were also carried out to determine the effect of vesicle lysis on
ELV vaccinations. To do this, ELV samples were lysed by adding 0.1% (v/v)
SDS, followed by three freeze/thaw cycles, whereby vesicles were frozen in
liquid nitrogen and thawed in a 37oC water bath. ELV samples were vortexed
between each step. Lysis was confirmed using dynamic light scattering (DLS,
see section 2.16). Mice were vaccinated subcutaneously with 3 μg of lysed ELV
material, followed by 1.5 μg of lysed ELV material 14 days later. Mice were
subcutaneously vaccinated with unlysed ELV samples (as described above) as
a positive control. Mice were infected with 25 T. muris eggs by oral gavage
14 days following the second vaccination and were sacrificed at day 32 p.i. to
assess worm burdens.

65
2.15 DNA synthesis, transfections and collection of recombinant
proteins
Immunogenic candidates were identified as described in Chapter 4. For each
candidate, cDNA synthesis and cloning into the pCepHis vector, which encodes
ampicillin and puromycin resistance genes, (see Figure A1.1 for full sequence)
were carried out by Thermo Fisher Scientific Geneart GmbH. One Shot®
TOP10 Chemically Competent Escherichia coli (Thermo) were transformed with
each construct according to the manufacturer’s instructions. The bacteria were
streaked out onto carbenicillin (Melford) antibiotic selection plates (0.1 mg/ml
carbencillin in Luria broth, LB, agar) and left to grow overnight at 37oC. For each
construct, a single colony was picked and this was grown overnight in LB.
Bacteria were lysed and DNA was collected using the PureLink® Quick Plasmid
Miniprep Kit (Thermo) according to the manufacturer’s instructions.
HEK293 EBNA cells were grown to 80% confluency in T75 flasks (Corning) for
transfections. DNA-lipid complexes were prepared for transfections by adding
20 to 30 µg DNA and 30 µl lipofectamine 2000 (Life Technologies) to 3 ml Opti-
MEM reduced serum media I (Thermo). DNA-lipid complexes were incubated
for 12 to 15 minutes. Cells were washed in PBS (sterile) and were incubated
with the DNA-lipid complexes for 3 days at 37oC, 5% CO2, after which the
transfection medium was collected and antbiotic selection with DMEM
supplemented with 10% FBS, 100 U/ml penicillin, 100 µg/ml streptomycin and
0.5 μg/μl puromycin was commenced. Antibiotic selection was applied for 2-3
weeks to kill non-transfected cells and selection media was replaced twice a
week. Following the 2-3 week antibiotic selection period, cells were grown in
DMEM supplemented with 10% FBS, 100 U/ml penicillin, 100 µg/ml
streptomycin and 0.05 μg/μl puromycin. Once cells were confluent, they were
split into six T75 flasks using trypsin-EDTA solution (Sigma). Once these were
confluent, cells were divided into three triple layer flasks to enable efficient
protein collection. Once cells were fully confluent in the triple layer flasks, cells
were washed with PBS and then the media was replaced with DMEM, high
glucose, GlutaMAX™ Supplement media (Thermo) for protein collection. Media

66
was collected in 50 ml falcon tubes (Corning) twice a week. Medium was
centrifuged at 2000 g for 5 minutes to pellet cells, following which medium was
transferred to sterile bottles and stored at -20oC until required.
2.16 Purification of recombinant proteins
DMEM media containing recombinant protein was dialysed overnight with 20
mM Tris, 400 mM NaCl, 10 mM imidazole (pH 7.9) using BioDesignDialysis
tubing with either 3,000 or 14,000 MWCO (Fisher Scientific). The buffer was
changed the following morning to allow for thorough dialysis. The buffer
containing the dialysed proteins was filtered using a 0.22 μm filter. Each protein
was purified by nickel affinity chromatography, followed by size exclusion
chromatography. For the nickel affinity step, protein was pumped through a
HisTrap FF 1 ml column to enable the His-tagged recombinant proteins to bind.
Bound protein was eluted using 200 mM imidazole followed by 500 mM
imidazole (both in 20 mM Tris, 400 mM NaCl, pH 7.9) and protein was collected
in 1 ml fractions. The fractions were separated by SDS-PAGE to determine in
which fractions the recombinant proteins were eluted. Size exclusion
chromatography was carried out on those fractions containing the recombinant
protein, in order to further purify the protein and remove the imidazole.
The size exclusion chromatography step was carried out using a Superdex 75
gel filtration column and the AKTAPurifier protein purification system, under the
control of Unicorn software. Fractions were analysed by SDS-PAGE and mass
spectrometry analysis was carried out to confirm the identity of each purified
recombinant protein. The fractions containing the recombinant protein of
interest were pooled and concentrated. Proteins were stored at -20oC until
required.
HEK293 cells were not able to express five of the recombinant proteins. In order
to determine whether the proteins were expressed but not secreted, transfected

67
cells were lysed with Radioimmunoprecipitation assay (Ripa) buffer (Thermo
Scientific). Western blotting was carried out to determine whether cell lysates
contained His-tagged proteins corresponding to the predicted molecular weight
for each protein.
2.17 TEM analysis of ELV samples
Samples were transferred to formvar-carbon-coated EM grids and negatively
stained with 2% (w/v) uranyl acetate. Samples were imaged using a Tecnai
BioTwin microscope, at 100 Kv under low-dose conditions. Images were
recorded using a Gatan Orius CCD camera at 3.5 Å/pixel. ImageJ v1.46r (NIH)
was used to view images and to add scale bars.
2.18 DLS measurements
DLS was used to identify the size distributions of the exosome preparations.
DLS measurements were performed using a Zetasizer Nano-S (Malvern) at a
controlled temperature of 25°C. Scattering at 90° gives rise to the z-average
particle scattering intensity, which is deduced from the correlation function
(186). Sample polydispersity was deduced from the slope of the correlation
function and mean particle diameter from the diffusion coefficient according to
Mie theory (186). The diffusion coefficient and therefore size of the particles
were calculated using a refractive index of 1.39. Particle size was measured 13
times for each sample. The number distribution of particles was used to report
the relative amounts of each peak distribution and this transformation assumes
that all particles are spherical and that the sample density is homogenous (186).

68
2.19 ELV fusion assay
In order to determine whether ELVs could fuse with mammalian colonic
epithelial cells, HT-29 cells were grown to 80% confluency in RPMI medium
supplemented with 10% (v/v) FBS, 1% (v/v) L-glutamine, 100 U/ml penicillin and
100 µg/ml streptomycin on μ-slide 8-well Ibitreat plates (Ibidi). Once the cells
were confluent, they were washed with sterile PBS (Sigma) and cells were
starved of FBS for 16 hours. During this period, cells were incubated with RPMI
supplemented with 1% (v/v) L-glutamine, 100 U/ml penicillin and 100 µg/ml
streptomycin.
ELVs were labeled with PKH26 red fluorescent cell linker (Sigma) and excess
dye was removed using exosome spin columns (Life Technologies), according
to the manufacturer’s instructions. The labeled ELVs were incubated with the
HT-29 cells for 4 hours. Cells were washed thoroughly in PBS to remove
extracellular PKH26 labeled vesicles, and then imaged using a Swept Field
confocal (Nikon), with a [60x/ 1.40 Plan Apo] objective on an inverted TE2000
inverted microscope. The settings were as follows, pinholes [30μm], scan speed
[1400Hz or 100fps], format [512 x 512]. The PKH26-stained ELVs were excited
with the 543 nm laser line and images were acquired on a Cascade 512B EM
CCD camera (Photometrics) through the Elements Software (Nikon).
2.20 Graphing and statistical analysis
All graphing and statistical analysis was carried out using GraphPad Prism
version 7. One-way ANOVA is a common way to compare three or more
variables, however in order to carry out such an analysis, data must be normally
distributed (187). For each experiment, a Shapiro-Wilk normality test was
carried out to confirm that data was normally distributed. All experiments
passed the normality test, which meant that one-way ANOVA (followed by a
Dunn’s Multiple Comparison Test) was appropriate. Unless otherwise stated,

69
error bars represent SEM. Statistical significance is represented by * (P< 0.01),
** (P<0.05), *** (P< 0.001) or **** (P< 0.0001).

70
Chapter 3: Defining the host protective
components within the soluble portion of
T. muris ES

71
3.1 Introduction
With over 465 million people infected worldwide, T. trichiura is one of the four
most clinically important STHs. Heavy worm burdens are associated with
Trichuris dysentery syndrome, symptoms of which include stomach pain,
diarrhoea and in extreme cases, rectal prolapse (2, 8). A combination of anti-
helminthic drugs, sanitation improvements and prophylactic vaccines are
predicted to reduce the morbidity associated with these infections (22).
However a recent meta-analysis showed that the benzamidazole drugs
currently used to treat STH infections have poor efficacy against T. trichiura,
and there are reports of drug resistance arising within parasite populations in
Vietnam and Zanzibar (20, 21, 188). In addition, there are no commercially
available vaccines against human STH species, and few against their veterinary
counterparts (3).
The naturally-occurring murine whipworm, T. muris, has been used for over 50
years as a model for T. trichiura (32). These parasites share extensive
homology at the genomic and transcriptomic levels, and the immune responses
associated with both acute and chronic infection have been well studied using
the T. muris mouse model (31, 33). Infection dose can influence the relative
resistance/susceptibility of mice to T. muris. Most standard laboratory strains
are capable of expelling a high dose infection (200 eggs), while a low dose
infection (10-25 eggs), which is more reflective of natural challenge, promotes
an IFN-γ rich CD4+ T helper cell type 1 (Th1) response, leading to chronic
infection and colitis (48). During acute infection, worm expulsion is driven by a
Th2 response, and the Th2 cytokines, IL-9 and IL-13, are known to stimulate
effector mechanisms that drive worm expulsion (31). These effector
mechanisms include increasing mucus production and epithelial turnover in the
caecum (IL-13), and inducing intestinal hypercontractility (IL-9) (57, 68, 189).
Natural immunity is acquired during acute infection, and mice are protected
against all subsequent infections (both high and low dose) (48).

72
Vaccination with ES collected from adult worms has previously been shown to
protect naturally susceptible AKR mice against a subsequent high dose
infection and promote earlier expulsion of an acute infection in NIH mice
(19, 28, 115). However, relatively little progress has been made towards
identifying the parasite antigens responsible for triggering expulsion, and
crucially the potential for ES vaccinations to protect mice against a low dose
infection, which is more reflective of natural infection and primes for
chronicity (48), has yet to be investigated.
In this chapter, a stringent vaccination protocol in which to test various
combinations of ES components was developed using a chronic infection model
(low dose infection in male C57BL/6 mice). This was used in combination with
an acute infection model (high dose infection in female C57BL/6 mice) in order
to assess cytokine production and antibody response to various ES
components in a setting where immunity is acquired in response to infection.
Various chromatography media were investigated to fractionate adult T. muris
ES, in order to narrow down the search for protective components.
Experiments were performed to investigate whether the protective properties of
adult ES are dependent on the protein content, and whether vaccination with
adult T. muris ES can induce long-lasting immunity. For the latter experiment, a
low dose infection was administered to male C57BL/6 mice a total of 50 days
following vaccine boost (as opposed to 14 days after, as was performed for the
other experiments described in this chapter). This time point was chosen as it is
generally accepted that in order for immunological memory to develop, a period
of at least 30 days should have elapsed (190-192). Larval ES was also
investigated as a source of protective antigens. For this experiment, ES was
collected from larvae at day 15 p.i. (L2 stage), which is around the time that
worms are expelled during acute infection (31). The protective capacity of larval
ES vaccinations was compared to that of the adult ES vaccinations and
proteomic analysis was also carried out in order to assess the similarities and
differences between the protein content of the two ES collections.

73
The results of these experiments showed that vaccination with ES proteins
(collected from the larval and adult stages) stimulates long-lasting protection
against a subsequent low dose infection in C57BL/6 mice, and a sub-fraction of
ES (pool 3) was identified as a potent source of protective antigens that will be
investigated further in Chapter 4.
3.2 Preparation of T. muris ES for fractionation using gel filtration
chromatography
T. muris ES contains a myriad of proteins, ranging from 10 to 250 kDa in size
(Figure 3.1A). Unpublished data from our laboratory shows that the major ES
component, a 43 kDa protein referred to as the P43 (indicated by red box on
Figure 3.1A), is poorly immunogenic, and therefore the work presented here
has focused on the other ES components. The P43 has a natural poly-histidine
tag, enabling the removal of this protein from ES using nickel affinity
chromatography (Allison Bancroft, University of Manchester, personal
communication). To do this, ES was incubated with nickel beads on a rotator for
two hours, after which it was centrifuged to pellet the beads (with captured P43,
Figure 3.1B), and the supernatant (containing the other ES proteins) was
removed for further study. For the purpose of this thesis, ‘ES’ refers to the
material remaining after P43 removal (Figure 3.1C), while ‘ES+P43’ refers to the
native form (Figure 3.1A). VivaSpin concentrators were used to concentrate and
buffer exchange the ES to 25 mM Tris, 10 mM NaCl (pH 7.4), a buffer suitable
for both size exclusion and anion exchange chromatography.

74
Figure 3.1. Preparation of ES by removing P43 using nickel affinity chromatography. The major ES component, the P43 (red box, A), was removed from ES+P43 (A) using nickel beads. The protein was eluted using 250 mM imidazole in 20 mM Tris, 400 mM NaCl, pH 7.9 (B) and the remaining ES proteins (C, referred to from hereon in as ‘ES’) were concentrated using a Vivaspin concentrator. The left hand lane of each gel shows the molecular weight markers in kilodaltons (kDa).
3.2.1 Investigating the suitability of gel filtration media to fractionate ES
into smaller sub-groups
Tandem mass spectrometry analysis of tryptic peptides showed that ES
contains over 460 proteins (see Appendix 1, Table A1.1), and therefore it was
necessary to identify a method to divide ES into smaller sub-groups in order to
refine the search for protective material. A number of gel filtration media,
including Superdex 75, Superdex 200 and Superose 12 were investigated, in
order to determine which would most effectively divide ES into different size
groups. Anion exchange chromatography was also investigated, as discussed
in section 3.7. The Superdex 75 gel filtration resin was able to divide the ES into
two broad groups – the higher molecular weight (Mw) group (80 to 120 kDa)
was eluted between 0.9 and 1 ml, while the lower Mw group (10 to 60 kDa) was
eluted between 1.2 and 1.3 ml (Figure 3.2A). Similarly, the Superdex 200 gel
filtration resin was able to separate larger ES proteins from medium-sized and
smaller proteins – the larger proteins (120 to 160 kDa) were eluted between
1.15 and 1.25 ml, and continued to be eluted up until 1.5 ml, along with the
medium-sized proteins (30 to 70 kDa), while the smaller proteins (10 to 70 kDa)
250
10
150
100
75
50
37
25 20
15
kDa ES
A) ES+P43
B) C)
250
150
100
75
50
37
25 20 15 10
kDa ES
250
150 100
75
50 37
25
20
15
10
kDa 250
Fractions containing P43 eluted using 250 mM imidazole
100
75
50
37
25 20
15
10
kDa 250
150
100
75
50
37
25 20 15 10
kDa
1 2 3 4 5 6 7 8

75
were eluted between 1.6 and 1.7 ml (Figure 3.2B). In contrast, the ES proteins
were fractionated more effectively by the Superose 12 gel filtration resin, so that
the protein composition of the fractions progressively decreased in size
(Figure 3.2C).
Figure 3.2. Investigating the suitability of Superdex 75, Superdex 200 and Superose 12 gel filtration media for fractionating ES by size exclusion chromatography. The UV trace (absorption measured at 280 nm) for each fractionation is shown in (A), (C) and (E). The resulting fractions were separated by SDS-PAGE and visualised by silver nitrate staining as shown in (B), (D) and (F). The left hand lane of each gel shows the molecular weight markers in kDa.
0.8
1.0
5
0.8
5
0.9
0.9
5
1.0
1.1
0
1.1
5
250 150 100 75
50 37
25 20 15 10
kDa 250
150 100 75
50 37
25 20 15 10
kDa 1.2
1.2
5
1.3
1.3
5
1.5
1.4
5
Volume (ml) Volume (ml)
250
150 100 75 50 37
25 20 15 10
kDa
1.2
5
1.3
0
1.3
5
1.5
0
1.1
5
1.2
0
1.4
0
1.5
5
1.6
0
1.6
5
1.7
0
1.7
5
1.8
0
A) Superdex 75 UV trace B) SDS-PAGE gel for Superdex 75
C) Superdex 200 UV trace D) SDS-PAGE gel for Superdex 200
Volume (ml) Volume (ml)
E) Superose 12 UV trace F) SDS-PAGE gel for Superose 12
0.0 0.5 1.0 1.5 2.0 2.5 3.0
0
50
100
150
200
Volume (ml)
Ab
so
rban
ce
0.0 0.5 1.0 1.5 2.0 2.5 3.0
0
20
40
60
80
Volume (ml)
Ab
so
rban
ce
0.0 0.5 1.0 1.5 2.0
0
25
50
75
100
Small Sup12
Volume (ml)
Ab
so
rban
ce
0.8
1.0
5
0.8
5
0.9
0.9
5
1.0
1.1
0
1.1
5
0.7
5
1.1
5
Volume (ml)
250
150 100 75
50 37
25 20 15 10
kDa
250
150 100 75
50 37
25 20 15 10
kDa
Volume (ml)
Volume (ml)
Volume (ml)
Ab
so
rban
ce
A
bso
rban
ce
Ab
so
rba
nce

76
Based on these preliminary studies, Superose 12 gel filtration media appeared
to be the most effective for fractionating ES into distinct sub-groups. These
initial studies were carried out using 2.4 ml columns. The next step was to
confirm that the 24 ml Superose 12 column fractionated ES in a manner similar
to the 2.4 ml column, as the larger column would enable fractionation of a
greater amount of material in each chromatography run.
3.2.2 Division of ES into four sub-groups using Superose 12 gel filtration
media
The 24 ml Superose 12 column appeared to fractionate ES in a similar manner
to the smaller column. For both columns, the first few protein containing
fractions contained material of 30 to 150 kDa in size, the middle fractions
contained material of between 10 and 100 kDa in size, and the last fractions
contained material less than 50 kDa in size (Figure 3.3). Protein was eluted in
25 mM Tris, 10 mM NaCl (pH 7.4) and 0.5 ml fractions were collected. Fractions
were pooled into four groups based on the UV trace (absorption at 280 nm),
which gives an indication of when protein was eluted from the column
(Figure 3.3A), and their protein composition was visualised by SDS-PAGE
(Figure 3.3B). Pool 1 consisted of fractions 16-18, pool 2 consisted of fractions
20-22, fractions 24-27 were pooled to make a third group, and fractions 29-32
were pooled to make a fourth. There was some overlap in protein size between
these groups, however it is clear from both SDS-PAGE gels and mass
spectrometry analysis (see Appendix 1, Tables A1.2-5) that each pool is
enriched in a different set of proteins. As pool 1 contained very little material,
most of the experiments shown in this thesis have involved pools 2 to 4.

77
Figure 3.3. Fractionation of ES using the 24 ml Superose 12 column. (A) Shows UV trace (measured at 280 nm) for fractionation. (B) Shows SDS-PAGE separation of fractions (visualised by Coomassie blue staining). Labels on (A) and (B) indicate which fractions were pooled to make pools 1-4 and the numbers above the lanes in (B) indicate fraction number. The left hand lanes of the gels in (B) show the molecular weight markers in kDa.
Next, the cellular and humoral immune responses to pools 2 to 4 were
investigated during acute T. muris infection. For these studies, female C57BL/6
mice were infected with 200 T. muris eggs. Mice were sacrificed at day 20 p.i.,
and the MLNs were collected to assay for cytokine production following antigen
re-stimulation of lymphocytes, while blood was collected to measure serum
anti-parasite IgG1 and IgG2a antibody levels. Cytokine production by infection-
primed lymphocytes was compared to that of naïve lymphocytes harvested from
age-matched mice on the same day, and anti-parasite IgG1 antibody levels in
the serum of infected and naïve mice was also compared. Interestingly, these
analyses showed that the cellular and humoral arms of the immune system
responded differently to pools 2 to 4. This will be discussed in more detail in
sections 3.2.3 and 3.2.4.
B) Fractions eluted from Superose 12
250 150 100
75
50
37
25 20 15 10
kDa
250 150 100
75
50
37
25 20 15 10
kDa 16 17 18 19 20 21 22 23 24 25 26 28 29 30 31 32
Pool 4 Pool 3 Pool 2 Pool 1
A) UV trace for Superose 12 column
5 10 15 200
50
100
150
Volume (ml)
Ab
so
rban
ce a
t 280 m
AU
10 15 20 25 30 350
20
40
60
80
100
Fraction2
80
ab
so
rba
nc
e (
mA
U)
1
2
34
Ab
so
rba
nc
e
Volume (ml)
10 15 20 25 30 35
Fraction

78
3.2.3 Investigating the cellular immune response to pools 2-4 MLN cells pooled from T. muris infected (day 20 p.i.) and naïve female C57BL/6
mice were stimulated with pools 2 to 4 or unfractionated ES, and supernatant
cytokine levels were measured by cytometric bead array after 42 hours.
Samples were assayed for Th2 cytokines (IL-9 and IL-13), Th1/Th17 cytokines
(IFN-γ and IL-17A), inflammatory cytokines (IL-6 and TNF-α), and the
anti-inflammatory cytokine, IL-10.
The most noticeable difference in cytokine production in response to pools 2
to 4 was for IL-13 and IL-9. Infection-primed lymphocytes produced high levels
of these cytokines in response to stimulation with pools 3 and 4, but little in
response to pool 2. In fact, IL-13 production was 30 to 40-fold higher when
primed lymphocytes were stimulated with pools 3 and 4 compared to pool 2,
whereas IL-9 production was 100-fold higher (Figure 3.4A and B). Stimulation of
infection-primed lymphocytes with unfractionated ES resulted in similarly high
levels of these cytokines. Naïve lymphocytes released relatively less IL-13 and
IL-9 in response to unfractionated ES and pools 2-4, suggesting that these
cytokines are released in response to antigenic peptides that are encountered
specifically during T. muris infection.
Other than IL-13 and IL-9, there was little difference in the level of cytokines
produced by infection-primed lymphocytes in response to pools 2 to 4.
However, with the exception of TNF-α, much greater cytokine levels were
measured following stimulation of infection-primed lymphocytes compared to
the naïve controls (Figure 3.4). This experiment was repeated three times, and
the results shown here are representative of the three experiments. Based on
these results, pools 3 and 4 appear to stimulate the strongest Th2 response,
suggesting that these may be good sources of potential protective immunogens.
The protective properties of pools 2 to 4 were evaluated in vivo by vaccinating
male C57BL/6 mice with these sub-groups (plus adjuvant) and assessing
whether vaccination protected these mice against a subsequent low dose
infection. These data are presented in section 3.3.

79
Figure 3.4. Cellular immune response to pools 2-4 and unfractionated ES. Lymphocytes harvested from the MLNs of infected C57BL/6 mice (female, n=3, day 20 p.i.) were pooled and re-stimulated with pools 2 to 4 or unfractionated ES. MLN lymphocytes were also harvested from age-matched naïve mice (female, n=2) and stimulated in the same way. Supernatants were taken after 42 hours and levels of the following cytokines were measured by cytometric bead array: IL-6, 9, 10, 13, 17A, TNF-α and IFN-γ. Cytokine levels detected in supernatants taken from infection-primed (black) and naïve (grey) lymphocytes were plotted (A-G). This experiment was repeated 3 times and the results displayed are representative of all three repeats.
A) IL-13 production in response to stimulation with pools 2 to 4
2 3 4ES
0
200
400
600
800
Pool
IL-1
3 (
pg
/ml)
Infected
Naive
2 3 4ES
0
50
100
150
Pool
IL-9
(p
g/m
l)2 3 4
ES
0
500
1000
1500
PoolIF
N-y
(p
g/m
l)2 3 4
ES
0
200
400
600
800
Pool
IL-1
0 (
pg
/ml)
2 3 4ES
0
500
1000
1500
2000
2500
Pool
IL-6
(p
g/m
l)
2 3 4ES
0
200
400
600
800
Pool
TN
F-a
(p
g/m
l)
2 3 4ES
0
200
400
600
Pool
IL-1
7A
(p
g/m
l)
B) IL-9 production in response to stimulation with pools 2 to 4
C) IL-10 production in response to stimulation with pools 2 to 4
E) IL-17A production in response to stimulation with pools 2 to 4
G) TNF-α production in response to stimulation with pools 2 to 4
F) IL-6 production in response to stimulation with pools 2 to 4
D) IFN-Υ production in response to stimulation with pools 2 to 4

80
3.2.4 Assessment of anti-parasite IgG serum antibody response during
acute T. muris infection
The strategy used in this thesis to identify potential immunogens has focussed
on material that induces Th2 cytokine release from infection-primed
lymphocytes. However, assessing antibody response may also provide valuable
information about the immune response to ES sub-groups during acute
infection. The anti-parasite IgG serum antibody response to pools 2 to 4 was
assessed both quantitatively (by ELISA) and qualitatively (by Western blotting)
for female C57BL/6 mice at day 20 p.i.
The highest anti-parasite IgG1 serum antibody levels were measured in
response to pool 2 (Figure 3.5). This was greater than the response to
unfractionated ES, suggesting that pool 2 is enriched for proteins targeted by
the humoral response. The anti-parasite IgG1 serum antibody levels measured
in response to pools 3 and 4 were similar to those measured in response to
unfractionated ES (Figure 3.5E). No anti-parasite IgG1 antibodies were
detected in the serum of naïve mice, suggesting that there is little cross-
reactivity with other antigens that the mice may be exposed to (Figure 3.5). In
addition, no anti-parasite IgG2a antibodies were detected in the serum of
infected or naïve mice at this time point (data not shown).

81
Figure 3.5. Anti-parasite IgG1 serum antibody response to pools 2 to 4 and unfractionated ES. The sera of three infected (female, day 20 p.i.) and two naïve mice were pooled and the anti-parasite IgG1 antibody response to pools 2 to 4 was measured by ELISA. Immunograde plates were coated with 5 µg/ml of pools 2 to 4 or unfractionated ES and were incubated with serially diluted serum from infected or naïve mice (A to D, circles and squares represent infected and naïve respectively). Anti-parasite IgG1 levels were measured by adding biotinylated anti-mouse IgG1 secondary antibody, followed by streptavidin peroxidase, and lastly ABTS substrate. Optical density was measured at 405 nm. (E) Shows the O.D. for each sub-group measured at 1:40 serum dilution. Black and grey bars represent infected and naïve serum respectively. This experiment was repeated 3 times and the results displayed are representative of all three repeats.
A) Serum dose response curve for pool 2
E) Mean O.D. (1:40 serum dilution)
2 3 4ES
0.0
0.2
0.4
0.6
O.D
.
Pool
Infected
Naive
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.2
0.4
0.6
0.8
Dilution
O.D
.
Infected
Naive
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.2
0.4
0.6
0.8
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.2
0.4
0.6
0.8
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.2
0.4
0.6
0.8
Dilution
O.D
.
B) Serum dose response curve for pool 3
C) Serum dose response curve for pool 4
D) Serum dose response curve for unfractionated ES

82
Qualitative assesment of the anti-parasite IgG serum antibody response for
infected (day 20 p.i.) and naïve female C57BL/6 mice was carried out by
Western blotting after separation of ES by SDS-PAGE. Sera for three infected
mice were pooled, and sera two naïve mice were pooled. Bound antibody was
detected using an anti-mouse IgG (whole molecule) alkaline phosphatase
antibody, and proteins were visualised using BCIP and NBT. The anti-parasite
serum IgG antibody response targets a wide range of ES proteins, however, it
appears to favour larger proteins (40-250 kDa, Figure 3.6A). This is in line with
the ELISA data presented in Figure 3.5, since pool 2 contains proteins of a
similar size range (Figure 3.3B). No anti-parasite IgG antibodies were detected
in naïve sera, confirming that there is no cross-reactivity from normal mouse
sera.
Figure 3.6. Western blots showing anti-parasite IgG serum antibody response for T. muris infected and naïve C57BL/6 mice. ES+P43 was separated by SDS-PAGE and the proteins were transferred onto nitrocellulose membrane, along with 10 to 250 kDa marker proteins (left of each panel). Sera was pooled from three infected and two naïve mice. (A) Shows membrane probed with infected sera, while (B) shows membrane pooled with naïve sera. (C) Shows SDS-PAGE separation of ES (Coomassie blue staining).
3.3 Vaccination with pools 2 to 4 stimulates protective immunity
The data presented in section 3.2 demonstrated that re-stimulating
infection-primed lymphocytes with pools 3 and 4 resulted in greater levels of
IL-13 and IL-9 production than pool 2. In order to investigate whether these
protective properties translated in vivo, a vaccination protocol was developed,
250
150
100
75
50
37
25 20
15 10
kDa
A) Infected B) Naïve
250
150
100
75
50
37
25 20
15 10
kDa
250
100
75
50
37
25 20 15 10
kDa
150
C) SDS-PAGE separation of ES
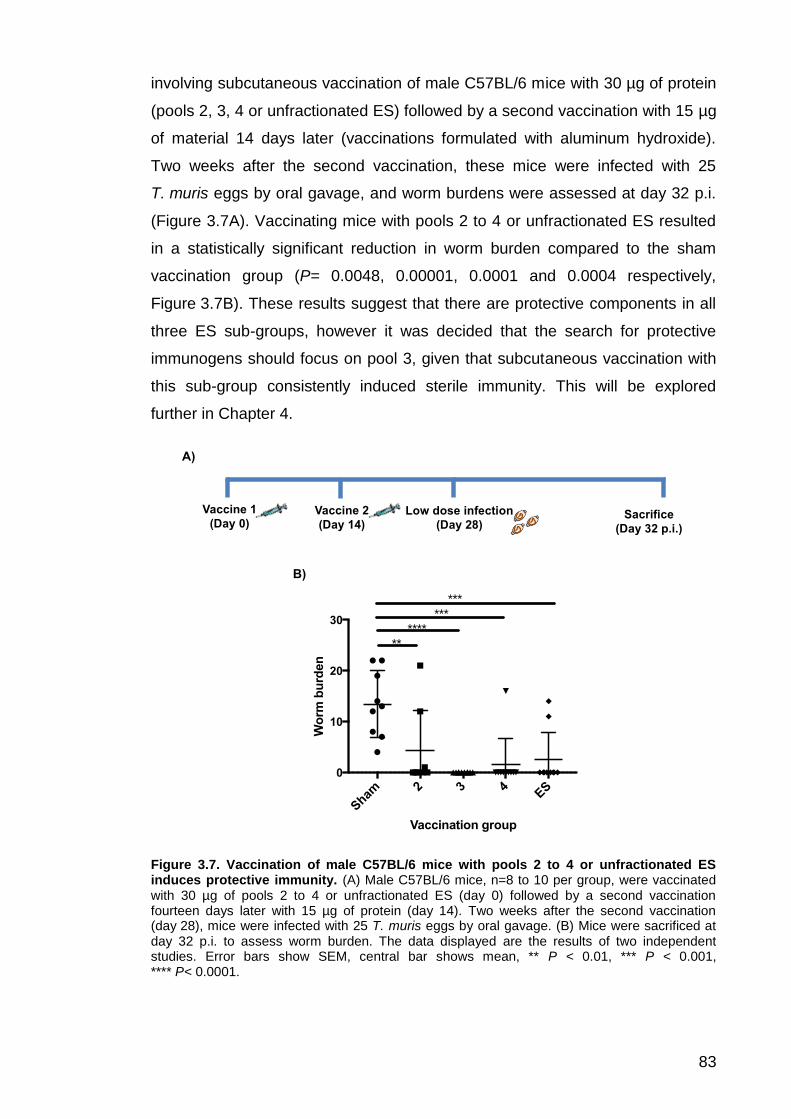
83
involving subcutaneous vaccination of male C57BL/6 mice with 30 µg of protein
(pools 2, 3, 4 or unfractionated ES) followed by a second vaccination with 15 µg
of material 14 days later (vaccinations formulated with aluminum hydroxide).
Two weeks after the second vaccination, these mice were infected with 25
T. muris eggs by oral gavage, and worm burdens were assessed at day 32 p.i.
(Figure 3.7A). Vaccinating mice with pools 2 to 4 or unfractionated ES resulted
in a statistically significant reduction in worm burden compared to the sham
vaccination group (P= 0.0048, 0.00001, 0.0001 and 0.0004 respectively,
Figure 3.7B). These results suggest that there are protective components in all
three ES sub-groups, however it was decided that the search for protective
immunogens should focus on pool 3, given that subcutaneous vaccination with
this sub-group consistently induced sterile immunity. This will be explored
further in Chapter 4.
Figure 3.7. Vaccination of male C57BL/6 mice with pools 2 to 4 or unfractionated ES induces protective immunity. (A) Male C57BL/6 mice, n=8 to 10 per group, were vaccinated with 30 µg of pools 2 to 4 or unfractionated ES (day 0) followed by a second vaccination fourteen days later with 15 µg of protein (day 14). Two weeks after the second vaccination (day 28), mice were infected with 25 T. muris eggs by oral gavage. (B) Mice were sacrificed at day 32 p.i. to assess worm burden. The data displayed are the results of two independent studies. Error bars show SEM, central bar shows mean, ** P < 0.01, *** P < 0.001, **** P< 0.0001.
A)
B)
Sham
2 3 4ES
0
10
20
30
Vaccination group
Wo
rm b
urd
en
**
****
***
***
Sacrifice
(Day 32 p.i.)
Low dose infection
(Day 28)
Vaccine 2
(Day 14)
Vaccine 1
(Day 0)

84
3.3.1 Assessment of antibody response following vaccination with pools 2
to 4 and subsequent infection
Anti-parasite IgG1 and IgG2a serum antibody levels were measured by ELISA
following vaccination (and subsequent infection) of mice with pools 2 to 4 or
unfractionated ES. Anti-parasite IgG1 serum antibody levels were highest for
the pool 2 vaccination group, however all vaccinations stimulated a statistically
significant increase in anti-parasite IgG1 serum antibody production compared
to the sham vaccination group (Figure 3.8; P= 0.0001, 0.0469, 0.0031 and
0.0004 for pool 2, 3, 4 and unfractionated ES vaccination groups respectively).
This suggested that all three ES sub-groups contain material capable of
promoting anti-parasite IgG1 antibody production. The results shown here are
from the first vaccination study, however these are representative of the two
experiments performed.

85
Figure 3.8. Anti-parasite IgG1 serum antibody response for mice vaccinated with pools 2 to 4 or unfractionated ES. Mice, n=5 per group, were vaccinated with pools 2, 3, 4 or unfractionated ES. Two weeks following the second vaccination, mice were infected with 25 T. muris eggs and were sacrificed at day 32 p.i. The anti-parasite IgG1 serum antibody response to ES+P43 was measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccination group (A-E). The symbols on each graph represent individual mice within the same vaccination group. (F) Shows the mean O.D. for each vaccination group at 1:320 serum dilution. Sham= mice vaccinated with aluminum hydroxide only. Data representative of two experiments. Error bars show SEM, * P <0.05, ** P < 0.01, *** P < 0.001, **** P < 0.0001.
A) Serum dose response curve for sham vaccination group
B) Serum dose response curve for pool 2 vaccination group
C) Serum dose response curve for pool 3 vaccination group
D) Serum dose response curve for pool 4 vaccination group
E) Serum dose response curve for ES vaccination group
F) Mean O.D. (1:320 serum dilution)
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
00.0
0.2
0.4
0.6
0.8
1.0
1.2
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Dilution
O.D
.
Sham
2 3 4ES
0.0
0.2
0.4
0.6
0.8
1.0
Vaccination group
O.D
.
****
***
***

86
Mice that were vaccinated with pools 3 or 4 had lower anti-ES IgG2a serum
antibody levels compared to the sham vaccination group, however this was not
statistically significant (Figure 3.9). There was significant variation in the anti-ES
IgG2a serum antibody levels measured between individuals within the pool 2
and unfractionated ES vaccination groups. In both cases, there were some
individuals whose anti-ES IgG2a serum antibody levels were comparable to the
sham group, while some had much higher levels (Figure 3.9). The results
shown here are from the first vaccination study, however these are
representative of the two experiments performed.

