correlation of protoporphyrinogen oxidase inhibition by o-phenyl pyrrolidino- and...
TRANSCRIPT

Pestic. Sci. 1992, 35, 2277235
Correlation of Protoporphyrinogen Oxidase Inhibition by 0-Phenyl Pyrrolidino- and Piperidino-Carbarnates with their Herbicidal Effects Ujjana B. Nandihalli," Timothy D. Sherman,a Mary V. Duke," James D. Fisher,b Vincent A. MUSCO,~ Josk M. Becerrilc & Stephen 0. Dukeu* 5 o u t h e r n Weed Science Laboratory, Agricultural Research Service, United States Department of Agriculture, PO Box 350, Stoneville, Mississippi 38776, USA 'Rohm and Haas Company, 727 Norristown Road, Spring House, Pennsylvania 19477, USA "niversidad del Pais Vasco, Facultad de Ciencias, Dep. Biologia Vegetal y Ecologia, Apdo. Correo 644, 48080 Bilbao, Spain
(Revised manuscript received 27 January 1992; accepted 6 April 1992)
Abstract : Phenopylatet (2,4-dichlorophenyl 1-pyrrolidinecarboxylate) and 13 of its 0-phenyl pyrrolidino- and piperidino-carbamate analogues were tested for their capacity to inhibit protoporphyrinogen oxidase (Protox), to cause ac- cumulation of protoporphyrin IX (Proto IX), and to cause herbicidal injury. All 14 compounds inhibited Protox of barley etioplast preparations. The I,, values ranged from 21 nM to 4 mM. Highly significant correlations were found between Protox I,, values and herbicidal activity of the more active of the compounds on greenhouse-grown weeds (pitted morningglory, barnyardgrass, prickly sida, and velvetleaf), pre- and/or post-emergence, and on cucumber cotyledon discs or barley leaf sections in cellular leakage bioassays. Proto IX accumulation in barley leaf sections treated with any of the 14 compounds correlated well with both the I,, values for Protox inhibition and with herbicidal damage to the tissue sections. These data indicate most or all of the herbicidal damage caused by these compounds is due to inhibition of Protox.
1 INTRODUCTION normal porphyrin pathway to Proto IX.6~20 Proto IX accumulates in or near the plasmalemma, and, in the presence of molecular oxygen and light, generates highly destructive singlet oxygen. 21-23 The process is similar to the accumulation and cellular damage in humans and yeast with genetic defects in pro to^.^*,^^
No comprehensive study has been published, using a wide range of analogues of a Protox-inhibiting class of herbicides, to determine the relationship between in-vitro enzyme inhibition and herbicidal activity. In this paper we extend the list of chemical classes which inhibit Protox to include a range of phenyl pyrrolidine-l- carboxylate analogues which belong to the piperidino- and pyrrolidino-carbamate family (Table 1). These and similar compounds have been patented as 27
They were reported to cause herbicidal symptoms similar
Photo-bleaching herbicides from several different chemi- cal classes, including certain oxadiazoles, diphenyl ethers, pyrazole phenyl ethers, and N-phenylimides are potent inhibitors of protoporphyrinogen oxidase (Protox).l-' Protox is the last enzyme common to synthesis of both haem and chlorophyll. Inhibition of Protox leads to accumulation of high concentrations of protoporphyrin IX (Proto IX).1"-19 Apparently, when Protox is inhibited, protoporphyrinogen IX (Protogen IX), its substrate, leaves the chloroplast and is rapidly oxidized outside the
*To whom correspondence should be addressed. tPhenopylate is not a BSI/ISO-approved common name.
Pestic. Sci. 0031-613)3/92/$05.00 @ 1992 SCI. Printed in Great Britain 227

228 Ujjana B. Nandihalli et al.
TABLE 1 Structures and Protox Is, values for Compounds used in Studies
Compound 2 4 5 R' R2
Phenopylate RH 0710 RH 0978 RH 1224 RH 1422 RH 1908 RH 1909 RH 1911 RH 1964 RH 1965 RH 4663 RH 6251 RH 7160 RH 9611
CI c1 c1 CI F H CI H c1 CI CI CI c1 CI
C1 H
C1 OCH,C-CH CI OCH,C:CH CI CO,CH(CH,), H H H H C1 H CI CO,CH(CH,), C1 CO,CH, C1 OCH,C:CH CI OCH,CXH C1 OCH,C-CH CI OH
-OCH,CO.N(CH,CXH)-
Protox I,, (w) 2.1 0.15 0.030 0.045 0.02 1
~-
4000 240
11.50 0.40 0.70 0.12 0.28 7.50 4.70
to those of other Protox-inhibiting herbicides.2s How- ever, neither their effects on porphyrin accumulation nor on Protox activity have been reported preyiously. In this study, we report that some of these compounds are potent inhibitors of Protox and that their activity as Protox inhibitors correlates well with their capacity to cause Proto IX accumulation and their herbicidal activity at the tissue or whole plant level.
