cooperative cluster formation in metallothionein
TRANSCRIPT

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 250, No. 1, October, pp. 233-237, 1986
Cooperative Cluster Formation in Metallothionein’
JONATHAN BYRD AND DENNIS R. WINGE’
University of Utah, Medical Center, Salt Lake City, Utah 84132
Received May 1, 1986
An ion-exchange chromatography procedure was used to resolve apometallothionein from the metallo- form in a study of metal-thiolate cluster formation. Chromatography of metallothionein reconstituted with Cd(II), Zn(II), or Cu(1) at neutral pH on carboxy- methyl-cellulose led to removal of apoprotein from a solution without effect on recovery of the metalloprotein. Analysis of the effluent revealed apparent cooperative binding of these metal ions to the protein. Addition of l-4 mol eq Cd(I1) ions led to the recovery of metallothionein with around 4 mol eq Cd bound. The yield of this form increased with increasing starting metal ion equivalency. These results were obtained with two different ion-exchange resins. The cooperativity of binding was not total, but was initially confined to the carboxyl-terminal o( domain. The results of metal and protein yields are inconsistent with random, noninteractive binding. Similar data were obtained with Zn(I1) and Cu(1) ions although Cu(1) exhibited initial cooperative binding within the amino- terminal fi domain with over 5 mol eq Cu(1) bound. o 198s Academic PMS, IIK.
-
Metal ions are bound by mammalian metallothionein within two clusters en- folded by separate portions of the polypep- tide (1,2). Cluster B within the amino-ter- minal segment (residues l-29) is formed by nine cysteinyl thiolates ligating either three divalent ions such as Cd or Zn or six univalent ions such as Cu or Ag (3-5). Cluster A (resides 30-61) within the car- boxyl-terminal segment is formed by 11 cysteinyl thiolates ligating either four Cd(I1) or Zn(I1) ions or six Cu(1) ions (6, 7). Cd and Zn atoms bind with tetrahedral geometry whereas Cu atoms exhibit tri- gonal coordination (3, 8-10). Other ions that appear to bind to metallothionein in a fashion analogous to Cd(I1) include Bi(II), Co(II), Hg(II), Ni(II), Pb(II), In(III), and Sb(II1) (4, 8, 11-13).
These steps leading to the formation of the metal-thiolate clusters are not com- pletely understood. Using mild proteolysis
1 This work was supported by NIH Grants ES03817 and ES0014’7.
’ To whom correspondence should be addressed.
to “trap” intermediates we demonstrated that Cd(I1) and Cu(1) bind in an ordered fashion (5, 6). Addition of Cd(I1) to apo- metallothionein, followed by incubation with subtilisin, led to increased recoveries of the carboxyl-terminal a-domain peptide with 4 mol eq Cd(I1) bound (6). Following saturation of the 01 domain, Cd(I1) binding occurred within the P-domain cluster. Cu(1) binding within the two domains also oc- curred sequentially but with this metal initial binding occurred in the p domain (5). Cluster formation appeared to occur in a cooperative manner but no cooperativity was evident between the two clusters. One limitation of these studies was the possi- bility that the cooperative and ordered formation was induced by action of the protease. Although the observations were invariant with different protein concen- trations, an effect of the protease could not be rigorously dismissed. In this report metal binding to apometallothionein was studied using ion-exchange chromatogra- phy capable of resolving apo- from holo- metallothionein. No proteases were em-
233 0003-9861/86 $3.00 Copyright 0 1986 by Academic Press, Inc. All rights of reproduction in any form reserved.

234 BYRD AND WINGE
ployed. Results of these studies confirm our earlier findings that binding of Cd(II), Cu(I), and Zn(I1) occurs with apparent cooperativity.
MATERIALS AND METHODS
Metallothionein II purified from liver of rats in- jected with CdClz was used throughout in these studies (6). Metals were removed to prepare apometallothi- onein by gel filtration in 0.02 N HCl (4). The deme- tallized protein was deaerated and stored in the acid at -70°C. Apometallothionein was quantified by amino acid analysis and thiol titration using dithio- dipyridine (5).
