cross-resistance and herbicide metabolism in grass weeds in europe: biochemical and physiological...
TRANSCRIPT

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
Cross-resistance and herbicide metabolism in grass weeds in Europe: biochemicaland physiological aspectsAuthor(s): Rafael A. De Prado and Antonio R. FrancoSource: Weed Science, 52(3):441-447. 2004.Published By: Weed Science Society of AmericaDOI: http://dx.doi.org/10.1614/P2002-168AURL: http://www.bioone.org/doi/full/10.1614/P2002-168A
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

De Prado and Franco: Grass weed resistance in Europe • 441
Weed Science, 52:441–447. 2004
Cross-resistance and herbicide metabolism in grass weedsin Europe: biochemical and physiological aspects
Rafael A. De PradoCorresponding author. Departamento de QuımicaAgrıcola, Universidad de Cordoba, Campus deRabanales, Edificio Marie Curie, 14071-Cordoba,Spain; [email protected]
Antonio R. FrancoDepartamento de Bioquımica y Biologıa Molecular,Universidad de Cordoba, Campus de Rabanales,Edificio Severo Ochoa, 14071-Cordoba, Spain
In Europe, 18 weedy grass species had been confirmed to have biotypes with resis-tance to herbicides. The most frequent is that of atrazine resistance, with nine re-sistant biotypes found. These biotypes are mainly resistant because of changes in theD1 protein of photosystem II. All atrazine-resistant biotypes, except that of bristlyfoxtail, show cross-resistance to s-triazine and as-triazines. From an agriculture pointof view, the most important cases of resistance are those found in blackgrass, wildoat, Italian ryegrass, rigid ryegrass, and barnyardgrass. In these species, cross- andmultiple resistances were observed due to metabolism or changes in the target proteinby genetic mutations or both. These biotypes are extremely difficult to control withalternative herbicides.
Nomenclature: EPSF synthase, 5-enolpyruvylshikimate-3-phosphate synthase;barnyardgrass, Echinochloa crus-galli (L.) Beauv. ECHCG; blackgrass, Alopecurus myo-suroides Huds. ALOMY; bristly foxtail, Setaria verticillata (L.) Beauv. SETVE; Italianryegrass, Lolium multiflorum Lam. LOLMU; rigid ryegrass, Lolium rigidum GaudinLOLRI; wild oat, Avena fatua L. AVEFA.
Key words: Herbicide, mode of action, resistance, cross-resistance, multiple resis-tance, natural tolerance, resistance mechanisms.
The most important herbicides used nowadays are thosecapable of controlling a broad spectrum of weeds withoutaffecting the crops. This selectivity may be because of phys-ical, or more frequently, biochemical and genetic causes (Ja-ger 1983). This means that certain species of cultivatedplants and weeds can survive the recommended doses ofherbicides. However, they can still be controlled at higherherbicide doses. This is generally known as natural toleranceor the inheritable ability or aptitude of a plant populationto survive and reproduce after a treatment and can be con-sidered as a population characteristic (Retzinger and Mal-lory-Smith 1997). However, genetic variability also occursintraspecifically. Because of these factors and as a result ofthe selection pressure imposed by continuous application ofherbicides, some individual plants develop the capability tosurvive a herbicide application that the same species weresusceptible to. This response is known as resistance. Thisnewly acquired trait of a weed population (or biotype) hasbeen defined as the ability or aptitude of a plant species tosurvive and reproduce itself after treatment with a herbicideat doses normally lethal to that population. In a plant, re-sistance can occur naturally or can be induced by techniquessuch as genetic engineering or the selection of resistant var-iants obtained from tissue cultures or mutagenesis (Retzingerand Mallory-Smith 1997). Although fairly complete, this issomehow a problematic definition because it presumes thatresistance is only associated with factors of physiological ormorphological nature or both. In reality, a broader defini-tion of herbicide resistance should include a more functionalviewpoint, including phenological changes such as dorman-cy (Maxwell and Mortimer 1994). Unlike tolerant plants,resistant biotypes usually not only survive commercial her-bicide doses but also substantially higher herbicide doses(De Prado et al. 1996).
The concepts of cross-resistance and multiple resistance
have also arisen. Definitions of these terms vary but aregenerally associated with resistance mechanisms. Jutsum andGraham (1995) postulates that cross-resistance happenswhen an individual is resistant to more than one herbicidebecause of a single resistance mechanism and multiple resis-tance when an individual has more than one mechanism ofresistance to one, several, or several herbicides belonging tothe same or different chemical family. Rubin (1997) definedcross-resistance as a case where a weed population is resistantto two or more herbicides acting at the same primary siteof action and multiple resistance, when a population is re-sistant to two or more herbicides acting at a different site.
The term tolerance is frequently used not only to refer tovariations in ability to withstand a herbicide application be-tween different species but also when there is variabilitywithin a population of the same species (Gasquez 1997).Tolerance and resistance are also regarded as terminologiesdenoting differences in the intensity of the same phenom-enon. Resistance is considered as an extreme, but less fre-quent, case of tolerance (Holt and LeBaron 1990; Mazurand Falco 1989). Some consider tolerance as being a poly-genic mechanism and resistance as monogenic (Gressel2002). For this article, the definitions of Jutsum and Gra-ham (1995) in relation to cross-resistance and multiple re-sistance will be used because they fit best to the type of casesbeing discussed.
The development of resistance is associated, inevitably,with the introduction of different types of pesticides, themost blatant case being that of insecticides. Today, there areover 400 known species of insects, some of them agriculturalpests, that have developed resistance to one or several ofthese chemicals (Ware 1994). A similar situation has beenfound with other types of pesticides, such as antibiotics orrodenticides, in which resistance appeared shortly after theirintroduction. Although this was also predicted for herbi-

442 • Weed Science 52, May–June 2004
cides, resistance to herbicides generally developed over a lon-ger period of time than that to other pesticides. For exam-ple, the triazines were introduced in the 1950s. It was notuntil the end of the 1960s that the first case of herbicideresistance was recorded: a biotype of common groundsel (Se-necio vulgaris L.) resistant to s-triazines, atrazine and sima-zine, isolated from a conifer field in Washington state wheresimazine was repeatedly used for many years (Ryan 1970).Since then, the number of herbicide-resistant weeds detectedhas notoriously increased. A review by Ian Heap(www.weedscience.com) shows that more than 258 resistantweed biotypes exist. Most of these biotypes were discoveredin Europe and North America, but the number of casesdetected in other regions of the world is also surprisinglyhigh.
