corticosterone regulation of ovarian follicular … levels of baseline corticosterone (cort), the...
TRANSCRIPT

1860 Journal of Lipid Research Volume 54, 2013
Copyright © 2013 by the American Society for Biochemistry and Molecular Biology, Inc.
This article is available online at http://www.jlr.org
secretion of gonadotropin-releasing hormone (GnRH) from the hypothalamus stimulates the release of the pituitary gonadotropins, luteinizing hormone (LH), and follicle-stimulating hormone (FSH), which in turn activate go-nadal development and release of sex steroids, including estradiol (E2) and testosterone. Stress is a common prob-lem disrupting breeding in either wild birds or domestic chickens ( 1 ). Glucocorticoids (GC), as the fi nal effectors of the hypothalamic-pituitary-adrenal axis, participate in the control of whole body homeostasis and the arousal of stress responses. GCs transmit information about environ-mental conditions to the HPG axis, which ultimately infl u-ences the timing of breeding.
Female kittiwakes with low baseline LH levels and ele-vated levels of baseline corticosterone (CORT), the main form of GCs in avians, were more likely to skip breeding ( 2 ). An experimental reduction of CORT release during the prelaying period was associated with an advancement of egg laying in female kittiwakes ( 3 ). In domestic chick-ens, an acute infusion of CORT resulted in a pause in lay-ing and a severely reduced ovarian weight ( 4 ). Chronic and repeated exposure to CORT during the rearing phrase suppressed reproductive performance, resulting in a delay of fi rst egg laid and a reduction of egg production ( 5 ).
In avians, the development of ovarian follicles is accom-panied by the deposition of a large amount of yolk. During a laying cycle, follicular development is matched with a supply of yolk precursors. Triacylglycerols (TG), the main
Abstract Glucocorticoids participate in the arousal of stress responses and trigger physiological adjustments that shift energy away from reproduction toward survival. Ovar-ian follicular development in avians is accompanied by the supply of yolk precursors, which are mainly synthesized in the liver. Therefore, we hypothesized energy status and he-patic lipogenesis are involved in the induction of reproduc-tive disorders by glucocorticoids in laying hens. The results show that corticosterone decreased the laying performance by suppressing follicular development in energy-defi cit state, rather than in energy-suffi cient state. In corticosterone-treated hens, the suppressed follicular development was as-sociated with the reduced availability of yolk precursor, indicated by the plasma concentration of VLDL and vitello-genin and the decreased proportion of yolk-targeted VLDL (VLDLy). Corticosterone decreased the expression of apoli-poprotein B and apolipoprotein VLDL-II in the liver. A drop in VLDL receptor content and an increase in the expression of tight junction proteins occludin and claudin1 were also observed in hierarchical follicles. The results suggest corticosterone-suppressed follicular development is energy dependent. The decreased apolipoprotein synthesis and VLDLy secretion by liver are responsible for the decreased availability of circulating yolk precursor, and the upregula-tion of occludin and claudin expression further prevents yolk deposition into oocytes. —Wang, X-J., Y. Li, Q-Q. Song, Y-Y. Guo, H-C. Jiao, Z-G. Song, and H. Lin. Corticosterone regulation of ovarian follicular development is dependent on the energy status of laying hens. J. Lipid Res. 2013. 54: 1860–1876.
Supplementary key words glucocorticoids • ovarian development • energy state • apolipoprotein synthesis • tight junction proteins
In birds, the onset of breeding involves the activation of the hypothalamic-pituitary-gonadal (HPG) axis. The
This work was supported by grants from the China Agriculture Research System (CARS-41-K14) and the Specifi c Fund for Agro-scientifi c Research in the Public Interest (201003011).
Manuscript received 29 January 2013 and in revised form 18 April 2013.
Published, JLR Papers in Press, April 18. 2013 DOI 10.1194/jlr.M036301
Corticosterone regulation of ovarian follicular development is dependent on the energy status of laying hens
Xiao-Juan Wang, 1 Yan Li, 1 Qun-Qing Song, 1 Ying-Ying Guo, Hong-Chao Jiao, Zhi-Gang Song, and Hai Lin 2
Department of Animal Science, Shandong Agricultural University , Taian, Shandong 271018, People’s Republic of China
Abbreviations: ACC1, acetyl CoA carboxylase 1; BM, body mass; CORT, corticosterone; E2, estradiol; ESR, estrogen receptor; FSH, folli-cle-stimulating hormone; FSHR, FSH receptor; GC, glucocorticoid; GnRH, gonadotropin-releasing hormone; HF, hierarchical follicle; HPG, hypothalamic-pituitary-gonadal; LH, luteinizing hormone; LHR, LH re-ceptor; LXR � , liver X receptor � ; ME, malic enzyme; MTP, microsomal triglyceride transfer protein; OVR, VLDL receptor; SCD1, stearoyl-CoA desaturase 1; SREBP-1c, sterol response element-binding protein-1c; SYF, small yellow follicle; TG, triacylglycerol; VLDLy, yolk-targeted VLDL; VTG, vitellogenin; 18SrRNA, 18S ribosomal RNA.
1 X-J. Wang, Y. Li, and Q-Q. Song contributed equally to this work. 2 To whom correspondence should be addressed. e-mail: [email protected]
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from

Corticosterone suppresses follicular growth of hen 1861
Experiment 1. At 26 weeks of age, 48 hens with similar body mass (BM) and egg production were randomly divided into eight groups of six chickens and subjected to one of the two following treatments ( Fig. 1A ): subcutaneous injection of corticosterone dissolved in corn oil at a dose of 2 mg/kg/day BM for 7 days (CORT) or sham injection of the same volume of vehicle-corn oil (Control). BM, feed intake, egg number, and egg mass were re-corded daily for each group. Hen-day egg production was calcu-lated. Eggs laid were counted daily and laying rate was expressed as a percentage of total hen amounts for each group. Before the sampling day (day 7), half of the hens from each group were randomly selected with or without 12 h overnight feed with-drawal. On day 7, eight chickens from each treatment were se-lected for a blood sample. The sample was obtained within 30 s with a heparinized syringe from a wing vein and collected in iced tubes. Plasma was obtained after centrifugation at 400 g for 10 min at 4°C and was stored at � 20°C for further analysis. Immediately after blood sampling, the hens were sacrifi ced by cervical disloca-tion and exsanguination ( 17 ). Thereafter, the liver, ovaries and follicles, and abdominal fat were harvested and weighed. The num-ber and weight of hierarchical follicles (HF, >12 mm) and small yellow follicles (SYF, 5–12 mm) were recorded. The liver and the follicular membranes of HF were sampled and washed with ice-cold sterilized saline, cooled down in liquid nitrogen, and stored at � 80°C for further analysis.
Experiment 2. Forty-eight 30-week-old hens with similar BM and egg production were randomly divided into eight groups of six chickens and given ad libitum access to either a normal diet (2,654 kcal/kg) or a high-energy diet (2,917 kcal/kg). After a three-week dietary treatment period, half of the hens in each
components of yolk lipid ( 6 ), are mainly synthesized in the liver and secreted in the form of yolk-targeted very-low density lipoprotein (VLDLy). An elevated circulating VLDLy concentration is coincident with rapid yolk devel-opment and matched variation in total yolky follicle mass in European starlings ( 7 ). VLDLy has two associated apoli-poproteins, apolipoprotein B (apoB) and apolipoprotein VLDL-II (apoVLDL-II), which are involved in the assembly of TG-rich lipoprotein particles. Avian apoB is a compo-nent of specialized VLDL particles that are produced by the liver in response to estrogen ( 8 ). apoVLDL-II mRNA was specifi cally expressed in the liver and upregulated af-ter laying ( 9, 10 ).
When domestic laying hens were infused with CORT, food consumption remained high and body weight re-mained unchanged, although the liver weight doubled ( 4 ). In our previous work on immature chickens, GCs stimu-lated energy intake ( 11 ) and enhanced hepatic lipogenesis and fat deposition in adipose tissues, indicating the redis-tribution of energy stores in CORT-challenged chickens ( 12–14 ). GCs trigger physiological and behavioral adjust-ments that shift energy investment away from reproduction and toward survival ( 15 ). CORT secretion during the pre-laying period mediates the timing of breeding in kittiwakes, possibly through the dynamics of energy reserves ( 3 ). High-level CORT exposure does not affect the timing of breed-ing in Florida scrub-jays under normal feeding conditions ( 16 ). Hence, we hypothesized that energy status and he-patic lipogenesis are involved in the physiological responses of the reproductive system to stress perturbations.
In the present study, two experiments were conducted to evaluate the effect of GCs on ovarian function in laying hens ( Gallus gallus domesticus ). In Experiment 1 and Exper-iment 2, the effects of CORT on laying performance, folli-cular development and hepatic lipogenesis were evaluated for two feeding states (fasting and feeding) and for diets with two different energy levels, respectively. The present study suggests that exogenous GCs perturbed the repro-duction of laying hens in an energy-dependent manner.
METHODS
Ethics statement All animal experiments were reviewed and approved by the
Institutional Animal Care and Use Committee of Shandong Agri-cultural University and performed in accordance with the “Guide-lines for Experimental Animals” of the Ministry of Science and Technology (Beijing, China). Animal suffering was minimized as much as possible.
Birds and treatments Hy-Line Brown laying hens were housed in an environmentally
controlled room. The hens were fed a layer diet based on a corn-soybean meal diet (containing 16% crude protein, 2,702 kcal/kg metabolizable energy, 3.6% calcium, and 0.4% available phospho-rus). The lighting regime was 16 h light (from 05:00 to 21:00) and 8 h dark. All birds had free access to feed and water during the rearing period.
Fig. 1. Schematic diagrams of design of Experiment 1 (A) and Experiment 2 (B).
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from

1862 Journal of Lipid Research Volume 54, 2013
18 h at 14°C in a Sorvall 45.6 TFT rotor (Sorvall-DuPont, Wilming-ton, DE). The VLDL diameters were determined optically by dy-namic light scattering using a Wyatt DynaPro Nanostar instrument (Santa Barbara, CA) at 22°C, according to a modifi cation of the method from Walzem et al. ( 24 ). Centrifugally isolated VLDLs were suspended as a 1:30 dilution in a NaCl solution (density = 1.0063 g/ml) and placed into the sample well. The system soft-ware and 3 mW, � = 658 nm laser beam were activated. Light scattering was recorded from the lipoprotein particles for 4 min. The data were analyzed using Dynamics 6.0 software, which auto-matically calculates the particle size distribution according to the characteristics of the scattered light within the measurement re-gion. VLDL particle diameters of 25–44 nm were positively associ-ated with egg production ( 25, 26 ).
Plasma variables. The concentration of glucose (no. F006), nonesterifi ed fatty acids (NEFA, no. A042), and TG (no. F001) were measured spectrophotometrically using commercial diag-nostic kits (Jiancheng Bioengineering Institute, Nanjing, China). The concentrations of VLDL were determined as previously de-scribed ( 27 ).
Plasma insulin was measured by radioimmunoassay with a guinea pig anti-porcine insulin serum (3 V Biochemical Engineer-ing Co., Weifang, China). A large cross-reaction was observed between chicken insulin and this anti-serum (porcine) ( 28 ). The insulin in this study is referred to as immunoreactive insulin. The sensitivity of the assay was 6.9 pmol/l, and all samples were in-cluded in the same assay to avoid interassay variability. The in-traassay coeffi cient of variation was 6.9%. FSH, LH, and E2 levels were measured by radioimmunoassay with a rabbit anti-human serum (Jiuding Bioproduction Co., Tianjin, China). The sensi-tivities of the assays for FSH, LH, and E2 were 0.9 IU/l, 1.0 IU/l, and 2.1 ng/l, respectively. All the plasma samples were included in the same assay to avoid interassay variability. The intraassay coeffi cients of variation were 5.5, 5.4, and 7.7% for FSH, LH, and E2, respectively.