2 MATERIALS AND METHODS
2.1 Plant material
For in-vitro and tissue section experiments, barley (Hordeum vulgare L. [cv. Morex]) and cucumber (Cucumis sativus L. [cv. Straight Eight]) were grown in flats in a commercial greenhouse substrate (Jiffy-Mix ; JPA, West Chicago, IL) and watered with distilled water. Green plants were grown at 25" C under white light of 500 pmol m-' s-l photosynthetically active radiation (PAR) and > 90 YO relative humidity before tissues were harvested for use at 6 to 8 days of growth. Some plants were germinated in darkness at 25°C and grown in far- red light.29
For whole plant studies, prickly sida (Sida spinosa L.), velvetleaf (Abutilon theophrastii Medicus), barnyardgrass
(Echinochloa crus-galli (L.) Beauv.), and pitted morning- glory (Zpomoea lacunosa L.) plants were grown in greenhouses supplemented with artificial lighting pro- viding 11,000 to 13,000 lux at bench level at night. Plants were grown with a 16: 8 h light: dark photoperiod and temperatures ranged from 22 to 35°C. For pre-emergence studies, plants were grown in silty loam soil (1-1*5% organic matter) in 20 x 25 x 10 or 25 x 35 x 10 cm (width x length x depth) trays. Five to 20 seeds of each species were planted in each tray, depending on the seed size (fewer seeds for those with larger seeds).
For post-emergence greenhouse studies, plants were grown as for pre-emergence studies in preliminary studies and in 7.5- to 10-cm pots in later studies. Plants were treated at 14 to 21 days after planting. At this time, grasses were in the two- to four-leaf stage and broadleaf weeds were at the one- to two-true-leaf stage.
2.2 Herbicide treatment
In tissue section experiments, tissues were treated with the herbicides of Table 1 as before3' by cutting 504- mm diameter cucumber cotyledon discs with a cork borer, or 4-mm barley leaf squares with a razor blade, and then placing them in a polystyrene petri dish (6 crn diam.) in 5 ml of sucrose (10 g litref'), 2-[N- morpholino]ethanesulfonic acid (MES ; 1 mM, pH 6.5)

Protoporphyrinogen oxidase inhibition and herbicidal efects 229
medium with or without technical-grade herbicide dissolved in acetone. The final concentration of acetone in all dishes was 1.0 ml litre-'. The discs were then incubated at 25°C in darkness for 20 h before exposure to 500 pmol m-' s-' PAR for up to 24 h.
In pre-emergence greenhouse studies, acetone solu- tions of various concentrations of technical-grade test compounds were sprayed onto the surface of soil immediately after planting. In post-emergence green- house studies, acetone solutions of technical-grade test compounds were sprayed onto appropriately sized plants. Spraying was conducted with an endless belt sprayer in which test plants moved under a fixed spray nozzle at a rate to deliver spray volumes of 234 or 467 litre ha-'. After spray application, plants were placed in a vented cabinet to dry before being returned to the greenhouse. Pre-emergence treatments were provided with overhead watering and post-emergence treatments were sub-irrigated.
Two to three weeks after treatment, injury to the plants was evaluated on a scale of 0 (no injury) to 100 (complete plant death). The injury values were based on a composite of factors, including chlorosis, necrosis, and inhibition of growth. The scores were converted to LC,, values using probit analysis. The highest application rates in greenhouse studies were 4.44 kg ha-'. LC,, values of up to 11.2 kg ha-' were extrapolated from these studies.
2.3 Porphyrin determinations
All extractions for HPLC were made under a dim, green light source. Samples (0-2 g of barley leaf sections) were homogenized in HPLC-grade methanol + 0.1 M am- monium hydroxide (9 + 1 , by volume; 3 ml) with a Brinkmann Polytron at 30,000 rev min-l for 30 s. The homogenate was centrifuged at 30,OOOg for 10min at 0°C and the supernatant was saved. The pellet was resuspended in the basic methanol homogenization solution (3 ml), sonicated for 5 min, and centrifuged at 30,OOOg for 10 min at 0°C. Supernatants were combined and evaporated to dryness at 40°C with a rotary evaporator. The residue was dissolved in HPLC-grade basic methanol (2 ml) and filtered through a 0.2-pm syringe filter. Samples were stored in light-tight glass (wrapped in aluminium foil) vials at - 20°C until analysis by HPLC.
HPLC determinations of Proto IX, Mg-Proto IX, Mg- Proto IX monomethyl ester, and protochlorophyllide (PChlide) were made as b e f ~ r e . ~ All porphyrin com- pound levels were expressed on a molar basis per gram of fresh weight. All treatments for porphyrin samples were triplicated.
2.4 Protox assays
Leaves of etiolated barley were homogenized with a
Sorvall Omnimixer for 30 s at full speed using a fresh- weight-to-volume ratio of 1 : 5. Homogenization buffer consisted of 2-[4-(2-hydroxyethyl)piperazin- 1 -yl]- ethanesulfonic acid (HEPES; 10 mM, pH 7.7 at 2°C); sorbitol (330 mM); ethylenediaminetetra(acetic acid) (EDTA; 1 mM); magnesium chloride ( 1 mM); and cysteine ( 5 mM). Homogenates were filtered through two layers of Miracloth, and crude cell debris was removed by centrifugation at 150g for 5 min at 4°C. Plastids were pelleted from this supernatant by centrifugation at 6000g for 20 min at 4°C. On ice, plastids were resuspended in assay buffer and disrupted by sonication in two 10-sec bursts, with intermittent cooling. The assay buffer was the same as the homogenization buffer except that dithiothreitol (DTT; 1 mM) was substituted for cysteine (5 mM), and phenylmethylsulfonyl fluoride (PMSF; I mM), leupeptin ( ~ O P M ) , pepstatin ( ~ O P M ) , and 1 , lO- phenanthroline (5 mM) were added as protease inhibi- tors. Protein concentration was determined using the method of Bradford,31 with bovine serum albumin as a standard. Extracts were resuspended to a concentration of 4 mg protein ml-' in assay buffer and stored at - 80°C until use.
