cooperative breeding and human cognitive …faculty.bennington.edu/~sherman/sex/hrdy...
TRANSCRIPT

Cooperative Breeding and Human CognitiveEvolutionJ. M. BURKART, S. B. HRDY, AND C. P. VAN SCHAIK
Despite sharing a recent common ancestor, humans are surprisingly differentfrom other great apes. The most obvious discontinuities are related to our cogni-tive abilities, including language, but we also have a markedly different, coopera-tive breeding system. Among many nonhuman primates and mammals in gen-eral, cooperative breeding is accompanied by psychological changes leading togreater prosociality, which directly enhances performance in social cognition.Here we propose that these cognitive consequences of cooperative breedingcould have become more pervasive in the human lineage because the psycho-logical changes were added to an ape-level cognitive system capable of under-standing simple mental states, albeit mainly in competitive contexts. Once moreprosocial motivations were added, these cognitive abilities could also be usedfor cooperative purposes, including a willingness to share mental states, therebyenabling the emergence of shared intentionality. Shared intentionality has beenidentified as the original source of many uniquely human cognitive abilities,including cumulative culture and language. Shared intentionality rests on a fun-damentally prosocial disposition that is strikingly absent in chimpanzees, butpresent in cooperatively breeding primates. Thus, our hypothesis is that whilechimpanzees and perhaps all great apes exhibit many of the important cognitivepreconditions for uniquely human mental capacities to evolve, they lack the psy-chological preconditions. In humans, we argue, the two components merged,the cognitive component due to common descent from ape ancestors and themotivational component due to convergent evolution of traits typical of many co-operative breeders.
As recently as 6 to 7 million yearsago, the hominin lineage split off fromthe rest of the great ape (hominid) line-age1 and consequently shares many bi-ological traits and behavioral and cog-nitive similarities with great apes.2,3
Nevertheless, humans also exhibit re-markable differences from our closestrelatives. First, we not only live far lon-ger lives and reproduce at faster rates
than do the other great apes, but our
offspring take much longer to mature
and women cease reproduction well
before somatic senescence sets in.4 Sec-
ond, our ecological niche involves spe-
cialization on large, valuable food pack-
ages that have to be acquired together,
as well as shared, and mandatory reli-
ance on techniques acquired through
cumulative culture.5 This niche and
our social relationships are based on
an un-apelike selflessness, a degree of
hypersociality reflected in a concern for
others, eagerness to share food and in-
formation with others, and cooperation
in a wide array of contexts, even with
nonrelatives and near-strangers.5–8 Ourmode of life facilitates our spread intonew habitats, resulting in a ubiquitousgeographic distribution.9 Third, withregard to intellectual performance,humans differ from the other greatapes, which, as a group, show relativelyhomogeneous cognitive abilities.10–12
Whenever a species exhibits multi-ple derived traits, it may be helpful tobegin by asking if these are causallyconnected. Many anthropologistshave argued that humankind’s pecu-liar package of traits coevolved withthe rise of the genus Homo around 2Ma,5,13 which supports the possibilityof a causal connection. The coopera-tive breeding hypothesis claims thatthe emergence in the genus Homo ofallomaternal care and provisioning ofyoung by a range of helpers accountsfor many of these species-specifictraits.9,14,15 The life-history, ecologi-cal, and demographic dimensions ofthe cooperative breeding hypothesisare beginning to be well documented,but much less work has been done ondeveloping the psychological and cog-nitive dimensions of the model, par-ticularly as they apply to allomothers.While Hrdy9,14,16 has explored theemotional implications for childrenand mothers, she has glossed over thecognitive implications for allo-mothers, our main focus here. Byextrapolating from the general pat-tern of psychological consequences ofcooperative breeding in other taxa,17
we elaborate the role of this breedingsystem in the emergence of uniquelyhuman cognition.
ARTICLES
Judith M. Burkart is a senior post-doc at theAnthropological Institute and Museum (AIM)of the University of Zurich and is interestedin the cognitive evolution of primates. E-mail:[email protected] B. Hrdy is Professor Emerita at the Uni-versity of California-Davis. Her recent book,Mothers and Others, deals with the cognitiveand emotional implications of humankind’sdeep legacy of cooperative breeding.Carel P. van Schaik is Professor and Direc-tor of the AIM. His main interests are soci-oecology and social evolution in primates,especially the primate foundations ofhuman culture and intelligence. He wroteAmong Orangutans and has co-edited var-ious volumes on primate behavior andconservation. E-mail: [email protected]
Key words: allomaternal care; prosociality; callitri-
chids; shared intentionality; food sharing
VVC 2009 Wiley-Liss, Inc.DOI 10.1002/evan.20222Published online in Wiley InterScience(www.interscience.wiley.com).
Evolutionary Anthropology 18:175–186 (2009)

GLOSSARY
Allomaternal care—care forimmatures by individuals female ormale group members other than themother, including, in many cases,care by the genetic father. Ideally,the term is reserved for those casesin which paternity is known, as instrictly monogamous breeding sys-tems or ones where DNA data areavailable.Cognitive performance versus
cognitive potential—the distinctionbetween cognitive performance andability refers to the cognitive poten-tial inherent in an organism (ability)and its actual implementation inreal-life problem-solving situations(performance), Performance can bereduced relative to potential by vari-ous factors. A well-known discrep-ancy between performance andpotential occurs in social learning,where social dynamics can inhibitperformance even if individualswould, in principle, be able to per-form it.80 Cooperative breedingremoves restrictions on performancein many ways,17 making it likely thatincreased performance in socio-cog-nitive contexts in cooperativelybreeding primates, compared to theirindependently breeding sister taxa, isnot necessarily linked to greater cog-nitive potential or ability per se.26
Cooperative breeding & inde-pendent breeding—breeding sys-tems can be described along a gra-dient that specifies who is responsi-ble for infant care. At one end ofthis continuum, we have independ-ent breeders with exclusive mater-nal care. Typical cases includechimpanzees or orangutans. Themother is very possessive of herinfant. Although she may allowmale or female group members(allomothers) to touch or handle(but not remove) her infant, for atleast the first six months. she resistsall attempts by others to take orcarry her infant. In species withexclusive maternal care, allomotherscan take infants only under unusualcircumstances, as when the motheris incapacitated or a dominantfemale ‘‘kidnaps’’ her infant. Coop-erative breeders fall at the other end
of the continuum, where mothersvoluntarily permit access to theirinfants and many group membersare actively engaged in activecare and provisioning of infants,thereby increasing their growth andsurvival.Cultural intelligence—in its
broad version,83–85 the culturalintelligence hypothesis seeks toexplain why species engaging insocial learning are more likely thanothers to evolve large brains. It pro-poses that optimization of sociallearning favors the evolution oflarger brains and increased generalcognitive potential because organ-isms capable of social learning canmore easily respond to selectivepressures to enhance cognitive skillsand brain size. Because social guid-ance improves the signal-to-noiseratio in available environmentalinputs necessary for brain develop-ment relative to individual explora-tion and learning as required forindividual skill acquisition, the effi-ciency with which brain tissue cangenerate adaptive cognitive skillscan thus be increased if skill acqui-sition is socially guided. The humanversion of the cultural intelligencehypothesis refers to the processwhereby our species-typical socio-cognitive abilities, (including sharedintentionality) which emerge anddevelop in early childhood, triggersthe emergence of uniquely humancognition.12
Prosocial behavior—behaviorsthat produce benefits to others. Atthe proximate level, such behaviorscan be motivated by impulses tohelp others (spontaneous prosocial-ity), but they can also result fromother psychological processes, suchas the calculation of one’s ownfuture benefits by calculated reci-procity, enforcement by the recipi-ent through harassment or intimi-dation or, more subtly, be elicitedby tolerated theft and begging bythe recipient and high social toler-ance for these activities by thedonors.Spontaneous prosociality—refers
to a motivational predisposition toperform acts that benefit others,
even in the absence of the expecta-tion of reciprocation and solicita-tion by the recipient, such as beg-ging and harassment. Importantly,such spontaneous prosociality ismore than a quantitative extensionof social tolerance, which is a per-missive, but passive attitudetowards various behaviors of socialpartners. This is because prosocial-ity crucially includes a motivationaldrive actively to affect others’ cir-cumstances in a positive way, aspontaneous helping impulse thatdoes not have to be elicited throughexternal signals like begging. Spon-taneous prosociality corresponds tothe concept of ‘‘other-regardingpreferences’’ commonly invoked byeconomists to describe behavioraloutcomes that are not only moti-vated by the maximization of one’sown benefits, but also increasesbenefits to others.7
Shared intentionality—refers tothe ‘‘ability to participate withothers in collaborative activitieswith shared goals and intentions’’along with the desire to do so.22:675
This capacity emerges much earlierduring human ontogeny than does afully fledged theory of mind, but itsroutine expression is strikinglyabsent in other apes.12 It is basedon a cognitive component, or theunderstanding of others, goalsand intentions, and on a motiva-tional component, the desire to dothis. In a proposal central to thispaper, Tomasello and Carpenter25
specified how shared intentionalityis crucial in transforming thebasic understanding of other minds,as found in great apes, into jointattention, cooperative communica-tion, collaboration, and instructedlearning. These, in turn, give riseto further cognitive developments,for example by cultural construc-tion,113,128 but also by releasingco-evolutionary processes betweensocial and nonsocial cognitive abil-ities whereby the increased effi-ciency of brains due to sociallearning also facilitates the develop-ment and evolution of nonsocialcognitive abilities (see also ‘‘culturalintelligence’’).85
176 Burkart et al. ARTICLES

Before we can explain the cognitiveevolution of the human lineage, wehave to identify the cognitive differen-ces between humans and the otherapes. While earlier accounts have oftenstressed the key role of theory ofmind,18–20 more recent results suggestthat apes do have simple elements ofthat theory, such as an emergingunderstanding of what others areattending to and intending, althoughthese capacities are primarilyexpressed in competitive contexts11
(but see Penn and Povinelli21). Suchfindings shift the focus onto the appa-rently central role played by sharedintentionality,22,23 which has now beenidentified as the fundamental sourcefor the majority, if not all, of ourunique cognitive achievements. Thoseinclude language, complex technolo-gies, and art, as well as formalizednorms and institutions, which in turngave rise to religion. These achieve-ments permit the formation of large,structurally complex societies, such asstates.23–25
The crucial question is thereforewhat precipitated the evolution ofshared intentionality. Shared inten-tionality is critically based on a proso-
cial motivational predisposition thatinvolves an interest in sharing psycho-logical states with others.22 The rudi-mentary understanding of psychologi-cal states in apes seems to be restrictedto competitive contexts,11 not coopera-tive contexts. Hence, the preconditionfor shared intentionality that apes lackis not the capacity to grasp mentalstates per se, but the motivational pre-disposition actively to share mentalstates with others, to care about others’mental states, and thus to use mentalstate understanding systematically incooperative contexts.