FAS and ME activities and OVR protein content. The activi-ties of fatty acid synthase (FAS) and malic enzyme (ME) in the liver were measured. The liver samples were homogenized in ice-cold 0.25 mol/l sucrose, 1 mmol/l dithiothreitol, and 1 mmol/l EDTA at pH 7.4. The cytosolic fractions were obtained by cen-trifugation at 100,000 g for 1 h at 48°C and used for the enzy-matic assays. The FAS activity was measured according to the method of Halestrap and Denton ( 29 ). One unit of FAS is de-fi ned as 1 nmol reduced NADPH consumed by 1 mg protein in 1 min. ME activity was determined by a method modifi ed from Hsu and Lardy ( 30 ). One unit of ME is defi ned as 1 � mol NADPH produced by 1 mg protein in 1 min. The VLDL/vitellogenin re-ceptor (OVR) protein content in the follicular membranes of HF was measured using an ELISA method (Cusabio Biotech Co.). The sensitivity of the assay for OVR protein content was 3.9 pg/ml. All the samples were included in the same assay to avoid interas-say variability. The intraassay coeffi cients of variation were less than 8%.
mRNA expression. Gene expression was measured using real-time RT-PCR. Briefl y, total RNA was extracted from the liver tissue and the HF membranes using Trizol (Invitrogen, San Diego, CA). The quantity and quality of the isolated RNA were determined us-ing a biophotometer (Eppendorf, Hamburg, Germany) and aga-rose gel electrophoresis, respectively. Reverse transcription was performed in RT reactions (10 � l) consisting of 500 ng total RNA, 5 mmol/l MgCl 2 , 1 � l RT buffer, 1 mmol/l dNTP, 2.5 U AMV, 0.7 nmol/l oligo d(T), and 10 U RNase inhibitor (TaKaRa, Dalian, China). mRNA was amplifi ed using cDNA as template in a 20 � l
group were randomly assigned to either CORT administration or sham treatment for 7 days as in Experiment 1 ( Fig. 1B ). At the end of the experiment, eight chickens from each treatment were selected and sampled after a 12 h feed withdrawal. The measure-ments were performed in the same way as in Experiment 1.
Measurements Histological analysis. The degrees of hepatic steatosis were
observed with hematoxylin and eosin staining as described in Wang et al. ( 18 ). Briefl y, the liver tissue was fi xed conventionally in 4% formaldehyde, dehydrated, and embedded in paraffi n. De-paraffi nated sections with a thickness of 4 � m were stained with Harris hematoxylin and eosin (Sigma-Aldrich, St. Louis, MO). Sections were examined under an Olympus CX-41 phase contrast microscope (Olympus, Tokyo, Japan).
The accumulation of cytoplasmic lipid droplets in liver was vi-sualized by Oil Red O staining according to the protocol of Lillie and Fullmer ( 19 ). Briefl y, tissues were immediately frozen in liq-uid nitrogen and cut in a Leica CM-1850 cryostat microtome (Leica, Wetzlar, Germany). Then 18 � m thick sections were fi xed in 4% formaldehyde for 60 min and stained with fi ltered 0.5% Oil Red O (Sigma-Aldrich, St Louis, MO), which was made by dissolving in isopropyl alcohol for 15 min at room temperature. Sections were photographed using an Olympus CX-41 phase con-trast microscope (Olympus, Tokyo, Japan).
Hepatic TG content. Total lipids were extracted according to the method used by Folch et al. ( 20 ). A 100 mg liver sample was homogenized with 2 ml of chloroform-methanol (2:1, v/v). After a centrifugation at 400 g for 10 min, the supernatant was obtained, and then saline (4:1, v/v) was added and vortexed. The organic phase was isolated by a centrifugation at 400 g for 10 min, then transferred to a new tube and evaporated under a stream of nitro-gen. The extracted lipids were then dissolved with 1 ml isopropyl alcohol, and the levels of hepatic TG content were determined spectrophotometrically using commercial kits (GPO-PAP) pur-chased from the Jiancheng Bioengineering Institute (Nanjing, China).
In vivo hepatic VLDL secretion. VLDL secretion was mea-sured according to Jong et al. ( 21 ). The laying hens that under-went Experiment 1 were injected intravenously with 15% (w/v) Triton WR1339 (Sigma, St Louis, MO) (500 mg/kg BM) dis-solved in 0.9% NaCl. Plasma VLDL clearance is virtually com-pletely inhibited under these circumstances ( 22 ). Immediately before Triton administration and 45 and 90 min after Triton ad-ministration, blood samples were drawn, and TG content was measured using commercially available colorimetric assays. The TG accumulation in plasma was linear during this time period. Because the vast majority of TG is associated with the apoB-con-taining VLDL particles, we refer to these TG measurements as VLDL-TG. Hepatic VLDL-TG secretion rate was calculated from the slope of the line and expressed as � mol/h per kg BM.
Determination of VLDL particle size distribution. Plasma VLDL was isolated by centrifugation to determine particle size distributions by dynamic laser light scattering as described below. Lipoproteins were isolated from 1 ml plasma using sequential density gradient centrifugation ( 23 ). The plasma activity of lecithin:cholesterol acyltransferase was inhibited by the addition of 15 � l dithionitrobenzoic acid/ml plasma. Density solutions were prepared by adding increasing amounts of NaBr to a basal NaCl solution (density = 1.0063 g/ml) containing 0.01% EDTA and 50 kU/l each of streptomycin and penicillin. VLDL was iso-lated as the plasma fraction of density <1.019 g/ml by aspiration with a narrow-bore pipette after centrifugation at 148,600 g for
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
Corticosterone suppresses follicular growth of hen 1863
the BCA assay kit (Beyotime, Jiangsu, China). Samples were boiled at 100°C for 5 min in 5× sample buffer. Protein extracts (80 � g) were electrophoresed in 7.5–10% SDS polyacrylamide gels (Bio-Rad, Richmond, CA) according to the Laemmli method ( 32 ). Separated proteins were then transferred onto a nitrocellulose membrane at 100 V for 1 h at 4°C in tris-glycine buffer contain-ing 20% methanol. Membranes were blocked for 1 h and im-munoblotted overnight at 4°C with the following primary antibodies: apoB100 antibody (1:500, Sigma, St. Louis, MO) for liver samples and occludin antibody (1:100, Invitrogen, Cama-rillo, CA) and claudin1 antibody (1:125, Invitrogen, Camarillo, CA) for HF membrane samples. Protein was detected using goat anti-rabbit IgG (H+L)-HRP conjugated secondary antibody (1:2000, Bio-Rad, Richmond, CA) with enhanced chemilumi-nescence plus western blot detection reagents (Beyotime, Ji-angsu, China). � -actin was used as an internal control (Beyotime, Jiangsu, China). Western blots were developed and quantifi ed using BioSpectrum 810 with VisionWorksLS 7.1 software (UVP LLC, California).
Statistical analysis All the data were subjected to one-way ANOVA analysis with
the Statistical Analysis Systems statistical software package (Version 8e, SAS Institute, Cary, NC), and the main effect of CORT treatment was evaluated. Multiple comparisons between means were conducted using the Tukey’s honestly signifi cant difference test. Means were considered signifi cantly different at P < 0.05.
PCR reaction containing 0.2 µmol/l of each primer (Sangon, Shanghai, China) and SYBR green master mix (TaKaRa, Dalian, China). Real-time PCR was performed at 95°C for 10 s of predena-turation, followed by 40 cycles, with each cycle consisting of denatur-ation at 95°C for 5 s and annealing and extension at 60°C for 40 s. mRNA expression was quantifi ed according to the compara-tive CT method (2 � � � CT ) ( 31 ). The mRNA levels of target genes were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA, 18S rRNA (18SrRNA), and � -actin ( � CT). The � CT was calibrated against the average of the control chickens. The number of target molecules relative to the control was calcu-lated using 2 � � � C . Therefore, all gene transcription results are re-ported as the n-fold difference relative to the control. The primer sequences are listed in Table 1 . The PCR products were verifi ed by electrophoresis on a 0.8% agarose gel and by DNA sequencing. Standard curves were generated using pooled cDNA from the sam-ples that were assayed. All samples were run in duplicate, and primers were designed to span an intron to avoid contamination from genomic DNA.
Western blotting analysis. Samples from the liver tissue and the HF membranes were homogenized on ice in radioimmuno-precipitation assay buffer (50 mmol/l Tris-HCl at pH 7.4, 1% NP-40, 0.25% sodium deoxycholate, 150 mmol/l NaCl, 1 mmol/l EDTA, 1 mmol/l phenylmethylsulfonyl fl uoride, 1 � g/ml aproti-nin, 1 � g/ml leupeptin, 1 � g/ml pepstatin, 1 mmol/l sodium ortho-vanadate, 1 mmol/l sodium fl uoride) and centrifuged at 12,000 g for 5 min at 4°C. The protein concentration was determined using
TABLE 1. Gene-specifi c primers of related genes
Gene Name GenBank Number Primers Sequences(5 ′ → 3 ′ ) Product Size (bp)
18SrRNA AF173612 Forward: ataacgaacgagactctggca 136Reverse: cggacatctaagggcatcaca
ACC1 NM_205505 Forward: aatggcagctttggaggtgt 136Reverse: tctgtttgggtgggaggtg
apoB100 M18421 Forward: cacgcctcacagaccaagta 407Reverse: ccagtcaaacggcacatcta
apoVLDL-II NM-205483 Forward: agggctgaactggtaccaacaaac 140Reverse: ggatgaccagccagtcacga
� -actin AW05994 Forward: tgcgtgacatcaaggagaag 300Reverse: tgccagggtacattgtggta
Claudin1 AY750897 Forward: tcctgggtctggttggtgtg 199Reverse: acgggtgtgaaagggtcataga
ESR NM_205183 Forward: gtctggtcttgtgagggctg 435Reverse: gcttcgttgaatggtctattggg
FAS NM-205155 Forward: cgcagtttgttgatggtgag 179Reverse: tccttggtgttcgtgacg
FSHR NM_205079 Forward: caccaatgccacagaactgagat 290Reverse: gcaccttatggacgacgggt
GAPDH NM_204305 Forward: ctacacacggacacttcaag 244Reverse: acaaacatgggggcatcag
LHR NM_204936 Forward: agccttcctgctttgtctg 359Reverse: atcgttgtgtatccgcctg
LPL NM_205282 Forward: cagtgcaacttcaaccatacca 150Reverse: aaccagccagtccacaacaa
LXR � AJ507202 Forward: gtccctgaccctaataaccgc 185Reverse: gtctccaacaacatcacctctatg
ME NM204303 Forward: tgccagcattacggtttagc 175Reverse: ccattccataacagccaaggtc
MTP XM_420662 Forward: ttcacagtaccccttcctagtctgt 138Reverse: ccttccaacatttctgctttcc
Occludin NM_205128 Forward: caccgaggacagccctcaata 141Reverse: accatgcgcttgatgtggaa
OVR NM_205229 Forward: actgtgaggatgggtctgacga 101Reverse: tgggatacactgggttgactgag
SCD1 NM_204890 Forward: accacagccttcatcgacct 104Reverse: ccatctccagtccgcatttt
SREBP-1c AY029224 Forward: gccctctgtgcctttgtcttc 130Reverse: actcagccatgatgcttcttcc
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from

1864 Journal of Lipid Research Volume 54, 2013
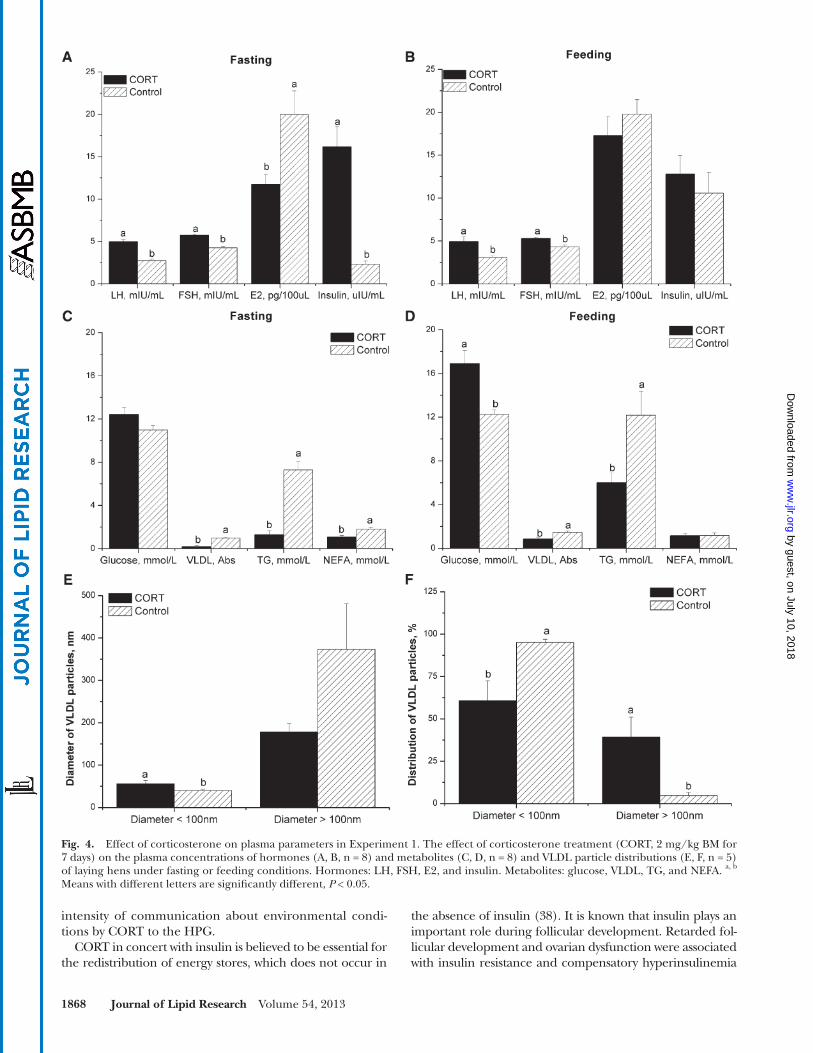
proportion of VLDL particles with a large diameter (>100 nm) was increased by CORT ( P < 0.05), while their di-ameter was not signifi cantly changed ( P > 0.05, Fig. 4E, F ).