Reconstitution of metallothionein was accomplished by mixing the apoprotein with an aliquot of a metal stock solution prepared in 0.1 N HCl and stepwise neutralization to pH 7 with Hepes3 base (Chelex fil- tered). The final Hepes concentration was about 30 mM. In experiments in which sodium dithionite was used, the final concentration of the reagent was 0.58 mM. The Cu(1) stock solution was prepared fresh daily from CuCl in thoroughly degassed 0.1 N HCl contain- ing 4% NaCl by weight. The entire reconstitution pro- cedure was performed in an anaerobic chamber.
Chromatography was carried out with either Chelex 100 or carboxymethyl-cellulose. In experiments uti- lizing Chelex 100, 250 ng protein reconstituted with varying mole equivalencies of Cd(I1) was mixed batchwise with 1 ml of a Chelex slurry (1 g/10 ml) that had been degassed. After a 25-min anaerobic in- cubation at 23°C the samples were centrifuged and metal and protein analyses were carried out on the supernatant. In experiments with carboxymethyl- cellulose as the separation resin, a column with a 3- ml bed volume was prepared and washed initially with 1 ml 50 mM EDTA followed by several bed volumes of 5 mM Hepes-Cl, pH ‘7.0 (Chelex filtered). After ap- plication of the sample, two 5-ml fractions were eluted with 5 mM Hepes-Cl followed by a 5-ml fraction with 0.1 N HCI as the eluant. Metal and protein analyses were performed on these elution fractions. Since the metalloprotein eluted in the first 5-ml fraction, data presented in the text are on this fraction.
Carboxymethylation of metallothionein was per- formed by incubating the protein at pH 7 with io- doacetate (1.2 mol/mol cysteinyl sulfur). The solution also contained 1.3 mM sodium dithionite. After 20 min at 23°C subtilisin was added to yield a metallothio- nien/protease weight ratio of 30. After 2 h at 37’C, the sample was chromatographed on a Sephadex G- 50 column, equilibrated, and eluted with 10 mM po- tassium phosphate, pH 7.
a Abbreviation used: Hepes, 4-(2-hydroxyethyl)-l- piperazineethanesulfonic acid.
RESULTS
The initial experiments were designed to determine conditions which would selec- tively remove apometallothionein from so- lution. The net ionic charge of rat liver metallothionein II can be estimated from amino acid sequence data. At neutral pH the net charge of the metal-free protein is +3, whereas the Cd, form is -3. Saturation of only the four metal sites in the (Y domain would yield a neutral molecule. Therefore, ion-exchange resins should be capable of resolving holo- from apometallothionein. Various exchange resins were evaluated; Chelex 100 and carboxymethyl-cellulose were found to remove 97 and 92%) respec- tively, of the apoprotein from solution without appreciably affecting the recovery of the fully metallo- form. Neither resin depleted the Cd(I1) content of the metal- saturated protein. Under these chromato- graphic conditions we attempted recovery of partially metal-saturated molecules. The prediction was that cooperative binding would result in only three states of the protein: apo-, Cdl-, and Cd7-metallothi- onein. Only the two metallo- forms should elute from the ion-exchange resin. Alter- natively, if binding were random and non- interactive, intermediate metallo- species with unsaturated (Y and p domains would be obtained. These intermediate species would have sufficient charge and would be retained by the resin.
