correlaÇÃo entre caracterÍsticas de eficiÊncia …iz.sp.gov.br/pdfs/1440006053.pdf · direto,...
TRANSCRIPT

INSTITUTO DE ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO ANIMAL SUSTENTÁVEL
CORRELAÇÃO ENTRE CARACTERÍSTICAS DE EFICIÊNCIA
ALIMENTAR, DE CRESCIMENTO E DE CARCAÇA EM BOVINOS
NELORE
Thaís Matos Ceacero
Nova Odessa
Março – 2015


GOVERNO DO ESTADO DE SÃO PAULO
SECRETARIA DE AGRICULTURA E ABASTECIMENTO
AGÊNCIA PAULISTA DE TECNOLOGIA DOS AGRONEGÓCIOS
INSTITUTO DE ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO ANIMAL SUSTENTÁVEL
CORRELAÇÃO ENTRE CARACTERÍSTICAS DE EFICIÊNCIA
ALIMENTAR, DE CRESCIMENTO E DE CARCAÇA EM BOVINOS
NELORE
Thaís Matos Ceacero
Orientadora: Dra. Maria Eugênia Zerlotti Mercadante
Nova Odessa
Fevereiro - 2015
Dissertação apresentada ao
Programa de Pós-graduação do
Instituto de Zootecnia, APTA/SAA,
como parte dos requisitos para
obtenção do título de Mestre em
Produção Animal Sustentável.

Ficha catalográfica elaborada pelo
Núcleo de Informação e Documentação do Instituto de Zootecnia
Bibliotecária: Tatiane Helena Borges de Salles – CRB 8/8946
C387c Ceacero, Thaís Matos
Correlação entre características de eficiência alimentar, de crescimento e de
carcaça em bovinos Nelore / Thaís Matos Ceacero.
Nova Odessa, SP: [s.n.], 2015.
48 p.: il.
Dissertação (mestrado) – Instituto de Zootecnia. APTA/SAA,
Nova Odessa.
Orientadora: Dra. Maria Eugênia Zerlotti Mercadante
Bibliografia
1. Espessura de gordura. 2. Herdabilidade. 3. Parâmetros genéticos. I. Mercadante,
Maria Eugênia Zerlotti. II. Titulo.
CDD 636.213

GOVERNO DO ESTADO DE SÃO PAULO
SECRETARIA DA AGRICULTURA E ABASTECIMENTO
AGÊNCIA PAULISTA DE TECNOLOGIA DOS AGRONEGÓCIOS
INSTITUTO DE ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM PRODUÇÃO ANIMAL SUSTENTÁVEL
CERTIFICADO DE APROVAÇÃO
TÍTULO: CORRELAÇÃO ENTRE CARACTERÍSTICAS DE EFICIÊNCIA
ALIMENTAR, DE CRESCIMENTO E DE CARCAÇA EM BOVINOS NELORE
Autor: THAÍS MATOS CEACERO
Orientadora: Dra. Maria Eugênia Zerlotti Mercadante
Aprovado como parte das exigências para obtenção de título de MESTRE em Produção
Animal Sustentável, pela Comissão Examinadora:
Prof(a). Dr(a). Maria Eugênia Zerlotti Mercadante
(Orientadora)
Prof(a). Dr(a). Lenira El Faro
(Instituto de Zootecnia)
Dr. Marcos Eli Buzanskas
(Banca)
Data da realização: 24 de Fevereiro de 2015.
Presidente da Comissão Examinadora
Prof. Dr. Maria Eugênia Zerlotti Mercadante


Dedico:
Aos meus pais todos os bons exemplos transmitidos,
Pelas muitas palavras e estímulos e
Pelo incondicional amor e carinho


“Que a felicidade não dependa do
tempo, nem da paisagem, nem da
sorte, nem do dinheiro.
Que ela possa vir com toda
simplicidade, de dentro pra fora,
de cada um para todos”.
(Carlos Drummond de Andrade)


ix
CORRELAÇÃO ENTRE CARACTERÍSTICAS DE EFICIÊNCIA
ALIMENTAR, DE CRESCIMENTO E DE CARCAÇA EM BOVINOS NELORE
RESUMO: A seleção de bovinos de corte para a eficiência alimentar levará a redução
dos custos de produção, no entanto, ela não deve ter impactos negativos sobre o peso e a
qualidade da carcaça. O objetivo do presente estudo foi estimar correlações fenotípicas
(rf) e genéticas (rg) entre características de eficiência alimentar [eficiência alimentar
bruta (EAB), consumo alimentar residual (CAR), consumo alimentar residual ajustado
para espessura de gordura no lombo (CARegl) e para espessura de gordura no lombo e
na garupa (CAReg), ganho de peso residual (GANR), consumo e ganho residual (CGR),
consumo e ganho residual utilizando o CARegl (CGRegl) e o CAReg (CGReg)] e
características de interesse econômico como peso à seleção (PSEL), altura (ALT),
perímetro torácico (PTOR), área de olho de lombo (AOL), espessura de gordura no
lombo (EGL) e na garupa (EGG). O banco de dados continha 955 registros de consumo
de matéria seca (CMS), ganho médio diário (GMD) e peso vivo metabólico (PV0,75
)
obtidos pós desmame em 21 testes de desempenho de 83±15 dias. Para PSEL foram
utilizados 8078 registros de machos e fêmeas, ajustados para 378 dias e 550 dias de
idade, respectivamente. Para ALT, PTOR, AOL, EGL e EGG foram utilizados,
respectivamente, registros de 6548, 3876, 2283, 2285 e 1817 machos ao ano e fêmeas
ao sobreano. O arquivo de genealogia continha 8478 animais. Os componentes de
variância foram estimados pelo método da máxima verossimilhança restrita, com
modelo animal em análises bicaracterística. Os modelos para as características de
eficiência alimentar, CMS, GMD e PV0,75
incluíram efeitos aleatórios genéticos aditivos
direto, efeitos fixos de grupo de contemporâneo (GC), e idade da vaca (efeito linear e
quadrático) e do animal (efeito linear) como covariáveis. Para as demais características
os modelos incluíram efeitos aleatórios genético aditivo direto e de ambiente
permanente materno (somente para PSEL), além dos efeitos fixos de GC, mês de
nascimento do animal, e idade da vaca (efeitos linear e quadrático) e do animal (efeito
linear) como covariáveis. As estimativas de herdabilidade (h2) de CMS, GMD e PV
0,75
foram 0,45, 0,41e 0,53, respectivamente. As estimativas de h2 de EAB, CAR, CARegl,
CAReg, GANR, CGR, CGRegl e CGReg foram 0,14; 0,24; 0,20; 0,22; 0,19; 0,15; 0,11 e
0,11, respectivamente. Todas as rf entre as características estudadas foram próximas de
zero, exceto entre as características de eficiência alimentar com CMS e GMD. As rg
entre CMS, GMD e PV0,75
foram altas com as características de crescimento (maior que
0,61) e baixas a médias com as características de carcaça (entre 0,15 e 0,48). Dentre as
características de eficiência alimentar, o GANR foi o que apresentou maior rg com
PSEL, ALT e PTOR (0,34, 0,25 e 0,34, respectivamente) e menor rg com as espessuras
de gordura subcutânea (-0,17 e 0,18). As rg entre CAR, CARegl e CAReg com PSEL
foram desfavoráveis (0,17, 0,23 e 0,22) assim como com EGL (0,37, 0,33 e 0,33) e
EGG (0,30, 031 e 0,32). As rg entre EAB, CGR, CGRegl e CGReg e características de
crescimento foram baixas e favoráveis (0,07 a 0,22) e entre espessuras de gordura foram
médias e desfavoráveis (-0,22 a -0,45). A inclusão da espessura de gordura subcutânea
nos modelos de cálculo do CAR não diminuiu a rg entre essas características. A seleção
de animais mais eficientes na utilização de alimentos implicará em mudança genética
baixa ou nula para crescimento e negativa para espessura de gordura subcutânea
Palavras-chave: espessura de gordura, herdabilidade, parâmetros genéticos.


CORRELATION BETWEEN OF FEED EFFICIENCY, GROWTH AND
CARCASS TRAITS IN NELLORE CATTLE
Beef cattle selection for feed efficiency will lead to reduction of costs production,
however it should not have negative impacts on weight and quality of carcasses. This
study aimed to estimate genetic (rg) and phenotypic (rp) correlations among feed
efficiency traits [gross feed efficiency (G:F), residual feed intake (RFI), residual feed
intake adjusted for backfat thickness (RFIbf) and for backfat thickness and rump fat
thickness (RFIft), residual average daily gain (RG), residual intake and gain (RIG),
residual intake and gain using RFIbf (RIGbf) and RFIft (RIGft)] with selection weight
(SW), hip height (HH), girth circumference (GC), longissimus muscle area (LMA),
backfat (BFT) and rump fat thickness (RFT). Feed efficiency data set consisted of 955
records of dry matter intake (DMI), average daily gain (ADG) and metabolic body
weight (BW0.75
) obtained after post-weaning in 21 performance tests of 83±15 days
long. The SW data set consisted of 8078 records of males and females adjusted to 378
and 550 d of age, respectively. Additionally, records of 6548, 3876, 2283, 2285 and
1817 males and females were available for HH, GC, LMA, BFT and RFT, respectively.
Pedigree file contained 8478 animals. (Co)variance components were estimated through
two-trait animal models by restricted maximum likelihood. The feed efficiency traits
and DMI, ADG and BW0.75
models included random effects of direct additive genetic,
fixed effect of test group, linear effect of animal age in the middle of performance test,
linear and quadratic effects of dam age. For growth and carcass traits the models
included random effects of direct additive genetic, permanent environmental effects of
dam (only SW) and fixed effects of contemporary group and month of birth, linear
effect of animal age, linear and quadratic effects of dam age. The heritability (h2)
estimates for DMI, ADG and BW0,75
were 0.45; 0.41 and 0.53 respectively. Estimates
h2 of feed efficiency traits were 0.14, 0.24, 0.20, 0.22, 0.19, 0.15, 0.11 and 0.11,
respectively for G:F, RFI, RFIbf, RFIft, RG, RIG, RIGbf, RIGft. All rp among studied
traits were close to zero, excepting some of them between feed efficiency traits with
DMI and ADG. The rg among DMI, ADG and PV0.75
were strong and positive with
growth traits (higher than 0.61), whereas rg were weak to medium, also positive, with
carcass traits (0.15 to 0.48). Among feed efficiency traits, RG showed the highest and
positive rg with SW, HH and GC (0.34, 0.25 and 0.34, respectively) and the lowest rg
with subcutaneous fat thickness (-0.17 with BFT and 0.18 with RFT). The rg between
RFI, RFIbf and RFIft with SW were unfavorable (0.17, 0.23 and 0.22) as well as with
BFT (0.37, 0.33 and 0.33) and RFT (0.30, 031 and 0.32). The rg between G:F, RIG,
RIGbf and RIGft with growth traits were weak and favorable (0.07 to 0.22), but rg were
medium and unfavorable with subcutaneous fat thickness (-0.22 to -0.45). The inclusion
of BFT and RFT in the RFI calculation model did not decrease rg between them.
Selection for improved feed efficient in growing Nellore cattle is expected to decrease
carcass fat deposition without effect on growth traits
Keywords: fat thickness, genetic parameters, heritability, residual feed intake.