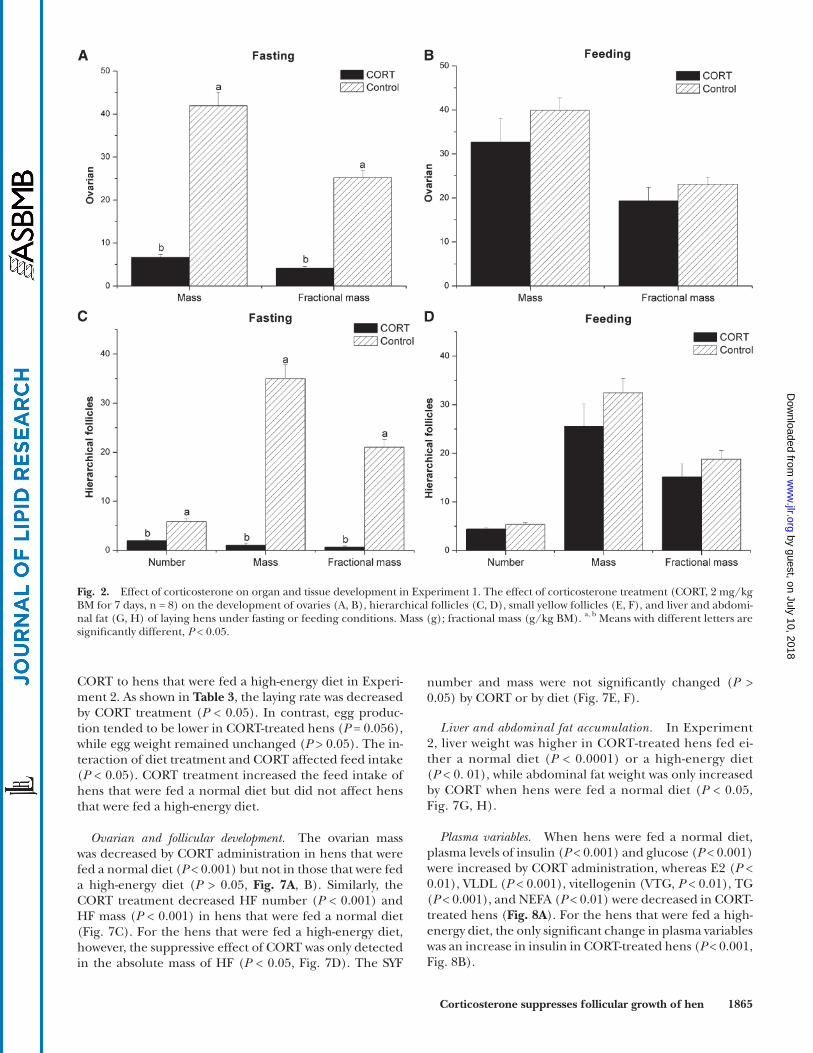
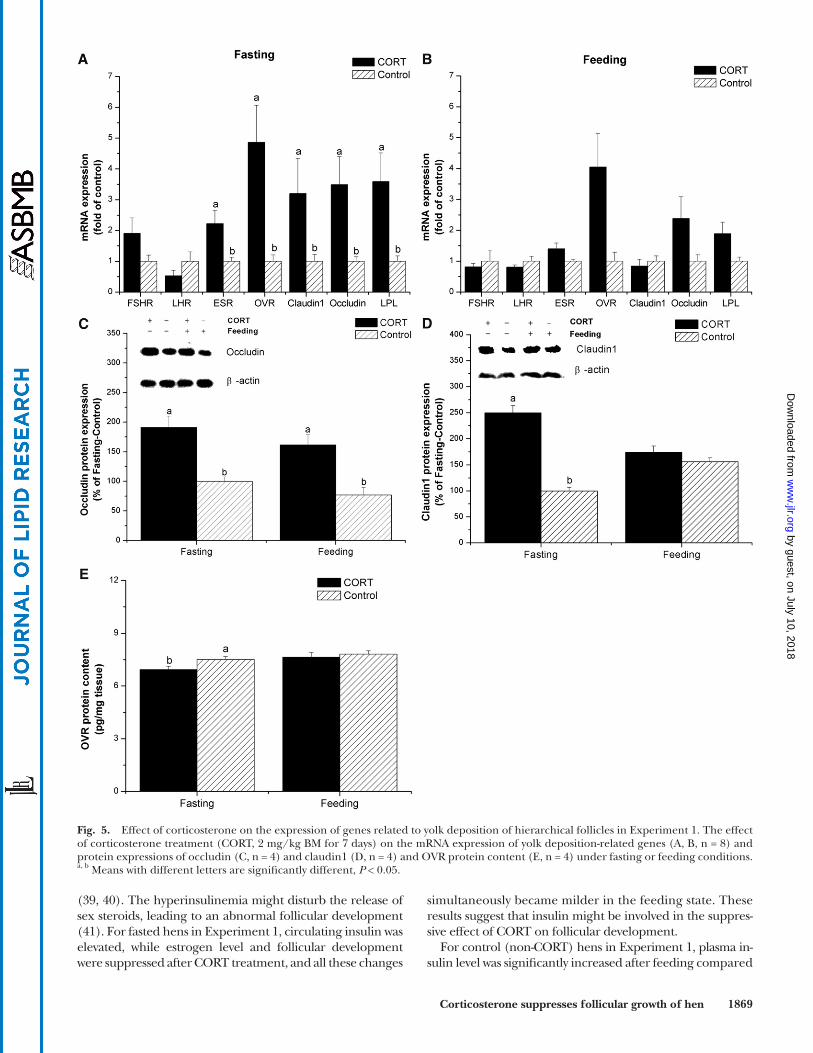
mRNA and protein expression of genes related to yolk deposition in HF membranes. In the fasting state, there were remark-able increases in the mRNA levels of estrogen receptor (ESR, P < 0.01), OVR ( P < 0.01), claudin1 ( P < 0.05), occlu-din ( P < 0.01), and lipoprotein lipase (LPL, P < 0.001) in CORT-treated hens compared with control hens, while no signifi cant changes were observed in the mRNA levels of FSH receptor (FSHR) and LH receptor (LHR) ( P > 0.05, Fig. 5A ). In the feeding state, however, the expression levels of all these genes were not signifi cantly infl uenced by CORT treatment ( P > 0.05, Fig. 5B ). The protein expression of occludin was depressed by CORT ( P < 0.01) independent of the feeding state ( Fig. 5C ). CORT-treated fasting hens exhibited higher claudin1 protein expression ( P < 0.001, Fig. 5D ) and lower OVR protein content ( P < 0.05, Fig. 5E ).
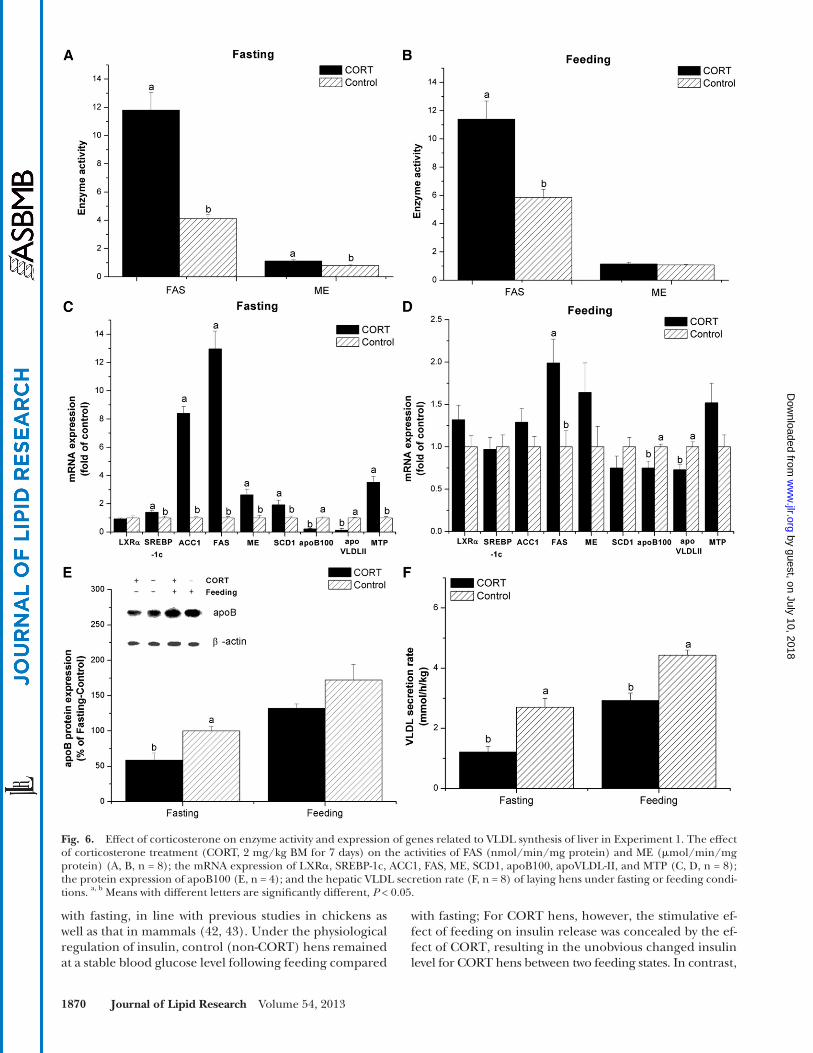
mRNA and protein expression and enzymatic activity of genes re-lated to VLDL synthesis in liver. FAS activity was higher in CORT-treated hens than in control hens in both the feeding ( P < 0.0001) and fasting states ( P < 0.01). In contrast, ME ac-tivity was only increased by CORT in the fasting state ( P < 0.01, Fig. 6A , B ). The mRNA levels of sterol response ele-ment-binding protein-1c (SREBP-1c, P < 0.05), acetyl CoA carboxylase 1 (ACC1, P < 0.001), ME ( P < 0.001), stearoyl-CoA desaturase 1 (SCD1, P < 0.05), and microsomal triglycer-ide transfer protein (MTP, P < 0.001) were upregulated by CORT in the fasting state, but they were not signifi cantly changed in the feeding state ( P > 0.05). The gene expression of liver X receptor � (LXR � ) was not altered by CORT in either the feeding or fasting state ( P > 0.05). In contrast, the mRNA levels of apoB100 ( P < 0.001) and apoVLDL-II ( P < 0.001) were downregulated, and FAS was upregulated ( P < 0.001) by CORT treatment in both the feeding and fasting states ( Fig. 6C, D ). In the fasting state, the protein expression of apoB100 was decreased in CORT-treated hens compared with control hens ( P < 0.05), but no effect was observed in the feeding state ( P > 0.05, Fig. 6E ). The hepatic VLDL secretion rate was reduced by CORT in both the fasting and feeding states ( P < 0.05, Fig. 6F ).