Metallothionein reconstituted with in- creasing quantities of Cd(I1) was incubated with Chelex 100 in an anaerobic batch pro- cedure. The recovery of the protein in the supernatant increased linearly with rising equivalency (Fig. 1). Analysis of the su- pernatant for Cd content revealed that re- gardless of the starting equivalency, the resulting molecules contained at least 4 mol eq Cd (Fig. 1A). Specifically, when mole- cules were reconstituted with 0.7 g atom Cd(I1) per mole metallothionein there was a 12.5% recovery from Chelex of a molecule with 4.2 f 0.3 mol eq Cd (n = 4). If metals associate in clusters within metallothi- onein in a random fashion, the probability calculated from a binomial distribution that a molecule would be recovered with 4 mol eq metal when only 1 mol eq was re-
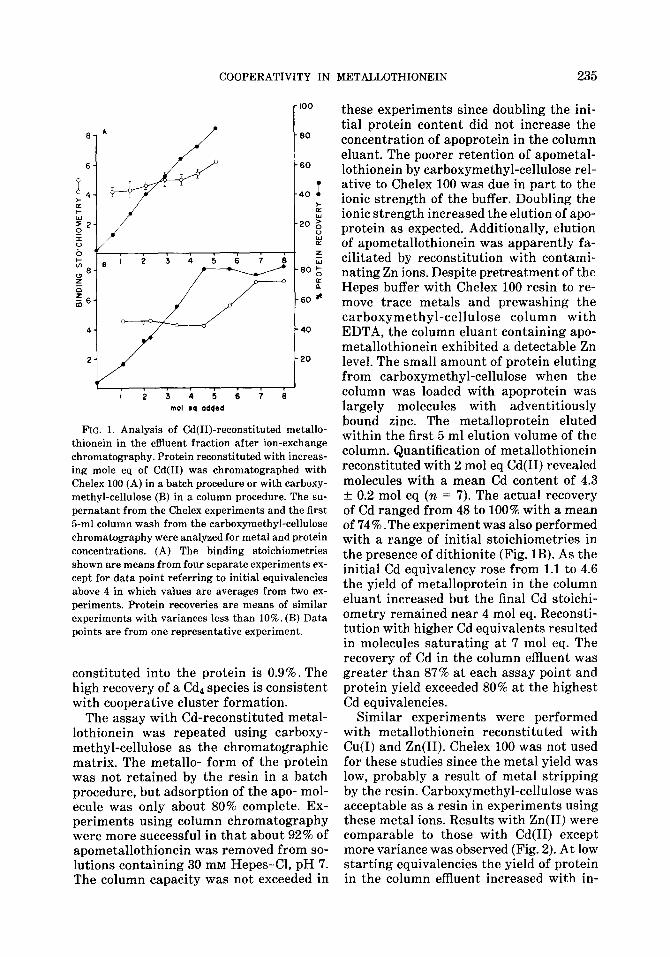
COOPERATIVITY IN METALLOTHIONEIN 235
mol eq added
FIG. 1. Analysis of Cd(II)-reconstituted metallo- thionein in the effluent fraction after ion-exchange chromatography. Protein reconstituted with increas- ing mole eq of Cd(I1) was chromatographed with Chelex 100 (A) in a batch procedure or with carboxy- methyl-cellulose (B) in a column procedure. The su- pernatant from the Chelex experiments and the first &ml column wash from the carboxymethyl-cellulose chromatography were analyzed for metal and protein concentrations. (A) The binding stoichiometries shown are means from four separate experiments ex- cept for data point referring to initial equivalencies above 4 in which values are averages from two ex- periments. Protein recoveries are means of similar experiments with variances less than 10%. (B) Data points are from one representative experiment.
constituted into the protein is 0.9%. The high recovery of a Cdl species is consistent with cooperative cluster formation.