SUMÁRIO
RESUMO .................................................................................................................................. ix
ABSTRACT ............................................................................................................................. xi
1 INTRODUÇÃO ................................................................................................................... 15
2 REVISÃO DE LITERATURA ........................................................................................... 17
2.1 Medidas de eficiencia alimentar ..................................................................................... 17
2.2 Estimativa de herdabilidade de características de eficiência alimentar .......................... 18
2.3 Correlações entre características de eficiência alimentar e características de
crescimento ........................................................................................................................... 20
2.4 Correlações entre características de eficiência alimentar e características de
carcaça .................................................................................................................................. 21
3 MATERIAL E MÉTODOS ................................................................................................ 25
3.1 Características de eficiência alimentar ........................................................................... 25
3.2 Características de crescimento........................................................................................ 28
3.3 Características de carcaça ............................................................................................... 28
3.4 Análises estatísticas e genéticas ..................................................................................... 29
4 RESULTADOS E DISCUSSÃO ........................................................................................ 31
5 CONCLUSÃO ...................................................................................................................... 41
6 REFERÊNCIAS .................................................................................................................. 43


15
1 INTRODUÇÃO
O Brasil possui o segundo maior rebanho bovino, com aproximadamente 200
milhões de animais (MAPA, 2013). O clima tropical e a vasta extensão territorial
contribuem e permitem que a atividade seja exercida em todos os estados do território
brasileiro. Desse modo, o país é líder nas exportações de carne desde 2004, e
responsável pela produção de um quinto da carne bovina comercializada e vendida em
mais de 180 países (MAPA, 2013). Para que se alcançasse este patamar foram
necessários grandes avanços em técnicas de manejo e criação, nutrição e melhoramento
animal.
Os produtores de carne têm como principal objetivo maximizar a lucratividade
do sistema por meio do aprimoramento do produto final “outputs” e redução do uso de
insumos “inputs”. A alimentação dos animais é o maior contribuinte para a elevação dos
custos, apresentando influencia significativo sobre a lucratividade da produção.
Segundo Arthur e Herd (2008), a melhoria na utilização dos alimentos pelos animais em
todo o sistema pode contribuir para a redução dos custos. A busca pela otimização deve
depender não apenas de maior produção, mas também da eficiência alimentar do
animal. O aumento da produção de carne bovina por unidade de alimentação ao longo
de todo o sistema de produção deve resultar em benefícios econômico-ambientais
significativos.
Além da menor quantidade de alimento consumido, os animais mais eficientes
podem reduzir o impacto ambiental, uma vez que ao aproveitarem melhor os alimentos
a demanda por uso de terra para pastagens é menor, assim como a emissão de gases de
efeito estufa, como metano e gás carbônico (ALMEIDA, 2005).
Há diversas medidas de eficiência alimentar estudadas e aplicadas em sistemas
de produção, dentre elas, eficiência alimentar bruta (EAB) e sua inversa, a conversão
alimentar (CA), ambas calculadas por razão direta do ganho de peso diário (GMD) e
consumo de matéria seca (CMS). A desvantagem da utilização dessas características
como critério de seleção em programas de melhoramento é que as duas são
correlacionadas com ganho de peso e taxa de crescimento e, isso implicaria em animais
maiores, que resultariam em um maior consumo para a mantença (ARCHER et al.,
1999).
Um importante indicador de eficiência alimentar que foi proposto por Koch et al.
(1963) é o consumo alimentar residual (CAR), que vem sendo amplamente estudado.

16
Esta característica pode ser definida como a diferença do consumo observado e o
consumo estimado em função do peso vivo metabólico (PV0,75
) e do GMD. Com o
mesmo propósito, os autores sugeriram o ganho de peso residual (GANR), ajustando o
ganho de peso para o CMS e PV0,75
. Recentemente, Berry e Crowley (2012)
apresentaram outra característica indicadora de eficiência alimentar, denominada
consumo e ganho residual (CGR), que une CAR e GANR em uma só característica.
Estudos revelam que uma das ferramentas para a melhoria da eficiência
alimentar é a seleção para CAR, pois a partir dela é possível obter animais mais
eficientes, sem alterar o peso e ganho de peso. Esses animais ingerem menor quantidade
de alimento para obter o mesmo ganho de peso e peso corporal, indicando variação
genética na utilização de nutrientes (ARTHUR e HERD, 2008; CASTILHOS et al.,
2011).
A seleção deve ser baseada no equilíbrio entre diversas características
importantes para o sistema de produção. Em programas de melhoramento genético de
bovinos de corte no Brasil, um desafio é a escolha das características de eficiência
alimentar para serem implementadas como critério de seleção. As associações
fenotípicas e genéticas entre as características de eficiência alimentar e outras
características de interesse econômico em bovinos de corte, como características de
crescimento, carcaça, qualidade de carne e reprodutivas, devem ser bem conhecidas
para dar suporte para a definição desses critérios de seleção. Se houver antagonismos
entre as características, estes podem reduzir o benefício econômico alcançado pela
seleção para diminuição dos custos com alimentação animal.
Mais pesquisas são necessárias para estabelecer claramente as correlações
fenotípicas e genéticas das características de eficiência alimentar com características de
crescimento e carcaça, principalmente em animais Bos indicus, já que grande parte dos
resultados publicados é proveniente de Bos taurus criados em sistemas de produção
vigentes em países de clima temperado (DEL CLARO et al., 2012; BERRY e
CROWLEY, 2013).
Os objetivos desse estudo foram estimar as correlações fenotípicas e genéticas
de características de eficiência alimentar e características de crescimento e de carcaça
em bovinos da raça Nelore.

17
2 REVISÃO DE LITERATURA
2.1 Medidas de eficiência alimentar
Animais com melhor eficiência de conversão beneficiam de várias maneiras os
sistemas de produção, pois além de contribuírem com diminuição dos custos, com
menor consumo de matéria seca, contribuem também para diminuir o impacto ambiental
produzindo menor quantidade de dejetos e menor emissão de metano (NKRUMAH et
al., 2006; MOORE et al., 2008).
A eficiência da utilização do alimento não é uma característica mensurável
diretamente, mas em geral, calculada em função do consumo individual de alimento e
do ganho de peso corporal em determinado período de tempo. Alterações nas exigências
de mantença, composição do ganho e consumo alimentar podem variar de acordo com
intervalos de tempo, idades e diferentes pesos corporais. Essas alterações influenciam as
variações das características de eficiência alimentar e dificultam as comparações entre
os animais (CASTILHO et al., 2011).
Ao longo do tempo, várias maneiras de medir a eficiência alimentar foram
propostas, sendo as principais: conversão alimentar (CA), eficiência alimentar bruta
(EAB), eficiência parcial de crescimento (EPC) e o consumo alimentar residual (CAR).
Essas características são calculadas pelo consumo de matéria seca (CMS), que
representa o “input” e características de crescimento, como ganho médio diário (GMD)
e o peso vivo metabólico (PV0,75
), representando os “outputs” (BULLE et al., 2007;
ROLFE et al., 2011; GRION et al., 2014). A eficiência alimentar bruta (EAB) é
calculada pela razão entre o ganho de peso e o consumo de matéria seca e, portanto,
animais com EAB superior são considerados mais eficientes. Por ser uma razão direta
entre ganho de peso e consumo de matéria seca, esta característica, pode ocasionar
seleção de animais mais pesados, que não necessariamente apresentam menor consumo
de matéria seca, e o contrário, animais de crescimento lento com baixo consumo
alimentar, que também não são interessantes para a indústria da carne (ARTHUR et al.,
2001).
Atualmente, a característica de eficiência alimentar mais utilizada é o consumo
alimentar residual (CAR), sugerido por Koch et al. (1963). Essa característica é
independente da taxa de crescimento e do peso corporal e é definida pela diferença entre
o consumo real e o consumo predito, por meio de uma equação de regressão linear do
consumo individual real em função do peso vivo metabólico médio (PV0,75
) e ganho de

18
peso diário (GMD), sendo o resíduo dessa equação o valor do CAR. Animais
considerados eficientes na utilização de alimentos possuem CAR negativo (consumo
real menor que o consumo predito) e animais considerados ineficientes possuem CAR
positivo (consumo real maior que o consumo predito).
Com princípio semelhante ao utilizado na definição do CAR, Koch et al. (1963)
propuseram o ganho de peso ajustado para o consumo, conhecido como ganho de peso
residual (GANR), calculado como o resíduo da regressão linear do GMD em função do
PV0,75
e do consumo de matéria seca (CROWLEY et al., 2010). Neste caso, os fenótipos
desejados são os animais com GANR positivos, ou seja, aqueles que ganham mais peso
que o esperado para o PV0,75
e o CMS observados e, portanto, são mais eficientes que
seus contemporâneos (BERRY e CROWLEY, 2013).
Com a finalidade de melhorar a eficiência alimentar em bovinos, Berry e
Crowley (2012) propuseram recentemente a característica consumo e ganho residual
(CGR) como indicadora de eficiência alimentar, quando realizaram um estudo com
informações de 2605 bovinos puros da raça Angus, Charolês, Hereford, Limousin e
Simental, provenientes de testes de desempenho realizados na Irlanda. Esta
característica é calculada pela soma do CAR (resíduo da equação de ajuste do CMS) e
do GANR (resíduo da equação de ajuste do GMD), após inversão do sinal do CAR. O
CGR é usado para identificar animais que possuem o crescimento mais rápido e que
consomem menor quantidade de alimento por dia.
2.2 Estimativas de herdabilidade de características de eficiência alimentar
O objetivo do melhoramento genético é obter animais com genótipos superiores
por meio da seleção ou de cruzamento entre indivíduos geneticamente superiores para
determinado conjunto de características. Entretanto, o ambiente influencia na expressão
de grande número de caracteres, podendo mascarar os efeitos dos genótipos (PEREIRA,
2012).
Estimativas de herdabilidades para as características de eficiência alimentar são
mais frequentes para animais Bos taurus, em países de clima temperado (DEL CLARO
et al., 2012; BERRY e CROWLEY, 2013).
Para a característica conversão alimentar (CA), as estimativas de herdabilidade
variam de 0,17, como relatado por Herd e Bishop (2000), avaliando 542 machos da raça
Hereford na Escócia, até 0,65 como foi estimado por Shojo et al. (2005), avaliando

19
3250 machos Wagyu, consumindo alimento volumoso. Entretanto, as estimativas mais
relatadas na literatura para esta característica são de moderada magnitudes, em torno de
0,35 (SCHENKEL et al., 2004; MAO et al., 2013). Rolfe et al. (2011), utilizando dados
de 1141 bovinos cruzados (taurinos britânicos e continentais) nos Estados Unidos,
estimaram herdabilidade de 0,27 para a eficiência alimentar bruta.
Estimativas de herdabilidade do CAR, obtidos em estudos com animais de raças
taurinas, variam de moderadas a altas magnitudes, com valores de, 0,39; 0,18; 0,24;
0,47; 0,45 e 0,52, relatadas por ARTHUR et al., 2001; ROBINSON e ODDY et al.,
2004; HOQUE et al., 2006; LANCASTER et al., 2009; BOUQUET et al., 2010;
ROLFE et al., 2011, respectivamente. Poucos são os trabalhos que relatam estimativas
de herdabilidades em animais Bos indicus. Santana et al. (2014), em experimento com
1038 machos da raça Nelore, estimaram herdabilidade de 0,37 para CAR.
Schenkel et al. (2004), trabalhando com machos de raças taurinas em
crescimento (Charolês, Limousin, Angus, Simental, Hereford e Blonde d’Aquitaine),
reportaram que as características de eficiência alimentar devem ser utilizadas como
critério de seleção, porém devem ser verificadas as mudanças de características relativas
ao tamanho do animal e à espessura de gordura subcutânea. Os autores calcularam CAR
ajustando para a espessura de gordura no lombo (EGL) no modelo da regressão do
CMS, e estimaram herdabilidade de 0,39, valor bem próximo ao estimado para CAR
(0,38) sem a inclusão de EGL no modelo. Mao et al. (2013) também ajustaram CMS
para EGL e reportaram estimativas de herdabilidade de 0,47 e 0,64 para animais Angus
e Charolês, respectivamente.
Koch et al. (1963), propuseram o GANR, porém, essa característica só tem sido
utilizada em estudos recentes (CROWLEY et al., 2010; BERRY e CROWLEY, 2012,
2013). Devido a isso, existem poucas estimativas de herdabilidade para essa
característica na literatura, e estas foram apresentadas por Koch et al. (1963) (0,62) e
Crowley et al. (2010) (0,28) para animais taurinos e para zebuínos, Santana et al. (2014)
estimaram herdabilidade de 0,40. O GANR é altamente correlacionado com o ganho de
peso, e, sendo assim, pode ser confundido por sua correlação forte com outras
características de produção. Crowley et al. (2010) relataram correlação genética entre
GANR e CMS de -0,03, indicando que essa característica de eficiência alimentar é
independente geneticamente, do consumo dos animais e não é útil para identificar
diferenças no consumo de alimentos entre os indivíduos, que também é uma variável