Experiment 2
Laying performance. We further investigated the effect of energy state on follicular development by administering
RESULTS
Experiment 1 Laying performance. We investigated the effect of CORT
treatment on follicular development and the involvement of energy state in Experiment 1. Table 2 shows that CORT decreased BM ( P < 0.001), laying rate ( P < 0.05), egg pro-duction ( P < 0.05), and egg weight ( P < 0.05). Feed intake, however, was not affected by CORT administration ( P > 0.05).
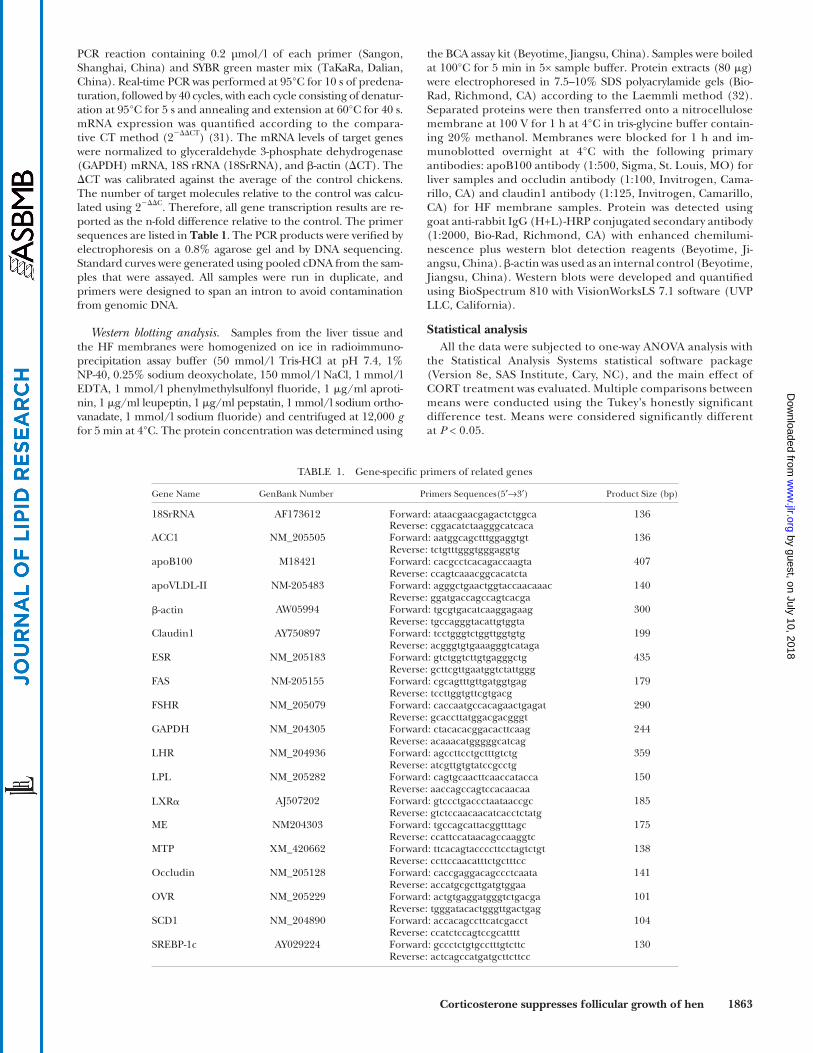
Ovarian and follicular development. In the fasting state, CORT administration decreased ovarian mass ( P < 0.001), HF number and mass ( P < 0.001), and SYF number and mass ( P < 0.05, Fig. 2A , C, E ). In contrast, none of these variables were altered in the feeding state ( P > 0.05, Fig. 2B, D, F ).
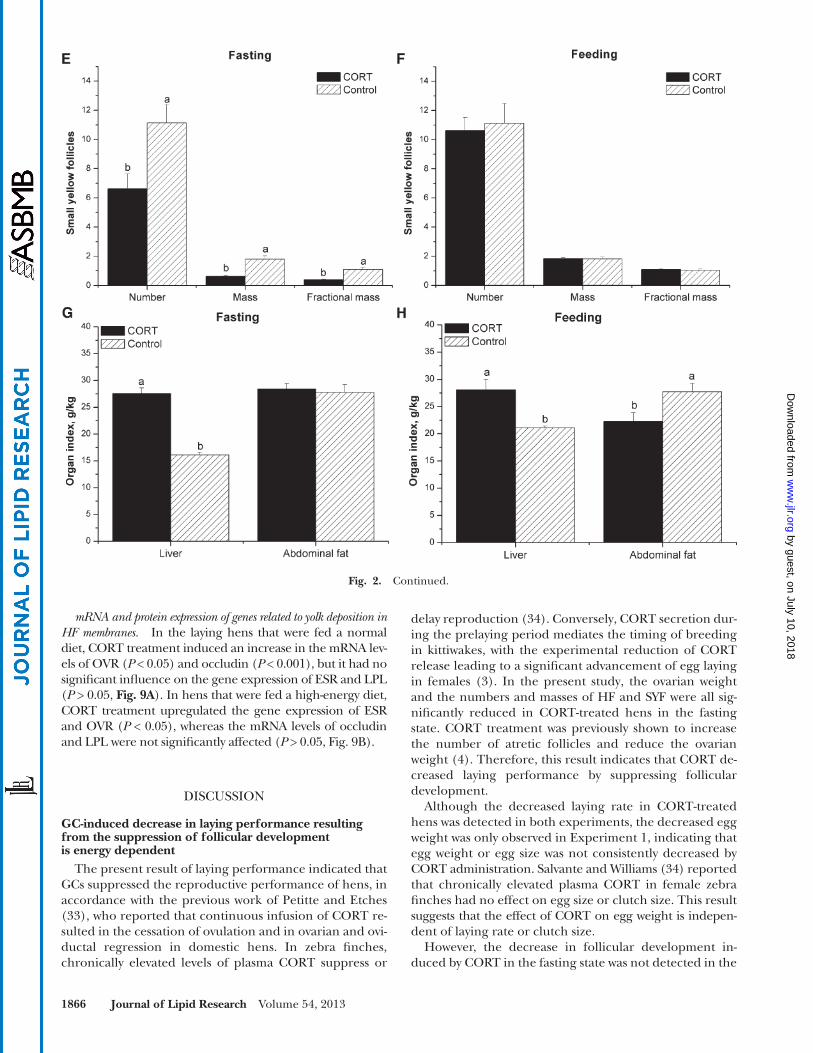
Liver and abdominal fat accumulation. The liver fat ac-cumulation was measured to evaluate the hepatic lipogen-esis. CORT-treated hens had a higher liver mass compared with control hens in either the fasting ( P < 0.0001) or feed-ing state ( P < 0.01). In contrast, CORT only decreased ab-dominal fat weight in the feeding state ( P < 0.05, Fig. 2G, H ). CORT treatment increased lipid deposition in the liver, as indicated by both histological analysis ( Fig. 3A –D ) and the hepatic TG content ( P < 0.01, Fig. 3E ).
Plasma variables. We checked the availability of yolk precursors by testing the circulating VLDL content and diameter of VLDL particles. In the fasting state, CORT ad-ministration signifi cantly increased the plasma concentra-tions of FSH ( P < 0.001), LH ( P < 0.01), and insulin ( P < 0.001), while the E2 level tended to decrease ( P = 0.075, Fig. 4A ). In the feeding state, CORT treatment also in-creased LH ( P < 0.05) and FSH concentrations ( P < 0.01). However, CORT administration did not signifi cantly affect E2 and insulin levels ( P > 0.05, Fig. 4B ). Plasma concentra-tions of glucose were signifi cantly increased in the feeding state ( P < 0.01) but not in the fasting state ( P > 0.05, Fig. 4C, D ). The reverse is true for the NEFA level, which was only decreased by CORT in the fasting state ( P < 0.01). Plasma concentrations of TG and VLDL were decreased by CORT administration in both the fasting ( P < 0.001) and feeding states ( P < 0.05). For small diameter VLDL par-ticles (< 100 nm), CORT treatment signifi cantly increased their diameter ( P < 0.05) and decreased their proportion ( P < 0.05) compared with control hens. In contrast, the
TABLE 2. Effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days) on laying performances of hens (Experiment 1)
Item CORT Control Probability
BM, kg 1.65 ± 0.02 b 1.72 ± 0.01 a < 0.001Feed intake, kcal/day 288.3 ± 13.8 295.1 ± 30.3 NSLaying rate, % 53.6 ± 12.8 b 95.8 ± 2.0 a 0.017Egg production, g/day/hen 28.6 ± 6.8 b 53.7 ± 0.8 a 0.011Egg weight, g 53.5 ± 0.3 b 56.1 ± 0.9 a 0.034
Mean ± SE (n = 4). NS, not signifi cant. a , b Means within the same item with different superscript are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
Corticosterone suppresses follicular growth of hen 1865
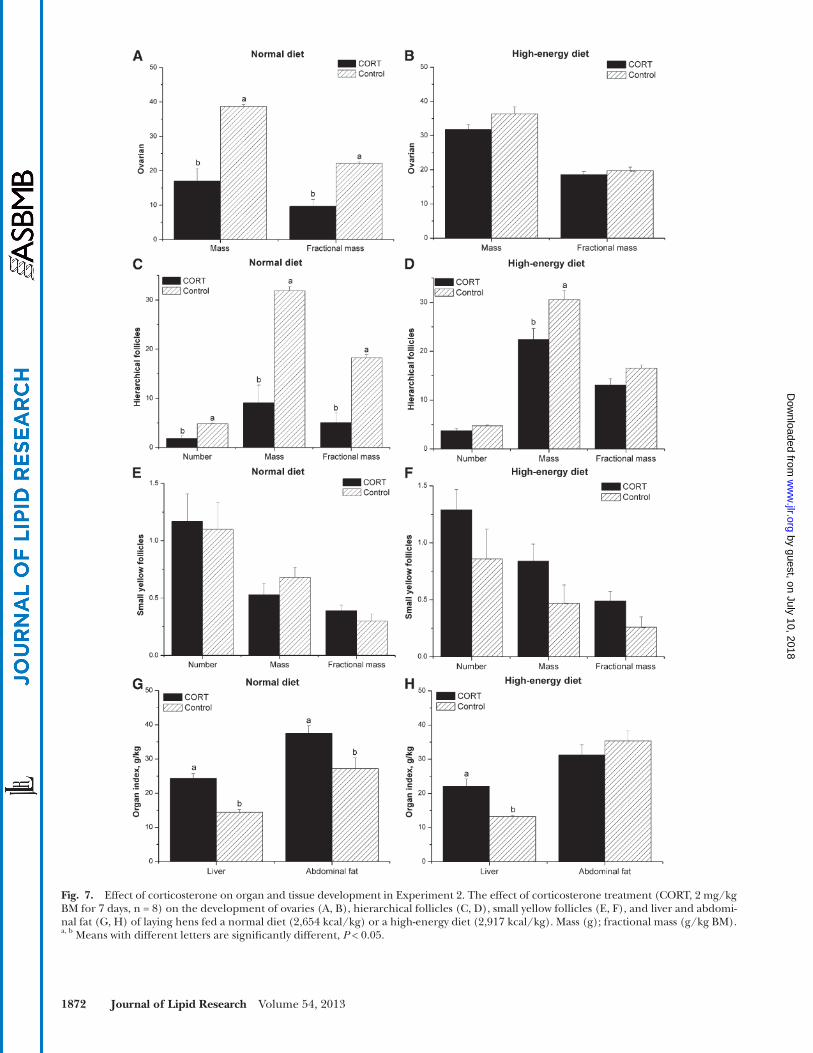
number and mass were not signifi cantly changed ( P > 0.05) by CORT or by diet ( Fig. 7E, F ).