This article discusses the occurrence of herbicide-resistantgrass weeds in Europe and an overview of resistance mech-anisms. Discussion on resistance mechanisms focuses on cas-es of metabolic-based resistance.
Grass Weeds Resistant to Herbicides in Europe
The number of herbicide-resistant dicotyledonous plants(94) is 1.5 times greater than that of monocots (62). Thiscould be because of the large number of broadleaf speciesinfesting corn (Zea mays) fields, one of Europe’s main cul-tivated crops. Many gramineae, such as those of the subfam-ily Panicoides (giant foxtail, Setaria faberi Herrm,; bristlyfoxtail; and giant green foxtail, S. viridis var major Gaudin),show different tolerance levels (Jensen et al. 1977). Thesespecies detoxify triazines through conjugation with gluta-thione as corn does (De Prado et al. 1995a; Lopez-Martınezet al. 1997). This natural tolerance mechanism in mono-cotyledonous plants is responsible for the less frequent ap-pearance of resistant biotypes in these plants than in dicot-yledonous species (Gimenez-Espinosa et al. 1996).
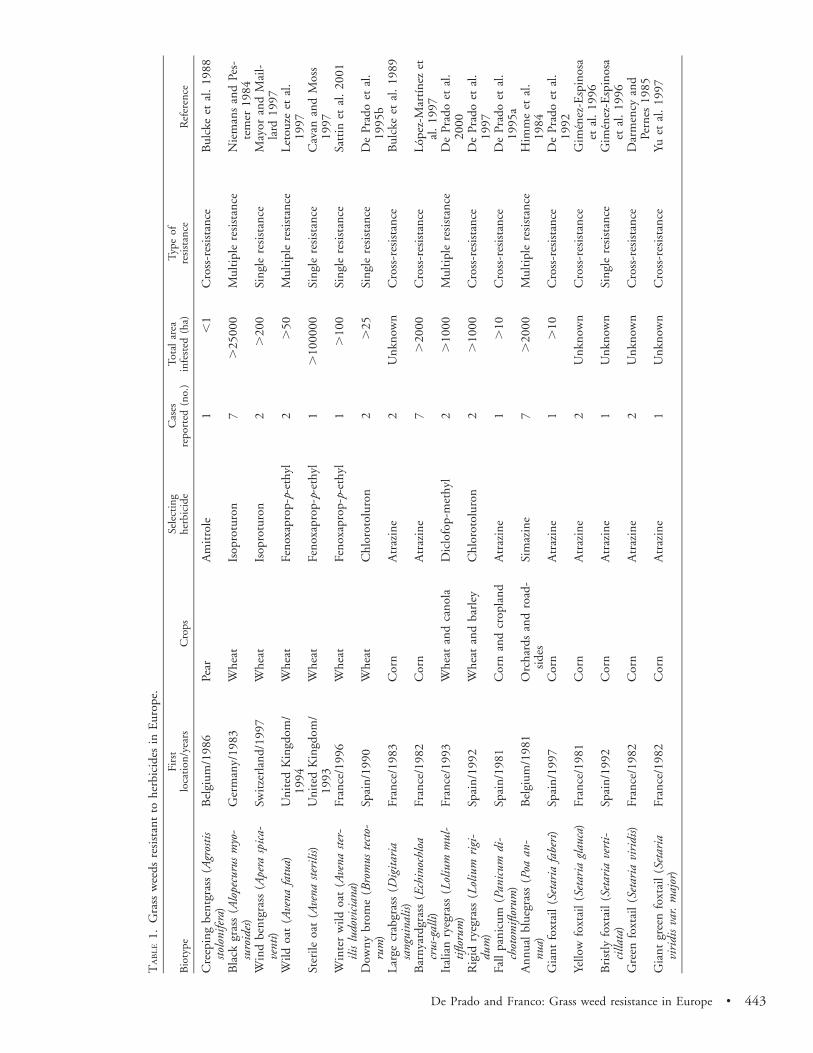
Of the weed species listed in Table 1, perhaps the mostwidespread and troublesome are blackgrass, barnyardgrass,and a number of Lolium species, particularly Italian ryegrassand rigid ryegrass. In Germany, blackgrass infesting wheat(Triticum aestivum) fields acquired resistance to ureas andamides in 1983 (Niemans and Pestemer 1984). Currently,resistant blackgrass infests more than 25,999 ha in most partof Europe (Belgium, France, Germany, Spain, Switzerland,Netherlands, and United Kingdom). Blackgrass populationshave been reported resistant to acetyl co-A carboxylase (AC-Case) and acetolactate synthase (ALS) inhibitors, dinitroan-ilines, photosystem II (PS II) inhibitors, ureas, and amides.Thus, cross- and multiple resistance in blackgrass is one ofEurope’s most serious weed problem. In some instances, fewherbicides remain that can control the resistant biotype.
The Echinochloa spp. are major weeds wherever rice (Ory-za sativa) is grown and are the most intractable resistanceproblem in this cultivar. In Greece, barnyardgrass firstevolved resistance to ureas and amides in 1989. Resistantbiotypes have been detected in 20 sites in Greece and Spainand are spread over approximately 100 ha. Barnyardgrassresistance to ACCase and ALS inhibitors, to dinitroanilines,and to synthetic auxins has been studied in Italy and Spain(Lopez-Martınez et al. 1997; Sattin et al. 1999). Triazine-resistant barnyardgrass has achieved particular notoriety be-cause it infests large areas, being detected in cornfields of
seven countries. A fewer number of weed species resistantto ALS-inhibitor herbicide have appeared in Europe becauseof less usage of ALS herbicides in Europe compared withNorth America. The majority of Echinochloa spp. resistantbiotypes can still be easily controlled by alternative herbi-cides (Heap and LeBaron 2001).