Protogen IX was prepared according to Jacobs and To slow the rate of autooxidation, DTT was
added to a final concentration of 2 mM. Prior to assay, plastid extracts were thawed and held
on ice. In the herbicide treatments, herbicides were added in a volume of 2p l of acetone to 2 0 0 ~ 1 of extract. Acetone alone was added to control treatments. The assay mixture consisted of HEPES (25 mM, pH 7.5, 3OoC), EDTA (5mM), DTT ( 2 m ~ ) , and Protogen IX (c. 2 PM). The reaction was initiated by addition of 0.2 ml of extract (k herbicide) to 0.8 ml of assay mixture and monitored for Proto IX fluorescence for 2 min at 30°C. Fluorescence was monitored directly from the assay mixture using a Shimadzu RF-5000 U, temperature- controlled, recording spectrofluorometer with excitation at 395 nm and emission monitored at 622 nm. The reaction rate was essentially constant over this 2-min period. Autooxidation of Protogen IX to Proto IX in the presence of heat-inactivated extract was negligible.
3 RESULTS
3.1 Herbicidal activity
The LC,, values for several of the more active com- pounds in Table 1 for four weed species applied both pre- emergence and post-emergence in greenhouse studies are presented in Table 2. LC,, values ranged over almost two orders of magnitude, despite the fact that the upper values could not be determined, Pre-emergence treatment was more effective than was post-emergence treatment against prickly sida. However, in velvetleaf and barnyardgrass, some of the compounds were more active

230 Ujjunu B. Nundihalli et al.
TABLE 2 Pre- and Post-emergence Whole-Plant Activity of Some of the Compounds of Table 1
Compound
Phenopylate RH 0978 RH 1224 RH 1422 RH 1964 RH 1965 RH 4663 RH 6251
Pre-emergence LC,, (kg ha-') Post-emergence LC,, (kg ha-'). _ _
BYG" PSI VEL BYG MG PSI VEL
0.62 0.44 1.85 7.19 5.99 3.00 0.49 0.20 0.056 0.056 1.12 0.26 0.13 0.079 019 0.045 0.13 0.64 0.30 - 0.3 5 0.42 0.11 011 1.20 0.47 0.056 0.056 4.49 0.28 0.26 3.60 1.80 0.69 0.53
> 11.2 0.70 2.25 449 1.01 0.20 0.056 0.34 0.067 0.21 1.00 0.30 0.24 0.21 0.28 0.17 0.54 > 11.2 8.99 0.93 0.67
"BYG = Barnyardgrass, PSI = Prickly sida, VEL = Velvetleaf, MG = Morningglory.
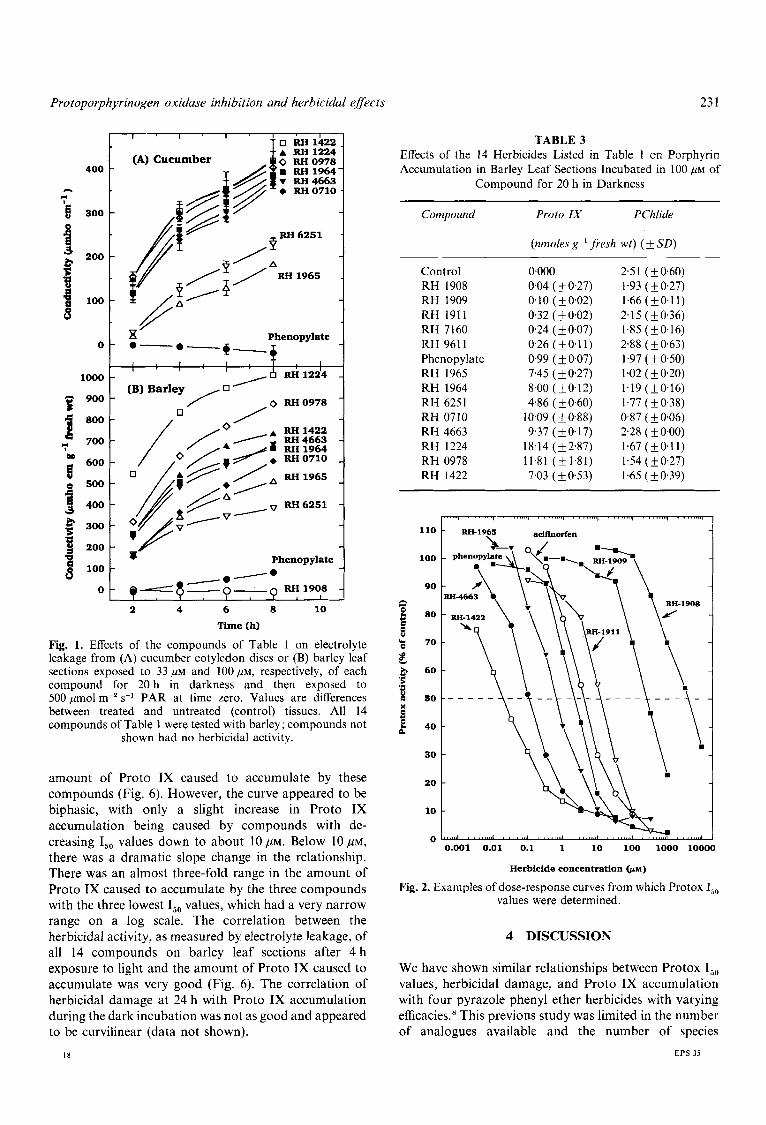
as a post-emergence than a pre-emergence treatment. The relative efficacies of the different compounds were similar for both pre- and post-emergence applications within a species. However, the relative efficacies varied somewhat between species. In cucumber cotyledon disc and barley leaf section studies, the relative efficacies were similar to those for whole plants (Fig. 1). Phenopylate and compounds with Protox I,, values higher than that of phenopylate were not herbicidal during the first 8 h of light exposure; however, some of these compounds were herbicidal after 24 h (data not shown).