Liver and abdominal fat accumulation. In Experiment 2, liver weight was higher in CORT-treated hens fed ei-ther a normal diet ( P < 0.0001) or a high-energy diet ( P < 0. 01), while abdominal fat weight was only increased by CORT when hens were fed a normal diet ( P < 0.05, Fig. 7G, H ).
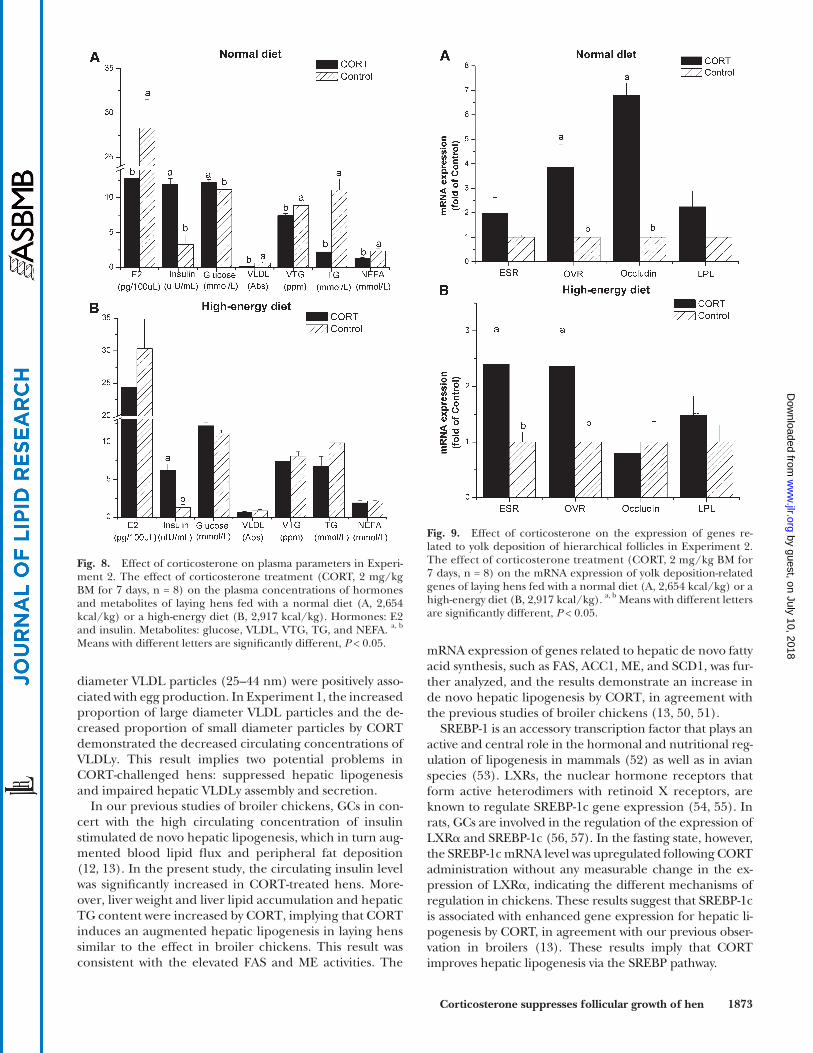
Plasma variables. When hens were fed a normal diet, plasma levels of insulin ( P < 0.001) and glucose ( P < 0.001) were increased by CORT administration, whereas E2 ( P < 0.01), VLDL ( P < 0.001), vitellogenin (VTG, P < 0.01), TG ( P < 0.001), and NEFA ( P < 0.01) were decreased in CORT-treated hens ( Fig. 8A ). For the hens that were fed a high-energy diet, the only signifi cant change in plasma variables was an increase in insulin in CORT-treated hens ( P < 0.001, Fig. 8B ).
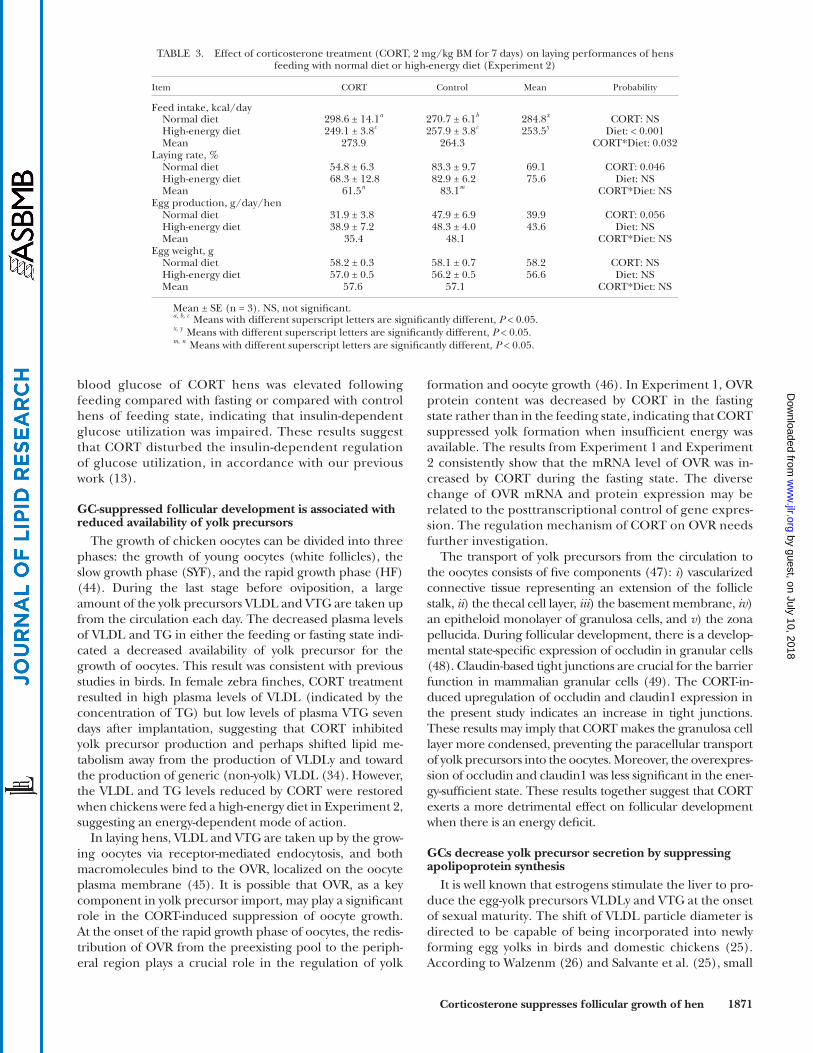
CORT to hens that were fed a high-energy diet in Experi-ment 2. As shown in Table 3 , the laying rate was decreased by CORT treatment ( P < 0.05). In contrast, egg produc-tion tended to be lower in CORT-treated hens ( P = 0.056), while egg weight remained unchanged ( P > 0.05). The in-teraction of diet treatment and CORT affected feed intake ( P < 0.05). CORT treatment increased the feed intake of hens that were fed a normal diet but did not affect hens that were fed a high-energy diet.
Ovarian and follicular development. The ovarian mass was decreased by CORT administration in hens that were fed a normal diet ( P < 0.001) but not in those that were fed a high-energy diet ( P > 0.05, Fig. 7A , B ). Similarly, the CORT treatment decreased HF number ( P < 0.001) and HF mass ( P < 0.001) in hens that were fed a normal diet ( Fig. 7C ). For the hens that were fed a high-energy diet, however, the suppressive effect of CORT was only detected in the absolute mass of HF ( P < 0.05, Fig. 7D ). The SYF
Fig. 2. Effect of corticosterone on organ and tissue development in Experiment 1. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days, n = 8) on the development of ovaries (A, B), hierarchical follicles (C, D), small yellow follicles (E, F), and liver and abdomi-nal fat (G, H) of laying hens under fasting or feeding conditions. Mass (g); fractional mass (g/kg BM). a, b Means with different letters are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
1866 Journal of Lipid Research Volume 54, 2013
mRNA and protein expression of genes related to yolk deposition in HF membranes. In the laying hens that were fed a normal diet, CORT treatment induced an increase in the mRNA lev-els of OVR ( P < 0.05) and occludin ( P < 0.001), but it had no signifi cant infl uence on the gene expression of ESR and LPL ( P > 0.05, Fig. 9A ). In hens that were fed a high-energy diet, CORT treatment upregulated the gene expression of ESR and OVR ( P < 0.05), whereas the mRNA levels of occludin and LPL were not signifi cantly affected ( P > 0.05, Fig. 9B ).
DISCUSSION
GC-induced decrease in laying performance resulting from the suppression of follicular development is energy dependent
The present result of laying performance indicated that GCs suppressed the reproductive performance of hens, in accordance with the previous work of Petitte and Etches ( 33 ), who reported that continuous infusion of CORT re-sulted in the cessation of ovulation and in ovarian and ovi-ductal regression in domestic hens. In zebra fi nches, chronically elevated levels of plasma CORT suppress or
delay reproduction ( 34 ). Conversely, CORT secretion dur-ing the prelaying period mediates the timing of breeding in kittiwakes, with the experimental reduction of CORT release leading to a signifi cant advancement of egg laying in females ( 3 ). In the present study, the ovarian weight and the numbers and masses of HF and SYF were all sig-nifi cantly reduced in CORT-treated hens in the fasting state. CORT treatment was previously shown to increase the number of atretic follicles and reduce the ovarian weight ( 4 ). Therefore, this result indicates that CORT de-creased laying performance by suppressing follicular development.
Although the decreased laying rate in CORT-treated hens was detected in both experiments, the decreased egg weight was only observed in Experiment 1, indicating that egg weight or egg size was not consistently decreased by CORT administration. Salvante and Williams ( 34 ) reported that chronically elevated plasma CORT in female zebra fi nches had no effect on egg size or clutch size. This result suggests that the effect of CORT on egg weight is indepen-dent of laying rate or clutch size.
However, the decrease in follicular development in-duced by CORT in the fasting state was not detected in the
Fig. 2. Continued.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
Corticosterone suppresses follicular growth of hen 1867
feeding state. In mammals, many researchers have demon-strated the suppressive effect of fasting or food depriva-tion. A fi ve-day fast during the luteal phase reduced the total number of follicles, causing a delay in the develop-ment of ovarian follicles in ewes ( 35 ). Food deprivation also led to the death of oocytes and attenuated glucose consumption by cumulus-oocyte complexes ( 36 ). In con-trast, the blockage of mouse ovulation induced by food deprivation could be reversed by glucose, oil, or short-term feeding ( 36 ). We found that the decrease in ovarian and HF development in CORT-treated hens fed the con-trol diet was restored by the high-energy diet. Similarly, when supplemented with food, exposure to high circulating
CORT levels alone did not affect the timing of breeding in Florida scrub-jays ( 16 ). Moreover, as CORT release was ex-perimentally reduced during the prelaying period, the in-crease in mass in female kittiwakes was suggested as a mechanism by which CORT mediates the timing of breed-ing in the seabird ( 3 ). In mammals, a short-term intermit-tent nutritional stimulus (high-energy diet) in the luteal phase increased the total number of ovulatory follicles and the ovulation rate in goats ( 37 ). GCs shift energy away from reproduction toward survival ( 15 ). These results to-gether demonstrated that the effects of GC on follicular development are energy-state dependent and that the un-limited availability of food or energy intake attenuates the
Fig. 3. Effect of corticosterone on lipid accumulation of livers in Experiment 1. (A, B) Representative liver sections stained with hematoxylin and eosin (400×). Enlarged lipid-containing vacuoles in degenerating hepatocytes are more evident in hens treated with corticosterone (CORT, 2 mg/kg BM for 7 days, n = 8). (C, D) Oil Red O staining of liver sections for cytoplasmic lipid droplets (indi-cated by arrows, 400×) showing effect of corticosterone treatment on lipid accumulation in liver. (E) The effect of corticosterone treatment on the TG content in the livers of hens. a, b Means with different letters are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
1868 Journal of Lipid Research Volume 54, 2013
the absence of insulin ( 38 ). It is known that insulin plays an important role during follicular development. Retarded fol-licular development and ovarian dysfunction were associated with insulin resistance and compensatory hyperinsulinemia
intensity of communication about environmental condi-tions by CORT to the HPG.