These findings indicate that discover-ing what really made us human largelyboils down to answering the questionof what was responsible for the evolu-tion of such fundamentally prosocialattitudes. Based on comparative datasuggesting that cooperative breedingplays a central role in the evolution ofprosociality (Box 1), we argue thatengaging in routine allomaternal careand provisioning was the impetusbehind the emergence of prosocial dis-positions and thus, eventually, uniquelyhuman cognition (Fig. 1). Hence, wewill first briefly summarize the psycho-logical and resulting cognitive conse-quences of cooperative breeding basedon studies of nonhuman primates and
other mammals (reviewed elsewhere indetail17,26). We will then return to thespecific human case by asking readersto engage in a thought experiment:What happens if we take a clever apewith incipient tool manufacturing andtool using potentials, rudimentarytheory of mind, and some empathiccapacity, then introduce cooperativebreeding, a novel mode of child-rear-ing? We conclude by proposing a set ofpredictions and steps required for fur-ther testing of these ideas.
THE PSYCHOLOGY OFCOOPERATIVE BREEDING
Broadly defined, cooperative breed-ing refers to any breeding system inwhich individuals other than parents(alloparents) help to care for and provi-sion offspring.27,28 Some researchersrequire that breeding-age helpers tem-porarily forgo independent reproduc-tion,29,30 as has been argued to be thecase among humans (mid-life meno-pause).9,31 As will become evident, thispoint is not relevant to the psychologi-cal and cognitive dimension of the co-operative breeding hypothesis elabo-rated here. The crucial feature here isthe availability in the group of a num-
Figure 1. The role of cooperative breeding in the transition from ape-like to uniquely human cognition. In many species, engaging in shared careplus provisioning is likely tobeaccompaniedbypsychological adaptations suchas increased social toleranceand spontaneous prosociality. Thesecan increase cognitive performance in the social domain, as seen in callitrichids. In humans, however, spontaneous prosociality was added to analready ape-like cognitive system, among others capable of basic mental state understanding. In addition to the cognitive consequencesobserved in other cooperatively breeding species, this enabled the emergence of shared intentionality. Shared intentionality (see glossary) hasbeen identifiedasa keydifferencebetweenhumansandother greatapes; it is responsible for theemergenceof uniquely humancognitive systemsbothphylogenetically andontogenetically.22 [Color figurecanbeviewed in theonline issue,which is availableatwww.interscience.wiley.com.]
ARTICLES Cooperative Breeding and Human Cognitive Evolution 177
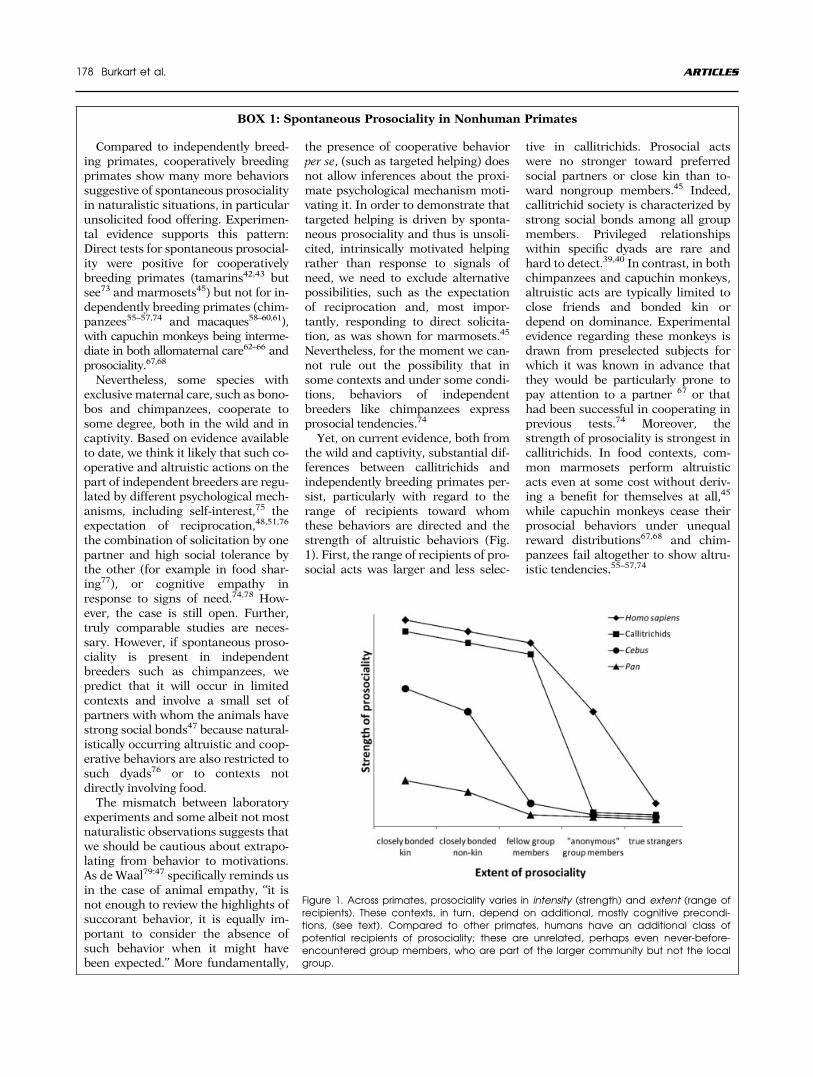
BOX 1: Spontaneous Prosociality in Nonhuman Primates
Compared to independently breed-ing primates, cooperatively breedingprimates show many more behaviorssuggestive of spontaneous prosocialityin naturalistic situations, in particularunsolicited food offering. Experimen-tal evidence supports this pattern:Direct tests for spontaneous prosocial-ity were positive for cooperativelybreeding primates (tamarins42,43 butsee73 and marmosets45) but not for in-dependently breeding primates (chim-panzees55–57,74 and macaques58–60,61),with capuchin monkeys being interme-diate in both allomaternal care62–66 andprosociality.67,68
Nevertheless, some species withexclusive maternal care, such as bono-bos and chimpanzees, cooperate tosome degree, both in the wild and incaptivity. Based on evidence availableto date, we think it likely that such co-operative and altruistic actions on thepart of independent breeders are regu-lated by different psychological mech-anisms, including self-interest,75 theexpectation of reciprocation,48,51,76
the combination of solicitation by onepartner and high social tolerance bythe other (for example in food shar-ing77), or cognitive empathy inresponse to signs of need.74,78 How-ever, the case is still open. Further,truly comparable studies are neces-sary. However, if spontaneous proso-ciality is present in independentbreeders such as chimpanzees, wepredict that it will occur in limitedcontexts and involve a small set ofpartners with whom the animals havestrong social bonds47 because natural-istically occurring altruistic and coop-erative behaviors are also restricted tosuch dyads76 or to contexts notdirectly involving food.The mismatch between laboratory
experiments and some albeit not mostnaturalistic observations suggests thatwe should be cautious about extrapo-lating from behavior to motivations.As deWaal79:47 specifically reminds usin the case of animal empathy, ‘‘it isnot enough to review the highlights ofsuccorant behavior, it is equally im-portant to consider the absence ofsuch behavior when it might havebeen expected.’’ More fundamentally,
the presence of cooperative behaviorper se, (such as targeted helping) doesnot allow inferences about the proxi-mate psychological mechanism moti-vating it. In order to demonstrate thattargeted helping is driven by sponta-neous prosociality and thus is unsoli-cited, intrinsically motivated helpingrather than response to signals ofneed, we need to exclude alternativepossibilities, such as the expectationof reciprocation and, most impor-tantly, responding to direct solicita-tion, as was shown for marmosets.45
Nevertheless, for the moment we can-not rule out the possibility that insome contexts and under some condi-tions, behaviors of independentbreeders like chimpanzees expressprosocial tendencies.74
Yet, on current evidence, both fromthe wild and captivity, substantial dif-ferences between callitrichids andindependently breeding primates per-sist, particularly with regard to therange of recipients toward whomthese behaviors are directed and thestrength of altruistic behaviors (Fig.1). First, the range of recipients of pro-social acts was larger and less selec-
tive in callitrichids. Prosocial actswere no stronger toward preferredsocial partners or close kin than to-ward nongroup members.45 Indeed,callitrichid society is characterized bystrong social bonds among all groupmembers. Privileged relationshipswithin specific dyads are rare andhard to detect.39,40 In contrast, in bothchimpanzees and capuchin monkeys,altruistic acts are typically limited toclose friends and bonded kin ordepend on dominance. Experimentalevidence regarding these monkeys isdrawn from preselected subjects forwhich it was known in advance thatthey would be particularly prone topay attention to a partner 67 or thathad been successful in cooperating inprevious tests.74 Moreover, thestrength of prosociality is strongest incallitrichids. In food contexts, com-mon marmosets perform altruisticacts even at some cost without deriv-ing a benefit for themselves at all,45