3.2 Porphyrins
No Proto IX was found in untreated barley tissues after 20 h in darkness, and Proto IX was found in all tissues treated with any of the 14 compounds (Table 3). Proto IX accumulation ranged from 0.04 to 18.14 nmoles Proto IX g-' fresh weight for RH 1908 and RH 1224, re- spectively. PChlide content was unaffected or reduced by the herbicides. The highest levels of Proto IX caused to accumulate by any of the herbicides (c. 18 nmol g-l fresh wt with RH 1224) was about seven- to eight-fold the amount of PChlide found in untreated tissues. Concen- trations of Mg-Proto IX monomethyl ester and Mg- Proto IX were much lower than those of Proto IX and PChlide and were unaffected by the herbicides (data not shown).
3.3 Protox inhibition
All compounds inhibited Protox activity. Dose-response curves for inhibition of Protox activity by individual compounds generally ranged over about three orders of magnitude and were similar in shape to the dose- response curve of acifluorfen (Fig. 2) . Protox activity of the etioplast preparation used was almost completely inhibited by sufficiently high levels of all compounds. The I,, values of the compounds ranged from 21 nM to 4000 p~ (Table 1).
3.4 Correlation of Protox inhibition with herbicidal damage
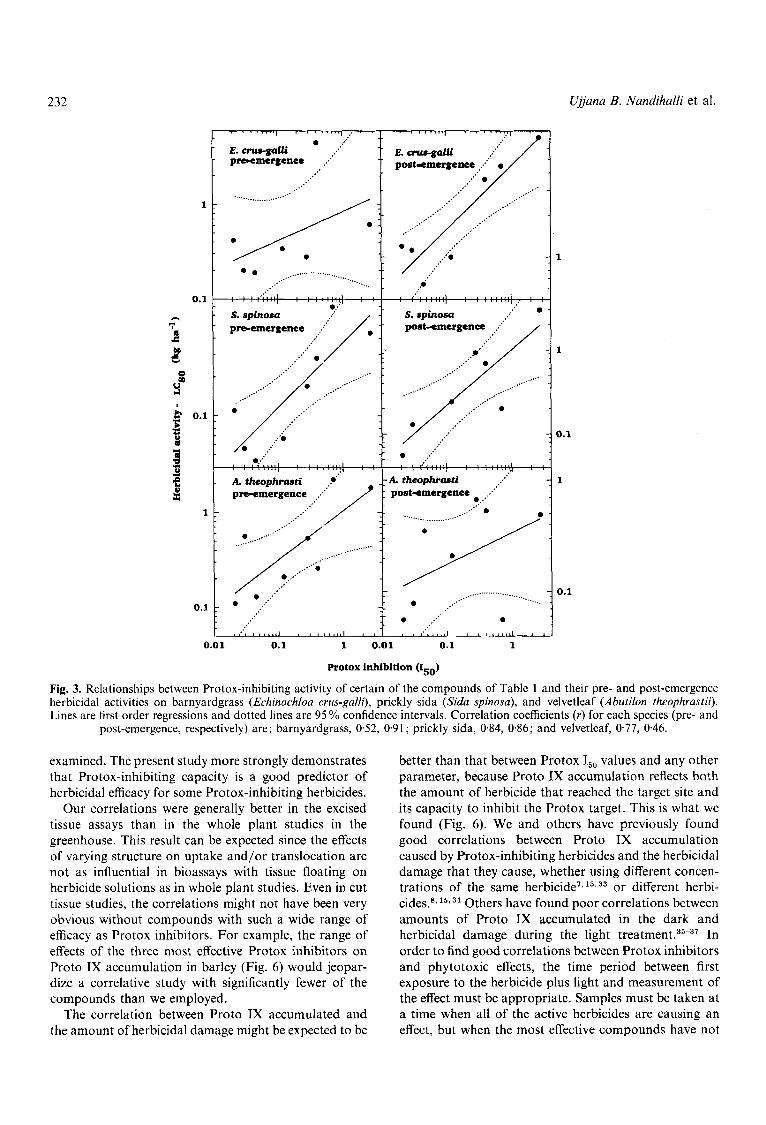
Linear regression plots of the log of Protox I,, values versus the log of LC,, values of the active compounds for various weed species are provided in Figs 3 & 4. Those compounds for which LC,, values were too high to be determined could not be included in these correlation studies. The correlation of I,, values with pre-emergence activity against morningglory is not shown because the values were greater than the highest possible extrapolated application rates (1 1.2 kg ha-l) for a number of the compounds. The degree of fit was better for some species than for others, and it was better with pre-emergence treatment than post-emergence with some species and vice uersa with others. The correlations of Protox I,, values and electrolyte leakage of cucumber cotyledon discs (Fig. 4) or cut leaf sections of barley (Fig. 5) of a broader array of compounds was better than the correlation with whole plant efficacy. Nevertheless, linear relationships with r values indicative of a greater than 95% chance of a correlation were obtained with both whole plants (Figs 3 & 4) and tissue pieces (Figs 4 & 5 ) for all species tested, except for the pre-emergence barnyardgrass treatments and the post-emergence velvet- leaf treatments. A separate regression was calculated for the effects of those compounds with high I,, values on barley after 4 h of light because these compounds did not meet the threshold requirements for herbicidal activity at this time (Fig. 5). In some cases, particular compounds fell outside the confidence intervals of linear relation- ships. However, the positive relationship between Protox-inhibiting capacity and herbicidal damage was clear.