87
Figure 3.9. Anti-parasite IgG2a serum antibody levels for mice vaccinated with pools 2 to 4 or unfractionated ES. Mice, n=5 per group, were vaccinated with pools 2 to 4 or unfractionated ES. Two weeks following the vaccine boost, mice were infected with 25 T. muris eggs and were sacrificed at day 32 p.i. The anti-parasite IgG2a serum antibody response to ES+P43 was measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccination group (A-E). The symbols on each graph represent individual mice within the same vaccination group. (F) Shows the mean O.D. for each vaccination group at 1:80 serum dilution. Sham= mice vaccinated with aluminum hydroxide only. Data representative of two experiments. Sham= mice vaccinated with aluminum hydroxide only. Error bars show SEM.
A) Serum dose response curve for sham vaccination group
B) Serum dose response curve for pool 2 vaccination group
C) Serum dose response curve for pool 3 vaccination group
D) Serum dose response curve for pool 4 vaccination group
E) Serum dose response for ES vaccination group
F) Mean O.D. (1:80 serum dilution)
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
Dilution
O.D
.
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1 in
612
0
0.0
0.2
0.4
0.6
Dilution
O.D
.
Sham
2 3 4ES
0.00
0.05
0.10
0.15
Vaccination group
O.D
.

88
In order to qualitatively assess the anti-parasite serum IgG antibody response of
male C57BL/6 mice following vaccination with pools 2, 3 or 4 (and subsequent
low dose infection), ES was separated by SDS-PAGE and transferred onto
nitrocellulose membrane. The membrane was probed with pooled sera (taken at
day 32 p.i.) from mice vaccinated with aluminum hydroxide only (sham), pools
2, 3, or 4 and unfractionated ES (Figure 3.10). Bound antibody was detected
using an anti-mouse IgG (whole molecule) alkaline phosphatase antibody, and
proteins were visualised using BCIP and NBT. Each of the pooled sera bound
to a diverse array of proteins, however the IgG antibody response for mice
vaccinated with pools 2, 3 or 4 was biased towards higher molecular weight
proteins (30 to 250 kDa, Figure 3.10B-D). This result is particularly surprising for
mice vaccinated with pool 4, given that this sub-group is enriched in smaller
proteins (10-60 kDa, Figure 3.3). The anti-parasite IgG antibody response for
the sham vaccination group and mice vaccinated with unfractionated ES
targeted a wide range of proteins, including smaller sized proteins (Figure 3.10A
and E).
Figure 3.10. Western blots showing anti-ES serum antibody response for the sham, pool 2, 3, 4 or unfractionated ES vaccination groups. ES+P43 was separated by SDS-PAGE and the proteins were blotted onto nitrocellulose membrane, along with 10 to 250 kDa marker proteins (left of each panel). Membranes were probed with serum from the sham, pools 2, 3, 4 or ES vaccination groups taken at day 32 p.i. (left to right). Bound antibody was detected using an anti-mouse IgG (whole molecule) alkaline phosphatase antibody and proteins were visualised using BCIP and NBT. Sham= mice vaccinated with aluminum hydroxide only. (F) Shows SDS-PAGE separation of adult ES.
250
150
100
75
50
37
25 20
15 10
kDa
A) Sham B) Pool 2 C) Pool 3 D) Pool 4 E) ES
250
100
75
50
37
25
20
15 10
kDa
150
F) SDS-PAGE separation of ES

89
3.4 Vaccination with ES induces long-lasting protection against a
subsequent low dose infection
In order to determine whether vaccination with ES could induce longer-lasting
protection against a subsequent T. muris infection, male C57BL/6 mice were
subcutaneously vaccinated with either pool 3 or unfractionated ES formulated
with aluminum hydroxide as described in section 3.3. However, for this
experiment mice were infected with 25 T. muris eggs 50 days after the second
vaccination (see Figure 3.11A). Vaccination with either pool 3 or unfractionated
ES led to a statistically significant reduction in worm burden by day 32 p.i.
compared to the sham vaccination group (P= 0.0001 for both). Notably, sterile
immunity was induced in 4 out of 5 mice vaccinated with pool 3, and 3 out of 5
mice vaccinated with unfractionated ES. This was comparable to the positive
control group, who were infected 14 days after the second vaccination, as
described in Figure 3.7. These results suggest that vaccination with ES material
can induce immunological memory that is able to protect against future
infections.
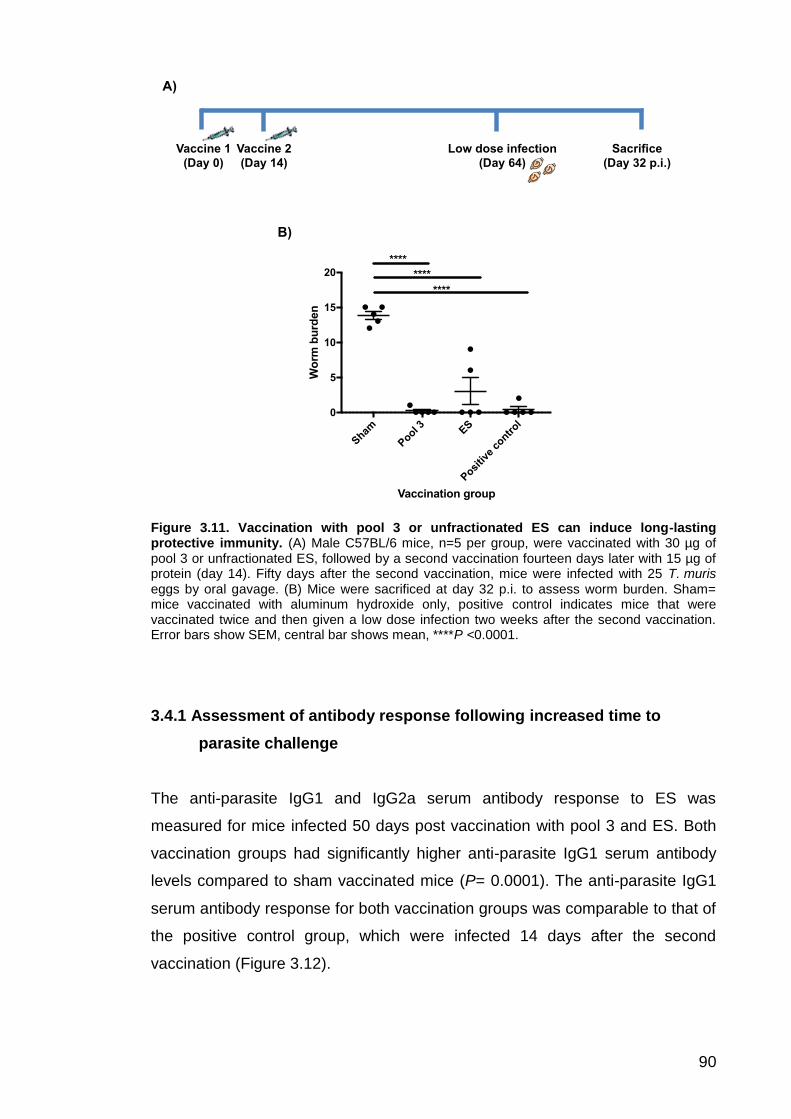
90
Figure 3.11. Vaccination with pool 3 or unfractionated ES can induce long-lasting protective immunity. (A) Male C57BL/6 mice, n=5 per group, were vaccinated with 30 µg of pool 3 or unfractionated ES, followed by a second vaccination fourteen days later with 15 µg of protein (day 14). Fifty days after the second vaccination, mice were infected with 25 T. muris eggs by oral gavage. (B) Mice were sacrificed at day 32 p.i. to assess worm burden. Sham= mice vaccinated with aluminum hydroxide only, positive control indicates mice that were vaccinated twice and then given a low dose infection two weeks after the second vaccination. Error bars show SEM, central bar shows mean, ****P <0.0001.
3.4.1 Assessment of antibody response following increased time to
parasite challenge
The anti-parasite IgG1 and IgG2a serum antibody response to ES was
measured for mice infected 50 days post vaccination with pool 3 and ES. Both
vaccination groups had significantly higher anti-parasite IgG1 serum antibody
levels compared to sham vaccinated mice (P= 0.0001). The anti-parasite IgG1
serum antibody response for both vaccination groups was comparable to that of
the positive control group, which were infected 14 days after the second
vaccination (Figure 3.12).
A)
B)
Sacrifice
(Day 32 p.i.)
Low dose infection
(Day 64)
Vaccine 2
(Day 14)
Vaccine 1
(Day 0)
Sham
Pool 3 E
S
Posi
tive
contr
ol0
5
10
15
20
Vaccination group
Wo
rm b
urd
en
****
****
****

91
Figure 3.12. Anti-parasite IgG1 serum antibody levels following long-term vaccination and subsequent infection. Mice, n=5 per group, were vaccinated with pool 3 or unfractionated ES. Fifty days following the second vaccination, mice were infected with 25 T. muris eggs and were then sacrificed at day 32 p.i. The anti-parasite IgG1 serum antibody response to ES+P43 was measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccination group (A-D). The symbols on each graph represent individual mice within the same vaccination group. (E) Shows the mean O.D. for each vaccination group at 1:40 serum dilution. Sham= mice vaccinated with aluminum hydroxide only, positive control indicates mice that were vaccinated twice and then given a low dose infection two weeks after the second vaccination. Error bars show SEM, **** P <0.0001.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
long-term pool 3 vaccination group
C) Serum dose response curve for
long-term ES vaccination group
D) Serum dose response curve for
positive control group
E) Mean O.D.(1:40 dilution)
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
Sham
Pool 3
long-t
erm
ES lo
ng-term
Posi
tive
contr
ol0.0
0.5
1.0
1.5
2.0
Vaccine group
O.D
.
****
****
****

92
Both vaccination groups had significantly lower anti-parasite IgG2a serum
antibody levels compared to the sham vaccination group (P= 0.0056 for pool 3
vaccination group, P= 0.0158 for ES vaccination group). The anti-parasite
IgG2a serum antibody response for both vaccination groups was comparable to
that of the positive control group (Figure 3.13).

93
Figure 3.13. Anti-parasite IgG2a serum antibody levels following long-term vaccination and subsequent infection. Mice, n=5 per group, were vaccinated with pool 3 or unfractionated ES. Fifty days following the second vaccination, mice were infected with 25 T. muris eggs and were then sacrificed at day 32 p.i. The anti-parasite IgG2a serum antibody response to ES+P43 was measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccination group (A-D). The symbols on each graph represent individual mice within the same vaccination group. (E) Shows the mean O.D. for each vaccination group at 1:40 serum dilution. Sham= mice vaccinated with aluminum hydroxide only, positive control indicates mice that were vaccinated twice and then given a low dose infection two weeks after the second vaccination. Error bars show SEM, * P <0.05, ** P < 0.01.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
long-term pool 3 vaccination group
C) Serum dose response curve for
long-term ES vaccination group
D) Serum dose response curve for
positive control group
E) Mean O.D. (1:40 dilution)
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
Sham
Pool 3
long-t
erm
ES lo
ng-ter
m
Posi
tive
contr
ol0.0
0.5
1.0
1.5
Vaccine group
O.D
.
*
**
**

94
3.5 Proteinase K degradation of ES abrogates its protective properties
ES was treated with 100 µg of proteinase K in order to determine whether
degradation of proteinaceous material within ES abrogated its protective
properties. Peptides were collected using a VivaSpin concentrator (10 kDa
MWCO) and separated by SDS-PAGE in order to assess degradation
(Figure 3.14A). Male C57BL/6 mice were vaccinated with 30 µg of treated ES
(protein concentration determined before degradation) or untreated ES,
followed by 15 µg of material two weeks later. These mice were infected with 25
T. muris eggs by oral gavage and worm burdens were assessed at day 32 p.i.
(Figure 3.14B). The mean worm burden for the group vaccinated with untreated
ES was significantly lower than that of the sham vaccination group (P= 0.0001),
however there was no significant difference between the mean worm burden for
mice vaccinated with treated ES compared to the sham vaccination group
(P= 0.2527).
Figure 3.14. Proteinase K treatment of ES abrogates its protective properties. The protein content of ES was degraded using 100 µg proteinase K. Peptides were collected using a Vivaspin concentrator and separated by SDS-PAGE to assess degradation (A). Male C57BL/6 mice, n=5 per group, were vaccinated with 30 µg of treated or untreated ES (protein concentration assessed prior to degradation) formulated with aluminum hydroxide, followed by a second vaccination fourteen days later with 15 µg of material plus adjuvant. Mice were infected with 25 T. muris eggs by oral gavage and were sacrificed at day 32 p.i. to assess worm burden (B). Untreated ES= native ES. Treated ES= degradation with proteinase K. Error bars show SEM, central bar shows mean. **** P < 0.0001, NS= non-significant.
A)
Sham
Untr
eate
d ES
Trea
ted E
S
0
5
10
15
20
Vaccination group
Wo
rm b
urd
en
NS
****B)
250
150
100
75
50
37
25 20 15 10
kDa Untr
eate
d ES
Treat
ed ES

95
3.6 Transfer of serum from ES vaccinated mice does not confer
resistance to unvaccinated mice
In order to determine the importance of serum antibodies in driving protective
immunity following vaccination with ES, 500 µl of heat-treated serum from ES
vaccinated male C57BL/6 mice were transferred to unvaccinated mice of the
same sex and strain. For comparison, another group received serum from
sham-vaccinated mice (vaccinated with aluminum hydroxide only). Both groups
were infected with 25 T. muris eggs by oral gavage immediately after the serum
transfer and worm burdens were assessed at day 32 p.i. There was no
significant difference between the mean worm burden of the Transfer (sham)
group, who received serum from sham vaccinated mice, and the Transfer (ES)
group, who received serum from ES vaccinated mice (Figure 3.15, P= 0.5724).
In contrast, most mice in the positive control group, who were vaccinated with
ES and infected as described in Section 3.3 had expelled worms by day 32 p.i.
(Figure 3.15, P= 0.0001).
Figure 3.15. Worm burdens following transfer of serum from sham or ES vaccinated mice. Transfer (sham) group received 500 µl of serum from sham vaccinated male C57BL/6 mice. Transfer (ES) group received 500 µl of serum from ES vaccinated mice. The negative control group received two sham vaccinations, while individuals in the positive control group were vaccinated with 30 µg of ES formulated with aluminum hydroxide, followed by 15 µg two weeks later. Mice were infected with 25 T. muris eggs by oral gavage on the same day as the serum transfer (2 weeks following the second vaccination for the control groups). Error bars show SEM, central bar shows mean. **** P < 0.0001, NS= non-significant.
Tran
sfer
(sham
)
Tran
sfer
(ES)
Neg
ativ
e co
ntrol
Posi
tive
contr
ol0
5
10
15
20
Experimental group
Wo
rm b
urd
en
****
NS
NS

96
The anti-parasite IgG1 and IgG2a serum antibody response was measured for
each experimental group as well as for the pooled transfer serum (Figure 3.16).
Groups 1 and 3 represent pooled sera from mice given sham and ES
vaccinations respectively. Groups 2 and 4 received sera from Groups 1 and 3
respectively, and were then infected with T. muris (these represent Transfer
(sham) and Transfer (ES) groups in Figure 3.15). The negative control group
(Group 5) received two sham vaccinations followed by a low dose infection,
while the positive control group (Group 6) received two ES vaccinations
followed by a low dose infection.
The serum from sham vaccinated naïve animals (Group 1) contained no
detectable anti-parasite IgG1 and IgG2a antibodies. Low levels of anti-parasite
IgG1 and high levels of anti-parasite IgG2a were measured for Group 2, who
received serum from Group 1 and were infected with T. muris. This was similar
to Group 5, who received two sham vaccinations followed by a low dose
infection (Figure 3.16).
The serum from ES vaccinated naïve mice (Group 3) contained high levels of
anti-parasite IgG1 and no detectable anti-parasite IgG2a antibodies. Group 4
(who received sera from Group 3) had low levels of anti-parasite IgG1 and high
levels of anti-parasite IgG2a. This was also similar to Group 5 (negative control
group, Figure 3.16).
The positive control group (Group 6), who received two ES vaccinations
followed by a low dose infection, had high levels of anti-parasite IgG1 and low
levels of IgG2a (Figure 3.16).

97
Figure 3.16. Anti-parasite serum IgG1 and IgG2a antibody response for serum transfer groups. (A) and (B) Show anti-parasite serum IgG1 and IgG2a response at 1:320 and 1:40 serum dilution respectively. Group 1: Sham vaccination only. Group 2: Transfer of serum from Group 1 plus infection. Group 3: ES vaccination only. Group 4: Transfer of serum from Group 3 plus infection. Group 5: Negative control – sham vaccination plus infection. Group 6: Positive control – ES vaccination plus infection.
The results in Figures 3.15 and 3.16 may suggest that the protection conferred
by vaccination with ES is not mediated by anti-parasite IgG1 alone. However
further experiments are required in order to confirm this, since low levels of
IgG1 were detected in the sera of pool 4, despite high levels of IgG1 being
transferred in the sera from group 3.
3.7 Fractionation of ES by anion exchange chromatography
In addition to fractionating ES using size exclusion chromatography, anion
exchange chromatography was investigated as a method to divide ES into
smaller sub-groups. A Mini Q column was employed and elution of proteins was
achieved with a salt gradient of 0-300 mM NaCl over 15 column volumes
followed by 300-1000 mM NaCl over 8 column volumes (Figure 3.17A). Protein
was eluted in 0.05 ml fractions and the content of these fractions was separated
by SDS-PAGE and visualised by Coomassie blue staining. Fractions were
pooled into six groups (A to F, Figure 3.17B) and these were used for
vaccinations.
A) Serum IgG1 response B) Serum IgG2a response
1 2 3 4 5 6
0.0
0.5
1.0
1.5
2.0
2.5
Experimental group
O.D
.
1 2 3 4 5 6
0.0
0.5
1.0
1.5
2.0
Experimental group
O.D
.

98
Figure 3.17. Fractionation of ES by anion exchange chromatography. ES was fractionated by anion exchange chromatography using a Mini Q column. Protein was eluted in 25 mM Tris (pH 7.4) using a salt gradient of 0-300 mM NaCl over 15 column volumes, followed by 300-1000 mM NaCl over 8 column volumes and 1 M salt for 8 column volumes. The UV trace for this fractionation process is shown in (A), while (B) shows separation of the resulting fractions by SDS-PAGE (proteins visualised by Coomassie blue staining). The left hand lanes in (B) shows the molecular weight markers in kDa.
Male C57BL/6 mice were vaccinated subcutaneously with 30 µg of A to F,
followed by 15 µg two weeks later (vaccinations formulated with aluminum
hydroxide). Mice were then infected with 25 T. muris eggs two weeks following
the second vaccination (as described in Figure 3.7) and worm burdens were
assessed at day 32 p.i. Vaccination with pools A to E prior to a low dose
challenge resulted in a significant reduction in worm burden compared to the
sham vaccination group (P=0.0001 for A to C, P=0.0063 for D and 0.0061
for E), however vaccination with F did not significantly reduce the worm burden
of subsequently infected mice (Figure 3.18).
250
150
100
75
37
25 20 15 10
50
250 150
100
75
37
25 20 15 10
50
A B C D D (cont.)
E F
0.5
0
2.3
0
3.0
0
3.1
5
3.3
0
3.4
5
3.6
0
3.9
5
4.1
0
4.2
5
3.7
5
Volume (ml)
4.4
5
4.6
0
4.7
5
5.0
5
5.3
0
5.5
0
5.7
0
6.1
0
6.2
5
7.3
5
5.8
5
A) UV trace for MiniQ fractionation of ES
(absorbance measured at 280 nm)
B) SDS-PAGE separation of MiniQ fractions
kDa kDa
0 2 4 6 80
500
1000
0
20
40
60
80
100
Volume (ml)
Ab
so
rban
ce
% N
aC
l
% NaCl
Absorance1000
800
600
400
200
[Na
Cl] m
M
0 2 4 6 80
500
1000
0
20
40
60
80
100
Volume (ml)
Ab
so
rban
ce
% N
aC
l
[NaCl]
Absorance
Ab
so
rba
nc
e
Absorbance
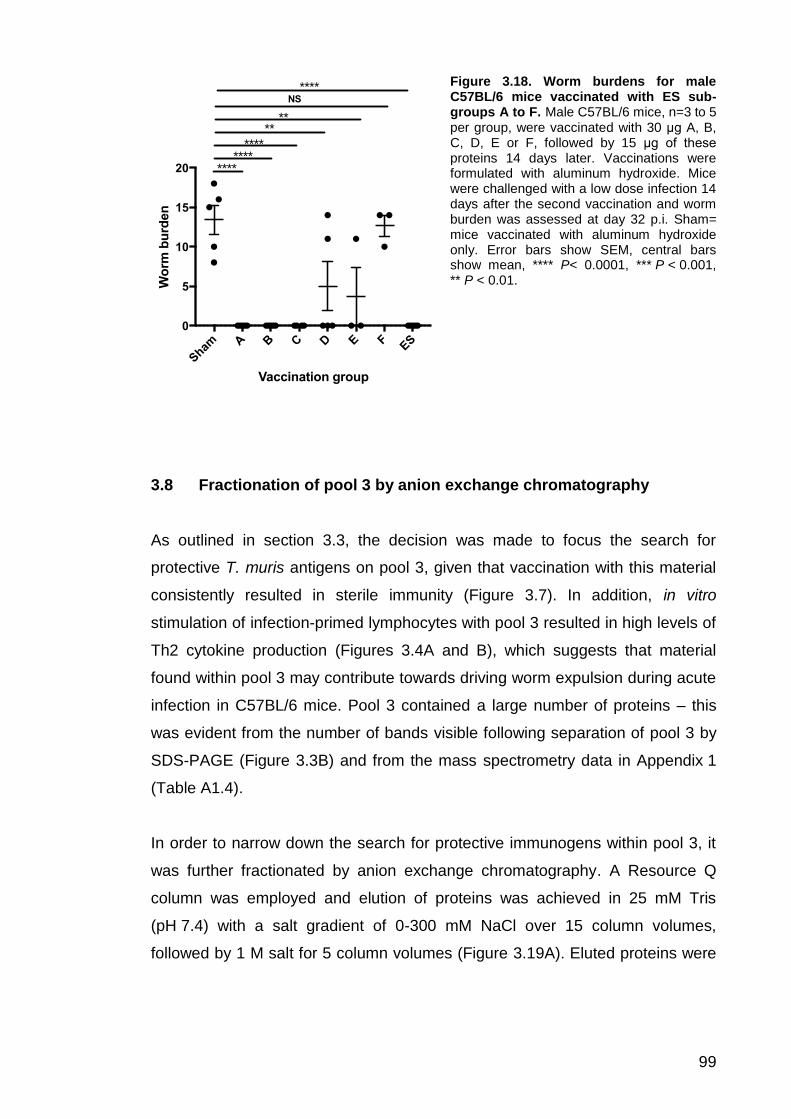
99
Figure 3.18. Worm burdens for male C57BL/6 mice vaccinated with ES sub-groups A to F. Male C57BL/6 mice, n=3 to 5 per group, were vaccinated with 30 μg A, B, C, D, E or F, followed by 15 μg of these proteins 14 days later. Vaccinations were formulated with aluminum hydroxide. Mice were challenged with a low dose infection 14 days after the second vaccination and worm burden was assessed at day 32 p.i. Sham= mice vaccinated with aluminum hydroxide only. Error bars show SEM, central bars show mean, **** P< 0.0001, *** P < 0.001, ** P < 0.01.
3.8 Fractionation of pool 3 by anion exchange chromatography
As outlined in section 3.3, the decision was made to focus the search for
protective T. muris antigens on pool 3, given that vaccination with this material
consistently resulted in sterile immunity (Figure 3.7). In addition, in vitro
stimulation of infection-primed lymphocytes with pool 3 resulted in high levels of
Th2 cytokine production (Figures 3.4A and B), which suggests that material
found within pool 3 may contribute towards driving worm expulsion during acute
infection in C57BL/6 mice. Pool 3 contained a large number of proteins – this
was evident from the number of bands visible following separation of pool 3 by
SDS-PAGE (Figure 3.3B) and from the mass spectrometry data in Appendix 1
(Table A1.4).
In order to narrow down the search for protective immunogens within pool 3, it
was further fractionated by anion exchange chromatography. A Resource Q
column was employed and elution of proteins was achieved in 25 mM Tris
(pH 7.4) with a salt gradient of 0-300 mM NaCl over 15 column volumes,
followed by 1 M salt for 5 column volumes (Figure 3.19A). Eluted proteins were
Sham
A B C D E FES
0
5
10
15
20
Vaccination group
Wo
rm b
urd
en
********
****
****
NS****

100
collected in 0.5 ml fractions. These fractions were pooled as shown in
Figure 3.19B to make four sub-groups (A to D). Vaccination studies were
carried out as described in section 3.3 (two vaccinations followed by infection
with 25 T. muris eggs). All vaccinations were formulated with aluminum
hydroxide and mice were sacrificed at day 32 p.i. to assess worm burdens.
Figure 3.19. Fractionation of pool 3 by anion exchange chromatography. Pool 3 was further fractionated by anion exchange chromatography using a Resource Q column. Protein was eluted in 25 mM Tris (pH 7.4) using a salt gradient of 0-300 mM NaCl over 15 column volumes, followed by 1 M salt for 5 column volumes. The UV trace for this fractionation process is shown in (A), while (B) shows the protein content of various fractions eluted from the column (SDS-PAGE gel visualised by Coomassie blue staining). The left hand lane in (B) shows the molecular weight markers in kDa.
250
150
100
75
37
25
20
15 10
50
A B C D
Volume (ml)
8.0
22.5
23.5
24.5
25.5
26.5
27.5
28.5
31.0
32.0
A) UV trace for fractionation of pool 3 by anion exchange chromatography
B) SDS-PAGE gel for fractionation of pool 3 by anion exchange chromatography
kDa
0 10 20 30 400
100
200
300
400
500
600
0
20
40
60
80
100
Volume (ml)
Ab
so
rban
ce
% N
aC
l
% NaCl
Absorance1000
800
600
400
200
[NaC
l] mM
0 2 4 6 80
500
1000
0
20
40
60
80
100
Volume (ml)
Ab
so
rban
ce
% N
aC
l
[NaCl]
AbsoranceAbsorbance

101
Vaccination with any of the four vaccination preparations protected mice against
a subsequent T. muris infection, with a statistically significant reduction in worm
burden compared to the sham vaccination group (P= 0.0028, 0.0080, 0.0015
and 0.0006 for A, B, C and D respectively, Figure 3.20).
Figure 3.20. Worm burdens for male C57BL/6 mice vaccinated with pool 3 sub-groups A to D. Male C57BL/6 mice, n=3 to 5 per group, were vaccinated subcutaneously with 30 μg of A, B, C, D, or pool 3, followed by 15 μg of these proteins 14 days later. All vaccinations were formulated with aluminum hydroxide. Mice were challenged with a low dose infection 14 days after the second vaccination and worm burden was assessed at day 32 p.i. Sham= mice vaccinated with aluminum hydroxide only. Error bars show SEM, central bars show mean, *** P < 0.001, ** P < 0.01.
3.9 Vaccination with L2 ES induces protective immunity
The experiments carried out thus far have involved vaccination with adult
T. muris ES. However, during acute infection, worms are expelled before they
reach patency (31), and therefore the host is not ordinarily exposed to adult ES.
Instead, larval components must be recognised by the host immune system in
order to trigger worm expulsion. To investigate this, a vaccination experiment
was carried out whereby male C57BL/6 mice were subcutaneously vaccinated
with 30 μg of ES collected from L2 larvae (L2 ES). Mice were vaccinated a
second time 14 days later (with 15 μg), and a low dose infection was
administered two weeks after. Mice that were vaccinated with L2 ES had
significantly lower worm burdens compared to the sham vaccination group
(P= 0.0001), and sterile immunity was achieved in 4 out of 5 mice. The level of
protection conferred by vaccination with L2 ES was similar to that conferred by
Sham
A B C D
Pool 3
0
10
20
30
Vaccination group
Wo
rm b
urd
en
****
**
***
***

102
vaccinating with adult ES, suggesting that both types of ES contain protective
material (Figure 3.21).
Figure 3.21. Vaccination with L2 or adult ES can induce protective immunity. Male C57BL/6 mice, n=3 to 5 per group, were vaccinated with 30 µg of L2 or adult ES formulated with aluminum hydroxide, followed by a second vaccination fourteen days later with 15 µg of protein plus adjuvant. Two weeks later, mice were infected with 25 T. muris eggs by oral gavage. Mice were sacrificed at day 32 p.i. to assess worm burden. Error bars show SEM, central bar shows mean, **** P < 0.0001.
Western blotting was carried out in order to determine whether there was much
overlap between the antibody repertoire of mice vaccinated with L2 and adult
ES. L2 and adult ES were separated by SDS-PAGE and proteins were
transferred onto nitrocellulose membrane. The membrane was probed with
pooled sera from mice vaccinated with either L2 or adult ES, and antibody
binding was detected using an anti-mouse IgG antibody. Both sera were able to
bind a range of L2 and adult ES components (Figure 3.22), suggesting that
there is significant overlap between the antibody repertoire of mice vaccinated
with L2 and adult ES.
Sham
L2 ES
Adult
ES
0
5
10
15
20
Vaccination group
Wo
rm b
urd
en
****
****

103
Figure 3.22. Western blots showing IgG antibody response to L2 and adult ES. L2 (A, B) and adult (C, D) ES was separated by SDS-PAGE and the proteins were transferred onto nitrocellulose membrane, along with 10 to 250 kDa marker proteins (left of each panel). The membrane was probed with pooled sera from mice that were vaccinated with either L2 (A, C) or adult (B, D) ES, followed by a low dose infection. After incubating with an anti-mouse IgG antibody, proteins were visualised using BCIP and NBT.
L2 and adult ES were also separated by SDS-PAGE in order to compare their
protein content. Whilst it is clear that some bands are shared between the two
ES samples, for example the bands at approximately 80, 100, 150 and
200 kDa, there are also some differences (Figure 3.23). Notably, the P43
(TMUE_s0083002300), which is amongst the most abundant adult ES proteins,
appeared to be absent from L2 ES, and this was confirmed by mass
spectrometry (Appendix 1, Table A1.6). One hundred and fourteen proteins
were identified within L2 ES by mass spectrometry (identification criteria set to
95% protein threshold, 50% peptide threshold, minimum 2 peptides identified
per sample and proteins must be identified in 2 or more samples), while 468
proteins were identified within adult ES (list generated by collating the proteins
identified in Superose 12 fractions 16 to 32, see Appendix 1, Table A1.1). A
comparison of the two samples showed that 76% of L2 proteins were identified
within adult ES.
250
150
100
75
50
37
25 20 15 10
kDa kDa
A) B)
L2 ES probed with sera from mice vaccinated with L2 (A) or adult (B) ES.
Adult ES probed with sera from mice vaccinated with L2 (C) or adult (D) ES.
C) D)
250
150
100
75
50
37
25 20 15 10
kDa
250
150
100
75
50
37
25 20 15 10
kDa
250
150
100
75
50
37
25 20 15 10

104
Figure 3.23. SDS-PAGE separation of L2 and adult ES. L2 (A) and adult (B) ES were separated by SDS-PAGE. Gels were stained with Coomassie blue to visualise proteins. The position of the P43 is indicated in blue on (B). The left hand lane on both gels shows marker proteins in kDa.
3.10 Evaluation of the AKR mouse model for vaccination studies
Previous vaccination studies have used the naturally susceptible AKR mouse
strain to assess protective immunity following vaccination with T. muris ES (19).
In order to investigate the utility of pools 2 to 4, a similar vaccination study was
carried out, whereby AKR mice were vaccinated with pools 2 to 4 or
unfractionated ES, and subsequently infected with 200 T. muris eggs. Whilst all
three ES sub-groups showed potential (sterile immunity was achieved for all
mice vaccinated with pools 2 to 4, Figure 3.24), this model did not enable
discrimination between different sub-groups in terms of immunogenicity. The
C57BL/6 (low dose) model used earlier in this chapter is arguably more
stringent, since a low dose infection naturally primes for susceptibility (31), and
strong vaccine candidates are more likely to be identified using a model that
can distinguish between more and less immunogenic candidates, and therefore,
the low dose C57BL/6 model was used going forward.
250
150
100
75
50
37
25 20
15 10
A) L2 ES
kDa
B) Adult ES
250
150
100
75
50
37
25 20
15 10
kDa
P43

105
Figure 3.24. Vaccination of male AKR mice with ES components protects against a subsequent high dose infection. Male AKR mice were vaccinated with 30 µg of pools 2 to 4 or unfractionated ES formulated with aluminum hydroxide, followed by a second vaccination 2 weeks later with 15 µg of protein plus adjuvant. Two weeks after the second vaccination, mice were given a high dose infection (200 eggs) by oral gavage. Worm burden was assessed at day 32 p.i. Sham= mice vaccinated with aluminum hydroxide only. Error bars show SEM, central bars show mean, **** P= 0.0001.
3.11 Discussion
This chapter describes experiments carried out to fractionate ES and to test the
immunogenicity of various sub-fractions in vitro and in vivo. Prior to these
studies, little was known about the immunogenic components of T. muris ES.
The first experiments to demonstrate that ES contained protective material were
carried out in the 1970s and 1980s by Jenkins and co-workers (28, 115). They
showed that vaccinating NIH mice with T. muris ES formulated with CFA lead to
earlier expulsion of a subsequent high dose infection – by day 9 p.i. 80% of
worms had been expelled. Jenkins and colleagues also attempted to divide ES
into smaller groups of proteins in order to narrow down the search for protective
antigens. They did this using size exclusion and anion exchange
chromatography, as well as isoelectric focussing, and were able to divide the
ES into different sub-groups of varying immunogenicity (28). Similar approaches
were used in this thesis, experimenting with both gel filtration and anion
exchange chromatography.
The first experiments carried out in this chapter investigated the suitability of
three different gel filtration media to divide the ES into smaller sub-groups.
Superose 12 gel filtration media appeared to be most effective, and material
Sham
2 3 4ES
0
100
200
300
400
Vaccine group
Wo
rm b
urd
en
****
********
****

106
was divided into four sub-groups (pools 1 to 4). The amount of material in pool 1
was minimal, and therefore the rest of this chapter mainly focussed on pools 2
to 4. In vitro assays showed that during acute infection, the strongest anti-
parasite serum IgG1 response was directed towards pool 2 (which contains
larger sized material), while re-stimulating infection-primed lymphocytes with
pools 3 and 4 resulted in high levels of IL-13 and IL-9 production. This suggests
that ES may be targeted differentially by the humoral and cellular immune
response during acute T. muris infection. Vaccination with any of these ES sub-
groups (pools 2 to 4) afforded mice some protection against a subsequent low
dose infection, however vaccination with pool 3 consistently induced sterile
immunity (Figure 3.7). These results suggest that the efficacy of experimental
vaccinations can be predicted by assaying for Th2 cytokine production by
infection-primed lymphocytes in vitro.
The majority of vaccination experiments carried out in this chapter have
involved subcutaneous vaccination of male C57BL/6 mice with ES material
formulated with aluminum hydroxide. The decision to administer material
subcutaneously was based on data published by Jenkins and co-workers in the
1980s, who showed that subcutaneous vaccination with ES (formulated without
adjuvant) induced better protection than intraperitoneal vaccination (70% mean
worm expulsion at day 9 p.i. following subcutaneous vaccination of NIH mice,
compared to 33% following intraperitoneal vaccination) (25). Jenkins and
colleagues showed that vaccinations formulated with aluminum hydroxide
conferred similar levels of protection as those formulated with CFA (68% worm
expulsion for aluminum hydroxide versus 77% expulsion for CFA) (25).
Aluminum hydroxide is safer than CFA and is licensed for use in
humans (193-196), which was an important consideration since the ultimate
goal is to identify vaccine candidates for the human whipworm, T. trichiura, and
therefore it was decided that aluminum hydroxide would be used for the
vaccination experiments described in this chapter.
The vaccination model used by Jenkins and colleagues is based on assessing
protection in NIH mice by looking for earlier expulsion (day 9 p.i.) of a high dose

107
infection (28). This challenge dose would ordinarily be completely expelled
before the worms reach patency (31). The approach used to investigate the
efficacy of vaccinations in this thesis was arguably more scientifically rigorous,
since it involved administering a low dose infection (25 eggs), which would
ordinarily progress to chronicity in C57BL/6 mice (197). If, following vaccination,
mice expel a low dose infection, it would suggest that the vaccination induces a
protective Th2 response in an environment where a Th1 response would
normally prevail (197). Male C57BL/6 mice were used for these vaccination
studies to ensure that expulsion was not influenced by gender differences in
cytokine production (see section 1.2.2) (43).
The suitability of the AKR mouse model for vaccination studies was also
assessed in this chapter. A high dose infection ordinarily progresses to
chronicity in this strain, as AKR mice mount a strong Th1 response (198). Data
presented here showed that vaccinating male AKR mice with unfractionated ES
or pools 2 to 4 resulted in sterile immunity for all vaccination groups
(Figure 3.24). Although this is an interesting result and largely confirmed the
data from the C57BL/6 vaccination model, these results made it difficult to
discriminate between the efficacies of different combinations of proteins. Not
only were the vaccination studies carried out using the C57BL/6 (low dose)
model more informative than the AKR (high dose) model in terms of narrowing
down the search for protective immunogens, low dose infection is more
reflective of natural challenge (48), and therefore it was decided that the
C57BL/6 (low dose) model would be utilised for the rest of the thesis.
As well as developing a stringent vaccination protocol in which to assess the
protective capacity of various ES sub-fractions, a number of important
observations were made relating to the immune response induced by
vaccination. Firstly, vaccination with pool 3 or unfractionated ES was able to
protect mice from a low dose infection administered 50 days after the second
vaccination (Figure 3.11). This suggests that vaccination with ES material can
induce longer-lasting protection. To our knowledge, this is the first study to
investigate whether immunological memory can be achieved by vaccinating with

108
T. muris ES. Future work should include further investigation of this memory
response, including characterisation of the immune components involved. For
example, Smith and colleagues have isolated memory T cells
(CD45+/CD3+/CD4+/CD11ahi/CD69+/CD62Llo/CD44hi) from Streptococcus
pneumoniae infected mice using flow cytometry, and demonstrated that these
cells secrete inflammatory cytokines upon re-stimulation with heterotypic
pneumococcal strains (199). Similar approaches could be used in our
experimental vaccination model, perhaps to determine whether the transfer of
memory T cells from vaccinated mice can protect naïve mice from a subsequent
low dose infection.
Secondly, data presented in this chapter demonstrated that treating ES with
proteinase K abrogated its protective properties, suggesting that the protective
properties of ES are likely dependent on the protein content (Figure 3.14).
Similar findings were reported by Jenkins and colleagues, who showed that
vaccinating NIH mice with peptides derived from treating ES with trypsin was
not as effective as vaccinating mice with untreated ES (28). These data support
the rationale for focussing on the protein content of ES in Chapter 4.
Thirdly, data presented here suggest that transferring serum from ES
vaccinated naïve mice does not confer protective immunity to unvaccinated
naïve mice, despite high levels of anti-parasite IgG1 antibodies being
transferred. However, since low levels of anti-parasite IgG1 were measured in
the serum of these mice following transfer, this requires further investigation in
order to confirm that antibody production is not required for vaccine-induced
immunity. Future experiments could also investigate whether the transfer of
CD4 T cells from vaccinated mice can protect against infection in naïve mice.
Nevertheless, this data provides further justification for focussing on ES material
that stimulates Th2 cytokine production, rather than using antibody-based
screening methods to identify potential vaccine candidates, as have been used
for other helminths (27, 108, 109). These approaches will be discussed in more
detail in Chapter 4.

