control of human viral infections by natural killer cells · simplex virus (hsv), ... hspg, heparan...
TRANSCRIPT

IY31CH07-Altfeld ARI 14 February 2013 21:16
Control of Human ViralInfections by NaturalKiller CellsStephanie Jost and Marcus AltfeldRagon Institute of MGH, MIT and Harvard, Boston, Massachusetts 02129;email: [email protected], [email protected]
Annu. Rev. Immunol. 2013. 31:163–94
First published online as a Review in Advance onJanuary 3, 2013
The Annual Review of Immunology is online atimmunol.annualreviews.org
This article’s doi:10.1146/annurev-immunol-032712-100001
Copyright c© 2013 by Annual Reviews.All rights reserved
Keywords
HIV-1, HCV, influenza, CMV, KIR, NKG2D, NK cells
Abstract
Natural killer (NK) cells are effector cells of the innate immune systemand are important in the control of viral infections. Their relevance isreflected by the multiple mechanisms evolved by viruses to evade NKcell–mediated immune responses. Over recent years, our understandingof the interplay between NK cell immunity and viral pathogenesis hasimproved significantly. Here, we review the role of NK cells in thecontrol of four important viral infections in humans: cytomegalovirus,influenza virus, HIV-1, and hepatitis C virus.
163
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
INTRODUCTION
Natural killer (NK) cells are large granular lym-phocytes that play a central role in the controlof viral infections. NK cells were initially iden-tified through their ability to mediate cellularcytotoxicity against tumor cells (1). Subsequentstudies illustrating activation of NK cells inresponse to type I interferons (IFNs) alsoidentified them as a component of the innateimmune response against pathogens. The con-tribution of NK cells to the antiviral immuneresponse has been extensively studied in mousemodels of viral infections, demonstrating thatNK cells not only contain viral replication bykilling infected cells during the earliest stages ofinfection, prior to the development of adaptiveimmunity, but also play a critical immunoregu-latory role during the development of adaptiveimmunity (2, 3). Although NK cells areconsidered part of the innate immune systemand mediate their effector functions througha number of germ-line-encoded receptors, re-cent studies have suggested that a subset of NKcells in mice can mediate immunological recallresponses to a variety of different haptens andviral pathogens (4–6). These data indicate thatNK cells might comprise a more heterogeneouscell population than initially assumed.
In humans, several rare NK cell deficiencieshave been described (7). Biron et al. (8) first re-ported a case of a young girl who lacked func-tional NK cells and experienced a series of viralinfections during childhood and adolescence,including infections by multiple herpes viruses.Several subsequent studies have associated hu-man immunodeficiency syndromes that resultin complete or partial impairment of NK cellnumbers and functions with an increased sus-ceptibility to viral infections, including herpessimplex virus (HSV), varicella zoster virus, cy-tomegalovirus (CMV), and human papillomavirus, as well as more severe and progressivediseases during childhood (9–11). These stud-ies established the critical role of functionalNK cells in the control of viral infections inhumans. Here, we review the mechanisms bywhich NK cells participate in the control of viralinfections, with particular focus on four major
human viral infections: CMV, influenza virus,hepatitis C virus (HCV), and human immuno-deficiency virus-1 (HIV-1).
NK CELLS AND THEIRRECEPTORS IN MICEAND HUMANS
NK cells are generally described as large lym-phocytes that lack the expression of the T cellreceptor (i.e., CD3 negative), but there is nounifying receptor that identifies all NK cells indifferent species. NK1.1 can be used to isolateNK cells in some mouse strains, but not all, inwhich case CD49b (DX5) represents an alterna-tive. Murine NK cells can be further subdividedinto four subsets with distinct maturation levelsaccording to their surface expression of CD27and CD11b (12). Human NK cells have beenclassically defined as CD3−CD56+ lympho-cytes, representing about 15% of peripheralblood lymphocytes. CD3−CD56+ NK cellscan be further subdivided into CD56bright NKcells, which lack the expression of CD16 (Fcγ-receptor) and KIR (killer immunoglobulin-likereceptor), and CD56dim NK cells, whichexpress CD16 and KIR (1, 13). In addition,a subset of CD56−CD16+ NK cells appearsto be expanded in chronic viral infections andseems to represent an exhausted/anergic subsetof NK cells (14, 15).
The most prominent function of NK cellsis to lyse malignant or virally infected cells.NK cells mediate cytolysis by a number of dif-ferent mechanisms, including (a) exocytosis ofcytoplasmic granules containing perforin andgranzyme, (b) Fas ligand–mediated induction ofapoptosis, and (c) antibody-dependent cellularcytotoxicity (ADCC). NK cells, particularlyCD56bright NK cells, also play an importantrole as immune regulatory cells, bridging theinnate and adaptive immune responses as theyproduce soluble factors including cytokinesand chemokines (16). In addition, NK cells candirectly modulate adaptive immune responses,mainly via the interaction with dendritic cells(reviewed in Reference 2). The precise mech-anisms by which NK cells recognize virally
164 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.
IY31CH07-Altfeld ARI 14 February 2013 21:16
infected target cells are complex and still notentirely understood. Unlike B and T cells,NK cells do not appear to express uniqueclonally distributed receptors for specificantigens, but they do express an arsenal ofdifferent stimulatory and inhibitory receptors
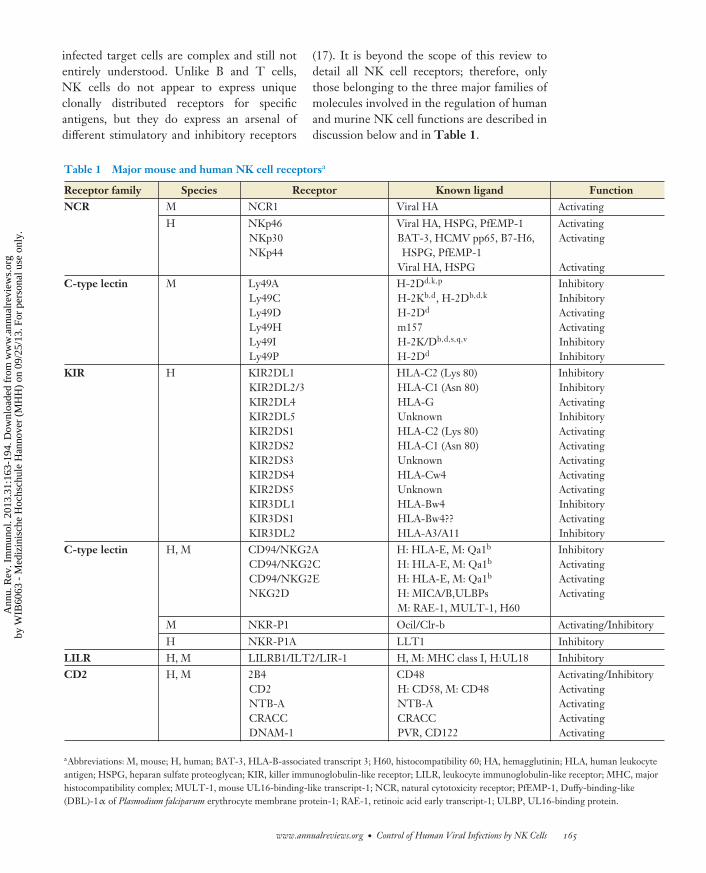
(17). It is beyond the scope of this review todetail all NK cell receptors; therefore, onlythose belonging to the three major families ofmolecules involved in the regulation of humanand murine NK cell functions are described indiscussion below and in Table 1.
Table 1 Major mouse and human NK cell receptorsa
Receptor family Species Receptor Known ligand FunctionNCR M NCR1 Viral HA Activating
H NKp46NKp30NKp44
Viral HA, HSPG, PfEMP-1BAT-3, HCMV pp65, B7-H6,HSPG, PfEMP-1
Viral HA, HSPG
ActivatingActivating
ActivatingC-type lectin M Ly49A
Ly49CLy49DLy49HLy49ILy49P
H-2Dd,k,p
H-2Kb,d, H-2Db,d,k
H-2Dd
m157H-2K/Db,d,s,q,v
H-2Dd
InhibitoryInhibitoryActivatingActivatingInhibitoryInhibitory
KIR H KIR2DL1KIR2DL2/3KIR2DL4KIR2DL5KIR2DS1KIR2DS2KIR2DS3KIR2DS4KIR2DS5KIR3DL1KIR3DS1KIR3DL2
HLA-C2 (Lys 80)HLA-C1 (Asn 80)HLA-GUnknownHLA-C2 (Lys 80)HLA-C1 (Asn 80)UnknownHLA-Cw4UnknownHLA-Bw4HLA-Bw4??HLA-A3/A11
InhibitoryInhibitoryActivatingInhibitoryActivatingActivatingActivatingActivatingActivatingInhibitoryActivatingInhibitory
C-type lectin H, M CD94/NKG2ACD94/NKG2CCD94/NKG2ENKG2D
H: HLA-E, M: Qa1b
H: HLA-E, M: Qa1b
H: HLA-E, M: Qa1b
H: MICA/B,ULBPsM: RAE-1, MULT-1, H60
InhibitoryActivatingActivatingActivating
M NKR-P1 Ocil/Clr-b Activating/Inhibitory
H NKR-P1A LLT1 InhibitoryLILR H, M LILRB1/ILT2/LIR-1 H, M: MHC class I, H:UL18 InhibitoryCD2 H, M 2B4
CD2NTB-ACRACCDNAM-1
CD48H: CD58, M: CD48NTB-ACRACCPVR, CD122
Activating/InhibitoryActivatingActivatingActivatingActivating
aAbbreviations: M, mouse; H, human; BAT-3, HLA-B-associated transcript 3; H60, histocompatibility 60; HA, hemagglutinin; HLA, human leukocyteantigen; HSPG, heparan sulfate proteoglycan; KIR, killer immunoglobulin-like receptor; LILR, leukocyte immunoglobulin-like receptor; MHC, majorhistocompatibility complex; MULT-1, mouse UL16-binding-like transcript-1; NCR, natural cytotoxicity receptor; PfEMP-1, Duffy-binding-like(DBL)-1α of Plasmodium falciparum erythrocyte membrane protein-1; RAE-1, retinoic acid early transcript-1; ULBP, UL16-binding protein.
www.annualreviews.org • Control of Human Viral Infections by NK Cells 165
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
Table 2 Associations between KIR/HLA interactions and human viral diseases
Infection KIR/HLA association Observation Reference(s)HIV KIR3DS1 and HLA-Bw4-80I
KIR3DL1high and HLA-Bw4-80ISlower progression to AIDSSlower progression to AIDS
7071
KIR3DS1 Protection from infection 73, 78HCV KIR2DL3 and HLA-C1 homozygosity Resolution of infection 126
KIR3DS1 and HLA-Bw4-80I Protection against development of HCC 138CMV Fewer than one activating KIR in donor Reduced risk of CMV reactivation in
recipient following bone marrowtransplant
52, 53
KIR2DL1 expression on NK cells Recurrent CMV infection 51Influenza virus KIR2DL2/3 and HLA-C1
KIR3DL1/S1 and HLA-Bw6 homozygositySevere influenza infection 198
KIR2DL1 and HLA-C1 homozygosityKIR3DS1, KIR2DS5, KIR2DL5
199
Abbreviations: CMV, cytomegalovirus; HCC, hepatocellular carcinoma; HCV, hepatitis C virus; HIV, human immunodeficiency virus; HLA, humanleukocyte antigen; KIR, killer immunoglobulin-like receptor.