while capuchin monkeys cease theirprosocial behaviors under unequalreward distributions67,68 and chim-panzees fail altogether to show altru-istic tendencies.55–57,74
Figure 1. Across primates, prosociality varies in intensity (strength) and extent (range ofrecipients). These contexts, in turn, depend on additional, mostly cognitive precondi-tions, (see text). Compared to other primates, humans have an additional class ofpotential recipients of prosociality; these are unrelated, perhaps even never-before-encountered group members, who are part of the larger community but not the localgroup.
178 Burkart et al. ARTICLES

ber of reliable helpers. Among prim-ates, allomaternal care, whereby groupmembers other than the mother pro-tect, keep warm, or otherwise care forthe young, is widespread. However, thestrongest reliance on allomaternal careand provisioning is found in humansand callitrichids, where many groupmembers contribute to infant-rearing.9
Callitrichid monkeys, (marmosetsand tamarins),32 live in family groupstypically composed of a single breedingpair and its adult and immature off-spring, although polygynous and espe-cially polyandrous constellations occuras well.33 Helpers tend not to be repro-ductively active in the group. All groupmembers, including unrelated or eveninitially unfamiliar individuals,34 sys-tematically engage in many cooperativebehaviors, ranging from infant carry-ing, shared vigilance, and systematicprovisioning with high-value food itemsto collective action such as communalgroup defense (Fig. 2).32,35 Zahed andcoworkers36 demonstrated that adultmale marmosets in general possess aspontaneous motivation to care for anyinfant, related or not. Helpers oftenprovision offspring by giving food callsand actively offering high-value fooditems rather than in response to infantbegging.37 Callitrichid infants sponta-neously transfer to allomothers. This isunusual among primates and suggeststhat there has been a sufficiently long
history of benevolent attention andeffective allomaternal care to produceselection for this self-transfer byinfants. Allomothers, meanwhile, neverforcefully retrieve an infant fromanother helper or the mother. Rather,transfers occur in a highly orchestratedmanner, indicating that each care-taker’s behaviors and intentions arecontinuously monitored and theirbehaviors are adjusted accordingly.Finally, callitrichids show high levels ofsocial tolerance (Fig. 3), are highly re-sponsive to signals from other groupmembers,38–40 and do not appear topunish underperforming helpers.41
Based on this natural history, one candeduce that callitrichid caregivers havespontaneously prosocial motivationsthat render them eager to perform actsthat benefit others, even in the absenceof either solicitation by the recipient,(such as begging or harassment), or ex-pectation of reciprocation.
Experimental tests show that cotton-top tamarins42–44 and commonmarmo-sets45 have a spontaneous helpingimpulse when they are given the oppor-tunity to provide food to group mem-bers, even if they don’t receive anythingfor themselves if recipients cannotreciprocate, and in the absence of beg-ging. Moreover, prosocial food dona-tions in the marmoset experimentswere not exclusively directed at infantsor other immatures but to any group
members; equally strong toward rela-tives as to unrelated group members orunrelated potential group members;not solicited by the recipients; and notselectively directed at preferred socialpartners. If spontaneous prosocialitywere to occur in any primate with morenearly exclusively maternal care ofyoung infants, we would expect itamong species such as bonobos, chim-panzees, or capuchins, since all threeoccasionally share food and exhibitmore cooperative and altruistic behav-iors than is typical of most primates.46–49
However, their food sharing is usuallypreceded by begging or resembles toler-ated taking.49–52 Bonobo allomothersvery occasionally allow immature indi-viduals to take vegetable food or meat,and in rare instances offer it.46,53
Among chimpanzees, even food shar-ing between mother and infant resem-bles tolerated theft. Rare cases of dona-tion typically involve low-value fooditems or discarded remains such asnonedible husks and only occur afterbegging by the infant.54 The infrequentand often grudging nature of these fooddeliveries differs from the spontaneous,unsolicited, and routine offering ofhigh-value food that is observed amongcallitrichids and humans, includingchildren.37 In accordance with thesedifferences, experiments have so farfailed to show evidence of prosocialityin species with exclusive maternal care
Figure 2. A family group of cooperatively breeding golden lion tamarins (Leontopithecus rosalia). Females can give birth to twins twice ayear without experiencing lactational amenorrhea. The mother can afford this high energetic investment because the infants are carried,and after weaning, provisioned by all group members, mostly fathers and older siblings, but also nonrelatives. Drawing by Sarah Landry.
ARTICLES Cooperative Breeding and Human Cognitive Evolution 179

of infants, such as chimpanzees55–57 ormacaques.58–61
Capuchin monkeys show more ele-ments of cooperative breeding than dothe Pan species, with occasional alloma-ternal carrying, suckling of older infantsbetween the ages of 3 and 6 months,and low levels of allomaternal provi-sioning.62–66 The majority of sharedfood is transferred from allomothers toimmatures. In one-fifth of all 18 obser-vations of food transfer, it was activelyoffered by one monkey to another. Halfof these involved offerings by nullipar-ous females to infants49:740. On rareoccasions, food is actively sharedbetween adults.52 Indeed, experimentssuggest some prosociality among capu-chins.67,68 A broader discussion is pro-vided in Box 1.Some canids, elephants, and other
nonprimate cooperative breedersexhibit similar natural history, suggest-ing analogous prosocial motivations.17
Spontaneous prosociality is suggestedby a variety of behaviors of African wilddogs (Lycaon pictus). These includespontaneous provisioning through re-gurgitation and, less often, the carryingof carcass portions back to the den toprovision pregnant and lactating moth-ers or babysitters as well as pups. Inaddition, pups enjoy feedingpriority. Such extreme allomaternalinvestment is not the result of coercion.