Italian ryegrass and rigid ryegrass are major weeds inwheat, canola (Brassica napus), and barley (Hordeum vulgare)fields in France, Greece, Italy, Spain, and United Kingdom(Table 1). Although ryegrass species are not so widespreadas blackgrass, they show the same cross- and multiple resis-tance problems in Europe. In southern Spain, a populationof Italian ryegrass with high resistance to glyphosate hasbeen detected. This occurred in olive (Olea europea L.) or-chards where glyphosate has been used repeatedly to controlgrass weeds.
Resistance Mechanisms in PlantsThere are at least four mechanisms identified by which
plants become resistant to a herbicide. The two most com-mon mechanisms are those involving metabolic reactionsand changes in the deoxyribonucleic acid sequence (muta-tions) that alter the structure and features of the target pro-teins (Gressel 2002; Hatzios 2001). Metabolism eventuallychanges the chemical nature of the herbicide, and hence itsproperties. These processes account for more cases of naturalresistance than any other mechanism. Tolerant plants candetoxify the herbicide quickly enough to avoid accumula-tion up to toxic levels. With persistent herbicide pressure,resistant biotypes acquire the capability of detoxifying theherbicide. The quicker the plant can alter or modify thesemolecules, the higher the degree of resistance that will ensue.Mutations on the target site modify the interactions of thetarget proteins with the herbicides, resulting in inefficientbinding or total loss of binding capacity. Thus far, targetsite mutation is the most common cause of resistance toherbicides. Alteration of target site will not be discussed indetail in this study.
The other two mechanisms involve either an alteration inthe penetration or translocation of the herbicides within theplant or the overexpression of the target protein as a con-sequence of other gene modifications (gene duplication orenhanced transcription). These latter processes mainly alterthe ratio of binding site to herbicide molecules. If morebinding sites are available relative to herbicide molecules,then the target biochemical pathway will not be totally im-paired. These mechanisms have been reported to occur(Donn et al. 1984; Shyr et al. 1992; Watanabe et al. 1998)in a few cases only, and therefore, will not be discussed inthis study.
Resistance Mediated by Metabolic ReactionsThe transformation of herbicides in plants can be roughly
attributed to nonsynthetic and synthetic metabolic reac-tions. Nonsynthetic reactions comprise redox reactions(mainly oxygenation and hydroxylation) and hydrolysis.These are classified as Phase-I metabolic reactions (Devineet al. 1993). In synthetic reactions, the herbicides conjugatewith glutathione, sugars, or other molecules as amino acids(Hatzios 1997). These are classified as Phase-II reactions. Inboth cases, the chemical nature of the herbicide changes to
De Prado and Franco: Grass weed resistance in Europe • 443
TA
BL
E1.
Gra
ssw
eeds
resi
stan
tto
herb
icid
esin
Eur
ope.
Bio
type
Firs
tlo
cati
on/y
ears
Cro
psSe
lect
ing
herb
icid
eC
ases
repo
rted
(no.
)To
tal
area
infe
sted
(ha)
Typ
eof
resi
stan
ceR
efer
ence
Cre
epin
gbe
ntgr
ass
(Agr
osti
ssto
loni
fera
)B
elgi
um/1
986
Pear
Am
itro
le1
,1
Cro
ss-r
esis
tanc
eB
ulck
eet
al.
1988
Bla
ckgr
ass
(Alo
pecu
rus
myo
-su
roid
es)
Ger
man
y/19
83W
heat
Isop
rotu
ron
7.
2500
0M
ulti
ple
resi
stan
ceN
iem
ans
and
Pes-
tem
er19
84W
ind
bent
gras
s(A
pera
spic
a-ve
nti)
Swit
zerl
and/
1997
Whe
atIs
opro
turo
n2
.20
0Si
ngle
resi
stan
ceM
ayor
and
Mai
l-la
rd19
97W
ildoa
t(A
vena
fatu
a)U
nite
dK
ingd
om/
1994
Whe
atFe
noxa
prop
-p-e
thyl
2.
50M
ulti
ple
resi
stan
ceL
etou
zeet
al.
1997
Ster
ileoa
t(A
vena
steri
lis)
Uni
ted
Kin
gdom
/19
93W
heat
Feno
xapr
op-p
-eth
yl1
.10
0000
Sing
lere
sist
ance
Cav
anan
dM
oss
1997
Win
ter
wild
oat
(Ave
naste
r-ili
slu
dovi
cian
a)Fr
ance
/199
6W
heat
Feno
xapr
op-p
-eth
yl1
.10
0Si
ngle
resi
stan
ceSa
ttin
etal
.20
01
Dow
nybr
ome
(Bro
mus
tect
o-ru
m)
Spai
n/19
90W
heat
Chl
orot
olur
on2
.25
Sing
lere
sist
ance
De
Prad
oet
al.
1995
bL
arge
crab
gras
s(D
igit
aria
sang
uina
lis)
Fran
ce/1
983
Cor
nA
traz
ine
2U
nkno
wn
Cro
ss-r
esis
tanc
eB
ulck
eet
al.
1989
Bar
nyar
dgra
ss(E
chin
ochl
oacr
us-g
alli)
Fran
ce/1
982
Cor
nA
traz
ine
7.
2000
Cro
ss-r
esis
tanc
eL
opez
-Mar
tıne
zet
al.
1997
Ital
ian
ryeg
rass
(Lol
ium
mul
-ti
floru
m)
Fran
ce/1
993
Whe
atan
dca
nola
Dic
lofo
p-m
ethy
l2
.10
00M
ulti
ple
resi
stan
ceD
ePr
ado
etal
.20
00R
igid
ryeg
rass
(Lol
ium
rigi
-du
m)
Spai
n/19
92W
heat
and
barl
eyC
hlor
otol
uron
2.
1000
Cro
ss-r
esis
tanc
eD
ePr
ado
etal
.19
97Fa
llpa
nicu
m(P
anic
umdi
-ch
otom
iflor
um)
Spai
n/19
81C
orn
and
crop
land
Atr
azin
e1
.10
Cro
ss-r
esis
tanc
eD
ePr
ado
etal
.19
95a
Ann
ual
blue
gras
s(P
oaan
-nu
a)B
elgi
um/1
981
Orc
hard
san
dro
ad-
side
sSi
maz
ine
7.