CORT in concert with insulin is believed to be essential for the redistribution of energy stores, which does not occur in
Fig. 4. Effect of corticosterone on plasma parameters in Experiment 1. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days) on the plasma concentrations of hormones (A, B, n = 8) and metabolites (C, D, n = 8) and VLDL particle distributions (E, F, n = 5) of laying hens under fasting or feeding conditions. Hormones: LH, FSH, E2, and insulin. Metabolites: glucose, VLDL, TG, and NEFA. a, b Means with different letters are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
Corticosterone suppresses follicular growth of hen 1869
Fig. 5. Effect of corticosterone on the expression of genes related to yolk deposition of hierarchical follicles in Experiment 1. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days) on the mRNA expression of yolk deposition-related genes (A, B, n = 8) and protein expressions of occludin (C, n = 4) and claudin1 (D, n = 4) and OVR protein content (E, n = 4) under fasting or feeding conditions. a, b Means with different letters are signifi cantly different, P < 0.05.
( 39, 40 ). The hyperinsulinemia might disturb the release of sex steroids, leading to an abnormal follicular development ( 41 ). For fasted hens in Experiment 1, circulating insulin was elevated, while estrogen level and follicular development were suppressed after CORT treatment, and all these changes
simultaneously became milder in the feeding state. These results suggest that insulin might be involved in the suppres-sive effect of CORT on follicular development.
For control (non-CORT) hens in Experiment 1, plas ma in-sulin level was signifi cantly increased after feeding compared
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
1870 Journal of Lipid Research Volume 54, 2013
with fasting; For CORT hens, however, the stimulative ef-fect of feeding on insulin release was concealed by the ef-fect of CORT, resulting in the unobvious changed insulin level for CORT hens between two feeding states. In contrast,
with fasting, in line with previous studies in chickens as well as that in mammals ( 42, 43 ). Under the physiological regulation of insulin, control (non-CORT) hens remained at a stable blood glucose level following feeding compared
Fig. 6. Effect of corticosterone on enzyme activity and expression of genes related to VLDL synthesis of liver in Experiment 1. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days) on the activities of FAS (nmol/min/mg protein) and ME ( � mol/min/mg protein) (A, B, n = 8); the mRNA expression of LXR � , SREBP-1c, ACC1, FAS, ME, SCD1, apoB100, apoVLDL-II, and MTP (C, D, n = 8); the protein expression of apoB100 (E, n = 4); and the hepatic VLDL secretion rate (F, n = 8) of laying hens under fasting or feeding condi-tions. a, b Means with different letters are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
Corticosterone suppresses follicular growth of hen 1871
TABLE 3. Effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days) on laying performances of hens feeding with normal diet or high-energy diet (Experiment 2)
Item CORT Control Mean Probability
Feed intake, kcal/day Normal diet 298.6 ± 14.1 a 270.7 ± 6.1 b 284.8 x CORT: NS High-energy diet 249.1 ± 3.8 c 257.9 ± 3.8 c 253.5 y Diet: < 0.001 Mean 273.9 264.3 CORT*Diet: 0.032Laying rate, % Normal diet 54.8 ± 6.3 83.3 ± 9.7 69.1 CORT: 0.046 High-energy diet 68.3 ± 12.8 82.9 ± 6.2 75.6 Diet: NS Mean 61.5 n 83.1 m CORT*Diet: NSEgg production, g/day/hen Normal diet 31.9 ± 3.8 47.9 ± 6.9 39.9 CORT: 0.056 High-energy diet 38.9 ± 7.2 48.3 ± 4.0 43.6 Diet: NS Mean 35.4 48.1 CORT*Diet: NSEgg weight, g Normal diet 58.2 ± 0.3 58.1 ± 0.7 58.2 CORT: NS High-energy diet 57.0 ± 0.5 56.2 ± 0.5 56.6 Diet: NS Mean 57.6 57.1 CORT*Diet: NS
Mean ± SE (n = 3). NS, not signifi cant . a, b, c Means with different superscript letters are signifi cantly different, P < 0.05. x, y Means with different superscript letters are signifi cantly different, P < 0.05. m, n Means with different superscript letters are signifi cantly different, P < 0.05.
blood glucose of CORT hens was elevated following feeding compared with fasting or compared with control hens of feeding state, indicating that insulin-dependent glucose utilization was impaired. These results suggest that CORT disturbed the insulin-dependent regulation of glucose utilization, in accordance with our previous work ( 13 ).
GC-suppressed follicular development is associated with reduced availability of yolk precursors
The growth of chicken oocytes can be divided into three phases: the growth of young oocytes (white follicles), the slow growth phase (SYF), and the rapid growth phase (HF) ( 44 ). During the last stage before oviposition, a large amount of the yolk precursors VLDL and VTG are taken up from the circulation each day. The decreased plasma levels of VLDL and TG in either the feeding or fasting state indi-cated a decreased availability of yolk precursor for the growth of oocytes. This result was consistent with previous studies in birds. In female zebra fi nches, CORT treatment resulted in high plasma levels of VLDL (indicated by the concentration of TG) but low levels of plasma VTG seven days after implantation, suggesting that CORT inhibited yolk precursor production and perhaps shifted lipid me-tabolism away from the production of VLDLy and toward the production of generic (non-yolk) VLDL ( 34 ). However, the VLDL and TG levels reduced by CORT were restored when chickens were fed a high-energy diet in Experiment 2, suggesting an energy-dependent mode of action.
In laying hens, VLDL and VTG are taken up by the grow-ing oocytes via receptor-mediated endocytosis, and both macromolecules bind to the OVR, localized on the oocyte plasma membrane ( 45 ). It is possible that OVR, as a key component in yolk precursor import, may play a signifi cant role in the CORT-induced suppression of oocyte growth. At the onset of the rapid growth phase of oocytes, the redis-tribution of OVR from the preexisting pool to the periph-eral region plays a crucial role in the regulation of yolk
formation and oocyte growth ( 46 ). In Experiment 1, OVR protein content was decreased by CORT in the fasting state rather than in the feeding state, indicating that CORT suppressed yolk formation when insuffi cient energy was available. The results from Experiment 1 and Experiment 2 consistently show that the mRNA level of OVR was in-creased by CORT during the fasting state. The diverse change of OVR mRNA and protein expression may be related to the posttranscriptional control of gene expres-sion. The regulation mechanism of CORT on OVR needs further investigation.
The transport of yolk precursors from the circulation to the oocytes consists of fi ve components ( 47 ): i ) vascularized connective tissue representing an extension of the follicle stalk, ii ) the thecal cell layer, iii ) the basement membrane, iv ) an epitheloid monolayer of granulosa cells, and v ) the zona pellucida. During follicular development, there is a develop-mental state-specifi c expression of occludin in granular cells ( 48 ). Claudin-based tight junctions are crucial for the barrier function in mammalian granular cells ( 49 ). The CORT-in-duced upregulation of occludin and claudin1 expression in the present study indicates an increase in tight junctions. These results may imply that CORT makes the granulosa cell layer more condensed, preventing the paracellular transport of yolk precursors into the oocytes. Moreover, the overexpres-sion of occludin and claudin1 was less signifi cant in the ener-gy-suffi cient state. These results together suggest that CORT exerts a more detrimental effect on follicular development when there is an energy defi cit.
GCs decrease yolk precursor secretion by suppressing apolipoprotein synthesis
It is well known that estrogens stimulate the liver to pro-duce the egg-yolk precursors VLDLy and VTG at the onset of sexual maturity. The shift of VLDL particle diameter is directed to be capable of being incorporated into newly forming egg yolks in birds and domestic chickens ( 25 ). According to Walzenm ( 26 ) and Salvante et al. ( 25 ), small
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
1872 Journal of Lipid Research Volume 54, 2013
Fig. 7. Effect of corticosterone on organ and tissue development in Experiment 2. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days, n = 8) on the development of ovaries (A, B), hierarchical follicles (C, D), small yellow follicles (E, F), and liver and abdomi-nal fat (G, H) of laying hens fed a normal diet (2,654 kcal/kg) or a high-energy diet (2,917 kcal/kg). Mass (g); fractional mass (g/kg BM). a, b Means with different letters are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from
Corticosterone suppresses follicular growth of hen 1873
mRNA expression of genes related to hepatic de novo fatty acid synthesis, such as FAS, ACC1, ME, and SCD1, was fur-ther analyzed, and the results demonstrate an increase in de novo hepatic lipogenesis by CORT, in agreement with the previous studies of broiler chickens ( 13, 50, 51 ).
SREBP-1 is an accessory transcription factor that plays an active and central role in the hormonal and nutritional reg-ulation of lipogenesis in mammals ( 52 ) as well as in avian species ( 53 ). LXRs, the nuclear hormone receptors that form active heterodimers with retinoid X receptors, are known to regulate SREBP-1c gene expression ( 54, 55 ). In rats, GCs are involved in the regulation of the expression of LXR � and SREBP-1c ( 56, 57 ). In the fasting state, however, the SREBP-1c mRNA level was upregulated following CORT administration without any measurable change in the ex-pression of LXR � , indicating the different mechanisms of regulation in chickens. These results suggest that SREBP-1c is associated with enhanced gene expression for hepatic li-pogenesis by CORT, in agreement with our previous obser-vation in broilers ( 13 ). These results imply that CORT improves hepatic lipogenesis via the SREBP pathway.
diameter VLDL particles (25–44 nm) were positively asso-ciated with egg production. In Experiment 1, the increased proportion of large diameter VLDL particles and the de-creased proportion of small diameter particles by CORT demonstrated the decreased circulating concentrations of VLDLy. This result implies two potential problems in CORT-challenged hens: suppressed hepatic lipogenesis and impaired hepatic VLDLy assembly and secretion.
In our previous studies of broiler chickens, GCs in con-cert with the high circulating concentration of insulin stimulated de novo hepatic lipogenesis, which in turn aug-mented blood lipid fl ux and peripheral fat deposition ( 12, 13 ). In the present study, the circulating insulin level was signifi cantly increased in CORT-treated hens. More-over, liver weight and liver lipid accumulation and hepatic TG content were increased by CORT, implying that CORT induces an augmented hepatic lipogenesis in laying hens similar to the effect in broiler chickens. This result was consistent with the elevated FAS and ME activities. The
Fig. 9. Effect of corticosterone on the expression of genes re-lated to yolk deposition of hierarchical follicles in Experiment 2. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days, n = 8) on the mRNA expression of yolk deposition-related genes of laying hens fed with a normal diet (A, 2,654 kcal/kg) or a high-energy diet (B, 2,917 kcal/kg). a, b Means with different letters are signifi cantly different, P < 0.05.
Fig. 8. Effect of corticosterone on plasma parameters in Experi-ment 2. The effect of corticosterone treatment (CORT, 2 mg/kg BM for 7 days, n = 8) on the plasma concentrations of hormones and metabolites of laying hens fed with a normal diet (A, 2,654 kcal/kg) or a high-energy diet (B, 2,917 kcal/kg). Hormones: E2 and insulin. Metabolites: glucose, VLDL, VTG, TG, and NEFA. a, b Means with different letters are signifi cantly different, P < 0.05.
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from

1874 Journal of Lipid Research Volume 54, 2013
feeding state or the high-energy diet state, suggesting that the effect of CORT on the release of estrogen is en-ergy dependent. Taken together, the decreased estro-gen level, downregulated apolipoprotein expression, and reduced VLDLy proportion raise the possibility that im-paired VLDLy secretion is related to the reduced induc-tion by estrogen.
It is generally known that GCs regulate the secretion of sex steroids via indirect or/and direct mechanisms ( 69 ): The indirect mechanism is by inhibition at the suprago-nadal level. GCs induce an interruption of hypothalamic GnRH release and in turn pituitary gonadotropins secre-tion, which suppress sex steroids synthesis. An endocrine feedback loop is present in this HPG axis, as high levels of sex steroids suppress the release of GnRH through its re-turn to the hypothalamus. The direct mechanism is at the gonadal level by inhibiting sex steroids production in granulosa cells. In the present study, estrogen secretion was reduced by CORT, while the blood concentrations of FSH and LH remained higher and the amounts of FSHR and LHR in HF were not affected. According to the previ-ous studies ( 70–72 ), the increased plasma concentration of FSH and LH may be feedback compensation in re-sponse to low estrogen secretion. Another possibility is that CORT directly affected estrogen production in granu-losa cells in a gonadotropin-independent manner. This speculation is supported by several studies. Heat stress ex-erted a direct effect on ovarian tissue, which was suggested as a factor contributing to the decline in egg production ( 73, 74 ). CORT altered the responsiveness of ovarian tis-sues to gonadotropins ( 75 ). In rats, GCs inhibited FSH-induced estrogen production in cultured rat granulosa cells ( 76 ).
