decreased sodium influx and abnormal red cell membrane lipids in a patient with familial plasma...
TRANSCRIPT

American Journal of Hematology 16:129-137 (1984)
Decreased Sodium Influx and Abnormal Red Cell Membrane Lipids in a Patient With Familial Plasma Lecithin:Cholesterol Acyltransferase Deficiency Naoki Murayama, Yasushi Asano, Saichi Hosoda, Masaji Maesawa, Masaki Saito, Fumimaro Takaku, Takashi Sugihara, Kosuke Miyashima, and Yoshihito Yawata
Department of Internal Medicine, Jichi Medical School, Kawachimachi, Third Department of Internal Medicine, University of Tokyo, Tokyo, and Division of Hematology, Department of Medicine, Kawasaki Medical School, Kurashiki, Japan
Red cell membrane metaholism in familial 1ecithin:cholesterol acy Itransferase t LCAT) deficiency uas incestigated. The family presented here is the third case discobered in Japan. An increase of free cholesterol was observed in the red cell membranes, concomitant with increased phosphatidyl choline. Osmotic fragility of the patient's red cells was diminished rather than increased. Red cell survical (5'Cr T1 2 ) wds shortened (15 days). Sodium influx was mdrkedly decreased, although sodium efflux, both ouabain-sensitive and ouahain-insensitive, was normal. The actibity of acety I- cholinesterase a5 a marker of the outer Leaflet of the red cell membrane\ uas decreased, while the dctivitq af glqcerdldehyde-3-pho~phate dehqdrogendse as a marker of the inner leaflet was normal. N o abnormalities of adenosine triphosphatases in red cell membranes were ohserced. These results suggest that the alteration of cholesterol metabolism in the plasma of LCAT deficiency incream the red cell membrane cholesterol and affects the functions of the red cell membranes, especially of the outer leaflet, which may result in decreased sodium influx.
Key word5: I f i T deficiencq, decreased %a influx, abnormal red cell membrane
INTRODUCTION
Since familial 1ecithin:cholesterol acyltransferase (LCAT) deficiency was first described in three Norwegian sisters by Norum and Gjone in 1967 [ 11, more than 30 patients with this disease have been reported. We have previously reported the siblings of the present case, which represents the third case in Japan [2].
Received for publication Februarq 24, 1083; accepted September 15, 1983.
Addre55 reprint requests to Naoki Murayama, MD, Division of Xephrology , Department of Vardiologq, Jichi Medical School, 3311-1 Minamikavcachi-Machi, Kawachi-Gun, Tochigi-Ken, 329-04, Japan.
1984 Alan R. Lir5, Inc.

130 Murayama et a1
This disease is characterized clinically by corneal infiltration and proteinuria, and biochemically by several plasma lipoprotein abnormalities with the absence or the near-absence of plasma LCAT activity [l]. Mild hemolytic anemia with altered lipid composition of the red cells is also one of the features of this disease [3]. It has been reported that the content of free cholesterol is markedly increased in the red cell membranes. Although the content of total membrane phospholipids is normal, phos- phatidyl choline (lecithin) is markedly increased concomitantly with a decrease in phosphatidyl ethanolamine and sphingomyelin [4,5].
The abnormalities in the composition of red cell membrane lipids with familial LCAT deficiency are accompanied by an alteration of red cell morphology. Thus, the metabolic relationship between the membrane lipid alteration and cellular functions was studied in these red cells.
This communication describes the altered red cell membrane functions in LCAT deficiency, and also reports the effects of fresh-frozen plasma transfusions on red cell membrane lipids and morphology.
MATERIALS AND METHODS
Red cell membrane ghosts were prepared by the method of Dodge et a1 [6 ] . Red cells were washed three times with physiological saline, and 1 vol of packed cells was lysed by adding 20 vol of 30 mOsm Veronal buffer (pH 7.4), and centrifuged at 15,000 rpm for 30 min. Sedimented ghosts were washed four times with the same Veronal buffer. All these procedures were performed at 4°C.