2000
Mul
tipl
ere
sist
ance
Him
me
etal
.19
84G
iant
foxt
ail
(Set
aria
fabe
ri)
Spai
n/19
97C
orn
Atr
azin
e1
.10
Cro
ss-r
esis
tanc
eD
ePr
ado
etal
.19
92Ye
llow
foxt
ail
(Set
aria
glau
ca)
Fran
ce/1
981
Cor
nA
traz
ine
2U
nkno
wn
Cro
ss-r
esis
tanc
eG
imen
ez-E
spin
osa
etal
.19
96B
rist
lyfo
xtai
l(S
etar
iave
rti-
cilla
ta)
Spai
n/19
92C
orn
Atr
azin
e1
Unk
now
nSi
ngle
resi
stan
ceG
imen
ez-E
spin
osa
etal
.19
96G
reen
foxt
ail
(Set
aria
viri
dis)
Fran
ce/1
982
Cor
nA
traz
ine
2U
nkno
wn
Cro
ss-r
esis
tanc
eD
arm
ency
and
Pern
es19
85G
iant
gree
nfo
xtai
l(S
etar
iavi
ridi
sva
r.m
ajor
)Fr
ance
/198
2C
orn
Atr
azin
e1
Unk
now
nC
ross
-res
ista
nce
Yuet
al.
1997

444 • Weed Science 52, May–June 2004
FIGURE 1. Schematic summary of reactions dealing with herbicide detoxi-fication in a plant cell. (A) denotes nonsynthetic reactions such as oxidation,reduction, and hydrolysis. (B) denotes synthetic reactions. In both cases,the chemical nature of the herbicide is modified. (C) denotes cases whereherbicides are sequestrated into organelles or other compartments prevent-ing their interactions with the target. Putative fates of altered herbicides arealso presented. CytP450, Cytochrome P450 monoxygenase; GT, glucosyl-transferases; GST, glutathione-S-transferase. Other transferases includetransaminases or transferases other than GST. Enzymes with esterase, phos-phatase, or amidase activity are grouped as hydrolases. The ring moleculesrepresent the herbicide molecules. These detoxification reactions are of vary-ing complexity, and in many cases simultaneously involve more than onetype of reaction.
a form that has reduced or no toxicity. The eventual fate ofaltered or original herbicide molecules is variable. Some ofthem are retained in the cytosol, compartmentalized in thevacuole, or excreted outside the cell where the molecules aredeposited in the cell walls. These are classified as Phase-IIIreactions. In the vacuole, these molecules can be furthertransformed or hydrolyzed. A schematic diagram is present-ed in Figure 1. Note that plants will show resistance if un-altered herbicide molecules are kept from interacting withthe target site (Fuerst and Vaughn 1990).
Among the oxidative enzymes that facilitate detoxificationof herbicides, the most widely studied are the cytochromeP-450 monooxygenases (CytP450s). These comprise severalenzyme families able to catalyze more than 60 reactions thatexhibit distinctive specificity in tissue and substrate. Theseenzymes are the main focus of research nowadays becausethey seem to be involved in more cases of resistance thanany other type of metabolic reactions. The number ofcloned genes encoding for these enzymes has dramaticallyincreased in recent years (Gressel 2002). However, manydifferent aspects remain to be unraveled, such as the actualfunction of a specific type of CytP450 and how many ofthese enzymes are actually involved in a specific case of re-sistance (Hatzios 1997).
Recent attention has also shifted toward characterizationof hydrolytic enzymes, mainly esterases. Hydrolysis can oc-cur if the herbicide is a carboxylic acid ester or contains anamide or phosphate group. Hydrolases split the herbicidemolecule producing generally nonherbicidal metabolites,which are further processed in the plant. If the activity ofthese hydrolases increases with repeated exposure to the her-bicide or by selection of individuals with higher level of
esterase activity, it can account for resistance. However, insome cases, these activities are responsible for the conversionof a proherbicide to its active form. If this is the case, lackof hydrolytic activity leads to resistance (Devine et al. 1993).
Research on conjugation or Phase-II reactions are devotedlargely to the study of glutathione S-transferases (GST)(Marrs 1996). These enzymes are present in all stages ofplant development and in every tissue examined to date.They encompass a gene family with varying levels of spec-ificity, although only some of them are involved in detoxi-fication (McGonigl et al. 1997). In some cases, homoglu-tathione is the molecule transferred to the herbicide (Skipseyet al. 1997). Other processes involve the formation of gly-coside and amino acid conjugates. Many different types ofreactions, such as peptide hydrolysis, sulfur oxygenation,and malonylation, can further metabolize all types of con-jugated or modified herbicide molecules. Most glycosilatedherbicides enter the vacuole, whereas amino acid–herbicideconjugates are mainly excreted outside the cell and depositedin the cell wall. A plant can be resistant to a particularherbicide by one or more of these mechanisms.
Cross- and Multiple Resistances inGrass Weeds
Understanding the mechanisms and genetics of resistancecan lead to more effective management of herbicide-resistantweeds. This information is important in determining whichherbicides should be used in combinations or in rotationsto prevent either target site– or metabolism-based resistance.In addition, as new herbicides are discovered, we may beable to use the information obtained in the laboratory topredict the mechanisms and the genetics of resistance beforeit occurs in the field. This information can then be used tointegrate new herbicides into the weed management pro-gram in a manner that minimizes the chances for selectingherbicide-resistant biotypes (Shaner 1995).
Grass Weeds Resistant to Photosynthesis-Inhibiting Herbicides
The first grass weed that evolved resistance to atrazinewas a biotype of yellow foxtail (Setaria glauca (L.) Beauv.SETLU) in France (Table 1). All cases of resistance were asa result of amino acid substitution in the D1 protein of PSII (Gronwald 1997). The most noticeable cases are those ofgreen (Setaria viridis (L.) Beauv. SETVI) and yellow foxtails.Both species have developed resistance to atrazine becauseof combined mechanisms of enhanced metabolism and tar-get site modification. The resistant biotypes showed en-hanced glutathione S-transferase activity and are able to con-jugate atrazine with glutathione (De Prado et al. 1999). Inaddition, the resistant biotypes also displayed changes in theaffinity of D1 protein to atrazine. These biotypes showcross-resistance to s-triazine and as-triazines and did nothave resistance to other herbicide families.