In conclusion, exogenous GC administration signifi -cantly decreased the laying performance of hens in an energy-dependent manner. These results suggest that the decreased apolipoprotein synthesis and VLDLy secretion by the liver might be responsible for the decreased avail-ability of circulating yolk precursor in CORT-treated hens. In HF, the upregulated expression of occludin and clau-din in granulosa cells further prevented yolk deposition into oocytes.
The authors thank Ms. Jinying Wang for care of animals.
REFERENCES
1 . Schoech , S. J. , M. A. Rensel , E. S. Bridge , R. K. Boughton , and T. E. Wilcoxen . 2009 . Environment, glucocorticoids, and the timing of reproduction. Gen. Comp. Endocrinol. 163 : 201 – 207 .
2 . Goutte , A. , F. Angelier , C. C. Chastel , C. Trouvé , B. Moe , C. Bech , G. W. Gabrielsen , and O. Chastel . 2010 . Stress and the timing of breeding: glucocorticoid-luteinizing hormones relationships in an arctic seabird. Gen. Comp. Endocrinol. 169 : 108 – 116 .
3 . Goutte , A. , C. Clément-Chastel , B. Moe , C. Bech , G. W. Gabrielsen , and O. Chastel . 2011 . Experimentally reduced corticosterone release promotes early breeding in black-legged kittiwakes. J. Exp. Biol. 214 : 2005 – 2013 .
4 . Williams , J. B. , R. J. Etches , and J. Rzasa . 1985 . Induction of a pause in laying by corticosterone infusion or dietary alterations: effects
As hepatic lipogenesis remained at a higher level in CORT-treated hens, we further investigated whether CORT affected hepatic VLDL assembly and secretion. A reduction in the VLDL secretion rate in Experiment 1 im-plies that CORT decreased circulating VLDL content by inhibiting the assembly of lipoprotein by the liver. ApoB and apoVLDL-II are required for VLDLy assembly in the liver, and they provide structural support that accommo-dates greater amounts of TG on the surface of the colloi-dal particles ( 58 ). The expressions of apoVLDL-II and apoB100 were downregulated by CORT in Experiment 1. Successful assembly of VLDL requires the cotranslational association of apoB100 with polar and neutral lipids ( 59 ). ApoVLDL-II functions as an inhibitor of LPL to ensure the delivery of VLDL to the oocyte membranes without sub-stantial lipolysis ( 60 ). These results suggest that decreased expression of apolipoprotein could be responsible for the suppressed VLDLy assembly and secretion in CORT-treated hens.
MTP is involved in delivering TG to nascent apoB mol-ecules during the assembly of lipoprotein particles ( 61, 62 ). Experiment 1 shows that CORT treatment elevated MTP mRNA expression. The accumulated TG in the liver may play a positive regulatory role in MTP activity ( 63 ). This result may imply that MTP-mediated lipid transfer is not the main factor responsible for decreased VLDLy secretion.
ApoVLDL-II is responsible for the decrease in VLDLy diameter ( 64 ). The present result suggests that the down-regulated gene expression for apoVLDL-II is associated with the altered VLDL particle diameter by CORT. More-over, apoVLDL-II is well known to confer LPL resistance to VLDL ( 60, 65 ). This characteristic enables VLDL to re-sist the lipolytic activity of LPL, thereby securing a high TG content for the egg yolk. In the present study, an increase in LPL mRNA expression was observed in HF upon CORT treatment in the fasting state. Further investigation is re-quired to determine whether upregulated LPL expression and increased VLDL particle diameter are involved in the suppression of follicular development.
GC-induced decrease in estrogen level is energy dependent
Estrogen is recognized as playing the primary role in the conversion of carbohydrate to TG and in the production of apoB and apoVLDL-II, leading to the hypersecretion of these lipoproteins into the circulation ( 8, 66 ). ApoVLDL-II and apoB were shown to be rapidly induced by exogenous estrogen in the rooster liver and to accumulate within the VLDL fraction ( 67 ). With the onset of egg production, es-trogen shifts hepatic lipoprotein production from generic VLDL to VLDLy, which is resistant to LPL and to intact uptake by oocyte receptors ( 64 ). This dramatic change oc-curs in both domestic laying hens selected for high egg production ( 68 ) and wild birds [Passerine bird ( 25 )].
Estrogen is synthesized in the granulosa cells of folli-cles and exerts regulatory activity by binding to the ESR. In the present study, the circulating concentration of es-trogen was decreased in the fasting state but not in the
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from

Corticosterone suppresses follicular growth of hen 1875
25 . Salvante , K. G. , G. Lin , R. L. Walzem , and T. D. Williams . 2007 . Characterization of very-low density lipoprotein particle diameter dynamics in relation to egg production in a passerine bird. J. Exp. Biol. 210 : 1064 – 1074 .
26 . Walzenm , R. L. 1996 . Lipoproteins and the laying hen: form fol-lows function. Poult. Avian Biol. Rev. 7 : 31 – 64 .
27 . Griffi n , H. D. , and C. C. Whitehead . 1982 . Plasma lipoprotein con-centration as an indicator of fatness in broilers: development and use of a simple assay for plasma very low density lipoproteins. Br. Poult. Sci. 23 : 307 – 313 .
28 . Simon , J. , P. Freychet , and G. Rosselin . 1974 . Chicken insulin: ra-dioimmunological characterization and enhanced activity in rat fat cells and liver plasma membranes. Endocrinology . 95 : 1439 – 1449 .
29 . Halestrap , A. P. , and R. M. Denton . 1973 . Insulin and the regu-lation of adipose tissue acetyl-coenzyme A carboxylase. Biochem. J. 132 : 509 – 517 .
30 . Hsu , R. Y. , and H. A. Lardy . 1969 . Malic enzyme. In Methods in Enzymology. Vol. 13. J. M. Lowenstein, editor. Academic Press, New York, NY. 230–235.
31 . Livak , K. J. , and T. D. Schmittgen . 2001 . Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods . 25 : 402 – 408 .
32 . Laemmli , U. K. 1970 . Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature . 227 : 680 – 685 .
33 . Petitte , J. N. , and R. J. Etches . 1991 . Daily infusion of corticosterone and reproductive function in the domestic hen (Gallus domesti-cus). Gen. Comp. Endocrinol. 83 : 397 – 405 .
34 . Salvante , K. G. , and T. D. Williams . 2003 . Effects of corticosterone on the proportion of breeding females, reproductive output and yolk precursor levels. Gen. Comp. Endocrinol. 130 : 205 – 214 .
35 . Alexander , B. M. , Z. Kiyma , M. McFarland , E. A. Van Kirk , D. M. Hallford , D. E. Hawkins , K. K. Kane , and G. E. Moss . 2007 . Infl uence of short-term fasting during the luteal phase of the estrous cycle on ovarian follicular development during the ensuing proestrus of ewes. Anim. Reprod. Sci. 97 : 356 – 363 .
36 . Yan , J. , B. Zhou , J. Yang , P. Tai , X. Chen , H. Zhang , M. Zhang , and G. Xia . 2008 . Glucose can reverse the effects of acute fasting on mouse ovulation and oocyte maturation. Reprod. Fertil. Dev. 20 : 703 – 712 .
37 . Zabuli , J. , T. Tanaka , W. Lu , and H. Kamomae . 2010 . Intermittent nutritional stimulus by short-term treatment of high-energy diet promotes ovarian performance together with increases in blood levels of glucose and insulin in cycling goats. Anim. Reprod. Sci. 122 : 288 – 293 .
38 . Strack , A. M. , C. J. Horsley , R. J. Sebastian , S. F. Akana , and M. F. Dallman . 1995 . Glucocorticoids and insulin: complex interaction on brown adipose tissue. Am. J. Physiol. 268 : R1209 – R1216 .
39 . Salley , K. E. , E. P. Wickham , K. I. Cheang , P. A. Essah , N. W. Karjane , and J. E. Nestler . 2007 . Glucose intolerance in polycys-tic ovary syndrome - a position statement of the Androgen Excess Society. J. Clin. Endocrinol. Metab. 92 : 4546 – 4556 .
40 . Brothers , K. J. , S. Wu , S. A. DiVall , M. R. Messmer , C. R. Kahn , R. S. Miller , S. Radovick , F. E. Wondisford , and A. Wolfe . 2010 . Rescue of obesity-induced infertility in female mice due to a pituitary-spe-cifi c knockout of the insulin receptor. Cell Metab. 12 : 295 – 305 .
41 . Diamanti-Kandarakis , E. 2008 . Polycystic ovarian syndrome: patho-physiology, molecular aspects and clinical implications. Expert Rev. Mol. Med. 10 : e3 .
42 . Millward , D. J. , D. O. Nnanyelugo , W. P. James , and P. J. Garlick . 1974 . Protein metabolism in skeletal muscle: the effect of feeding and fasting on muscle RNA, free amino acids and plasma insulin concentrations. Br. J. Nutr. 32 : 127 – 142 .
43 . Dupont , J. , M. Derouet , J. Simon , and M. Taouis . 1999 . Corti cos-terone alters insulin signalling in chicken muscle and liver at differ-ent steps. J. Endocrinol. 162 : 67 – 76 .
44 . Griffi n , H. D. , M. M. Perry , and A. B. Gilbert . 1984 . Yolk forma-tion. In Physiology and Biochemistry of the Domestic Fowl. B. M. Freeman, editor. Academic Press, London. 345–380.
45 . Nimpf , J. , and W. J. Schneider . 1991 . Receptor-mediated lipopro-tein transport in laying hens. J. Nutr. 121 : 1471 – 1474 .
46 . Shen , X. , E. Steyrer , H. Retzek , E. J. Sanders , and W. J. Schneider . 1993 . Chicken oocyte growth: receptor-mediated yolk deposition. Cell Tissue Res. 272 : 459 – 471 .
47 . Schneider , W. J. 2009 . Receptor-mediated mechanisms in ovarian follicle and oocyte development. Gen. Comp. Endocrinol. 163 : 18 – 23 .
48 . Schuster , M. K. , B. Schmierer , A. Shkumatava , and K. Kuchler . 2004 . Activin A and follicle-stimulating hormone control tight junctions
on the reproductive system, food consumption and body weight. Br. Poult. Sci. 26 : 25 – 34 .
5 . Shini , S. , A. Shini , and G. R. Huff . 2009 . Effects of chronic and re-peated corticosterone administration in rearing chickens on physi-ology, the onset of lay and egg production of hens. Physiol. Behav. 98 : 73 – 77 .
6 . Kuksis , A. 1992 . Yolk lipids. Biochim. Biophys. Acta . 1124 : 205 – 222 . 7 . Challenger , W. O. , T. D. Williams , J. K. Christians , and F. Vézina .
2001 . Follicular development and plasma yolk precursor dynam-ics through the laying cycle in the European starling (Sturnus vul-garis). Physiol. Biochem. Zool. 74 : 356 – 365 .
8 . Kirchgessner , T. G. , C. Heinzmann , K. L. Svenson , D. A. Gordon , M. Nicosia , H. G. Lebherz , A. J. Lusis , and D. L. Williams . 1987 . Regulation of chicken apolipoprotein B: cloning, tissue distribu-tion, and estrogen induction of mRNA. Gene . 59 : 241 – 251 .
9 . Berkowitz , E. A. , and M. I. Evans . 1992 . Functional analysis of reg-ulatory regions upstream and in the fi rst intron of the estrogen-responsive chicken very low density apolipoprotein II gene. J. Biol. Chem. 267 : 7134 – 7138 .
10 . Yen , C. F. , Y. N. Jiang , T. F. Shen , I. M. Wong , C. C. Chen , K. C. Chen , W. C. Chang , Y. K. Tsao , and S. T. Ding . 2005 . Cloning and expression of the genes associated with lipid metabolism in Tsaiya ducks. Poult. Sci. 84 : 67 – 74 .