3.5 Correlation of Proto IX accumulation with Protox inhibition and herbicidal damage
There was a negative correlation between the log of the I,,, values of the 11 most active compounds and the
Protoporphyrinogen oxidase inhibition and herbicidal effects 23 1
r - " ' l ' l ' l T 0 RH 1422
400 - - H
'I 300 -
200 1 F e 4 100 -
(A) Cucumber I
+ r RH 1224 o RHO^^ rn RH 1964 v RH4663 + RHO71C
z-6251 / RH1965
Phenopylate
I---4 0-
1000
'3 a P
600
500
400
300
M
B
'ff 200 jj 100
0
(B) Barley
RH 1422 R H 4663 RH 1964 RH 0710
RH 1965
RH 6251
Phenopylate
2 4 6 8 10
Time (h)
Fig. 1. Effects of the compounds of Table 1 on electrolyte leakage from (A) cucumber cotyledon discs or (B) barley leaf sections exposed to 33 L(M and 100 p ~ , respectively, of each compound for 20 h in darkness and then exposed to 500 pmol m-'s-' PAR at time zero. Values are differences between treated and untreated (control) tissues. All 14 compounds of Table 1 were tested with barley; compounds not
shown had no herbicidal activity.
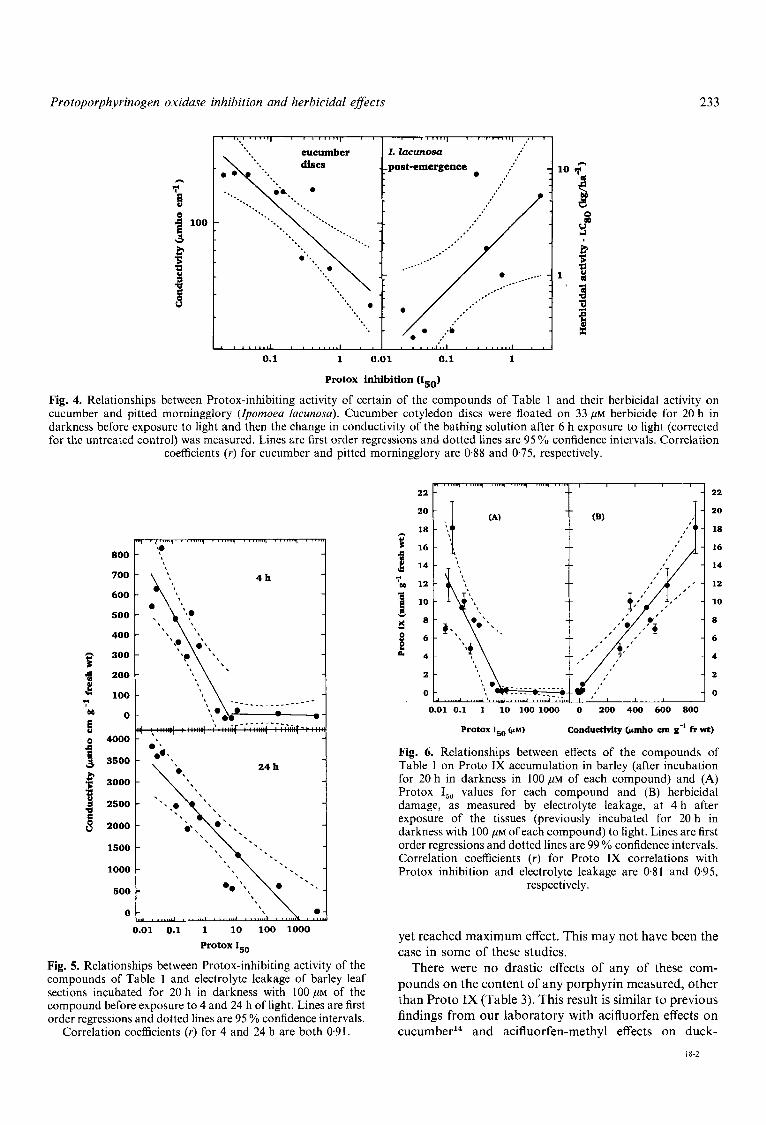
amount of Proto IX caused to accumulate by these compounds (Fig. 6) . However, the curve appeared to be biphasic, with only a slight increase in Proto IX accumulation being caused by compounds with de- creasing I,, values down to about 1 0 , ~ ~ . Below ~ O , U M , there was a dramatic slope change in the relationship. There was an almost three-fold range in the amount of Proto IX caused to accumulate by the three compounds with the three lowest I,, values, which had a very narrow range on a log scale. The correlation between the herbicidal activity, as measured by electrolyte leakage, of all 14 compounds on barley leaf sections after 4 h exposure to light and the amount of Proto IX caused to accumulate was very good (Fig. 6). The correlation of herbicidal damage at 24 h with Proto IX accumulation during the dark incubation was not as good and appeared to be curvilinear (data not shown).