Indeed, it is not even restricted toclosely related pups, underscoring justhow intrinsic motivations to nurtureand provision are. Canid prosocial careeven extends to adult group members,as suggested by tolerance at kills andcases in which incapacitated and olderpack members are provisioned byothers. Similar results are reported forelephants among which allomaternalcare improves calf growth and sur-vival.17 Thus, although no experimentshave been conducted yet, the naturalhistory of these nonprimate coopera-tive breeders suggests the presence of asimilar helping impulse.
When such spontaneous prosocialityextends from the donation of food tothat of information, we enter the realmof teaching. As Rapaport emphasized,69
teaching, which is rare in nature, isstrikingly overrepresented in speciesrelying on cooperative breeding; posi-tive evidence is limited to ants, piedbabblers, meerkats, callitrichids, andsolitary felids.69 Thus, teaching is lim-ited to species showing unsolicited fooddonation, and hence is overrepresentedamong cooperative breeders.70 Its pres-ence in independently breeding felinecarnivores derives from the need toprovision offspring who require muchtime to learn to capture their ownprey.71 Despite the impressive socio-cognitive potential of nonhuman apes,
such as simple mental state attribu-tion,72 it is worth noting that compara-ble observations of teaching are notreported for these taxa.In sum, in nonhuman primates,
canids, and elephants, cooperativebreeding is associated not only withincreased levels of social toleranceand responsiveness to the signals andneeds of others, but also with thepresence of spontaneous prosocialmotivations, which extend beyondinfants and sometimes beyond food toinformation, resulting in teaching.
COOPERATIVE BREEDING ANDCOGNITION
There is no a priori reason whycooperative breeders would requiregreater cognitive skills than do otherspecies. However, cooperative breeding
seems to result in increased cognitiveperformance as a side effect.9,16,38
Indeed, a comparison17,26 of callitri-chids and their more independentlybreeding sister taxa, capuchin andsquirrel monkeys, in their performanceon cognitive tasks found that callitri-chids systematically outperformed theirsister taxa in the social domain, butscored lower in nonsocial cognitivetasks (Table 1). These effects, however,need not reflect fundamentallyenhanced cognitive ability. Instead,they are more likely the result of moti-vational changes associated with coop-erative breeding.26 Social learning, forexample, which has been documentedmore consistently in primates that
Figure 3. Common marmosets (Callithrix jacchus). A sexually mature helper watches howanother adult processes an unusual food. Relaxed, close monitoring of each others’ activities isnot restricted to infant-caregiver dyads. It occurs in all dyad types and in a variety of contexts.[Color figurecanbeviewed in theonline issue,which is availableatwww.interscience.wiley.com.]
There is no a priori reasonwhy cooperativebreeders would requiregreater cognitive skillsthan do other species.However, cooperativebreeding seems to resultin increased cognitiveperformance as a sideeffect.
180 Burkart et al. ARTICLES

breed cooperatively than in primatesthat do not, is well known to dependnot only on the cognitive skills involved,but at least as much on the level ofsocial tolerance.80 Such tolerance facili-tates close-range attention to conspe-cific behavior and even permits mutualgazing without provoking attack (Bur-kart, unpublished; personal communi-cation from Karen Bales). Indeed, evenif the cognitive prerequisites for imita-tional learning are present in othermonkey species,81,82 their applicationto imitative learning may be hamperedby a lack of mutual tolerance.26 Thus,the more effective performance of co-operative breeders in socio-cognitivetasks is likely to be a side effect of moti-vational changes directly related to thedeployment and coordination of care-giving activities,17 as well as a sideeffect of developmental adjustments byimmature individuals, who must moni-tor the whereabouts and intentions oftheir mother and allomothers.9,16
Cooperative breeding may alsoremove obstacles to the evolution ofbrain size. First, because social learningenhances the efficient use of brain tis-sue, a social system with increasedopportunities for social learning can,over evolutionary time, favor the evolu-tion of larger brains, as detailed in thegeneral version of the cultural intelli-gence hypothesis.83–85 This hypothesispredicts that many cooperativebreeders have evolved larger brainsthan have their independently breedingcounterparts. Second, Isler and vanSchaik86,87 have shown the existence ofa maximum sustainable brain size in agiven lineage. This ‘‘gray ceiling’’ is aresult of a strong reduction in maxi-mum reproductive rate due toincreased brain size (principally as aconsequence of delayed maturation).88
However, these researchers also foundthat this rule does not hold among co-operative breeders, probably becauseenergy inputs to mothers and newlyweaned infants from allomaternal pro-visioning allows these species to escapefrom under this so-called gray ceilingand thus evolve larger brains.
These kinds of effects may explainwhy cooperative breeding is linked notonly to psychological dispositions, butalso to cognitive performance in prima-tes and other mammals. Althoughdetails of cooperative breeding systemsare bound to differ between taxadepending, for example, on the degreeof reproductive skew or presence ofnonkin, other cooperatively breedingmammals, such as canids and ele-phants, show similarly strong socio-cognitive performance without a con-comitant increase in nonsocial cogni-tive tasks.17 The socio-cognitive per-formance of elephants parallels the cal-litrichid pattern. With the exception oftheir memory capacity, their perform-ance in many nonsocial cognitive tasksis unimpressive for a mammal withsuch a large brain, yet they excel insocio-cognitive contexts, including mir-ror self-recognition and vocal imita-tion.
Recent experiments with dogs haverevealed socio-cognitive performancesthat rival those of apes. These includethe understanding of visual perspectiveand mental states, victim-directedthird-party postconflict affiliation(‘‘consolation’’), reasoning by exclusionin a social context, sophisticated pas-sive and active communicative abilities,cooperation skills, and imitation,including selective inferential imitation.Yet in the nonsocial and physicalrealm, dogs perform less impressivelythan do great apes. However, such
studies are rare and thus difficult toevaluate.17 Superior socio-cognitiveperformance in dogs cannot beexplained by domestication alone89,90
since comparable effects are not foundin other domesticated animals exceptthose that are also cooperativebreeders. For example, rapid experi-mentally induced domestication of sil-ver foxes (Vulpes vulpes), which alsobreed cooperatively,91,92 produced asimilar increase in socio-cognitive per-formance.93 Thus, socio-cognitive abil-ities that were already present in wildancestors are likely to have been ampli-fied and directed by humans throughselective breeding, or domestication.A positive effect of cooperative breed-
ing on inter-individual social tolerance,including maternal tolerance of otherspostpartum, spontaneous prosociality(the helping impulse) and socio-cogni-tive performance is well documentedfor nonhuman primates and likely forother taxa. An extrapolation of thesegeneral findings to the hominin lineagesuggests that humans also fit this pat-tern.