RECEPTORS THAT CONTROLNK CELL FUNCTION
Natural Cytotoxicity Receptors
The majority of human NK cells constitutivelyexpress NKp46 and NKp30, two activating re-ceptors that belong to the family of natural cy-totoxicity receptors (NCRs). NCRs comprise athird member, NKp44, which is only detectedon the cell surface upon IL-2-mediated NK cellactivation (18). The precise ligands for thesereceptors remain largely undefined, yet severalviral- or tumor-associated molecules that caninteract with NCRs have been identified (re-viewed in Reference 19) (Table 1). Of partic-ular interest, NKp46 and NKp44 can interactwith influenza hemagglutinin (HA) and medi-ate cytolysis of influenza-infected NK cells (20,21). Only NKp46 has a mouse ortholog, NCR1,which seems to share the same antiviral and an-titumoral properties as its human counterpart.
Ly49 Receptors and KillerImmunoglobulin-Like Receptors
Two large families of structurally distinct butfunctionally similar receptors in human andmice play an important role in regulating NKcell function. The C-type lectin-like Ly49 re-ceptors in mice and the killer immunoglobulin-
like receptors (KIRs) in humans share crucialcharacteristics, including common signalingpathways, that can lead to inhibitory oractivating outcomes depending on the trans-membrane residues or cytoplasmic motifs in thereceptors, and the ability to primarily recognizemajor histocompatibility complex (MHC) classI and related surface molecules (reviewed inReference 22). Activation or inhibition of NKcells is significantly affected by the strengthof the binding between each Ly49 and KIRand their respective ligands, which itself candepend on the MHC class I–presented peptideand, at least in the case of Ly49 receptors, onthe existence of cis rather than trans interactionswith their ligands. Some of these molecules,in conjunction with their ligands, have beenassociated with protection against viral infec-tion and other diseases (Table 2) and are thusmore extensively described in the followingparagraph.
Ly49 receptors bind specific allotypes ofH2D and H2K murine MHC class I molecules(22) (Table 1). One of these receptors, theactivating Ly49H, does not have any knownMHC class I ligand but has a high affin-ity for the murine cytomegalovirus (MCMV)protein m157 (reviewed below). KIRs inter-act with specific allotypes of the classical (-A,-B, -C) and nonclassical (-G) human leukocyte
166 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
antigen (HLA) class I receptors. In particular,KIR3DL1, and potentially KIR3DS1, binds toHLA-B molecules that carry a distinctive sero-logical motif, w4, that can be distinguished fromthe w6 motif on the basis of amino acids 77to 83 (23). HLA-Bw6 molecules do not serveas known ligands for any KIR (24). Other re-ceptors involved in the control of human vi-ral infections are KIR2DL1, KIR2DL2, andKIR2DL3, all of which bind HLA-C molecules.HLA-C alleles can carry either an Asn at posi-tion 80, and are classified as group 1 (HLA-C1),or a Lys, and then belong to group 2 (HLA-C2). KIR2DL2/3 preferentially bind HLA-C1,and KIR2DL1 preferentially binds HLA-C2,with the binding affinity of the latter combi-nation being stronger than the former. Gen-erally, engagement of inhibitory Ly49/KIRs totheir MHC class I ligands protects cells fromNK cell–mediated lysis, and engagement of ac-tivating Ly49/KIRs provides an activating sig-nal. Nevertheless, thus far only KIR2DS1 hasbeen shown to interact directly with HLA-C,and ligands for the remaining activating KIRsstill remain to be identified (25). Finally, genesencoding Ly49 and KIR receptors display agreat diversity in number and sequence poly-morphisms and are expressed in a stochasticmanner on NK cells, leading to unique subsetsof NK cells with distinct properties and activa-tion thresholds. Expression of Ly49 and KIRnot only depends on the repertoire of genespresent in the genome, which altogether forma haplotype, but also seems to be influenced bythe presence of the MHC ligand (26).
NKG2 Receptors
Murine and human NK cells also expressCD94-NKG2A/C/E heterodimers, whichbelong to another family of C-type lectins.These highly conserved receptors can de-liver activating or inhibitory signals. Humaninhibitory CD94-NKG2A and activatingCD94-NKG2C/E heterodimers bind the non-classical HLA-E loaded with peptides derivedfrom other HLA class I molecules, therebymonitoring the overall expression level of
HLA class I (Table 1). The murine receptorsexert a similar function by binding to Qa1b.The activating receptor NKG2D is highlyexpressed on human and murine NK cells andis distinct from NKG2A/C/E based on se-quence alignments with other members of thesuperfamily and because it forms a homodimerinstead of a heterodimer with CD94. It existsin a long and a short isoform in mice, favoringcytotoxicity and cytokine production, respec-tively, upon stimulation, but only the longform of NKG2D is present in humans. HumanNKG2D recognizes human cytomegalovirus(HCMV) glycoprotein UL16-binding proteins(ULBPs) and the MHC class I chain–relatedmolecules MICA and MICB. Ligands for themouse NKG2D include the retinoic acid earlytranscript-1 (RAE-1) molecules α, β, γ, δ,and ε, mouse UL16-binding-like transcript-1(MULT-1) molecules, and histocompatibility60 (H60) molecules. All the receptors recog-nized by NKG2D are typically expressed bystressed or malignant cells, and viruses haveevolved multiple mechanisms to evade recogni-tion by NKG2D+ NK cells (reviewed below).
NK CELLS IN CMV INFECTION
HCMV is a large double-stranded DNA virusof the subfamily Betaherpesvirinae and ishighly prevalent in humans, with 50–90% ofindividuals in the United States, and as manyas 100% of individuals in developing countries,being seropositive. Infection is usually acquiredearly in life, and most immunocompetent hostsare asymptomatic. However, CMV can causesevere disease in immunocompromised individ-uals, such as patients with AIDS or recipientsof transplants on immunosuppressive therapy,resulting from the reactivation of latent infec-tion. Furthermore, maternal CMV reactivationor primary infection during pregnancy causescongenital CMV, the most common congenitalinfection in the developed world, and is asso-ciated with long-term sequelae such as hearingloss and developmental delay in about 50% ofthese infants (27). HCMV establishes latentpersistent infection in monocyte precursors
www.annualreviews.org • Control of Human Viral Infections by NK Cells 167
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
and tissue stroma cells, and persistent immunecontrol is thought to be responsible for thecontrol of the latent infection. HCMV hascoevolved with its host for millions of years(28), resulting in the evolution of a largenumber of genes that prevent recognition ofinfected cells by the immune system, includingNK cells, and enable the virus to establishpersistent infection and dissemination.
NK Cells in the Mouse Model ofMCMV Infection
Because CMV infection is species specific,no good animal models for HCMV infectioncurrently exist. As a result, most animal studiesof CMV infection have been performedusing MCMV in mice. Although MCMVand HCMV have similar tissue tropisms andsimilar pathogenesis, they have developed dif-ferences in the mechanisms they use to evadehost-specific immunity (reviewed in Reference29). This is reflected by significant primarygenomic sequence diversity and less than50% homology in the open reading framesbetween MCMV and HCMV. Furthermore,the highest sequence diversity is observed inthose regions of CMV that encode for genesdedicated toward immune evasion. The timeto complete a life cycle also distinguishes theseviruses, with MCMV replicating significantlyfaster (approximately 30 h) than HCMV(72–96 h). Nevertheless, studies focusing onMCMV have provided critical insights into thebasic mechanisms by which CMV evades thehost’s immune response (30).
In an effort to avoid recognition of MHCclass I–presented viral epitopes by CD8+ Tcells, CMV encodes several gene products thatinterfere with the presentation of viral pep-tides (reviewed in References 29 and 30). TheMCMV proteins m04, m06, and m152 are in-volved in the disruption of the MHC class Isynthesis pathway, resulting in the retention ordegradation of MHC molecules, and modulateantigen presentation. Although the reductionof MHC class I on the cell surface can allowevasion from virus-specific CD8+ T cells, it
also renders infected cells more susceptible torecognition by NK cells, owing to the loss of lig-ands for inhibitory NK cell receptors. To com-pensate for this, MCMV possesses several genesthat encode for MHC class I–like molecules andcan engage inhibitory Ly49 receptors, therebyproviding an inhibitory signal to NK cells.These MHC class I–like molecules includem157 and m144, yet in the case of m144, thecognate Ly49 ligand and the precise mechanismof action have not been identified. MCMV in-fection also results in the expression of stressmolecules on infected cells that serve as ligandsfor activating NK cell receptors, particularlyNKG2D ligands (reviewed in References 29and 30) (Table 1). Several of the MCMV pro-teins interfere with the expression of NKG2Dligands, including m152, which reduces the ex-pression of the RAE-1 gene products; m145,which downregulates MULT-1; m155, whichreduces H60 expression; and m138, which ap-pears to reduce the expression of several ofthese NKG2D ligands. Overall, MCMV ded-icates multiple gene products toward evasionfrom recognition by NKG2D+ NK cells.
In the process of coevolution with MCMV,some mouse NK cell receptors appear to havebeen selected for their ability to specificallyrecognize MCMV proteins expressed oninfected cells. Genes encoding these receptors(Cmv1r) provide resistance to otherwise lethalMCMV infection and are found in C57BL/6and MA/My strains of mice. In C57BL/6 mice,the activating Ly49H receptor on NK cellsconfers resistance to MCMV (31–33). NKcells from Ly49H-deficient C57BL/6 mice areunable to respond to MCMV, and these miceare susceptible to infection. Ly49H+ NK cellsrecognize the MCMV-encoded glycoproteinm157, which is expressed on infected cells. Insupport of this critical role of Ly49H-mediatedrecognition of m157 in the immune protectionfrom MCMV, mutants that lack m157 aresignificantly more virulent in C57BL/6 mice(34). In addition to serving as a ligand forthe activating Ly49H receptor, m157 is anMHC class I homolog that compensates forMHC class I downregulation. For example,
168 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
m157 is recognized by high-affinity inhibitoryLy49I receptors expressed on NK cells in somestrains of mice (33), potentially explaining whythis MCMV gene product has persisted duringevolution despite the presence of an activatingreceptor directed against it. Finally, the evolu-tion of an antigen-specific receptor recognizingm157 in C57BL/6 mice appears to have enabledmice to develop NK cell memory responses toMCMV, as described by Sun et al. (6).
Similar to Ly49H in C57BL/6 mice, Ly49Pprovides resistance to MCMV infection inMA/My mice that do not possess the Ly49Hgene (35). In contrast to Ly49H, the Ly49P geneis not sufficient to confer resistance but needs tobe encoded together with the MHC class I genefor H2-Dk. Functional studies showed that theLy49P response to MCMV is restricted by H2-Dk and can be blocked by antibodies againstH2-Dk. Furthermore, it was dependent on theMCMV-encoded m04 protein, as m04-deletedmutants abrogated resistance to MCMV infec-tion in MA/My mice (36). However, the precisemechanisms by which Ly49P recognizes m04 inthe context of H2-Dk are not understood, as ad-ditional elements conferred by MCMV infec-tion are required for this interaction. Taken to-gether, these detailed studies of the interactionbetween MCMV-encoded gene products andactivating and inhibitory murine NK cell recep-tors have identified multiple immune recogni-tion and immune evasion pathways and provideimportant insights into the possible strategiesevolved in humans to control HCMV infection.