20
determinante de lucro. Porém, os autores encontraram diferença significativa na média
(P<0,05) de CMS de animais alto GANR e baixo GANR.
Assim como o CAR e o GANR, o CGR apresenta herdabilidade moderada.
Berry e Crowley (2012) e Retallick (2013) relataram estimativas de herdabilidade de
0,36±0,06 e 0,22±0,10, respectivamente, para animais taurinos. Oliveira Júnior et al.
(2013) e Santana et al. (2014) relataram estimativas de herdabilidade de 0,62 e 0,54 para
CGR em animais da raça Nelore.
2.3 Correlações entre características de eficiência alimentar e características de
crescimento
A inclusão do CAR em programas de seleção depende de sua relação com outras
características de interesse econômico. Muitos estudos têm demonstrado que CAR e
ganho de peso possuem correlações próximas de zero, ou seja, que o CAR é
independente geneticamente de ganho de peso. Berry e Crowley (2013) realizaram
metanálise e obtiveram estimativa combinada de 0,02 para correlação genética entre
CAR e GMD, envolvendo 13 trabalhos científicos com animais taurinos. As estimativas
de Santana et al. (2014), com animais da raça Nelore, estão de acordo com as relatadas
na metanálise citada acima.
Entretanto, existem trabalhos que demonstram resultados contrários da maioria
dos trabalhos encontrados na literatura. Arthur et al. (2001) conduziram um trabalho
com registros de 1.180 touros Angus e novilhas envolvidos em teste de desempenho
com idade média de 268 dias no início do teste. As características estudadas incluíram o
peso aos 200 dias, peso aos 400 dias, e características de carcaça, e as correlações
genéticas entre CAR e os respectivos pesos foram de -0,45 e -0,26, respectivamente.
Bouquet et al. (2010), estimaram correlação genética entre CAR e peso ajustado aos 450
dias de idade de -0,40 para animais Blonde d'Aquitaine. De acordo com os autores, o
CAR pode ser uma característica de eficiência alimentar utilizada para o melhoramento
genético, pois está correlacionada com características de crescimento.
Apesar de o CAR ser independente das características de desenvolvimento
ponderal, poucos autores têm estudado as relações entre CAR e outras características
economicamente importantes. Existem trabalhos que correlacionam características de
eficiência alimentar com características relativas ao tamanho esquelético. Basarab et al.
(2003), Nkrumah et al. (2006), e Smith et al. (2010) não encontraram diferenças de

21
altura na garupa (ALT) em classes divergentes de CAR. Assim como os autores citados
acima, Schenkel et al. (2004), relataram que o CAR e CA são independentes
geneticamente da ALT. Kelly et al. (2011), em estudo com 2.605 touros relataram
correlação genética entre CAR e GANR com perímetro torácico (PTOR) de -0,11 e
0,23, respectivamente. Essas associações, no entanto, podem ser influenciadas pela raça,
idade do animal no teste de eficiência, e interação genótipo x tipo de dieta (BERRY e
CROWLEY, 2013).
De acordo com a maioria dos trabalhos, o CAR é independente do ganho de
peso, porém essa característica de eficiência alimentar apresenta correlações genéticas
altas com CMS, de acordo com a estimativa combinada (0,72) envolvendo 18 trabalhos
científicos (BERRY e CROWLEY, 2013).
Crowley et al. (2010) e Berry e Crowley (2012) relataram correlações genéticas
próximas de zero entre o GANR e CMS em animais de raças taurinas. O GANR foi
geneticamente (0,70) associado com taxas de crescimento mais rápidas, porém, não foi
possível detectar diferenças de consumo de alimentos entre as classes de GANR
(CROWLEY et al., 2010). Santana et al. (2014) estimaram correlações genéticas
negativas (-0,12) entre GANR e CMS em animais Nelore.
O CGR é fortemente correlacionado com CAR e GANR (-0,90 e 0,82,
respectivamente), já que ele é um índice que inclui essas duas características. CGR foi
altamente correlacionado com CMS (-0,87) e positivamente com GMD (0,20), ambas
correlações favoráveis. Portanto, a seleção dos animais com base em CGR, não
compromete o tamanho adulto do rebanho e diminui a quantidade de alimento
consumida, levando à maior lucratividade da produção (SANTANA et al., 2014).
2.4 Correlações entre características de eficiência alimentar e características de
carcaça
O aumento da gordura está relacionado com a concentração de leptina (JI et al.,
1997; MINTON et al., 1998). Este hormônio apresentou correlação genética positiva
(0,31) com CAR, ou seja, animais com CAR positivo apresentaram maior concentração
de leptina (RICHARDSON et al., 2004). Assim, animais menos eficientes apresentam
maior porcentagem de gordura na carcaça.
Alguns autores têm relatado associações entre CAR e composição corporal, com
intuito de avaliar se a seleção para CAR acarretará em mudanças genéticas

22
correlacionadas em outras características de produção. Segundo Richardson et al.
(2001), as correlações entre os valores genéticos estimados para CAR e para
características de carcaça obtidas por ultrassom, como espessura de gordura subcutânea
no lombo (EGL) e na garupa (EGG) e área de olho de lombo (AOL), sugerem que a
seleção baseada somente em CAR pode ocasionar em mudanças na composição
corporal e composição do ganho dos animais. De acordo com Basarab et al. (2003), a
variação nas características de carcaça medidas por ultrassonografia pode contribuir
com 9% da variação do CAR.
Alguns estudos com Bos taurus (BASARAB et al., 2003; LANCASTER et al.,
2009; MAO et al., 2013) indicam que o CAR é positivamente correlacionado com taxa
de deposição de gordura na carcaça, ou seja, animais mais eficientes depositam menos
gordura intramuscular. Berry e Crowley (2013) relataram estimativa de correlação
genética entre CAR e gordura de 0,20, estimada por meio de metanálises envolvendo 12
trabalhos científicos. Robinson e Oddy (2004) estimaram correlação genética alta e
positiva (0,72 ± 0,17) entre CAR e CA com EGG para animais mestiços de raças
temperadas e tropicais adaptados. Oliveira Júnior et al. (2013) estimaram correlação
positiva entre CAR e EGG (0,39) para animais Bos indicus. Por outro lado, existem
trabalhos presentes na literatura relatando a independência genética de CAR e medidas
de gordura subcutânea (SCHENKEL et al., 2004; BOUQUET et al., 2010; BAKER et
al., 2006; AHOLA et al., 2011; SANTANA et al., 2014).
Crowley et al. (2011), avaliando 2605 animais de seis raças taurinas distintas em
teste de desempenho na Irlanda, relataram correlação genética alta entre CAR e GANR
com EGL (0,39 e -0,44), e correlação nula entre essas com GANR e AOL. A partir
desses resultados, os autores recomendaram que a seleção para a melhoria da eficiência
alimentar tanto para o CAR quando para o GANR, resultariam em animais com carcaça
mais magra.
Diante das evidências da relação entre CAR e espessura de gordura, a
composição corporal tem sido incluída nos modelos de estimativa do CAR. Alguns
pesquisadores incluíram medidas de gordura (EGL, ganho de EGL) e AOL no modelo
para o cálculo do CAR (BASARAB et al., 2003; SCHENKEL et al., 2004; NKRUMAH
et al., 2007; KELLY et al., 2010; MAO et al., 2013). Quando as variáveis de carcaça
são incluídas no modelo, o ajuste tende a melhorar e as associações fenotípicas e
genéticas do CAR com a composição da carcaça diminuem. Schenkel et al. (2004)

23
estudaram CAR ajustado para EGL, e obtiveram correlações nulas (-0,01) entre CAR
ajustado com espessura de gordura, diferentemente da correlação de CAR (calculado
como a regressão do CMS em função do GMD e PV0,75
) com espessura de gordura
(0,16), em diferentes raças taurinas. Em contrapartida, Lancaster et al. (2009) e Mao et
al. (2013) obtiveram correlações genéticas similares tanto para CAR quanto para CAR
ajustado para EGL com espessura de gordura.
Mao et al. (2013) estimaram as correlações fenotípicas e genéticas do CAR,
CAR (ajustado para EGL) e CAR (ajustado para EGL e AOL) com características de
crescimento e de carcaça em 551 novilhos Angus e 417 novilhos Charolês. As
estimativas de correlação genética de CAR e CAR ajustado para EGL com AOL e EGL
foram similares. A inclusão da EGL no modelo para predizer o CMS explicou, em
média, variação de 0,5% do CMS em animais Angus e variação de 2,3% de CMS em
animais Charolês. A inclusão da EGL e AOL explicou apenas 0,7% da variância
adicional no CMS em Angus e 0,6% no Charolês.
Estudo realizado com animais da raça Nelore foi desenvolvido por Oliveira
Júnior (2013), em que foram avaliados 1058 animais machos nascidos entre 2005 e
2011. Os animais foram mantidos em sistema de confinamento em três diferentes tipos
de instalações: baias individuais, Calan Gates e GrowSafe, tendo em média 519 ± 168
dias de idade e 366 ± 59 kg de peso no início dos testes de desempenho. Foram
estimadas as correlações genéticas entre características de eficiência alimentar e
características de carcaça. As estimativas de correlação genética de CAR com AOL,
EGL e EGG foram -0,06, 0,10 e 0,39, respectivamente. Entre GANR com as mesmas
características de carcaça, as correlações genéticas foram 0,13, 0,13 e -0,09,
respectivamente, e entre CGR e as mesmas características de carcaça foram 0,08, -0,05
e -0,35. O CGR apresentou as correlações genéticas mais favoráveis com características
de carcaça dentre as medidas de eficiência alimentar.
Em um estudo com 147 machos castrados e 779 novilhos da raça Nelore,
Santana et al. (2014) relataram correlações genéticas entre CAR com AOL, EGL e EGG
de -0,38, 0,02 e 0,17, respectivamente, e correlações genéticas entre CGR com AOL,
EGL e EGG de 0,02, -0,03, 0,03, respectivamente. A seleção para CGR está
inteiramente relacionada com aumento no ganho de peso do animal, mas não com as
características de carcaça.

24
Berry e Crowley (2012) foram os primeiros a relatar correlações genéticas de
CGR com características de desempenho de novilhos Bos taurus, mas os autores não
apresentaram as associações entre CGR e características de carcaça. Há necessidade de
mais estudos com CGR, pois se existirem antagonismos entre CGR, características de
carcaça e de qualidade de carne, esses devem ser identificados, pois podem reduzir o
benefício econômico alcançado pela seleção de animais eficientes.