109
Vaccination experiments were also carried out using L2 ES, demonstrating that
subcutaneous vaccination with this material induced sterile immunity in the
majority of mice. The rationale for this experiment was that larval components
must be responsible for driving worm expulsion during acute infection, since
worms are normally expelled before they reach patency (31). Adult worms
produce much larger quantities of ES than larval stages (data not shown),
making fractionation and vaccination experiments much more practical, hence
why this material was used for the majority of experiments described in this
thesis. However, it would appear that the levels of protection induced by
vaccination with adult ES were comparable to those generated by larval ES,
and comparing the protein content of larval and adult ES may inform vaccine
design.
Mass spectrometry analysis showed that there is significant overlap between
the protein content of L2 and adult ES (76% of L2 ES proteins were identified in
adult ES, Appendix 1, Table A1.6). This is reflected in the serum antibody
response of vaccinated mice. Western blot analysis showed that serum from
mice vaccinated with L2 ES contains IgG antibodies that bind a range of adult
ES proteins and vice versa. These results reflect those reported by Dixon and
colleagues, who showed that serum from AKR mice vaccinated with adult
T. muris ES could bind L3 ES, suggesting that there was significant overlap
between larval and adult ES antigens (19). Hewitson and colleagues used a
similar approach, comparing the protein content of L4 and adult H. polygyrus
ES to that of H. polygyrus eggs. Vaccination with L4 or adult ES protected mice
from a subsequent infection, however vaccination with egg material did not. The
authors used this information to inform vaccine design, by focussing on material
shared between the L4 and adult ES but absent from H. polygyrus egg
content (200).
Many more proteins were identified within adult T. muris ES compared to L2 ES
(468 versus 114 proteins). This is likely due to the process by which the list of
identified proteins was compiled, rather than a reflection of the complexity of the
samples. The list of proteins identified within adult ES was compiled by collating

110
the mass spectrometry data for multiple gel filtration fractions (Superose 12
fractions 16 to 32). This may enable the detection less abundant proteins, which
would ordinarily be below the detection threshold in unfractionated ES. Far
fewer proteins were identified following mass spectrometry analysis of
unfractionated ES (data not shown), which is why this approach was used. A
similar approach could be carried out in future to enable a more definitive list of
L2 ES proteins to be generated.
In addition to gel filtration chromatography, experiments were carried out to
fractionate ES and pool 3 by anion exchange chromatography. The resulting
fractions were pooled into 7 groups for the ES fractionation experiment (A-F)
and 4 groups for the fractionation experiment with pool 3 (A-D). All of the mice
that were vaccinated with ES pools A, B or C were protected from subsequent
infection. Vaccination with pool 3 subgroups A to D was also very effective at
protecting mice from subsequent infection, and sterile immunity was achieved
for the majority of mice. It was hoped that these fractionation/vaccination
experiments would enable us to refine the search for protective immunogens
within pool 3, or to focus on proteins shared between pool 3 and various
immunogenic sub-groups resulting from the ES anion exchange step. However,
given that almost all of the sub-fractions stimulated protective immunity, this
was not possible. As a result, the focus of Chapter 4 was to develop methods to
fractionate ES and pool 3 using gel filtration chromatography, assess the
immunogenicity of these sub-fractions in vitro using a measure of T cell cytokine
production, and identify potential immunogenic candidates within the sub-
fractions that induced the highest IL-13 and IL-9 production.
In summary, the data presented here describe the methods by which T. muris
ES can be fractionated by gel filtration and anion exchange chromatography to
produce smaller sub-groups of proteins in which to search for immunogenic
components. The immunogenicity of various ES sub-groups was assessed
using in vitro methods, and a vaccination protocol was developed to assess the
protective properties of these sub-groups in vivo. Data presented in this chapter
showed that long-lasting immunity can be induced by vaccination with T. muris

111
ES, and that proteins within ES are likely to be responsible for this protective
immunity. Pool 3, a group of proteins resulting from fractionating ES using
Superose 12 gel filtration media, will form the basis of the search for protective
immunogens described in Chapter 4.

112
Chapter 4: Identification of immunogenic candidates within T. muris ES

113
4.1 Chapter introduction
The data presented in Chapter 3 showed that the protective properties of
T. muris ES are dependent on protein material and that the efficacy of
experimental vaccinations can be predicted by assaying for Th2 cytokine
production by infection-primed lymphocytes in response to re-stimulation with
ES products. Pool 3, which arose from the pooling of fractions resulting from a
size exclusion chromatography step using Superose 12 gel filtration media,
consistently induced sterile immunity. Likewise, pool 4 induced high levels of
Th2 cytokine release from infection-primed lymphocytes, and was also very
effective at inducing protective immunity in vivo.
The following chapter describes the proteomics-based approach used to identify
immunogenic candidates within pool 3. Recombinant forms of these candidate
proteins were expressed using HEK293 cells, since other laboratories have
successfully used this expression system to synthesise helminth proteins with
comparable antigenic and/or immunogenic activity to their native forms
(Maizels, personal communication). The immunogenicity of these proteins was
assessed both in vitro and in vivo. Recombinant proteins were tested in the low
dose/C57BL/6 vaccine model described in Chapter 3, both in isolation and in
combination with the other recombinant proteins. The initial vaccines were
formulated with aluminum hydroxide, however, when these vaccines failed to
protect against subsequent infection, alternative adjuvants were investigated.
Freund’s adjuvants were selected on the basis of their ability to stimulate potent
T cell responses (135), and based on previous studies, which showed that
vaccination of NIH and AKR mice with ES formulated with Freund’s adjuvants
induced protective immunity (19, 28, 29). Montanide ISA 720 was also selected,
since it is considered a safer alternative to Freund’s adjuvant and is licenced for
use in the clinic (135), which is an important consideration, considering the end
goal is to produce a vaccine against T. trichiura that is suitable for use in
humans. Montanide ISA 720 has also been used in pre-clinical vaccines against
S. mansoni, demonstrating that it is an effective adjuvant for use in helminth
vaccines (142).

114
4.2 Identification of immunogenic candidates using a size exclusion
chromatography and proteomics approach
The approach used to identify immunogenic candidates involved two size
exclusion chromatography steps, in combination with in vitro cytokine assays
and tandem mass spectrometry of tryptic peptides. The two size exclusion
chromatography steps involved fractionating ES using Superose 12 gel filtration
media and subsequently further fractionating pool 3 (fraction numbers 24-27;
Figure 3.3) using Superdex 75 media. Infection-primed (high dose) and naïve
lymphocytes were stimulated for 42 hours with the fractions resulting from each
chromatography step and supernatant cytokine production was measured by
cytometric bead array. Tandem mass spectrometry of tryptic peptides was used
to analyse the protein content of each fraction. The total number of spectral
counts for each protein was used as a surrogate marker of protein
abundance (201). The abundance of proteins was mapped across the size
exclusion chromatography fractions, and proteins whose abundance profile
matched the pattern of IL-13 production from infection-primed lymphocytes
were considered immunogenic candidates. The immunogenic candidates
identified in both chromatography steps were investigated further.
4.2.1 Fractionating ES using Superose 12 gel filtration media
ES was fractionated using Superose 12 gel filtration media. Protein elution was
assessed by UV absorption measured at 280 nm (Figure 4.1A), and the
proteins in each fraction were separated by SDS-PAGE (gel stained with
Coomassie blue, Figure 4.1B). Infection-primed and naïve lymphocytes were
stimulated with fractions 22 to 32 and supernatant cytokine levels were
measured after 42 hours by cytometric bead array. These fractions were
selected based on data presented in Chapter 3, showing that pools 3 (fractions
24-27) and 4 (fractions 29-32) stimulated the highest levels of Th2 cytokine
release from infection-primed lymphocytes. There were two major peaks of
IL-13 production by infection-primed lymphocytes – the first was between

115
fractions 24 and 27 and the second was between fractions 29 and 31
(Figure 4.1C). IL-13 production was also high in response to fraction 28 – this is
likely due to an overlap in the immunogenic material found in the first and
second IL-13 peaks. IL-9 production by infection-primed lymphocytes was also
highest in response to fractions 25 to 28, with a second peak at fraction 30
(Figure 4.1D). There was very little IL-13 and IL-9 produced by naïve
lymphocytes in response to stimulation with these fractions, suggesting that
these fractions contain parasite-specific antigens that drive Th2 cytokine
release during acute infection. Supernatant levels of IL-10, IFN-γ, TNF-α, IL-6
and IL-17A were also measured across fractions 22 to 32, however no obvious
pattern emerged for these cytokines (Figure 4.1E-I).

116

117
Figure 4.1. Cytokine production by infection-primed and naïve lymphocytes in response to stimulation with Superose 12 fractions 22 to 32. (A) Shows UV trace (absorbance measured at 280 nm) from Superose 12 fractionation of ES. (B) Shows separation of fractions 22-32 by SDS-PAGE (gel stained with Coomassie blue). Molecular weight markers (in kDa) are shown on left hand side of gel and numbers above gel indicate fraction number. (C-I) Infection-primed and naïve lymphocytes were stimulated with these fractions and supernatant cytokine production was measured by cytometric bead array. Black and grey bars indicate cytokine production by infection-primed and naïve lymphocytes respectively. Striped bars indicate cytokine production in response to stimulation with unfractionated ES.
The protein content of fractions 22 to 32 was analysed by tandem mass
spectrometry of tryptic peptides, which identified 325 proteins across the eleven
fractions. For each fraction, a standardised amount of protein (10 μg) was used
for mass spectrometry analysis to enable comparison of protein abundance
between fractions, and total spectral count (the total number of spectra detected
for each protein) was used as a surrogate for protein abundance (201). Proteins
whose spectral count peaked around fractions 24 to 27 were considered
potential immunogenic candidates, as high levels of IL-13 were detected in
these fractions, and vaccinating mice with these fractions (in the form of pool 3)
protected mice against a subsequent T. muris infection (see Chapter 3). The
number of potential immunogenic candidates identified in this step was 63, and
these are shown in Table 4.1.

118
Spectral count
Accession number Protein Mw
(kDa) Frac 23
Frac 24
Frac 25
Frac 26
Frac 27
Frac 28
Frac 29
Frac 30
Frac 31
Frac 32
TMUE_s0015001100 Pancreatic alpha amylase 66
7 18 19 8 7
TMUE_s0005004200 Malate dehydrogenase 39
3 5 17 9 3
4
TMUE_s0256000200 Hypothetical protein 70 3 4 7 13 12 12 8 6 5 2
TMUE_s0086000700 78 kDa glucose regulated protein
73 11 11 10 12 5
2 3 5 3
TMUE_s0016004100 Serpin protein 177 3 3 7 12 2 5
3 3 2
TMUE_s0013010700 NADH dependent fumarate reductase
57
8 11 7 7
TMUE_s0055003300 Elongation factor 2 91
11 6 5
2
3
TMUE_s0084003500 CAP-domain containing protein 39 3 6 9 10 7 6
3
2
TMUE_s0058002100 Conserved hypothetical protein 26
4 9 7 6
4 3 3
TMUE_s0093000600 Protein disulfide isomerase A6 48
5 6 8 3 5 2 4 4 2
TMUE_s0003017400 Rab GDP dissociation inhibitor alpha
48
6 8 5 4
TMUE_s0016007600 Hypothetical protein 23
3 8 5 4
4
TMUE_s0036003800 Parathyroid hormone 2 receptor 45 2 6 5 7 6 4
3
TMUE_s0066001200 Ion trans 2 and Pfam-B 17708-domain containing protein
132
3 6 3 6 3 5 3 5
TMUE_s0076003900 Trans sialidase 156 3
2 6 3 3
4 4
TMUE_s0074001100 Hypothetical protein 53
4 6 3 5
2 2
TMUE_s0051004900 Sulfhydryl oxidase 1 70
6 5
2
TMUE_s0083000200 Inorganic pyrophosphatase 63
2 5 6 3
TMUE_s0009001300 Translationally controlled tumor protein
21
6 5 4
2
TMUE_s0165000700 Cysteine glycine protein 2 12
3 5 4 3
5 5 5
TMUE_s0293000700 Hypothetical protein 23
2 3 5 4 5
5 3 3
TMUE_s0004017800 Porin 41
2 5 4 3 4 3 2 2
TMUE_s0106000600 Moesin:ezrin:radixin 1 69 4 2 4 5 3 3
2
TMUE_s0085005000 Venom allergen 5 39
3 5
2
3

119
TMUE_s0146001900 Trypsin-domain containing protein
52
2 5 3 2
TMUE_s0072000900 ADP ribose pyrophosphatase, mitochondrial
29
5 2 3
TMUE_s0012011300 Neurogenic locus notch protein 2
103
2 2 4 2 2
TMUE_s0002013500 Conserved hypothetical protein 56
3 4 3
2
TMUE_s0072003400 Independent phosphoglycerate mutase
46
4 3 4
TMUE_s0098000400 Protein wos2 21
4
2
3
TMUE_s0217000400 Pfam-B 11092 and MBOAT-domain containing protein
269
4
TMUE_s0104003200 NUC194 and PI3 PI4 kinase and FATC-domain containing protein
442 3
3
TMUE_s0048003100 Vinculin 123
2 2 3
TMUE_s0037007000 Pfam-B_927_and_Beach_and_WD40-domain_containing_protein
369
3
TMUE_s0241001300 Pkinase-domain containing protein
43
3
TMUE_s0010008400 Disks large 5 208
3
TMUE_s0019001900 DUF21-domain containing protein
63
3
TMUE_s0428000200 Lactoylglutathione lyase 20
3
2
TMUE_s0132001200 Gut specific cysteine proteinase 39
3 2
TMUE_s0006005400 Autophagy protein 2 protein B 225
3
TMUE_s0326000200 PAN 1-domain containing protein
34
3
TMUE_s0071003300 TPD52 domain containing protein
17
3
TMUE_s0175001100 E3 SUMO protein ligase RanBP2
235
2
TMUE_s0189000300 Eukaryotic initiation factor 4A 50
2
TMUE_s0145001100 Pfam-B 10329 and zf-CCHC- 57
2

120
domain containing protein
TMUE_s0004005500 Trypsin-domain containing protein
33
2
TMUE_s0009011100 la protein 45
2
2
TMUE_s0017002000 rve and Pfam-B 10329 and RVT 1 and Pfam-B 2707-domain containing protein
159
2
TMUE_s0138001500 Arginine glutamic acid dipeptide repeats
148
2
TMUE_s0004000500 Gut specific cysteine proteinase 48 2
2
TMUE_s0013011900 Glyco hydro 38 and Alpha-mann mid and Glyco hydro 38C-domain containing protein
126
2
TMUE_s0037005700 Thioredoxin 18
2
2
TMUE_s0005009900
Pfam-B 1842 and Pfam-B 3141 and tRNA int end N2 and uDENN and DENN and dDENN and Pfam-B 2124-domain containing protein
231
2
TMUE_s0071006300 Pfam-B 16788-domain containing protein
123
2
TMUE_s0049001100 Peptidase S9 prolyl oligopeptidase active site
150
2
TMUE_s0011000700 Dsrm-domain containing protein
25
2
TMUE_s0062003000 Nascent polypeptide associated complex protein
22
2
TMUE_s0033004300 Probable nuclear transport factor nuclear transport factor
15
2
TMUE_s0090002300 DEAD and Helicase C and dsRNA bind-domain containing protein
74
2
TMUE_s0026000400 Alpha amylase 69
2
TMUE_s0082003700 Pfam-B 2621-domain containing protein
34
2
TMUE_s0304000500 Phenylalanine 4 hydroxylase 59
2

121
Table 4.1. List of identified proteins with peak abundance around Superose 12 fractions 24 to 27. The protein content of fractions 23 to 32 was analysed by mass spectrometry. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Proteins were sorted by spectral count in fraction 26 and those with peak abundance around fractions 24 to 27 are displayed here. Bold indicates proteins that were identified in both the Superose 12 and Superdex 75 chromatography steps. Mw= molecular weight in kDa. Frac = fraction.

122
4.2.2 Fractionation of pool 3 using Superdex 75 gel filtration media
The second size exclusion chromatography step involved fractionation of
Superose 12 fractions 24-27 using Superdex 75 gel filtration media, as these
fractions were consistent with pool 3 (Figure 3.3). The Superdex 75 and
Superose 12 gel filtration media have different molecular size resolution
capabilities (202, 203), and therefore they are likely to fractionate material
differently. Protein was eluted from the Superdex 75 column across seven
fractions (Figure 4.2A and B). Infection-primed lymphocytes were re-stimulated
with each of these fractions and supernatant cytokine production was measured
by cytometric bead array.
The pattern of IL-13 production by infection-primed lymphocytes in response to
the Superdex 75 fractions was very clear; there was a steady increase in IL-13
in response to stimulation with fractions 17 to 21. IL-13 production peaked at
fraction 21, and this was followed by a decrease in IL-13 production in response
to fractions 22 and 23 (Figure 4.2C). The pattern of IL-9 production by infection-
primed lymphocytes was very similar, although cytokine production peaked
slightly earlier, at fraction 20 (Figure 4.2D). Naïve lymphocytes produced very
little IL-13 and IL-9 in response to stimulation with fractions 17 to 23, again
demonstrating that these fractions contain parasite-specific antigens capable of
driving Th2 cytokine production during acute infection. The pattern of cytokine
production across these fractions was similar for IL-10 and IFN-γ, however,
there was no obvious pattern of cytokine production for TNF-α, IL-6 or IL-17A
(Figure 4.2).

123
17 18 19 20 21 22 23
0
500
1000
1500
2000
2500
Fraction
IL-6
(p
g/m
l)
17 18 19 20 21 22 23
0
500
1000
1500
2000
Fraction
IFN
-y (p
g/m
l)
17 18 19 20 21 22 23
0
200
400
600
800
1000
Fraction
TN
F-a
(p
g/m
l)
10 12 14-20
0
20
40
60
80
100
Volume (ml)Ab
so
rban
ce a
t 280 m
AU
250
150 100 75
50
37
25
20
15
10
17 18 19 20 21 22 23 kDa
16 18 20 22 24 26 280
20
40
60
80
Fraction
Ab
so
rbtio
n a
t 280 m
AU
17 18 19 20 21 22 23
0
100
200
300
400
500
Fraction
IL-1
3 (
pg
/ml)
17 18 19 20 21 22 23
0
50
100
150
200
Fraction
IL-9
(p
g/m
l)
17 18 19 20 21 22 23
0
200
400
600
800
Fraction
IL-1
0 (p
g/m
l)
C) IL-13 production in response to fractions 17-23
E) IL-10 production in response to fractions 17-23
D) IL-9 production in response to fractions 17-23
F) IFN-γ production in response to fractions 17-23
G) TNF-α production in response to fractions 17-23
H) IL-6 production in response to fractions 17-23
A) UV trace from Superdex 75 fractionation of pool 3
B) Protein profile of fractions 17-23

124
Figure 4.2. Cytokine production by infection-primed and naïve lymphocytes in response to Superdex 75 fractions 17 to 23. (A) Shows UV trace (absorbance measured at 280 nm) from Superdex 75 fractionation of pool 3. (B) Shows separation of fractions 17-23 by SDS-PAGE (gel stained with Coomassie blue). Molecular weight markers (in kDa) are shown on left hand side of gel and numbers above gel indicate fraction number. (C-I) Infection-primed and naïve lymphocytes were stimulated with these fractions and supernatant cytokine production was measured by cytometric bead array. Black and grey bars represent cytokine levels secreted by infection-primed and naïve lymphocytes respectively.
The protein content of fractions 17 to 23 was analysed by tandem mass
spectrometry of tryptic peptides, and 190 proteins were identified across the
seven fractions. For each fraction, a standardised amount of protein (10 μg)
was used for mass spectrometry analysis to enable comparison of protein
abundance between fractions and spectral count was used as a surrogate for
protein abundance (201). A total of 70 proteins were identified whose
abundance peaked around fractions 20 to 22, matching the peak in IL-13 and
IL-9 levels. These proteins are listed in Table 4.2.
17 18 19 20 21 22 23
0
200
400
600
800
1000
Fraction
IL-1
7A
(p
g/m
l)
I) IL-17A production in response to fractions 17-23

125
Spectral count
Accession number Protein Mw (kDa) Frac 17
Frac 18
Frac 19
Frac 20
Frac 21
Frac 22
Frac23
TMUE_s0016011400 MULE and Pfam-B 516 and WAP-domain containing protein
71
50 68 77 67 13
TMUE_s000300760 WAP type 'four disulfide core' 41 6 11 34 45 46 37 15
TMUE_s0175001500 Porin 54 13 19 39 34 41 9
TMUE_s0256000200 Hypothetical protein 70
2 5 16 27 10 7
TMUE_s0090001300 Porin 19
26 27 9
TMUE_s0269000200 Triosephosphate isomerase 27
2 11 12 19 14 10
TMUE_s0191000800 Trypsin domain containing protein 79
3 8 13 17 4
TMUE_s0256000600 Conserved hypothetical protein 48
7 17 7 4
TMUE_s0048003500 Serine protease 45
6 13 16 6 5
TMUE_s0103000900 Glutathion S transferase 23
3 14 4 2
TMUE_s0014006600 Motile sperm domain containing protein 15
7 12 9 5
TMUE_s0016004100 Serpin 177
2 3 9 12 2
TMUE_s0033003400 Trypsin domain containing protein 37
12
TMUE_s0023000300 Pfam-B 9093-domain containing protein 32
5 10 11 9 5
TMUE_s0208001600 Pathogenesis protein 1B 43 5 4 8 5 11 2
TMUE_s0003017400 Rab GDP dissociation inhibitor alpha 48
10 17 11 3 0
TMUE_s0012009900 Hypothetical protein 45 2 4 7 5 10
TMUE_s0009001300 Translationally controlled tumour protein 21
2 10 4 3
TMUE_s0013010700 NADH dependent fumarate reductase 57
11 11 10 2
TMUE_s0177000800 Heat shock protein 70 71
6 9 9 4
TMUE_s0072003400 Independent phosphoglycerate mutase 46
2 7 9 2 2
TMUE_s0078002100 Conserved hypothetical protein 41
2 9
TMUE_s0047003900 Cap domain containing protein 58
4 4 8 2
TMUE_s0006008700 Thioredoxin 22
3 8 2
TMUE_s0031002800 Hypothetical protein 17
8 2
TMUE_s0005016100 Peptidy proly cis trans isomerase FKBP4 52
2 3 8
TMUE_s0042004400 Low density lipo receptor repeat 164
2 5 7 3

126
TMUE_s0204000100 Motile sperm domain containing protein 21
4 3 7 3 2
TMUE_s0003007400 Peptidase, S1 subfamily 49
2
7 2 2
TMUE_s0023000500 Pfam-B 9093-domain containing protein 26
2 7 7 2
TMUE_s0137001000 Hypothetical protein 43
5 7 5
TMUE_s0016007600 Hypothetical protein 23
7 7 2
TMUE_s0078004700 Copper:zinc superoxide dismutase 20
7
TMUE_s0037003400 Pathogenesis protein 1B 41
2 6 6 4 2
TMUE_s0078001400 DUF290-domain containing protein 11
3 5 6
TMUE_s0010006100 Calmodulin 18
5 6 3
TMUE_s0007005500 Peroxiredoxin 2 121
2 6
TMUE_s0085003400 Motile sperm domain containing protein 20
6
TMUE_s0042005600 Motile sperm domain containing protein 24
4 5 5 3
TMUE_s002200550 Eukaryotic elongation factor 1 delta 21
3 5 5 2
TMUE_s0013012700 Motile sperm domain containing protein 14
5
TMUE_s0074002600 Alcohol dehydrogenase NADP+ A 75
5
TMUE_s0281000600 CBM 14 and TIL-domain containing protein 202
5
TMUE_s0132001200 Gut specific cysteine proteinase 39
4 4 4
TMUE_s0004002000 Chymotrypsin inhibitor 16
4
2
TMUE_s0014013200 Heat shock protein 90 81
2 4 2
TMUE_s0066002300 Hypothetical protein 16
4 4
TMUE_s0133001900 Heat shock 70 kDa protein 4 95
3 3 4
TMUE_s0328000400 Hypothetical protein 19
4 4
TMUE_s0004019100 Major sperm protein 1 14
2 4
TMUE_s0038007500 DDE Tnp IS1595 and Pfam-B 9093-domain containing protein
25
3 3
TMUE_s0003006100 Hypothetical protein 225
2 3
TMUE_s0009000200 Pathogenesis protein 1B 19
3
3
TMUE_s0428000200 Lactoylglutathione lyase 20
2 3
TMUE_s0007006700 Chymotrypsin inhibitor 15
3
TMUE_s0071006300 Pfam-B 16788-domain containing protein 123
2 3
TMUE_s0071003300 TPD52 domain containing protein 17
3

127
TMUE_s0146001900 Trypsin domain containing protein 52
3
TMUE_s0074002400 Alcohol dehydrogenase NADP+ A 37
3
TMUE_s0066001200 Ion trans 2 and Pfam-B 17708-domain containing protein
132
3
TMUE_s0045002900 Serine protease inhibitor Kazal type 4 8
2
TMUE_s0024004400 Conserved hypothetical protein 38
2
TMUE_s0003007500 Coagulation factor IX 15
2
TMUE_s0031002200 Uncharacterised transposase protein 39
2
TMUE_s0136001600
Pfam-B 18137 and Pfam-B 15771 and Pfam-B 9463 and Pfam-B 8674 and Pfam-B 2154 and Pfam-B 244 and Pfam-B 19705 and Pfam-B 3092 and ubiquitin and Pfam-B 9754-domain containing protein
63
2
TMUE_s0009009900 Protein asteroid 28
2
TMUE_s0208000600 WAP domain containing protein 25
2
TMUE_s0005009800 Aminomethyltransferase, mitochondrial 91
2
TMUE_s0024001300 TSP 1 and Reeler and Spond N and Peptidase M23 and Kunitz BPTI-domain containing protein
124
2
TMUE_s0071007100 Cathepsin F 55
2
TMUE_s0078001800 DUF290-domain containing protein 15
2
Table 4.2. List of identified proteins with peak abundance around Superdex 75 fractions 20 to 22. The protein content of fractions 17 to 23 was analysed by mass spectrometry. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Proteins were sorted by spectral count in fraction 21 and those with peak abundance around fractions 20 to 22 are displayed here. Bold text indicates proteins that were identified in both the Superose 12 and Superdex 75 chromatography steps. Mw= molecular weight in kDa. Frac = fraction.

128
4.3 Identification and synthesis of potential immunogenic candidates
Eleven potential immunogenic candidates were selected for further investigation
based on the overlap between the 63 candidates identified in the first size
exclusion chromatography step (using Superose 12 gel filtration media) and the
70 candidates identified in the second step (using Superdex 75 gel filtration
media, Figure 4.3). These proteins are listed in Table 4.3 and highlighted in bold
in Tables 4.1 and 4.2.
Figure 4.3. Strategy for selecting immunogenic candidates for further investigation. Eleven potential immunogenic candidates were identified based on the overlap between the proteins identified from the Superose 12 (step 1) and Superdex 75 (step 2) fractionation steps.
Accession number Protein Mw
(kDa)
TMUE_s0016004100 Serpin protein 41
TMUE_s0009001300 Translationally controlled tumour protein 21
TMUE_s0066001200 Ion trans 2 and Pfam-B 17708-domain containing protein
132
TMUE_s0146001900 Trypsin domain containing protein 52
TMUE_s0016007600 Hypothetical protein 23
TMUE_s0072003400 Independent phosphoglycerate mutase 46
TMUE_s0071003300 TPD52 domain containing protein 17
TMUE_s0132001200 Gut specific cysteine proteinase 39
TMUE_s0428000200 Lactoylglutathione lyase 20
TMUE_s0003017400 Rab GDP dissociation inhibitor alpha 48
TMUE_s0013010700 NADH dependent fumarate reductase 57
Table 4.3. List of potential immunogenic candidates. The list was compiled based on the overlap between the candidates identified in the two size exclusion chromatography steps (with Superose 12 and Superdex 75 gel filtration media). Proteins that were identified in L2 ES are indicated in bold text, while those with no homologue in T. trichiura are displayed in grey. Mw= molecular weight.
Step 2
70 proteins
Step 1
63 proteins 11

129
Ten of the eleven potential immunogenic candidates had homologues in
T. trichiura; the Trypsin domain containing protein was the only protein without a
direct homologue (indicated in grey, Table 4.3). Four of the eleven candidate
proteins were identified in L2 ES: Serpin, Translationally controlled tumour
protein, Ion trans 2 and Pfam-B 17708-domain containing protein and Trypsin-
domain containing protein (see Appendix 1, Table A1.6 for full list of L2 ES
proteins). These are indicated in bold in Table 4.3. The reason for investigating
which of these candidates are secreted by L2 larvae is that during acute
infection, worms are expelled before they reach patency, suggesting that
expulsion is triggered in response to larval components. The data presented in
Chapter 3 showed that vaccinating C57BL/6 mice with either L2 or adult ES
induced protective immunity. Identifying material shared between L2 and adult
ES should aid the search for immunogenic material, and the identification of
four of the candidates in L2 larval ES suggests that these proteins could be
important immunogenic components secreted by multiple life cycle stages.
Although the other candidates were not identified in L2 ES, the possibility that
these proteins are present at low levels in L2 ES (below the cut off for mass
spectrometry detection) cannot be excluded.
The cDNA for each candidate protein was synthesised and sub-cloned into a
pCepHis expression vector (see Appendix 1, Figure A1.1 for sequence). The
constructs were amplified in E. coli, purified using a commercially available kit,
and HEK293 cells were transfected with each of the constructs. Secreted
proteins were purified from the cell culture media collected from transfected
HEK293 cells by nickel affinity chromatography, since the expression vector
encodes a poly-histidine tag. Fractions eluted from the nickel affinity step were
separated by SDS-PAGE and gels were stained with Coomassie blue (left of
each panel, Figure 4.4). The fractions containing bands corresponding to the
predicted molecular weight for each protein were pooled and the proteins were
further purified using Superdex 75 gel filtration media. Protein elution was
assessed by UV absorption measured at 280 nm (middle of each panel in
Figure 4.4). The resulting fractions were also separated by SDS-PAGE (gels
stained with Coomassie blue), as shown on the right of each panel. The identity

130
of each recombinant protein was confirmed by tandem mass spectrometry of
tryptic peptides from gel digests (Appendix 2, Tables A2.1-5) and the fractions
containing the protein of interest were pooled.