18
TABLE 3 Effects of the 14 Herbicides Listed in Table 1 on Porphyrin Accumulation in Barley Leaf Sections Incubated in 100 C(M of
Compound for 20 h in Darkness ~ ~ ______- ~ _ _ _ _ _ _ ______
Compound Proto IX PChlide
(nmoles g-l fresh wt) (2 SD)
Control 0.000 2 3 1 ( + 0.60) 004 (k 0.27) 1.93 ( f 0.27) RH 1908 0.10 (+0.02) 1.66(+0.11) RH 1909 0.32 (2 0.02) 2.15 ( 5 0.36) RH 1911
RH 7160 0.24 (50.07) 1.85 (f0.16) 026 ( f 0.1 1) 2.88 (& 0.63) RH 9611
Phenopyla te 0.99 (k 0.07) 1.97 ( f 0.50) RH 1965 7.45 ( f 0.27) 1.02 ( k 0.20) RH 1964 8.00 (fO12) 1.19 (k0.16) RH 6251 4.86 (& 0.60) 1.77 ( f 0.38)
0.87 ( k 0.06) RH 0710 2.28 ( 5 0.00) RH 4663
RH 1224 8.14 (52.87) 1.67 (+O.ll) RH 0978 1.81 (+1.81) 1.54(+0.27)
1.65 ( f 0.39) RH 1422
0.09 ( f 0.88) 9.37 ( k 0.17)
7.03 (k 053)
100
90
ao
70
60
50
40
30
20
10
n " 0.001 0.01 0.1 1 10 100 1000 10000
Herbicide concentration &MI)
Fig. 2. Examples of dose-response curves from which Protox I,, values were determined.
4 DISCUSSION
We have shown similar relationships between Protox I,, values, herbicidal damage, and Proto IX accumulation with four pyrazole phenyl ether herbicides with varying efficacies.8 This previous study was limited in the number of analogues available and the number of species
EPS 35
232 Ujjana B. Nandihalli et al.
1
1
0.1
1
0.1
Protox inhibition (Iso)
Fig. 3. Relationships between Protox-inhibiting activity of certain of the compounds of Table 1 and their pre- and post-emergence herbicidal activities on barnyardgrass (Echinochloa crus-galli), prickly sida (Sida spinosa), and velvetleaf (Abutilon theophrastii). Lines are first order regressions and dotted lines are 95 % confidence intervals. Correlation coefficients ( r ) for each species (pre- and
post-emergence, respectively) are : barnyardgrass, 0.52, 0.91 ; prickly sida, 0.84, 0.86; and velvetleaf, 0.77, 0.46.
examined. The present study more strongly demonstrates that Protox-inhibiting capacity is a good predictor of herbicidal efficacy for some Protox-inhibiting herbicides.
Our correlations were generally better in the excised tissue assays than in the whole plant studies in the greenhouse. This result can be expected since the effects of varying structure on uptake and/or translocation are not as influential in bioassays with tissue floating on herbicide solutions as in whole plant studies. Even in cut tissue studies, the correlations might not have been very obvious without compounds with such a wide range of efficacy as Protox inhibitors. For example, the range of effects of the three most effective Protox inhibitors on Proto IX accumulation in barley (Fig. 6) would jeopar- dize a correlative study with significantly fewer of the compounds than we employed.
The correlation between Proto TX accumulated and the amount of herbicidal damage might be expected to be
better than that between Protox I,, values and any other parameter, because Proto IX accumulation reflects both the amount of herbicide that reached the target site and its capacity to inhibit the Protox target. This is what we found (Fig. 6). We and others have previously found good correlations between Proto IX accumulation caused by Protox-inhibiting herbicides and the herbicidal damage that they cause, whether using different concen- trations of the same herbicide'^'^.^^ or different herbi- cides.Ms 1 5 9 5 4 Others have found poor correlations between amounts of Proto IX accumulated in the dark and herbicidal damage during the light treatment.35-37 In order to find good correlations between Protox inhibitors and phytotoxic effects, the time period between first exposure to the herbicide plus light and measurement of the effect must be appropriate. Samples must be taken at a time when all of the active herbicides are causing an effect, but when the most effective compounds have not
Protoporphyrinogen oxidase inhibition and herbicidal effects 233
cucumber
I. 0
, I I . . . a 1 I I 3 I I I*.I I I t 1 1 1 . I , 1 1 1 1 1 1 I , 0.1 1 0.01 0.1 1
Protox inhibition (I50)
Fig. 4. Relationships between Protox-inhibiting activity of certain of the compounds of Table 1 and their herbicidal activity on cucumber and pitted morningglory (Zpomoea lacunosa). Cucumber cotyledon discs were floated on 33 ,UM herbicide for 20 h in darkness before exposure to light and then the change in conductivity of the bathing solution after 6 h exposure to light (corrected for the untreated control) was measured. Lines are first order regressions and dotted lines are 95 YO confidence intervals. Correlation
coefficients (Y) for cucumber and pitted morningglory are 0.88 and 0.75, respectively.