25
3 MATERIAL E MÉTODOS
As informações utilizadas nas análises foram provenientes de 8078 animais da
raça Nelore, nascidos de 1978 a 2013, filhos de 320 touros e 2078 matrizes,
pertencentes ao Centro APTA Bovinos de Corte, unidade de pesquisa do Instituto de
Zootecnia, localizada em Sertãozinho, Estado de São Paulo (21°10’ latitude sul e 47°
57’ longitude oeste), Brasil. O rebanho Nelore do referido centro de pesquisa é
composto de três linhas de seleção iniciadas em 1978, denominadas Nelore Seleção e
Nelore Tradicional, nas quais foi aplicada seleção baseada em maior diferencial do peso
pós desmame, e outra denominada controle (NeC), cuja seleção foi baseada em
diferencial em torno de zero (seleção estabilizadora) (MERCADANTE e RAZOOK,
2010). Neste contexto, os machos foram submetidos à seleção para peso ajustado aos
378 dias de idade (P378), obtido após prova de ganho de peso em confinamento de 168
dias, de acordo com a metodologia descrita por Razook et al. (1997), e as fêmeas foram
submetidas à seleção para peso ao sobreano ajustado aos 550 dias de idade (P550),
obtido em pastagem.
A seleção dos machos (P378) é realizada em idade anterior à das fêmeas (P550),
pois os mesmos permanecem em confinamento (prova de ganho de peso) após a
desmama, que coincide com o período de seca, e as fêmeas permanecem em pastagem
com ganhos relativamente baixos e muito variáveis. Portanto, o peso ao ano das fêmeas
não reflete o real potencial para crescimento, o que é obtido após o período das águas,
ao sobreano.
3.1 Características de eficiência alimentar
As características de eficiência alimentar foram obtidas para machos e fêmeas
nascidos de 2004 a 2013, filhos de 78 touros e 542 matrizes. O consumo de matéria seca
(CMS) e o ganho médio diário (GMD) foram obtidos em 21 testes de desempenho
(Tabela 1), após os animais serem desmamados em manejo de pastagem. Anteriormente
ao início dos testes, os animais foram submetidos a período de adaptação à dieta e
instalações, de pelo menos, 21 dias. Os animais permaneceram em baias individuais e
em dois piquetes coletivos com sistema automático de alimentação GrowSafe®
(GrowSafe Systems Ltd., Airdrie, Alberta, Canadá), com acesso ad libitum à dieta e
água. A dieta fornecida teve alterações ao longo dos anos quanto à inclusão de

26
alimentos, mas foram formuladas de modo a permanecerem equivalentes, em torno de
62% de nutrientes digestíveis totais e 13% de proteína bruta, conforme descrito por
Grion et al. (2014).
Tabela 1. Descrição dos grupos de teste de desempenho
Ano de teste Sexo1 n
Duração do
teste (dias)
Idade inicial,3 (dias) PV inicial,
3 (kg)
2005i F 32 56 263 ± 18 164 ± 28
2005i F 32 56 341 ± 18 233 ± 34
2006i F 32 70 267 ± 24 155 ± 30
2006i
F 32 70 350 ± 21 242 ± 48
2007i M 61 112 268 ± 14 205 ± 31
2008i M 62 112 249 ± 25 194 ± 36
2009i M 60 70 300 ± 21 295 ± 32
2009i F 32 85 272 ± 25 203 ± 28
2009i F 32 84 391 ± 22 292 ± 26
2010i M 60 73 262 ± 22 248 ± 40
2010i F 32 84 294 ± 19 212 ± 28
2010i F 24 84 281 ± 22 208 ± 25
2011i M 62 72 261 ± 23 243 ± 40
2011i F 31 84 294 ± 23 220 ± 33
2011i F 25 84 286 ± 30 212 ± 28
2012i M 23 91 276 ± 19 248 ± 32
2012c M 85 91 271 ± 22 240 ± 40
2012i F 21 86 326 ± 24 260 ± 29
2012i
F 30 86 323 ± 25 258 ± 29
2013c
M 128 86 283 ± 25 311 ± 48
2014c M 59 99 290 ± 20 295 ± 37
Geral – 955 83 ± 163 287 ± 38 237 ± 50
1 F= fêmeas; M= machos;
3 Média ± Desvio Padrão;
i= baias individuais e
c= baias coletivas.
Nas baias individuais a quantidade de alimento fornecida foi ajustada
diariamente de acordo com as sobras, para corresponderem a 5 a 10% do total oferecido.
Nas baias coletivas, os cochos foram mantidos sempre com alimento, e o consumo
diário de matéria natural individual, registrado automaticamente pelo software do
GrowSafe®, foi multiplicado pela matéria seca (MS) do alimento fornecido.
Amostragens semanais da dieta fornecida e do conteúdo das sobras foram analisadas em
amostras compostas por períodos a cada 28 dias para determinação da MS. Valores
diários de CMS foram excluídos quando não havia sobras nas baias individuais e
quando constatada taxa de desaparecimento de alimento sem causa ≤ 10% pelo sistema
GrowSafe®.
Os animais foram pesados a cada 14 dias após jejum de 12 horas (2005 a 2006),
a cada 28 dias com jejum para os machos (2007 e 2008) e fêmeas (2009 a 2011). De
2009 a 2012 os machos foram pesados semanalmente sem jejum, sendo três pesagens
por semana em dias consecutivos em 2009 e 2010, duas pesagens por semana em dias

27
consecutivos em 2011, e uma pesagem por semana em 2012. Em 2013 e 2014 esses
foram pesados sem jejum a cada 15 dias. Em 2012, as fêmeas foram pesadas dois dias
consecutivos a cada 15 dias.
O CMS foi obtido pela média de todos os dias de consumo válidos durante o
período de teste. O GMD foi estimado como o coeficiente de regressão linear dos pesos
em função dos dias em teste (DET):
yi = α + β * DETi + εi
em que yi é o peso do animal na iésima observação; α é o intercepto, que representa o
peso inicial; β é o coeficiente de regressão linear que representa o GMD; DETi
representa os dias em teste na iésima observação e εi é o erro aleatório associado a cada
observação. O peso vivo metabólico (PV0,75
) no meio do teste foi calculado como PV0,75
= [α + β * (DET/2)]0,75
.
A eficiência alimentar bruta (EAB) foi calculada como a razão entre GMD e
CMS. O consumo alimentar residual (CAR) foi estimado como o resíduo da equação de
regressão linear do CMS sobre o GMD e PV0,75
e, o ganho de peso residual (GANR) foi
estimado como o resíduo da equação de regressão do GMD sobre CMS e PV0,75
(Koch
et al., 1963), dentro de cada grupo contemporâneo, utilizando o programa SAS (SAS
Inst. Inc., Cary, NC). O consumo-ganho residual (CGR) foi calculado a partir do CAR e
GANR (Berry e Crowley, 2012).
Três modelos de regressão linear foram ajustados para estimar o CAR:
𝐶𝑀𝑆 = 𝛽1𝐺𝑀𝐷 + 𝛽2 𝑃𝑉0,75 + 𝜀 (1)
𝐶𝑀𝑆 = 𝛽1𝐺𝑀𝐷 + 𝛽2 𝑃𝑉0,75 + 𝛽3𝐸𝐺𝐿 + 𝜀 (2)
𝐶𝑀𝑆 = 𝛽1𝐺𝑀𝐷 + 𝛽2 𝑃𝑉0,75 + 𝛽3𝐸𝐺𝐿 + 𝛽4𝐸𝐺𝐺 + 𝜀 (3)
em que β1 é o coeficiente de regressão parcial sobre GMD; β2 é o coeficiente de
regressão parcial sobre PV0,75
, β3 é o coeficiente de regressão parcial da espessura de
gordura subcutânea no lombo (EGL), β4 é o coeficiente de regressão parcial da
espessura de gordura subcutânea na garupa (EGG), e o ε no modelo 1, 2 e 3 é o CAR,
CAR ajustado para EGL (CARegl) e CAR ajustado para EGL e EGG (CAReg),
respectivamente.
O modelo a seguir foi ajustado para estimar o GANR:
𝐺𝑀𝐷 = 𝛽1𝐶𝑀𝑆 + 𝛽2 𝑃𝑉0,75 + 𝜀

28
em que β1 e β2 são, respectivamente, os coeficientes de regressão parcial do CMS e
PV0,75
, e ε é GANR.
O CGR foi calculado, como: 𝐶𝐺𝑅 =𝐺𝐴𝑁𝑅
𝜎𝐺𝐴𝑁𝑅 −
𝐶𝐴𝑅
𝜎𝐶𝐴𝑅,
em que: 𝜎𝐺𝐴𝑁𝑅 é o desvio padrão da média do GANR do grupo contemporâneo e 𝜎𝐶𝐴𝑅 é
o desvio padrão da média do CAR do grupo contemporâneo. O consumo-ganho residual
utilizando o CARegl (CGRegl) e o CAReg (CGReg) também foram calculados,
considerando os desvios padrão da média do grupo contemporâneo das respectivas
características.
3.2 Características de crescimento
O peso à seleção (PSEL) foi considerado como os pesos padronizados P378
(machos) e P550 (fêmeas), analisados como uma única característica. A altura da garupa
(ALT), por ser também obtida em machos e fêmeas, respectivamente ao ano e ao
sobreano, foi analisada como uma única característica. O mesmo foi feito para o
perímetro torácico (PTOR). ALT foi obtida por meio de fitas métricas instaladas no
interior da balança e representada por uma vertical baixada da porção anterior do sacro
ao solo, e PTOR foi obtido pelo contorno do tórax, passando pelo cilhadouro e voltando
perpendicularmente à linha do dorso. PSEL foi obtido para animais nascidos de 1978 a
2013, ALT para animais nascidos de 1985 a 2013 e PTOR para animais nascidos de
1989 a 2013.
3.3 Característica de carcaça
As características de carcaça foram obtidas para animais nascidos de 1996 a
2002 em machos (com exceção dos anos 1998, 2000 e 2003), e em machos (ao ano) e
fêmeas (ao sobreano) nascidos de 2004 a 2013, utilizando ultrassonografia [Pie Medical
401347-Aquila, com sonda linear de 18cm, de 3,5 MHz (Esaote Europe B.V.)]. Para a
obtenção da área de olho de lombo (AOL) e EGL, o transdutor foi colocado
perpendicularmente à coluna vertebral, transversalmente sobre o músculo Longissimus
dorsi, entre a 12ª e 13ª costelas, do lado esquerdo do animal, utilizando-se um acoplador
acústico. Para a obtenção da imagem da EGG, o transdutor foi colocado na intersecção
dos músculos Gluteus medius e Biceps femoris, localizados entre o ílio e o ísquio
(PINHEIRO et al., 2012). Posteriormente, as imagens foram mensuradas utilizando o
programa Echo Image Viewer 1.0 (Pie Medical Equipament B.V., 1996).