Membrane Lipids Lipids were extracted from the red cell ghosts into chloroform/methanol (2: 1
v/v) by the method of Folch et a1 [7]. Cholesterol was measured by the method of Zak [8], and the lipid phosphorus was determined as described by Bartlett [9]. Individual phospholipids were fractionated and measured by thin-layer chromatogra- phy on a basic Silica Gel G plate using chloroform/methanol/acetic acidlwater (25:15:4:2 v/v) as the developing solvent [lo]. Each spot of phospholipids was identified by spraying Dittmer’s regent, and taken into chloroform/methanol/acetic acid/water (25: 10:3:2 v/v) eluent. The content of lipids phosphorus in the eluents was determined by the method of Bartlett.
Membrane Proteins
Red cell membrane proteins were examined in the red cell ghosts. Protein bands after sodium dodecyl sulfate (SDS)-polyacrylamide disc gel electrophoresis (SDS- PAGE) [I l l were stained with Coomassie blue, and examined with a Gilford Spectro- photometer with a gel scanner.
Sodium Influx and Efflux
Sodium transport was examined in the washed red cells [ 121. Red cells sus- pended in 0.154 M Na/K phosphate-buffered saline solution (pH 7.4) with glucose (250 mg/dl) were incubated with 22Na (New England Nuclear NEZ-081) at 37°C for 2 hr, and then washed three times at 4°C with 0.1 M MgC12 solution. The radioactiv- ity of 22Na remaining in the red cells was determined by the y-counter (sodium influx). Sodium efflux was determined by incubating the 22Na-labeled red cells further at 37°C for 2 hr in a medium not containing 22Na. After this incubation, the

Familial LCAT Deficiency 131
radioactivity of 22Na in the incubated red cells was determined. Red cell sodium and potassium were determined by flame photometry.
Membrane-bound Enzymes
Enzymatic activities of adenosine-triphosphatase, acetylcholinesterase, and gly- ceraldehyde-3-phosphate dehydrogenase in red cell ghosts were determined by the method of Hanahan et a1 [ 131, Ellman et a1 [ 141, and McDaniel et a1 [ 151, respectively.
Enzyme Replacement Therapy by Fresh-frozen Plasma Transfusions
The patient was treated with a total of 2200 ml fresh-frozen plasma during 10 days. Red cell membrane lipids and red cell morphology were investigated before and after this procedure. LCAT activities of the fresh-frozen plasma for the transfu- sion were 32.1 +_ 11.6 pmol/ml/hr (N=6, by the method of Stokke and Norum [16]).
RESULTS Case Report
The propositus (M.Y.) is a 49-year-old man. Corneal opacities with marked peripheral lipoid arcus were detected in his childhood, and proteinuria was found at age 34. Edema in his legs was first noticed at age 45. He was admitted to the Jichi Medical School Hospital with complaints of edema in his legs and body weight gain. On admission in October 198 1, marked corneal opacities, edema, hepatosplenomeg- aly, and nephrotic syndrome were noted.
Laboratory findings are shown in Table I. Normal red cell counts with moderate reticulocytosis suggested the presence of compensated hemolysis, as supported by a slight increase of indirect bilirubin in the plasma. Plasma obtained in the fasting state was turbid. The concentration of cholesteryl esters in plasma was markedly reduced, while the total cholesterol concentration was normal. High density lipoprotein (HDL)- cholesterol was markedly reduced. Lipoprotein X was detected as well. The LCAT activity was decreased below 10% of normal level. Glomerular filtration rate was slightly diminished with BUN level just above normal level. Other plasma chemistries did not show overt liver dysfunction.
The morphological study by scanning electron microscopy revealed marked poihlocytosis with occasional target cells in addition to slight anisocytosis (Fig. 2).
Osmotic fragility of red cells was decreased and the red cell life span (51Cr 1/2) was shortened (15 days).
The family tree of this case is shown in Figure 1. The elder sister of the propositus, 59-year-old, also showed marked corneal opacities and slight anemia with target cells, which had been detected in her childhood. She had also very low plasma cholesteryl ester concentration and undetectable HDL-cholesterol with a markedly decreased plasma LCAT activity (below 10 % of normal value). No renal dysfunctions were observed in the elder sister.