COOPERATIVE BREEDING ANDTHE EVOLUTION OF HUMAN
COGNITION
Given the motivational and cognitiveconsequences of cooperative breedingin other taxa what might have hap-pened when our ape-like ancestorsadopted such breeding system? Wecan extrapolate from the comparativefindings and provide a first, tentativesketch of how this new mode of child-rearing could have led to key featuresof uniquely human cognition. Fittingthe general pattern, humans clearlyshow the psychological dispositionsassociated with cooperative breeding,particularly strong social tolerance andspontaneous prosociality. The cognitivedifferences between our great-ape likeancestors and ourselves, however, arefar more pervasive than those betweencallithrichids and their sister taxa. Weargue that the cognitive consequencesof cooperative breeding were more pro-nounced in our ancestors than in othertaxa because the selection pressuresassociated with cooperative breedingwere acting on an already ape-like cog-nitive system, allowing for the emer-
TABLE 1. Cognitive Domains in Which Cooperatively Breeding Callitrichids Do or
Do Not Outperform Their Independently Breeding Sister Taxaa
Increased in cooperatively
breeding primates
Not increased in cooperatively
breeding primates
Socio-cognitive abilities Nonsocial cognitive abilities� Social learning � General cognitive ability� Vocal communication � Working memory of actions� Teaching-like behaviors � Innovation rates� Gaze understanding � Tool-use rates� Cooperative problem solving � Patience
� Inhibitory control
a For a full description, see Burkart and van Schaik.17
ARTICLES Cooperative Breeding and Human Cognitive Evolution 181

gence of shared intentionality (Fig.1).22 Our hypothesis is that while chim-panzees and, perhaps, all great apes10,11
may have many of the relevant cogni-tive preconditions for uniquely humancognition to evolve, they lack the moti-vational preconditions. In humansalone, these two components havecome together, the cognitive compo-nent due to common descent and themotivational component due to conver-gent evolution resulting from the selec-tion pressures associated with coopera-tive breeding (Fig. 4).Understanding the role of cooperative
breeding in the emergence of humancognition first requires that we delineatethe point of departure; that is, the cogni-
tive system that was in place when theselection pressures of cooperativebreeding were added during humanevolution. Because cooperative breedingarose after the split between homininsand great apes, a conservative estimateof the cognitive endowment of the homi-nin that first adopted cooperative breed-ing would be that it was rather similarto that of the last common ancestor.Given the cognitive similarities amonggreat apes, we postulate that the cogni-tive potential for early hominins wassimilar to that of extant great apes; thatis, more complex than the cognitivepotential of monkeys in both social andnonsocial domains.10,11,94
Two consequences follow directlyfrom adding cooperative breeding tosuch a cognitive system. First, existingcognitive skills become available fordeployment in cooperative contexts. Inthe nonsocial domain, cognitive per-formance tends to be more pronouncedin great apes than in other nonhumanprimates as shown by a meta-analysisbased on a variety of tasks such aslearning sets, patterned-string prob-lems, reversal learning, delayedresponse, and invisible displacement.10
Great apes also use tools more often,95
do so based on causal understand-ing,96,97 and plan ahead by anticipatingtheir future needs, such as need for atool.98,99 Such skills, in particular sim-ple planning, can greatly improve thecoordination of activities among groupmembers. Second, cooperative breed-ing amplifies opportunities for social
learning. Immature individuals in coop-eratively breeding species haveincreased opportunities for social learn-ing because of the availability of multi-ple, highly tolerant role models, as wellas potentially longer juvenile learningperiods,16,100 thus expanding individualskill repertoires.Such immediate consequences can
also occur in other cooperatively breed-ing species, but will be less pronouncedbecause ape-level cognitive potentialwas not present to begin with. Thus,the cognitive consequences to be dis-
Figure 4. Origin of key components of uniquely derived human cognition as elaboratedin the text. The components highlighted in italics indicate sequential evolutionary pro-cesses further specified in the text. [Color figure can be viewed in the online issue, whichis available at www.interscience.wiley.com.]
Our hypothesis is thatwhile chimpanzees and,perhaps, all great apesmay have many of therelevant cognitivepreconditions foruniquely humancognition to evolve, theylack the motivationalpreconditions. Inhumans alone, thesetwo components havecome together...
182 Burkart et al. ARTICLES

cussed could have emerged only inhumans. Intentional teaching requiresan understanding of another individu-al’s knowledge state.101,102 Such anunderstanding is present in great apes,but is predominantly restricted to com-petitive contexts.11,25,72,103 Further-more, some understanding of causalrelationships in general, which is pres-ent and, to all appearances, deeper ingreat apes than in other primates,10,97
is an important precondition for theactive transmission of more complexskills. However, all these cognitive com-ponents will fail to result in intentionalteaching unless some willingness toshare information is present, a willing-ness derived from an extension of spon-taneous prosociality in food-sharingcontexts to information sharing. Inhumans, sharing information withimmature individuals seems to haveplayed a particularly important role, ashighlighted by Csibra and Gergely104
who propose that we have evolved addi-tional adaptations facilitating skilltransfer, summarized under the con-cept of pedagogy.The sporadic occurrence of targeted
helping among chimpanzees under-scores their ability to understand thegoals, intentions and needs ofothers.74,105 Overall, however, tar-geted helping is rare; it is entirelyabsent when food is at issue, suggest-ing that chimpanzees lack any strongmotivation to help, even if they pre-sumably understand how their tar-geted actions could benefit others inspecific situations. Adding a helpingimpulse would further increase themotivation to improve others’ situa-tions when preexisting cognitivemechanisms correctly identified thatand why an individual is in need ofhelp, thus resulting in the systematicoccurrence of targeted helping.Chimpanzees may have the cognitive
prerequisites for recognizing inequit-able distributions and responding toegocentric or ‘‘disadvantageous’’inequity if they themselves are affectedby unfair offers.106 However, this issueis controversial.107 If such a basic abil-ity to detect inequity is coupled with aconcern not only for one’s own well-being, but also for that of others, ego-centric inequity aversion can turn intoallocentric inequity aversion; that is, an-
tipathy against unfair treatment ofothers, or at least in-group members.6
The rudimentary ability to graspothers’ mental states, which has beendocumented for great apes72 but notfor cooperatively breeding primates(such as marmosets),108 makes itpossible that in humans the helpingimpulses are no longer confined tofood or information, but extend to awillingness to share mental states.This, in turn, enables the emergenceof shared intentionality, which is crit-ically based on a prosocial motiva-tional predisposition that encom-passes an interest in sharing psycho-logical states with others.22 Sharedintentionality is considered to be thebasis for many aspects of uniquely
human cognition.25 Through sharedintentionality, the adoption of coop-erative breeding might have influ-enced the emergence of an array ofother capacities, including languageand cumulative cultural evolution.Let us briefly consider these.
Experiments with enculturated greatapes illustrate that they possess the cog-nitive capacities for acquiring simplelanguage systems.109 However, the useof such systems among great apesremains predominantly imperative;that is they use their language skills pri-marily to request things rather than forsharing information with others in de-clarative modes.18 The same limitationis apparent in their use of pointing ges-tures, which are also restricted to im-
perative contexts.110,111 The absence ofa helping impulse is manifest in thislack of interest in sharing informationwith others. This strongly contrastswith the way humans use language.Even from a very young age, childrenuse language for declarative pur-poses.112 Correspondingly, many devel-opmental psychologists have empha-sized the role of joint attention andshared intentionality for language de-velopment.23,113–116
Any scenario for the evolution oflanguage needs to explain where thisfundamental prosocial and coopera-tive attitude essential for languagecame from and how it originated. Lan-guage could only evolve in a commu-nication system characterized by pro-sociality and, as Zahavi117 pointed outlong ago, would disappear if cheaterspredominated. Indeed, it is a commonnotion in linguistic pragmatics thathuman communication is based onthe principle of cooperation.118,119 Thecooperative breeding hypothesis pro-vides a simple and biologically validsolution to this longstanding problem.The psychological endowment of co-operative breeders provides preciselythe motivational preconditions neededfor the evolution of honest, low-costcommunication signals.120
Finally, shared care and provisioningcan have the potential to promote cul-tural evolution in multiple ways. Cul-tural evolution requires innovations andtheir subsequent social transmission.121
While cultural variation in behavior isknown in great apes, the contents ofthese cultures tend to be hardly morecomplex than what could be independ-ently invented. Moreover, they are onlymarginally cumulative.122,123 Comparedto other apes, more active, accurate,and reliable transmission of skills andknowledge are favored in humans byincreased opportunities for social learn-ing, the presence of intentional teach-ing, shared intentionality, and declara-tive communicative activities. It is alsoconceivable that prosociality promotesinnovation, but this topic is as yetlargely unexplored. However, thecapacity to solve problems cooperativelycan result in pooling of individualstrengths, particularly when individualsjointly participate with others in collab-orative activities with shared goals andintentions.124
... it is a common notionin linguistic pragmaticsthat humancommunication is basedon the principle ofcooperation. Thecooperative breedinghypothesis provides asimple and biologicallyvalid solution to thislongstanding problem.
ARTICLES Cooperative Breeding and Human Cognitive Evolution 183

It is important to note that thetransitions we have described arelikely to be achieved on a popula-tion-wide basis only if the helpingimpulse, as expressed in willingnessto share information or mentalstates, is also ubiquitous in the popu-lation rather than restricted to spe-cific dyads. This could help explainwhy the unusual cognitive potentialsuniversally present in Homo sapiensevolved in a line of cooperativelybreeding apes while failing to evolvein independently breeding apes likechimpanzees, no matter how intelligenttheir ancestors were to begin with.