131
250
150
100 75
50
37
25 20 15 10
250 150
100
75
50
37
25 20 15 10
kDa kDa
20 50 100 200 500
Conc. Imidazole (mM)
500 500 6.5
7.0
7.5
10.5
11.0
11
.5 12.0
12.5
13.0
Volume (ml)
0 5 10 15 200
500
1000
1500
2000
Volume (ml)
Ab
so
rban
ce a
t 280 n
m
Serpin
Serpin
Lactoylglutathione_lyase
Nickel affinity chromatography UV trace for Sx75 purification step Gel for Sx75 purification step
Nickel affinity chromatography UV trace for Sx75 purification step Gel for Sx75 purification step
250
150 100
75
50
37
25 20 15 10
250
150
100 75
50
37
25 20 15 10
kDa kDa 20 50 100 200 500
Conc. Imidazole (mM)
500 500 8.0
8.5
9.0
10.5
11
.0
12.0 13.0
Volume (ml)
0 5 10 15 200
1000
2000
3000
Volume (ml)
Ab
so
rban
ce a
t 280 n
mLacto
Nickel affinity chromatography UV trace for Sx75 purification step Gel for Sx75 purification step
Nickel affinity chromatography UV trace for Sx75 purification step Gel for Sx75 purification step
Translationally controlled tumour protein
TPD52 domain containing protein
250
150
100
50
37
25 20 15 10
250
150
100 75
50
37
25 20 15 10
kDa
20 50 100 200 500
Conc. Imidazole (mM)
500 500 7.5
8.0
8.5
9.0
9.5
10.0
10.5 11
.0
Volume (ml)
0 5 10 15 200
100
200
300
400
Volume (ml)
Ab
so
rban
ce a
t 280 n
m
Trans
250
150
100 75
50
37
25 20 15 10
kDa
250
150
100
75
50
37
25 20 15 10
kDa
20 50 100 200 500
Conc. Imidazole (mM)
500 500 7.5
8.0 8.5
9.5
10.0
Volume (ml)
0 5 10 150
50
100
150
200
Volume (ml)
Ab
so
rban
ce a
t 280 n
m
TPD

132
Figure 4.4. Purification of T. muris recombinant proteins. Histidine-tagged recombinant forms of the candidate proteins were expressed using HEK293 cells. These proteins were purified by nickel affinity followed by size exclusion chromatography. Each panel shows SDS-PAGE separation of nickel affinity fractions (left), UV trace from Superdex 75 purification step (middle) and SDS-PAGE separation of Superdex 75 fractions (right). Sx75= Superdex 75.
In these studies, the HEK293 cells did not express four of the immunogenic
candidate proteins (Ion trans 2 and Pfam-B 17708-domain containing protein,
Gut specific cysteine proteinase, Independent phosphoglycerate mutase and
NADH dependent fumarate reductase), despite multiple transfection attempts.
Rab GDP dissociation inhibitor alpha was only expressed at low levels, and
therefore it was not possible to test this protein in any vaccination studies,
although it did show potential in in vitro assays.
Hypothetical protein
Nickel affinity chromatography UV trace for Sx75 purification step Gel for Sx75 purification step
250
150
100
75
50
37
25 20 15 10
kDa 250 150
100
75
50
37
25 20 15
10
kDa 100 200 500
Conc. Imidazole (mM)
500 500 12.5
13.5 14.5
16.0 17.0
18.0
19.0
0 5 10 15 200
100
200
300
Volume (ml)
Ab
so
rban
ce a
t 280 n
m
Hypo Volume (ml)

133
4.4 In vitro assessment of the immunogenicity of candidate proteins
The recombinant forms of Translationally controlled tumour protein, TPD52
domain containing protein, Lactoylglutathione lyase and Rab GDP dissociation
inhibitor alpha induced IL-13 production by infection-primed lymphocytes
(Figure 4.5A), suggesting that these proteins may be recognised by
lymphocytes during acute infection and are able to induce Th2 cytokine
production. These recombinant proteins also induced IFN-γ, IL-10, IL-6, IL-17A
and TNF-α release from infection-primed lymphocytes. For the majority of these
proteins, higher levels of each cytokine were measured in the supernatants
from infection-primed lymphocytes compared to the naïve supernatants. The
exceptions were IL-13 and IL-17A production in response to recombinant
Lactoylglutathione lyase. In addition, all of the recombinant proteins induced
TNF-α release from naïve lymphocytes.

134
Figure 4.5. Cytokine release by infection-primed and naïve lymphocytes following stimulation with recombinant proteins. Infection-primed lymphocytes were re-stimulated with recombinant Translationally controlled tumour protein (Trans), TPD52 domain containing protein (TPD), Lactoylglutathione lyase (Lacto) and Rab GDP dissociation inhibitor alpha (Rab). Cells were also incubated with native pool 3 and ES proteins for comparison. Supernatant cytokine levels were measured by cytometric bead array and were compared to those of naïve lymphocytes. Black and grey bars represent cytokine levels secreted by infection-primed and naïve mice respectively.
A) IL-13 production in response to stimulation with recombinant T. muris
proteins
ES
ES
B) IFN-γ production in response to stimulation with recombinant T. muris
proteins
C) IL-10 production in response to stimulation with recombinant T. muris
proteins
D) IL-6 production in response to stimulation with recombinant T. muris
proteins
E) IL-17A production in response to stimulation with recombinant T. muris
proteins
F) TNF-α production in response to stimulation with recombinant T. muris
proteins
ES
ES
ES
ES

135
4.5 In vivo assessment of immunogenicity of candidate proteins
In order to assess the protective properties of the recombinant T. muris proteins
in vivo, a vaccination study was carried out, whereby male C57BL/6 mice were
vaccinated with each protein, either singularly (30 μg of protein, followed by a
second vaccination with 15 μg) or a multi-protein vaccination (30 μg of each of
the five recombinant proteins, 150 μg in total, followed by a second vaccination
with 75 μg of material). All vaccinations were formulated with aluminum
hydroxide and mice were infected with 25 T. muris eggs two weeks after the
second vaccination (Figure 4.6A). Worm burdens were assessed at day 32 p.i.,
and were compared to the sham vaccinated mice (vaccinated with aluminum
hydroxide diluted in Tris buffer). None of the recombinant proteins were able to
protect mice from a subsequent infection when administered as single or multi-
protein vaccinations formulated with aluminum hydroxide. In contrast, sterile
immunity was achieved for all individuals in the positive control group
(vaccinated with ES formulated with aluminum hydroxide, Figure 4.6B).

136
Figure 4.6. Vaccination with recombinant T. muris proteins formulated with aluminum hydroxide does not induce protective immunity in male C57BL/6 mice. (A) Mice, n=5 per group, were vaccinated with recombinant T. muris proteins (either singularly or a combination of all 5 proteins) formulated with aluminum hydroxide. For the single protein vaccinations, mice were vaccinated with 30 μg of protein, followed by a 15 μg boost two weeks later. For the multi-protein vaccinations, mice were vaccinated with a total of 150 μg protein (30 μg of each protein), followed by a 75 μg boost (15 μg of each protein). Mice were infected with 25 T. muris eggs 14 days following the second vaccination. (B) The mean worm burden at day 32 p.i. was compared to that of the sham vaccination group. Sham vaccinations were performed with aluminum hydroxide diluted in Tris buffer. The ‘ES’ group (positive control) represents mice that were vaccinated with 30 μg of ES, followed by a 15 μg boost two weeks later (both formulated with aluminum hydroxide). Error bars represent SEM, **** P < 0.0001. Trans= Translationally controlled tumour protein, TDP= TPD52 domain containing protein, Lacto= Lactyoglutathione lyase, Hypo= Hypothetical protein.
Sacrifice
(Day 32 p.i.)
Low dose infection
(Day 28)
Vaccine 2
(Day 14)
Vaccine 1
(Day 0)
Single protein vaccinations: 30 µg of recombinant protein
Multi-protein vaccinations: 30 µg of each protein, 150 µg in total
Single protein vaccinations: 15 µg of recombinant protein
Multi-protein vaccinations: 15 µg of each protein, 75 µg in total
A)
B)
Sham
Tran
sTP
D
Lacto
Ser
pin
Hyp
o
Multi
-pro
tein ES
0
5
10
15
20
Vaccine group
Wo
rm b
urd
en
****

137
Experiments were also carried out to determine whether formulating
recombinant proteins with other adjuvants could boost protective immunity. For
these experiments, multi-protein vaccinations were administered, consisting of
150 μg of recombinant proteins (30 μg of each of the five proteins), followed by
a second and third vaccination with 75 μg of protein, each ten days apart
(Figure 4.7A). Vaccinations were formulated with Freund’s adjuvants (CFA for
first vaccination, IFA for subsequent vaccinations, Figure 4.7B) or Montanide
ISA 720 (Figure 4.7C). These multi-protein vaccinations were unable to protect
mice against a subsequent low dose infection (Figure 4.7B and C).
Alongside each of these recombinant protein vaccinations, a group of mice
were vaccinated with 30 μg of ES formulated with the appropriate adjuvant, in
order to determine the efficacy of these adjuvants when used in combination
with a known source of immunogenic material. ES vaccinations formulated with
Freund’s adjuvants were able to induce protective immunity, albeit not sterile
immunity (Figure 4.7B). ES vaccinations formulated with Montanide ISA 720
afforded mice some protection, however these were not as effective as ES
vaccinations formulated with aluminum hydroxide (Figure 4.7C).

138
Figure 4.7. Vaccination with recombinant T. muris proteins formulated with Freund’s adjuvants or Montanide ISA 720 does not induce protective immunity in male C57BL/6 mice. (A) Mice, n=5 per group, were vaccinated with recombinant T. muris proteins formulated with Freund’s adjuvants or Montanide ISA 720. For both experiments, mice were vaccinated with 150 μg of the pooled recombinant proteins (30 μg of each protein, blue bars), followed by two further vaccinations with 75 μg of protein, each ten days apart. Mice were infected with 25 T. muris eggs by oral gavage ten days after the last vaccination, and were sacrificed at day 32 p.i. (B) and (C) show mean worm burden at day 32 p.i. for vaccinations formulated with Freund’s adjuvants and Montanide ISA 720 respectively. Sham vaccinations (grey bars) were performed with adjuvant diluted in Tris buffer, and alongside these vaccinations, a group of mice was vaccinated with 30 μg ES formulated with either Freund’s or Montanide ISA 720 (purple bars). Another group of mice was vaccinated with 30 μg ES formulated with aluminum hydroxide (alum, green bars), as a positive control. The mean worm burden at day 32 p.i. was compared to that of the sham vaccination group, **** P < 0.0001, *** P< 0.001, ** P< 0.01. Error bars indicate SEM.
Although the recombinant protein vaccinations did not induce protective
immunity, vaccination with recombinant Translationally controlled tumour
protein, TPD52 domain containing protein, Lactoglutathione lyase and Serpin all
led to significant induction of IgG1 antibodies specific for that particular protein
compared to acutely infected mice, or mice vaccinated with pool 3
(Figure 4.8A-D). This suggests that these recombinant proteins are
immunogenic, despite a lack of protective immunity. However, since a high
C) Montanide vaccination study B) Freund’s vaccination study
A)
Sham
Rec
ombin
ants
ES +
Fre
und's
ES +
alu
m
0
5
10
15
Vaccination group
Wo
rm b
urd
en
***
***
Sham
Rec
ombin
ants
ES +
Monta
nide
ES +
alu
m
0
5
10
15
20
25
Vaccination group
Wo
rm b
urd
en
****
**
Sacrifice (Day 32 p.i.)
Low dose infection (Day 30)
Vaccine 2 (Day 10)
Vaccine 1 (Day 0)
Vaccine 3 (Day 20)

139
dose infection does not generate IgG1 antibodies capable of binding these
recombinant proteins (nor does vaccination with pool 3), the conformation of the
recombinant proteins may be different to the native proteins.
Figure 4.8. Vaccination with T. muris recombinant proteins induced production of IgG1 antibodies specific for these proteins. IgG1 antibodies specific for Translationally controlled tumour protein (A), TPD52 domain containing protein (B), Lactoylglutathione lyase (C) and Serpin (D) were measured in sera from infected (high dose) mice, pool 3 vaccinated mice, or mice vaccinated with each of the recombinant proteins (single protein vaccinations formulated with aluminum hydroxide). IgG1 serum antibody levels were measured for each individual by ELISA (reading at 405 nm) and graphs show mean antibody titres at 1:40 serum dilution. Error bars show SEM, * P < 0.05, ** P < 0.01, **** P < 0.0001. Trans= Translationally controlled tumour protein, TDP= TPD52 domain containing protein, Lacto= Lactyoglutathione lyase.
A) IgG1 antibody response to
Translationally controlled tumour protein
B) IgG1 antibody response to TPD52
domain containing protein
C) IgG1 antibody response to
Lactoylglutathione lyase
D) IgG1 antibody response to Serpin
Hig
h d
ose
infe
ctio
n
Pool 3
v
acci
nated
Lac
to v
acci
nated
0.0
0.5
1.0
1.5
Serum response against Lacto
Source of serum
O.D
.
****
Hig
h d
ose in
fect
ion
Po
ol 3 v
acci
nated
Tr
ans
vacc
inat
ed
0.0
0.1
0.2
0.3
0.4
0.5
Serum response against Trans
Source of serum
O.D
.
**
Hig
h dose
infe
ctio
n
Po
ol 3
vac
cinat
e
d
TPD v
acci
nate
d
0.0
0.2
0.4
0.6
0.8
1.0
Serum response against TPD52
Source of serum
O.D
.
*
High d
ose
in
fect
ion
Pool 3
v
acci
nated
Serp
in
vac
cinat
ed
0.0
0.5
1.0
1.5
Serpin
Source of serum
O.D
.
****

140
4.6 Discussion
This chapter describes the process by which immunogenic candidates were
identified using a combination of gel filtration chromatography, in vitro
re-stimulation assays and mass spectrometry. Similar approaches were used by
Jenkins and colleagues to search for T. muris antigens (28), and by Santos and
co-workers to identify immunomodulatory material within T. trichiura
homogenate (204). Vaccine candidates for other gastrointestinal helminths have
been identified using antibody-based screening methods, whereby immune sera
(and other sources of protective antibodies) were used to probe for antigenic
material. For example, Pearson and colleagues identified a number of
S. mansoni vaccine candidates using sera from DIR individuals and naturally
resistant rhesus macaques (27). These candidates included Sm-TSP-1, which
is currently undergoing Phase I clinical trials (27). Redmond and co-workers
identified potential vaccine candidates within T. circumcincta larval secretions,
including a cathepsin F protein (Tci-CF-1), a 20 kDa protein of unknown
function (Tci-ES20), an astacin-like metalloproteinase-1 (Tci-MEP) and an
activation-associated secretory protein-1 (Tci-ASP-1) by probing with IgA
antibodies in the gastric lymph of infected sheep (205, 206). These proteins
have been used as part of an effective multivalent vaccination for sheep, where
vaccinated animals showed significant reductions in worm burden and egg
output compared to unvaccinated controls (206). Highly effective vaccines
against Taenia parasites have also been developed using antibody-based
screening methods (98). Clearly, antibody-based screening approaches have
been greatly successful for identifying vaccine candidates for these parasites,
however such an approach may not be effective for T. muris, given that worm
expulsion is not antibody mediated (39, 42, 207).
Instead of using antibodies from pre-immune animals to identify protective
antigens, the approach used in this thesis has focussed on identifying material
capable of inducing Th2 cytokine release in the context of acute infection. The
experiments carried out in this chapter have centred on material that had
previously been demonstrated to induce protective immunity both in vitro and

141
in vivo (pool 3). Eleven immunogenic candidates were identified, and ten of
these candidates had direct homologues in T. trichiura, which emphasises the
potential for the T. muris model to inform vaccine design for T. trichiura. For
most proteins, the amino acid sequence was highly conserved between
T. muris and T. trichiura, with the majority of proteins sharing between 40 and
90% sequence homology over 85 to 95% of the sequence (see Appendix 2,
Table A2.6). Five of the ten recombinants were expressed and purified without
difficulty, however four of the proteins were not expressed at all, and the Rab
GDP dissociation inhibitor alpha protein (TMUE_s0003017400) was expressed
at low levels. For those proteins lacking expression by HEK293 cells, the
transfections were repeated a total of three times, however, expression did not
improve with these attempts. The Ion trans 2 and Pfam-B 17708-domain
containing protein was very large (an estimated 132 kDa), which may explain
why it was difficult to express. The protein sequence of Ion trans 2 and Pfam-B
17708-domain containing protein (TMUE_s0066001200) was searched against
all organisms using the BLAST protein search tool, and the results revealed that
the predicted protein sequence listed in the T. muris database may have arisen
by incorrectly pairing together two proteins (see Appendix 2, Figure A2.1). The
search results revealed homology between the C-terminal portion
(approximately amino acids 780 to 1200) and ion transporter proteins of other
trichocephalid parasites (Appendix 2, Table A2.7). In future, attempts should be
carried out to express the C-terminal portion of the protein.
Another reason why expression may have failed was that the proteins were
toxic to the cells, although for the most part the cells appeared to be viable. To
rule out that these proteins were being expressed but not released from cells,
transfected cells were lysed with Ripa buffer and Western blotting was carried
out with a His-probe to determine whether the His-tagged proteins were present
within the lysis material, however no His-tagged proteins were detected (data
not shown). In future, alternative expression systems should be investigated, to
determine whether these cells are capable of expressing the four proteins.
Yeast, insect cells and tobacco plants all warrant further investigation, as these
expression systems have been used to express recombinant proteins for other

142
parasitic helminths (102, 105). For example, recombinant Na-GST-1 from the
hookworm N. americanus was expressed using Pichia pastoris, while the
S. mansoni protein, Sm14, was expressed using Saccharomyces cerevisiae
(23, 104). Spodpotera frugiperda Sf9 insect cells were used to express
N. americanus ASP-2, and tobacco plants were used to express Na-APR-1
(100, 101). Protein yield is also an important consideration when choosing a
protein expression system (208). This is of particular importance for T. muris
recombinant proteins, given that there were issues with protein yield for the Rab
GDP dissociation inhibitor alpha protein. Goud and colleagues report that 1.5 g
of Na-GST-1 were purified from 20 litres of P. pastoris culture, demonstrating
that this is an effective expression system for purifying large quantities of
recombinant helminth proteins (104). E. coli have also been used to express
recombinant T. circumcincta proteins (206), although bacterial expression is not
always suitable for expression of helminth proteins, as exemplified by
O. ostertagi (discussed below) (209).
The recombinant proteins were first tested for their ability to stimulate Th2
cytokine release from infection-primed lymphocytes in vitro. Four proteins
induced IL-13 release from infection-primed cells – these were Translationally
controlled tumour protein, TPD52 domain containing protein, Lactoylglutathione
lyase and Rab GDP dissociation inhibitor alpha. Equally high levels of IL-13
were produced in response to each recombinant protein compared to pool 3.
This is particularly striking given that there are likely to be relatively fewer T
cells specific for each recombinant protein, compared to the number of T cells
capable of recognising material within pool 3. The recombinant proteins were
then tested in vaccination studies, however none of the proteins could protect
mice from a subsequent low dose infection, when administered as single or
multi-protein vaccinations formulated with a range of adjuvants. One possibility
for the lack of protection conferred by vaccination with the recombinant proteins
was that the post-translational modification of proteins by HEK293 cells was
different to the modifications added to the native proteins. Little is known about
the post-translational modifications of Trichuris parasites, however a recent
report found that T. suis had some distinctive post-translational modifications,

143
for example glycans carrying N,N′-diacetyllactosamine (LacdiNAc) modified by
phosphorylcholine and/or fucose, as well a number of typical invertebrate
glycan modifications such as paucimannosidic and oligomannosidic N-glycans
(210, 211). Further characterisation of the glycome of Trichuris parasites is
required to enable researchers to choose the most appropriate expression
system (210).
Other gastrointestinal nematode vaccine projects have encountered similar
problems translating native antigens into an effective recombinant vaccine.
These include vaccines for H. contortus, the barber’s pole worm of sheep, and
O. ostertagi, the brown stomach worm of cattle (209). Following the partial
success of an irradiated larvae vaccine for H. contortus, work began on the
so-called ‘hidden’ gut membrane antigens (212-214). H. contortus is a blood-
feeding parasite and therefore the rationale for this vaccination approach was
based on inducing antibodies to gut membrane antigens, leading to damage of
these intestinal components upon ingestion of host blood, reduced worm
viability and parasite expulsion (209). Initial studies showed that native antigen
combinations were highly protective (213, 214). Detailed studies were then
carried out on the major components of this gut membrane preparation, an
aminopeptidase called H11 and a galactose containing glycoprotein complex
H-gal-GP (215-217). Vaccination with the native H11 protein lead to 95%
reduction in egg output and around 90% reduction in worm burden, while
vaccination with native H-gal-GP lead to 93% reduction in egg output and 72%
reduction in worm burden (215-217). These studies clearly show the therapeutic
potential of native forms of these proteins, however, recombinant forms of these
proteins failed to induce consistent protection when administered as vaccines
(218-220). Since then, a vaccine based on the native gut proteins, Barbervax,
has launched in Australia, and H11 and H-gal-GP are predicted to be the key
protective components (209). The vaccine requires purifying these components
from worms extracted from experimentally infected sheep (209). H. contortus
ES proteins are now being explored as potential vaccine candidates, in the
hope that this will identify alternative vaccine candidates that will be more
effective as recombinant protein vaccines (209).

144
The main focus for vaccine design for O. ostertagia has been the activation-
associated secretory proteases (ASPs) (221, 222). However, despite success
with native ASP proteins purified from O. ostertagi and Cooperia oncophora (a
related gastrointestinal nematode), recombinant ASPs expressed by E. coli
conferred poor protection against infection challenge (209). The authors
postulate that this could be due to inappropriate folding of the protein backbone
and a lack of protein glycosylation (209). However, technologies to engineer
P. pastoria to add complex mammalian glycosylation patterns during
recombinant protein expression are becoming available (223, 224), and this
technology could be applied to helminth recombinant protein expression,
although this would require more in depth knowledge of N- and O-linked
glycosylation patterns in helminths (209). In addition, attempts have been made
to express helminth proteins (the H11 protein of H. contortus) using the
free-living nematode, C. elegans (225). However, immunization with
recombinant H11 purified from C. elegans did not reduce egg output or worm
burden following infection challenge, and low quantities of protein were
expressed (225), which may prevent this strategy being used for other helminth
vaccines.
The first recombinant vaccinations described in this chapter were formulated
with aluminum hydroxide, since data presented in Chapter 3 showed that it is an
effective adjuvant for vaccinations involving native T. muris proteins. Aluminum
hydroxide has also been used for experimental H. polygyrus ES and ELV
vaccinations, for Phase I clinical trials for the hookworm vaccine candidates,
Na-GST-1 and Na-APR-1, and for clinical studies with the S. mansoni vaccine
candidate, rSh28GST, suggesting that it is a suitable adjuvant for inducing Th2
immunity against helminths (23, 100, 155). After finding that the recombinant
T. muris protein vaccines formulated with aluminum hydroxide failed to induce
protective immunity, alternatives were sought. Freund’s adjuvants were
selected as Jenkins and colleagues showed that ES vaccinations formulated
with CFA were highly effective (28). CFA is considered one of the most potent
adjuvants, inducing strong T and B cell responses (135), and therefore it was
decided that CFA should be investigated as an adjuvant for the recombinant

145
protein vaccines, despite it not being licenced for use in humans (135). CFA is
an oil-in-water emulsion, however, recently water-in-oil alternatives (such as
Montanide ISA 51 and ISA 720), which have better safety profiles due to the
reduced oil content, have been developed (135). Since Montanide ISA 720 is
licenced for use in humans (226) and was used in pre-clinical vaccine studies
involving schistosome cathepsin B, where vaccination reduced worm burden by
60% compared to sham vaccinated control mice (142), the potential for this
novel adjuvant to boost antigenicity when formulated with a mixture of the
T. muris recombinant proteins was investigated.
Montanide ISA 720 is thought to exert its adjuvant effects through slowly
releasing antigen over time, aiding diffusion of antigen to draining lymph nodes
and recruiting antigen presenting cells (227). Montanide ISA 720 has been used
in several phase I clinical trials for malaria vaccine candidates, which have
emphasised the safety and immunogenicity of this adjuvant, inducing
anti-recombinant IgG1 and IgG3 antibodies in healthy volunteers with no
previous exposure to Plasmodium parasites (139-141, 226, 228). Montanide
ISA 720 has also been used as an adjuvant for Phase I and pre-clinical
Epstein-Barr, hepatitis C and simian immunodeficiency virus (SIV) peptide
vaccinations (136-138). Interestingly, four of the adults who volunteered for the
Epstein-Barr Phase I trial later encountered the virus and seroconverted
asymptomatically, while the SIV vaccination protected a third of rhesus
macaques from a subsequent SIV infection (136, 138).
These studies demonstrate that Montanide ISA 720 is a safe and effective
adjuvant in the context of these infections, however when this adjuvant was
formulated with T. muris ES, it did not perform as well as aluminum hydroxide or
Freund’s adjuvants (Figure 4.7). Despite this, ES vaccinations formulated with
Montanide ISA 720 induced a robust anti-parasite IgG1 antibody response, as
well as a strong anti-parasite IgG2a antibody response (Appendix 2, Figures
A2.2 and A2.3). However, the multi-protein recombinant vaccinations
formulated with either Montanide ISA 720 or Freund’s adjuvants were unable to
protect mice from a subsequent T. muris infection (Figure 4.7).

146
The lack of protective immunity following vaccination with recombinant T. muris
antigenic candidates was surprising given that some of the proteins induced
IL-13 production from infection-primed lymphocytes in vitro, which would
suggest that these proteins are capable of inducing Th2 cytokine production in
the context of an acute infection. Vaccination did, however, induce production of
IgG1 antibodies specific for each of the recombinant proteins, which further
suggesting that these proteins are antigenic. These studies emphasise the
difference between antigenicity and protection. Clearly, the recombinant
proteins are antigenic, as they induce Th2 cytokine release in the context of
acute infection, however administration of these recombinant proteins in
vaccines did not protect mice against subsequent infection.
The anti-parasite IgG antibody response is mainly skewed towards IgG2a
following vaccination with the recombinant proteins (Appendix 2, Figure A2.4 to
A2.7), and therefore it is surprising that IgG1 antibodies were generated against
each of the recombinant proteins following vaccination, particularly as IgG1
class switching is largely regulated by a Th2 cytokine environment (75). Little or
no anti-recombinant IgG1 antibodies were measured in the serum of infected
(high dose) or pool 3 vaccinated mice, which may suggest that the conformation
of the recombinant proteins is different to that of the native proteins. However,
conformation is arguably less important for stimulating T cells, as antigen
presenting cells present peptides to T cells (229). Low levels of anti-
recombinant IgG2a antibodies were detected in the sera of recombinant-
vaccinated animals, and could also be detected (in some cases) in the sera of
infected (high dose) and pool 3 vaccinated mice (Appendix 2, Figure A2.8).
Future experiments should investigate whether sera from recombinant vaccine
recipients (prior to infection) recognises native parasite antigen, as this may
shed light on whether the recombinant proteins are folded correctly.
Antigenic homologues of Gut specific cysteine proteinase and Independent
phosphoglycerate mutase, both of which HEK293 cells were unable to express,
have been identified in other helminths. Vermeire and colleagues showed that
treating A. ceylanicum infected hamsters orally with cysteine proteinase

147
inhibitors reduced worm burdens by over 90%, while an older study showed that
vaccinating dogs with a recombinant A. caninum cysteine protease induced IgG
antibodies capable of binding to (and presumably neutralising) native cysteine
proteases in the gut of worms (230, 231). Little is known about the feeding
processes of Trichuris parasites (33), however gut cysteine proteinases may be
involved in the breakdown of nutrients by the worm, and therefore targeting
these may reduce the viability of the parasite.
The immunogenic potential of B. malayi Independent phosphoglycerate mutase
is highlighted by vaccination and RNA interference experiments (232, 233).
Vaccination with recombinant B. malayi Independent phosphoglycerate mutase
protected BALB/c mice from a subsequent infection, with a 58% reduction in
worm burden, while RNA silencing of Independent phosphoglycerate mutase in
female adult worms resulted in a 90% decrease in worm motility and an 80%
reduction in the number of microfilariae released (232, 233). Furthermore, only
55% of L3 larvae treated with Independent phosphoglycerate mutase-specific
siRNA survived to adulthood, suggesting that this protein plays an important
role across several life cycle stages (233).
In addition to investigating alternative expression systems in order to identify a
suitable platform for expression of those antigenic candidates which HEK293
cells were unable to express, another round of antigen selection could be
carried out in order to identify other potential candidates. For example pool 4
could be investigated further, since this material was also effective at
stimulating protective immunity as shown in Chapter 3. Pool 4 arose from
pooling Superdex 12 fractions 29 to 32, which is where the second peak in
IL-13 production was observed (Figure 4.1C), and these fractions also induce
IL-9 production from infection-primed lymphocytes (Figure 4.1D). It may also be
advantageous to investigate Superose 12 fraction 28 as this stimulated high
levels of IL-9 and IL-13 production from infection-primed lymphocytes, however
a larger amount of starting material (ES) would be required to do so. Another
potential source of antigens are T. muris ELVs – this will be discussed in more
detail in Chapter 5.

148
Chapter 5: T. muris ELVs as a source of immunogenic material

149
5.1 Chapter introduction
ELVs have been isolated from the secretions of several parasitic nematodes,
including Trichuris species, H. polygyrus and B. malayi (154, 165, 179, 180).
There are numerous reports describing the ability of parasite-derived ELVs to
stimulate and/or modulate host immunity (165, 169, 173). In addition, recent
studies have described the potential for helminth-derived ELVs as vaccine
candidates (155, 156). The majority of pre-clinical helminth vaccinations are
based on material identified within parasite homogenates or the soluble portion
of parasite secretions (27, 28, 108, 109, 115), while the potential for parasite-
derived ELVs as a source of protective antigens remains relatively unexplored.
This chapter aims to address this, showing for the first time that vaccination with
T. muris ELVs can protect mice against a subsequent low dose infection in the
absence of adjuvant.
5.2 ELVs can be isolated from T. muris ES by ultracentrifugation
ELVs were isolated from adult T. muris ES by ultracentrifugation at 100,000 g
for 2 hours. Pelleted material was viewed by transmission electron microscopy,
and a heterogeneous population of cup-shaped vesicles, approximately
30-100 nm diameter, was observed (Figure 5.1).
Figure 5.1. Visualisation of ELVs isolated from T. muris ES. ELVs were isolated from T. muris ES by ultracentrifugation at 100,000 g. The pelleted material was transferred to formvar-carbon-coated EM grids and negatively stained with 2% uranyl acetate for TEM analysis. Images displayed are representative of three preps. Scale bars represent 100 nm.

150
The size range of a typical T. muris ELV sample as measured by DLS is shown
in Figure 5.2. The raw scattering intensity as defined by the z-average intensity
plot showed two major peaks below 1 µM (those above this value are deemed
to be aggregates or dust particles) (160). The major scattering peak was at
~ 200 nm in diameter with a smaller intensity peak at ~ 40 nm (Figure 5.2A).
De-convolution to a distribution by number, which is a direct comparison of the
number of particles of each peak distribution according to Mei Theory (186),
showed that the majority of vesicles (82%) are between 37 and 60 nm in
diameter (Figure 5.2B). The raw correlation data showed a smooth single
exponential decay function (Figure 5.2C), indicating that ELVs were evenly
dispersed within the sample with low overall polydispersity (186).
Figure 5.2. Size range of a typical T. muris ELV sample. DLS was used to measure the diameter of particles within ELV samples. (A) Shows the z-average intensity plot, (B) Shows size distribution (diameter) by number, and (C) shows raw correlation data.

151
5.3 Mass spectrometry analysis shows that T. muris ELVs contain
typical exosome markers and are enriched in proteins lacking a
signal peptide
A number of known of exosome markers were identified within T. muris ELV
samples (Table 5.1). These include tetraspanins (tetraspanin 9 and TSP-1
domain containing protein), heat shock proteins, enolase, Rab proteins and
apoptosis linked gene 2 interacting protein X 1 (Alix) (157, 165). These data
strongly suggest that the vesicles isolated by ultracentrifugation of adult ES are
exosomes.
Table 5.1. List of exosome markers identified in T. muris ELV samples. The protein content of T. muris ELVs was analysed by mass spectrometry. Table shows known exosome markers identified within T. muris ELV samples. Mw= molecular weight. No. of peptides= number of unique peptides identified in each ELV sample (samples 1-3, criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified).
Comparison of the protein content of T. muris ES and ELVs revealed that 77%
of ELV proteins were also identified within ES depleted of ELVs, and 65% of
these lack a signal peptide (Table 5.2, see Appendix 3, Table A3.1 for full list of
ELV proteins). This suggests that ELVs may be an important mechanism by
which proteins lacking a signal peptide are released by parasites into the
external environment.
No. of peptides
Accession Number Protein Mw
(kDa) Sample
1 Sample
2 Sample
3
TMUE_s0037005100 Tetraspanin 9 43 0 5 4
TMUE_s0070003500 TSP-1 domain containing protein
46 3 5 3
TMUE_s0177000800 Heat shock protein 70 71 4 9 5
TMUE_s0014013200 Heat shock protein 90 81 2 6 2
TMUE_s0203001300 Small heat shock protein
16 0 2 6
TMUE_s0102000900 Enolase 48 3 5 2
TMUE_s0163002000 Ras protein Rab 11B 31 0 2 2
TMUE_s0078002300 Apoptosis linked gene 2 interacting protein X 1 (Alix)
122 0 2 0

152
Accession Number Protein Mw
(kDa)
Signal peptide
(Y/N)
TMUE_s0245000500 VWD and Vitellogenin N and DUF1943-domain containing protein
190 Y
TMUE_s0134000500 CAP domain containing protein 32 Y
TMUE_s0049005700 Protein crumbs 382 N
TMUE_s0004007100 Trypsin domain containing protein 107 Y
TMUE_s0052001100 Hypothetical protein 38 Y
TMUE_s0093001800 Vacuolar protein sorting associated protein 52 175 N
TMUE_s0175000100 Pfam-B 9093 domain containing protein 34 N
TMUE_s0003000700 Histone H4 11 N
TMUE_s0029005700 Actin 42 N
TMUE_s0071005500 Angiotensin converting enzyme 153 N
TMUE_s0201000900 Pfam-B 9093 domain containing protein 39 Y
TMUE_s0028000200 Phosphoenolpyruvate carboxykinase GTP 80 N
TMUE_s0033001500 CAP domain containing protein 40 Y
TMUE_s0175000200 Pfam-B 9093 domain containing protein 37 Y
TMUE_s0091002200 Fasciclin domain containing protein 81 Y
TMUE_s0177000800 Heat shock protein 70 71 N
TMUE_s0005010900 Neurogenic locus notch protein 367 Y
TMUE_s0003006100 Hypothetical protein 225 Y
TMUE_s0033006300 Hypothetical protein 40 Y
TMUE_s0094001000 Glyceraldehyde 3 phosphate dehydrogenase 41 N
TMUE_s0004006800 Trypsin domain containing protein 129 Y
TMUE_s0042005600 Motile sperm domain containing protein 24 N
TMUE_s0173000900 Delta protein 4 54 N
TMUE_s0002015700 Vitellogenin N and VWD and C8 and DUF1943 domain containing protein
354 Y
TMUE_s0001012400 Phosphoprotein phosphatase 1 86 N
TMUE_s0003006600 Pfam B 13663 domain containing protein 164 Y
TMUE_s0147001700 Hypothetical protein 15 Y
TMUE_s0302000300 Fructose bisphosphate aldolase class I 44 N
TMUE_s0022011000 Hypothetical protein 21 N
TMUE_s0119001000 Membrane metallo endopeptidase 1 like protein 173 N
TMUE_s0001012300 Hypothetical protein 39 Y
TMUE_s0062004700 Ubiquitin domain containing protein 95 N
TMUE_s0049001500 Trypsin domain containing protein 139 N
TMUE_s0037005100 Tetraspanin 9 43 N
TMUE_s0117003000 Eukaryotic translation elongation factor 1A 49 N
TMUE_s0014013200 Heat shock protein 90 81 N
TMUE_s0027006600 ASP domain containing protein 44 N
TMUE_s0102000900 Enolase 48 N
TMUE_s0106000600 Moesin:ezrin:radixin 1 69 N
TMUE_s0019006300 Hypothetical protein 39 Y

153
TMUE_s0008014600 Hypothetical protein 34 N
TMUE_s0157001800 Trans 2 enoyl coenzyme A reductase 32 N
TMUE_s0053003800 Trypsin domain containing protein 33 Y
TMUE_s0022000400 Na+,K+ ATPase alpha subunit 1 118 N
TMUE_s0037003800 NADP dependent malic enzyme, mitochondrial 71 N
TMUE_s0327000100 Pfam-B 9093 domain containing protein 39 Y
TMUE_s0042008400 Peptidase M2 domain containing protein 40 N
TMUE_s0070003500 TSP-1 domain containing protein 46 N
TMUE_s0024002900 Hypothetical protein 32 Y
TMUE_s0049001600 Trypsin domain containing protein 215 N
TMUE_s0120000600 Tubulin alpha chain 50 N
TMUE_s0028001200 VAB 10a protein 829 Y
TMUE_s0060000200 Prominin domain containing protein 76 Y
TMUE_s0117002800 Trypsin and CUB domain containing protein 71 N
TMUE_s0201000800 Conserved hypothetical protein 51 N
TMUE_s0191000800 Trypsin domain containing protein 79 Y
TMUE_s0005001100 Kunitz protease inhibitor 25 Y
TMUE_s0006000700 Solute carrier family 2, facilitated glucose 62 N
TMUE_s0281000600 CBM 14 and TIL domain containing protein 202 N
TMUE_s0116002200 Ubiquitin associated and SH3 41 N
TMUE_s0064002700 Hypothetical protein 27 N
TMUE_s0037003400 Pathogenesis protein 1B 41 N
TMUE_s0189001400 Neurogenic locus notch protein 54 Y
TMUE_s0012010600 EGF domain containing protein 78 N
TMUE_s0014006600 Motile sperm domain containing protein 15 Y
TMUE_s0086000700 78 kDa glucose regulated protein 73 Y
TMUE_s0009007500 14-3-3 protein 28 N
TMUE_s0013012700 Motile sperm domain containing protein 14 N
TMUE_s0033006400 CAP domain containing protein 35 Y
TMUE_s0096005200 Ubiquitin associated and SH3 protein 39 N
TMUE_s0007005500 Peroxiredoxin 2 121 N
TMUE_s0001022900 Conserved hypothetical protein 40 N
TMUE_s0089001100 Conserved hypothetical protein 54 Y
TMUE_s0006008700 Thioredoxin 22 Y
TMUE_s0022008100 BTB domain containing protein 41 N
TMUE_s0038006700 32 kDa beta galactoside binding lectin 39 N
TMUE_s0058002600 Hypothetical protein 25 N
TMUE_s0030008500 Conserved hypothetical protein 36 N
TMUE_s0005011400 Peptidyl prolyl cis trans isomerase 7 23 N
TMUE_s0023004600 Pfam-B 9093 domain containing protein 27 N
TMUE_s0081001900 Protein disulfide isomerase 55 Y
TMUE_s0015006100 Peptidase M8 domain containing protein 50 N
TMUE_s0058002100 Conserved hypothetical protein 26 N
TMUE_s0122001100 Pol poly protein 43 N

154
TMUE_s0002004000 Pfam-B 18698 and PLAT and DCX domain containing protein
118 N
TMUE_s0291000100 Prominin domain containing protein 36 N
TMUE_s0012010900 Neurogenic locus notch protein 43 Y
TMUE_s0131003400 Hypothetical protein 23 N
TMUE_s0005004200 Malate dehydrogenase 39 N
TMUE_s0036001700 Conserved hypothetical protein 37 N
TMUE_s0104001300 Hypothetical protein 10 N
TMUE_s0119002100 Galectin 31 N
TMUE_s0004012300 LIM domain containing protein 24 N
TMUE_s0041006400 Hypothetical protein 33 Y
TMUE_s0034006900 T complex protein 1 subunit beta 58 N
TMUE_s0172001800 CH domain containing protein 41 N
Table 5.2. List of shared ELV and adult ES proteins. A comparison of the proteins identified within T. muris ELVs and ES (depleted on ELVs) was carried out. Table shows list of shared proteins. Table also indicates whether each protein has a signal peptide (Y/N = yes and no respectively). Mw = molecular weight in kDa.
5.4 Exosomes are able to fuse with colonic epithelial cells in vitro
Marcilla and colleagues demonstrated that E. caproni ELVs were able to fuse
with rat intestinal epithelial cells in vitro, suggesting that these ELVs could
deliver protective antigens and/or immunomodulatory material to host cells
in vivo (152). T. muris is in constant contact with the caecal epithelium during
infection (35), and therefore the potential for T. muris ELVs to fuse with
intestinal epithelial cells in vitro was investigated. This involved labeling ELVs
with a red fluorescent dye, PKH26, which integrates into the phospholipid
bilayer of vesicles (234). The fluorescently labeled ELVs were then incubated
with HT-29 cells (a human colonic epithelial cell line) for 4 hours and vesicle
uptake was visualised by confocal microscopy. Fluorescently labeled vesicles
were observed inside HT-29 cells (Figure 5.3), suggesting that uptake of
T. muris ELVs by caecal epithelial cells may be possible during infection.