NK Cells in HCMV Infection
As described above, NK cells play a critical rolein the protection from herpesvirus infection inhumans. Similar to MCMV, HCMV encodesfor several gene products that enable it toevade the immune response in its natural host(reviewed in References 29 and 30). HCMVdownregulates HLA class I expression by usingseveral different proteins in an effort to evadeCMV-specific CD8+ T cell responses. US2and US11 shuttle HLA class I from the endo-plasmic reticulum to the cytoplasm, resulting
in HLA degradation; US3 causes retentionof HLA class I in the endoplasmic reticulum;and US6 blocks the TAP-dependent transportof processed peptides into the endoplasmicreticulum for loading on HLA class I. HCMValso encodes for several HLA class I homologsthat engage inhibitory KIRs to avoid NK cellactivation. Additional HCMV proteins serve asligands for other inhibitory NK cell receptors,such as UL18, which binds to the inhibitoryleukocyte immunoglobulin-like receptor1 (LIR-1) expressed on NK cells, therebydampening LIR-1+ NK cell function (37, 38).
Furthermore, HCMV infection results inthe upregulation of stress molecules, includingMICA, MICB, and ULBPs, on human cells.These molecules serve as ligands for the acti-vating NKG2D receptor and trigger strong ac-tivation of NKG2D+ NK cells (29, 30). HCMVexpresses several proteins that downregulatethe expression of NKG2D ligands, includingUL16, which reduces the expression of MICBand ULBP1/2; UL142, which reduces the ex-pression of MICA; and several additional pro-teins (UL18, UL40, UL83, UL141) (as re-viewed in Reference 30). In addition, HCMVencodes for microRNAs that target transcriptsof NKG2D ligands for degradation. Mandel-boim and colleagues (39, 40) demonstrated thatthe HCMV-encoded microRNA miR-UL112downregulates the expression of MICB on in-fected cells, leading to decreased binding ofNKG2D and reduced killing by NK cells. Thedescription of viral microRNAs that participatein evading immune recognition has opened anew area of immune evasion pathways that iscurrently under intense investigation.
Whereas specific NK cell receptors thatrecognize MCMV proteins have been foundin mice, no such receptors have been identifiedin humans to date. HCMV encodes a peptidewithin the UL40 protein that is similar to thosefound in many HLA class I molecules and leadsto the upregulation of HLA-E on HCMV-infected cells, resulting in the inhibition ofNKG2A+ NK cells (41). In contrast, NK cellsexpressing the activating counterpart NKG2C,which binds to HLA-E with less affinity than
www.annualreviews.org • Control of Human Viral Infections by NK Cells 169
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
NKG2A does, expand in response to HCMVinfection. Cross-sectional studies showedthat NKG2C+ NK cells, accounting for lessthan 2% of NK cells in CMV-seronegativeindividuals, can represent more than 25% ofperipheral NK cells in individuals infected withCMV (42, 43). More recently, Lopez-Vergeset al. (44) characterized the changes in NKcells longitudinally in a group of individualswith acute CMV infection and showed apreferential expansion of NKG2C+ NK cellsthat eventually acquired the expression ofthe maturation marker CD57 but lackedthe expression of the inhibitory receptorsNKG2A and KIR3DL1 (in those subjectsalso encoding for the ligand HLA-Bw4).Furthermore, expansion kinetics of NKG2C+
NK cells has been studied in individuals thatreceived umbilical cord blood transplantationand subsequently reactivated CMV infection.Della Chiesa et al. (45) observed an expansionof NKG2C+ NK cells following HCMVreactivation that was associated with a morerapid acquisition of the mature KIR+NKG2A−
NK cell phenotype. Similarly, Foley et al. (46)described an expansion of NKG2C+CD57+
NK cells after CMV reactivation in adultsreceiving hematopoietic stem cell transplanta-tion. Taken together, these studies consistentlydescribe the expansion of NKG2C+ NK cellsin response to CMV and demonstrate thatthese NKG2C+ NK cells acquire a moredifferentiated and mature phenotype.
However, the precise mechanism by whichthis subset of NK cells recognizes HCMVremains unknown, and there is currently noevidence that NKG2C directly binds to anHCMV-encoded protein. Furthermore, it re-mains unclear whether these mature NK cellsrepresent a subset of memory NK cells inhumans, similar to the recent description ofMCMV-specific memory NK cells expressingLy49H in mice (6). In this context, it is of inter-est that a rapid expansion and long-term per-sistence of NKG2C+ NK cells were also ob-served following human hantavirus infection(47). However, this expansion was less impor-tant in the three individuals that were HCMV
negative. Similarly, an expansion of NKG2C+
NK cells was observed in HIV-1 infection (48)and chikungunya virus infection (49), but againonly in those individuals also infected withHCMV, whereas such an expansion was not ob-served in recurrent HSV-2 infection (50). It re-mains to be elucidated whether prior exposureto HCMV primes NKG2C+ NK cells to ex-pand more rapidly in response to other viruses,or whether different viral infections can trig-ger the expansion of these NKG2C+ NK cells.Finally, some studies have suggested that spe-cific KIR genotypes are associated with protec-tion from CMV reactivation (51–53) (Table 2);however, their functional consequences remainunclear. Overall, CMV has developed severalevasion strategies against NK cell–mediatedimmune pressure that have helped identify theprincipal mechanisms by which NK cells rec-ognize virally infected cells.
NK CELLS IN HIV-1 INFECTION
HIV-1 is a retrovirus that has infected morethan 60 million individuals since its initialdescription in the mid-1980s and has killedmore than 30 million individuals. Although allparts of the world are affected by the epidemic,most infected individuals live in sub-SaharanAfrica. HIV-1 is transmitted mainly throughmucosal routes during sexual exposure byan infected partner, by sharing needles withinfected individuals during intravenous druguse, and from mother to child at the time ofbirth and through breast-feeding. Followingacute infection, the virus establishes a chronicinfection and a persistent viral reservoir.HIV-1 preferentially infects CD4+ T cellsand macrophages and eventually leads to theloss of CD4+ T cells and the developmentof AIDS in most infected individuals in theabsence of antiretroviral therapy. However,the speed of HIV-1 disease progression varieslargely between infected individuals, with someindividuals developing AIDS within one year ofinfection, whereas others remain healthy withno manifestations of HIV-1 disease even aftermore than 25 years of infection. Apart fromdifferences in viral virulence and host genetic
170 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
factors, differences in immune responsesto the virus have been associated with theheterogeneity of HIV-1 disease outcome (54).
NK Cells in Animal Modelsof HIV-1 Infection
HIV-1 infects humans, and there is currentlyno good animal model for HIV-1 infection.However, several groups have developedhumanized mouse models, including irradiatedNOD/SCID/IL-2Rγc−/− mice transplantedwith human fetal liver hematopoietic stemcells and autologous thymic grafts (55), andthese models for HIV-1 infection have becomemore robust over the past few years (56). Theprecise role of different immune cell subsets,including NK cells, in the restriction of HIV-1replication in these mouse models remains tobe determined, but these models will probablybecome increasingly important tools for study-ing immune responses to HIV-1 in upcomingyears. In the absence of a well-characterizedanimal model for HIV-1 infection, simian im-munodeficiency virus (SIV) infection of rhesusmonkeys has been used to study AIDS viruspathogenesis in nonhuman primates and to testvaccine efficacy (57). SIV infection of rhesusmonkeys leads to an acute infection with highviremia that eventually declines to a subsequentset point of viral replication that depends onthe SIV clone used for infection and on theMHC class I genotype of the infected rhesusmonkeys (58). In addition, the KIR genotypeof infected rhesus monkeys can be associatedwith the level of viral control and the speedof SIV disease progression (59), suggestinga potential role for NK cells in that model.Studies of the consequences of KIR and MHCclass I genotypes for SIV disease outcome havebeen complicated by the complexity of theKIR genotype in rhesus monkeys (reviewed inReference 60). Inhibitory and activating KIRshave been characterized as ligands for MHCclass I molecules in rhesus monkeys, and poly-morphisms within these KIRs can modulate thepeptide-dependent binding of KIRs to MHCclass I molecules (61, 62). The consequenceof these KIR interactions with MHC class
I/peptide complexes for NK cell function andviral control in rhesus monkeys during SIVinfection requires further investigation.
Of interest, almost all NK cells in rhesusmonkeys express CD8, in contrast to humans,in which only a relatively small subset of NKcells expresses this marker (63). CD8 expressionon NK cells in rhesus monkeys has complicatedthe interpretation of studies that used anti-CD8antibodies for the depletion of CD8+ T cells,as these interventions also resulted in the re-moval of NK cells. Several studies have demon-strated that CD8 depletion in SIV-infected rhe-sus monkeys results in a significant increase inviral replication, suggesting a critical role ofCD8+ T cells, and potentially CD8+ NK cells,in the control of SIV viremia (64, 65). In con-trast, the depletion of CD16+ cells in rhesusmonkeys has not resulted in changes in viralreplication (66). However, anti-CD16 antibod-ies depleted most (up to 90%) but not all pe-ripheral blood NK cells, for a very short pe-riod of time, and potentially spared CD16− NKcells endowed with immunoregulatory func-tions. Novel approaches aimed at more specif-ically depleting the NK cell population in rhe-sus monkeys are currently being evaluated andwill hopefully provide some insights into therole of NK cells in the control of SIV replica-tion in this model. Furthermore, recent studieshave suggested that NK cells in rhesus monkeysmediate ADCC against SIV-infected cells, andthat these ADCC responses might be inverselycorrelated with SIV viral loads (67, 68). Takentogether, the precise role of NK cells in the con-trol of SIV replication in rhesus monkeys re-mains unclear. Optimizing the nonhuman pri-mate and humanized mouse models for thestudies of the antiviral activity of NK cells mightprovide new insights into AIDS virus pathogen-esis and the correlates of protective immunity.
Genetic Population Studies Suggesta Role of KIR+ NK Cells inHIV-1 Disease
Genetic studies in populations of HIV-1-infected individuals have provided evidencethat NK cells might play a role in the control
www.annualreviews.org • Control of Human Viral Infections by NK Cells 171
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
of HIV-1 replication and disease outcome.Initial studies by Flores-Villanueva et al. (69)showed that HIV-1-infected individuals withHLA class I alleles encoding for moleculescarrying the Bw4 serological motif experiencesignificantly better control of HIV-1 viremiaand slower HIV-1 disease progression. Insubsequent studies, Carrington and colleagues(70) showed that HIV-1-infected individualsexpressing KIR3DS1 and a subset of HLA-Bw4alleles encoding for an isoleucine in position80 (HLA-Bw4-80I) experienced significantlyslower progression to AIDS compared with in-dividuals encoding for either HLA-Bw4-80I orKIR3DS1. Similarly, the interaction betweenHLA-Bw4 molecules and their inhibitory lig-and KIR3DL1 appears to have an impact on theoutcome of HIV-1 disease. Studies by Martinet al. (71) showed that the protective effect ofHLA-Bw4 alleles is further modulated by theKIR3DL1 genotype. KIR3DL1 is highly poly-morphic, and expression of KIR3DL1 on NKcells varies depending on the KIR3DL1 subtype(71). A subset of KIR3DL1 alleles is associatedwith high expression of the receptor on the NKcell surface, whereas other KIR3DL1 alleles areexpressed at low levels. Individuals encodingfor HLA-Bw4 and highly expressed KIR3DL1allotypes had significantly lower levels of HIV-1 replication than did individuals encodingfor the same HLA-Bw4 alleles but KIR3DL1subtypes associated with low expression (71).Several studies have since then reported as-sociations between the HLA-Bw4/KIR3DL1genotype and protection from HIV-1 infectionor differential outcome of infection (72–78).The number of KIR3DS1 genes, which varybetween individuals as a consequence of geneduplication, has also been associated withbetter HIV-1 disease outcome in the contextof HLA-Bw4 but not HLA-Bw6 (79).