29
3.4 Análises estatísticas
Foram retirados do banco de dados os animais que apresentaram mais de um
registro para as características estudadas fora do intervalo de ±3,5 desvios-padrão da
média do grupo contemporâneo (GC) (Tabela 2). Para as características de crescimento
e de carcaça, os GC foram definidos como ano de nascimento dos animais, rebanho e
sexo e para as características de eficiência alimentar o GC foi definido como ano,
instalação do teste e sexo.
Tabela 2. Número de animais com registro (N), média e desvio-padrão (DP), valores
mínimo e máximo, e número de grupos de contemporâneos (GC)
Característica1 N Média ± DP Mínimo Máximo Nº GC
CMS (kg.dia)
GMD (kg)
PV0,75
(kg)
EAB (kg ganho kg MS-1
)
955
955
955
955
6,83 ± 1,33
1,00 ± 0,26
68,15 ± 10,12
0,148 ± 0,033
2,15
0,17
37,40
0,04
12,63
1,71
94,60
0,27
21
21
21
21
CAR (kg.dia-1
) 955 -0,011 ± 0,60 -2,31 4,96 21
CARegl (kg.dia-1
) 894 -0,0091 ± 0,60 -2,36 4,96 21
CAReg (kg.dia-1
) 891 -0,0090 ± 0,59 -2,24 4,91 21
GANR (kg.dia-1
) 955 0,002 ± 0,12 -0,70 0,33 21
CGR 955 0,044 ± 1,71 -5,93 4,92 21
CGRegl 894 0,042 ± 1,70 -6,35 4,85 21
CGReg 891 0,043 ± 1,69 -5,94 4,80 21
PSEL (kg) 8078 299,70 ± 48,81 160,03 486,76 201
ALT (cm) 6548 132,26 ± 5,43 100 149 169
PTOR (cm) 3876 164,13 ± 8,72 128 192 100
AOL (cm2) 2283 51,43 ± 8,89 21,4 83,40 65
EGL (mm) 2285 1,75 ± 1,42 0 10,5 65
EGG (mm) 1817 5,08 ± 2,54 0 19,2 51
1CMS = consumo de matéria seca; GMD = ganho médio diário; PV
0,75 = peso vivo metabólico; EAB =
eficiência alimentar bruta; CAR = consumo alimentar residual; CARegl = consumo alimentar residual
ajustado para espessura de gordura no lombo; CAReg = consumo alimentar residual ajustado para
espessura de gordura; GANR = ganho de peso residual; CGR = consumo e ganho residual; CGRegl =
consumo e ganho residual utilizando CARegl; CGReg = consumo e ganho residual utilizando CAReg; PSEL
= peso à seleção; ALT = altura da garupa de machos e fêmeas; PTOR = perímetro torácico de machos e
fêmeas; AOL = área de olho de lombo; EGL = espessura de gordura no lombo; EGG = espessura de
gordura na garupa.

30
Para as características de eficiência alimentar e para CMS, GMD e PV0,75
foram
incluídos no modelo os efeitos aleatórios genéticos aditivos direto e os efeitos fixos de
GC e idade da vaca (efeito linear e quadrático) e do animal (efeito linear), como
covariáveis. Para as demais características (crescimento e carcaça) foram incluídos no
modelo os efeitos aleatórios genético aditivo direto e os efeitos de ambiente permanente
materno (somente para a característica PSEL), além dos efeitos fixos de GC e mês de
nascimento do animal, idade da vaca (efeito linear e quadrático) e do animal (efeito
linear) como covariáveis. Para todas as análises foi utilizado um arquivo de genealogia
contendo 8478 animais.
Os componentes de (co)variâncias, necessários para a estimação dos parâmetros
genéticos, foram estimados pelo método da máxima verossimilhança restrita, em
modelo animal bicaracterística, utilizando o programa WOMBAT (MEYER, 2007),
com critério de convergência de 10-9
. O modelo geral utilizado pode ser representado
matricialmente por:
𝒚 = 𝑿𝜷 + 𝒁𝜶 + 𝑾𝒄 + 𝜺
em que: y é o vetor das observações; βi é o vetor dos efeitos fixos associados à matriz de
incidência X; α é o vetor dos efeitos aleatórios genéticos aditivo direto associados à
matriz de incidência Z; c é o vetor dos efeitos de ambiente permanente materno
associados à matriz de incidência W, e ε é o vetor dos resíduos.
Neste estudo, foi assumido que:
E[y] = Xβ; Var(α) = A Sα; Var(c) = I Sc e Var(e) = I Se
em que: Sα = é a matriz de covariâncias genéticas aditivas diretas; Sc = é a matriz de
variâncias de ambiente permanente materno; Se = é a matriz de covariância residual; A
= matriz do numerador de relações genético aditivas; I = matriz identidade e =
produto direto entre matrizes. Foi assumido ainda que α, c, e são não correlacionados
entre si.

31
4 RESULTADOS E DISCUSSÃO
A duração média dos testes foi de 83 ± 16 dias (intervalo 56 a 112 dias) (Tabela
1). Os machos entraram no teste, em média, com 274 ± 26 (CV= 9%) dias de idade e
247 ± 48 (CV= 19%) kg de peso vivo, e fêmeas com 307 ± 44 (CV= 15%) dias de idade
e 221 ± 49 (CV= 22%) kg de peso vivo.
A estatística descritiva das características estudadas é apresentada na Tabela 2 e
os componentes de variância e estimativas de herdabilidade das características de
eficiência alimentar são apresentados na Tabela 3. As variâncias genéticas aditivas
estimadas para as características de eficiência alimentar nesse estudo estão de acordo
com as relatadas para animais zebuínos (BARWICK et al., 2009; SANTANA et al.,
2014). Em geral, as estimativas de herdabilidade estimadas em análises
unicaracterísticas foram maiores que as estimadas em análises multicaracterísticas.
Segundo Crews Jr. et al. (2010) pode haver essas diferenças em razão do maior número
de informações nas análises multicaracterísticas, pois as covariâncias entre as
características são levadas em consideração, proporcionando estimativas mais acuradas.
As estimativas de herdabilidade para as características de eficiência alimentar
foram de magnitude baixa (Tabela 3). A herdabilidade estimada para EAB foi inferior á
obtida para animais taurinos cruzados (0,27), estimados por Rolfe et al. (2011).
Durunna et al. (2011) estimaram herdabilidade de EAB de 0,14 e 0,40 para bovinos
Angus e Charolês cruzados em dietas de crescimento e terminação, respectivamente,
concordando com o valor estimado no presente trabalho para animais em dieta de
crescimento.
A estimativa de herdabilidade do CAR foi de baixa magnitude, quando
comparado valores relatados na literatura para Bos taurus, de 0,07 a 0,68 (FAN et al.,
1995; HERD e BISHOP; 2000; HOQUE et al., 2006; LANCASTER et al., 2009;
BERRY e CROWLEY, 2012; MAO et al., 2013; STEYN et al., 2014), e próxima à
estimativa combinada de 0,26 obtida por metanálise a partir de 21 estudos também em
Bos taurus (DEL CLARO et al., 2012). Para Bos indicus, Barwick et al. (2009) e
Santana et al. (2014) relataram estimativas de herdabilidade do CAR de 0,24 e 0,37,
respectivamente.

32
Tabela 3. Componentes de variância genética aditiva (σ²a), residual (σ²e) e herdabilidade
(h² ± erro-padrão) das características de eficiência alimentar, estimadas em
análises bicaracterísticas
Características Componentes de variância
σ2
a σ2
e h2 ± EP
EAB 0,06 0,38 0,14 ± 0,06
CAR 0,08 0,28 0,24 ± 0,06
CARegl 0,07 0,29 0,20 ± 0,06
CAReg 0,07 0,28 0,22 ± 0,06
GANR 0,003 0,01 0,19 ± 0,06
CGR 0,45 2,44 0,15 ± 0,05
CGRegl 0,34 2,53 0,11 ± 0,05
CGReg 0,31 2,51 0,11 ± 0,05
EAB = eficiência alimentar bruta; CAR = consumo alimentar residual; CARegl = consumo alimentar
residual ajustado para espessura de gordura no lombo; CAReg = consumo alimentar residual ajustado para
espessura de gordura; GANR = ganho de peso residual; CGR = consumo e ganho residual; CGRegl =
consumo e ganho residual utilizando CARegl; CGReg = consumo e ganho residual utilizando CAReg.
Modelos diferentes para cálculo do CAR podem contribuir para diferenças nas
estimativas de herdabilidade (ROLFE et al., 2011). As estimativas de herdabilidade do
CAR diminuíram com a inclusão das características de espessura de gordura no modelo
(CAR, CARegl e CAReg). Lancaster et al. (2009), em um estudo com fêmeas da raça
Brangus, e Mao et al. (2013), que estudaram machos da raça Charolês, calcularam CAR
ajustado para AOL e EGL e relataram estimativas de herdabilidade menores (0,42 e
0,60) que as observadas para CAR (0,47 e 0,68), diferentemente de Schenkel et al.
(2004) e Mao et al. (2013), que relataram estimativas similares para CAR e CAR
ajustado para AOL e EGL.
Apesar da característica GANR ter sido proposta concomitantemente ao CAR
por Koch et al. 1963, estimativas de parâmetros genéticos para essa característica só
foram relatadas nos últimos anos. Diferentemente do CAR, em que os animais mais
eficientes são aqueles com valores negativos, os animais mais eficientes para GANR
são os que têm valores positivos. A estimativa de herdabilidade para GANR foi inferior
à relatada por Koch et al. (1963) e Crowley et al. (2010) em animais taurinos (0,64 e
0,28, respectivamente). Oliveira Júnior et al. (2013) e Santana et al. (2014), com dados
de animais da raça Nelore, estimaram herdabilidades superiores (0,40 e 0,56,
respectivamente) à do presente estudo. O GANR é essencialmente uma característica de

33
crescimento, já que é o GMD ajustado para CMS e PV0,75
. Entretanto, a estimativa de
herdabilidade é substancialmente menor que a estimada para o GMD de 0,41 (Tabela 4),
possivelmente porque grande parte da variação genética do GMD é devida às diferenças
genéticas de CMS entre animais.
O CGR foi proposto recentemente por Berry e Crowley (2012). A justificativa
da proposta dessa característica, segundo os autores, é que a independência fenotípica
entre CAR e GMD é uma razão para os criadores não adotarem o CAR nos programas
de seleção, uma vez que animais com crescimento lento (característica indesejável)
podem ser classificados como eficientes (CAR negativo). Além disso, a independência
fenotípica entre GANR e CMS cria situação similar. Portanto, uma solução para isso é a
combinação de CAR e GANR, como o CGR, que garante a independência dessa
característica com o PV0,75
e correlação fenotípica negativa com CMS e correlação
fenotípica positiva com GMD (Berry e Crowley, 2013), seria o desejável. Portanto,
CGR reduz (mas não elimina) a probabilidade de um animal de crescimento lento ser
muito bem classificado com base em CGR (Berry e Crowley, 2013). Entretanto, essas
relações fenotípicas criadas por construção das características CAR, GANR e CGR, não
implicam em relações genéticas similares. Berry e Crowley (2012) relataram estimativa
de herdabilidade de CGR magnitude moderada (0,36) em animais taurinos, enquanto
que Oliveira Júnior (2013) e Santana et al. (2014) relataram estimativas de
herdabilidade de alta magnitude (0,54 e 0,62, respectivamente), em estudos com
animais da raça Nelore. Entretanto, a estimativa de herdabilidade do CGR obtida no
presente estudo foi bem abaixo, variando de 0,11 a 0,15 (Tabela 3).
Dentre as características de eficiência alimentar, o CAR foi o que apresentou a
maior herdabilidade (0,24), em análises bicaracterísticas. Portanto, a resposta à seleção
deve ser mais rápida para CAR se comparada com as outras características de eficiência
alimentar.
As estimativas de herdabilidade e as correlações genéticas e fenotípicas do
CMS, GMD, PV0,75
, características de crescimento (PSEL, ALT e PTOR) e
características de carcaça (AOL, EGL e EGG) estão apresentados na Tabela 4.
As três características principais (CMS, GMD e PV0,75
) que compõem as
características de eficiência alimentar apresentaram estimativas de herdabilidade de
maiores magnitudes em relação ás apresentadas na Tabela 3 para as características de
eficiência alimentar. Em geral, as estimativas de herdabilidade de CMS, GMD e PV0,75