Resistance to photosystem II–inhibiting herbicides, otherthan triazines, is rare. Four weed species have developedresistance to phenylurea herbicides, and one has acquiredresistance to the amide herbicide propanil (Table 1). Thewidespread occurrence of chlorotoluron- and isoproturon-resistant populations of blackgrass and rigid ryegrass are ofparticular concern because these populations have the ability

De Prado and Franco: Grass weed resistance in Europe • 445
to metabolize a wide range of herbicides with differentmodes of action through ring-hydroxylation (De Prado etal. 1997). Both biotypes are able to detoxify five herbicides(chlorotoluron, chlorsulfuron, diclofop-methyl, simazine,and pendimethalin). Preston et al. (1996) have proposedthat this can be attributed to at least four different CytP450enzymes.
Several populations of barnyardgrass have gained resis-tance to propanil in rice fields in Greece and Spain. Resis-tance of barnyardgrass to propanil is due to higher level ofhydrolysis of propanil in the resistant compared with thesusceptible biotype (Carey et al. 1997). The rate of conver-sion of propanil to its nonherbicidal metabolite 3,4-di-chloroaniline is similar between the resistant barnyardgrassand rice (Carey et al. 1997). It was concluded that barn-yardgrass resistance to propanil is due to elevated activity ofarylacylamidase in the resistant biotype, although this en-zyme was strongly inhibited by carbamate or organophos-phate insecticides recommended for use in rice (Lopez-Mar-tınez et al. 2001). Arylacylamidase is the enzyme that cat-alyzes the hydrolysis of propanil in rice (Giannopolitis andVassiliou 1989).
Grass Weeds Resistant to Membrane DisruptersThe first report of resistance to paraquat was a biotype of
annual bluegrass (Poa annua L. POAAN) isolated in Bel-gium and United Kingdom (Clay 1989; Himme et al.1984). This population showed multiple resistance to ami-trole and simazine. In general, two theories have been pro-posed to explain the resistance to the bipyridilium herbi-cides: detoxification of highly reactive oxygen species andsequestration of the herbicide in nonchloroplast sites (Fuerstand Vaughn 1990).
Grass Weeds Resistant to ACCase-InhibitingHerbicides
The aryloxyphenoxypropionate (AOPP) and the cyclo-hexanedione (CHD) herbicides provide excellent control ofgrass weeds in dicotyledonous and some grass crops. In themajority of AOPP- and CHD-resistant weed species ana-lyzed to date, resistance is conferred by an ACCase alterationthat results in reduced herbicide sensitivity (Devine 1997).Recent studies suggest that different patterns of cross-resis-tance to ACCase inhibitors could be because of differentACCase mutations in the resistant biotypes. Cross-resistanceto various AOPP and CHD herbicides has been describedin blackgrass, wild oat, Italian ryegrass, and rigid ryegrass(Devine 1997; De Prado et al. 1998, 2000).
Resistance to diclofop-methyl has been discovered inmany populations of blackgrass and some populations ofrigid ryegrass, which are generally resistant to chlorotoluronas well (De Prado et al. 1998; Menendez and De Prado1997). Diclofop-methyl is metabolized more rapidly in allthe resistant biotypes than in the susceptible one. This met-abolic detoxification is catalyzed in both the resistant andsusceptible biotypes of these species by CytP450 monoox-ygenases. Inhibition of CytP450 by ABT (1-aminobenzo-triazole) results in a strong reduction in the hydroxylationof diclofop (Menendez and De Prado 1997). Research re-sults indicate that monooxygenase activity seems to be high-er in resistant than in susceptible biotypes. The same mech-
anism of resistance to substituted ureas, ALS- and ACCase-inhibitor herbicides in blackgrass and rigid ryegrass makesit extremely difficult to control the resistant biotypes withalternative herbicides. Therefore, it is difficult to predict theeffectiveness of herbicides on these biotypes, as these pop-ulations develop resistance to herbicides even after short pe-riods of repeated herbicide application (De Prado and Me-nendez 1996).
Grass Weeds Resistant to ALS-InhibitingHerbicides
Resistance to acetolactate synthase inhibitors has evolvedquickly in both crop and noncrop situations after 5 to 10applications of these herbicides. In Europe, there have beenfour grass weeds resistant to ALS inhibitors: blackgrass, wildoat, Italian ryegrass, and rigid ryegrass (Table 1). Some pop-ulations of blackgrass and wild oat found in the UnitedKingdom showed multiple resistance to chlorotoluron,chlorsulfuron, diclofop-methyl, fluazifop-butyl, imazameth-abenz-methyl, pendimethalin, simazine, tribenuron, andtriallate herbicides. In both species, resistance to ALS-inhib-iting herbicides is because of a less sensitive ALS (Hall et al.1994), whereas resistance to other herbicides is due to en-hanced metabolism (Cocker et al. 1999). It is postulatedthat CytP450 and GST are involved in the enhanced de-toxification reactions. Some populations of these ALS-resis-tant biotypes showed cross-resistance to ALS inhibitorsthrough enhanced detoxification by CytP450 (Durst et al.1997; Gressel 2002).
Grass Weeds Resistant to Mitotic Disrupters
The dinitroaniline herbicides provide . 90% control ofweeds for up to 10 wk in many environments and season-long control under some situations. Despite the extensiveand sustained use of these herbicides, only a few cases ofresistance have been reported in Europe. In two cases, re-sistance to dinitroaniline herbicides has been discovered ingrass weeds with cross-resistance to other herbicide families(De Prado and Menendez 1997; Smeda and Vaughn 1997).Blackgrass showed the highest level of tolerance to pendi-methalin, but it was as sensitive to trifluralin as the suscep-tible biotype, whereas rigid ryegras showed high-resistancelevel to both herbicides (De Prado and Menendez 1997).Biochemical studies done on both resistant biotypes indicatethat resistance to substituted ureas, ALS- and ACCase-in-hibitor herbicides is because of the enhanced ability of thesebiotypes to degrade herbicides through oxidative processes(Moss 1990).