These results suggest that the shortened red cell survival with compensated hemolysis may be attributed to abnormal red cell membrane lipids due to familial LCAT deficiency.
Red Cell Membrane Metabolism in Familial LCAT Deficiency
Red cell membrane metabolism was studied in the propositus. The results of red cell membrane lipid analyses are shown in Table 11. The lipid composition of the

132 Murayarna et at
TABLE I. Laboratory Findings in LCAT Deficiency
Patient Normal range
Peripheral blood Red cells Hemoglobin Hematacrit MCV MCH MCHC Reticulocytes
Direct Indirect
LDH Haptoglobin Red cell life span ("Cr T112) Osmotic fragility test
Total bilirubin
Initial Complete
GOT GPT AI-P-tase Cholinesterase lndocyanine green test (I5 min)
Renal function tests BUN Creatinine Glomerular filtration rate Urinary protein
Glomsct-Wright (nmolinil/hr)
Stokkc-Norum (nmolinil/ hr)
Liver function tests
LCAT activities
Plasma lipids Cholesterol
Total Free Estcrified
HDL-cholesterol Lipoprotein X Tridvcerides
418 x 104imm" 410-530 X 104/rn~n' 13.8 gm/dl 40.9% 97.0 p3 33.2 Fpg 33.8% 6% I .4 mg/dl 0.6 mgidl 0.8 mgidl 258 IU 70 nigidl 15 days
0.40% 0.18%
11.7 IU 14.9 IU 210 IU 0.8 ApH 1.4%
28.0 mgidl I .3 mg/dl 69.0 mlimin 9.6 @day
9.62
2.3
194 mgidl 90.6% 9.4% 3 .O mgidl Present 284 m d d l
14.0-18.0 gmidl
80-90 pL3 27-32 ppg 32-36% 0.5-1.0% 0.2-0.8 mgidl 0.0-0.4 mgidl 0.2-0.6 mgidl 150-350 1U 96-350 mgldl 25-32 days
39-52 %
0.44-0.4274 0.34-0.32 '%
3-28 IU 3-28 IU 6 1-225 IU 0.7- I . 1 ApH Below 10%
8-20 mgidl 0.6-1.6 mg/dl 70- I30 mlimin 20-60 mgiday
93.3 16.8
66.3 f 1 1 . 1 (Normal means f SD,N= 1 I )
130-250 mgidl 20-30% 70-80 % 42.1-61.7 mgidl Not detected 76- 172 m d d l
red cell membrane was abnormal. Particularly, the red cell cholesterol was increased up to 1.3 times of the normal mean. The content of total phospholipids was normal, so that the molar ratio of cholesterol to phospholipid was considerably increased. Concomitantly, phosphatidyl choline was increased in the red cell membranes, con- trary to a slight decrease of sphingomyelin. The ratio of the phospholipid contents of the outer surface of red cell membranes (phosphatidyl choline + sphingomyelin) to those of the inner surface (phosphatidyl ethanolamine + phosphatidyl serine) was also elevated chiefly due to the higher concentration of phosphatidyl choline.

Familial LCAT Deficiency 133
I
I d m 4 b/ propositus
10
m 1 2 b 3+J 4&3 Fig. I . A family pedigree of LCAT deficiency: , 0 examined (no anemia, no corneal opacities); , @ examined (slight anemia + corneal opacities); , 0 deceased.
TABLE 11. Red Cell Membrane Lipids in LCAT Deficiency
Patient Control Normala
Cholesterol (&lo8 RBC) 13.30 10.90 10.28 k 1.50 Phospholipids (pgllO* RBC) 23.00 24.00 22.64 f 4.00 Lysophosphatidyl choline (%) 3.0 2.6 2.56 f 0.85 Sphingomyelin (%) 18.0 23.2 22.96 -I 2.38 Phosphatidyl choline (%) 49.9 27.9 28.71 2.58 Phosphatidyl serine (%) 12.0 17.0 17.79 1.27 Phosphatidyl ethanolamine (%) 17.1 29.4 28.00 k 1.33 Cholesterollphospholipids 1.16 0.91 0.91 f 0.11
(molar ratio)
aMean 5 SD from 10 healthy adults.