In addition to the research that has revealedthe protective interactions between HLA-Bw4and a subset of KIR3DS1/KIR3DL1 alleles,several genome-wide association studies aimedat identifying host genetic polymorphismsassociated with better control of HIV-1 viremiaor disease outcome have led to the description
of a single nucleotide polymorphism (SNP)35 kb upstream of HLA-C (rs9264942) that isassociated with the control of HIV-1 (80, 81).Further, SNP rs9264942 is associated withlevels of HLA-C transcription and cell surfaceexpression. Kulkarni et al. (82) elegantlydemonstrated the underlying mechanism,showing that this SNP modulates the abilityof microRNA hsa-miR-148 to bind to thepromoter area of HLA-C and thereby en-hances HLA-C mRNA and protein expression.HLA-C molecules serve as ligands for KIR2Dreceptors, and variations in HLA-C expressionmight therefore have an impact on the functionof NK cells. Studies of HIV-1-specific CD8+
T cell responses have consistently shownthat most virus-specific T cell responses arerestricted by HLA-B and HLA-A and thatHLA-C-restricted CD8+ T cells contributeonly a little to the total HIV-1-specific CD8+ Tcell response (83). In this context, it is of interestthat HIV-1 Nef (84) results in the downregula-tion of HLA-A molecules, which do not serve asligands for inhibitory KIRs (except for HLA-A3and A11, which might serve in some situationsas ligands for KIR3DL2) (85), and HLA-Bmolecules, which serve as ligands for KIR3DL1(24). However, HLA-C molecules, which inter-act with the large family of inhibitory KIR2DLmolecules, are not downregulated (84). HIV-1Nef might therefore have evolved to evadestrong virus-specific CD8+ T cell responses re-stricted by HLA-A and HLA-B while maintain-ing the HLA-C ligand for the broadly expressedand inhibitory KIR2DL molecules on the sur-face of infected cells to provide an inhibitorysignal to NK cells. Taken together, thesegenetic studies suggest that KIR+ NK cells andtheir interactions with the ligands, HLA classI molecules of the HLA-B and HLA-C family,might play a critical role in the outcome ofHIV-1 disease and have stimulated intensiveresearch into the underlying mechanisms.
Functional Studies of Human NKCells in HIV-1 Infection
Although population genetic studies haveprovided strong indications suggesting that
172 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
KIR/HLA interactions play a role in HIV-1disease outcome, the functional correlates ofthese interactions are less well understood. NKcells are expanded during acute HIV-1 infec-tion, particularly prior to the development ofdetectable adaptive immunity, and this expan-sion of NK cells appears to be regulated to someextent by KIR and HLA genotypes (86, 87).KIR3DS1+ NK cells are highly enriched in theperipheral blood of individuals with acute in-fection, yet only in individuals that also encodefor HLA-Bw4 alleles and not in HLA-Bw6 ho-mozygous individuals (86). Furthermore, sev-eral studies have suggested that KIR3DL1+ NKcells from HLA-Bw4+ individuals are function-ally superior to KIR3DL1+ NK cells from Bw6homozygous individuals (74, 75). These differ-ences might reflect differences in NK cell “li-censing” during development. Current modelsof NK cell development suggest that NK cellsneed to receive an inhibitory signal throughan inhibitory KIR during development to be-come functionally competent (88–91). In linewith this model, KIR3DL1+ NK cells fromindividuals that also encode HLA-Bw4 allelesmight be better licensed to kill infected cellsthan KIR3DL1+ NK cells from HLA-Bw6+
individuals. As the licensing of NK cells hasalso been suggested as being modulated by theexpression levels of KIRs and/or HLA class Iligands, this model provides an attractive ex-planation for the protective effect of KIR3DL1alleles associated with higher KIR3DL1 expres-sion in the context of HLA-Bw4 and of the SNPrs9264942 associated with higher expression ofHLA-C molecules. However, convincing func-tional data demonstrating such a correlate arestill pending. Furthermore, some studies havedemonstrated that KIR3DS1+ NK cells havestronger activity in HIV-1-infected individu-als encoding for HLA-Bw4 (77) and can in-hibit in vitro HIV-1 replication more potently(92), but whether this is due to the ability ofthese KIR3DS1+ NK cells to better recognizeHIV-1-infected cells or to a general higherfunctional activity is not known. The identi-fication of functional correlates of the geneticassociations described above is further compli-
cated by several factors. First, a direct inter-action of KIR3DS1 with HLA-Bw4 moleculeshas not been shown to date, and several studieshave failed to show any binding of KIR3DS1to HLA class I molecules (24, 93–95). Second,in individuals expressing both KIR3DS1 andKIR3DL1 receptors, it is currently difficult toevaluate how each of these molecules influencesNK cell function. For instance, while inhibitoryKIR receptors have been clearly demonstratedas playing a determining role in the licensingof NK cells, activating KIR receptors mightalso be involved in this process (96). However,in contrast to what has been observed for in-hibitory KIR receptors, interaction between ac-tivating KIRs and their cognate ligand duringNK cell development might lead to decreasedresponsiveness of NK cells. Third, the genesencoding KIR3DL1 and KIR3DS1 are foundin variable copy numbers among individuals,and it appears that protection against HIV-1increases with the number of gene copies ofKIR3DL1, provided that at least one copy ofthe KIR3DS1 gene and HLA-Bw4-80I are alsopresent (79). Finally, KIR genes are located veryclose to each other in the KIR locus, leading tosubstantial linkage disequilibrium, notably be-tween KIR3DS1 and the activating KIR2DS1receptor (97).
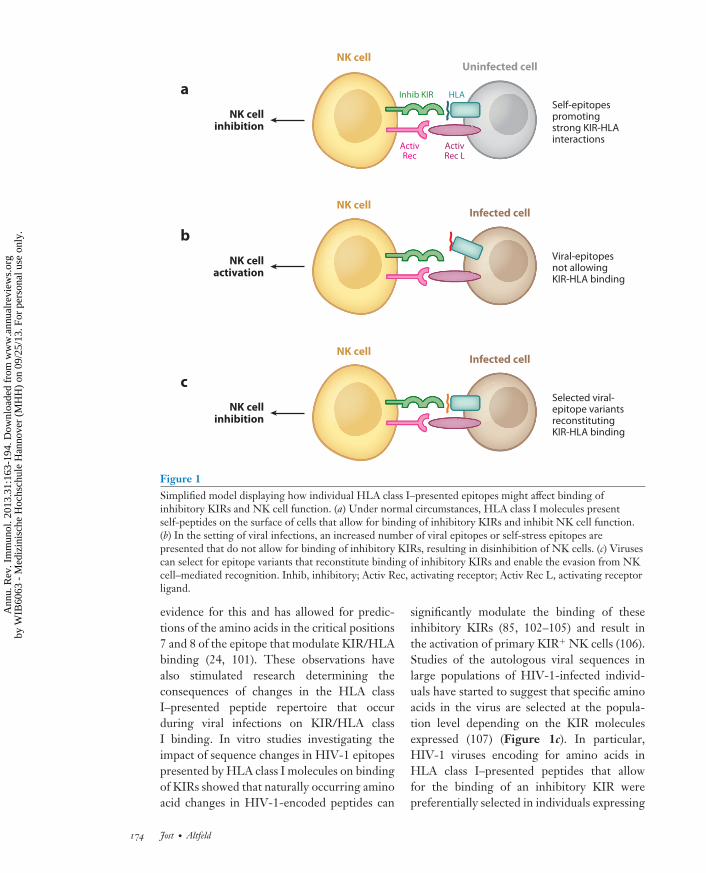
Apart from the role of KIR/HLA class Iinteractions for the functional licensing ofNK cells, the binding of KIR molecules toHLA class I is dependent on the sequenceand structure of the HLA class I–presentedepitopes (Figure 1). The role of the epitopesequence in this interaction was postulated byfunctional studies demonstrating that sequencechanges, particularly in the C-terminal part ofthe epitope, significantly modulated the abilityof NK cells expressing the respective inhibitoryKIR receptors to kill peptide-pulsed cell lines(98, 99). These data suggested that HLA classI–presented peptides that did not allow for thebinding of the inhibitory KIR resulted in a “dis-inhibition” of NK cells and target cell lysis (100)(Figure 1a,b). The recent characterization ofcrystal structures of HLA class I/peptide/KIRcomplexes has provided further structural
www.annualreviews.org • Control of Human Viral Infections by NK Cells 173
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.
IY31CH07-Altfeld ARI 14 February 2013 21:16
NK cell
NK cell
NK cell
Inhib KIR
ActivRec
ActivRec L
NK cellinhibition
Self-epitopespromotingstrong KIR-HLAinteractions
aUninfected cell
Infected cell
Infected cell
HLA
NK cellactivation
Viral-epitopesnot allowingKIR-HLA binding
b
NK cellinhibition
Selected viral-epitope variantsreconstitutingKIR-HLA binding
c
Figure 1Simplified model displaying how individual HLA class I–presented epitopes might affect binding ofinhibitory KIRs and NK cell function. (a) Under normal circumstances, HLA class I molecules presentself-peptides on the surface of cells that allow for binding of inhibitory KIRs and inhibit NK cell function.(b) In the setting of viral infections, an increased number of viral epitopes or self-stress epitopes arepresented that do not allow for binding of inhibitory KIRs, resulting in disinhibition of NK cells. (c) Virusescan select for epitope variants that reconstitute binding of inhibitory KIRs and enable the evasion from NKcell–mediated recognition. Inhib, inhibitory; Activ Rec, activating receptor; Activ Rec L, activating receptorligand.
evidence for this and has allowed for predic-tions of the amino acids in the critical positions7 and 8 of the epitope that modulate KIR/HLAbinding (24, 101). These observations havealso stimulated research determining theconsequences of changes in the HLA classI–presented peptide repertoire that occurduring viral infections on KIR/HLA classI binding. In vitro studies investigating theimpact of sequence changes in HIV-1 epitopespresented by HLA class I molecules on bindingof KIRs showed that naturally occurring aminoacid changes in HIV-1-encoded peptides can
significantly modulate the binding of theseinhibitory KIRs (85, 102–105) and result inthe activation of primary KIR+ NK cells (106).Studies of the autologous viral sequences inlarge populations of HIV-1-infected individ-uals have started to suggest that specific aminoacids in the virus are selected at the popula-tion level depending on the KIR moleculesexpressed (107) (Figure 1c). In particular,HIV-1 viruses encoding for amino acids inHLA class I–presented peptides that allowfor the binding of an inhibitory KIR werepreferentially selected in individuals expressing
174 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
the respective KIR but not in individuals whodo not encode for the respective KIR, suggest-ing that viruses encoding for these inhibitorysequences might have a selective benefit ininfected individuals (107; C.F. Thobakgale &M. Altfeld, unpublished data). However, theprecise functional consequences of these viralsequence polymorphisms for the recognitionof infected cells by NK cells remain to beelucidated.