34
estão próximas às estimativas combinadas de herdabilidade á partir de estimativas de
vários estudos, obtidas principalmente em Bos taurus (DEL CLARO et al., 2012;
BERRY e CROWLEY, 2013). Os valores estimados estão de acordo com os relatados
para CMS (0,40) e GMD (0,35) em animais da raça Nelore (SANTANA et al., 2014).
As estimativas de herdabilidade para as características de crescimento (PSEL,
ALT e PTOR) foram de moderadas a altas magnitudes (acima de 0,31). As estimativas
de herdabilidade para PSEL e ALT foram, respectivamente, ligeiramente superiores e
bem superiores às relatadas na literatura em animais da raça Nelore, de 0,42
(BOLIGON et al., 2010) e de 0,44 e 0,46 (BOLIGON et al., 2010; YOKOO et al.,
2010). Para PTOR a herdabilidade foi inferior à relatada por Simielli et al. (2014),
estimada para bovinos Nelore, de 0,44, e similar à relatada por Crowley et al. (2011),
estimada em seis raças taurinas, de 0,31.
As herdabilidades para as características de carcaça foram moderadas, dentre
elas, a AOL apresentou a maior estimativa (0,44), sendo assim, deve responder à
seleção mais rapidamente quando comparada às demais características de carcaça. A
estimativa de herdabilidade da AOL foi superior às relatadas em trabalhos recentes com
bovinos Nelore no Brasil, com valor de 0,29, 0,30 e 0,35 (YOKOO et al., 2010;
SIMIELLI et al., 2014; SANTANA et al., 2014), respectivamente. As estimativas de
herdabilidade para as duas espessuras de gordura (EGL e EGG) foram de média
magnitude, e inferiores às relatadas por Yokoo et al. (2010) e Simielli Filho et al.
(2014). Mao et al. (2013) relataram estimativa de herdabilidade de 0,35 para EGL em
animais da raça Angus.
O CMS foi altamente correlacionado com as características de crescimento
(Tabela 4). Altas correlações fenotípicas e genéticas entre o CMS e características de
crescimento já foram bastante descritas e discutidas na literatura em várias espécies de
animais domésticos, além de bovinos de corte (Arthur et al., 2001) Assim, o aumento da
taxa de crescimento e do tamanho corporal do animal acarretam, invariavelmente,
aumento no consumo de alimentos. A vantagem do CAR e do GANR está justamente
em eliminar ou reduzir a alta dependência do CMS com a taxa de crescimento e
tamanho corporal dos animais.
As correlações genéticas de CMS e PSEL foram de média magnitude com AOL,
e de baixa magnitude com EGL e EGG (Tabela 4). As correlações genéticas de CMS e
GMD com espessura de gordura estão de acordo com o relatado por Berry e Crowley

35
(2013), cujas estimativas combinadas foram de 0,28 para CMS (envolvendo 10
trabalhos científicos) e 0,09 para GMD (envolvendo 7 trabalhos científicos).
As estimativas de correlações genéticas e fenotípicas entre as medidas de
eficiência alimentar com as características de crescimento e de carcaça estão
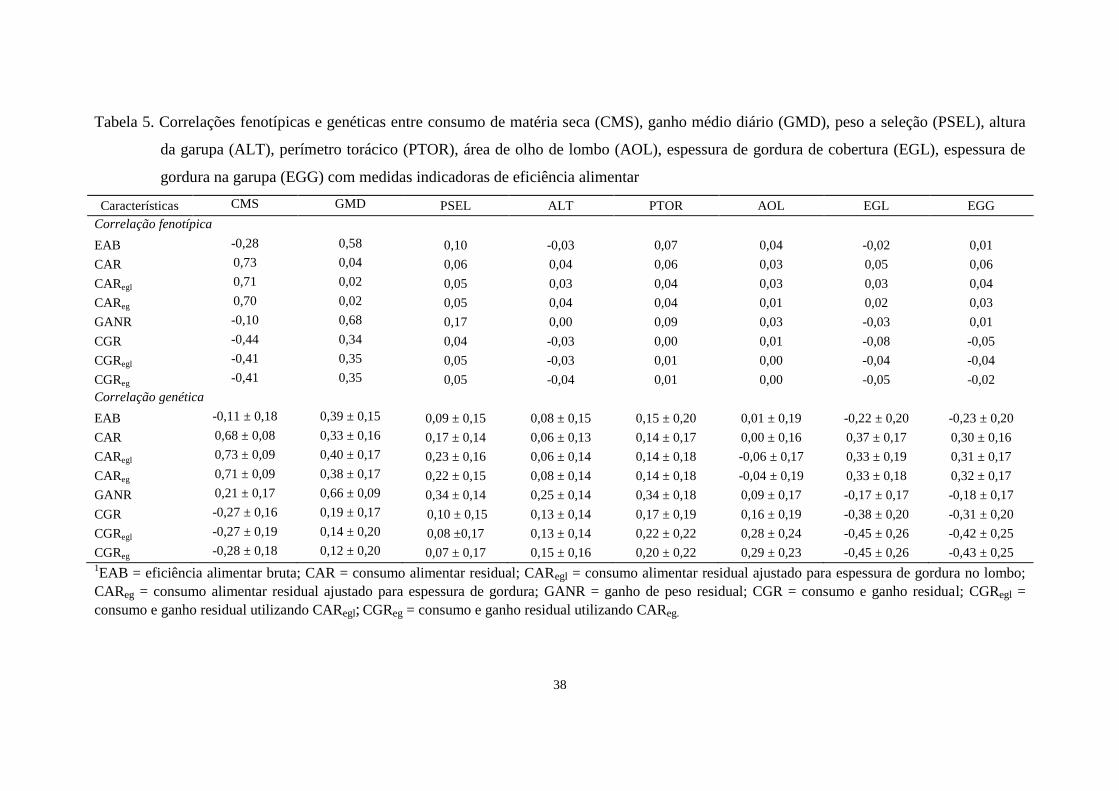
apresentadas na Tabela 5. Em geral, as correlações fenotípicas entre as características de
eficiência alimentar com as características de crescimento e de carcaça foram baixas e
próximas de zero. Hafla et al. (2012) estimaram correlação fenotípica entre EAB e EGL
de -0,03, valor igual ao estimado nesse estudo. Lancaster et al. (2009), em estudo com
novilhos Angus, relataram correlações fenotípicas baixas de CAR com AOL (-0,05) e
EGL (0,12) e de CAReg com AOL (0,01) e EGL (-0,05). Kelly et al. (2011) também
relataram correlações fenotípicas próximas de zero entre EAB e CAReg com EGL (-0,03
e 0,01) em animais de raças taurinas, semelhantes às estimadas nesse estudo.
As estimativas de correlação genética apresentaram altos erros-padrão devido ao
número de animais com registros de CMS, a característica mais difícil de ser obtida em
um estudo desse tipo. Desconsiderando o erro-padrão, as características de eficiência
alimentar apresentaram associação genética baixa com o PSEL, com exceção das três
definições de CAR (CAR, CARegl e CAReg) e de GANR, cujas correlações genéticas,
foram de sentido desfavorável para CAR e favorável para GANR.
A característica EAB apresentou correlações genéticas próximas de zero com
PSEL e ALT. Porém apresentou correlação baixa com AOL (0,18) e PTOR (0,15), e
correlação negativa com EGL (-0,22) e EGG (-0,23). Mao et al, (2013) encontraram
correlações genéticas semelhantes ao deste trabalho, de conversão alimentar (CA), o
contrário de EAB, com EGL (-0,18) em animais da raça Angus.
O CAR apresentou estimativas de correlações genéticas baixas com ALT, PTOR
e AOL indicando que não há associação entre essas características. Os resultados
diferem aos obtidos por Arthur et al. (2001), que estimaram correlações genéticas
moderadas entre o CAR e peso aos 200 dias (-0,45) e 400 dias (-0,26) de idade em

36
Tabela 4. Herdabilidade (diagonal), correlações fenotípicas (abaixo da diagonal) e genéticas (acima da diagonal) entre consumo de matéria seca
(CMS), ganho médio diário (GMD), peso vivo metabólico (PV0,75
), características de crescimento (PSEL, ALT e PTOR) e carcaça
(AOL, EGL e EGG)
CMS GMD PV0,75
PSEL ALT PTOR AOL EGL EGG
CMS 0,45 ± 0,05 0,83 ± 0,06 0,82 ± 0,05 0,8 ± 0,06 0,61 ± 0,07 0,79 ± 0,07 0,48 ± 0,10 0,29 ± 0,13 0,39 ± 0,11
GMD 0,56 0,41 ± 0,06 0,73 ± 0,08 0,84 ± 0,05 0,62 ± 0,08 0,80 ± 0,08 0,41 ± 0,11 0,15 ± 0,13 0,24 ± 0,12
PV0,75
0,65 0,42 0,53 ± 0,06 0,84 ± 0,05 0,80 ± 0,04 0,90 ± 0,03 0,63 ± 0,07 0,14 ± 0,12 0,30 ± 0,11
PSEL 0,60 0,61 0,83 0,40 ± 0,03 0,75 ± 0,02 0,87 ± 0,02 0,48 ± 0,06 -0,02 ± 0,10 0,11 ± 0,10
ALT 0,45 0,36 0,69 0,67 0,61 ± 0,02 0,64 ± 0,04 0,19 ± 0,07 -0,15 ± 0,09 -0,03 ± 0,09
PTOR 0,55 0,60 0,75 0,73 0,57 0,31 ± 0,03 0,38 ± 0,08 0,17 ± 0,11 0,29 ± 0,10
AOL 0,44 0,36 0,59 0,49 0,31 0,45 0,44 ± 0,05 0,09 ± 0,10 0,18 ± 0,10
EGL 0,23 0,15 0,29 0,22 0,65 0,28 0,20 0,30 ± 0,04 0,79 ± 0,05
EGG 0,25 0,21 0,29 0,26 0,07 0,30 0,25 0,61 0,38 ± 0,05