155
Figure 5.3. Uptake of PKH26 labeled ELVs by colonic epithelial (HT-29) cells. +Exo 1 and 2 show representative areas of the HT-29 monolayer after incubation with fluorescently labeled ELVs, while control image was taken after incubation with the fluorescent dye alone. Exo 1 and Exo 2 refer to two separate experiments. Left of each panel shows bright field Z-stack of HT-29 monolayer after addition of PKH26 labeled vesicles, middle of each panel shows fluorescence microscopy images (543 nm excitation), right of each panel show bright field and fluorescence channels merged. Scale bars represent 10 μm.
Brightfield 543 nm excitation Merge
+E
xo
1
Co
ntr
ol
+E
xo
2

156
5.5 Vaccination with T. muris ELVs can induce protective immunity and
protection is dependent on intact vesicles
In order to investigate whether T. muris ELVs contain antigenic material capable
of stimulating protective immunity, male C57BL/6 mice were subcutaneously
vaccinated with 3 μg of ELV material, followed by 1.5 μg of material two weeks
later (these vaccinations were formulated without adjuvant). Mice were infected
with 25 T. muris eggs by oral gavage and worm burden was assessed at day
32 p.i. Vaccination with T. muris ELVs lead to a statistically significant reduction
in worm burden compared to the sham vaccination group (vaccinated with PBS
only, P= 0.0001, Figure 5.4). However, vaccination did not induce sterile
immunity for all individuals. The data shown in Figure 5.4 is the result of
combining three individual vaccination studies. For each study, there were a
number of mice that expelled worms following vaccination, some individuals
who developed chronic infection, and one or two mice where some, but not all,
worms were expelled. As a result, the protection afforded by ELV vaccination
was more variable than that induced by vaccination with 30 μg ES formulated
with aluminum hydroxide, which was used as a positive control (Figure 5.4).
The mean worm burden for mice vaccinated with lysed ELVs was similar to that
of the sham vaccination group (P=0.0754, Figure 5.4), suggesting that the
protective properties of T. muris ELVs are dependent on intact vesicles.
Figure 5.4. Vaccination with T. muris ELVs induces protective immunity. Male C57BL/6 mice, n= 10 to 15 per group, were subcutaneously vaccinated with 3 μg of ELVs or lysed ELVs, followed by 1.5 μg of material 14 days later. Mice were infected with 25 T. muris eggs by oral gavage and sacrificed at day 32 p.i. to assess worm burden. The mean worm burden of ELV (and lysed ELV) vaccinated mice was compared to the sham vaccination group (vaccinated with PBS only). As a positive control, mice were vaccinated with 30 μg followed by 15 μg of ES (formulated with aluminum hydroxide). The results displayed are a combination of three independent experiments. Error bars indicate SEM, central bar shows
mean. **** P= 0.0001, NS= non-significant.
Sham
ELV
s
Lyse
d ELV
sES
0
5
10
15
20
Vaccination group
Wo
rm b
urd
en
****
****
NS

157
5.6 Vaccination with ELVs boosts IgG1 serum antibody response to
soluble ES components
As discussed previously, anti-parasite IgG1 and IgG2a/c serum antibodies are
often used as surrogate markers of resistance/chronicity during T. muris
infection (29). The serum IgG1 and IgG2a antibody response against ES
depleted of ELVs was measured for each vaccination group. Significantly higher
IgG1 antibody levels (which correlate with resistance) were measured for the
ELV vaccination group compared to the sham vaccination group (P= 0.0001,
Figure 3A). High levels of anti-parasite IgG2a/c, which correlates with chronicity,
were also measured for the ELV vaccination group (Figure 3B), which may
suggest that ELV vaccinated mice mount a mixed Th1/Th2 response. High
levels of anti-parasite IgG1 (Figure 3A) and low levels of anti-parasite IgG2a
(Figure 3B) were detected for the ES vaccination group, confirming that
successful vaccination stimulates Th2 immunity, while high levels of anti-
parasite IgG2a antibodies were measured for the sham vaccination group,
confirming that low dose infection naturally primes for chronicity (Figure 3B).
Figure 5.5. IgG1 and IgG2a serum antibody response against ES following vaccination with ELVs. The IgG1 (A) and IgG2a (B) serum antibody response targeting ES depleted of ELVs was measured by ELISA. The mean O.D. value (reading at 405 nm) for each vaccination group (sham, ELV or ES vaccinated mice, 10 per group) is shown at 1:320 (IgG1) and 1:40 (IgG2a) serum dilution. Error bars show SEM, **** P< 0.0001, *** P< 0.001, NS= non-significant.
A) IgG1 to response to ES depleted of ELVs B) IgG2a response to ES depleted of ELVs
Sham
ELV
sES
0.0
0.5
1.0
1.5
2.0
Vaccination group
O.D
.
****
****
Sham
ELV
sES
0.0
0.5
1.0
1.5
2.0
Vaccination group
O.D
.
NS
***

158
5.7 Identification of ELV components targeted by serum IgG antibodies
following vaccination
Western blotting was performed to investigate which ELV and ES components
are bound by serum IgG antibodies following vaccination of mice with PBS
(sham), ELVs or adult ES and subsequent T. muris infection (Figure 3A-C).
Infection alone does not generate IgG antibodies against ELV material
(Figure 5.6A), however vaccination with ELVs primes for IgG antibodies that
target a range of ELV components between 50 and 200 kDa in size (indicated
by asterisks on Figure 5.6B). Sera collected from the ES vaccination group
contained IgG antibodies that target 80 and 100 kDa ELV components
(indicated by asterisks on Figure 5.6C). Sera taken from all three groups also
recognised a wide range of ES components (Figure 5.6A-C).
Figure 5.6. Western blots showing anti-ELV and anti-ES serum IgG response for sham, ELV and ES vaccination groups. For each blot, ELV and ES components were separated by SDS-PAGE. Samples were electrotransferred onto nitrocellulose membrane and this was probed with sera from the sham (A), ELV (B) or ES (C) vaccination groups. Bound antibody was detected using an anti-mouse IgG (whole molecule) alkaline phosphatase antibody, and proteins were visualised using BCIP and nitro blue tetrazolium. * Indicates major ELV components bound by sera. (D) Shows ELV and ES material separated by SDS-PAGE. Left of each panel shows molecular weight markers in kDa. 1 and 2 refer to two separate ELV preparations.
250
150
100
75
37
25 20 15 10
50
250
150
100
75
37
25 20 15 10
50
250
150
100
75
37
25 20 15 10
50
ELVs
ES
250
150
100
75
37
25 20 15 10
50
ELVs ES
kDa kDa kDa kDa
ELVs
ES
ELVs
ES
*
**
*
*
*
A) Sham vaccination group B) ELV vaccination group C) ES vaccination roup D) SDS-PAGE separation of ELVs
and ES
*
*
1 2 1 2 1 2

159
Figure 5.6D shows SDS-PAGE separation of ELV and ES material. Bands
corresponding to 100, 80 and 70 kDa ELV components were excised from the
gel, since these were the most prominent bands in Figure 5.6B. The protein
composition of these bands was determined by mass spectrometry (Table 2).
Table 5.3. Possible identities of ELV components targeted by IgG antibodies following vaccination. Bands corresponding to 100 (Band 1), 80 (Band 2) and 70 kDa (Band 3) were excised from the SDS-PAGE gel shown in Figure 5.6D and their protein content was analysed by mass spectrometry. The proteins identified within these bands are listed. The number of unique peptides identified for each protein is displayed (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). *= proteins identified within ES depleted of ELVs.
The proteins identified within band 1 (100 kDa) include the Vacuolar protein
sorting associated protein 52 and Conserved hypothetical protein. The
predicted molecular weights of these proteins are 164 and 51 kDa respectively,
however fragments or dimers of these proteins may be present within this band.
This may also be the case for band 2 (80 kDa), where the Vacuolar protein
sorting associated protein 52 has also been identified, along with VWD and
Accession number Protein Band
1 Band
2 Band
3 Mw
(kDa)
*TMUE_s0245000500 VWD and Vitellogenin N and DUF1943-domain containing protein
9 17 190
*TMUE_s0093001800 Vacuolar protein sorting associated protein 52
5 4
164
*TMUE_s0070003500 TSP-1 domain containing protein
3 39
*TMUE_s0189001400 Neurogenic locus notch protein
3
53
TMUE_s0015006300 Peptidase M8 domain containing protein
4
60
TMUE_s0037004100 Conserved hypothetical protein 4
51
*TMUE_s0022000400 Na+ K+ ATPase alpha subunit 1
3 118
TMUE_s0117002800 Trypsin and CUB domain containing protein
3
69
TMUE_s0093001400 Nicastrin family protein
3 78
TMUE_s0106000600 Moesin:ezrin:radixin 1 3
64
TMUE_s0011007600 Anoctamin
2 96
TMUE_s0060000200 Prominin domain containing protein
2 75
TMUE_s0059000500 Neurogenic locus notch protein
2 62
TMUE_s0320000100 Neurogenic locus notch protein 2
2 49

160
Vitellogenin N and DUF1943-domain containing protein (predicted molecular
weight of 190 kDa). The VWD and Vitellogenin N and DUF1943-domain
containing protein was also identified in band 3 (70 kDa), along with the TSP-1
domain containing protein, a protein with multiple tetraspanin domains
(Appendix 3, Figure A3.1). The S. mansoni tetraspanins, Sm-TSP-1 and
Sm-TSP-2, have been identified as vaccine candidates (27), and there appears
to be significant homology between TMUE_s0070003500 and four S. mansoni
proteins (Appendix 3, Table A3.2).
Many of these proteins were also identified within the soluble portion of ES
(indicated by * on Table 5.3), confirming that although there are differences
between the protein content of T. muris ES and ELV samples, the two are not
mutually exclusive.
5.8 Discussion
The vesicles isolated from T. muris ES fit the size and shape characteristics for
classification as exosomes, and previously described exosome markers
(including tetraspanins, heat shock proteins and Alix (157) were identified within
these samples. These vesicles are described here as ELVs, since a lack of
commercially available antibodies against T. muris exosome markers prevents
unequivocal classification of these vesicles as exosomes by Western blotting.
Mass spectrometry analysis showed that the majority of T. muris ELV proteins
lack a signal peptide (68%) and that there was significant overlap between the
protein content of ELVs and ES (77% of ELV proteins were identified within
adult ES). This suggests that ELVs may be an important mechanism by which
these proteins are released into the external environment. Similarly, Marcilla
and colleagues reported significant overlap between the protein content of
F. hepatica and E. caproni ES and ELV samples (152). Data presented in this
chapter also showed that T. muris ELVs fused with human colonic epithelial
cells in vitro, suggesting that these vesicles may be able to fuse with host cells

161
in vivo. This is particularly relevant for T. muris since the parasite is in constant
contact with the caecal epithelium.
ELVs have been isolatated previously from T. muris ES by Tritten and
colleagues and Eichenberger and co-workers (180, 183). Tritten and colleagues
carried out proteomic analysis on the isolated ELVs, however there are
differences between the protein content of ELVs isolated for this project and
those isolated by Tritten and co-workers. Notably, Tritten and colleagues report
that the most abundant protein within T. muris ELV samples was the P43 (180),
whereas this protein was absent from our proteomic analyses. The P43 was
removed from T. muris ES following ELV isolation, and therefore the removal
step should not have effected the P43 content of ELVs. Tritten and colleagues
used a commercially available kit to isolate ELVs from ES samples (180),
whereas ultracentrifugation was used for this project. It is possible that the kit
used by Tritten and colleagues led to greater contamination of ELV samples
with ES proteins.
The total number of ELV proteins identified for this project was also greater than
the number identified by Tritten and colleagues (125 versus 73) (180). Again,
these differences may be due to variation in the methods used to isolate ELVs
and/or perform proteomic analysis. Ultracentrifugation is considered the ‘gold
standard’ within the field of EV research and the size of vesicles isolated at
different speeds has been well characterised (166). The commercially available
kit used by Tritten and colleagues may enrich for vesicles of a certain size
range whilst excluding others, which could affect the protein content of samples.
In addition, ELV samples were precipitated before mass spectrometry analysis
was carried out (as described by Marcilla and colleagues) (152), whereas
Tritten and colleagues did not perform a precipitation step (180). Another key
difference between the two data sets is that Tritten and colleagues analysed the
protein content of a single sample (180), whereas our data set is arguable more
stringent, since proteomic analysis was carried out on three biological replicates
and only proteins identified in two or more samples were included.

162
Data presented in this chapter showed that vaccination with T. muris ELVs can
induce protective immunity against a subsequent T. muris infection. The
protection afforded by these ELV vaccinations was variable; sterile immunity
was achieved for some individuals, while others developed chronic infection.
This has also been reported for H. polygyrus ELV vaccinations, although these
vaccinations were formulated with aluminum hydroxide (155), whereas the ELV
vaccinations performed for this thesis were formulated without adjuvant.
Vaccinating mice with 3 μg of T. muris ES formulated with aluminum hydroxide
induced sterile immunity against a subsequent low dose infection (Appendix 3,
Figure A3.2), demonstrating that the right combination of antigens and adjuvant
can stimulate protective immunity, even when low quantities of antigen are
used. Based on these observations, the potential for aluminum hydroxide to
boost immunity to T. muris ELVs should be investigated further.
The protective properties of T. muris ELVs are dependent on intact vesicles, as
vaccination with lysed ELVs did not protect mice from a subsequent infection.
Vaccine research using liposomes and microparticles may offer insight into why
ELVs make effective vaccines (235). It has been suggested that encapsulating
antigens in lipid spheres protects them from degradation and enables slow
release of antigen over time (235-237). In addition, Fifis and colleagues have
demonstrated that 40 to 50 nm-sized nanoparticles are preferentially taken up
by DEC205+ CD40+ CD86+ murine DCs compared to larger particles of up to
2 μm in size (238). Therefore it is reasonable to suggest that presentation of
Trichuris proteins within ELVs makes them better suited for uptake by antigen
presenting cells, thus increasing their antigenicity. This should be explored
further as encapsulating recombinant or purified native Trichuris antigens within
ELVs may be a viable alternative to traditional vaccinations formulated with
adjuvant.
The data presented here shows that vaccination with ELVs boosts IgG1
antibody production against soluble ES proteins. This may be explained by the
extensive overlap between proteins identified in ELV samples and ES depleted
of ELVs (Table 5.2). Coakley and colleagues report similar findings,

163
demonstrating that vaccinating mice with H. polygyrus exosomes prior to
infection boosted antibody response to ES depleted of ELVs, and that rats
vaccinated with H. polygyrus ELVs make antibodies against ELV and ES
material in the absence of infection (155). High levels of anti-parasite IgG2a/c
were measured in all mice that received the ELV vaccine, and anti-parasite
IgG2a/c did not appear to correlated with worm burden (data not shown), which
suggests that ELV vaccinated mice mount a mixed Th1/Th2 response to a low
dose T. muris infection.
Figure 5.6B shows that the IgG in sera of ELV vaccinated mice binds to a
number of components that are enriched within ELV samples. The strongest
antibody response was directed towards approximately 100, 80 and 70 kDa
components. Figure 5.6D shows SDS-PAGE separation of the lysed ELV
material; although it is difficult to identify distinct bands that correlate with these
molecular weights, mass spectrometry analysis of the protein content within
these regions revealed a number of potential antigens. These include VWD and
Vitellogenin N and DUF1943-domain containing protein, TSP-1 domain
containing protein and Vacuolar protein sorting associated protein 52, which are
amongst the most abundant ELV proteins (Appendix 3, Table A3.1). Although
antibody responses may not reflect protection, the therapeutic value of related
proteins has been demonstrated in other helminths (34-41), suggesting that
these proteins are major candidates for protective antigens. Vitellogenin
proteins isolated from the ES of gravid adult female Litomosoides sigmodontis,
a filarial nematode of rodents, and Ostertagia ostertagi, an intestinal nematode
of cattle, have been identified as novel vaccine candidates using
immunoscreening and proteomics approaches (239, 240). In addition,
Vitellogenin proteins have been recognised as potential vaccine candidates for
ectoparasites, such as ticks and mites (241-244). Tritten and colleagues also
identified this protein within T. muris ELV samples (180).
There are no published reports relating to Vacuolar sorting protein associated
protein 52, however there appear to be homologues of this protein in other
tricephalid parasites, including Trichuris and Trichinella species (Appendix 3,

164
Table A3.3), and this protein was also identified in pool 3 (Appendix 1, Table
A1.4), which formed the basis for the experiments carried out in Chapter 4. The
TSP-1 domain containing protein could also be a promising immunogenic
candidate, given that S. mansoni TSP proteins have shown great potential in
pre-clinical and Phase I clinical trials (23, 245). In future, 2D Western blotting
should be carried out in order to get better resolution and more certain protein
identification.
This is the first example of successful vaccination against a Trichuris parasite
using ELVs, and the first example of an ELV vaccination formulated without
adjuvant. Recent reports have demonstrated that vaccination with H. polygyrus
ELVs can protect mice against a subsequent infection, while vaccinating mice
with E. caproni ELVs can improve the clinical outcome of infection (155, 156).
There are also a number of examples of protective immunity induced by
vaccination with ELVs derived from host cells, for example, vaccinating CBA/J
mice with ELVs collected from splenic DCs pulsed with T. gondii antigens
before pregnancy induced protective immunity in pups, resulting in fewer brain
cysts and lower mortality following congenital exposure (246). Similarly, del
Cacho and co-workers demonstrated that immunising chickens with ELVs
derived from DCs pulsed with Eimeria parasites lead to reduced mortality,
intestinal inflammation and faecal oocyst shedding (247). Martin-Jaular and co-
workers also report a protective role for reticulocyte-derived ELVs containing
Plasmodium yoelii material, showing that vaccination with these ELVs
stimulated IgG antibodies capable of binding infected red blood cells, with 83%
of mice surviving an otherwise lethal P. yoelii infection. Previously described
S. mansoni vaccine candidates have also been identified in ELV
samples (173-177), supporting the data presented here, which suggest that
helminth ELVs may be an important source of protective material.
In conclusion, the data presented here show that vaccination with T. muris
ELVs can protect mice against a subsequent T. muris infection, and these
vaccinations boost antibody response to ES depleted of ELVs. A number of
potential immunogenic candidates were identified by Western blotting; these

165
include VWD and Vitellogenin N and DUF1943-domain containing protein,
Vacuolar protein sorting associated protein 52 and TSP-1 domain containing
protein. Future work should investigate recombinant forms of these proteins as
protective antigens and explore opportunities for ELVs to boost their
antigenicity.

166
6 Summary discussion

167
The aim of this thesis was to identify Trichuris-specific antigens that prime the
host for worm expulsion. Recent figures suggest that 800 million people are
infected with T. trichiura worldwide, and Trichuris parasites are also a huge
source of morbidity in livestock (1, 7). The need for vaccines against T. trichiura
is arguably greater than ever, due to reports showing that drug resistance has
arisen in some parasite populations (20, 21). Trichuris ES was identified as a
potent source of host protective material as far back as 1977 (115), however
relatively little progress has been made towards identifying the molecular
components involved. Over the course of this thesis, a number of important
observations were made relating to the characteristics of the immune response
induced by vaccination with ES (Chapter 3), and several immunogenic
candidates were identified using chromatography and mass spectrometry
methods combined with measures of T cell cytokine production (Chapter 4).
In addition, Chapter 5 explored the potential of T. muris ELVs as a source of
protective material. This research represents significant progress towards
identifying vaccine candidates for T. trichiura and other Trichuris parasites.
6.1 Identification of immunogenic candidates within T. muris ES
To date, there are no commercially available vaccines for human STH
infections, and very few against their veterinary counterparts (248). However, in
the last decade, remarkable progress has been made towards a vaccine for the
hookworm, N. americanus (100, 106, 107). The selection of vaccine candidates
was based on identifying parasite components involved in the breakdown of
host haemoglobin (Na-APR-1) and detoxification of haem (Na-GST-1) (100).
These processes are essential for parasite survival, and it was predicted that
vaccination with recombinant forms of these proteins would induce antibodies
capable of inhibiting the native hookworm proteins, thus affecting parasite
viability (100). These vaccine candidates are currently undergoing Phase II
clinical trials in Gabon, having successfully completed phase I trials in the US
and Brazil (107). Comparatively little is known about the biology of T. trichiura,

168
and unlike N. americanus, there were no immediately obvious vaccine
candidates for this parasite. A detailed analysis of the genomes and
transcriptomes of T. trichiura and T. muris published in 2014 identified a number
of key functional groups that are upregulated in the anterior end of the worm,
which was speculated to be the site of ES release (although precise details of
how ES is released are unknown) (33). These include chymotrypsin-like serine
proteases, protease inhibitors/WAP proteins and DNases, however, the large
number of proteins within these groups may be a barrier to investigating these
components as vaccine candidates (33).
Other researchers have used antibody-based screening methods to identify
potential immunogenic candidates for helminth parasites. One of the most
clinically advanced examples is S. haematobium Sh28GST, a glutathione-S-
transferase, which was shown to reduce worm viability (by 38%) and egg output
(by 55 to 74%) in a range of experimental animal models, and was deemed safe
and effective against urinary schistosomiasis in Phase I clinical trials
(23, 249, 250). Field studies demonstrated that resistance to schistosomiasis is
associated with anti-Sh28GST IgA and IgG3 neutralising antibodies,
re-enforcing the rationale for using antibody-screening methods to identify
vaccine candidates for this parasite (251, 252). Highly effective recombinant
protein vaccines have also been developed for a range of taeniid parasites
using antibody-based screening methods, and these studies demonstrate that it
is possible to induce protective immunity against metazoan parasites using a
single recombinant protein (although this may not be the case for all parasitic
helminths) (92).
The results presented in Chapter 3 showed that transfer of serum from ES
vaccinated naïve mice did not protect unvaccinated mice from a subsequent low
dose infection, despite the fact that high levels of anti-parasite IgG1 antibodies
were detected in the transfer serum. Based on these results, antibody-based
screening methods may not identify potent vaccine candidates for Trichuris
parasites, particularly as ES appears to be targeted differentially by the humoral
and cellular branches of the immune system (Chapter 3). Instead, the approach
used to identify vaccine candidates in this thesis focused on material that

169
stimulates Th2 cytokine release, particularly IL-13 and IL-9, since previous
studies have demonstrated a crucial role for these cytokines in resistance to
T. muris, through increasing intestinal mucus production, accelerating caecal
epithelial cell turnover and inducing intestinal hypercontractility (44, 56, 57).
This assumes that vaccination primes for effector mechanisms similar to those
induced by natural infection, however this should be investigated further.
Chapter 3 included other important observations relating to the immune
response following vaccination with ES products. Firstly, degradation of ES
using proteinase K abrogated its ability to stimulate protective immunity
following vaccination, suggesting that proteins are likely to be responsible for
the protective properties of ES. This reinforces the rationale for focussing on the
protein content of ES in terms of identifying immunogenic candidates.
Alternatively, this result may suggest that even though CD4 T cells recognise
small peptide fragments (as presumably proteinase K would generate), they do
not enter the processing/presentation pathway in antigen presenting cells to
effectively prime the T cells, or that the aluminum hydroxide adjuvant does not
function very effectively with small sized peptides.
Secondly, vaccination with ES products stimulates long-lasting immunity against
a subsequent challenge, suggesting that vaccination stimulates immunological
memory. It is generally accepted that in order to confirm that memory cells are
generated in mouse studies, at least 30 days should have elapsed post antigen
dosage (190-192). Here, mice were infected a total of 50 days after the second
vaccination, which strongly suggests that vaccination with T. muris ES products
stimulates immunological memory. This should be explored further in order to
characterise the immune components involved.
Thirdly, the experiments described in Chapter 3 suggest that vaccination at a
peripheral (subcutaneous) site is able to prime cells that can operate at an
intestinal site. This is surprising as it is believed that for primed T cells to move
into intestinal tissue, gut-specific homing molecules need to be upregulated on
the T cells during activation (253). The present data would suggest that this
may not be the case, or that a subcutaneous injection of antigen does prime

170
sufficient numbers of T cells in the MLN, where appropriate homing molecules
can be acquired (253).
Dixon and colleagues provide some insight into the immune response following
subcutaneous vaccination of AKR mice with T. muris ES (29). They showed that
the peripheral lymph node cells proliferate and produce Th2 cytokines and
IFN-γ following vaccination (in the absence of infection) in response to
re-stimulation with ES (29). However, when vaccination was followed by a high
dose infection, the MLN was the major source of Th2 cytokines, leading to
goblet cell hyperplasia and influx (and alternative activation) of
macrophages (29). Although the vaccinated AKR mice did expel a high dose
infection, the authors did not observe an increase in epithelial cell turnover,
suggesting that there are some differences between the mechanisms of
expulsion in mice that have acquired resistance through acute infection and in
response to vaccination (29). In future, further characterisation of the immune
response following vaccination should be carried out. Vaccination experiments
could be performed in IL-13 or IL-4R knock out mice in order to confirm that
vaccine-driven immunity is dependent on IL-13 production, and experiments
could be carried out to determine the role of IL-9, Muc5ac production and
epithelial turnover in driving worm expulsion in a low dose C57BL/6 vaccination
model.
Finally, vaccination with ES collected from both larval and adult stages
stimulates protective immunity, and there is significant overlap between the
protein content of these two sources of ES. The rationale for this experiment
was that during acute infection, worms are expelled before they reach patency
and therefore the host is not ordinarily exposed to adult ES (31). Instead,
expulsion in resistant mouse strains must be triggered by larval components.
The experiments carried out prior to this involved vaccination with adult ES as
practically, much larger quantities of ES can be collected from adult worms
compared to larval stages. However, the present experiments do confirm that
adult T. muris ES is a biologically relevant source of antigens that can induce
protection.

171
Additionally, data presented in Figure A3.2 showed that vaccination with 3 μg of
ES protected mice against a subsequent T. muris infection. The main reason for
carrying out this titration experiment was to compare the protection conferred by
vaccination with 3 μg of ELV material and 3 μg of ES (Figures 5.3 and A3.2).
Clearly, the ES vaccination was more potent, however this vaccination was
formulated with aluminum hydroxide, whereas the ELV vaccine was formulated
without adjuvant. Adjuvant usage often enables lower quantities of antigens to
be used (a phenomena known as ‘dose sparing’) (135), and therefore, the ES
titration experiment should be repeated both with and without adjuvant to
compare the quantity of material required to stimulate protection in the presence
and absence of adjuvant. ELV vaccinations formulated with and without
aluminum hydroxide should be carried out alongside this experiment, in order to
determine the optimum conditions for vaccination with ES and ELV material.
The ES titration experiment was carried out at the end of the project, which is
why all the experiments described in Chapters 3 and 4 involved vaccinations
with 30 μg of ES products. Based on initial experiments performed with native
ES products in Chapter 3, 30 μg of protein was also used for the recombinant
protein vaccines described in Chapter 4, although the amount of each protein
within 30 μg of ES is likely to be much lower.
6.2 Identification of immunogenic candidates within T. muris ELVs
Chapter 5 investigated the potential of T. muris ELVs as a source of
immunogenic candidates. ELVs have been isolated previously from T. suis and
T. muris ES, however this is the first report to include TEM images of the
vesicles, and to isolate ELVs from adult ES using differential ultracentrifugation.
Mass spectrometry was carried out on three ELV samples, and the list of
identified proteins was more comprehensive than the list published by Tritten
and colleagues earlier this year (180). Comparison of the protein content of
T. muris ELVs and ES samples showed that there was significant overlap.
Importantly, the present work also showed that ELV vaccinations (formulated

172
without adjuvant) induced protective immunity in male C57BL/6 mice. This is the
first example of a successful helminth ELV vaccination formulated without
adjuvant.
In future, work should be carried out in order to explore how an ELV vaccine
might work in the host and to determine how antigen presentation might be
affected. Encapsulating antigens in lipid spheres may increase their antigenicity,
much like how lipid-based adjuvants increase the antigenicity of vaccines (135).
Alternatively, ELV size could play a major role, promoting uptake of antigens by
DCs (238). QuilA, a commonly used adjuvant for vaccines against ruminant
gastrointestinal nematodes (209), encapsulates antigens in 40 nm cage-like
structures, and the size of these particles is thought to increase
immunostimulation (135). In addition, studies comparing the adjuvanticity of
112 nm and 9.3 μm aluminum hydroxide particles showed that antibody
production and antigenicity was higher in response to the nano-particles (254).
The authors suggest that this may be due to the larger surface area available
for absorption of antigen (254). These studies emphasise the importance of
particle size on adjuvanticity.
An antibody-based screening approach was used to identify potential
immunogenic candidates within ELVs in Chapter 5. Despite the discussions
throughout this project regarding the suitability of such approaches for
identifying host protective antigens for T. muris, limitations in the amount of ELV
material available would have made identifying immunogenic candidates using
chromatography methods similar to those employed in Chapter 4 impractical, if
not impossible. Antibody-based screening methods at least give an indication of
which components are targeted by the immune response following vaccination
and subsequent infection. The two immunogenic candidates that stood out were
VWD and Vitellogenin N and DUF1943-domain containing protein and TSP-1
domain containing protein. Both Vitegellin and TSP-1 proteins have been
identified as vaccine candidates for other helminth and ectoparasites (23, 27,
239-244), and therefore warrant further investigation.