No abnormalities in membrane protein fractions on SDS-PAGE were observed in the red cells.
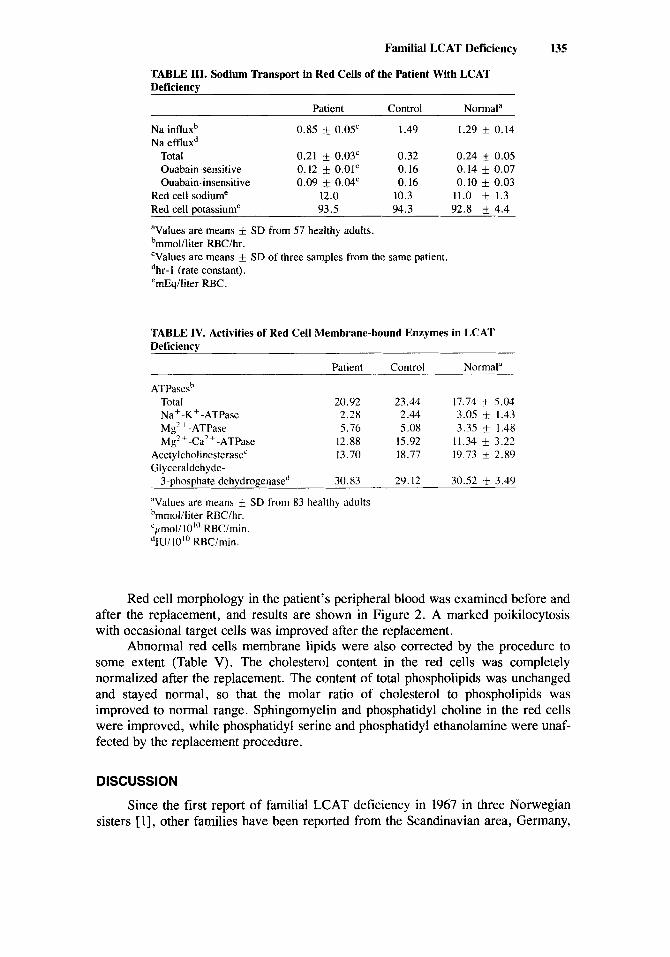
Sodium influx was markedly decreased, compared with the normal control, although sodium efflux, both ouabain-sensitive and ouabain-insensitive, was not altered (Table III). Red cell sodium (12.0 mEq/liter) and potassium (93.5 mEqfiter) were maintained normally during the incubation experiments. The activities of various ATPases were normal (Table IV).
The activities of acetylcholinesterase (AChE) as a marker of the outer surface of red cell membranes and the activities of glyceraldehyde-3-phosphate dehydrogen- ase (G-3-PD) as a marker of the inner surface were also investigated (Table N). The AChE activities in the red cells were slightly decreased, with normal G-3-PD activities.
Enzyme Replacement Therapy by Fresh-frozen Plasma Transfusions The effects of enzyme replacement by fresh-frozen plasma transfusions on red
cell membrane lipids and morphology were investigated. The patient’s plasma was replaced partially with a total of 2200 ml fresh-frozen plasma. During the procedure for 10 days, the patient was placed on an isocaloric diet to control the quality and quantity of lipids from external sources such as foods. The hospital diet was set at 20% of total calories (1900 Cal) as fats, 60% as carbohydrate, and 20% as proteins, respectively. After the replacement procedure, the LCAT activity in the patient’s plasma increased from 9.4% of normal value to 17.4%.