37
bovinos Angus avaliados em testes de desempenho na Austrália. Crowley et al. (2011)
estimaram correlação genética baixa, (-0,11) entre CAR e PTOR, o que difere do
resultado obtido no presente trabalho. A estimativa de correlação genética com ALT foi
próxima de zero (0,06). Segundo Arthur et al. (2001); Schenkel et al. (2004); Kelly et
al. (2011) e Steyn et al. (2014) a ALT não é correlacionada geneticamente com CAR.
As correlações genéticas estimadas de CAR com as medidas de gordura
subcutânea (Tabela 5) foram moderadas e indicam haver relação entre essas medidas,
porém no sentido antagônico. As estimativas de correlações genéticas estão próximas à
relatada por Berry e Crowley (2013) de 0,20, estimada por meio de metanálise
envolvendo 12 trabalhos científicos. Os resultados com animais da raça Nelore
apresentaram correlações genéticas baixas (0,02 e 0,17) para EGL e EGG,
respectivamente (SANTANA et al. 2014). Bouquet et al. (2010), em animais da raça
Limousin, encontraram correlações genéticas nulas entre CAR e escore de gordura na
carcaça, apontando independência entre essas características.
As estimativas de correlações genéticas de EGL e EGG com CARegl e CAReg,
não diferiram muito das estimativas destas com CAR. Observa-se que o fato das
características serem fenotipicamente independentes não garante baixas correlações
genéticas. Há trabalhos que relatam que animais mais eficientes depositam menos
gordura na carcaça, e a alternativa apontada pelos autores para diminuir esse efeito seria
a inclusão de uma medida de gordura subcutânea no modelo de cálculo do CAR. Desse
modo, a correlação dessas características com o CAR seria próxima de zero, e a seleção
com base no CAR não acarretaria em resposta correlacionada para deposição de gordura
na carcaça (BASARAB et al., 2003). Alguns pesquisadores incluíram medidas de
gordura no modelo de cálculo do CAR (RICHARDSON et al., 2001; BASARAB et al.,
2003; SCHENKEL et al., 2004; LANCASTER et al., 2009, KELLY et al., 2010; MAO
et al., 2013). Schenkel et al. (2004), relataram correlação genética nula (-0,01) entre
EGL e CAR quando esse foi ajustado para espessura de gordura, porém a correlação
entre CAR e AOL e CAReg e AOL apresentou leve aumento de -0,17 para -0,19. Mao et
al. (2013) também ajustaram o CAR para espessura de gordura e AOL, e observaram
leve diminuição das correlações genéticas entre CARegl com EGL, (0,17 para 0,12 e
0,33 para 0,19) tanto para animais Angus quanto para Charolês, respectivamente.
38
Tabela 5. Correlações fenotípicas e genéticas entre consumo de matéria seca (CMS), ganho médio diário (GMD), peso a seleção (PSEL), altura
da garupa (ALT), perímetro torácico (PTOR), área de olho de lombo (AOL), espessura de gordura de cobertura (EGL), espessura de
gordura na garupa (EGG) com medidas indicadoras de eficiência alimentar
Características CMS GMD PSEL ALT PTOR AOL EGL EGG
Correlação fenotípica
EAB -0,28 0,58 0,10 -0,03 0,07 0,04 -0,02 0,01
CAR 0,73 0,04 0,06 0,04 0,06 0,03 0,05 0,06
CARegl 0,71 0,02 0,05 0,03 0,04 0,03 0,03 0,04
CAReg 0,70 0,02 0,05 0,04 0,04 0,01 0,02 0,03
GANR -0,10 0,68 0,17 0,00 0,09 0,03 -0,03 0,01
CGR -0,44 0,34 0,04 -0,03 0,00 0,01 -0,08 -0,05
CGRegl -0,41 0,35 0,05 -0,03 0,01 0,00 -0,04 -0,04
CGReg -0,41 0,35 0,05 -0,04 0,01 0,00 -0,05 -0,02
Correlação genética
EAB -0,11 ± 0,18 0,39 ± 0,15 0,09 ± 0,15 0,08 ± 0,15 0,15 ± 0,20 0,01 ± 0,19 -0,22 ± 0,20 -0,23 ± 0,20
CAR 0,68 ± 0,08 0,33 ± 0,16 0,17 ± 0,14 0,06 ± 0,13 0,14 ± 0,17 0,00 ± 0,16 0,37 ± 0,17 0,30 ± 0,16
CARegl 0,73 ± 0,09 0,40 ± 0,17 0,23 ± 0,16 0,06 ± 0,14 0,14 ± 0,18 -0,06 ± 0,17 0,33 ± 0,19 0,31 ± 0,17
CAReg 0,71 ± 0,09 0,38 ± 0,17 0,22 ± 0,15 0,08 ± 0,14 0,14 ± 0,18 -0,04 ± 0,19 0,33 ± 0,18 0,32 ± 0,17
GANR 0,21 ± 0,17 0,66 ± 0,09 0,34 ± 0,14 0,25 ± 0,14 0,34 ± 0,18 0,09 ± 0,17 -0,17 ± 0,17 -0,18 ± 0,17
CGR -0,27 ± 0,16 0,19 ± 0,17 0,10 ± 0,15 0,13 ± 0,14 0,17 ± 0,19 0,16 ± 0,19 -0,38 ± 0,20 -0,31 ± 0,20
CGRegl -0,27 ± 0,19 0,14 ± 0,20 0,08 ±0,17 0,13 ± 0,14 0,22 ± 0,22 0,28 ± 0,24 -0,45 ± 0,26 -0,42 ± 0,25
CGReg -0,28 ± 0,18 0,12 ± 0,20 0,07 ± 0,17 0,15 ± 0,16 0,20 ± 0,22 0,29 ± 0,23 -0,45 ± 0,26 -0,43 ± 0,25
1EAB = eficiência alimentar bruta; CAR = consumo alimentar residual; CARegl = consumo alimentar residual ajustado para espessura de gordura no lombo;
CAReg = consumo alimentar residual ajustado para espessura de gordura; GANR = ganho de peso residual; CGR = consumo e ganho residual; CGRegl =
consumo e ganho residual utilizando CARegl; CGReg = consumo e ganho residual utilizando CAReg.

39
As estimativas de correlação genética entre CARegl e CAReg foram moderadas
com EGL e EGG (0,33 e 0,31; e 0,23 e 0,32, respectivamente), valores que estão de
acordo aos relatados por Mao et al. (2013).
As correlações genéticas entre GANR e CGR com as características de
crescimento e carcaça foram moderadas (Tabela 5). A estimativa de correlação genética
entre GANR e PTOR foi moderada e positiva, ou seja, seleção para maior GANR
acarretará em aumento no perímetro torácico. Esse resultado é semelhante ao
encontrado em animais taurinos (0,23), porém a correlação é moderada (CROWLEY et
al. 2011).
O GANR apresentou correlação genética baixa, enquanto o CGR correlação
genética moderada, ambas negativas, com EGL e EGG, ou seja, a seleção para animais
mais eficientes acarretará em mudanças no sentido contrário nas espessuras de gordura
subcutânea, isto é, diminuindo a gordura na carcaça do animal. Crowley et al. (2011)
estudando animais taurinos, estimaram correlação genética baixa entre GANR e AOL
(0,03), o que está de acordo com as estimativas obtidas no presente estudo, porém
correlação genética moderada e negativa com EGL (-0,44), valor superior ao estimado
nesse estudo.
Não existem pesquisas relacionando o impacto da seleção pelo CGR sobre as
características de crescimento, de carcaça e qualidade de carne em animais taurinos. Há
um estudo com animais da raça Nelore, o qual não corrobora os resultados obtidos no
presente estudo. Santana et al. (2014) relataram estimativas de correlação genética
próximas de zero entre CGR com AOL, EGL e EGG (0,02, -0,03 e 0,03,
respectivamente), indicando independência dessa medida de eficiência alimentar com a
deposição de gordura subcutânea.
A escolha da característica de eficiência alimentar para ser implementada como
critério de seleção deve ser baseada nas suas associações genéticas com as outras
características relevantes. Neste estudo com animais da raça Nelore, foi possível
observar que os animais mais eficientes apresentaram perda na qualidade da carcaça,
especificamente na deposição de gordura subcutânea. Entretanto, a espessura de gordura
foi medida durante o crescimento dos animais, fase em que há pouca deposição de
gordura subcutânea. Além disso, o ambiente/dieta não foi adequado à alta deposição de
gordura, como são as dietas de terminação. Portanto, a relação genética entre eficiência

40
alimentar e gordura subcutânea durante a fase pós-desmame, estimada no presente
trabalho, pode ser diferente da relação genética na fase de terminação.

41
5 CONCLUSÃO
As características de eficiência alimentar, crescimento e carcaça possuem
variabilidade genética suficiente para responder à seleção, entretanto, para as
características de eficiência alimentar o progresso genético será mais lento
comparativamente às demais características.
Há evidências de baixo antagonismo genético entre CAR e crescimento e médio
sinergismo genético entre GANR e crescimento.
A seleção de animais mais eficientes no consumo de alimentos implicará em
certa resposta correlacionada na espessura de gordura de cobertura no sentido contrário,
ou seja, há antagonismo de média magnitude entre o efeito dos genes para eficiência
alimentar e os genes para deposição de gordura na carcaça de bovinos Nelore.

42

43
REFERÊNCIAS
AHOLA, J.K.; SKOW, T.A.; HUNT, C.W.; HILL, R.A. Relationship between residual
feed intake and end product palatability in longissimus steaks form steers sired by
Angus bulls divergent for intramuscular fat expected progeny difference. The
Professional Animal Scientist, v.27, p.109-115, 2011.
ALMEIDA, R. Consumo e eficiência alimentar de bovinos em crescimento. 2005.
181 p. Tese (Doutorado em Ciência Animal e Pastagens) – Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2005.
ARCHER, J.A.; RICHARDSON, E.C.; HERD, R.M.; ARTHUR, P. Potential for
selection to improve efficiency of feed use in beef cattle: a review. Australian Journal
of Agricultural Science, v.50, p.147-161, 1999.
ARTHUR, P.F.; ARCHER, J.A.; JOHNSTON, D.J.; HERD, R.M.; RICHARDSON,
E.C.; PARNELL, P.F. Genetic and phenotypic variance and covariance components for
feed intake, feed efficiency, and other postweaning traits in Angus cattle. Journal of
Animal Science, v.79, p.2805-2811, 2001.
ARTHUR, P.F.; HERD, R.M. Residual feed intake in beef cattle. Revista Brasileira de
Zootecnia, v.37, suplemento especial, p.269-279, 2008.
BAKER, S.D.; SZASZ, J.I.; KLEIN, T.A.; KUBER, P.S. Residual feed intake of pubred
Angus steers: effects on meat quality and palatability. Journal of Animal Science,
v.84, p.38-945, 2006.
BARWICK, S.A.; WOLCOTT, M.L.; JOHNSTON, D.J.; BURROW, H.M.;
SULLIVAN, M.T. Genetics of steer daily and residual feed intake in two tropical beef
genotypes, and relationships among intake, body composition, growth and other post-
weaning measures. Animal Production Science, v.49, p.351-366, 2009.
BASARAB, J.A.; PRICE, M.A.; AALHUS, J.L.; OKINE, E.K.; SNELLING, W.M.;
LYLE, K.L. Residual feed intake and body composition in young growing cattle.
Canadian Journal of Animal Science, v.83, p.189-204, 2003.
BERRY, D.P.; CROWLEY, J. J. Residual intake and gain; a new measure of efficiency
in growing cattle. Journal of Animal Science, v.90, p.109-115, 2012.
BERRY, D.P.; CROWLEY, J.J. Genetics of feed efficiency in dairy and beef cattle.
Journal of Animal Science, v.91, p.1594-613, 2013.

44
BOLIGON, A.A.; MERCADANTE, M.E.Z.; FORNI, S.; LÔBO, R.B.;
ALBUQUERQUE, L.G. Covariance functions for body weight from birth to maturity in
Nellore cows. Journal of Animal Science, v.88, p.849–859, 2010.
BOUQUET, A.; FOUILLOUX, M.-N.; RENAND, G.; PHOCAS, F. Genetic parameters
for growth, muscularity, feed efficiency and carcass traits of young bulls. Livestock
Science, v.129, p.38-48, 2010.
BULLE, F.C.P.C.; PAULINO, P.V., SANCHES, A.C.; SAINZ, R.D. Growth, carcass
quality, and protein and energy metabolism in beef cattle with different growth
potentials and residual feed intakes. Journal of Animal Science, v.85, p.928–936,
2007.
CASTILHOS, A.M.; BRANCO, R.H.; RAZOOK, A.G.; BONILHA, S.F.M.;
MERCADANTE, M.E.Z.; FIGUEIREDO, L.A. Test post-weaning duration for
performance, feed intake and feed efficiency in Nellore cattle. Revista Brasileira de
Zootecnia, v.40, p.301-307, 2011.
CREWS, D.H. Jr; DIKEMAN, M.E.; NORTHCUTT, S.L.; GARRICK, D.J.;
MARSTON, T.T.; MACNEIL, M.D.; OLSON, L.W.; PASCHAL, J.C.; ROUSE, G.H.;
WEABER, R.; WHEELER, T.L.; SHACKELFORD, S.D.; WILLIAMS, R.E.;
WILSON, D.E. Animal Evaluation. In: CUNDIFF, L.V.; VAN VLECK, L.D.;
HOHENBOKEN, W.D. (Ed). Guidelines For Uniform Beef Improvement Programs.
Raleigh, NC: BIF, 2010. p. 16-55.
CROWLEY, J.J; McGEE, M.; KENNY, D.A.; CREWS, D.H. Jr; EVANS, R.D.;
BERRY; D.P. Phenotypic and genetic parameters for different measures of feed
efficiency in different breeds of Irish performance-tested beef bulls. Journal of Animal
Science, v.88, p.885-894, 2010.
CROWLEY, J J.; EVANS, R.D.; MC HUGH, N.; PABIOU, T.; KENNY, D.A.;
MCGEE JR, M.; CREWS, D.H. Jr; BERRY, D.P. Genetic associations between feed
efficiency measured in a performance test station and performance of growing cattle in
commercial beef herds. Journal of Animal Science, v.89, p.3382-3393, 2011.
DEL CLARO, A.C.; MERCADANTE, M.E.Z.; SILVA, J.A. II V. Meta-análise de
parâmetros genéticos relacionados ao consumo alimentar residual e as suas
características componentes em bovinos. Pesquisa Agropecuária Brasileira, v.47,
p.302-310, 2012.
DURUNNA, O.N.; WANG, Z.; BASARAB, J.A.; OKINE, E.K.; MOORE, S.S.
Phenotypic and genetic relationships among feeding behavior traits, feed intake, and