173
6.3 Other considerations for Trichuris vaccine design
If effective vaccine candidates were identified, it would be necessary to consider
the ‘target product profile’ of the vaccine. This would involve identifying the
target patient group and summarising the safety and efficacy of the vaccine.
Population studies show that the majority of low-level infections are
asymptomatic, while clinical symptoms are generally associated with heavy
worm burdens (8, 9). This suggests that a successful vaccine against
T. trichiura would need to reduce the worm burden such that clinical symptoms
are reduced, however, this may be possible without achieving complete worm
expulsion. In endemic areas, close to 90% of children are infected with
T. trichiura within the first twelve months (9), so ideally a successful vaccine
against this parasite should be administered in the first year of life, along with
other childhood vaccines. It would also be necessary to determine whether
vaccination can protect against a pre-existing infection. If this was the case,
older children and adults could be vaccinated along with infants. Bancroft and
colleagues showed that mice are susceptible to a high dose T. muris infection if
a low dose is administered first, however the authors also demonstrated that
worm expulsion occurs when mice are given multiple low dose infections (trickle
regime) (45). These data suggest that it might be possible to vaccinate people
with an ongoing chronic T. trichiura infection, however it may be necessary to
administer multiple vaccinations over time to achieve an appropriate level of
protection.
This could be investigated using the T. muris mouse model by allowing a low
dose infection to develop to chronicity in male C57BL/6 mice. These mice could
then be given three or more ES vaccinations and worm burdens should be
assessed a number of weeks post-vaccination. Similar experiments were
performed as part of this project, however when the mice were sacrificed at day
76 p.i., the negative control group (which was infected and then vaccinated with
aluminum hydroxide only) had very low worm burdens (around 5 worms per
mouse, data not shown), which meant that it was difficult to compare the mean
worm burden of sham versus ES vaccinated mice. This is likely due to the

174
viability of worms decreasing with age. In future, this experiment should be
repeated, sacrificing mice at an earlier time point to ensure that the worms are
still viable.
Gomez-Samblas and colleagues claim to have identified a vaccine candidate (a
serine/threonine phosphatase 2A from A. costaricensis) that can trigger
expulsion of a chronic T. muris infection in AKR mice (30). However, the worms
were 90 days old at the point of necroscopy, and therefore, based on the
observations described above, the viability of the worms is likely to be reduced
at this time point. Although the authors do report a statistically significant
reduction in worm burden following vaccinations with the serine/threonine
phosphatase compared to the sham vaccination group 12 days post
vaccination, the worm burden of the sham vaccine group was reduced by 50%
compared to the worm burden at 7 days post vaccination (30). This suggests
that the viability of the worms is reduced at this time point. This study should be
modified in order to assess worm burden at an earlier time point, when the
worms are less aged, in order to confirm that the serine/threonine phosphatase
is a potent vaccine candidate.
Another complicating factor for vaccinating humans, or indeed veterinary
species, against Trichuris parasites, is that individuals are exposed to multiple
pathogens from a young age. These may be other STH species, for example
A. lumbricoides and hookworms, however people are also likely to be exposed
to a range of bacterial, viral and protozoan pathogens (2, 255-257). This may
affect the type of immunity that is mounted in response to vaccination with
Trichuris components, and is something that needs to be taken into
consideration if a vaccine was to reach clinical trials (256, 257).
In addition, care should be taken to ensure that vaccine candidates are not
allergenic, as this may cause problems if the vaccine population has previously
been exposed to the parasite. This issue was brought to the attention of
researchers after the Phase II clinical trial for the N. americanus vaccine
candidate, Na-ASP-2, was terminated early, as three out of the seven

175
volunteers experienced allergic reactions, which are thought to have been
triggered by high levels of pre-existing anti-parasite IgE (103).
6.4 Conclusions and future perspectives
In summary, the work presented in this thesis represents significant progress
towards identifying vaccine candidates for Trichuris parasites and important
observations were made in relation to the immunity induced by vaccination with
ES products and ELVs in mice. Specifically, the work described in Chapter 3
showed that vaccination with T. muris ES proteins (collected from the larval and
adult stages) stimulates long-lasting protection against a subsequent low dose
infection in C57BL/6 mice. The data presented in Chapter 4 provided a
framework by which to identify vaccine candidates within T. muris ES. This
approach could be used in future to identify immunogenic candidates within
other groups of ES proteins (for example pool 4). Future work should also
include investigation of alternative expression systems, in order to find one that
will enable the expression of the five immunogenic candidates that HEK293
cells were unable to express. As discussed in Chapter 4, there is no clear way
to determine the most appropriate expression system for synthesis of
recombinant helminth proteins, since expression systems that are effective for
one protein may not enable expression of other proteins from the same species
(206). Recent work describing the ability to engineer yeast to add complex
mammalian glycans during recombinant protein expression could be applied to
helminth protein expression, providing that more information regarding the
glycosylation patterns of these species was available (223, 224).
Chapter 5 highlighted the potential of Trichuris ELVs as a source of
immunogenic material, and this is the first report, to our knowledge, to identify
novel immunogenic candidates within nematode ELVs. These studies
demonstrate that ELVs are an undervalued source of protective antigens that
warrant further exploration, and suggest that packaging of native or

176
recombinant antigens within ELVs may boost protection. Future work should
investigate the potential for recombinant forms of the ELV antigenic candidates
to stimulate protective immunity when administered as a vaccine formulated
with aluminum hydroxide, and also the potential to boost immunity through
encapsulating these proteins in synthetic ELV-sized lipid spheres.

177
References
1. Grencis RK. Cytokine regulation of resistance and susceptibility to intestinal nematode infection - from host to parasite. Veterinary Parasitology. 2001;100(1-2):45-50.
2. Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A, Diemert D, Hotez PJ. Soil-transmitted helminth infections: ascariasis, trichuriasis, and hookworm. Lancet. 2006;367(9521):1521-32.
3. Hotez PJ, Strych U, Lustigman S, Bottazzi ME. Human anthelminthic vaccines: Rationale and challenges. Vaccine. 2016;34(30):3549-55.
4. Ojha SC, Jaide C, Jinawath N, Rotjanapan P, Baral P. Geohelminths: public health significance. Journal of Infection in Developing Countries. 2014;8(1):5-16.
5. Utzinger J, Keiser J. Schistosomiasis and soil-transmitted helminthiasis: common drugs for treatment and control. Expert Opinion on Pharmacotherapy. 2004;5(2):263-85.
6. Hotez PJ, Molyneux DH, Fenwick A, Kumaresan J, Sachs SE, Sachs JD, Savioli L. Current concepts - Control of neglected tropical diseases. New England Journal of Medicine. 2007;357(10):1018-27.
7. Keiser J, Utzinger J. Efficacy of current drugs against soil-transmitted helminth infections - Systematic review and meta-analysis. Jama-Journal of the American Medical Association. 2008;299(16):1937-48.
8. Stephenson LS, Holland CV, Cooper ES. The public health significance of Trichuris trichiura. Parasitology. 2000;121:S73-S95.
9. Bundy DAP. Population ecology of intestinal helminth infections in human communities. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences. 1988;321(1207):405-20.
10. Bundy DAP, Cooper ES, Thompson DE, Didier JM, Anderson RM, Simmons I. Predisposition to Trichuris trichiura infection in humans. Epidemiology and Infection. 1987;98(1):65-71.
11. Stoltzfus RJ, Dreyfuss ML, Chwaya HM, Albonico M. Hookworm control as a strategy to prevent iron deficiency. Nutrition Reviews. 1997;55(6):223-32.
12. Stephenson LS, Latham MC, Kurz KM, Kinoti SN, Brigham H. Treatment with a single dose of albendazole improves growth of Kenyan school children with hookworm, Trichuris trichiura, and Ascaris lumbricoides infections. American Journal of Tropical Medicine and Hygiene. 1989;41(1):78-87.
13. Stephenson LS, Latham MC, Adams EJ, Kinoti SN, Pertet A. Physical fitness, growth and appetite of Kenyan school boys with hookworm, Trichuris

178
trichiura and Ascaris lumbricoides infections are improved 4 months after a single dose of albendazole. Journal of Nutrition. 1993;123(6):1036-46.
14. Smith JL, Brooker S. Impact of hookworm infection and deworming on anaemia in non-pregnant populations: a systematic review. Tropical Medicine & International Health. 2010;15(7):776-95.
15. Partridge FA, Murphy EA, Willis NJ, Bataille CJR, Forman R, Heyer-Chauhan N, Marinič B, Sowood DJC, Wynne GM, Else KJ, Russell AJ, Sattelle DB. Dihydrobenz[e][1,4]oxazepin-2(3H)-ones, a new antihelmintic chemotype immobilising whipworm and reducing infectivity in vivo. PLOS Neglected Tropical Diseases. 2017.
16. Albonico M1, Smith PG, Ercole E, Hall A, Chwaya HM, Alawi KS, Savioli L. Rate of reinfection with intestinal nematodes after treatment of children with mebendazole or albendazole in a highly endemic area. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1995;89(5):538-41.
17. Chan L, Bundy DAP, Kan SP. Aggregation and predisposition to Ascaris lumbricoides and Trichuris trichiura at the familial level. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1994;88(1):46-8.
18. Elkins DB, Haswellelkins M, Anderson RM. The importance of host age and sex to patterns of reinfection with Ascaris lumbricoides following mass anthelminthic treatment in a South Indian fishing community. Parasitology. 1988;96:171-84.
19. Dixon H, Johnston CE, Else KJ. Antigen selection for future anti-Trichuris vaccines: a comparison of cytokine and antibody responses to larval and adult antigen in a primary infection. Parasite Immunology. 2008;30(9):454-61.
20. Albonico M, Bickle Q, Ramsan M, Montresor A, Savioli L, Taylor M. Efficacy of mebendazole and levamisole alone or in combination against intestinal nematode infections after repeated targeted mebendazole treatment in Zanzibar. Bulletin of the World Health Organization. 2003;81(5):343-52.
21. Flohr C, Tuyen LN, Lewis S, Minh TT, Campbell J, Britton J, Williams H, Hien TT, Farrar J, Quinnell RJ. Low efficacy of mebendazole against hookworm in Vietnam: Two randomized controlled trials. American Journal of Tropical Medicine and Hygiene. 2007;76(4):732-6.
22. Z Zhan B, Beaumier CM, Briggs N, Jones KM, Keegan BP, Bottazzi ME, Hotez PJ. Advancing a multivalent 'Pan-anthelmintic' vaccine against soil-transmitted nematode infections. Expert Review of Vaccines. 2014;13(3):321-31.
23. Riveau G, Deplanque D, Remoue F, Schacht AM, Vodougnon H, Capron M, Thiry M, Martial J, Libersa C, Capron A. Safety and Immunogenicity of rSh28GST Antigen in Humans: Phase 1 Randomized Clinical Study of a

179
Vaccine Candidate against Urinary Schistosomiasis. Plos Neglected Tropical Diseases. 2012;6(7).
24. Zhang WD, Ahmad G, Le L, Rojo JU, Karmakar S, Tillery KA, Torben W, Damian RT, Wolf RF, White GL, Carey DW, Carter D, Reed SG, Siddiqui AA. Longevity of Sm-p80-specific antibody responses following vaccination with Sm-p80 vaccine in mice and baboons and transplacental transfer of Sm-p80-specific antibodies in a baboon. Parasitology Research. 2014;113(6):2239-50.
25. Le L, Zhang WD, Karmakar S, Ahmad G, Torben W, Siddiqui AA. Simultaneous priming with DNA encoding Sm-p80 and boosting with Sm-p80 protein confers protection against challenge infection with Schistosoma mansoni in mice. Parasitology Research. 2014;113(3):1195-200.
26. Karmakar S, Zhang WD, Ahmad G, Torben W, Alam MU, Le L, Damian RT, Wolf RF, White GL, Carey DW, Carter D, Reed SG, Siddiqui AA. Use of an Sm-p80-Based Therapeutic Vaccine to Kill Established Adult Schistosome Parasites in Chronically Infected Baboons. Journal of Infectious Diseases. 2014;209(12):1929-40.
27. Pearson MS, Becker L, Driguez P, Young ND, Gaze S, Mendes T, Li XH, Doolan DL, Midzi N, Mduluza T, McManus DP, Wilson RA, Bethony JM, Nausch N, Mutapi F, Felgner PL, Loukas A. Of monkeys and men: immunomic profiling of sera from humans and non-human primates resistant to schistosomiasis reveals novel potential vaccine candidates. Frontiers in Immunology. 2015;6.
28. Jenkins SN, Wakelin D. Functional antigens of Trichuris muris released during in vitro maintenace - their immunogenicity and partial purification. Parasitology. 1983;86(FEB):73-82.
29. Dixon H, Little MC, Else KJ. Characterisation of the protective immune response following subcutaneous vaccination of susceptible mice against Trichuris muris. International Journal for Parasitology. 2010;40(6):683-93.
30. Gomez-Samblas M, Garcia JJ, Rodriguez M, Trelis M, Bernal D, Lopez-Jaramillo FJ, Santoyo-Gonzalez F, Vilchez S, Espino AM, Bolas-Fernandez F. Self-adjuvating C18 lipid vinil sulphone-PP2a vaccine: study of the induced immunomodulation against Trichuris muris infection. Open Biology. 2017;7.
31. Klementowicz JE, Travis MA, Grencis RK. Trichuris muris: a model of gastrointestinal parasite infection. Seminars in Immunopathology. 2012;34(6):815-28.
32. Hurst RJM, Else KJ. Trichuris muris research revisited: a journey through time. Parasitology. 2013;140(11):1325-39.
33. F Foth BJ, Tsai IJ, Reid AJ, Bancroft AJ, Nichol S, Tracey A, Holryd N, Cotton JA, Stanley EJ, Zarowiecki M, Liu JZ, Huckvale T, Cooper PJ, Grencis RK, Berriman M. Whipworm genome and dual-species transcriptome analyses provide molecular insights into an intimate host-parasite interaction. Nature genetics. 2014;46(7):693-700.

180
34. Hayes KS, Bancroft AJ, Goldrick M, Portsmouth C, Roberts IS, Grencis RK. Exploitation of the Intestinal Microflora by the Parasitic Nematode Trichuris muris. Science. 2010;328(5984):1391-4.
35. Tilney LG, Connelly PS, Guild GM, Vranich KA, Artis D. Adaptation of a nematode parasite to living within the mammalian epithelium. Journal of Experimental Zoology Part a-Comparative Experimental Biology. 2005;303A(11):927-45.
36. Grencis RK, Humphreys NE, Bancroft AJ. Immunity to gastrointestinal nematodes: mechanisms and myths. Immunological Reviews. 2014;260(1):183-205.
37. Wakelin D, Selby GR. Thymus-dependency of the immune response of mice to a primary infection with the nematode Trichuris muris. International journal for parasitology. 1974;4:657-61.
38. Lee TDG, Wakelin D, Grencis RK. Cellular mechanisms of immunity to the nematode Trichuris muris. International Journal for Parasitology. 1983;13(4):349-53.
39. Ito Y. The absence of resistance in congenitally athymic nude mice toward infection with the intestina nematode, Trichuris muris - resistance restored by lymphoid cell transfer. International Journal for Parasitology. 1991;21(1):65-9.
40. Koyama K, Tamauchi H, Ito Y. The role of CD4+ and CD8+ T cells in protective immunity to the murine nematode parasite Trichuris muris. Parasite Immunology. 1995;17(3):161-5.
41. Koyama K. NK1.1(+) cell depletion in vivo fails to prevent protection against infection with the murine nematode parasite Trichuris muris. Parasite Immunology. 2002;24(11-12):527-33.
42. Betts CJ, deSchoolmeester ML, Else KJ. Trichuris muris: CD4(+) T cell-mediated protection in reconstituted SCID mice. Parasitology. 2000;121:631-7.
43. Else KJ, Finkelman FD, Maliszewski CR, Grencis RK. Cytokine mediated regulation of chronic intestinal helminth infection. Journal of Experimental Medicine. 1994;179(1):347-51.
44. Bancroft AJ, McKenzie ANJ, Grencis RK. A critical role for IL-13 in resistance to intestinal nematode infection. Journal of Immunology. 1998;160(7):3453-61.
45. Hayes KS, Bancroft AJ, Grencis RK. The role of TNF-alpha in Trichuris muris infection I: influence of TNF-alpha receptor usage, gender and IL-13. Parasite Immunology. 2007;29(11):575-82.

181
46. Hepworth MR, Hardman MJ, Grencis RK. The role of sex hormones in the development of Th2 immunity in a gender-biased model of Trichuris muris infection. European Journal of Immunology. 2010;40(2):406-16.
47. Helmby H, Takeda K, Akira S, Grencis RK. Interleukin (IL)-18 promotes the development of chronic gastrointestinal helminth infection by downregulating IL-13. Journal of Experimental Medicine. 2001;194(3):355-64.
48. Bancroft AJ, Else KJ, Humphreys NE, Grencis RK. The effect of challenge and trickle Trichuris muris infections on the polarisation of the immune response. International Journal for Parasitology. 2001;31(14):1627-37.
49. Else KJ, Hultner L, Grencis RK. Modulation of cytokine production and response phenotypes in murine Trichuriasis. Parasite Immunology. 1992;14(4):441-9.
50. Else KJ, Grencis RK. Antibody-independent effector mechanisms in resistance to the intestinal nematode parasite Trichuris muris. Infection and Immunity. 1996;64(8):2950-4.
51. Behnke JM, Wakelin D. The survival of Trichuris muris in wild populations of its natural hosts. Parasitology. 1973;67:157-64.
52. Faulkner H, Turner J, Kamgno J, Pion SD, Boussinesq M, Bradley JE. Age- and infection intensity-dependent cytokine and antibody production in human trichuriasis: The importance of IgE. Journal of Infectious Diseases. 2002;185(5):665-72.
53. Koyama K, Ito Y. Comparative studies on immune responses to infection in susceptible B10.BR mice infected with different strains of the murine nematode parasite Trichuris muris. Parasite Immunology. 1996;18(5):257-63.
54. Bellaby T, Robinson K, Wakelin D. Induction of differential T-helper-cell responses in mice infected with variants of the parasitic nematode Trichuris muris. Infection and Immunity. 1996;64(3):791-5.
55. D'Elia R, Behnke JM, Bradley JE, Else KJ. Regulatory T Cells: A Role in the Control of Helminth-Driven Intestinal Pathology and Worm Survival. Journal of Immunology. 2009;182(4):2340-8.
56. Hasnain SZ, Evans CM, Roy M, Gallagher AL, Kindrachuk KN, Barron L, Dickey BF, Wilson MS, Wynn TA, Grencis RK, Thornton DJ. Muc5ac: a critical component mediating the rejection of enteric nematodes. Journal of Experimental Medicine. 2011;208(5):893-900.
57. Cliffe LJ, Humphreys NE, Lane TE, Potten CS, Booth C, Grencis RK. Accelerated intestinal epithelial cell turnover: A new mechanism of parasite expulsion. Science. 2005;308(5727):1463-5.

182
58. Khan WI, Richard M, Akiho H, Blennerhasset PA, Humphreys NE, Grencis RK, Van Snick J, Collins SM. Modulation of intestinal muscle contraction by interleukin-9 (IL-9) or IL-9 neutralization: Correlation with worm expulsion in murine nematode infections. Infection and Immunity. 2003;71(5):2430-8.
59. Betts CJ, Else KJ. Mast cells, eosinophils and antibody-mediated cellular cytotoxicity are not critical in resistance to Trichuris muris. Parasite Immunology. 1999;21(1):45-52.
60. Svensson M, Bell L, Little MC, deSchoolmeester M, Locksley RM, Else KJ. Accumulation of eosinophils in intestine-draining mesenteric lymph nodes occurs after Trichuris muris infection. Parasite Immunology. 2011;33(1):1-11.
61. Dixon H, Blanchard C, deSchoolmeester ML, Yuill NC, Christie JW, Rothenberg ME, Else KJ. The role of Th2 cytokines, chemokines and parasite products in eosinophil recruitment to the gastrointestinal mucosa during helminth infection. European Journal of Immunology. 2006;36(7):1753-63.
62. Faulkner H, Renauld JC, Van Snick J, Grencis RK. Interleukin-9 enhances resistance to the intestinal nematode Trichuris muris. Infection and Immunity. 1998;66(8):3832-40.
63. Artis D, Mei LW, Keilbaugh SA, He WM, Brenes M, Swain GP, Knight PA, Donaldson DD, Lazar MA, Miller HR, Schad GA, Scott P, Wu GD. RELM beta/FIZZ2 is a goblet cell-specific immune-effector molecule in the gastrointestinal tract. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(37):13596-600.
64. Wakelin D. Immunity to intestinal parasites. Nature. 1978;273(5664):617-20.
65. Miller HRP. Gastrointestinal mucus, a medium for survival and for elimination or parasitic nematodes and protozoa. Parasitology. 1987;94:S77-S100.
66. Wahid FN, Behnke JM, Grencis RK, Else KJ, Bensmith AW. Immunological relationships during primary infection with Heligosomoides polygyrus - TH2 cytokines and primary response phenotypes. Parasitology. 1994;108:461-71.
67. Hasnain SZ, Wang H, Ghia J-E, Haq N, Deng Y, Velcich A, Grencis RK, Thornton DJ, Khan WI. Mucin Gene Deficiency in Mice Impairs Host Resistance to an Enteric Parasitic Infection. Gastroenterology. 2010;138(5):1763-U45.
68. Hasnain SZ, Thornton DJ, Grencis RK. Changes in the mucosal barrier during acute and chronic Trichuris muris infection. Parasite Immunology. 2011;33(1):45-55.
69. Hasnain SZ, Dawson PA, Lourie R, Hutson P, Tong H, Grencis RK, McGuckin MA, Thornton DJ. Immune-driven alterations in mucin sulphation is

183
an important mediator of Trichuris muris helminth expulsion. Plos Pathogens. 2017;13(2).
70. Nair MG, Guild KJ, Du Y, Zaph C, Yancopoulos GD, Valenzuela DM, Murphy A, Stevens S, Karow M, Artis D. Goblet cell-derived resistin-like molecule beta augments CD4(+) T cell production of IFN-gamma and infection-induced intestinal inflammation. Journal of Immunology. 2008;181(7):4709-15.
71. Herbert DR, Yang JQ, Hogan SP, Groschwitz K, Khodoun M, Munitz A, Orekov T, Perkins C, Wang Q, Brombacher F, Urban JF, Rothenberg ME, Finkelman FD. Intestinal epithelial cell secretion of RELM-beta protects against gastrointestinal worm infection. Journal of Experimental Medicine. 2009;206(13):2947-57.
72. Oudhoff MJ, Antignano F, Chenery AL, Burrows K, Redpath SA, Braam MJ, Perona-Wright G, Zaph C. Intestinal Epithelial Cell-Intrinsic Deletion of Setd7 Identifies Role for Developmental Pathways in Immunity to Helminth Infection. Plos Pathogens. 2016;12(9).
73. Lee TDG, Wakelin D. The use of host strain variation to assess the significance of mucosal mast cells in the spontaneous cure response of mice to the nematode Trichuris muris. International Archives of Allergy and Applied Immunology. 1982;67(4):302-5.
74. Koyama K, Ito Y. Mucosal mast cell responses are not required for protection against infection with the murine nematode parasite Trichuris muris. Parasite Immunology. 2000;22(1):13-20.
75. Blackwell NM, Else KJ. A comparison of local and peripheral parasite-specific antibody production in different strains of mice infected with Trichuris muris. Parasite Immunology. 2002;24(4):203-11.
76. Else KJ, Wakelin D, Wassom DL, Hauda KM. MHC-restricted antibody responses to Trichuris muris excretory secretory (E/S) antigen. Parasite Immunology. 1990;12(5):509-27.
77. Selby GR, Wakelin D. Transfer of immunity against Trichuris muris in the serum and cells. International Journal for Parasitology. 1973;3(6):717-22.
78. Blackwell NM, Else KJ. B cells and antibodies are required for resistance to the parasitic gastrointestinal nematode Trichuris muris. Infection and Immunity. 2001;69(6):3860-8.
79. Spits H, Cupedo T. Innate Lymphoid Cells: Emerging Insights in Development, Lineage Relationships, and Function. Annual Review of Immunology, Vol 30. 2012;30:647-75.
80. Perrigoue JG, Saenz SA, Siracusa MC, Allenspach EJ, Taylor BC, Giacomin PR, Nair MG, Du Y, Zaph C, van Rooijen N, Comeau MR, Pearce EJ, Laufer TM, Artis D. MHC class II-dependent basophil-CD4(+) T cell interactions

184
promote T(H)2 cytokine-dependent immunity. Nature Immunology. 2009;10(7):697-U45.
81. Sokol CL, Chu N-Q, Yu S, Nish SA, Laufer TM, Medzhitov R. Basophils function as antigen-presenting cells for an allergen-induced T helper type 2 response. Nature Immunology. 2009;10(7):713-U63.
82. Sokol CL, Barton GM, Farr AG, Medzhitov R. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nature Immunology. 2008;9(3):310-8.
83. Hepworth MR, Grencis RK. Disruption of Th2 Immunity Results in a Gender-Specific Expansion of IL-13 Producing Accessory NK Cells during Helminth Infection. Journal of Immunology. 2009;183(6):3906-14.
84. Humphreys NE, Xu D, Hepworth MR, Liew FY, Grencis RK. IL-33, a potent inducer of adaptive immunity to intestinal nematodes. Journal of Immunology. 2008;180(4):2443-9.
85. Hung L-Y, Lewkowich IP, Dawson LA, Downey J, Yang Y, Smith DE, Herbert DR. IL-33 drives biphasic IL-13 production for noncanonical Type 2 immunity against hookworms. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(1):282.
86. Yasuda K, Muto T, Kawagoe T, Matsumoto M, Sasaki Y, Matsushita K, Taki Y, Futatsugi-Yumikara S, Tsutsui H, Ishii KJ, Yoshimoto T, Akira S, Nakanishi K. Contribution of IL-33-activated type II innate lymphoid cells to pulmonary eosinophilia in intestinal nematode-infected mice. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(9):3451-6.
87. Owyang AM, Zaph C, Wilson EH, Guild KJ, McClanahan T, Miller HRP, Cua DJ, Goldschimdt M, Hunter CA, Kastelein RA, Artis D. Interleukin 25 regulates type 2 cytokine-dependent immunity and limits chronic inflammation in the gastrointestinal tract. Journal of Experimental Medicine. 2006;203(4):843-9.
88. Price AE, Liang H-E, Sullivan BM, Reinhardt RL, Eisley CJ, Erle DJ, Locksley RM. Systemically dispersed innate IL-13-expressing cells in type 2 immunity. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(25):11489-94.
89. Fallon PG, Ballantyne SJ, Mangan NE, Barlow JL, Dasvarma A, Hewett DR, McIlgorm A, Jolin HE, McKenzie AN. Identification of an interleukin (IL)-25-dependent cell population that provides IL-4, IL-5, and IL-13 at the onset of helminth expulsion. Journal of Experimental Medicine. 2006;203(4):1105-16.
90. Saenz SA, Siracusa MC, Perrigoue JG, Spencer SP, Urban JF, Tocker JE, Budelsky AL, Kleinschek MA, Kastelein RA, Kambayashi T, Bhandoola A, Artis D. IL25 elicits a multipotent progenitor cell population that promotes T(H)2 cytokine responses. Nature. 2010;464(7293):1362-U8.

185
91. Taylor BC, Zaph C, Troy AE, Du Y, Guild KJ, Comeau MR, Artis D. TSLP regulates intestinal immunity and inflammation in mouse models of helminth infection and colitis. Journal of Experimental Medicine. 2009;206(3):655-67.
92. Levison SE, McLaughlin JT, Zeef LAH, Fisher P, Grencis RK, Pennock JL. Colonic Transcriptional Profiling in Resistance and Susceptibility to Trichuriasis: Phenotyping a Chronic Colitis and Lessons for latrogenic Helminthosis. Inflammatory Bowel Diseases. 2010;16(12):2065-79.
93. Schopf LR, Hoffmann KF, Cheever AW, Urban JF, Wynn TA. IL-10 is critical for host resistance and survival during gastrointestinal helminth infection. Journal of Immunology. 2002;168(5):2383-92.
94. Collison LW, Chaturvedi V, Henderson AL, Giacomin PR, Guy C, Bankoti J, Finkelstein D, Forbes K, Workman CJ, Brown SA, Rehg JE, Jones ML, Ni HT, Artis D, Turk MJ, Vignali DA. IL-35-mediated induction of a potent regulatory T cell population. Nature Immunology. 2010;11(12):1093-U97.
95. Veldhoen M, Uyttenhove C, van Snick J, Helmby H, Westendorf A, Buer J, Martin B, Wilhelm C, Stockinger B. Transforming growth factor-beta 'reprograms' the differentiation of T helper 2 cells and promotes an interleukin 9-producing subset. Nature Immunology. 2008;9(12):1341-6.
96. Worthington JJ, Klementowicz JE, Rahman S, Czajkowska BI, Smedley C, Waldmann H, Sparwasser T, Grencis RK, Travis MA. Loss of the TGF beta-Activating Integrin alpha v beta 8 on Dendritic Cells Protects Mice from Chronic Intestinal Parasitic Infection via Control of Type 2 Immunity. Plos Pathogens. 2013;9(10).
97. Lightowlers MW. Control of Taenia solium taeniasis/cysticercosis: past practices and new possibilities. Parasitology. 2013;140(13):1566-77.
98. Lightowlers MW, Colebrook AL, Gauci CG, Gauci SM, Kyngdon CT, Monkhouse JL, Vallejo Rodriguez C, Read AJ, Rolfe RA, Sato C. Vaccination against cestode parasites: anti-helminth vaccines that work and why. Veterinary Parasitology. 2003;115(2):83-123.
99. Bottazzi ME. The human hookworm vaccine: recent updates and prospects for success. Journal of Helminthology. 2015;89(5):540-4.
100. Hotez PJ, Diemert D, Bacon KM, Beaumier C, Bethony JM, Bottazzi ME, Brooker S, Couto AR, Freire Mda S, Homma A, Lee BY, Loukas A, Loblack M, Morel CM, Oliveira RC, Russell PK. The Human Hookworm Vaccine. Vaccine. 2013;31:B227-B32.
101. Bethony J, Loukas A, Smout M, Brooker S, Mendez S, Plieskatt J, Goud G, Bottazzi ME, Zhan B, Wang Y, Williamson A, Lustigman S, Correa-Oliveira R, Xiao S, Hotez PJ. Antibodies against a secreted protein from hookworm larvae reduce the intensity of hookworm infection in humans and vaccinated laboratory animals. Faseb Journal. 2005;19(9):1743-+.

186
102. Goud GN, Zhan B, Ghosh K, Loukas A, Hawdon J, Dobardzic A, Deumic V, Liu S, Dobardzic R, Zook BC, Jin Q, Liu Y, Hoffman L, Chung-Debose S, Patel R, Mendez S, Hotez PJ. Cloning, yeast expression, isolation, and vaccine testing of recombinant Ancylostoma-secreted protein (ASP)-1 and ASP-2 from Ancylostoma ceylanicum. Journal of Infectious Diseases. 2004;189(5):919-29.
103. Diemert DJ, Pinto AG, Freire J, Jariwala A, Santiago H, Hamilton RG, Periago MV, Loukas A, Tribolet L, Mulvenna J, Correa-Oliveira R, Hotez PJ, Bethony. Generalized urticaria induced by the Na-ASP-2 hookworm vaccine: Implications for the development of vaccines against helminths. Journal of Allergy and Clinical Immunology. 2012;130(1):169-+.
104. Goud GN, Deumic V, Gupta R, Brelsford J, Zhan B, Gillespie P,Plieskatt JL, Tsao EI, Hotez PJ, Bottazzi ME. Expression, purification, and molecular analysis of the Necator americanus glutathione S-transferase 1 (Na-GST-1): A production process developed for a lead candidate recombinant hookworm vaccine antigen. Protein Expression and Purification. 2012;83(2):145-51.
105. Seid CA, Curti E, Jones RM, Hudspeth E, Rezende W, Pollet J, Center L, Versteeg L, Pritchard S, Musiychuk, Yusibov V, Hotez PJ, Bottazzi ME. Expression, purification, and characterization of the Necator americanus aspartic protease-1 (Na-APR-1 (M74)) antigen, a component of the bivalent human hookworm vaccine. Human Vaccines & Immunotherapeutics. 2015;11(6):1474-88.
106. Pearson MS, Jariwala AR, Abbenante G, Plieskatt J, Wilson D, Bottazzi ME, Hotez PJ, Keegan B, Bethony JM, Loukas A. New tools for NTD vaccines: A case study of quality control assays for product development of the human hookworm vaccine Na-APR-1M74. Human Vaccines & Immunotherapeutics. 2015;11(5):1251-7.
107. Hotez PJ, Beaumier CM, Gillespie PM, Strych U, Hayward T, Bottazzi ME. Advancing a vaccine to prevent hookworm disease and anemia. Vaccine. 2016;34(26):3001-5.
108. Tsuji N, Suzuki K, Kasuga-Aoki H, Matsumoto Y, Arakawa T, Ishiwata K, Isobe T. Intranasal immunization with recombinant Ascaris suum 14-kilodalton antigen coupled with cholera toxin B subunit induces protective immunity to A. suum infection in mice. Infection and Immunity. 2001;69(12):7285-92.
109. Tsuji N, Suzuki K, Kasuga-Aoki H, Isobe T, Arakawa T, Matsumoto Y. Mice intranasally immunized with a recombinant 16-kilodalton antigen from roundworm Ascaris parasites are protected against larval migration of Ascaris suum. Infection and Immunity. 2003;71(9):5314-23.
110. Islam MK, Miyoshi T, Tsuji N. Vaccination with recombinant Ascaris suum 24-kilodalton antigen induces a Th1/Th2-mixed type immune response and confers high levels of protection against challenged Ascaris suum lung-stage infection in BALB/c mice. International Journal for Parasitology. 2005;35(9):1023-30.