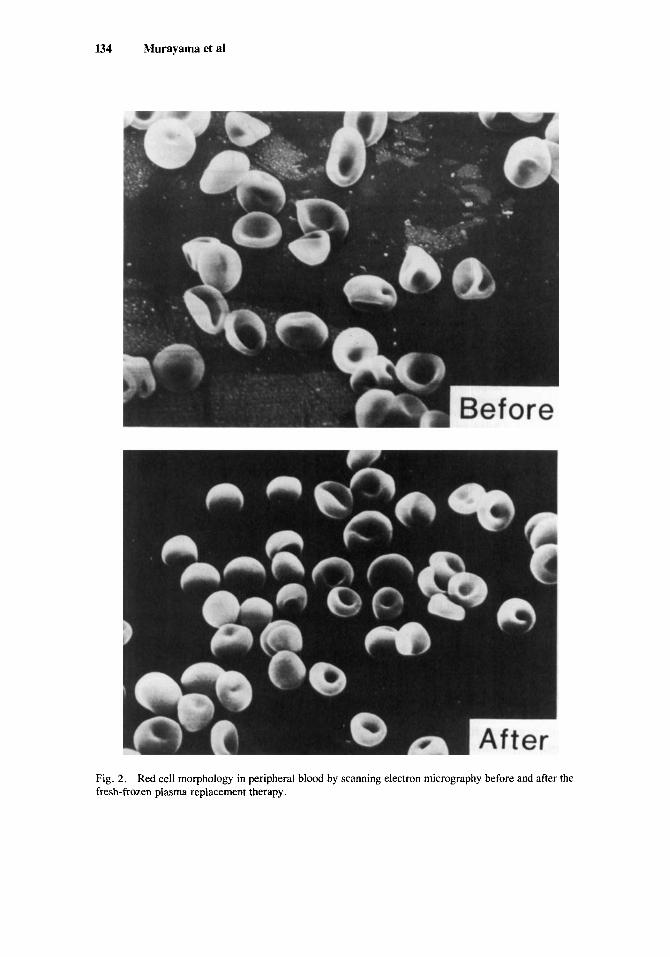
134 Murayama et a1
Fig. 2. fresh-frozen plasma replacement therapy.
Red cell morphology in peripheral blood by scanning electron micrography before and after the
Familial LCAT Deficiency 135
TABLE 111. Sodium Transport in Red Cells of the Patient With LCAT Deficiency
Patient Control Normala
Na influxb 0.85 k 0.05' 1.49 1.29 f 0.14 Na effluxd
Total 0.21 0.03' 0.32 0.24 k 0.05 Ouabain-sensitive 0.12 * 0.01' 0.16 0.14 f 0.07 Ouabain-insensitive 0.09 f 0.04' 0.16 0.10 0.03
Red cell sodiume 12.0 10.3 11.0 + 1.3 Red cell potassiume 93.5 94.3 92.8 f 4.4
Values are means f SD from 57 healthy adults. bmmol/liter RBC/hr. 'Values are means f SD of three samples from the same patient. dhr- 1 (rate constant). emEq/liter RBC.
TABLE IV. Activities of Red Cell Membrane-bound Enzymes in LCAT Deficiency
Patient Control Normal"
ATPasesh Total 20.92 23.44 17.74 k 5.04 Na+-K+-ATPase 2.28 2.44 3.05 +- 1.43 Mg'+-ATPase 5.76 5.08 3.35 k 1.48 Mg' + -Ca' + -ATPase 12.88 15.92 11.34 k 3.22
Acetylcholinesterase' 13.70 18.77 19.73 f 2.89 Glyceraldehyde-
3-phosphate dchydrogenased 30.83 29.12 30.52 f 3.49
"Values are means 'rnrnol/liter RBClhr. 'pnoli 10"' RBClmin. dIU/IO'" RBCimin.
SD from 83 healthy adults
Red cell morphology in the patient's peripheral blood was examined before and after the replacement, and results are shown in Figure 2. A marked poikilocytosis with occasional target cells was improved after the replacement.
Abnormal red cells membrane lipids were also corrected by the procedure to some extent (Table V). The cholesterol content in the red cells was completely normalized after the replacement. The content of total phospholipids was unchanged and stayed normal, so that the molar ratio of cholesterol to phospholipids was improved to normal range. Sphingomyelin and phosphatidyl choline in the red cells were improved, while phosphatidyl serine and phosphatidyl ethanolamine were unaf- fected by the replacement procedure.