800
700
600
500
400
300
4 200
100
D
4000 i 3500
# 3000
3 2500
8 2000
1500
1000
500
0
8
1
1 '7 ,,..,..1 '".'.., "'"''7 . . , I . ' I I 22 22
20
18
16
14
M 12 c
1 10 v
E 8 s 6 e h 4
a 0
0.01 0.1 1 10 100 1000 0 200 400 600 800
PrOtOX bM) Conductivlty (umho cm g-' fr wt)
Fig. 6. Relationships between effects of the compounds of Table 1 on Proto IX accumulation in barley (after incubation for 20 h in darkness in 100 ,UM of each compound) and (A) Protox I,, values for each compound and (B) herbicidal damage, as measured by electrolyte leakage, at 4 h after exposure of the tissues (previously incubated for 20 h in darkness with 100 ,UM of each compound) to light. Lines are first order regressions and dotted lines are 99 % confidence intervals. Correlation coefficients ( r ) for Proto IX correlations with Protox inhibition and electrolyte leakage are 0.81 and 0.95,
respectively.
0.01 0.1 1 10 100 1000 Protox Is0
yet reached maximum effect. This may not have been the case in some of these studies.
There were no drastic effects of any of these com- Fig. 5. Relationships between Protox-inhibiting activity of the
sections incubated for 20 h in darkness with 100 ,UM of the
order regressions and dotted lines are 95 % confidence intervals.
compounds of Table 1 and electrolyte leakage of barley leaf pounds on the content of any porphyrin measured, other than Proto IX (Tab1e 3). This to previous compound before exposure to 4 and 24 h of light, Lines are first is findings from Our laboratory with acifluorfen effects on
Correlation coefficients (Y) for 4 and 24 h are both 0.91. cucumber14 and acifluorfen-methyl effects on duck-
18-2

234 Ujjunu B. Nundihulli et al.
weed.lg Others have sometimes found increases in PChlide levels in plants treated with Protox inhibi- t o r ~ , " ' . ~ ~ and we found significant increases in PChlide in cucumber, but not barley, treated with Protox- inhibiting pyrazole phenyl ethers.8 We have speculated previously, based on kinetic data, that some of these effects may be due to re-entry of Proto IX formed as a result of herbicide action into the porphyrin pathway.2" This effect may not occur in barley.
Over seven-fold more Proto IX molecules were de- tected in tissues treated with the most effective Protox inhibitor than PChlide molecules found in untreated tissues (Table 3). Since it is unlikely that synthesis of other porphyrins would constitute a significant pro- portion of porphyrin synthesis compared to PChlide, our results suggest that the porphyrin pathway is deregulated by strong inhibition of Protox, resulting in more carbon flow into the pathway than in untreated tissues. Haem is a feedback inhibitor of the porphyrin pathway3' and haem levels are reduced by Protox inhibitor^.^^
Chemical structure/activity relationship analyses with a herbicidal effect as the activity parameter are com- plicated by the many ways in which chemical structure can affect herbicidal activity. These include uptake, translocation, metabolic modification, sequestration, activity at the primary molecular site, and confounding effects of activity at other molecular sites. Thus, structure/activity relationships should be simpler when only activity a t the molecular site of interest is con- sidered. Several qualitative relationships between the chemical structures of the compounds that we have studied and their capacity to inhibit Protox can be concluded.
Manipulation of the phenyl ring dramatically affected Protox-inhibiting activity. Halogen substituents a t the 2- and 4-positions of the phenyl ring were generally required for good Protox inhibition (Table I) . However, in the case of R H 0710, a heterocyclic, substituted ring between the 4 and 5 carbons resulted in increased inhibitory capacity over that of phenopylate. A methyl- or isopropyl-esterified carboxyl group a t the 5 carbon gave better activity than a non-substituted 5 carbon. However, a propyne ether substitution a t the 5 carbon produced the best activity.
With the amine portion of the molecule, a pyrrolidine ring was more active than a piperidine ring (compare R H 6251 with R H 4663). Difluoro substitution of the 2 o r 3 carbon of the pyrrolidine or the 3 carbon of the piperidine ring significantly increased activity. Breakage of the pyrrolidine ring between the 2 and 3 carbons (compare R H 71 60 and R H 4663) reduced activity more than 200- fold. The two compounds ( R H 1422 and R H 6251) with unexpectedly low effects on Proto IX accumulation (Tables 1 and 3), considering their I,, values, had no structural features that might predict low in-vivo activity.
Quantitative structure/activity aspects of the com- pounds reported here, using molecular and quantum
mechanics approaches and comparative binding studies, will be analyzed in a subsequent paper.
ACKNOWLEDGEMENT
The authors thank Colin M. Tice of Rohm and Haas Co. for his invaluable assistance in providing chemicals, advice, and constructive criticism.