FUTURE DIRECTIONS
The cooperative breeding hypothe-sis is compatible with most otherhypotheses for the origin of humanuniqueness in that it provides thecontext in which these various proc-esses could operate (discussed exten-sively by van Schaik and Burkart125).Moreover, it is consistent with ourcurrent state of knowledge of theevolution of human derived features.Nonetheless, more rigorous tests areneeded of both the broad interspe-cific version, which requires system-atic examination of various predic-tions of this hypothesis in a broadarray of species, and of the specificapplication to the human lineage.c Comparisons of the cognitive abil-
ities of cooperative and independentbreeders need to be extended. Are thesystematic cognitive effects of coopera-tive breeding observed in callitrichidsalso present in other taxa? If so, dothey involve social cognitive abilities,nonsocial ones, or both?c What is the exact link between
prosociality and cooperative breed-ing? So far only a few species havebeen tested to provide evidence ofthis link, often using diverse experi-mental paradigms (see Box 1). A sys-tematic assessment requires bothreliable estimates of the extent ofallomaternal care in different spe-cies9,126 and a standardized para-digm for testing for prosocialityacross a wide range of species toprovide truly comparable results.c If there is a causal connection
between prosociality and cognition,this should also be reflected in intra-
specific variation, for example amongindividual capuchins or chimpanzeesvarying in prosociality, perhaps as aresult of enculturation.
c The interpretation of the con-trasts between humans and greatapes with respect to division oflabor, collective action, and life his-tory would be strengthened if coop-erative breeders in general differfrom their independently breedingsister taxa in these characteristics.
c What dimensions of cooperativebreeding are relevant to the ques-tions we have posed? Cooperativebreeding systems are far from uni-form. They differ with regard to thedegree of reproductive skew, the kindof helping performed by allomothers,the age-sex classes that serve as allo-mothers, and the relative stress levelsof subordinates and dominants.
c The cooperative breeding hypoth-esis posits that alloparental care andprovisioning provided the context forthe evolution of many other derivedfeatures characterizing both humanlife histories and human sociocogni-tive and emotional traits. Conse-quently, the hypothesis predicts thatlife-history corollaries of cooperativebreeding, such as longer childhoods,and behavioral corollaries, such ascaring for disabled group members,should appear in the paleontologicalrecord at about the same time andearly in the evolution of the line lead-ing to Homo sapiens. Indeed, thesecorollaries should precede even morederived features such as fully sapient-sized brains, symbolic art, language,and cultural group selection. Althoughconsiderable debate persists abouthow to interpret the fossil record,there are certainly grounds to be opti-mistic about these predictions. 9,125,127
ACKNOWLEDGMENTS
This project was supported by theSwiss National Science Foundationprojects No. 105312-114107 and No.3100A0-111915.
REFERENCES
1 Glazko GV, Nei M. 2003. Estimation of diver-gence times for major lineages of primate spe-cies. Mol Biol Evol 20:424–434.
2 de Waal FBM. 2005. Our inner ape. New
York: Riverhead Books.
3 Boesch C. 2007. What makes us human(Homo sapiens)? The challenge of cognitivecross-species comparison. J Comp Psychol121:227–240.
4 Robson SL, Hawkes K, van Schaik CP. 2006.The derived features of human life history. In:Hawkes K, Paine RL, editors. The evolution ofhuman life History. Santa Fe: School of Ameri-can Research Press. p 17–44.
5 Kaplan HK, Hill K, Lancaster J, Hurtado AM.2000. A theory of human life history evolution:diet, intelligence and longevity. Evol Anthropol9:156–185.
6 Henrich J, Boyd R, Bowles S, Camerer C,Fehr E, Gintis H, McElreath R, Alvard M, BarrA, Ensminger J, and others. 2005. ‘‘Economicman’’ in cross-cultural perspective: behavioralexperiments in 15 small-scale societies. BehavBrain Sci 28:795–855.
7 Fehr E, Fischbacher U. 2003. The nature ofhuman altruism. Nature 423:785–791.
8 Batson CD. 1991. The altruism question: to-ward a social psychological answer. Hillsdale,NJ: Erlbaum.
9 Hrdy S. 2009. Mothers and others: the evolu-tionary origins of mutual understanding. Cam-bridge: Harvard University Press.
10 Deaner RO, van Schaik CP, Johnson V.2006. Do some taxa have better domain-generalcognition than others? A meta-analysis of non-human primate studies. Evol Psychol 4:149–196.
11 Call J. 2007. Social knowledge in primates.In: Dunbar RIM, Barrett L, editors. Handbookof evolutionary psychology. New York: OxfordUniversity Press. p 71–81.
12 Herrmann E, Call J, Hernandez-Lloreda MV,Hare B, Tomasello M. 2007. Humans haveevolved specialized skills of social cognition:the cultural intelligence hypothesis. Science317:1360–1366.
13 Aiello LC, Key C. 2002. Energetic conse-quences of being a Homo erectus female. Am JHum Biol 14:551–565.
14 Hrdy S. 2005. Evolutionary context ofhuman development: the cooperative breedingmodel. In: Carter CS, Ahnert L, Grossmann KE,Hrdy SB, Lamb ME, Porges SW, Sachser N,editors. Attachment and bonding: a new synthe-sis, from the 92nd Dahlem Workshop Report.Cambridge: MIT Press. p 9–32.
15 Hrdy S. 1999. Mother nature: a history ofmothers, infants, and natural selection. NewYork: Pantheon Books.
16 Hrdy S. 2005. Comes the child before theman: how cooperative breeding and prolongedpostweaning dependence shaped human poten-tials. In: Hewlett B, Lamb M, editors. Huntergatherer childhood. Psicataway: Transactions.p 65–91.
17 Burkart JM, van Schaik CP. n.d. Cognitiveconsequences of cooperative breeding in prima-tes. Anim Cogn. In press.
18 Tomasello M, Call J. 1997. Primate cogni-tion. New York: Oxford University Press.
19 Heyes C. 1998. Theory of mind in nonhu-man primates. Behav Brain Sci 21:101–184.
20 Povinelli DJ, Bering J, Giambrone S. 2000.Toward a science of other minds: escaping theargument by analogy. Cogn Sci 24:509–541.
21 Penn DC, Povinelli DJ. 2007. On the lack ofevidence that non-human animals possess any-thing remotely resembling a ‘‘theory of mind’’.Philos Trans R Soc B 362:731–744.
22 Tomasello M, Carpenter M, Call J, Behne T,Moll H. 2005. Understanding and sharing inten-tions: the origins of cultural cognition. Behav
184 Burkart et al. ARTICLES

Brain Sci 28:675–735.
23 Tomasello M, Rakoczy H. 2003. What makeshuman cognition unique? From individual toshared to collective intentionality. Mind Lan-guage 18:121–147.
24 Searle JR. 1995. The construction of socialreality. New York: Free Press.
25 Tomasello M, Carpenter M. 2007. Sharedintentionality. Dev Sci 10:121–125.
26 Burkart JM. 2009. Socio-cognitive abilitiesand cooperative breeding. In: Roska-Hardy LS,NeumannHeld EM, editors. Learning from ani-mals? Examining the nature of human unique-ness: Psychol Press. p 123–141.
27 Wilson EO. 1975. Sociobiology. Cambridge,MA: Harvard University Press.
28 Ligon JD, Burt DB. 2004. Evolutionary ori-gins. In: Koenig W, Dickinson J, editors. Ecologyand evolution of cooperative breeding in Birds.Cambridge: Cambridge University Press. p 5–34.
29 Solomon NG, French JA. 1997. Cooperativebreeding in mammals. New York: CambridgeUniversity Press.
30 Clutton-Brock T. 2006. Cooperative breedingin mammals. In: Kappeler PM, van Schaik CP,editors. Cooperation in primates and humans:mechanisms and evolution. Berlin: Springer. p173–190.
31 Foster KR, Ratnieks LW. 2005. A new euso-cial vertebrate? Trends Ecol Evol 20:363–364.
32 Digby LJ, Ferrari SF, Saltzman W. 2007.Callitrichines: the role of competition in coop-eratively breeding species. In: Campbell CJ,Fuentes A, MacKinnon KC, Panger MA,Bearder SK, editors. Primates in perspective.New York: Oxford University Press. p 85–105.
33 Garber PA, Rosenberger AL, Norconk MA.1995. Marmoset misconceptions. In: NorconkMA, Rosenberger AL, Garber PA, editors.Adaptive radiations of neotropical primates.New York: Plenum Press.
34 Dietz JM. 2004. Kinsip structure and repro-ductive skew in cooperatively breeding prima-tes. In: Chapais B, Berman C, editors. Kinshipand behavior in primates. Oxford: Oxford Uni-versity Press. p 223–241.