DISCUSSION
Since the first report of familial LCAT deficiency in 1967 in three Norwegian sisters [ 11, other families have been reported from the Scandinavian area, Germany,

136 Murayama et al
TABLE V. Effect of Plasma Replacement Therapy by Fresh-frozen Plasma Transfusions on Red Cell Membrane Lipids in LCAT Deficiency
Replacement procedure
Norm a 1 a Lipids Before After
Cholesterol (pg/lO* RBC) 13.30 10.90 10.28 f 1.50 Phospholipids (pg/ 10' RBC) 23.00 22.25 22.64 f 4.00 Lysophosphatidyl choline (%) 3.00 2.5 2.56 f 0.85 Sphingomyelin (%) 18.0 23.3 22.96 + 2.38 Phosphatidyl choline (%) 49.9 43.2 28.71 f 2.58 Phosphatidyl serine (%) 12.0 14.4 17.79 f 1.21 Phosphatidyl ethanolamine (%) 17. I 16.6 28.00 1.33 Cholesterol/phospholipids I . 16 0.97 0.91 f 0.11
(a molar ratio)
aMean f SD from 10 healthy adults.
Canada, France, England, and Japan [17]. In Japan, seven patients in three families including ours have already been found.
The alterations of red cell membrane lipids in the present patient were essentially identical to the results reported previously [4,5]. The content of free cholesterol was increased, with a concomitant increase of phosphatidyl choline. Instead, the contents of sphingomyelin and of phosphatidyl ethanolamine were diminished. The increment of red cell cholesterol is considered to be dependent upon the increased unesterified cholesterol in the plasma, because the red cell cholesterol in the patient was lost rapidly when incubated with normal plasma [ 181. In our fresh-frozen plasma replace- ment therapy, a marked improvement of cholesterol and phosphatidyl choline in red cell membranes was observed. In addition, a marked poihlocytosis with occasional target cells in the patient was also corrected to the nearly normal biconcave disc shape. These results suggest that the abnormal red cell morphology appears to be induced by the increased red cell cholesterol.
Godin et a1 [4] suggested that the functional abnormalities of red cell membrane may exist in this disorder in association with abnormal red cell lipid composition. However, it was not entirely clear whether the red cell membrane functions in this disease were actually altered. In the present study, it is interesting to note that sodium influx was markedly decreased, although sodium efflux, both ouabain-sensitive (Na+- K+ -ATPase dependent) and ouabain-insensitive (Na +-K+-ATPase independent), was normal. Furthermore, the activity of acetylcholinesterase, as the marker of the outer surface of red cell membranes, was decreased. These results suggest that the alteration of red cell membrane lipid may affect sodium influx. Especially, membrane lipids, such as cholesterol, at the outer surface of the cell membrane may have an important regulatory role in membrane permeability, such as sodium influx.
It has been shown that the balance of lipid constituents is important in the control of small molecule permeability in several model membrane systems [ 19-23]. Previous reports [ 19,221 have emphasized the importance of the ratio of cholesterol to phospholipids. It has also been reported that the insertion of cholesterol into a phospholipid array tightens up the array. This change is probably due to the action of intramolecular Van der Waal's forces between the long acyl groups of the adjacent lipid molecules. This alteration causes a contraction of membrane surface by so- called condensation effect of cholesterol [23], resulting in a reduction in membrane permeability [20]. The functional abnormalities of LCAT-deficient red cells may be

Familial LCAT Deficiency 137
related to this mechanism. Thus, it is possible that mild hemolytic anemia in this disease is due to the altered red cell membrane lipids secondary to LCAT deficiency.
ACKNOWLEDGMENTS
This investigation was supported in part by a grant for the Idiopathic Disorders of Hematopoietic Organs from the Ministry of Health and Welfare, the Japanese Government.
REFERENCES
1. Norum KR, Gjone E: Familial plasma 1ecithin:cholesterol acyltransferase deficiency. Biochemical study of a new inborn error of metabolism. Scand J Clin Lab Invest 20:231, 1967.