The assay with Cd-reconstituted metal- lothionein was repeated using carboxy- methyl-cellulose as the chromatographic matrix. The metallo- form of the protein was not retained by the resin in a batch procedure, but adsorption of the apo- mol- ecule was only about 80% complete. Ex- periments using column chromatography were more successful in that about 92% of apometallothionein was removed from so- lutions containing 30 mM Hepes-Cl, pH 7. The column capacity was not exceeded in
these experiments since doubling the ini- tial protein content did not increase the concentration of apoprotein in the column eluant. The poorer retention of apometal- lothionein by carboxymethyl-cellulose rel- ative to Chelex 100 was due in part to the ionic strength of the buffer. Doubling the ionic strength increased the elution of apo- protein as expected. Additionally, elution of apometallothionein was apparently fa- cilitated by reconstitution with contami- nating Zn ions. Despite pretreatment of the Hepes buffer with Chelex 100 resin to re- move trace metals and prewashing the carboxymethyl-cellulose column with EDTA, the column eluant containing apo- metallothionein exhibited a detectable Zn level. The small amount of protein eluting from carboxymethyl-cellulose when the column was loaded with apoprotein was largely molecules with adventitiously bound zinc. The metalloprotein eluted within the first 5 ml elution volume of the column. Quantification of metallothionein reconstituted with 2 mol eq Cd(I1) revealed molecules with a mean Cd content of 4.3 f 0.2 mol eq (n = 7). The actual recovery of Cd ranged from 48 to 100% with a mean of 74% .The experiment was also performed with a range of initial stoichiometries in the presence of dithionite (Fig. 1B). As the initial Cd equivalency rose from 1.1 to 4.6 the yield of metalloprotein in the column eluant increased but the final Cd stoichi- ometry remained near 4 mol eq. Reconsti- tution with higher Cd equivalents resulted in molecules saturating at 7 mol eq. The recovery of Cd in the column effluent was greater than 87% at each assay point and protein yield exceeded 80% at the highest Cd equivalencies.
Similar experiments were performed with metallothionein reconstituted with Cu(1) and Zn(I1). Chelex 100 was not used for these studies since the metal yield was low, probably a result of metal stripping by the resin. Carboxymethyl-cellulose was acceptable as a resin in experiments using these metal ions. Results with Zn(I1) were comparable to those with Cd(I1) except more variance was observed (Fig. 2). At low starting equivalencies the yield of protein in the column effluent increased with in-
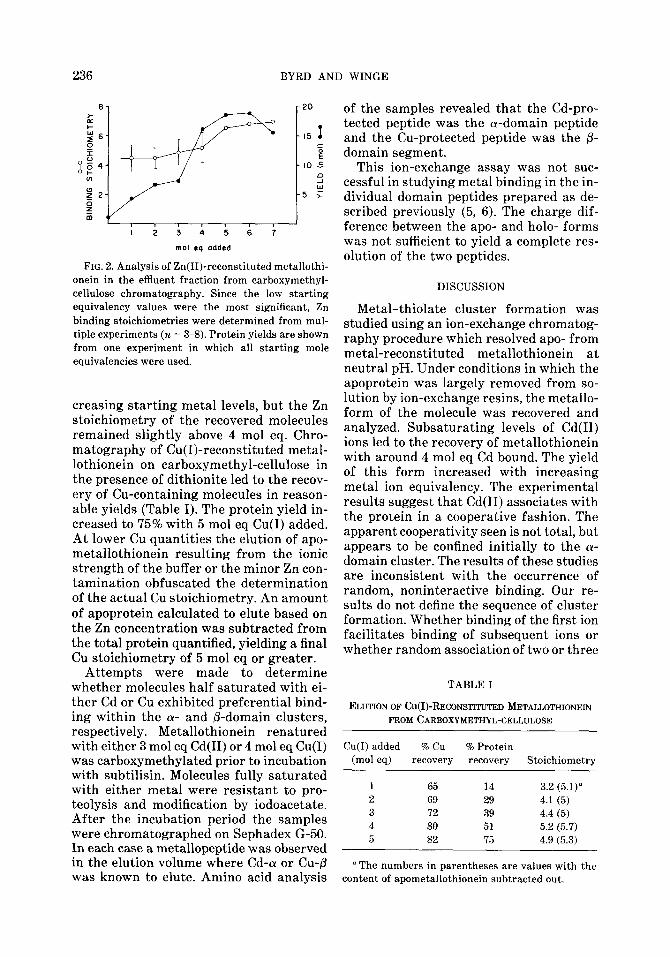
236 BYRD AND WINGE
-20
1234567 mol eq added
FIG. 2. Analysis of Zn(II)-reconstituted metallothi- onein in the effluent fraction from carboxymethyl- cellulose chromatography. Since the low starting equivalency values were the most significant, Zn binding stoichiometries were determined from mul- tiple experiments (n = 3-8). Protein yields are shown from one experiment in which all starting mole equivalencies were used.