Apart from the potential KIR-modulatedrecognition of HIV-1-infected cells by NKcells, receptors that are less polymorphic, in-cluding NKG2D, might play a role in the NKcell–mediated killing of HIV-1-infected cells.Several viruses, particularly herpesviruses, suchas CMV, have evolved elaborate mechanismsto evade this NKG2D-mediated recognition(as reviewed above). The HIV-1 Nef proteinappears to downregulate not only HLA-A andHLA-B but also NKG2D ligands expressedon infected cells (108), suggesting that thisspecific pathway might play a significant rolein the recognition of HIV-1-infected cells byNK cells.
Overall, several key NK cell receptorshave been associated with HIV-1 diseaseprogression and/or transmission, implying thatNK cells might contribute markedly to thecontrol of HIV-1 infection. Besides, HIV-1-encoded proteins can facilitate evasion fromNK cell recognition (as reviewed in Reference109). However, the extent to which theseviral evasion mechanisms modulate HIV-1pathogenesis and disease outcome remainsincompletely understood.
NK CELLS IN HEPATITIS CVIRUS INFECTION
Roughly 170 million people worldwide suf-fer from chronic HCV infection, a conditioncaused by an enveloped single-stranded RNAvirus of the Flaviviridae family (110, 111). Com-plications frequently arise from persistent HCVinfection, with liver cirrhosis and hepatocellu-lar carcinoma occurring in approximately 25%and 1–5% of cases, respectively. Although pro-
gression to chronic disease occurs in the major-ity of infected individuals, about one-fourth ofde novo HCV infections are cleared during theacute phase. Resolved infection is characterizedby the presence of HCV antibodies in the ab-sence of markers of active viral replication, suchas HCV RNA. Viral clearance is thought to relylargely on a broad, potent, and prolonged hostcellular immune response (112–114). Accord-ingly, defective T cell immunity is strongly as-sociated with viral persistence (115). No vaccineis currently available to prevent HCV infection,yet the rate of sustained virological responses totreatment has recently significantly increasedwith the administration of HCV protease in-hibitors in addition to the conventional combi-nation of pegylated INF-α and ribavirin (116).
In individuals who spontaneously clearHCV infection, viral control occurs within thefirst few months of infection. Before the onsetof the adaptive immune response, innate im-mune effector cells, such as NK and NKT cells,are thought to release IFN-γ, which is directlyresponsible for the noncytopathic inhibition ofHCV replication (117). IFN-γ-mediated clear-ance of the virus might be more important thandirect cytolysis of HCV-infected hepatocytesby NK cells, as nearly all hepatocytes becomeinfected and their elimination would lead toextensive liver damage. Type III IFN, such asIFN-λ, has received much attention recently,because not only does this cytokine inhibitHCV replication in vitro (118), but specificpolymorphisms located in the encoding gene,IL28B, are strongly associated with HCVclearance, response to therapy, and diseaseoutcome (119–122). Although the mechanismsunderlying IFN-λ-mediated eradication ofHCV replication in vivo remain to be deter-mined, it is possible that this cytokine plays acrucial role in modulating NK cell responsesto HCV, similarly to IFN-α (118).
NK cells constitute 30% of resident lym-phocytes in the normal liver, which is higherthan their representation in the peripheralblood, and percentages of NK cells in HCV-infected livers can reach as high as 90% (123).Compared to peripheral blood NK cells, NK
www.annualreviews.org • Control of Human Viral Infections by NK Cells 175
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
cells in healthy livers are characterized by adecreased ability to perform their cytotoxic andimmunoregulatory functions, a functional statethat might be maintained by high expressionof the inhibitory NKG2A receptor on NKcells, and an increased local production ofthe immunosuppressive cytokine IL-10 (124,125). These features are required to avoidimmunopathology of the liver, in which tolero-genic conditions have to be maintained toavoid inflammation in response to gut-derivedantigens.
Protective Effect Mediated byKIR2DL3 in Combination with ItsLigand HLA-C1
Compelling evidence for a role of human NKcells in HCV infection comes from epidemio-logical studies, which led to the identificationof an association between homozygosity forthe KIR/HLA genotype KIR2DL3/HLA-C1and viral clearance in intravenous drug users(126–128). This genotype not only has beensuggested to protect from the establishmentof chronic disease, but also predominates inHCV-exposed seronegative aviremic indi-viduals and in HCV-infected patients withsustained responses to IFN-α-based therapies(127, 129). Accordingly, the frequency ofindividuals encoding for HLA-C2 moleculeswas higher among patients with chronic HCVinfection than among healthy individuals (130),and individuals with resolved HCV infectionhad increased percentages of NK cells ex-pressing HLA-C-binding KIRs (includingKIR2DL1, -2DL2, -2DL3, -2DS1, -2DS2)compared to those with acute and chronicHCV infection or to healthy controls (131).
There are a few specific situations in whichthe positive effect of KIR2DL3 in conjunctionwith its HLA-C1 ligand seems to be lost.For instance, among all possible ligands forKIR2DL3, expression of HLA-Cw∗07 has beensuggested to be associated with KIR2DL3+
NK cells with an increased ability to produceIFN-γ, yet this particular HLA-C1 alleledoes not confer protection from progression
toward chronic HCV infection (132). A lack ofassociation between the KIR2DL3/HLA-C1genotype and HCV clearance in HIV-1-infected individuals has also been reported(133) and might reflect a general impairmentof NK cell function in these individuals. Inthe future, it will be important to evaluate howcoinfections and allelic polymorphisms affectthe ability of NK cells expressing protectiveKIRs to mediate their potentially beneficialeffect on HCV infection.
In support of a possible increased abilityof NK cells to eliminate virally infected cellsin the context of the KIR2DL3/HLA-C1genotype, KIR2DL2/3+ NK cells displayhigher levels of activation than KIR2DL1+
NK cells in the blood of individuals with acuteHCV infection, and particularly in those whosubsequently clear the infection (134). Becausethe affinity between the inhibitory KIR2DL3and its ligand HLA-C1 is weaker than thatbetween KIR2DL1 and HLA-C2 or KIR2DL2and HLA-C1, it is plausible that a lowerthreshold of activation is required to triggerKIR2DL3+ NK cell responses (135). However,HCV infection does not lead to a significantdownregulation of MHC class I molecules atthe surface of infected cells. Therefore, theinfluence of HLA-C1-presented epitopes onthe avidity of the KIR-HLA interaction, ratherthan HLA-C1 disappearance from the cellsurface, might explain the protective effectmediated by KIR2DL3+ NK cells, as suggestedfor HLA-Cw∗0102 (136). Further experimentsare required to determine the mechanismsunderlying KIR2DL3-mediated protection,including the influence of other KIRs.
Impact of Other KIRs on HCVDisease Outcome
Expression of additional KIRs has been pro-posed to also play a role in the prevention oroutcome of HCV infection. For instance, in-creased frequencies of the activating KIR2DS3receptor and homozygosity for KIR3DS1 inconjunction with HLA-Bw4 were associatedwith poor prognosis for HCV infection (137),
176 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
suggesting that liver damage in chronic HCVinfection might be explained, at least partly,by activating KIR-mediated NK cell overac-tivation. In contrast, in the context of hepa-tocellular carcinoma development in patientswith chronic HCV infection, expression ofKIR3DS1 or HLA-C1 has been suggested tobe protective (138), but the potential mecha-nisms underlying these protective associationsremain unknown. Studies comparing cohorts ofchronically HCV-infected subjects and high-risk aviremic seronegative individuals showedthat expression of the activating KIR2DS4 re-ceptor contributes to the KIR2DL3-mediatedprotection against infection in exposed butuninfected HLA-C1+ intravenous drug users(139). However, because the known ligand forKIR2DS4 is HLA-C2, the functional relevanceof this finding will have to be addressed (140).Finally, the failure of antiviral combinationtherapy against HCV has been associated withthe presence of KIR2DL5 (141). It is cur-rently difficult to provide a mechanistic expla-nation for this observation as the ligand forthe KIR2DL5 receptor has not been identi-fied. It will also be important to assess whetherKIR polymorphisms will be relevant for treat-ment outcome in the context of the recently im-proved HCV treatments, which have markedlyenhanced the percentages of patients with a sus-tained virological response.
To clarify the role of KIRs in the outcome ofnatural HCV infection, it will be important todetermine whether linkage disequilibrium withother genes accounts for the associations withliver disease or protection from infection ob-served for activating KIR receptors. The pres-ence of KIR/HLA compound genotypes thatallow for strong NK cell activation very earlyin HCV infection might significantly increasethe chances of clearing the virus, yet persistentinnate immune activation during chronic in-fection might contribute to pathology. For in-stance, HCMV infection leads to the expansionand long-term persistence of NK cells express-ing the activating NKG2C receptor (43). InHCMV-seropositive individuals with chronicHCV infection, this NKG2C+ NK cell sub-
population also tends to coexpress inhibitoryself-KIRs, including KIR2DL3, and might rep-resent an important mean to limit NK cell–mediated liver injury (142).
Studies of NK Cell Responses to HCVUsing In Vitro Systems
Scientific progress in understanding HCVpathogenesis has been hampered severely bythe lack of appropriate experimental in vivoor in vitro systems. Chimpanzees are the onlyimmunocompetent animals that can be reliablyinfected with HCV, but access to this modelhas been limited owing to ethical considera-tions, high costs, and difficulties inherent toworking with large animals and has thus beenstopped. As an attractive alternative, in vitroHCV culture systems and humanized mousemodels that are permissive to HCV infectionhave been recently developed (143). In 2005,a robust HCV culture system was establishedby infecting the Huh7.5 hepatoma-derived cellline with a full-length genotype 2a recombi-nant HCV. This in vitro system established theability of NK cells to abrogate HCV replicationin vitro (144) by demonstrating that incubationof HCV-infected hepatocytes with NK cellculture supernatant reduced HCV RNA andprotein production, an effect that depended onIFN-γ produced by NK cells. Furthermore,in vitro experiments conducted in cell culturemodels uncovered several potential mecha-nisms by which HCV might affect NK cellfunctions in vivo. One major finding was theability of the HCV envelope 2 (E2) proteinto interact directly with CD81, a cellularcoreceptor for the virus, thereby blockingantiviral functions of CD81+ NK cells (145–147). However, experimental settings of thesestudies involved plates coated with recombi-nant truncated HCV-E2 proteins. When NKcells are exposed to intact HCV virions, theirfunctions are preserved, whereas the presenceof anti-CD81 antibodies suppresses NK cell–mediated IFN-γ production and cytotoxicity(148). Therefore, it remains to be elucidatedwhether cross-linking of CD81 by HCV E2
www.annualreviews.org • Control of Human Viral Infections by NK Cells 177
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
affects NK cell function and provides a wayfor the virus to facilitate the establishment ofchronic infection. Alternatively, in vitro studiessuggested that NK cell function might beimpaired upon enhanced expression of ligandsfor inhibitory NK cell receptors on HCV-infected cells, including HLA class I (149), andspecifically HLA-E (150), or downregulationof MICA, a ligand for the activating NKG2DNK cell receptor (151). The contribution ofthese different pathways to the outcome ofHCV disease, however, remains incompletelyunderstood and requires further evaluation.