Grass Weeds Resistant to Auxinic Herbicides
Quinclorac is an herbicide that was developed for selec-tive control of Echinochloa spp. in rice. This was first usedin Spain in 1987, and it was only a few years later whenresistant biotypes of barnyardgrass were isolated from ricefields (Lopez-Martınez et al. 1998). It was proposed thatplants sensitive to quinclorac accumulate high levels ofHCN, which result from overstimulation of ethylene bio-synthesis by the herbicide (Grossmann and Scheltrup 1997;Lopez-Martınez et al. 1998). The mechanism of resistanceto quinclorac is not yet properly understood.

446 • Weed Science 52, May–June 2004
Grass Weeds Resistant to Glyphosate
Glyphosate is a nonselective herbicide used to controlweeds in orchards and in direct-seeded fields. Resistance toglyphosate has been detected in goosegrass (Eleusina indica)(Malaysia), Italian ryegrass (USA and Chile), and rigid rye-grass (Australia) after more than 15 yr of repeated use (http://www.weedscience.com). The only resistance mechanismknown is that of goosegrass, which contains an EPSP syn-thase enzyme with reduced sensitivity to glyphosate (Lee andNgim 2000). In Spain, glyphosate is used extensively forcontrolling Lolium spp. in olive fields, where 720 to 1,080g ai ha21 has been applied since 1985. To date, three pop-ulations of Lolium spp. had already developed resistance toglyphosate and are being characterized (De Prado, unpub-lished data). Resistance to glyphosate is generally not wellunderstood, and may differ between species. This is one areathat needs further study.
Conclusions
Similar to other developed regions, European agriculturerelies heavily on herbicides for weed management. Herbi-cide-resistant weeds will remain an important issue to re-solve. Understanding the mechanism(s) of resistance is im-portant to determine appropriate strategies for managementof resistant populations. Resistant biotypes with multiplemechanisms of resistance present a management challenge.Research tools in molecular biology have made it easier todetermine target-site alterations as a mechanism of resis-tance. This gives us an idea of possible cross-resistance andmultiple-resistance patterns. Metabolic-based resistancemechanisms could be more complicated to study because ofthe numerous types of enzymes involved in metabolic re-actions. There is a need to do more research on monooxy-genases, esterases, glutathione transferases, and other en-zymes that can modify xenobiotics in plants.
Acknowledgments
We thank Dr. Nilda Burgos (University of Arkansas, Fayetteville)for kindly reviewing the manuscript. This work has been in partsupported by Direccion General de Investigacion, Ministerio deCiencia y Tecnologıa, Spain, Grants AGL2000-1713-C03-01 andAGL2000-1713-C03-02.
Literature CitedBulcke, R., M. Himme van, J. Stryckers, and M. Van Himme. 1988. Tol-
erance to amitrole in weeds in long-term experiments in fruit plan-tations. Pages 287–295 in VIIIe Colloque International sur la Biologie,l’Ecologie et la Systematique des Mauvaises Herbes. Volume 1. Paris,France: Association National de Protection des Plantes.
Bulcke, R., M. Himme van, J. Stryckers, and M. Van Himme. 1989. Shiftsof a survey of weed biotypes resistant to atrazine. Pages 315–319 inProceedings 1992 Congress of the Spanish Weed Society. Valencia,Spain: Spanish Weed Society.
Carey, V. F., R. E. Hoagland, and R. E. Talbert. 1997. Resistance mecha-nism of propanil-resistant barnyardgrass: II. In-vivo metabolism of thepropanil molecule. Pestic. Sci. 49:333–338.
Cavan, G. and S. Moss. 1997. Herbicide resistance and gene flow in black-grass (Alopecurus myosuroides) and wild oats (Avena spp.). Pages 305–310 in Proceedings of the Brighton Crop Protection Conference—Weeds. Volume 1. Farnham, U.K.: British Crop Protection Council.
Clay, D. V. 1989. New developments in triazine and paraquat resistanceand co-resistance in weed species in England. Pages 317–324 in Pro-
ceedings of the Brighton Crop Protection Conference—Weeds. Vol-ume 1. Farnham, U.K.: British Crop Protection Council.
Cocker, K. M., S. R. Moss, and J.O.D. Coleman. 1999. Multiple mecha-nisms of resistance to fenoxaprop-P-ethyl in United Kingdom andother European populations of herbicide-resistant Alopecurus myosu-roides (black-grass). Pestic. Biochem. Physiol. 65:189–195.
Darmency, H. and J. Pernes. 1985. Use of wild Setaria viridis (L.) Beauv.to improve resistance in cultivated S. italica (L.) by hybridization.Weed Res. 25:175–179.
De Prado, J. L., M. D. Osuna, R. H. Shimabukuro, and R. De Prado.1998. Biochemical and physiological resistance mechanisms to diclo-fop-methyl in Lolium rigidum. Meded. Fac. Landbouwwet. 63:681–689.
De Prado, R., J. L. De Prado, and J. Menendez. 1997. Resistance to sub-stituted urea herbicide in Lolium rigidum biotypes. Pestic. Biochem.Physiol. 57:126–136.
De Prado, R., R. Gimenez-Espinosa, J. Gonzalez-Gutierrez, J. Menendez,J. Gasquez, and J. W. Gronwald. 2000. Resistance to acetyl CoA car-boxylase inhibiting-herbicides in a Lolium multiflorum biotype fromFrance. Weed Sci. 48:311–318.
De Prado, R., N. Lopez-Martınez, and J. Gonzalez-Gutierrez. 1999. Iden-tification of two mechanisms of atrazine resistance in Setaria faberiand Setaria viridis biotypes. Pestic. Biochem. Physiol. 67:114–124.
De Prado, R. and J. Menendez. 1996. Management of herbicide-resistantgrass weeds in Europe. Pages 393–398 in Second International WeedControl Congress. Copenhagen, Denmark: International Weed Sci-ence Society.