187
111. Islam MK, Miyoshi T, Yokomizo Y, Tsuji N. Molecular cloning and partial characterization of a nematode-specific 24 kDa protein from Ascaris suum. Parasitology. 2005;130:131-9.
112. Tsuji N, Kasuga-Aoki H, Isobe T, Arakawa T, Matsumoto Y. Cloning and characterisation of a highly immunoreactive 37 kDa antigen with multi-immunoglobulin domains from the swine roundworm Ascaris suum. International Journal for Parasitology. 2002;32(14):1739-46.
113. Chen N, Yuan Z-G, Xu MJ, Zhou D-H, Zhang XX, Zhang YZ, Wang XW, Yan C, Lin RQ, Zhu XQ. Ascaris suum enolase is a potential vaccine candidate against ascariasis. Vaccine. 2012;30(23):3478-82.
114. Chen N, Xu MJ, Nisbet AJ, Huang CQ, Lin RQ, Yuan ZG, Song HQ, Zhu XQ. Ascaris suum: RNAi mediated silencing of enolase gene expression in infective larvae. Experimental Parasitology. 2011;127(1):142-6.
115. Jenkins SA, Wakelin D. The source and nature of some functional antigens of Trichuris muris. Parasitology. 1977.
116. Robinson K, Bellaby T, Wakelin D. Efficacy of oral vaccination against the murine intestinal parasite Trichuris muris is dependent upon host genetics. Infection and Immunity. 1995;63(5):1762-6.
117. Hasnain SZ, McGuckin MA, Grencis RK, Thornton DJ. Serine Protease(s) Secreted by the Nematode Trichuris muris Degrade the Mucus Barrier. Plos Neglected Tropical Diseases. 2012;6(10).
118. Hewitson JP, Grainger JR, Maizels RM. Helminth immunoregulation: The role of parasite secreted proteins in modulating host immunity. Molecular and Biochemical Parasitology. 2009;167(1):1-11.
119. Fawzi EM, Bustos TC, Samblas MG, Gonzalez-Gonzalez G, Solano J, Gonzalez-Sanchez ME, De Pablos LM, Corral-Caridad MJ, Cuquerella M, Osuna A, Alunda JM. Intranasal Immunization of Lambs with Serine/Threonine Phosphatase 2A against Gastrointestinal Nematodes. Clinical and Vaccine Immunology. 2013;20(9):1352-9.
120. Tendler M, Almeida M, Simpson A. Development of the Brazilian anti Schistosomiasis vaccine based on the recombinant fatty acid binding protein Sm14 plus GLA-SE adjuvant. Frontiers in Immunology. 2015;6.
121. Rodriguezperez J, Rodriguezmedina JR, Garciablanco MA, Hillyer GV. Fasciola hepatica - molecular cloning, nucleotide sequence, and expression of a gene encoding a polypeptide homologous to a Schistosoma mansoni fatty acid binding protein. Experimental Parasitology. 1992;74(4):400-7.
122. Tendler M, Brito CA, Vilar MM, SerraFreire N, Diogo CM, Almeida MS, Delbem AC, Da Silva JF, Savino W, Garratt RC, Katz N, Simpson AS. A Schistosoma mansoni fatty acid-binding protein, Sm14, is the potential basis of

188
a dual-purpose anti-helminth vaccine. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(1):269-73.
123. Lee JS, Kim IS, Sohn WM, Lee J, Yong TS. A DNA vaccine encoding a fatty acid-binding protein of Clonorchis sinensis induces protective immune response in Sprague-Dawley rats. Scandinavian Journal of Immunology. 2006;63(3):169-76.
124. Chabalgoity JA, Harrison JA, Esteves A, deHormaeche RD, Ehrlich R, Khan CMA, Hormaeche CE. Expression and immunogenicity of an Echinococcus granulosus fatty acid-binding protein in live attenuated Salmonella vaccine strains. Infection and Immunity. 1997;65(6):2402-12.
125. Miller HM. Acquired immunity against a metazoan parasite by use of non-specific worm material. Proc, Soc Exp Biol and Med. 1932;29(1125-1126).
126. Khan K. Immunological studies of Cysticercus fasciolaris. Keio Igaka. 1935;14:663–87.
127. Campbell. The specific protective property of serum from rats infected with Cysticercus crassicollis. Journal of Immunology. 1938;35:195–204.
128. Harrison GB, Heath DD, Dempster RP, Gauci C, Newton SE, Cameron WG, Robinson CM, Lawrence SB, Lightowlers MW, Rickard MD. Identification and cDNA cloning of two novel low molecular weight host-protective antigens from Taenia ovis oncospheres. International Journal of Parasitology. 1996;26:195-204.
129. Harrison GB, Heath DD, Dempster RP, Lawrence SB, Lightowlers MW, Rickard MD. Identification of host-protective antigens of Taenia ovis oncospheres. International Journal of Parasitology. 1993;23:41-50.
130. Assana E, Kyngdon CT, Gauci CG, Geerts S, Dorny P, De Deken R, Anderson GA, Zoli AP, Lightowlers MW. Elimination of Taenia solium transmission to pigs in a field trial of the TSOL18 vaccine in Cameroon. International Journal for Parasitology. 2010;40(5):515-9.
131. Lightowlers MW. Eradication of Taenia solium cysticercosis: A role for vaccination of pigs. International Journal for Parasitology. 2010;40(10):1183-92.
132. Lightowlers MW, Rolfe R, Gauci CG. Taenia saginata: Vaccination against cysticercosis in cattle with recombinant oncosphere antigens. Experimental Parasitology. 1996;84(3):330-8.
133. Lightowlers MW, Lawrence SB, Gauci CG, Young J, Ralston MJ, Maas D, Heath DD. Vaccination against hydatidosis using a defined recombinant antigen. Parasite Immunology. 1996;18(9):457-62.
134. Lightowlers MW, Jensen O, Fernandez E, Iriarte JA, Woollard DJ, Cauci CG, Jenkins DJ, Heath DD. Vaccination trials in Australia and Argentina confirm

189
the effectiveness of the EG95 hydatid vaccine in sheep. International Journal for Parasitology. 1999;29(4):531-4.
135. Lebre F, Hearnden CH, Lavelle EC. Modulation of Immune Responses by Particulate Materials. Advanced Materials. 2016;28(27):5525-41.
136. Elliott SL, Suhrbier A, Miles JJ, Lawrence G, Pye SJ, Le TT, Rosenstengel A, Nguyen T, Allworth A, Burrows SR, Cox J, Pye D, Moss DJ, Bharadwaj M. Phase I trial of a CD8(+) T-Cell peptide epitope-based vaccine for infectious mononucleosis. Journal of Virology. 2008;82(3):1448-57.
137. Sabet LP, Taheri T, Memarnejadian A, Azad TM, Asgari F, Rahimnia R, Alavian SM, Rafati S, Samimi Rad K. Immunogenicity of Multi-Epitope DNA and Peptide Vaccine Candidates Based on Core, E2, NS3 and NS5B HCV Epitopes in BALB/c Mice. Hepatitis Monthly. 2014;14(10).
138. Koopman G, Beenhakker N, Nieuwenhuis I, Doxiadis G, Mooij P, Drijfhout JW, Koestler J, Hanke T, Fagrouch Z, Verschoor EJ, Bontrop RE, Wagner R, Bogers WM, Melief CJ. DNA/long peptide vaccination against conserved regions of SIV induces partial protection against SIVmac251 challenge. Aids. 2013;27(18):2841-51.
139. Hermsen CC, Verhage DF, Telgt DS, Teelen K, Bousema JT, Roestenberg M, Bolad A, Berzins K, Corradin G, Leroy O, Theisen M, Sauerwein RW.Glutamate-rich protein (GLURP) induces antibodies that inhibit in vitro growth of Plasmodium falciparum in a phase 1 malaria vaccine trial. Vaccine. 2007;25(15):2930-40.
140. Pierce MA, Ellis RD, Martin LB, Malkin E, Tierney E, Miura K, Fay MP, Marjason J, Elliott SL, Mullen GE, Rausch K, Zhu D, Long CA, Miller LH.
Phase 1 safety and immunogenicity trial of the Plasmodium falciparum blood-stage malaria vaccine AMA1-C1/ISA 720 in Australian adults. Vaccine. 2010;28(10):2236-42.
141. Audran R, Cachat M, Lurati F, Soe S, Leroy O, Corradin G, Druilhe P, Spertini F. Phase I malaria vaccine trial with a long synthetic peptide derived from the merozoite surface protein 3 antigen. Infection and Immunity. 2005;73(12):8017-26. 142. Ricciardi A, Visitsunthorn K, Dalton JP, Ndao M. A vaccine consisting of Schistosoma mansoni cathepsin B formulated in Montanide ISA 720 VG induces high level protection against murine schistosomiasis. Bmc Infectious Diseases. 2016;16.
143. Marrack P, McKee AS, Munks MW. Towards an understanding of the adjuvant action of aluminium. Nature Reviews Immunology. 2009;9(4):287-93.
144. Hutchison S, Benson R, Gibson V, Pollock A, Garside P, Brewer J. Antigen depot is not required for alum adjuvanticity. FASEB Journal. 2012;26(3):1272-9.

190
145. Marichal T1, Ohata K, Bedoret D, Mesnil C, Sabatel C, Kobiyama K, Lekeux P, Coban C, Akira S, Ishii KJ, Bureau F, Desmet CJ. DNA released from dying host cells mediates aluminum adjuvant activity. Nature Medicine. 2011;17(8):996-U118. 146. Sharp FA1, Ruane D, Claass B, Creagh E, Harris J, Malyala P, Singh M, O'Hagan DT, Pétrilli V, Tschopp J, O'Neill LA, Lavelle EC. Uptake of particulate vaccine adjuvants by dendritic cells activates the NALP3 inflammasome. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(3):870-5. 147. Li HF, Willingham SB, Ting JPY, Re F. Cutting edge: Inflammasome activation by alum and alum's adjuvant effect are mediated by NLRP3. Journal of Immunology. 2008;181(1):17-21.
148. Eisenbarth SC, Colegio OR, O'Connor W, Sutterwala FS, Flavell RA. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature. 2008;453(7198):1122-U13.
149. Duewell P1, Kisser U, Heckelsmiller K, Hoves S, Stoitzner P, Koernig S, Morelli AB, Clausen BE, Dauer M, Eigler A, Anz D, Bourquin C, Maraskovsky E, Endres S, Schnurr M. ISCOMATRIX Adjuvant Combines Immune Activation with Antigen Delivery to Dendritic Cells In Vivo Leading to Effective Cross-Priming of CD8(+) T Cells. Journal of Immunology. 2011;187(1):55-63. 150. Wilson NS1, Yang B, Morelli AB, Koernig S, Yang A, Loeser S, Airey D, Provan L, Hass P, Braley H, Couto S, Drane D, Boyle J, Belz GT, Ashkenazi A, Maraskovsky E. ISCOMATRIX vaccines mediate CD8(+) T-cell cross-priming by a MyD88-dependent signaling pathway. Immunology and Cell Biology. 2012;90(5):540-52. 151. Frazer IH1, Quinn M, Nicklin JL, Tan J, Perrin LC, Ng P, O'Connor VM, White O, Wendt N, Martin J, Crowley JM, Edwards SJ, McKenzie AW, Mitchell SV, Maher DW, Pearse MJ, Basser RL. Phase 1 study of HPV16-specific immunotherapy with E6E7 fusion protein and ISCOMATRIX (TM) adjuvant in women with cervical intraepithelial neoplasia. Vaccine. 2004;23(2):172-81. 152. Marcilla A, Trelis M, Cortes A, Sotillo J, Cantalapiedra F, Teresa Minguez M, Valero ML, Sanchez del Pino MM, Munoz-Antoli C, Toledo R, Bernal D. Extracellular Vesicles from Parasitic Helminths Contain Specific Excretory/Secretory Proteins and Are Internalized in Intestinal Host Cells. Plos One. 2012;7(9).
153. Coakley G, Maizels RM, Buck AH. Exosomes and Other Extracellular Vesicles: The New Communicators in Parasite Infections. Trends in Parasitology. 2015;31(10):477-89.

191
154. Hansen EP, Kringel H, Williams AR, Nejsum P. Secretion of RNA containing extracellular vesicles by the porcine whipworm, Trichuris suis. Journal of Parasitology. 2015;101(3):336-40.
155. Coakley G, McCaskill JL, Borger JG, Simbari F, Robertson E, Millar M, Harcus Y, McSorley HJ, Maizels RM, Buck AH. Extracellular Vesicles from a Helminth Parasite Suppress Macrophage Activation and Constitute an Effective Vaccine for Protective Immunity. Cell Reports. 2017;19:1545-57.
156. Trelis M, Galiano A, Bolado A, Toledo R, Marcilla A, Bernal D. Subcutaneous injection of exosomes reduces symptom severity and mortality induced by Echinostoma caproni infection in BALB/c mice. International Journal for Parasitology. 2016;46(12):799-808.
157. Mathivanan S, Ji H, Simpson RJ. Exosomes: Extracellular organelles important in intercellular communication. Journal of Proteomics. 2010;73(10):1907-20.
158. Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro - selective externalization of the receptor. Cell. 1983;33(3):967-77.
159. Harding C, Heuser J, Stahl P. Receptor mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. Journal of Cell Biology. 1983;97(2):329-39.
160. Raposo G, Nijman HW, Stoorvogel W, Leijendekker R, Harding CV, Melief CJM, Geuze HJ. B lymphocytes secrete antigen-presenting vesicles. Journal of Experimental Medicine. 1996;183(3):1161-72.
161. Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nature Cell Biology. 2007;9(6):654-U72.
162. Silverman JM, Reiner NE. Leishmania exosomes deliver preemptive strikes to create an environment permissive for early infection. Frontiers in Cellular and Infection Microbiology. 2012;1:1-8.
163. Silverman JM, Clos J, Horakova E, Wang AY, Wiesgigl M, Kelly I, Lynn MA, McMaster WR, Foster LJ, Levings MK, Reiner NE. Leishmania Exosomes Modulate Innate and Adaptive Immune Responses through Effects on Monocytes and Dendritic Cells. Journal of Immunology. 2010;185(9):5011-22.
164. Trocoli Torrecilhas AC, Tonelli RR, Pavanelli WR, da Silva JS, Schumacher RI, de Souza W, E Silva NC, de Almeida Abrahamsohn I, Colli W, Manso Alves MJ. Trypanosoma cruzi: parasite shed vesicles increase heart parasitism and generate an intense inflammatory response. Microbes and Infection. 2009;11(1):29-39.
165. Buck AH, Coakley G, Simbari F, McSorley HJ, Quintana JF, Le Bihan T, Kumar S, Abreu-Goodger C, Lear M, Harcus Y, Ceroni A, Babayan SA, Blaxter

192
M, Ivens A, Maizels RM. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nature Communications. 2014;5.
166. Cvjetkovic A, Lotvall J, Lasser C. The influence of rotor type and centrifugation time on the yield and purity of extracellular vesicles. Journal of Extracellular Vesicles. 2013. 167. Hanson PI, Cashikar A. Multivesicular Body Morphogenesis. Annual Review of Cell and Developmental Biology, Vol 28. 2012;28:337-62.
168. Coakley G, Buck AH, Maizels RM. Host parasite communications-Messages from helminths for the immune system Parasite communication and cell-cell interactions. Molecular and Biochemical Parasitology. 2016;208(1):33-40.
169. Wang L, Li Z, Shen J, Liu Z, Liang J, Wu X, Sun X, Wu Z. Exosome-like vesicles derived by Schistosoma japonicum adult worms mediates M1 type immune- activity of macrophage. Parasitology Research. 2015;114(5):1865-73.
170. Virginio VG, Monteiro KM, Drumond F, de Carvalho MO, Vargas DM, Zaha A, Ferreira HB. Excretory/secretory products from in vitro-cultured Echinococcus granulosus protoscoleces. Molecular and Biochemical Parasitology. 2012;183(1):15-22.
171. dos Santos GB, Monteiro KM, da Silva ED, Battistella ME, Ferreira HB, Zaha A. Excretory/secretory products in the Echinococcus granulosus metacestode: is the intermediate host complacent with infection caused by the larval form of the parasite? International Journal for Parasitology. 2016;46(13-14):843-56.
172. Bernal D, Trelis M, Montaner S, Cantalapiedra F, Galiano A, Hackenberg M, Marcilla A. Surface analysis of Dicrocoelium dendriticum. The molecular characterization of exosomes reveals the presence of miRNAs. Journal of Proteomics. 2014;105:232-41.
173. Sotillo J, Pearson M, Potriquet J, Becker L, Pickering D, Mulvenna J, Loukas A. Extracellular vesicles secreted by Schistosoma mansoni contain protein vaccine candidates. International Journal for Parasitology. 2016;46(1):1-5.
174. Espino AM, Hillyer GV. Molecular cloning of a member of the Fasciola hepatica saposin-like protein family. Journal of Parasitology. 2003;89(3):545-52.
175. Tran MH, Pearson MS, Jeffrey MB, Smyth DJ, Jones MK, Duke M, Don TA, McManus DP, Correa-Oliveira R, Loukas A. Tetraspanins on the surface of Schistosoma mansoni are protective antigens against schistosomiasis. Nature Medicine. 2006;12(7):835-40.
176. Cardoso FC, Pacifico RNA, Mortara RA, Oliveira SC. Human antibody responses of patients living in endemic areas for schistosomiasis to the

193
tegumental protein Sm29 identified through genomic studies. Clinical and Experimental Immunology. 2006;144(3):382-91.
177. Rezende CMF, Silva MR, Santos IGD, Silva GAB, Gomes DA, Goes AM. Immunization with rP22 induces protective immunity against Schistosoma mansoni: Effects on granuloma down-modulation and cytokine production. Immunology Letters. 2011;141(1):123-33.
178. Nowacki FC, Swain MT, Klychnikov OI, Niazi U, Ivens A, Quintana JF, Hensbergen PJ, Hokke CH, Buck AH, Hoffmann KF. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. Journal of Extracellular Vesicles. 2015;4.
179. Zamanian M, Fraser LM, Agbedanu PN, Harischandra H, Moorhead AR, Day TA, Bartholomay LC, Kimber MJ. Release of Small RNA-containing Exosome-like Vesicles from the Human Filarial Parasite Brugia malayi. Plos Neglected Tropical Diseases. 2015;9(9).
180. Tritten L, Tam M, Vargas M, Jardim A, Stevenson MM, Keiser J, Geary TG. Excretory/secretory products from the gastrointestinal nematode Trichuris muris. Experimental Parasitology. 2017;178:30-36.
181. Tzelos T, Matthews JB, Buck AH, Simbari F, Frew D, Inglis NF, McLean K, Nisbet AJ, Whitelaw CB, Knox DP, McNeilly TN. A preliminary proteomic characterisation of extracellular vesicles released by the ovine parasitic nematode, Teladorsagia circumcincta. Veterinary Parasitology. 2016;221:84-92.
182. Tritten L, Clarke D, Timmins S, McTier T, Geary TG. Dirofilaria immitis exhibits sex- and stage-specific differences in excretory/secretory miRNA and protein profiles. Veterinary Parasitology. 2016;232:1-7.
183. Eichenberger RM, Talukder MH, Field MA, Wangchuk P, Giacomin P, Loukas A, Sotillo J. Characterisation of Trichuris muris secreted proteins and extracellular vesicles provides new insights into host-parasite communication. BioRxiv. 2017. 184. McKenzie GJ, Bancroft A, Grencis RK, McKenzie ANJ. A distinct role for interleukin-13 in Th2-cell-mediated immune responses. Current Biology. 1998;8(6):339-42. 185. Hayes KS, Cliffe LJ, Bancroft AJ, Forman SP, Thompson S, Booth C,
Grencis RK. Chronic Trichuris muris infection causes neoplastic change in the intestine and exacerbates tumour formation in APC min/+ mice. PLoS Neglected Tropical Diseases. 2017.
186. Malvern. Dynamic light scattering common terms defined. 2011.
187. Hollander MaW, D.A. Nonparametric Statistical Methods: John Wiley & Sons.; 1973.

194
188. Keiser J, Utzinger J. The Drugs We Have and the Drugs We Need Against Major Helminth Infections. Advances in Parasitology, Vol 73: Important Helminth Infections in Southeast Asia: Diversity and Potential for Control and Elimination, Pt B. 2010;73:197-230.
189. Khan WI, Motomura Y, Blennerhassett PA, Kanbayashi H, Varghese AK, El-Sharkawy RT, Gauldie J, Collins SM. Disruption of CD40-CD40 ligand pathway inhibits the development of intestinal muscle hypercontractility and protective immunity in nematode infection. American Journal of Physiology-Gastrointestinal and Liver Physiology. 2005;288(1):G15-G22.
190. Amanna IJ, Slifka MK, Crotty S. Immunity and immunological memory following smallpox vaccination. Immunological Reviews. 2006;211:320-37.
191. Zhang X, Wang Q, Bi Y, Kou Z, Zhou J, Cui Y, Yan Y, Zhou L, Tan Y, Yang H, Du Z, Han Y, Song Y, Zhang P, Zhou D, Yang R, Wang X. Kinetics of Memory B Cell and Plasma Cell Responses in the Mice Immunized with Plague Vaccines. Scandinavian Journal of Immunology. 2014;79(3):157-62.
192. Pepper M, Linehan JL, Pagan AJ, Zell T, Dileepan T, Cleary PP, Jenkins MK. Different routes of bacterial infection induce long-lived T(H)1 memory cells and short-lived T(H)17 cells. Nature Immunology. 2010;11(1):83-U2.
193. Brady RC, Treanor JJ, Atmar RL, Keitel WA, Edelman R, Chen WH, Winokur P, Belshe R, Graham IL, Noah DL, Guo K, Hill H. Safety and immunogenicity of a subvirion inactivated influenza A/H5N1 vaccine with or without aluminum hydroxide among healthy elderly adults. Vaccine. 2009;27(37):5091-5. 194. Keitel WA1, Campbell JD, Treanor JJ, Walter EB, Patel SM, He F, Noah DL, Hill H. Safety and immunogenicity of an inactivated influenza A/H5N1 vaccine given with or without aluminum hydroxide to healthy adults: Results of a phase I-II randomized clinical trial. Journal of Infectious Diseases. 2008;198(9):1309-16. 195. Verdijk P, Rots NY, van Oijen MG, Weldon WC, Oberste MS, Okayasu H, Sutter RW, Bakker WA.
Safety and immunogenicity of a primary series of Sabin-IPV with and without aluminum hydroxide in infants. Vaccine. 2014;32(39):4938-44.
196. Schmidt CS, White CJ, Ibrahim AS, Filler SG, Fu Y, Yeaman MR, Edwards JE, Hennessey JP. NDV-3, a recombinant alum-adjuvanted vaccine for Candida and Staphylococcus aureus, is safe and immunogenic in healthy adults. Vaccine. 2012;30(52):7594-600.
197. Bancroft AJ, Else KJ, Grencis RK. Low level infection with Trichuris muris significantly affects the polarization of the CD4 response. European Journal of Immunology. 1994;24(12):3113-8.

195
198. Else KJ, Hultner L, Grencis RK. Cellular immune responses to the murine nematode parasite Trichuris muris. 2. Differential induction of the TH cell subsets in resistant versus susceptible mice. Immunology. 1992;75(2):232-7.
199. Smith NMS, Wasserman GA, Coleman FT, Hilliard KL, Yamamoto K, Lipsitz E, Malley R, Dooms H, Jones MR, Quinton LJ, Mizgerd JP. Regionally compartmentalized resident memory T cells mediate naturally acquired protection against pneumococcal pneumonia. Mucosal Immunology. 2017. 200. Hewitson JP, Ivens AC, Harcus Y, Filbey KJ, McSorley HJ, Murray J, Bridgett S, Ashford D, Dowle AA, Maizels RM. Secretion of Protective Antigens by Tissue-Stage Nematode Larvae Revealed by Proteomic Analysis and Vaccination-Induced Sterile Immunity. Plos Pathogens. 2013;9(8). 201. Neilson KA, Ali NA, Muralidharan S, Mirzaei M, Mariani M, Assadourian G, Lee A, van Sluyter SC, Haynes PA. Less label, more free: Approaches in label-free quantitative mass spectrometry. Proteomics. 2011;11(4):535-53.
202. Healthcare G. Superose 12 Overview [Available from: http://www.gelifesciences.com/webapp/wcs/stores/servlet/productById/en/GELifeSciences-uk/17517301.
203. Healthcare G. Superdex 75 10/300 GL and 5/150 G [Available from: http://www.gelifesciences.com/webapp/wcs/stores/servlet/productById/en/GELifeSciences-uk/17517401.
204. Santos LN, Gallo MBC, Silva ES, Figueiredo CAV, Cooper PJ, Barreto ML, Loureiro S, Pontes-de-Carvalho LC, Alcantara-Neves NM. A proteomic approach to identify proteins from Trichuris trichiura extract with immunomodulatory effects. Parasite Immunology. 2013;35(5-6):188-93.
205. Redmond DL, Smith SK, Halliday A, Smith WD, Jackson F, Knox DP, Matthews JB. An immunogenic cathepsin F secreted by the parasitic stages of Teladorsagia circumcincta. International Journal for Parasitology. 2006;36(3):277-86.
206. Nisbet AJ, McNeilly TN, Wildblood LA, Morrison AA, Bartley DJ, Bartley Y, Longhi C, McKendrick IJ, Palarea-Albaladejo J, Matthews JB. Successful immunization against a parasitic nematode by vaccination with recombinant proteins. Vaccine. 2013;31(37):4017-23.
207. Lee BH, Hwang DM, Palaniyar N, Grinstein S, Philpott DJ, Hu J. Activation of P2X(7) receptor by ATP plays an important role in regulating inflammatory responses during acute viral infection. PloS one. 2012;7(4):e35812-e.
208. Goncalves AM, Pedro AQ, Maia C, Sousa F, Queiroz JA, Passarinha LA. Pichia pastoris: A Recombinant Microfactory for Antibodies and Human Membrane Proteins. Journal of Microbiology and Biotechnology. 2013;23(5):587-601.

196
209. Matthews JB, Geldhof P, Tzelos T, Claerebout E. Progress in the development of subunit vaccines for gastrointestinal nematodes of ruminants. Parasite Immunology. 2016;38(12):744-53.
210. Wilson IBH, Paschinger K. Sweet secrets of a therapeutic worm: mass-spectrometric N-glycomic analysis of Trichuris suis. Analytical and Bioanalytical Chemistry. 2016;408(2):461-71.
211. Hokke C, van Diepen A. Helminth glycomics – glycan repertoires and host-parasite interactions. Molecular & Biochemical Parasitology. 2017;215:47-57. 212. Urquhart GM, Jarrett WFH, Jennings FW, McIntyre WI, Mulligan W. Immunity to Haemonchus contortus infection - relationship between age and successful vaccination with irradiated larvae. American Journal of Veterinary Research. 1966;27(121):1645-.
213. Smith WD. Protection in lambs immunized with Haemonchus contortus gut membrane proteins. Research in Veterinary Science. 1993;54(1):94-101.
214. Smith SK, Pettit D, Newlands GF, Redmond DL, Skuce PJ, Knox DP, Smith WD. Further immunization and biochemical studies with a protective antigen complex from the microvillar membrane of the intestine of Haemonchus contortus. Parasite Immunology. 1999;21(4):187-99. 215. Smith TS, Munn EA, Graham M, Tavernor AS, Greenwood CA. Purification and evaluation of the integral membrane protein H11 as a protective antigen against Haemonchus contortus. International Journal for Parasitology. 1993;23(2):271-80.
216. Smith SK, Smith WD. Immunisation of sheep with an integral membrane glycoprotein complex of Haemonchus contortus and with its major polypeptide components. Research in Veterinary Science. 1996;60(1):1-6.
217. Smith WD, Smith SK, Murray JM. Protection studies with integral membrane fractions of Haemonchus contortus. Parasite Immunology. 1994;16(5):231-41.
218. Knox DP, Redmond DL, Skuce PJ, Newlands GFJ. The contribution of molecular biology to the development of vaccines against nematode and trematode parasites of domestic ruminants. Veterinary Parasitology. 2001;101(3-4):311-35.
219. Newton SE, Meeusen ENT. Progress and new technologies for developing vaccines against gastrointestinal nematode parasites of sheep. Parasite Immunology. 2003;25(5):283-96.
220. Reszka N, Rijsewijk FAM, Zelnik V, Moskwa B, Bienkowska-Szewczyk K. Haemonchus contortus: Characterization of the baculovirus expressed form of aminopeptidase H11. Experimental Parasitology. 2007;117(2):208-13.

197
221. Geldhof P, Claerebout E, Knox D, Vercauteren I, Looszova A, Vercruysse J. Vaccination of calves against Ostertagia ostertagi with cysteine proteinase enriched protein fractions. Parasite Immunology. 2002;24(5):263-70.
222. Geldhof P, Vercauteren I, Gevaert K, Staes A, Knox DP, Vandekerckhove J, Vercruysse J, Claerebout E. Activation-associated secreted proteins are the most abundant antigens in a host protective fraction from Ostertagia ostertagi. Molecular and Biochemical Parasitology. 2003;128(1):111-4.
223. Jacobs PP, Geysens S, Vervecken W, Contreras R, Callewaert N. Engineering complex-type N-glycosylation in Pichia pastoris using GlycoSwitch technology. Nature Protocols. 2009;4(1):58-70.
224. Meuris L, Santens F, Elson G, Festjens N, Boone M, Dos Santos A, Devos S, Rousseau F, Plets E, Houthuys E, Malinge P, Magistrelli G, Cons L, Chatel L, Devreese B, Callewaert N. GlycoDelete engineering of mammalian cells simplifies N-glycosylation of recombinant proteins. Nature Biotechnology. 2014;32(5):485-U210.
225. Roberts B, Antonopoulos A, Haslam SM, Dicker AJ, McNeilly TN, Johnston SL, Dell A, Knox DP, Britton C. Novel expression of Haemonchus contortus vaccine candidate aminopeptidase H11 using the free-living nematode Caenorhabditis elegans. Veterinary Research. 2013;44.
226. Chitnis CE, Mukherjee P, Mehta S, Yazdani SS, Dhawan S, Shakri AR, Bhardwaj R, Gupta PK, Hans D, Mazumdar S, Singh B, Kumar S, Pandey G, Parulekar V, Imbault N, Shivyogi P, Godbole G, Mohan K, Leroy O, Singh K, Chauhan VS. Phase I Clinical Trial of a Recombinant Blood Stage Vaccine Candidate for Plasmodium falciparum Malaria Based on MSP1 and EBA175. Plos One. 2015;10(9).
227. Xu J, Bai X, Wang L, Shi HN, van der Giessen JWB, Boireau P, Liu MY, Liu XL. Influence of adjuvant formulation on inducing immune response in mice immunized with a recombinant serpin from Trichinella spiralis. Parasite Immunology. 2017. 228. Hu JH, Chen ZH, Gu J, Wan MB, Shen Q, Kieny MP, He J, Li Z, Zhang Q, Reed ZH, Zhu Y, Li W, Cao Y, Qu L, Cao Z, Wang Q, Liu H, Pan X, Huang X, Zhang Dm Xue X, Pan W. Safety and Immunogenicity of a Malaria Vaccine, Plasmodium falciparum AMA-1/MSP-1 Chimeric Protein Formulated in Montanide ISA 720 in Healthy Adults. Plos One. 2008;3(4). 229. Murphy K. Janeway's Immunobiology. 8th ed. London: Garland Science; 2012.
230. Loukas A, Bethony JM, Williamson AL, Goud GN, Mendez S, Zhan B, Hawdon JM, Bottazzi ME, Brindley PJ Hotez PJ. Vaccination of dogs with a recombinant cysteine protease from the intestine of canine hookworms

198
diminishes the fecundity and growth of worms. Journal of Infectious Diseases. 2004;189(10):1952-61.
231. Vermeire JJ, Lantz LD, Caffrey CR. Cure of Hookworm Infection with a Cysteine Protease Inhibitor. Plos Neglected Tropical Diseases. 2012;6(7).
232. Singh PK, Kushwaha S, Rana AK, Misra-Bhattacharya S. Cofactor Independent Phosphoglycerate Mutase of Brugia malayi Induces a Mixed Th1/Th2 Type Immune Response and Inhibits Larval Development in the Host. Biomed Research International. 2014.
233. Singh PK, Kushwaha S, Mohd S, Pathak M, Misra-Bhattacharya S. In vitro gene silencing of independent phosphoglycerate mutase (iPGM) in the filarial parasite Brugia malayi. Infectious Diseases of Poverty. 2013;2.
234. Feng D, Zhao WL, Ye YY, Bai XC, Liu RQ, Chang LF, Zhou Q, Sui SF. Cellular Internalization of Exosomes Occurs Through Phagocytosis. Traffic. 2010;11(5):675-87.
235. Guy B. The perfect mix: recent progress in adjuvant research. Nature Reviews Microbiology. 2007;5(7):505-17.
236. Chadwick S, Kriegel C, Amiji M. Nanotechnology solutions for mucosal immunization. Advanced Drug Delivery Reviews. 2010;62(4-5):394-407.
237. Gregoriadis G. Liposomes as Immunoadjuvants and Vaccine Carriers: Antigen Entrapment. ImmunoMethods. 1994;4(3):210-6.
238. Fifis T, Gamvrellis A, Crimeen-Irwin B, Pietersz GA, Li J, Mottram PL, McKenzie IF, Plebanski M. Size-dependent immunogenicity: Therapeutic and protective properties of nano-vaccines against tumors. Journal of Immunology. 2004;173(5):3148-54.
239. Armstrong SD, Babayan SA, Lhermitte-Vallarino N, Gray N, Xia D, Martin C, Kumar S, Taylor DW, Blaxter ML, Wastling JM, Makepeace BL. Comparative Analysis of the Secretome from a Model Filarial Nematode (Litomosoides sigmodontis) Reveals Maximal Diversity in Gravid Female Parasites. Molecular & Cellular Proteomics. 2014;13(10):2527-44.
240. Vercauteren I, Geldhof P, Peelaers I, Claerebout E, Berx G, Vercruysse J. Identification of excretory-secretory products of larval and adult Ostertagia ostertagi by immunoscreening of cDNA libraries. Molecular and Biochemical Parasitology. 2003;126(2):201-8.
241. Rodriguez PBR, Cruz RR, Garcia DID, Gutierrez RH, Quintanilla REL, Sahagun DO, Gonzalez Castillo C, Gutierrez Ortega A, Herrera Rodriguez SE, Vallejo Cardona A, Martinez Velazquez M. Identification of immunogenic proteins from ovarian tissue and recognized in larval extracts of Rhipicephalus (Boophilus) microplus, through an immunoproteomic approach. Experimental Parasitology. 2016;170:227-35.

199
242. Taheri M, Nabian S, Ranjbar M, Fard RMN, Sadeghian AG, Sazmand A. Study of vitellogenin in Boophilus annulatus tick larvae and its immunological aspects. Tropical Biomedicine. 2014;31(3):398-405.
243. Bartley K, Wright HW, Huntley JF, Manson EDT, Inglis NF, McLean K, Nath M, Bartley Y, Nisbet AJ. Identification and evaluation of vaccine candidate antigens from the poultry red mite (Dermanyssus gallinae). International Journal for Parasitology. 2015;45(13):819-30.
244. Zhang TT, Cui XJ, Zhang JC, Wang H, Wu M, Zeng H, Cao Y, Liu J, Hu J. Screening and Identification of Antigenic Proteins from the Hard Tick Dermacentor silvarum (Acari: Ixodidae). Korean Journal of Parasitology. 2015;53(6):789-93.
245. Pearson MS, Bethony JM, Pickering DA, de Oliveira LM, Jariwala A, Santiago H, Miles AP, Zhan B, Jiang D, Ranjit N, Mulvenna J, Tribolet L, Plieskatt J, Smith T, Bottazzi ME, Jones K, Keegan B, Hotez PJ, Loukas A. An enzymatically inactivated hemoglobinase from Necator americanus induces neutralizing antibodies against multiple hookworm species and protects dogs against heterologous hookworm infection. Faseb Journal. 2009;23(9):3007-19.
246. Beauvillain C, Juste MO, Dion S, Pierre J, Dimier-Poisson I. Exosomes are an effective vaccine against congenital toxoplasmosis in mice. Vaccine. 2009;27(11):1750-7.
247. del Cacho E, Gallego M, Lee SH, Lillehoj HS, Quilez J, Lillehoj EP, Quilez J, Lillehoj EP, Sanchez-Acedo C. Induction of Protective Immunity against Eimeria tenella, Eimeria maxima, and Eimeria acervulina Infections Using Dendritic Cell-Derived Exosomes. Infection and Immunity. 2012;80(5):1909-16.
248. McSorley HJ, Hewitson JP, Maizels RM. Immunomodulation by helminth parasites: Defining mechanisms and mediators. International Journal for Parasitology. 2013;43(3-4):301-10.
249. Boulanger D, Reid GDF, Sturrock RF, Wolowczuk I, Balloul JM, Grezel D, Pierce RJ, Otieno MF, Guerret MF, Grimaud A, Butterworth AE, Capron A. Immunization of mice and baboons with the recombinant SM28GST affects both worm viability and fecundity after experimental infection with Schistosome mansoni. Parasite Immunology. 1991;13(5):473-90.
250. Boulanger D, Warter A, Trottein F, Mauny F, Bremond P, Audibert F, Couret D, Kadri S, Codin C, Sellin E, Pierce RJ, Lecocq JP, Sellin B, Capron A. Vaccination of patas monkeys experimentally with Schistosoma haematobium using a recombinant glutathione-S-transferase cloned from Schistosoma mansoni. Parasite Immunology. 1995;17(7):361-9.
251. Grzych JM, Grezel D, Xu CB, Neyrinck JL, Capron M, Ouma JH, Butterworth AE, Capron A. IgA antibodies to a protective antigen in human Schistosomiasis mansoni. Journal of Immunology. 1993;150(2):527-35.

200
252. Remoue F, Rogerie F, Gallissot MC, Guyatt HL, Neyrinck JL, Diakkhate MM, Niang M, Butterworth AE, Auriault C, Capron A, Riveau G. Sex-dependent neutralizing humoral response to Schistosoma mansoni 28GST antigen in infected human populations. Journal of Infectious Diseases. 2000;181(5):1855-9.
253. Habtezion A, Nguyen LP, Hadeiba H, Butcher EC. Leukocyte Trafficking to the Small Intestine and Colon. Gastroenterology. 2016;150(2):340-54.
254. Li XR, Aldayel AM, Cui ZR. Aluminum hydroxide nanoparticles show a stronger vaccine adjuvant activity than traditional aluminum hydroxide microparticles. Journal of Controlled Release. 2014;173:148-57.
255. Salim N, Knopp S, Lweno O, Abdul U, Mohamed A, Schindler T, Rothen J, Masimba J, Kwaba D, Mohammed AS, Althaus F, Abdulla S, Tanner M, Daubenberger C, Genton B. Distribution and Risk Factors for Plasmodium and Helminth Co-infections: A Cross-Sectional Survey among Children in Bagamoyo District, Coastal Region of Tanzania. Plos Neglected Tropical Diseases. 2015;9(4):3660-.
256. Goncalves RM, Lima NF, Ferreira MU. Parasite virulence, co-infections and cytokine balance in malaria. Pathogens and Global Health. 2014;108(4):173-8.
257. George PJ, Anuradha R, Kumar NP, Sridhar R, Banurekha VV, Nutman TB, Babu S. Helminth Infections Coincident with Active Pulmonary Tuberculosis Inhibit Mono- and Multifunctional CD4(+) and CD8(+) T Cell Responses in a Process Dependent on IL-10. Plos Pathogens. 2014;10(9).