Changes in the NK Cell CompartmentDuring HCV Infection
One mechanism by which HCV establisheschronic persistent infection might involvevirus-induced alterations of NK cell functionsvery early in the course of infection. This hy-pothesis is supported by increasing evidencedemonstrating that patients with chronic HCVinfection have several peripheral blood and in-trahepatic NK cell abnormalities, such as de-creased numbers of NK cells, and altered NKcell subset distribution and/or NK cell receptorexpression (reviewed in Reference 152). How-ever, to what extent HCV-associated NK cellphenotypic changes affect NK cell function isnot entirely clear, and although initial stud-ies observed impaired natural cytotoxicity inchronic HCV infection (147, 153–155), morerecent reports strongly suggest that NK cellfunctions are not reduced (131, 148, 156–161).Moreover, the field has a limited understandingof intrahepatic NK cell function in HCV in-fection, and NK cells with decreased cytotoxicfunctions yet intact IFN-γ production mightstill be able to contribute to the clearance ofthe virus, as the antiviral effect mediated byIFN-γ has been proposed to be more efficientat controlling HCV replication than is directlysis of infected hepatocytes (162). Finally, be-cause acute HCV replication is mostly asymp-tomatic, access to acute infection samples hasbeen challenging, and the majority of studieshave focused on comparing peripheral blood
NK cell responses in resolved and chronic HCVinfection. These studies have generated a sur-prisingly high degree of controversial results,which are discussed below. Heterogeneity incohorts and experimental protocols applied tostudy NK cells might account for these discrep-ancies. However, the few existing studies focus-ing on NK cells in acute HCV infection pointtoward a pivotal role for NK cells in the earlycontrol of HCV replication and in predictingsubsequent disease outcome (131, 134, 163).
NK Cells in Acute HCV Infection
Redistribution of NK cell subsets occurs earlyin acute HCV infection and is characterizedby an increase in the percentages of CD56bright
and CD56neg NK cells, whereas percentagesof CD56dim NK cells are reduced. These datasuggest that the KIR-expressing cytotoxicCD56dim subpopulation might be recruitedtoward the acutely infected liver (131, 134).Overall, peripheral blood NK cells in acuteHCV infection display enhanced functionscompared with NK cells from uninfected orchronically infected subjects, and importantly,natural cytotoxicity does not differ betweensubjects who are subsequently able to clear thevirus and those who develop chronic infection(131, 134, 163). However, some differencesin NK cells during acute HCV infection havebeen suggested to be predictive of the outcomeof HCV infection. In particular, individualswho spontaneously clear HCV infection havesignificantly lower proportions of peripheralblood NK cells expressing NKp46, NKp30,NKG2D, and CD161 at the time of acuteinfection compared with those who developchronic infection (131). Further studies lookingat liver specimens during acute HCV infectionare required to conclude whether differencesin NK cells during the initial stage of infectioncan determine subsequent disease outcome.
NK Cell Phenotypes in ChronicHCV Infection
The distribution of circulating NK cell sub-sets is also skewed in chronic HCV infection,
178 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
with percentages of CD56neg and CD56bright
NK cells increased relative to CD56dim NKcells (reviewed in Reference 164). An alteredratio of CD56dim to CD56bright NK cells in theinfected liver has also been reported, yet it isnot clear whether this observation is directly re-lated to HCV infection as there are no convinc-ing data regarding the proportions of these NKcell subsets in the healthy liver (165). Further-more, chronic HCV infection is associated withan increased number of peripheral blood andintrahepatic NK cells bearing the inhibitoryheterodimeric receptor CD94/NKG2A (157,159, 165, 166). This feature has been pro-posed to result in reduced cytolytic activityand impaired dendritic cell activation, poten-tially facilitating viral persistence (167–169). Inaccordance with these observations, individu-als with spontaneously resolved HCV infectionhave diminished numbers of NK cells express-ing NKG2A, -2C, and -2E compared with pa-tients with chronic infection (161). At least oneHLA-E-restricted HCV peptide has been sug-gested to stabilize the ligand for NKG2A atthe surface of infected hepatocytes and mightbe responsible for the increased expression ofNKG2A on NK cells (150). However, expres-sion of NKG2A on CD56dim NK cells is in-versely correlated to HCV viral load, suggest-ing that the NKG2A+ NK cell subset, whosefunction is expected to be more inhibited, mightactually be protective (165). Further investiga-tions are required to determine whether otherreceptors on NKG2A+ NK cells might partlyaccount for this observation. In particular, coex-pression of CD94, which can protect NK cellsfrom cell death, might contribute to this de-scribed association (170). Higher pretreatmentfrequencies of NKG2A+KIR− NK cells occurin patients who subsequently present a success-ful response to antiviral therapy compared withthose who fail to respond, further suggestingthat a protective effect might be mediated by asubset of NKG2A+ NK cells that do not over-express inhibitory receptors (166).
Although the increase in percentages ofNKG2A+ NK cells in subjects with chronicHCV infection has been observed consistently
in several studies, contradictory data regardingthe frequency of NCR-bearing peripheralblood and intrahepatic NK cells have beenreported (131, 157–159, 166, 169). Of interest,a recent study showed that NKp46high NKcells have potent anti-HCV activity in vitroand accumulate in the liver of subjects withchronic HCV infection, suggesting that func-tionally competent NKp46+ NK cells mightbe recruited to the site of infection (171).Although the enhanced activity of this subsetmight contribute to liver damage, intrahepaticpercentages of NKp46high inversely correlatewith HCV viral load. A potential role forNKp30 in the control of HCV infection hasbeen further suggested by the fact that NK cellswith high expression levels of NKp30 mighthave an increased potential to protect againstHCV infection in vivo and in vitro (172).
Further investigations are required to linkalterations in NK cell phenotype to their anti-HCV function, yet it can be speculated that lowexpression of activating receptors on peripheralNK cells during chronic HCV infection reflectsanergy of NK cells, resulting from consistentactivation through these receptors. Alterna-tively, NK cells expressing activating receptorsmight be retained in the infected liver, and thishypothesis is supported by recent investigationslooking at both intrahepatic and peripheralblood NK cells in parallel (157, 159, 171).Taken together, the current literature suggeststhat increased and early expression of activatingreceptors on hepatic NK cells, associated withenhanced NK cell function, might provideprotection from infection in exposed individ-uals and might favor spontaneous resolution ofinfection, while the consequences of changesin the NK cell compartment during chronicinfection for disease outcome are less clear.
NK CELLS IN INFLUENZAVIRUS INFECTION
Orthomyxoviridae, a family of envelopedsingle-stranded segmented RNA viruses thatincludes influenza A, B, and C viruses, causeannual influenza disease outbreaks as well as
www.annualreviews.org • Control of Human Viral Infections by NK Cells 179
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
sporadic and devastating pandemics (reviewedin Reference 173). Seasonal influenza epi-demics result in an estimated 250,000–500,000deaths per year worldwide (174) and in anaverage of 200,000 hospitalizations (175) and36,000 deaths per year in the United Statesalone (176, 177). Seasonal epidemics areinstigated by the quick spread of influenzavirus variants generated by antigenic drift, aprocess that facilitates the escape of the virusfrom the host immune system. As a result,influenza vaccines need to be reformulatedbefore each annual epidemic, and althoughyearly immunization has been the most effi-cient way to prevent influenza so far, vaccinesdo not provide complete protection. Recoveryfrom acute influenza infection and resistanceto reinfection rely both on the production ofneutralizing antibodies targeting the HA andthe neuraminidase glycoproteins and on thekilling of infected cells by influenza-specificcytotoxic CD8+ T lymphocytes that mostlytarget conserved viral proteins (178, 179). Inaddition, several studies, reviewed below, havehighlighted the early and potentially importantrole of innate effector cells, particularly NKcells, in the control of influenza infection.
NK Cells in Mouse Models ofInfluenza Virus Infection
Most studies of influenza virus infection havebeen performed using mouse models. Mice arenaturally not susceptible to influenza infection.However, influenza disease and lethal infectioncan be achieved with viral strains that have beenadapted by serial passages in mice. In both hu-mans and mice, NK cells initially represent asubstantial portion of the lymphocyte popula-tion that resides in the healthy lungs, and withindays of influenza infection, these innate im-mune cells are further recruited to the respi-ratory tract (180–183). Experimental influenzainfection of NK cell–depleted mice, or differentmice models with defects in NK cell activity, re-sulted in a delayed clearance of the virus fromthe lungs and augmented morbidity and mortal-ity (184–188). However, the beneficial impact
of NK cells on the outcome of influenza diseasehas been recently questioned by a study report-ing an increased survival of mice devoid of IL-15, or depleted of NK cells using antibodies,and challenged with a lethal dose of influenzavirus (189). These discrepancies with previousobservations might be explained by the amountof virus used to infect the mice, and they sug-gest that at high infectious doses NK cells mightsignificantly contribute to influenza-mediatedimmunopathology by enhancing pulmonary in-flammation. Beyond its role in NK cell homeo-stasis, IL-15 also has chemotactic properties,and its influenza-induced release at the site ofinfection might participate in recruiting NKcells to the lung airways of infected mice (190).
Murine models and in vitro experimentsidentified NCR1, the murine receptor corre-sponding to human NKp46, as a potential NKcell receptor that can interact with influenzaHA in a sialic acid–dependent manner and is in-volved in the elimination of influenza-infectedcells (191, 192). Whereas the influence of otherNK cell receptors in the recognition and killingof influenza-infected cells remains unknown,mice lacking NCR1 fail to clear the virus anddo not survive infection with a lethal dose ofinfluenza. In this murine model, the presenceof the Ncr1 gene confers protection againstchallenge with influenza H1 subtypes and thehighly pathogenic avian influenza H5 subtype,although in vitro, interaction between NKp46and H5 was not sufficient to trigger killingof H5N1-infected cells (193). It is generallyaccepted that NCR1 can interact with influenzaHA, yet the relevance of NCR1 expression atthe surface of NK cells in mediating protectionagainst influenza infection in vivo remainscontroversial. Indeed, Noe mice, which carrya mutated Ncr1 gene that abolishes surfaceexpression of the receptor, display increasedresistance to influenza infection (194). Inaddition to binding to viral HA via NCR1, NKcells might recognize and eliminate influenza-infected cells via binding to influenza-specificantibodies (195, 196). ADCC has been identi-fied as the main mechanism providing protec-tion against lethal influenza challenge in mice
180 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
vaccinated with the extracellular part of the M2influenza protein (196). Taken together, stud-ies using mouse models demonstrated that NKcell function, particularly NCR1-mediated NKcell activation, can have a significant impact onthe outcome of influenza infection.
NK Cells in HumanInfluenza Infection
Humans are susceptible to several strains ofinfluenza, and the virus is normally eliminatedwithin a few days following infection inimmunocompetent individuals. Several studieshave suggested a role for NK cells in thisprocess. As described above, NK cells requirethe interaction between an inhibitory KIR andits ligand to become licensed during develop-ment. Applying this model to NK cell responsesagainst influenza, Ahlenstiel et al. (197) showedin in vitro experiments that KIR2DL3+ NKcells derived from individuals also encoding forits ligand HLA-C1 exhibited strong activityin response to influenza. However, to datethere is no evidence for a KIR2DL3-mediatedprotective effect at the population level. Somegenetic associations between KIR alleles andincreased control or susceptibility to severeand/or lethal influenza infection with the 2009pandemic H1N1 virus have been described(198, 199). For instance, KIR2DL2 and/orKIR2DL3 in conjunction with their HLA-C1ligands were enriched in subjects with severeinfluenza infection, whereas the absence ofHLA-Bw4 ligands for KIR3DL1/S1 or ofHLA-C2 ligands for KIR2DL1 prevailed inpatients with significant influenza-associatedpathology requiring intensive care treatment(Table 2) (198). Moreover, the frequencyof KIR3DS1, KIR2DS5, and KIR2DL5 wasassociated with increased severity in symptoms(199). However, these studies were conductedin relatively small patient cohorts, and furtherstudies involving larger numbers of subjectsare warranted to fully determine the clinicalrelevance of KIR alleles and their HLAligands in the immune response to influenzainfection.