35 Savage A, Snowdon CT, Giraldo H, Soto LH.1995. Parental care patterns and vigilance inwild cotton-top tamarins (Saguinus oedipus).In: Norconk MA, Rosenberger AL, Garber PA,editors. Adaptive radiations of neotropical pri-mates. New York: Plenum Press. p.187–199.
36 Zahed SR, Prudom SL, Snowdon CT, ZieglerTE. 2007. Male parenting and response toinfant stimuli in the common marmoset (Calli-thrix jacchus). Am J Primatol 70:84–92.
37 Brown GR, Almond REA, van Bergen Y.2004. Begging, stealing and offering: food trans-fer in non-human primates. Adv Study Behav34:265–295.
38 Snowdon CT. 2001. Social processes in com-munication and cognition in callitrichid mon-keys: a review. Anim Cogn 4:247–257.
39 Aureli F, Schaffner C. 2006. Causes, conse-quences and mechanisms of reconciliation: therole of cooperation. In: Kappeler PM, vanSchaik CP, editors. Cooperation in primatesand humans: mechanisms and evolution. Hei-delberg: Springer. p 121–136.
40 Schaffner CM, Caine NG. 2000. The peace-fulness of cooperatively breeding primates. In:Aureli F, De Waal FB, editors. Natural conflictresolution. Berkeley, California: University ofCalifornia Press. p 155–169.
41 Snowdon CT, Cronin KA. 2007. Coopera-tive breeders do cooperate. Behav Proc76:138–141.
42 Hauser MD, Chen MK, Chen F, Chuang E.2003. Give unto others: genetically unrelatedcotton-top tamarin monkeys preferentially givefood to those who altruistically give food back.Proc R Soc B 279:2363–2370.
43 Cronin KA, Snowdon CT. 2008. Theeffects of unequal reward distributions oncooperative problem solving by cottontoptamarins, Saguinus oedipus. Anim Behav 75:245–257.
44 Cronin KA, Schroeder KKE, Rothewell ES,Silk JB, Snowdon CT. n.d. Cooperatively breed-ing cottontop tamarins (Saguinus oedipus) donot donate rewards to their long-term mates.J Comp Psychol. In press.
45 Burkart JM, Fehr E, Efferson C, van SchaikCP. 2007. Other-regarding preferences in a non-human primate, the common marmoset (Calli-thrix jacchus). Proc Natl Acad Sci USA104:19762–19766.
46 Hohmann G, Fruth B. 1996. Food sharing andstatus in unprovisioned bonobos. In: Wiessner P,Schiefenhovel W, editors. Food and the statusquest. Providence: Berghahn Books. p 48–67.
47 de Waal FBM. 2006. Primates and philoso-phers. New Jersey: Princeton University Press.
48 de Waal FBM, Brosnan SF. 2006. Simpleand complex reciprocity in primates. In:Kappeler PM, van Schaik CP, editors. Coopera-tion in primates and humans: mechanisms andevolution. Heidelberg: Springer. p 85–105.
49 Rose LM. 1997. Vertebrate predation andfood-sharing in Cebus and Pan. Int J Primatol18:727–765.
50 Gilby IC. 2006. Meat sharing among theGombe chimpanzees: harassment and recipro-cal exchange. Anim Behav 71:953–963.
51 Boesch C, Boesch-Achermann H. 2000. Thechimpanzees of the Tay forest: behavioral ecologyand evolution. Oxford: Oxford University Press.
52 De Waal FBM. 1997. Food transfers throughmesh in brown capuchins. J Comp Psychol111:370–378.
53 Kano T. The last ape: pygmy chimpanzeebehavior and ecology, translated by E. Vine-berg. Stanford: Stanford University Press.
54 Ueno A, Matsuzawa T. 2004. Food transferbetween chimpanzee mothers and their infants.Primates 45:231–239.
55 Silk JB, Brosnan SF, Vonk J, Henrich J,Povinelli DJ, Richardson AS, Lambeth SP, Mas-caro J, Schapiro SJ. 2005. Chimpanzees areindifferent to the welfare of unrelated groupmembers. Nature 437:1357–1359.
56 Jensen K, Hare B, Call J, Tomasello M.2006. What’s in it for me? Self-regard precludesaltruism and spite in chimpanzees. Proc R SocB 273:1013–1021.
57 Vonk J, Brosnan SF, Silk JB, Henrich J,Richerdson AS, Lambeth SP, Schapiro SJ, Povi-nelli DJ. 2008. Chimpanzees do not takeadvantage of very low cost opportunities todeliver food to unrelated group members. AnimBehav 75:1757–1770.
58 Mason WA, Hollis JH. 1962. Communicationbetween young rhesus macaques. Anim Behav10:211–221.
59 Colman AD, Liebold KE, Boren JJ. 1969. Amethod for studying altruism in monkeys. Psy-cho Rec 19:401–405.
60 Wolfle DL, Wolfle HM. 1939. The develop-ment of cooperative behavior in monkeys andyoung children. J Gen Psychol 55:137–175.
61 Silk JB. 2007. Empathy, sympathy and pro-social preferences in primates. In: Dunbar RIM,Barrett L, editors. Handbook of evolutionarypsychology. New York: Oxford University Press.
p 115–126.
62 O’Brien TG, Robinson J. 1991. Allomaternalcare by female wedge-capped capuchin mon-keys: effects of age, rank and relatedness. Behav199:30–50.
63 Perry S. 1996. Female-female social rela-tionships in wild white-faced capuchin mon-keys, Cebus capucinus. Am J Primatol 40:167–182.
64 Fragaszy DM, Baer J, Adams-Curtis L. 1991.Behavioural development and maternal care intufted capuchin (Cebus apella) and squirrelmonkeys (Saimiri sciureus) from birth throughseven months. Dev Psychobiol: 375–394.
65 Baldovino MC, Bitetti MS. 2008. Allonursingin tufted capuchin monkeys (Cebus nigritus):milk or pacifier? Folia Primatol 79:79–92.
66 Fedigan L, Carnegie SD, Jack KM. 2008.Predictors of reproductive success in femalewhite-faced capuchins (Cebus capucinus). Am JPhys Anthropol: 82–90.
67 de Waal FBM, Leimgruber K, GreenbergAR. 2008. Giving is self-rewarding for monkeys.Proc Natl Acad Sci USA 105:13685–13689.
68 Lakshminarayanan VR, Santos LR. 2008.Capuchin monkeys are sensitive to others’ wel-fare. Curr Biol 18:R999–R1000.
69 Rapaport LG. 2006. Parenting behaviour:babbling bird teachers? Curr Biol 16:R675–R677.
70 Rapaport LG, Brown RB. 2008. Social influ-ences on foraging behavior in young nonhumanprimates: learning what, where, and how to eat.Evol Anthropol 17:189–201.
71 Caro TM, Hauser MD. 1992. Is there teach-ing in nonhuman animals? Q Rev Biol 67:151–172.
72 Call J, Tomasello M. 2008. Does the chim-panzee have a theory of mind? 30 years later.Trends Cogn Sci 12:187–192.
73 Cronin KA, Snowdon CT, Silk JB. 2008. Per-formance of cottontop tamarins (Saguinus oedi-pus) on a food donation task Primate Eye 96(Special Issue: Abstracts of the XXII Congressof IPS):17.
74 Warneken F, Hare B, Melis AP, Hanus D,Tomasello M. 2007. Spontaneous altruism bychimpanzees and young children. Plos Biol5:e184, 1–7.
75 Melis AP, Hare B, Tomasello M. 2006. Chim-panzees recruit the best collaborators. Science311:1297–1300.
76 Mitani JC. 2006. Reciprocal exchange inchimpanzees and other primates. In: KappelerPM, van Schaik CP, editors. Cooperation in pri-mates and humans: mechanisms and evolution.Berlin: Springer. p 107–119.
77 Stevens RC. 2003. The selfish nature of gen-erosity: harrassment and food sharing in prima-tes. Proc R Soc B 271:451–456.
78 de Waal FBM. 2008. Putting the altruismback into altruism: the evolution of empathy.Annu Rev Psychol 59:279–300.
79 De Waal FBM. 1996. Good natured: theorigins of right and wrong in humans andother animals. Cambridge: Harvard UniversityPress.
80 Coussi-Korbel S, Fragaszy DM. 1995. On therelation between social dynamics and sociallearning. Anim Behav 50:1441–1453.
81 Kumashiro M, Ishibashi H, Uchiyama Y,Itakura S, Murata A, Iriki A. 2003. Natural imi-tation induced by joint attention in Japanesemonkeys. Int J Psychophysiol 50:81–99.