11 . Lin , H. , E. Decuypere , and J. Buyse . 2004 . Oxidative stress induced by corticosterone administration in broiler chickens (Gallus gallus domesticus) 1. Chronic exposure. Comp. Biochem. Physiol. B . 139 : 737 – 744 .
12 . Cai , Y. , Z. Song , X. Zhang , X. Wang , H. Jiao , and H. Lin . 2009 . Increased de novo lipogenesis in liver contributes to the augmented fat deposition in dexamethasone exposed broiler chickens (Gallus gallus domesticus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 150 : 164 – 169 .
13 . Cai , Y. , Z. Song , X. Wang , H. Jiao , and H. Lin . 2011 . Dexamethasone Induced Hepatic Lipogenesis is Insulin Dependent in Chickens (Gallus gallus domesticus). Stress . 14 : 273 – 281 .
14 . Wang , X. , H. Lin , Z. Song , and H. Jiao . 2010 . Dexamethasone facilitates lipid accumulation and mild feed restriction improves fatty acids oxidation in skeletal muscle of broiler chicks (Gallus gal-lus domesticus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 151 : 447 – 454 .
15 . Wingfi eld , J. C. , and R. M. Sapolsky . 2003 . Reproduction and resis-tance to stress: when and how. J. Neuroendocrinol. 15 : 711 – 724 .
16 . Schoech , S. J. , R. Bowman , E. S. Bridge , G. M. Morgan , M. A. Rensel , T. E. Wilcoxen , and R. K. Boughton . 2007 . Corticosterone administration does not affect timing of breeding in Florida scrub-jays (Aphelocoma coerulescens). Horm. Behav. 52 : 191 – 196 .
17 . Close , B. , K. Banister , V. Baumans , E. M. Bernoth , N. Bromage , J. Bunyan , W. Erhardt , P. Flecknell , N. Gregory , H. Hackbarth , et al . 1997 . Recommendations for euthanasia of experimental animals: Part 2. DGXT of the European Commission. Lab. Anim. 31 : 1 – 32 .
18 . Wang , L. , Y. Han , C. S. Kim , Y. K. Lee , and D. D. Moore . 2003 . Resistance of SHP-null mice to bile acid-induced liver damage. J. Biol. Chem. 278 : 44475 – 44481 .
19 . Lillie , R. D. , and H. M. Fullmer . 1976 . Histopathologic Technic and Practical Histochemistry. 4 th edition. McGraw-Hill, New York.
20 . Folch , J. , M. Lees , and G. H. Sloane Stanley . 1957 . A simple method for the isolation and purifi cation of total lipids from animal tissues. J. Biol. Chem. 226 : 497 – 509 .
21 . Jong , M. C. , V. E. Dahlmans , P. J. van Gorp , K. W. van Dijk , M. L. Breuer , M. H. Hofker , and L. M. Havekes . 1996 . In the absence of the low density lipoprotein receptor, human apolipoprotein C1 overexpression in transgenic mice inhibits the hepatic uptake of very low density lipoproteins via a receptor- associated protein-sen-sitive pathway. J. Clin. Invest. 98 : 2259 – 2267 .
22 . Aalto-Setala , K. , E. A. Fisher , X. Chen , T. Chajek-Shaul , T. Hayek , R. Zechner , A. Walsh , R. Ramakrishnan , H. N. Ginsberg , and J. L. Breslow . 1992 . Mechanism of hypertriglyceridemia in human apolipoprotein (Apo) CIII transgenic mice. J. Clin. Invest. 90 : 1889 – 1900 .
23 . Fless , G. M. 1991 . General preparative ultracentrifugation and considerations for lipoprotein isolation. In Analyses of Fats, Oils and Lipoproteins. E. G. Perkins, editor. American Oil Chemists’ Society, Champaign, IL. 512–523.
24 . Walzem , R. L. , P. A. Davis , and R. J. Hansen . 1994 . Overfeeding increases very low density lipoprotein diameter and causes the ap-pearance of a unique lipoprotein particle in association with failed yolk deposition. J. Lipid Res. 35 : 1354 – 1366 .
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from

1876 Journal of Lipid Research Volume 54, 2013
62 . Sharp , D. , L. Blinderman , K. A. Combs , B. Kienzle , B. Ricci , K. Wager-Smith , C. M. Gil , C. W. Turck , M. E. Bouma , D. J. Rader , et al . 1993 . Cloning and gene defects in microsomal triglyceride transfer protein associated with abetalipoproteinaemia. Nature . 365 : 65 – 69 .
63 . Lu , S. , M. Huffman , Y. Yao , C. M. Mansbach 2nd , X. Cheng , S. Meng , and D. D. Black . 2002 . Regulation of MTP expression in de-veloping swine. J. Lipid Res. 43 : 1303 – 1311 .
64 . Walzem , R. L. , R. J. Hansen , D. L. Williams , and R. L. Hamilton . 1999 . Estrogen induction of VLDLy assembly in egg-laying hens. J. Nutr. 129 : 467S – 472S .
65 . Griffi n , H. , G. Grant , and M. Perry . 1982 . Hydrolysis of plasma triacylglycerol-rich lipoproteins from immature and laying hens (Gallus domesticus) by lipoprotein lipase in vitro. Biochem. J. 206 : 647 – 654 .
66 . Dashti , N. , J. L. Kelley , R. H. Thayer , and J. A. Ontko . 1983 . Concurrent inductions of avian hepatic lipogenesis, plasma lipids, and plasma apolipoprotein B by estrogen. J. Lipid Res. 24 : 368 – 380 .
67 . Williams , D. L. 1979 . Apoproteins of avian very low density lipopro-tein: demonstration of a single high molecular weight apoprotein. Biochemistry . 18 : 1056 – 1063 .
68 . Hermier , D. , P. Forgez , J. Williams , and M. J. Chapman . 1989 . Alterations in plasma lipoproteins and apolipoproteins associated with estrogen-induced hyperlipidemia in the laying hen. Eur. J. Biochem. 184 : 109 – 118 .
69 . Whirledge , S. , and J. A. Cidlowski . 2010 . Glucocorticoids, stress, and fertility. Minerva Endocrinol. 35 : 109 – 125 .
70 . Dubey , A. K. , and T. M. Plant . 1985 . A suppression of gonado-tropin secretion by cortisol in castrated male rhesus monkeys (Macaca mulatta) mediated by the interruption of hypotha-lamic gonadotropin-releasing hormone release. Biol. Reprod. 33 : 423 – 431 .
71 . Kamel , F. , and C. L. Kubajak . 1987 . Modulation of gonadotropin secretion by corticosterone: interaction with gonadal steroids and mechanism of action. Endocrinology . 121 : 561 – 568 .
72 . Briski , K. P. , and P. W. Sylvester . 1991 . Acute inhibition of pituitary LH release in the male rat by the glucocorticoid agonist decadron phosphate. Neuroendocrinology . 54 : 313 – 320 .
73 . Novero , R. P. , M. M. Beck , E. W. Gleaves , A. L. Johnson , and J. A. Deshazer . 1991 . Plasma progesterone, luteinizing hormone concentrations, and granulosa cell responsiveness in heat-stressed hens. Poult. Sci. 70 : 2335 – 2339 .
74 . Rozenboim , I. , E. Tako , O. Gal-Garber , J. A. Proudman , and Z. Uni . 2007 . The effect of heat stress on ovarian function of laying hens. Poult. Sci. 86 : 1760 – 1765 .
75 . Etches , R. J. , J. N. Petitte , and C. E. Anderson-Langmuir . 1984 . Interrelationships between the hypothalamus, pituitary gland, ovary, adrenal gland, and the open period for LH release in the hen (Gallus domesticus). J. Exp. Zool. 232 : 501 – 511 .
76 . Hsueh , A. J. , and G. F. Erickson . 1978 . Glucocorticoid inhibition of FSH-induced estrogen production in cultured rat granulosa cells. Steroids . 32 : 639 – 648 .
in avian granulosa cells by regulating occludin expression. Biol. Reprod. 70 : 1493 – 1499 .
49 . Furuse , M. , M. Hata , K. Furuse , Y. Yoshida , and A. Haratake . 2002 . Claudin-based tight junctions are crucial for the mammalian epi-dermal barrier: a lesson from claudin-1-defi cient mice. J. Cell Biol. 156 : 1099 – 1111 .
50 . Puvadolpirod , S. , and J. P. Thaxton . 2000a . Model of physiological stress in chickens 1. Response parameters. Poult. Sci. 79 : 363 – 369 .
51 . Puvadolpirod , S. , and J. P. Thaxton . 2000b . Model of physiological stress in chickens 2. Dosimetry of adrenocorticotropin. Poult. Sci. 79 : 370 – 376 .
52 . Yahagi , N. , H. Shimano , A. H. Hasty , T. Matsuzaka , T. Ide , T. Yoshikawa , M. Amemiya-Kudo , S. Tomita , H. Okazaki , Y. Tamura , et al . 2002 . Absence of sterol regulatory element-binding protein-1 (SREBP-1) ameliorates fatty livers but not obesity or insulin resis-tance in Lep(ob)/Lep(ob) mice. J. Biol. Chem. 277 : 19353 – 19357 .
53 . Assaf , S. , D. Hazard , F. Pitel , M. Morisson , M. Alizadeh , F. Gondret , C. Diot , A. Vignal , M. Douaire , and S. Lagarrigue . 2003 . Cloning of cDNA encoding the nuclear form of chicken sterol response element binding protein-2 (SREBP-2), chromosomal localization, and tissue expression of chicken SREBP-1 and -2 genes. Poult. Sci. 82 : 54 – 61 .
54 . Li , A. C. , and C. K. Glass . 2004 . PPAR- and LXR-dependent path-ways controlling lipid metabolism and the development of athero-sclerosis. J. Lipid Res. 45 : 2161 – 2173 .
55 . Kalaany , N. Y. , K. C. Gauthier , A. M. Zavacki , P. P. Mammen , T. Kitazume , J. A. Peterson , J. D. Horton , D. J. Garry , A. C. Bianco , and D. J. Mangelsdorf . 2005 . LXRs regulate the balance between fat storage and oxidation. Cell Metab. 1 : 231 – 244 .
56 . Paterson , J. M. , N. M. Morton , C. Fievet , C. J. Kenyon , M. C. Holmes , B. Staels , J. R. Seckl , and J. J. Mullins . 2004 . Metabolic syndrome without obesity: hepatic overexpression of 11 � -hydroxysteroid de-hydrogenase type 1 in transgenic mice. Proc. Natl. Acad. Sci. USA . 101 : 7088 – 7093 .
57 . Erhuma , A. , S. McMullen , S. C. Langley-Evans , and A. J. Bennett . 2009 . Feeding pregnant rats a low-protein diet alters the hepatic ex-pression of SREBP-1c in their offspring via a glucocorticoid-related mechanism. Endocrine . 36 : 333 – 338 .
58 . Boyle-Roden , E. , and R. L. Walzem . 2005 . Integral apolipoproteins increase surface-located triacylglycerol in intact native apoB-100-containing lipoproteins. J. Lipid Res. 46 : 1624 – 1632 .
59 . Manchekar , M. , P. E. Richardson , T. M. Forte , G. Datta , J. P. Segrest , and N. Dashti . 2004 . Apolipoprotein B-containing lipoprotein par-ticle assembly: lipid capacity of the nascent lipoprotein particle. J. Biol. Chem. 279 : 39757 – 39766 .
60 . Schneider , W. J. , R. Carroll , D. L. Severson , and J. Nimpf . 1990 . Apolipoprotein VLDL-II inhibits lipolysis of triglyceride-rich lipo-proteins in the laying hen. J. Lipid Res. 31 : 507 – 513 .
61 . Wetterau , J. R. , L. P. Aggerbeck , M. E. Bouma , C. Eisenberg , A. Munck , M. Hermier , J. Schmitz , G. Gay , D. J. Rader , and R. E. Gregg . 1992 . Absence of microsomal triglyceride transfer protein in individuals with abetalipoproteinemia. Science . 258 : 999 – 1001 .
by guest, on July 10, 2018w
ww
.jlr.orgD
ownloaded from