In support of the role of NCR1 in themouse NK cell response to influenza, NKp46predominantly mediates human NK cell lysisof influenza-infected cells in vitro by directlybinding to the HA proteins present at the sur-face of infected cells (20, 21, 200). α-2,6-linkedsialic acid residues play a crucial role in thebinding of both NKp46 and NKp44 to viral HA(18, 20, 21). Furthermore, NKp46 blockadeimpairs NK cell responses to influenza (201,202). In particular, expression of both NKp46and NKG2D has been suggested to be re-quired for the recognition of influenza-infecteddendritic cells and the subsequent activationof NK cells (201). Polyfunctional NK cellresponses to influenza rely on a combinationof signals provided by dendritic cells, whichinclude not only upregulation of NKp46- andNKG2D-ligands on dendritic cells but also theproduction of IFN-α and IL-12 (201). Finally,NKp46 downregulation following influenzaexposure has been described in different invitro experimental models, potentially reflect-ing influenza-induced activation of NKp46+
NK cells (202, 203). Altogether, these datasuggest the importance of activating NK cellreceptors in the response to influenza. Futureexperiments need to determine whether over-activation of NK cells during acute infectionwith highly pathogenic influenza viruses, suchas H5N1, might also contribute to increasingmorbidity and mortality.
Evasion from NK Cell–MediatedImmune Responses by Influenza Virus
NK cell contribution to the immune controlof influenza infection has also been reflectedby several studies describing strategies evolvedby influenza to specifically evade NK cell–mediated immune responses, specificallyNKp46-mediated responses. One strategyconsists of changing the glycosylation patternsof the HA protein (204, 205). These changesoccur over time as the virus rapidly acquiresmutations conferring resistance to antibody-mediated neutralization from one year toanother. For instance, H3N2 influenza viruses
www.annualreviews.org • Control of Human Viral Infections by NK Cells 181
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
that were isolated more than 16 years ago havea higher binding affinity for sialic acid, andtherefore an increased ability to trigger NKcell–mediated killing, than do contemporaryH3N2 influenza strains isolated between 1999and 2003 (204). Further studies are required todetermine whether this reduced recognition byNK cells is a reflection of direct adaptation toNK cell–mediated immune pressure, simply anindirect consequence of escape from antibodyresponses, or both.
Evasion from NK cell effector functions byincreasing the signals delivered via inhibitoryNK cell receptors is another strategy em-ployed by influenza viruses to escape NK cell–mediated lysis (206, 207). Prior to cell surfaceexpression of ligands for NK cell–activatingreceptors, such as NKp46, influenza infec-tion triggers the accumulation of MHC class Imolecules into lipid raft microdomains, therebyincreasing the binding of inhibitory NK cell re-ceptors, including KIR2DL1 and LIR-1, to theinfected cells (207). Altogether, these in vitrostudies suggest that influenza virus has devel-oped evasion strategies to escape recognitionby human NK cells.
Human NK Cell Responses toInfluenza in Vaccinatedand Infected Individuals
Several studies have assessed the NK cell re-sponses in individuals vaccinated with currentinfluenza vaccines or infected with influenza.About 30 years ago, increased NK cell cyto-toxic function and IFN-γ production were as-sociated for the first time with intranasal inocu-lation of volunteers with a live H1N1 influenzavirus (180). Since then, additional studies havereported an enhanced production of IFN-γ byNK cells in subjects vaccinated with live at-tenuated or inactivated influenza vaccines (208,209). Longitudinal analysis of NK cell pheno-types following immunization with influenzavaccines revealed a very early and transient de-crease in the percentage of NKp46+ NK cells,further supporting a downmodulation of thisNCR upon engagement with HA, and a role for
NKp46 in recognizing influenza-infected NKcells (203). In addition, influenza vaccinationwas associated with an expansion of NK cellsexpressing high levels of 2B4, which persistedfor up to 150 days.
Before the emergence of the 2009 pandemicH1N1 influenza virus, ex vivo studies charac-terizing changes in human NK cell phenotypeand function in acute influenza infection werescarce. However, increased mortality and mor-bidity associated with the pandemic strain rein-forced the need to further understand the roleplayed by immune effector cells in influenza-associated pathology and to shed further lighton human NK cell responses to influenza. No-tably, clinical investigations reported markeddecreases in peripheral blood NK cell numbersin influenza-infected individuals, with severesymptoms associated with a more dramatic de-crease or a complete absence of circulating NKcells (210–214). Reduced numbers of peripheralblood NK cells in patients with acute influenzainfection might reflect homing of these cellsto other tissues, including those of the respi-ratory tract. Alternatively, influenza-mediatedincreased apoptosis and activation-induced celldeath might explain the observed decrease incirculating NK cells (215, 216). In support ofthis latter hypothesis, lung inflammatory infil-trates from one subject with fatal H1N1 in-fluenza infection were devoid of NK cells (210).Further investigation is required to evaluate therole played by NK cell trafficking to infectedtissues and/or immune cell death in influenza-associated lymphopenia.
COMMON THEMES IN NKCELL–MEDIATEDRECOGNITION OF VIRALLYINFECTED CELLS
Viral infections trigger a panel of commonmodifications in their respective target cells,which can be sensed by NK cells. These includeligands of several different families of NK cellreceptors, probably to ensure that some willstill be efficient at eliciting NK cell responsesagainst viruses that have evolved multiple
182 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
mechanisms to evade NK cell recognition.NK cell activation depends, on the one hand,on the loss of inhibitory signals provided byMHC class I molecules and, on the other hand,on the expression of stress- or virally inducedligands for activating NK cell receptors. Thebest characterized of these ligands are thoseengaging NKG2D (Table 1 and Figure 2a).Interestingly, although many viruses expressproteins that counteract NKG2D-mediatedNK cell activation, no convincing escapemechanisms have been described for viral es-cape from NCR-mediated recognition to date.However, yet-to-be-discovered ligands for theNCRs may play a role in the NK cell responseto viruses (Table 1, Figure 2b). Similarly,natural ligands for most activating KIRs havenot been identified to date, and the precise roleof these receptors in the NK cell responsesagainst viruses remains to be determined. Hostgenetic-association studies with viral outcomehave led to the suggestion that activatingKIRs might recognize specific MHC class Imolecules when those molecules start present-ing epitopes derived from stress-induced orviral proteins (Table 1 and Figure 2c) or thatactivating KIRs might directly recognize viralantigens, as described for Ly49H in the contextof MCMV infection. Finally, both downmod-ulation of MHC class I and alterations of the
−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−→Figure 2Expected changes in the expression of ligands foractivating NK cell receptors at the surface of virallyinfected cells in the absence of viral escapemechanisms. (a) Upregulation of ligands forNKG2D. (b) Upregulation of cellular ligands and/orexpression of viral proteins recognized by NCRs.(c) Upregulation of viral and/or cellular ligandsand/or changes in the repertoire of peptidespresented by MHC class I that allow recognition byactivating KIRs. (Abbreviations: H, human; M,mouse; NCR, natural cytotoxicity receptor; HA,hemagglutinin; KIR, killer immunoglobulin-likereceptor; HLA, human leukocyte antigen;MULT-1, mouse UL16-binding-like transcript-1;RAE-1, retinoic acid early transcript-1; H60,histocompatibility 60; ULBP, UL16-bindingprotein; WNV/DV, West Nile virus/Dengue virus.)
a
NK cell
H
NKG
2D
NKG2D
Infected cell
ULBPs
M NKG2D MULT-1
RAE-1
H60
MICA/B
b
H
NCR
s
NCRsHCMV pp65
Viral HA
WNV/DVE glycoprotein
M Viral HA
?
c
H
KlRs
KIR2DS HLA-C
M
?
?
NCRs
KIR3DS
Ly49H
Ly49D
m157
H-2Dd
www.annualreviews.org • Control of Human Viral Infections by NK Cells 183
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
repertoire of MHC class I–presented peptidesmay contribute to reducing the interactionswith NK cell inhibitory receptors (Table 1 andFigure 1), resulting in NK cell disinhibition.
All the above-mentioned common featuresof virally infected cells might act in concert toenhance NK cell antiviral responses. Further-more, recent studies in Rag-deficient mice havesuggested that NK cells are endowed with theability to recognize virally infected cells in anantigen-specific manner, in some cases throughreceptors yet to be defined, allowing for subse-quent NK cell–mediated recall responses (4–6).To determine the precise receptors and mech-anisms underlying these NK cell–mediated re-call responses, and whether human NK cellspossess this faculty, will surely become an in-tense area of future investigation.
CONCLUSIONS
An increasing body of data suggests that NKcells play a critical role in the control of many vi-
ral diseases, including human infections causedby CMV, HIV-1, influenza virus, and HCV. Al-though NK cell–mediated immunity can con-tribute to viral control and viral clearance dur-ing the initial acute phase of these infections,overly extensive activation of NK cells dur-ing acute infection and persistent stimulationduring chronic infections might contribute tovirus-associated pathology. Furthermore, thereare significant limitations in our current under-standing of the mechanisms by which humanNK cells recognize different viruses and me-diate their antiviral function. In particular, thehuman NK cell receptors that might enable NKcells to directly recognize infected cells remainunknown. Future studies will need to elucidatein more detail the molecular mechanisms bywhich NK cells recognize infected cells, as wellas the receptor-ligand families involved. A moreprecise understanding of these interactions willrepresent the basis for the development of even-tual targeted interventions to harness the antivi-ral activity of human NK cells.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
Work by the authors has been supported by the National Institutes of Health (R01AI066031to M.A.), the Doris Duke Charitable Foundation (M.A.), the Philip T. and Susan M. RagonFoundation (M.A., S.J.), as well as the Swiss National Science Foundation and the Mas-sachusetts General Hospital Executive Committee on Research (ECOR) Fund for MedicalDiscovery (S.J.).
LITERATURE CITED
1. Cooper MA, Fehniger TA, Caligiuri MA. 2001. The biology of human natural killer-cell subsets. TrendsImmunol. 22:633–40
2. Altfeld M, Fadda L, Frleta D, Bhardwaj N. 2011. DCs and NK cells: critical effectors in the immuneresponse to HIV-1. Nat. Rev. Immunol. 11:176–86
3. Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. 2008. Functions of natural killer cells. Nat.Immunol. 9:503–10
4. O’Leary JG, Goodarzi M, Drayton DL, von Andrian UH. 2006. T cell- and B cell-independent adaptiveimmunity mediated by natural killer cells. Nat. Immunol. 7:507–16
184 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
5. Paust S, Gill HS, Wang BZ, Flynn MP, Moseman EA, et al. 2010. Critical role for the chemokinereceptor CXCR6 in NK cell-mediated antigen-specific memory of haptens and viruses. Nat. Immunol.11:1127–35
6. Sun JC, Beilke JN, Lanier LL. 2009. Adaptive immune features of natural killer cells. Nature 457:557–617. Orange JS. 2002. Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect.