creasing starting metal levels, but the Zn stoichiometry of the recovered molecules remained slightly above 4 mol eq. Chro- matography of Cu(I)-reconstituted metal- lothionein on carboxymethyl-cellulose in the presence of dithionite led to the recov- ery of Cu-containing molecules in reason- able yields (Table I). The protein yield in- creased to 75% with 5 mol eq Cu(1) added. At lower Cu quantities the elution of apo- metallothionein resulting from the ionic strength of the buffer or the minor Zn con- tamination obfuscated the determination of the actual Cu stoichiometry. An amount of apoprotein calculated to elute based on the Zn concentration was subtracted from the total protein quantified, yielding a final Cu stoichiometry of 5 mol eq or greater.
Attempts were made to determine whether molecules half saturated with ei- ther Cd or Cu exhibited preferential bind- ing within the (Y- and ,&domain clusters, respectively. Metallothionein renatured with either 3 mol eq Cd(H) or 4 mol eq Cu(1) was carboxymethylated prior to incubation with subtilisin. Molecules fully saturated with either metal were resistant to pro- teolysis and modification by iodoacetate. After the incubation period the samples were chromatographed on Sephadex G-50. In each case a metallopeptide was observed in the elution volume where Cd-cr or Cu-/I was known to elute. Amino acid analysis
of the samples revealed that the Cd-pro- tected peptide was the a-domain peptide and the Cu-protected peptide was the /3- domain segment.
This ion-exchange assay was not suc- cessful in studying metal binding in the in- dividual domain peptides prepared as de- scribed previously (5, 6). The charge dif- ference between the apo- and holo- forms was not sufficient to yield a complete res- olution of the two peptides.
DISCUSSION
Metal-thiolate cluster formation was studied using an ion-exchange chromatog- raphy procedure which resolved apo- from metal-reconstituted metallothionein at neutral pH. Under conditions in which the apoprotein was largely removed from so- lution by ion-exchange resins, the metallo- form of the molecule was recovered and analyzed. Subsaturating levels of Cd(I1) ions led to the recovery of metallothionein with around 4 mol eq Cd bound. The yield of this form increased with increasing metal ion equivalency. The experimental results suggest that Cd(I1) associates with the protein in a cooperative fashion. The apparent cooperativity seen is not total, but appears to be confined initially to the 01- domain cluster. The results of these studies are inconsistent with the occurrence of random, noninteractive binding. Our re- sults do not define the sequence of cluster formation. Whether binding of the first ion facilitates binding of subsequent ions or whether random association of two or three
TABLE I
ELUTIONOFCU(I)-RECONSTITUTED METALL~THIONEIN FROM CARBOXYMETH~L-CELLULOSE
Cu(1) added % cu % Protein Cm01 -4) recovery recovery Stoichiometry
1 65 14 3.2 (5.1)” 2 69 29 4.1 (5) 3 72 39 4.4 (5) 4 SO 51 5.2 (5.7) 5 82 75 4.9 (5.3)
o The numbers in parentheses are values with the content of apometallothionein subtracted out.

COOPERATIVITY IN METALLOTHIONEIN 237
Cd ions is necessary to create a structure favorable to binding of additional ions yielding a stable Cd4 LY cluster has not been determined.
The results of studies with Zn(I1) and Cu(1) ions were similar to those obtained in the Cd(I1) experiments although Cu(1) exhibits initial cooperative binding within the p domain. Experiments with all three metal ions confirm our earlier studies which used proteolysis to resolve apo- from holometallothionein (5, 6).