201
Appendix 1
GTTGACATTGATTATTGACTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAAAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTCCGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTACGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACACCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAATAACCCCGCCCCGTTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCGTTTAGTGAACCGTCAGATCTCTAGAAGCTGGGTACCAGCTGCTAGCAAGCTTCTTCCTGCCGCCAGCCTGCCTGCCACTGAGGGTTCCCAGCACCATGAGGGCCTGGATCTTCTTTCTCCTTTGCCTGGCCGGGAGGGCTCTGGCAGCCCCGCTAGCGCATCACCATCACCATCACGATCTGTACGACGATGACGATAAGCTCGAGGCCGGCAAGGCCGGATCCAGACATGATAAGATACATTGATGAGTTTGGACAAACCACAACTAGAATGCAGTGAAAAAAATGCTTTATTTGTGAAATTTGTGATGCTATTGCTTTATTTGTAACCATTATAAGCTGCAATAAACAAGTTAACAACAACAATTGCATTCATTTTATGTTTCAGGTTCAGGGGGAGGTGTGGGAGGTTTTTTAAAGCAAGTAAAACCTCTACAAATGTGGTATGGCTGATTATGATCCGGCTGCCTCGCGCGTTTCGGTGATGACGGTGAAAACCTCTGACACATGCAGCTCCCGGAGACGGTCACAGCTTGTCTGTAAGCGGATGCCGGGAGCAGACAAGCCCGTCAGGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCGCAGCCATGAGGTCGACTCTAGAGGATCGATGCCCCGCCCCGGACGAACTAAACCTGACTACGACATCTCTGCCCCTTCTTCGCGGGGCAGTGCATGTAATCCCTTCAGTTGGTTGGTACAACTTGCCAACTGGGCCCTGTTCCACATGTGACACGGGGGGGGACCAAACACAAAGGGGTTCTCTGACTGTAGTTGACATCCTTATAAATGGATGTGCACATTTGCCAACACTGAGTGGCTTTCATCCTGGAGCAGACTTTGCAGTCTGTGGACTGCAACACAACATTGCCTTTATGTGTAACTCTTGGCTGAAGCTCTTACACCAATGCTGGGGGACATGTACCTCCCAGGGGCCCAGGAAGACTACGGGAGGCTACACCAACGTCAATCAGAGGGGCCTGTGTAGCTACCGATAAGCGGACCCTCAAGAGGGCATTAGCAATAGTGTTTATAAGGCCCCCTTGTTAACCCTAAACGGGTAGCATATGCTTCCCGGGTAGTAGTATATACTATCCAGACTAACCCTAATTCAATAGCATATGTTACCCAACGGGAAGCATATGCTATCGAATTAGGGTTAGTAAAAGGGTCCTAAGGAACAGCGATATCTCCCACCCCATGAGCTGTCACGGTTTTATTTACATGGGGTCAGGATTCCACGAGGGTAGTGAACCATTTTAGTCACAAGGGCAGTGGCTGAAGATCAAGGAGCGGGCAGTGAACTCTCCTGAATCTTCGCCTGCTTCTTCATTCTCCTTCGTTTAGCTAATAGAATAACTGCTGAGTTGTGAACAGTAAGGTGTATGTGAGGTGCTCGAAAACAAGGTTTCAGGTGACGCCCCCAGAATAAAATTTGGACGGGGGGTTCAGTGGTGGCATTGTGCTATGACACCAATATAACCCTCACAAACCCCTTGGGCAATAAATACTAGTGTAGGAATGAAACATTCTGAATATCTTTAACAATAGAAATCCATGGGGTGGGGACAAGCCGTAAAGACTGGATGTCCATCTCACACGAATTTATGGCTATGGGCAACACATAATCCTAGTGCAATATGATACTGGGGTTATTAAGATGTGTCCCAGGCAGGGACCAAGACAGGTGAACCATGTTGTTACACTCTATTTGTAACAAGGGGAAAGAGAGTGGACGCCGACAGCAGCGGACTCCACTGGTTGTCTCTAACACCCCCGAAAATTAAACGGGGCTCCACGCCAATGGGGCCCATAAACAAAGACAAGTGGCCACTCTTTTTTTTGAAATTGTGGAGTGGGGGCACGCGTCAGCCCCCACACGCCGCCCTGCGGTTTTGGACTGTAAAATAAGGGTGTAATAACTTGGCTGATTGTAACCCCGCTAACCACTGCGGTCAAACCACTTGCCCACAAAACCACTAATGGCACCCCGGGGAATACCTGCATAAGTAGGTGGGCGGGCCAAGATAGGGGCGCGATTGCTGCGATCTGGAGGACAAATTACACACACTTGCGCCTGAGCGCCAAGCACAGGGTTGTTGGTCCTCATATTCACGAGGTCGCTGAGAGCACGGTGGGCTAATGTTGCCATGGGTAGCATATACTACCCAAATATCTGGATAGCATATGCTATCCTAATCTATATCTGGGTAGCATAGGCTATCCTAATCTATATCTGGGTAGCATATGCTATCCTAATCTATATCTGGGTAGTATATGCTATCCTAATTTATATCTGGGTAGCATAGGCTATCCTAATCTATATCTGGGTAGCATATGCTATCCTAATCTATATCTGGGTAGTATATGCTATCCTAATCTGTATCCGGGTAGCATATGCTATCCTAATAGAGATTAGGGTAGTATATGCTATCCTAATTTATATCTGGGTAGCATATACTACCCAAATATCTGGATAGCATATGCTATCCTAATCTATATCTGGGTAGCATATGCTATCCTAATCTATATCTGGGTAGCATAGGCTATCCTAATCTATATCTGGGTAGCATATGCTATCCTAATCTATATCTGGGTAGTATATGCTATCCTAATTTATATCTGGGTAGCATAGGCTATCCTAATCTATATCTGGGTAGCATATGCTATCCTAATCTATATCTGGGTAGTATATGCTATCCTAATCTGTATCCGGGTAGCATATGCTATCCTCATGCATATACAGTCAGCATATGATACCCAGTAGTAGAGTGGGAGTGCTATCCTTTGCATATGCCGCCACCTCCCAAGGGGGCGTGAATTTTCGCTGCTTGTCCTTTTCCTGCTGGTTGCTCCCATTCTTAGGTGAATTTAAGGAGGCCAGGCTAAAGCCGTCGCATGTCTGATTGCTCACCAGGTAAATGTCGCTAATGTTTTCCAACGCGAGAAGGTGTTGAGCGCGGAGCTGAGTGACGTGACAACATGGGTATGCCCAATTGCCCCATGTTGGGAGGACGAAAATGGTGACAAGACAGATGGCCAGAAATACACCAACAGCACGCATGATGTCTACTGGGGATTTATTCTTTAGTGCGGGGGAATACACGGCTTTT

202
AATACGATTGAGGGCGTCTCCTAACAAGTTACATCACTCCTGCCCTTCCTCACCCTCATCTCCATCACCTCCTTCATCTCCGTCATCTCCGTCATCACCCTCCGCGGCAGCCCCTTCCACCATAGGTGGAAACCAGGGAGGCAAATCTACTCCATCGTCAAAGCTGCACACAGTCACCCTGATATTGCAGGTAGGAGCGGGCTTTGTCATAACAAGGTCCTTAATCGCATCCTTCAAAACCTCAGCAAATATATGAGTTTGTAAAAAGACCATGAAATAACAGACAATGGACTCCCTTAGCGGGCCAGGTTGTGGGCCGGGTCCAGGGGCCATTCCAAAGGGGAGACGACTCAATGGTGTAAGACGACATTGTGGAATAGCAAGGGCAGTTCCTCGCCTTAGGTTGTAAAGGGAGGTCTTACTACCTCCATATACGAACACACCGGCGACCCAAGTTCCTTCGTCGGTAGTCCTTTCTACGTGACTCCTAGCCAGGAGAGCTCTTAAACCTTCTGCAATGTTCTCAAATTTCGGGTTGGAACCTCCTTGACCACGATGCTTTCCAAACCACCCTCCTTTTTTGCGCCTGCCTCCATCACCCTGACCCCGGGGTCCAGTGCTTGGGCCTTCTCCTGGGTCATCTGCGGGGCCCTGCTCTATCGCTCCCGGGGGCACGTCAGGCTCACCATCTGGGCCACCTTCTTGGTGGTATTCAAAATAATCGGCTTCCCCTACAGGGTGGAAAAATGGCCTTCTACCTGGAGGGGGCCTGCGCGGTGGAGACCCGGATGATGATGACTGACTACTGGGACTCCTGGGCCTCTTTTCTCCACGTCCACGACCTCTCCCCCTGGCTCTTTCACGACTTCCCCCCCTGGCTCTTTCACGTCCTCTACCCCGGCGGCCTCCACTACCTCCTCGACCCCGGCCTCCACTACCTCCTCGACCCCGGCCTCCACTGCCTCCTCGACCCCGGCCTCCACCTCCTGCTCCTGCCCCTCCTGCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTGCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTCCTGCTCCTGCCCCTCCTGCTCCTGCCCCTCCCGCTCCTGCTCCTGCTCCTGTTCCACCGTGGGTCCCTTTGCAGCCAATGCAACTTGGACGTTTTTGGGGTCTCCGGACACCATCTCTATGTCTTGGCCCTGATCCTGAGCCGCCCGGGGCTCCTGGTCTTCCGCCTCCTCGTCCTCGTCCTCTTCCCCGTCCTCGTCCATGGTTATCACCCCCTCTTCTTTGAGGTCCACTGCCGCCGGAGCCTTCTGGTCCAGATGTGTCTCCCTTCTCTCCTAGGCCATTTCCAGGTCCTGTACCTGGCCCCTCGTCAGACATGATTCACACTAAAAGAGATCAATAGACATCTTTATTAGACGACGCTCAGTGAATACAGGGAGTGCAGACTCCTGCCCCCTCCAACAGCCCCCCCACCCTCATCCCCTTCATGGTCGCTGTCAGACAGATCCAGGTCTGAAAATTCCCCATCCTCCGAACCATCCTCGTCCTCATCACCAATTACTCGCAGCCCGGAAAACTCCCGCTGAACATCCTCAAGATTTGCGTCCTGAGCCTCAAGCCAGGCCTCAAATTCCTCGTCCCCCTTTTTGCTGGACGGTAGGGATGGGGATTCTCGGGACCCCTCCTCTTCCTCTTCAAGGTCACCAGACAGAGATGCTACTGGGGCAACGGAAGAAAAGCTGGGTGCGGCCTGTGAGGATCAGCTTATCGATGATAAGCTGTCAAACATGAGAATTCTTGAAGACGAAAGGGCCTCGTGATACGCCTATTTTTATAGGTTAATGTCATGATAATAATGGTTTCTTAGACGTCAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTCCGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAAACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAACTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATGATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTGTTGACGCCGGGCAAGAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTCACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACCATGAGTGATAACACTGCGGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTAACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAGCTGAATGAAGCCATACCAAACGACGAGCGTGACACCACGATGCCTGCAGCAATGGCAACAACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATAGACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGCTGGTTTATTGCTGATAAATCTGGAGCCGGTGAGCGTGGGTCTCGCGGTATCATTGCAGCACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGGAGTCAGGCAACTATGGATGAACGAAATAGACAGATCGCTGAGATAGGTGCCTCACTGATTAAGCATTGGTAACTGTCAGACCAAGTTTACTCATATATACTTTAGATTGATTTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTCCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCC

203
AGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGCCTTTTGCTGGCCTTGAAGCTGTCCCTGATGGTCGTCATCTACCTGCCTGGACAGCATGGCCTGCAACGCGGGCATCCCGATGCCGCCGGAAGCGAGAAGAATCATAATGGGGAAGGCCATCCAGCCTCGCGTCGCGAACGCCAGCAAGACGTAGCCCAGCGCGTCGGCCCCGAGATGCGCCGCGTGCGGCTGCTGGAGATGGCGGACGCGATGGATATGTTCTGCCAAGGGTTGGTTTGCGCATTCACAGTTCTCCGCAAGAATTGATTGGCTCCAATTCTTGGAGTGGTGAATCCGTTAGCGAGGTGCCGCCCTGCTTCATCCCCGTGGCCCGTTGCTCGCGTTTGCTGGCGGTGTCCCCGGAAGAAATATATTTGCATGTCTTTAGTTCTATGATGACACAAACCCCGCCCAGCGTCTTGTCATTGGCGAATTCGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCGAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCCGGGGGGCAATGAGATATGAAAAAGCCTGAACTCACCGCGACGTCTGTCGAGAAGTTTCTGATCGAAAAGTTCGACAGCGTCTCCGACCTGATGCAGCTCTCGGAGGGCGAAGAATCTCGTGCTTTCAGCTTCGATGTAGGAGGGCGTGGATATGTCCTGCGGGTAAATAGCTGCGCCGATGGTTTCTACAAAGATCGTTATGTTTATCGGCACTTTGCATCGGCCGCGCTCCCGATTCCGGAAGTGCTTGACATTGGGGAATTCAGCGAGAGCCTGACCTATTGCATCTCCCGCCGTGCACAGGGTGTCACGTTGCAAGACCTGCCTGAAACCGAACTGCCCGCTGTTCTGCAGCCGGTCGCGGAGGCCATGGATGCGATCGCTGCGGCCGATCTTAGCCAGACGAGCGGGTTCGGCCCATTCGGACCGCAAGGAATCGGTCAATACACTACATGGCGTGATTTCATATGCGCGATTGCTGATCCCCATGTGTATCACTGGCAAACTGTGATGGACGACACCGTCAGTGCGTCCGTCGCGCAGGCTCTCGATGAGCTGATGCTTTGGGCCGAGGACTGCCCCGAAGTCCGGCACCTCGTGCACGCGGATTTCGGCTCCAACAATGTCCTGACGGACAATGGCCGCATAACAGCGGTCATTGACTGGAGCGAGGCGATGTTCGGGGATTCCCAATACGAGGTCGCCAACATCTTCTTCTGGAGGCCGTGGTTGGCTTGTATGGAGCAGCAGACGCGCTACTTCGAGCGGAGGCATCCGGAGCTTGCAGGATCGCCGCGGCTCCGGGCGTATATGCTCCGCATTGGTCTTGACCAACTCTATCAGAGCTTGGTTGACGGCAATTTCGATGATGCAGCTTGGGCGCAGGGTCGATGCGACGCAATCGTCCGATCCGGAGCCGGGACTGTCGGGCGTACACAAATCGCCCGCAGAAGCGCGGCCGTCTGGACCGATGGCTGTGTAGAAGTACTCGCCGATAGTGGAAACCGACGCCCCAGCACTCGTCCGGATCGGGAGATGGGGGAGGCTAACTGAAACACGGAAGGAGACAATACCGGAAGGAACCCGCGCTATGACGGCAATAAAAAGACAGAATAAAACGCACGGGTGTTGGGTCGTTTGTTCATAAACGCGGGGTTCGGTCCCAGGGCTGGCACTCTGTCGATACCCCACCGAGACCCCATTGGGGCCAATACGCCCGCGTTTCTTCCTTTTCCCCACCCCACCCCCCAAGTTCGGGTGAAGGCCCAGGGCTCGCAGCCAACGTCGGGGCGGCAGGCCCTGCCATAGCCACTGGCCCCGTGGGTTAGGGACGGGGTCCCCCATGGGGAATGGTTTATGGTTCGTGGGGGTTATTATTTTGGGCGTTGCGTGGGGTCAGGTCCACGACTGGACTGAGCAGACAGACCCATGGTTTTTGGATGGCCTGGGCATGGACCGCATGTACTGGCGCGACACGAACACCGGGCGTCTGTGGCTGCCAAACACCCCCGACCCCCAAAAACCACCGCGCGGATTTCTGGCGTGCCAAGCTAGTCGACCAATTCTCATGTTTGACAGCTTATCATCGCAGATCCGGGCAACGTTGTTGCCATTGCTGCAGGCGCAGAACTGGTAGGTATGGAAGATCTATACATTGAATCAATATTGGCAATTAGCCATATTAGTCATTGGTTATATAGCATAAATCAATATTGGCTATTGGCCATTGCATACGTTGTATCTATATCATAATATGTACATTTATATTGGCTCATGTCCAATATGACCGCCAT
Figure A1.1. Sequence for pCep-His vector used for expression of recombinant T. muris proteins.

204
The following Tables can be found on the CD provided with this thesis.
Table A1.1. List of proteins identified within adult T. muris ES. List was compiled by collating the mass spectrometry data for fractions 16 to 32. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. Frac = fraction.
Table A1.2. List of proteins identified within pool 1. List was compiled by collating the mass spectrometry data for fractions 16 to 18. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. Frac = fraction.
Table A1.3. List of proteins identified within pool 2. List was compiled by collating the mass spectrometry data for fractions 20 to 22. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. Frac = fraction. Table A1.4. List of proteins identified within pool 3. List was compiled by collating the mass spectrometry data for fractions 24 to 27. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. Frac = fraction. Table A1.5. List of proteins identified within pool 4. List was compiled by collating the mass spectrometry data for fractions 29 to 32. The total spectral count is displayed for each protein in each fraction (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. Frac = fraction.
Table A1.6. List of L2 larval ES protein. List was compiled based on proteins identified in unfractionated L2 larval ES. The total spectral count is displayed for each protein in each sample (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. Frac = fraction.

205
Appendix 2
Accession number Protein No. unique peptides
TMUE_s0016004100 Serpin 20
Table A2.1. Mass spectrometry analysis of recombinant Serpin. Once the recombinant Serpin protein had been expressed and purified, its identity was confirmed by tandem mass spectrometry of tryptic peptides.
No. unique peptides
Accession number Protein 45 kDa band
30 kDa band
25 kDa band
TMUE_s0428000200 Lactoylglutathione lyase 21 18 20
TMUE_s0029005700 Actin 2 3 3
Table A2.2. Mass spectrometry analysis of recombinant Lactoglutathione lyase. Once the recombinant Lactoglutathione lyase protein had been expressed and purified, its identity was confirmed by tandem mass spectrometry of tryptic peptides from gel digests. The proteins identified in the 45, 30 and 25 kDa bands are shown.
Accession number Protein No. unique peptides
TMUE_s0009001300 Translationally controlled tumour protein 13
Table A2.3. Mass spectrometry analysis of recombinant Translationally controlled tumour protein. Once the recombinant Translationally controlled tumour protein had been expressed and purified, its identity was confirmed by tandem mass spectrometry of tryptic peptides.
No. unique peptides
Accession number Protein 30 kDa band
20 kDa band
TMUE_s0071003300 TPD52 domain containing protein 12 12
Table A2.4. Mass spectrometry analysis of recombinant TPD52 domain containing protein. Once the recombinant TPD52 domain containing protein had been expressed and purified, its identity was confirmed by tandem mass spectrometry of tryptic peptides from gel digests. The proteins identified in the 30 and 20 kDa bands are shown.

206
Accession number Protein No. unique peptides
TMUE_s0016007700 Hypothetical protein 5
Table A2.5. Mass spectrometry analysis of recombinant Hypothetical protein. Once the recombinant Hypothetical protein had been expressed and purified, its identity was confirmed by tandem mass spectrometry of tryptic peptides.
Comparison to T. trichiura homologue
Accession number Protein Query
coverage (%) Identity
(%)
TMUE_s0016004100 Serpin protein 98 44
TMUE_s0009001300 Translationally controlled tumour protein 95 91
TMUE_s0066001200 Ion trans 2 and Pfam-B 17708-domain containing protein
35 90
TMUE_s0016007600 Hypothetical protein 98 40
TMUE_s0072003400 Independent phosphoglycerate mutase 93 88
TMUE_s0071003300 TPD52 domain containing protein 84 89
TMUE_s0132001200 Gut specific cysteine proteinase 85 71
TMUE_s0428000200 Lactoylglutathione lyase 97 86
TMUE_s0003017400 Rab GDP dissociation inhibitor alpha 99 92
TMUE_s0013010700 NADH dependent fumarate reductase 85 79
Table A2.6. Comparison of protein sequences for T. muris immunogenic candidates with T. trichiura homologues. A BLAST search was performed whereby the protein sequence for each of the immunogenic candidates was searched against all organisms. The top T. trichiura protein hit was identified and the query coverage (%) and identity (%) was recorded.

207
Figure A2.1. BLAST search results for Ion trans 2 and Pfam-B 17708-domain containing protein (TMUE_s0066001200). The protein sequence for Ion trans 2 and Pfam-B 17708-domain containing protein (TMUE_s0066001200) was searched against all organisms using the BLAST protein search tool. Protein sequences (from other organisms) with shared homology are aligned against the protein sequence for TMUE_s0066001200. The results suggest that the predicted protein sequence may have arisen by incorrectly pairing together two proteins (protein 1: approximately amino acids 1-620, protein 2: approximately amino acids 780-1200).
Protein Organism Query
coverage (%) Identity
(%)
Hypothetical protein T. suis 35 90
Ion trans 2 domain containing protein T. trichiura 35 90
Potassium channel subfamily K member 18 T. zimbabwensis 35 71
Potassium channel subfamily K member 18 T. pseudospiralis 35 71
Potassium channel subfamily K member 18 T. papuae 35 71
Potassium channel subfamily K member 18 T. murelli 35 71
Potassium channel subfamily K member 18 T. spiralis 35 63
Table A2.7. BLAST search results for T. muris Ion trans 2 and Pfam-B 17708-domain containing protein (TMUE_s0066001200). The protein sequence of T. muris Ion trans 2 and Pfam-B 17708-domain containing protein (TMUE_s0066001200) was searched against all organisms using the BLAST protein search tool. The results revealed that there are homologues within other tricephalid parasites, including Trichuris and Trichinella species.
Amino acid position
Alig
ned
se
qu
en
ce
s

208
Figure A2.2. Anti-parasite IgG1 serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Montanide ISA 720. Mice, n=5 per group, were subcutaneously vaccinated with recombinant T. muris proteins administered together as a single vaccination (30 μg of each protein, 150 μg protein in total). Mice were vaccinated a second and third time with 15 μg of each protein (75 μg protein in total). All vaccinations were formulated with Montanide ISA 720, and 30 days after the third vaccination, mice were infected with 25 T. muris eggs. Mice were sacrificed at day 32 p.i. Anti-parasite IgG1 serum antibody levels were measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccine group (A-C). The symbols on each graph represent individual mice within the same vaccination group. (D) Shows mean anti-parasite IgG1 serum antibody levels for each vaccine group at 1:320 serum dilution. Sham= mice vaccinated with aluminum hydroxide only, ES vaccinated mice were used as positive control. Error bars show SEM, **** P <0.0001.
A) Serum dose response curve
for sham vaccination group
B) Serum dose response curve for
recombinants vaccination group
C) Serum dose response curve
for ES vaccination group
D) Mean O.D. (1:320 serum dilution)
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
1.0
1.5
2.0
2.5
Dilution
O.D
.
Sham
Rec
ombin
ants
ES +
Monta
nide
ES +
alu
m
0.0
0.5
1.0
1.5
2.0
2.5
Vaccination group
O.D
.
****
****

209
Figure A2.3. Anti-parasite IgG2a serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Montanide ISA 720. Mice, n=5 per group, were subcutaneously vaccinated with recombinant T. muris proteins administered together as a single vaccination (30 μg of each protein, 150 μg protein in total). Mice were vaccinated a second and third time with 15 μg of each protein (75 μg protein in total). All vaccinations were formulated with Montanide ISA 720, and 30 days after the third vaccination, mice were infected with 25 T. muris eggs. Mice were sacrificed at day 32 p.i. Anti-parasite IgG2a serum antibody levels were measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccine group (A-C). The symbols on each graph represent individual mice within the same vaccination group. (D) Shows mean anti-parasite IgG2a serum antibody levels for each vaccine group at 1:40 serum dilution. Sham= mice vaccinated with aluminum hydroxide only, ES vaccinated mice were used as positive control. Error bars show SEM, **** P <0.0001, NS= non-significant.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
recombinants vaccination group
C) Serum dose response curve for
ES vaccination group
D) Mean O.D. (1:40 serum dilution)
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
Sham
Rec
ombin
ants
ES +
Monta
nide
ES +
alu
m
0.0
0.5
1.0
1.5
2.0
Vaccination group
O.D
.
NS
********
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.

210
Figure A2.4. Anti-parasite IgG1 serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with aluminum hydroxide. Mice, n=5 per group, were subcutaneously vaccinated with recombinant T. muris proteins administered together as a single vaccination (30 μg of each protein, 150 μg of protein in total) formulated with aluminum hydroxide. Two weeks following the second vaccination, mice were infected with 25 T. muris eggs and were sacrificed at day 32 p.i. The anti-parasite IgG1 serum antibody response was measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccination group (A-C). The symbols on each graph represent individual mice within the same vaccination group. (D) Shows mean anti-parasite IgG1 serum antibody levels for each vaccine group at 1:320 serum dilution. Sham vaccination group= mice vaccinated with aluminum hydroxide only. ES vaccination group= mice vaccinated with ES formulated with aluminum hydroxide. Error bars show SEM **** P < 0.0001, NS= non-significant.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
recombinants vaccination group
C) Serum dose response curve for
ES vaccination group
D) Mean O.D. (1:320 serum dilution)
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Dilution
O.D
.
Sham
Rec
ombin
ants E
S
0.0
0.5
1.0
1.5
2.0
2.5
Vaccination group
O.D
.
****
NS

211
Figure A2.5. Anti-parasite IgG2a serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with aluminum hydroxide. Mice, n=5 per group, were subcutaneously vaccinated with recombinant T. muris proteins administered together as a single vaccination (30 μg of each protein, 150 μg of protein in total) formulated with aluminum hydroxide. Two weeks following the second vaccination, mice were infected with 25 T. muris eggs and were sacrificed at day 32 p.i. The anti-parasite IgG2a serum antibody response was measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccination group (A-C). The symbols on each graph represent individual mice within the same vaccination group. (D) Shows mean anti-parasite IgG2a serum antibody levels for each vaccine group at 1:40 serum dilution. Sham vaccination group= mice vaccinated with aluminum hydroxide only. ES vaccination group= mice vaccinated with ES formulated with aluminum hydroxide. Error bars show SEM, NS= non-significant.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
recombinants vaccination group
C) Serum dose response curve for
ES vaccination group
D) Mean O.D. (1:40 serum dilution)
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
0
0.0
0.5
1.0
1.5
2.0
2.5
Dilution
O.D
.
Sham
Rec
ombin
ants E
S
0.0
0.5
1.0
1.5
Vaccination group
O.D
.
NS
NS

212
Figure A2.6. Anti-parasite IgG1 serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Freund’s adjuvants. Mice, n=5 per group, were subcutaneously vaccinated with recombinant T. muris proteins administered together as a single vaccination (30 μg of each protein, 150 μg of protein in total, formulated in CFA). Mice were vaccinated a second and third time with 15 μg of each protein (75 μg protein in total, formulated in IFA). 30 days after the third vaccination, mice were infected with 25 T. muris eggs. Mice were sacrificed at day 32 p.i. Anti-parasite IgG1 serum antibody levels were measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccine group (A-C). The symbols on each graph represent individual mice within the same vaccination group. (D) Shows mean anti-parasite IgG1 serum antibody levels for each vaccine group at 1:320 serum dilution. Sham= mice vaccinated with Freund’s adjuvants only (CFA for the first vaccination, IFA for the second and third). ES= mice vaccinated with ES formulated with Freund’s adjuvants. Error bars show SEM, * P <0.05, **** P <0.0001.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
recombinants vaccination group
C) Serum dose response curve for
ES vaccination group
D) Mean O.D. (1:320 serum dilution)
1 in 2
0
1 in 4
0
1 in 8
0
1 in 1
60
1 in 3
20
1 in 6
40
1 in 1
280
1 in 2
560
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Freund's negative control IgG1
Dilution
O.D
.
A
B
C
D
E
1 in 2
0
1 in 4
0
1 in 8
0
1 in 1
60
1 in 3
20
1 in 6
40
1 in 1
280
1 in 2
560
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Freund's recombinants IgG1
Dilution
O.D
.
A
B
C
D
E
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Dilution
O.D
.
Sham
Rec
ombin
ants E
S
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Vaccine group
O.D
.
*
****

213
Figure A2.7. Anti-parasite IgG2a serum antibody levels for mice vaccinated with recombinant T. muris proteins formulated with Freund’s adjuvants. Mice, n=5 per group, were subcutaneously vaccinated with recombinant T. muris proteins administered together as a single vaccination (30 μg of each protein, 150 μg of protein in total, formulated in CFA). Mice were vaccinated a second and third time with 15 μg of each protein (75 μg protein in total, formulated in IFA). 30 days after the third vaccination, mice were infected with 25 T. muris eggs. Mice were sacrificed at day 32 p.i. Anti-parasite IgG2a serum antibody levels were measured for each individual by ELISA (reading at 405 nm) and these are displayed for each vaccine group (A-C). The symbols on each graph represent individual mice within the same vaccination group. (D) Shows mean anti-parasite IgG2a serum antibody levels for each vaccine group at 1:40 serum dilution. Sham= mice vaccinated with Freund’s adjuvants only (CFA for the first vaccination, IFA for the second and third). ES= mice vaccinated with ES formulated with Freund’s adjuvants. Error bars show SEM, * P <0.05, NS= non-significant.
A) Serum dose response curve for
sham vaccination group
B) Serum dose response curve for
recombinants vaccination group
C) Serum dose response curve for
ES vaccination group
D) Mean O.D. (1:40 serum dilution)
Sham
Rec
ombin
ants E
S
0.0
0.5
1.0
1.5
2.0
2.5
Vaccine group
O.D
.
*
NS
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Dilution
O.D
.
1 in
20
1 in
40
1 in
80
1 in
160
1 in
320
1 in
640
1 in
128
0
1 in
256
00.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Dilution
O.D
.

214
Figure A2.8. Vaccination with T. muris recombinant proteins induced production of IgG2a antibodies specific for these proteins. IgG2a antibodies specific for Translationally controlled tumour protein (A), TPD52 domain containing protein (B), Lactoylglutathione lyase (C) and Serpin (D) were measured in sera from infected (high dose) mice, pool 3 vaccinated mice, or mice vaccinated with each of the recombinant proteins (single protein vaccinations formulated with aluminum hydroxide). IgG2a serum antibody levels were measured for each individual by ELISA (reading at 405 nm) and graphs show mean antibody titres at 1:40 serum dilution. Error bars show SEM, * P < 0.05, ** P < 0.01, **** P < 0.0001. Trans= Translationally controlled tumour protein, TDP= TPD52 domain containing protein, Lacto= Lactyoglutathione lyase.
A) IgG2a antibody response to
Translationally controlled tumour protein
B) IgG2a antibody response to TPD52
domain containing protein
C) IgG2a antibody response to
Lactoylglutathione lyase
D) IgG2a antibody response to Serpin
Hig
h d
ose in
fect
io
n
Pool 3
v
ac
cinat
ed
Lac
to
v
ac
cinat
ed
0.0
0.5
1.0
1.5
Serum response against Lacto
Source of serum
O.D
.
****
Hig
h dose
infe
ctio
n
Pool 3
vac
cinat
ed
Lacto
vac
cinat
ed
0.0
0.1
0.2
0.3
Source of serum
O.D
.
*
Hig
h d
ose in
fe
ctio
nPo
ol 3 v
ac
ci
nate
d
Serp
in v
ac
ci
nate
d
0.0
0.5
1.0
1.5
Summary 1:40 IgG2a
Vaccination group
O.D
.
**
Hig
h dose
infe
ctio
n
Pool 3
vac
cinat
edTr
ans
vacc
in
ated
0.00
0.02
0.04
0.06
0.08
0.10
1:40 dilution
Source of serum
O.D
.
*
Hig
h d
ose in
fe
ctio
nPo
ol 3
vac
cinat
ed
TPD
52
v
ac
cinat
ed
0.00
0.01
0.02
0.03
0.04
0.05
1:40 dilution
Source of serum
O.D
.
NS

215
Appendix 3 The following table can be found on the CD provided. Table A3.1. List of T. muris ELV proteins. Mass spectrometry analysis was carried out on three biological replicates (samples 1 to 3). The total spectral count is displayed for each protein in replicate (criteria set to 95% protein threshold, 50% peptide threshold, minimum 2 peptides identified). Mw= molecular weight in kDa. The table indicates whether each protein is identified in T. muris ES and whether there is a signal peptide present (Y/N= yes and no respectively).
Figure A3.1. BLAST search results for T. muris TSP-1 domain containing protein (TMUE_s0070003500). The protein sequence of T. muris TSP-1 domain containing protein (TMUE_s0070003500) was searched against all organisms using the BLAST protein search tool. The results revealed that this protein contains severaltetraspanin (TSP) domains.
Species Genomic Location Overlapping Gene(s)
E-val Identity (%)
S. mansoni Smp.Chr_1:27143252-27143419 Smp_060030 0.0034 28.6
S. mansoni Smp.Chr_1:53413414-53413488 Smp_158550 0.045 56
S. mansoni Smp.SC_0142:339943-340005 Smp_158620 1.30E-06 52.4
S. mansoni Smp.SC_0142:339829-339930 Smp_158620 1.30E-06 47.1
Table A3.2. BLAST search results show that there is significant homology between the T. muris TSP-1 domain containing protein (TMUE_s0070003500) and S. mansoni proteins. The WormBase ParaSite BLAST tool was used to compare the protein sequence of TMUE_s0070003500 to all Schistosoma sequences. The low E values (<0.05) suggest that there is significant homology between the sequences.

216
Figure A3.2. Worm burden and anti-parasite IgG1/IgG2a serum antibody response for ES titration experiment. (A) Worm burdens for mice vaccinated with 3, 10 or 30 μg ES, followed by a second vaccination with half the amount of material 2 weeks later. Mice were infected with 25 T. muris eggs and sacrificed at day 32 p.i.. (B) Shows mean anti-parasite IgG1 serum antibody levels for each vaccine group at 1:320 serum dilution. (C) Shows mean anti-parasite IgG2a serum antibody levels for each vaccine group at 1:40 serum dilution. Sham= mice vaccinated with aluminum hydroxide only. Error bars show SEM, **** P < 0.0001.
A) Worm burden
B) Anti-parasite IgG1 antibody
response for ES titration groups
(1:320 serum dilution)
C) Anti-parasite IgG2a antibody
response for ES titration groups
(1:40 serum dilution)
Sham 3
ug
10 u
g
30 u
g0.0
0.2
0.4
0.6
Vaccination group
O.D
.
********
****
Sham 3
ug
10 u
g
30 u
g
0.0
0.5
1.0
1.5
2.0
Vaccination group
O.D
.
****
****
****
Sham 3
ug
10 u
g
30 u
g
0
5
10
15
20
Vaccination group
Wo
rm b
urd
en
****
********

217
Table A3.3. BLAST search results for T. muris Vacuolar protein sorting associated protein (TMUE_s0093001800). The protein sequence of T. muris Vacuolar protein sorting associated protein (TMUE_s0093001800) was searched against all organisms using the BLAST protein search tool. The results revealed that there are homologues within other tricephalid parasites, including Trichuris and Trichinella species.
Protein Organism Query
coverage (%)
Identity (%)
Vacuolar sorting associated protein 52 T. trichiura 26 80
Vacuolar sorting associated protein 52-like protein T. pseudospiralis
26 58
Vacuolar sorting associated protein 52-like protein T. patagoniensis
26 59
Vacuolar sorting associated protein 52-like protein T. papuae
26 59
Vacuolar sorting associated protein 52-like protein T. zimbabwensis
26 59
Vacuolar sorting associated protein 52-like protein T. nelsoni
26 58