45
residual feed intake in steers fed grower and finisher diets. Journal of Animal Science,
v.89, p.3401-3409, 2011.
FAN, L.Q.; BAILEY, D.R.C.; SHANNON, N.H. Genetic parameter estimation of
postweaning gain, feed intake, and feed efficiency for Hereford and Angus bulls fed two
different diets. Journal of Animal Science, v.73, p.365-372, 1995.
GRION, A.L., MERCADANTE, M.E.Z., CYRILLO, J.N.S.G., BONILHA, S.F.M.,
MAGNANI, E., BRANCO, R.H. Selection for feed efficiency traits and correlated
genetic responses in feed intake and weight gain of Nellore cattle. Journal of Animal
Science, v.92, p.955–965, 2014.
HAFLA, A. N.; LANCASTER, P. A.; CARSTENS, G. E.; FORREST, D. W.; FOX, J.
T.; FORBES, T. D. A.; DAVIS, M. E.; RANDEL AND J. R. D.; HOLLOWAY, W.
Relationships between feed efficiency, scrotal circumference, and semen quality traits in
yearling bulls. Journal of Animal Science, v.90, p.3937-3944, 2012.
HERD, R. M.; BISHOP, S. C. Genetic variation in residual feed intake and its
association with other production traits in British Hereford cattle. Livestock
Production Science, v.63, p.111-119, 2000.
HOQUE, M.A.; ARTHUR, P.F.; HIRAMOTO, K.; OIKAWA, T. Genetic relationship
between different measures of feed efficiency and its component traits in Japanese
Black (Wagyu) bulls. Livestock Science, v.99, p.111-118, 2006.
JI, S.Q.; SCOTT, R.R.; SPURLOCK, M.E. Partial cloning of the bovine leptin gene and
its expression in adipose depots in cattle before and after finishing. Journal of Animal
Science, v.75, p.167-179, 1997.
KELLY, A.K.; McGEE, M.; CREWS, D.H. Jr; FAHEY, A.G.; WYLIE A.R.; KENNY,
D.A. Effect of divergence in residual feed intake on feeding behavior, blood metabolic
variables, and body composition traits in growing beef heifers. Journal of Animal
Science, v.88, p.109-123, 2010.
KELLY, A.K.; McGEE, M.; CREWS, D.H. Jr; LYNCH, C.O.; WUYLIE, A.R.;
EVANS, R.D.; KELLY, A.K. Relationship between body measurements, metabolic
hormones, metabolites and residual feed intake in performance tested pedigree beef
bulls. Livestock Science, v.135, p.8 -16, 2011.
KOCH, R.M., SWIGER, L.A., CHAMBERS, D., GREGORY, K.E. Efficiency of feed
use in beef cattle. Journal of Animal Science, v.22, p.486–494, 1963.

46
LANCASTER, P.A.; CARSTENS, G.E.; RIBEIRO, F.R.B.; TEDESCHI, L.O.;
CREWS, D.H. Jr Characterization of feed efficiency traits and relationships with
feeding behavior and ultrasound carcass characteristics in growing bulls. Journal of
Animal Science, v.87, p.1528-1539, 2009.
MAO, F.; CHEN, L.; VINSKY, M.; OKINE, E.; WANG, Z.; BASARAB, J.; CREWS,
D.H. Jr; LI, C. Phenotypic and genetic relationships of feed efficiency with growth
performance, ultrasound, and carcass merit traits in Angus and Charolais steers.
Journal of Animal Science, v.91, p.2067-2076, 2013.
BRASIL. MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO.
Disponível em: <http://www.agricultura.gov.br/>. Acesso em: 27 nov. 2014.
MERCADANTE, M.E.Z.; RAZOOK, A.G. Programa de Melhoramento Genético do
Instituto de Zootecnia. In: PIRES, A.V. (Org.). Bovinocultura de Corte. Piracicaba:
FEALQ, 2010, v. 2, p. 847-860.
MEYER, K. WOMBAT - A tool for mixed model analyses in quantitative genetics by
restricted maximum likelihood. Journal of Zhejiang University SCIENCE B. v.8, p.
815-821, 2007.
MINTON, J.E.; BINDEL, D.J.; DROUILLARD, J.S.; TITGEMEYER, E.C.;
GRIEGER, D.M. and HIIL, C.M. Serum leptin is associated with carcass traits in
finishing cattle. Journal of Animal Science, v.76, 1998. Suplemento, 1.
MOORE, S.S.; MUJIBI, F.D.; SHERMAN, E.L. Molecular basis for residual feed
intake in beef cattle. Journal of Animal Science, v. 87, p.41-47, 2008.
NKRUMAH, J.D.; OKINE, E.K.; MATHISON, G.W.; SCHMID, K.; LI, C.;
BASARAB, J. A.; PRICE, M.A.; WANG, Z.; MOORE S.S. Relationship of residual
feed intake with metabolic rate, methane production and energy partitioning in beef
cattle. Journal of Animal Science, v.84, p.145–153, 2006.
NKRUMAH, J.D.; BASARAB, J.A.; WANG, Z.; LI, C.; P RICE, M.A.; OKINE, E.K.;
CREWS, D.H. Jr; MOORE, S.S. Genetic and phenotypic relationships of feed intake
and different measures of feed efficiency with growth and carcass merit of beef cattle.
Journal of Animal Science, v.85, p.2711–2720, 2007.
OLIVEIRA JUNIOR, G.A.; SANTANA, M.H.A.; GOMES, R.C.; LEME, P.R.;
MATTOS, E.C.; FERRAZ, J.B.S. Parâmetros genéticos e correlações da ingestão e
eficiência alimentar em bovinos da raça Nelore. In: SIMPÓSIO DA SOCIEDADE

47
BRASILEIRA DE MELHORAMENTO ANIMAL, 5., 2013. Uberaba. Anais...
Uberaba: SBMA, 2013.
PEREIRA, J.C.C. Melhoramento genético aplicado à produção animal. 6.ed. Belo
Horizonte: FEPMVZ Editora, 2012, 758p.
PINHEIRO, T.R.; MERCADANTE, M.E.Z.; ALBUQUERQUE, L.G.; BONILHA,
S.F.M.; MONTEIRO, F.M. Selection for higher body weight in Nelore cattle is
effective in achieving an increase of longissimus muscle area without reducing
subcutaneous fat thickness. Revista Brasileira de Zootecnia, v.41, p.1426-1432, 2012.
RAZOOK, A.G.; FIGUEIREDO, L.A.; CYRILLO, J.N.S.G.; PACOLA, L.J.;
BONILHA NETO, L.M.; TROVO, J.B.F.; RUGGIERI, A.C.; MERCADANTE, M.E.Z.
Prova de Ganho de Peso: normas adotadas pela estação experimental de Zootecnia de
Sertãozinho: Instituto de Zootecnia, 1997. 33p. (Boletim Técnico, 40)
RETALLICK, K. Evaluation of feedlot feed efficiency relationships as well as
genetic and phenotypic performance, carcass, and economic outcomes. 2013. 102p.
Dissertation (PhD in Animal Sciences) University of Illinois, Urbana-Champaign, 2013.
Disponível em:<https://www.ideals.illinois.edu/handle/2142/42433>. Acesso em: 16
Ago. 2014.
RICHARDSON, E.C.; HERD, R.M.; ODDY, V.H.; THOMSPON, J.M.; ARCHER,
J.A.; ARTHUR, P.F. Body composition and implications for heat production of Angus
steer progeny of parents selected for and against residual feed intake. Australian
Journal of Experimental Agriculture, v.41, p.1065-1072, 2001.
RICHARDSON, E.C.; HERD, R.M.; ARCHER, J.A.; ARTHUR, P.F. Metabolic
differences in Angus steers divergently selected for residual feed intake. Australian
Journal of Experimental Agriculture, v.44, p.443-454, 2004.
ROBINSON, D.L.; ODDY, V.H. Genetic parameters for feed efficiency, fatness,
muscle area and feeding behavior of feedlot finished beef cattle. Livestock Production
Science, v.90, p.255-270, 2004.
ROLFE, K.M.; SNELLING, W.M.; NIELSEN, M.K.; FREETLY, H.C.; FERRELL,
C.L.; JENKINS, T.G. Genetic and phenotypic parameter estimates for feed intake and
other traits in growing beef cattle, and opportunities for selection. Journal of Animal
Science, v.89, p.3452-3459, 2011.

48
SANTANA, M.H.A.; OLIVEIRA JUNIOR, G.A.; GOMES, R.C.; SILVA, S.L.; LEME,
P.R.; STELLA, T.R.; MATTOS, E.C.; ROSSI JUNIOR, P.; BALDI, F.S.; ELER, J.P.;
FERRAZ, J.B.S. Genetic parameter estimates for feed efficiency and dry matter intake
and their association with growth and carcass traits in Nellore cattle. Livestock Science,
v.167, p.80-85, 2014.
SCHENKEL, F.S.; MILLER, S.P.; WILTON, J.W. Genetic parameters and breed
differences for feed efficiency, growth, and body composition traits of young beef bulls.
Canadian Journal of Animal Science, v.84, p.177-185, 2004.
SHOJO, M.; YOUNG, J.; ANADA, K.; OYAMA, K.; MUKAI, F. Estimation of
genetic parameters for growth and feed utilization traits in Japanese Black cattle.
Journal Animal Science, v.76, p.115-119, 2005.
SIMIELLI FILHO, E.A.; MERCADANTE, M.E.Z.; SILVA, J.A. II V.; JOSAHKIAN,
L.A. Ranking of Nellore animals in cattle championships: genetic parameters and
correlations with production traits. Genetics and Molecular Research, v.13, p.5722-
573, 2014.
SMITH, S.N.; DAVIS, M.E.; LOERCH, S.C. Residual feed intake of Angus beef cattle
divergently selected for feed conversion ratio. Livestock Science, v.132, p.41–47, 2010.
STEYN, Y.; VAN MARLE-KOSTER, E.; THERON, H.E. Residual feed intake as
selection tool in South African Bonsmara cattle. Livestock Science, v.164, p.35–38,
2014
YOKOO, M.J.; LOBÔ, R.B.; ARAUJO, F.R.C.; BEZERRA, L.A.F.; SAINZ, R.D.;
ALBUQUERQUE, L.G. Genetic associations between carcass traits measured by real-
time ultrasound and scrotal circumference and growth traits in Nelore cattle. Journal of
Animal Science, v.88, p.52–58, 2010.