Bernhard et al. (14) suggested that at pH 8.6 there is no preferred binding with low Cd equivalencies, but preferred and coop- erative formation occurs in the a domain with Cd mole equivalencies between 3 and 5. We were unable to investigate a pH effect since the apoprotein is not retained by the exchange resin at pH 8.6. Binding of metal ions by protein with ionized cysteine thiol- ates may lead to subtle differences. The same investigators have also demonstrated that Co(I1) binding to metallothionein and to its a-domain peptide occurred noncoop- eratively (15, 16). Antiferromagnetic cou- pling of the Co(I1) ions was not evident un- til greater than 4 mol eq was bound to the intact protein (15). Cooperative binding would yield a diamagnetic species. We have obtained evidence in support of this con- clusion using both the ion-exchange and proteoylsis procedures but the difficulty of maintaining complete anaerobiasis com- plicated the studies (data not shown). It is unclear as to the reason for cooperative cluster formation with Cd(I1) and not Co(I1).
The significance of cooperative metal binding to metallothionein is unclear. If an unloading process is likewise cooperative it may be of importance in the yet unre- solved function of the protein. In addition to cooperative cluster formation there is a second enigma concerning the metal- thiolate clusters. The clusters have been demonstrated to be kinetically labile in that metal ions undergo facile exchange
between molecules (17). In the exchange of Cd(I1) ions with Zn-metallothionein the exchange occurs in a noncooperative and nonordered fashion. There is no selective occupancy of Cd ions within a particular domain (17). Once cluster formation occurs the final distribution of Cd and Zn ions ap- pears to be influenced by these exchange reactions and the stability of a mixed Cd, Zn cluster in the p domain.
REFERENCES
1. OTVOS, J. D., AND ARMITAGE, I. M. (1980) Proc. Natl. Acad. Ski. USA 77,7094-‘7098.
2. WINGE, D. R., AND MIKLOSSY, K.-A. (1982) J. Biol. Chem 257,3471-34’76.
3. FUREY, W. F., ROBBINS, A. H., CLANCY, L. L.,
WINGE, D. R., WANG, B. C., AND STOUT, C. D.
(1986) Science 231, 704-710. 4. NIELSON, K. B., ATKIN, C. L., AND WINGE, D. R.
(1985) J. Biol. Chem. 260,5342-5350. 5. NIELSON, K. B., AND WINGE, D. R. (1984) J. Biol.
Chem. 259,4941-4946. 6. NIELSON, K. B., AND WINGE, D. R. (1983) J. Biol.
Chem. 258,13063-13069. 7. BOLJLANGER, Y., ARMITAGE, I. M., MIKLOSSY, K. A.,
AND WINGE, D. R. (1982) J. Biol. Chem. 257, 13717-13719.
8. VASAK, M., AND KAGI, J. H. R. (1983) in Metal Ions in Biological Systems (Sigel, H., eds.) pp. 213- 273, Dekker, New York.
9. GEORGE, G. N., WINGE, D. R., STOUT, C. D., AND CRAMER, S. P. (1986) .I Inorg. Biochem. 27,213- 220.
10. GARNER, C. D., HASAIN, S. S., BREMNER, I., AND BORDAS, J. (1982) J. Inoqg. Biochem. 16, 253- 256.
11. VASAK, M., KAGI, J. H. R., AND HILL, H. A. 0.
(1981) Biochemistry 20,2852-2856. 12. BERNHARD, W., GOOD, M., VASAK, M., AND KAGI,
J. H. R. (1983) Inorg. Chim. Acta 79,154-155. 13. VASAK, M., KAGI, J. H. R., HOLMQUIST, B., AND
VALLEE, B. L. (1981) Bi~hemi&y20,6659-6664. 14. BERNHARD, W. R., VASAK, M., AND KAGI, J. H. R.
(1986) Experientia Suppl. in press. [Abstract] 15. VASAK, M., AND KAGI, J. H. R. (1981) Proc. Natl.
Acad. Sci. USA 78,6709-6713. 16. VASAK, M. (1986) Experierdia Suppl, in press.
[Abstract] 17. NETTESHEIM, D. G., ENGESETH, H. R., AND OTVOS,
J. D. (1985) Biochemistry 24,6744-6751.