De Prado, R. and J. Menendez. 1997. Cross-resistance and herbicide me-tabolism in Alopecurus myosuroides Huds. Pages 351–366 in K. K.Hatzios, ed. Regulation of Enzymatic Systems Detoxifying Xenobioticsin Plants. NATO ASI Series. High Technology. Volume 37. Dor-drecht, The Netherlands: Kluwer.
De Prado, R., K. L. Plaisence, J. Menendez, and J. W. Gronwald. 1996.Effect of graminicide herbicides on growth and ACCase activity in achlorotoluron-resistant biotype of Alopecurus myosuroides. Pages 59–61in Proceedings of the International Symposium on Weed and CropResistance to Herbicides. Cordoba, Spain: University of Cordoba.
De Prado, R., E. Romera, and J. Menendez. 1995a. Atrazine detoxificationin Panicum dichotomiflorum and target site Polygonum lapathifolium.Pestic. Biochem. Physiol. 52:1–11.
De Prado, R., E. Romera, and J. Menendez. 1995b. Chorotoluron resis-tance in a Bromus tectorum L. biotype is due to enhanced detoxificationprocesses. Pages 62–64 in International Symposium on Weed andCrop Resistance to Herbicides, Cordoba, Spain. Dordrecht, The Neth-erlands: Kluwer.
De Prado, R., E. Romera, J. Menendez, and M. Tena. 1992. Mechanismof resistance to atrazine in Setaria verticillata and Setaria faberi. Pages457–463 in IXe Colloque International sur la Biologie des MauvaisHerbes, Dijon, France: Association Francaise de Protection des Plantes.
Devine, M. D., S. O. Duke, and C. Fedtke. 1993. Physiology of HerbicideAction. Englewood Cliffs, NJ: PTR Prentice Hall.
Devine, M. D. 1997. Target-site based resistance to ACCase inhibitors.Pages 61–69 in R. De Prado, J. Jorrın, and L. Garcıa-Torres, eds.Weed and Crop Resistance to Herbicides. Dordrecht, The Nether-lands: Kluwer.
Donn, G., E. Tischer, J. A. Smith, and H. M. Goodman. 1984. Herbicide-resistant alfalfa cells: an example of gene amplification in plants. J.Mol. App. Genet. 2:621–635.
Durst, F., J. P. Salaun, D. Werck-Reichhart, and F. Zimmerlin. 1997. Cy-tochrome P450 endowed herbicide metabolism. Pages 101–108 in R.De Prado, J. Jorrın, and L. Garcıa-Torres, eds. Weed and Crop Resis-tance to Herbicides. Dordrecht, The Netherlands: Kluwer.
Fuerst, E. P. and K. C. Vaughn. 1990. Mechanism of paraquat resistance.Weed Technol. 4:150–156.
Gasquez, J. 1997. Genetic of herbicides resistance within weeds. Factors ofevolution, inheritance and fitness. Pages 181–189 in R. De Prado, J.Jorrın, and L. Garcıa-Torres, eds. Weed and Crop Resistance to Her-bicides. Dordrecht, The Netherlands: Kluwer.
Giannopolitis, C. N. and G. Vassiliou. 1989. Propanil tolerance in Echin-ochloa crus-galli (L.) Beauv. Trop. Pest Manag. 35:6–7.
Gimenez-Espinosa, R., E. Romera, M. Tena, and R. De Prado. 1996. Fateof atrazine in treated and pristine accessions of three Setaria species.Pestic. Biochem. Physiol. 56:196–207.
Gressel, J. 2002. Molecular Biology of Weed Control. New York: Taylor &Francis.
Gronwald, J. W. 1997. Resistance to PS II inhibitors herbicides. Pages 53–

De Prado and Franco: Grass weed resistance in Europe • 447
59 in R. De Prado, J. Jorrın, and L. Garcıa-Torres, eds. Weed andCrop Resistance to Herbicides. Dordrecht, The Netherlands: Kluwer.
Grossmann, K. and F. Scheltrup, 1997. Selective induction of 1-aminocy-clopropane-1-carboxylic acid (ACC) synthase gene in transgenicplants. Plant Growth Regul. 16:183–188.
Hall, L. M., F. J. Tardif, and S. B. Powles. 1994. Mechanism of cross andmultiple herbicide resistance in Alopecurus myosuroides and Lolium rig-idum. Phytoprotection 75:17–23.
Hatzios, K. K. 1997. Regulation of enzymatic systems detoxifying xeno-biotics in plants: a brief overview and directions for future research.Pages 1–5 in K. K. Hatzios, ed. Regulation of Enzymatic SystemsDetoxifying Xenobiotics in Plants. NATO ASI Series. High Technol-ogy. Volume 37. Dordrecht, The Netherlands: Kluwer.
Hatzios, K. K. 2001. Mechanism of resistance to herbicides. Pages 275–287 in R. De Prado and J. V. Jorrın, eds. Uso de Herbicidas en laAgricultura del Siglo XXI. Cordoba, Spain: Servicio PublicacionesUniversidad de Cordoba.
Heap, I. and H. LeBaron. 2001. Introduction and overview of resistance.Pages 1–22 in S. B. Powles and D. L. Shaner, eds. Herbicide Resis-tance and World Grains. Boca Raton, FL: CRC.
Himme, M. Van, J. Stryckers, and R. Bulke. 1984. Herbicide-resistant bio-types, of meadow-grass, Poa annua L. and fat hen, Chenopodium albumL. Meded. Fac. Landbouwwet. 40:151–155.
Holt, J. S., and H. M. LeBaron. 1990. Significance and distribution ofherbicide resistance. Weed Technol. 4:141–149.
Jager, G. 1983. Herbicides. Pages 332–392 in K. H. Buchel, ed. Chemistryof Pesticides. New York: J Wiley.
Jensen, K.I.N., G. R. Stephenson, and L. A. Hunt. 1977. Detoxificationof atrazine in three gramineae subfamilies. Weed Sci. 25:212–220.
Jutsum, A. R., and J. C. Graham. 1995. Managing weed resistance: therole of the agrochemical industry. Pages 783–790 in Proceedings ofthe Brighton Crop Protection Conference—Weeds. Volume 3. Farn-ham, U.K.: British Crop Protection Council.