2. Murayama N, Kato K, Asano Y, Hosoda S, Sakamoto Y, Maesawa M, Yamada N, Kodama T, Murase T, Akanuma Y: Siblings of familial LCAT deficiency; Lipoprotein metabolism with fresh- frozen plasma replacement therapy, red cell membrane lipid composition, and effect of PHLA. Proc JCBL 24:397, 1982, (in Japanese).
3. Gjone E, Torsuik M, Norum KR: Familial plasma cholesterol ester deficiency. A study of the erythrocytes. Scand J Clin Lab Invest 21:327, 1968.
4. Godin DV, Gray GR, Frohlich J: Erythrocyte membrane alterations in 1ecithin:cholesterol acyltrans- ferase deficiency. Scand J Clin Lab Invest 38: 126, 1978.
5. Jain SK, Mohandas N, Sensabaugh GH, Shojania AM, Shohet SB: Hereditary plasma lecithin- cholesterol acyltransferase deficiency. A heterozygous variant with erythrocyte membrane abnor- malities. J Lab Clin Med 99:816, 1982.
6. Dodge JT, Mitchell C, Hanahan D: The preparation and chemical characteristics of hemoglobin free ghosts of human erythrocytes. Arch Biochem Biophys 100:119, 1963.
7. Folch J, Lees M and Sloane Stanley GH: A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:479, 1957.
8. Zak B: Simple rapid microtechnic for serum total cholesterol. Am J Clin Pathol 27:583, 1957. 9. Bartlett GR: Phosphorus assay in column chromatography. J Biol Chem 234:466, 1959.
10. Siakotos AN, Rouser G: Quantitative thin-layer chromatography of lipids by photographic densito- metry. Anal Biochem 14: 162, 1966.
11. Fairbanks G, Steck TL, Wallach DFH: Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 10:2606, 1971.
12. Jacobs HS, Karnovsky ML: Concomitant alterations of sodium flux and membrane phospholipid metabolism in red blood cells: Studies in hereditary spherocytosis. J Clin Invest 46:173, 1967.
13. Hanahan DJ, Ekholm JE: The expression of optimum ATPase activities in human erythrocytes. A comparison of different lytic procedures. Arch Biochem Biophys 187: 170. 1978.
14. Ellman GL, Courtney KD, Andres V Jr, Featherstone RM: A new and rapid colorimetric determi- nation of acetylcholinesterase activity. Biochem Pharmacol 7:88, 1961.
15. McDaniel CF, Kirtley ME, Tanner MJ: The interaction of glyceraldehyde-3-phosphate dehydrogen- ase with human erythrocyte membrane. J Biol Chem 249:6478, 1974.
16. Stokke KT, Norum KR: Determination of 1ecithin:cholesterol acyltransferase in human blood plasma. Scand J Clin Lab Invest 27:21, 1971.
17. Myant NB: “The Biology of Cholesterol and Related Steroids.” London: William Heinemann Medical Books Ltd, 1981, p 796.
18. Demel RA, Kinsky SC, Kinsky CB, van Deenen LLM: Effects of temperature and cholesterol on the glucose permeability of liposomes prepared with natural and synthetic lecithins. Biochim Biophys Acta 150:655, 1968.
19. Gier J, Mandersloot JC, van Deenen LLM: Lipid composition and permeability of liposomes. Biochim Biophys Acta 150:666, 1968.
20. Deuticke B, Gruber W: Anion permeability of mammalian red blood cells: Possible relation to membrane phospholipid patterns. Biochim Biophys Acta 211:369, 1970.
21. Chen LF, Lund DB, Richardson T: Essential fatty acids and glucose permeability of lecithin membranes. Biochim Biophys Acta 225:89, 1971.
22. Vanclenheurel FA: Study of biological structure at the molecular level with stereomodel projections. I. The lipid in the myelin sheath of nerve. J Am Oil Chem Soe 40:455, 1963.
23. Demel RA, van Deenen LLM, Pethica BA: Monolayer interactions of phospholipids and cholesterol. Biochim Biophys Acta 135: 11, 1967.