REFERENCES
1. Matringe, M., Camadro, J.-M., Labbe, P. & Scalla, R., FEBS Lett., 245 (1989) 35-8.
2. Matringe, M., Camadro, J.-M., Labbe, P. & Scalla, R., Biochem. J., 269 (1989) 231-5.
3. Witkowski, D. A. & Halling, B. P., Plant Physiol., 90 (1989) 123942.
4. Varsano, R., Matringe, M., Magnin, N., Mornet, R. & Scalla, R., FEBS Lett., 272 (1990) 1068.
5. Jacobs, J. M., Jacobs, N. J., Borotz, G. E. & Guerinot, M. L., Arch. Biochem. Biophys., 280 (1990) 369-75.
6 . Jacobs, J. M., Jacobs, N. J., Sherman, T. D. & Duke, S . O., Plant Physiol., 97 (1991) 197-203.
7. Sherman, T. D., Becerril, J. M., Matsumoto, H., Duke, M. V., Jacobs, J. M., Jacobs, N. J. & Duke, S. O., Plant Physiol., 97 (1991) 280-7.
8. Sherman, T. D., Duke, M. V., Clark, R. D., Sanders, E. F., Matsumoto, H. & Duke, S. O., Pestic. Biochem. Physiol., 40 (1991) 236-45.
9. Camadro, J.-M.. Matringe, M., Scalla, R. & Labbe, P., Biochem. J., 277 (1991) 17-21.
10. Matringe, M. & Scalla, R., Plant Physiol., 86 (1988) 16&72.
1 1 . Lydon, J. & Duke, S. O., Pestic. Biochem. Physiol., 31 (1988) 74-83.
12. Witkowski, D. A. & Halling, B. P., Plant Physiol., 87
13. Matringe, M. & Scalla, R., Pestic. Biochem. Physiol., 32
14. Becerril, J. M. & Duke, S. O., Pestic. Biochem. Physiol., 53
15. Becerril, J. M. & Duke, S. O., Plant Physiol., 90 (1989)
16. Nicolaus, B., Sandmann, G., Watanabe, H., Wakabayashi, K., & Boger, P., Pestic. Biochem. Physiol., 35 (1989)
17. Yanase, D. & Andoh, A,, Pestic. Biochem. Physiol., 35
18. Duke, S. O., Lydon, J. & Paul, R. N., Weed Sci., 37 (1989)
19. Matsumoto, H. & Duke, S. O., J . Agric. Food Chem., 38
20. Lehnen, L. P. Jr, Sherman, T. D., Becerril, J. M., & Duke, S. O., Pestic. Biochem. Physiol., 37 (1990) 23948.
21. Duke, S. O., Becerril, J. M., Sherman, T. D. & Matsumoto, H., Amer. Chem. Soc. Symp. Ser., 449 (1991) 371-86.
22. Duke, S. O., Becerril, J. M., Sherman, T. D., Lydon, J. & Matsumoto, H., Pestic. Sci., 30 (1990) 367-78.
23. Scalla, R., Matringe, M., Camadro, J.-M., & Labbe, P., Z . Naturforsch., 45c (1990) 503-1 1.
24. Deybach, J. C., de Verneuil, H., & Nordmann Y . , Hum. Genet. 58 (1981) 425-8.
25. Camadro, J. M., Urban-Grimal, D., & Labbe, P., Biochem. Biophys. Res. Commun., 106 (1982) 724-30.
26. Kishikawa, T. Japanese Patent 73 19, 626 (1973).
(1988) 632-7.
(1988) 162-72.
( 1 989) 1 19-26.
1 175-8 1.
192-20 1.
(1989) 70-80.
152-60.
(1 990) 2066-7 1.

Pro t oporph y r in ogen oxidase inhibit ion and herbic idaE efects 235
27. Takematsu, T., Konnai, M., Hosogai, T. & Nishida, T., US Patent 4, 521, 242 (1985).
28. Coe, P. L., Swithenbank, C. & Tice, C. M., 7th Znternat. Congr. Pestic. Chem., I(1990) 70.
29. Duke, S. 0. & Kenyon, W. H., Plant Physiol., 81 (1986)
30. Kenyon, W. H., Duke, S. O., & Vaughn, K. C., Pestic. Biochem. Physiol., 24 (1985) 240-50.
31. Bradford, M. M., Anal. Biochem., 72 (1976) 248-54. 32. Jacobs, N. J. & Jacobs, J. M., Enzyme, 28 (1982) 206-19. 33. Nandihalli, U. B., Liebl, R. A. & Rebeiz, C. A., Pestic.
882-8.
Sci., 31 (1991) 9-23.
34. Matringe, M., Clair, D. & Scalla, R., Pestic. Biochem. Physiol., 36 (1990) 300-7.
35. Sandmann, G. & Boger, P., Z . Naturforsch., 43c (1988)
36. Sandmann, G., Nicolaus, B. & Boger, P., 2. Naturforsch.,
37. Mayasich, J. M., Nandihalli, U. B., Liebl, R. A. & Rebeiz, C. A., Pestic. Biochem. Physiol., 36 (1990) 259-68.
38. Kouji, H., Masuda, T. & Matsunaka, S., Pestic. Biochem. Physiol., 33 (1989) 230-8.
39. Padmanaban, G., Ventkateswar, V. & Rangarajan, P. N., Trends Biochem. Sci., 14 (1990) 492-6.
699-704.
4 5 ~ (1990) 512-17.