82 Rizzalotti G, Fadiga L, Fogassi L, Gallese V.2002. From mirror neurons to imitation: factsand speculation. In: Meltzoff AN, Prinz W, edi-
ARTICLES Cooperative Breeding and Human Cognitive Evolution 185

tors. The imitative mind: development, evolu-tion, and brain bases. Cambridge: CambridgeUniversity Press. p 247–266.
83 van Schaik CP. 2006. Why are some animalsso smart? Sci Am 294:64–71.
84 Whiten A, van Schaik CP. 2007. The evolu-tion of animal ‘‘cultures’’ and social intelligence.Philos Trans R Soc B 362:603–620.
85 van Schaik CP, Burkart JM. n.d. Why cul-ture makes smart: the cultural intelligence hy-pothesis. Ethology.
86 Isler K, van Schaik CP. 2009. Why are thereso few smart mammals (but so many smartbirds)? Biol Lett 5:125–129.
87 Isler K, van Schaik CP. n.d. The expensivebrain: a framework for explaining evolutionarychanges in brain size. J Hum Evol. In press.
88 Barrickman NL, Bastian ML, Isler K, vanSchaik CP. 2008. Life history costs and benefitsof encephalization: a comparative test usingdata from long-term studies of primates in thewild. J Hum Evol 54:568–590.
89 Hare B, Brown M, Williamson C, TomaselloM. 2002. The domestication of social cognitionin dogs. Science 298:1634–2636.
90 Topal J, Gacsi M, Miklosi A, Viranyi Z,Kubinyi E, Csanyi V. 2005. Attachment tohumans: a comparative study on hand-rearedwolves and differently socialized dog puppies.Anim Behav 70:1367–1375.
91 Baker PJ, Harris S. 2004. Red foxes: thebehavioural ecology of red foxes in urban Bris-tol. In: Macdonald DW, Sillero-Zubiri C, edi-tors. Biology and conservation of wild canids.New York: Oxford University Press. p 207–216.
92 Macdonald DW. 1979. ‘‘Helpers’’ in fox soci-ety. Nature 282:69–71.
93 Trut L. 1999. Early canid domestication: thefarm-fox experiment. Am Sci 87:160–169.
94 Russon AE. 2004. Great ape cognitive sys-tems. In: Russon AE, Begun DR, editors. Theevolution of thought: evolutionary origins ofgreat ape intelligence. Cambridge: CambridgeUniversity Press. p 76–99.
95 van Schaik CP, Deaner RO, Merrill MY.1999. The conditions for tool use in primates:implications for the evolution of material cul-ture. J Hum Evol 36:719–741.
96 Visalberghi E, Fragaszy DM, Savage-Rum-baugh S. 1995. Performance in a tool-using taskby common chimpanzees (Pan troglodytes),bonobos (Pan paniscus), an orangutan (Pongopygmaeus), and capuchin monkeys (Cebusapella). J Comp Psychol 109:52–60.
97 Mulcahy NJ, Call J. 2006. How great apes
perform on a modified trap-tube task. AnimCogn 9:193–199.
98 Mulcahy NJ, Call J. 2006. Apes save tools forfuture use. Science 312:1038–1040.
99 Boesch C, Boesch H. 1984. Mental map inwild chimpanzees: an analysis of hammer trans-ports for nut cracking. Primates 25:160–170.
100 Rapaport LG. 2006. Provisioning in wildgolden lion tamarins (Leontopithecus rosalia):benefits to omnivorous young. Behav Ecol17:212–221.
101 Cheney DL, Seyfarth RM. 2007. Baboon met-aphysics. Chicago: University of Chicago Press.
102 Premack D. 2007. Human and animal cog-nition: continuity and discontinuity. Proc NatlAcad Sci USA 104:13861–13867.
103 Hare B. 2001. Can competitive paradigmsincrease the validity of experiments on primatesocial cognition? Anim Cogn 4:269–280.
104 Gergely G, Csibra G. 2005. The social con-struction of the cultural mind: Imitative learn-ing as a mechanism of human pedagogy. Inter-action Stud 6:463–481.
105 De Waal FBM. 2008. Putting the altruismback into altruism: The evolution of empathy.Annu Rev Psychol 59:4.1–4.22.
106 Brosnan SF, Schiff HC, De Waal FBM.2005. Tolerance for inequity may increase withsocial closeness in chimpanzees. Proc R Soc B272:253–258.
107 Brauer J, Call J, Tomasello M. 2006. Are apesreally inequity averse? Proc R Soc B 273:2123–3128.
108 Burkart JM, Heschl A. 2007. Perspectivetaking or behaviour reading? Understanding ofvisual access in common marmosets (Calithrixjacchus). Anim Behav 73:457–469.
109 Savage-Rumbaugh S, Shanker SG, TaylorJT. 1998. Apes, language, and the human mind.New York: Oxford University Press.
110 Tomasello M, Carpenter M. 2007. Theemergence of social cognition in three youngchimpanzees. Boston: Blackwell.
111 Leavens DA, Hopkins WD. 1998. Inten-tional communication by chimpanzees: a cross-sectional study of the use of referential ges-tures. Dev Psychol 34:813–822.
112 Tomasello M, Carpenter M, Liszkowski U.2007. A new look at infant pointing. Child Dev78:705–722.
113 Tomasello M. 1999. The cultural origins ofhuman cognition. Cambridge MA: Harvard Uni-versity Press.
114 Carpendale J, Lewis C. 2006. How childrendevelop social understanding. Oxford: Blackwell.
115 Tomasello M. 2003. Constructing a language:a usage-based theory of language acquisition.Cambridge MA: Harvard University Press.
116 Richardson DC, Dale R, Krickham NZ.2007. The art of conversation is coordination.Psychol Sci 18:407–413.
117 Zahavi A. 1975. Mate selection: a selectionfor a handicap. J Theor Biol 53:205–214.
118 Grice HP. 1975. Logic and conversation.In: Cole P, editor. Syntax and semantics, vol 3.New York: Academic Press. p 41–58.
119 Hurford J. 2007. The origins of meaning:language in the light of evolution. Oxford:Oxford University Press.
120 Fitch WT. 2004. Kin selection and ‘‘mothertongues’’: a neglected component in languageevolution. In: Oller DK, Griebel U, editors. Theevolution of communication systems: a compar-ative approach. Cambridge: MIT Press. p 275–296.
121 Ramsey G, Bastian ML, van Schaik CP.2007. Animal innovation defined and operation-alized. Behav Brain Sci 30:393–437.
122 Whiten A, Goodall J, McGrew WC, NishidaT, Reynolds V, Sugiyama Y, Tutin CEG, Wrang-ham RW, Boesch C. 1999. Cultures in chimpan-zees. Nature 399:682–685.
123 van Schaik CP, Ancrenaz M, Borgen G,Galdikas B, Knott CD, Singleton I, Suzuki A,Utami SS, Merrill M. 2003. Orangutan culturesand the evolution of material culture. Science299:102–105.
124 Wilson DS, Timmel JJ, Miller RR. 2004.Cognitive cooperation: when the going getstough, think as a group. Hum Nat 15:1–26.
125 van Schaik CP, Burkart JM. n.d. Mind thegap: cooperative breeding and the evolution ofour unique features. In: Kappeler PM, Silk J,editors. Mind the gap: tracing the origins ofhuman universals. In press.
126 Details in prep for 2010 posting at http://www.alltheworldsprimates.com/contributor/Social.aspx.
127 Walker A, Shipman P. 1996. The wisdomof the bones. New York: Alfred Knopf.
128 Behne T, Carpenter M, Grafenhain M, Lie-bal K, Liszkowski U, Moll H, Rakoczy H, Toma-sello M, Warneken F, Wyman E. 2008. Culturallearning and creation. In: Muller U, CarpendaleJ, Budwig N, Sokol B, editors. Social life andsocial knowledge: toward a process account ofdevelopment. Mahwah, NJ: Erlbaum. p 65–101.
VVC 2009 Wiley-Liss, Inc.
ERRATUM
In Volume 16, Issue 5 (2007) of Evolutionary Anthropology ‘‘Evolutionary transformation of the hominin shoulder,’’by Susan G. Larson. Figure 7, displaying box and whisker plots of humeral torsion for comparative samples ofapes, modern humans, and fossil, the torsion estimates for KNM-ER 739 and Omo 119-73-2718 had been inadver-tently switched.
186 Burkart et al. ARTICLES