Lee, L. J. and J. Ngim. 2000. A first report of glyphosate-resistant (Eleusinaindica) in Malaysia. Pest Manag. Sci. 56:336–339.
Letouze, A., J. Gasquez, D. Vaccara, D. Orlando, J. Leterrier, L. Roy, andE. Bouvard. 1997. Development of new reliable quick tests and stateof grass-weed herbicide resistance in France. Pages 783–790 in Pro-ceedings of the Brighton Crop Protection Conference—Weeds. Vol-ume 3. Farnham, U.K.: British Crop Protection Council.
Lopez-Martınez, N., G. Marshall, and R. De Prado. 1997. Resistance ofbarnyardgrass (Echinochloa crus-galli) to atrazine and quinclorac. Pestic.Sci. 51:171–175.
Lopez-Martınez, N., J. Gonzalez, and R. De Prado. 2001. Propanil activity,uptake and metabolism in resistant Echinochloa spp. biotypes. WeedRes. 41:187–196.
Lopez-Martınez, N., R. H. Shimabukuro, and R. De Prado. 1998. Effectof quinclorac on auxin-induced growth, transmembrane proton gra-dient and ethylene biosynthesis in Echinochloa spp. Aust. J. Plant Phy-siol. 25:851–857.
Marrs, K. A. 1996. The function and regulation of glutathione S-transfer-ases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47:127–158.
Maxwell, B. D. and A. M. Mortimer. 1994. Selection for herbicide resis-tance. Pages 1–26 in S. B. Powles and J.A.M. Holtum, eds. HerbicideResistance in Plants. Boca Raton, FL: Lewis.
Mayor, J. P. and A. Maillard. 1997. A wind bentgrass biotype resistant tothe herbicide isoproturon found in Changins. Rev. Suisse Agric. 29:9–44.
Mazur, B. J. and S. C. Falco. 1989. The development of herbicide resistantcrops. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:441–470.
McGonigl, B., S. C. Lau, and D. P. O’Keefe. 1997. Endogenous reactionsand substrate specificity of herbicide metabolizing enzymes. Pages 9–18 in K. K. Hatzios, ed. Regulation of Enzymatic Systems DetoxifyingXenobiotics in Plants. NATO ASI Series. High technology. Volume37. Dordrecht, The Netherlands: Kluwer.
Menendez, J. and R. De Prado. 1997. Diclofop-methyl cross-resistance ina chlorotoluron-resistant biotype of Alopecurus myosuroides. Pestic.Biochem. Physiol. 56:123–133.
Moss, S. R. 1990. Herbicide cross-resistance in slender foxtail (Alopecurusmyosuroides). Weed Sci. 38:492–496.
Niemans, P. and W. Pestemer. 1984. Resistance of blackgrass (Alopecurusmyosuroides) from different sites to herbicides. Nachrbl. dtsch. Pflan-zenschutzd. 36:113–118.
Preston, C., F. J. Tardif, J. T. Christopher, and S. B. Powles. 1996. Multipleresistance to dissimilar herbicide chemistries in a biotype of Loliumrigidum due to enhanced activity of several herbicide-degrading en-zymes. Pestic. Biochem. Physiol. 54:123–134.
Retzinger, E. J., Jr. and C. A. Mallory-Smith. 1997. Classification of her-bicides by site of action for weed resistance management strategies.Weed Technol. 11:384–393.
Rubin, B. 1997. Herbicide resistance outside North America and Europe:causes and significance. Pages 39–50 in R. De Prado, J. Jorrın, andL. Garcıa-Torres, eds. Weed and Crop Resistance to Herbicides. Dor-drecht, The Netherlands: Kluwer.
Ryan, G. F. 1970. Resistance of common groundsel to simazine and atra-zine. Weed Sci. 18:614–616.
Sattin, M., D. Berto, G. Zanin, and M. Tabacchi. 1999. Resistance to ALSinhibitors in weeds of rice in north-western Italy. Pages 783–790 inProceedings of the Brighton Crop Protection Conference—Weeds.Volume 3. Farnham, U.K.: British Crop Protection Council.
Sattin, M., M. A. Gasparetto, and C. Campagna. 2001. Situation andmanagement of Avena spp. ludoviciana and Phalaris paradoxa resistantto ACCase inhibitors in Italy. Pages 755–762 in Proceedings of theBrighton Crop Protection Conference—Weeds. Volume 3. Farnham,U.K.: British Crop Protection Council.
Shaner, D. L. 1995. Studies on mechanisms and genetics of resistance: theircontribution to herbicide resistance management. Pages 537–545 inProceedings of the Brighton Crop Protection Conference—Weeds.Volume 2. Farnham, U.K.: British Crop Protection Council.
Shyr, Y.Y.J., A. G. Hepburn, and J. M. Widholm. 1992. Glyphosate se-lected amplification of the 5-enolpyruvylshikimate-3-phosphate syn-thase gene in cultured carrot cells. Mol. Gen. Genet. 232:377–382.
Skipsey, M., C. J. Andrews, J. K. Towson, I. Jepson, and R. Edwards. 1997.Substrate and thiol specificity of a stress-inducible glutathione trans-ferase from soybean. FEBS Lett. 409:370–374.
Smeda, R. J., and K. C. Vaughn. 1997. Mechanism of resistance to dini-troaniline herbicides. Pages 89–99 in R. De Prado, J. Jorrın, and L.Garcıa-Torres, eds. Weed and Crop Resistance to Herbicides. Dor-drecht, The Netherlands: Kluwer.
Ware, G. W. 1994. Pesticide resistance. Pages 197–205 in G. W. Ware, ed.The Pesticide Book. Fresno, CA: Thomson.
Watanabe, N., Che, F-S., M. Iwano, S. Takayama, T. Nakano, S. Yoshida,and A. Isogai. 1998. Molecular characterization of photomixotrophictobacco cells resistant to protoporphyrinogen oxidase-inhibiting her-bicides. Plant Physiol. 118:751–758.
Yu, W. T., H. Darmency, and T. Y. Wang. 1997. Dinitroaniline herbicidecross-resistance in resistant Setaria italica lines selected from interspe-cific cross with S. viridis. Pestic. Sci. 49:277–283.
Received January 11, 2002, and approved April 4, 2003.