4:1545–588. Biron CA, Byron KS, Sullivan JL. 1989. Severe herpesvirus infections in an adolescent without natural
killer cells. N. Engl. J. Med. 320:1731–359. de Vries E, Koene HR, Vossen JM, Gratama JW, von dem Borne AE, et al. 1996. Identification of an
unusual Fcγ receptor IIIa (CD16) on natural killer cells in a patient with recurrent infections. Blood88:3022–27
10. Jawahar S, Moody C, Chan M, Finberg R, Geha R, Chatila T. 1996. Natural killer (NK) cell deficiencyassociated with an epitope-deficient Fc receptor type IIIA (CD16-II). Clin. Exp. Immunol. 103:408–13
11. de Haas M, Koene HR, Kleijer M, de Vries E, Simsek S, et al. 1996. A triallelic Fcγ receptor type IIIApolymorphism influences the binding of human IgG by NK cell FcγRIIIa. J. Immunol. 156:3948–55
12. Hayakawa Y, Andrews DM, Smyth MJ. 2010. Subset analysis of human and mouse mature NK cells.Methods Mol. Biol. 612:27–38
13. Caligiuri MA, Zmuidzinas A, Manley TJ, Levine H, Smith KA, Ritz J. 1990. Functional consequences ofinterleukin 2 receptor expression on resting human lymphocytes. Identification of a novel natural killercell subset with high affinity receptors. J. Exp. Med. 171:1509–26
14. Mavilio D, Benjamin J, Daucher M, Lombardo G, Kottilil S, et al. 2003. Natural killer cells in HIV-1infection: dichotomous effects of viremia on inhibitory and activating receptors and their functionalcorrelates. Proc. Natl. Acad. Sci. USA 100:15011–16
15. Alter G, Teigen N, Davis BT, Addo MM, Suscovich TJ, et al. 2005. Sequential deregulation of NK cellsubset distribution and function starting in acute HIV-1 infection. Blood 106:3366–69
16. Cooper MA, Fehniger TA, Turner SC, Chen KS, Ghaheri BA, et al. 2001. Human natural killer cells:a unique innate immunoregulatory role for the CD56bright subset. Blood 97:3146–51
17. Lanier LL. 2008. Up on the tightrope: natural killer cell activation and inhibition. Nat. Immunol. 9:495–502
18. Arnon TI, Markel G, Mandelboim O. 2006. Tumor and viral recognition by natural killer cells receptors.Semin. Cancer Biol. 16:348–58
19. Jost S, Altfeld M. 2012. Evasion from NK cell-mediated immune responses by HIV-1. Microbes Infect.14:904–15
20. Arnon TI, Lev M, Katz G, Chernobrov Y, Porgador A, Mandelboim O. 2001. Recognition of viralhemagglutinins by NKp44 but not by NKp30. Eur. J. Immunol. 31:2680–89
21. Mandelboim O, Lieberman N, Lev M, Paul L, Arnon TI, et al. 2001. Recognition of haemagglutininson virus-infected cells by NKp46 activates lysis by human NK cells. Nature 409:1055–60
22. Pegram HJ, Andrews DM, Smyth MJ, Darcy PK, Kershaw MH. 2011. Activating and inhibitory receptorsof natural killer cells. Immunol. Cell Biol. 89:216–24
23. Bashirova AA, Thomas R, Carrington M. 2011. HLA/KIR restraint of HIV: surviving the fittest. Annu.Rev. Immunol. 29:295–317
24. Vivian JP, Duncan RC, Berry R, O’Connor GM, Reid HH, et al. 2011. Killer cell immunoglobulin-likereceptor 3DL1-mediated recognition of human leukocyte antigen B. Nature 479:401–5
25. Stewart CA, Laugier-Anfossi F, Vely F, Saulquin X, Riedmuller J, et al. 2005. Recognition of peptide-MHC class I complexes by activating killer immunoglobulin-like receptors. Proc. Natl. Acad. Sci. USA102:13224–29
26. Jonsson AH, Yokoyama WM. 2009. Natural killer cell tolerance licensing and other mechanisms. Adv.Immunol. 101:27–79
27. Kenneson A, Cannon MJ. 2007. Review and meta-analysis of the epidemiology of congenital cy-tomegalovirus (CMV) infection. Rev. Med. Virol. 17:253–76
28. McGeoch DJ, Dolan A, Ralph AC. 2000. Toward a comprehensive phylogeny for mammalian and avianherpesviruses. J. Virol. 74:10401–6
www.annualreviews.org • Control of Human Viral Infections by NK Cells 185
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
29. Loewendorf A, Benedict CA. 2010. Modulation of host innate and adaptive immune defenses by cy-tomegalovirus: Timing is everything. J. Intern. Med. 267:483–501
30. Sun JC, Lanier LL. 2009. The natural selection of herpesviruses and virus-specific NK cell receptors.Viruses 1:362
31. Daniels KA, Devora G, Lai WC, O’Donnell CL, Bennett M, Welsh RM. 2001. Murine cytomegalovirusis regulated by a discrete subset of natural killer cells reactive with monoclonal antibody to Ly49H.J. Exp. Med. 194:29–44
32. Dokun AO, Kim S, Smith HR, Kang HS, Chu DT, Yokoyama WM. 2001. Specific and nonspecific NKcell activation during virus infection. Nat. Immunol. 2:951–56
33. Arase H, Mocarski ES, Campbell AE, Hill AB, Lanier LL. 2002. Direct recognition of cytomegalovirusby activating and inhibitory NK cell receptors. Science 296:1323–26
34. Bubic I, Wagner M, Krmpotic A, Saulig T, Kim S, et al. 2004. Gain of virulence caused by loss of a genein murine cytomegalovirus. J. Virol. 78:7536–44
35. Desrosiers MP, Kielczewska A, Loredo-Osti JC, Adam SG, Makrigiannis AP, et al. 2005. Epistasisbetween mouse Klra and major histocompatibility complex class I loci is associated with a new mechanismof natural killer cell-mediated innate resistance to cytomegalovirus infection. Nat. Genet. 37:593–99
36. Kielczewska A, Pyzik M, Sun T, Krmpotic A, Lodoen MB, et al. 2009. Ly49P recognition ofcytomegalovirus-infected cells expressing H2-Dk and CMV-encoded m04 correlates with the NK cellantiviral response. J. Exp. Med. 206:515–23
37. Saverino D, Ghiotto F, Merlo A, Bruno S, Battini L, et al. 2004. Specific recognition of the viralprotein UL18 by CD85j/LIR-1/ILT2 on CD8+ T cells mediates the non-MHC-restricted lysis ofhuman cytomegalovirus-infected cells. J. Immunol. 172:5629–37
38. Prod’homme V, Griffin C, Aicheler RJ, Wang EC, McSharry BP, et al. 2007. The human cytomegalovirusMHC class I homolog UL18 inhibits LIR-1+ but activates LIR-1− NK cells. J. Immunol. 178:4473–81
39. Stern-Ginossar N, Elefant N, Zimmermann A, Wolf DG, Saleh N, et al. 2007. Host immune systemgene targeting by a viral miRNA. Science 317:376–81
40. Nachmani D, Lankry D, Wolf DG, Mandelboim O. 2010. The human cytomegalovirus microRNAmiR-UL112 acts synergistically with a cellular microRNA to escape immune elimination. Nat. Immunol.11:806–13
41. Tomasec P, Braud VM, Rickards C, Powell MB, McSharry BP, et al. 2000. Surface expression of HLA-E,an inhibitor of natural killer cells, enhanced by human cytomegalovirus gpUL40. Science 287:1031
42. Guma M, Angulo A, Vilches C, Gomez-Lozano N, Malats N, Lopez-Botet M. 2004. Imprint of humancytomegalovirus infection on the NK cell receptor repertoire. Blood 104:3664–71
43. Guma M, Budt M, Saez A, Brckalo T, Hengel H, et al. 2006. Expansion of CD94/NKG2C+ NK cellsin response to human cytomegalovirus-infected fibroblasts. Blood 107:3624–31
44. Lopez-Verges S, Milush JM, Schwartz BS, Pando MJ, Jarjoura J, et al. 2011. Expansion of a uniqueCD57+NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl.Acad. Sci. USA 108:14725–32
45. Della Chiesa M, Falco M, Podesta M, Locatelli F, Moretta L, et al. 2012. Phenotypic and functionalheterogeneity of human NK cells developing after umbilical cord blood transplantation: a role for humancytomegalovirus? Blood 119:399–410
46. Foley B, Cooley S, Verneris MR, Pitt M, Curtsinger J, et al. 2012. Cytomegalovirus reactivation afterallogeneic transplantation promotes a lasting increase in educated NKG2C+ natural killer cells withpotent function. Blood 119:2665–74
47. Bjorkstrom NK, Lindgren T, Stoltz M, Fauriat C, Braun M, et al. 2011. Rapid expansion and long-termpersistence of elevated NK cell numbers in humans infected with hantavirus. J. Exp. Med. 208:13–21
48. Guma M, Cabrera C, Erkizia I, Bofill M, Clotet B, et al. 2006. Human cytomegalovirus infection isassociated with increased proportions of NK cells that express the CD94/NKG2C receptor in aviremicHIV-1-positive patients. J. Infect. Dis. 194:38–41
49. Petitdemange C, Becquart P, Wauquier N, Beziat V, Debre P, et al. 2011. Unconventional repertoireprofile is imprinted during acute chikungunya infection for natural killer cells polarization toward cyto-toxicity. PLoS Pathog. 7:e1002268
186 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
50. Bjorkstrom NK, Svensson A, Malmberg KJ, Eriksson K, Ljunggren HG. 2011. Characterization ofnatural killer cell phenotype and function during recurrent human HSV-2 infection. PLoS ONE 6:e27664
51. Gazit R, Garty BZ, Monselise Y, Hoffer V, Finkelstein Y, et al. 2004. Expression of KIR2DL1 on theentire NK cell population: a possible novel immunodeficiency syndrome. Blood 103:1965–66
52. Stern M, Elsasser H, Honger G, Steiger J, Schaub S, Hess C. 2008. The number of activating KIR genesinversely correlates with the rate of CMV infection/reactivation in kidney transplant recipients. Am. J.Transplant. 8:1312–17
53. Cook M, Briggs D, Craddock C, Mahendra P, Milligan D, et al. 2006. Donor KIR genotype has a majorinfluence on the rate of cytomegalovirus reactivation following T-cell replete stem cell transplantation.Blood 107:1230–32
54. Virgin HW, Walker BD. 2010. Immunology and the elusive AIDS vaccine. Nature 464:224–3155. Brainard DM, Seung E, Frahm N, Cariappa A, Bailey CC, et al. 2009. Induction of robust cellular and hu-
moral virus-specific adaptive immune responses in human immunodeficiency virus-infected humanizedBLT mice. J. Virol. 83:7305–21
56. Dudek TE, No DC, Seung E, Vrbanac VD, Fadda L, et al. 2012. Rapid evolution of HIV-1 to functionalCD8+ T cell responses in humanized BLT mice. Sci. Transl. Med. 4:143ra98
57. Watkins DI, Burton DR, Kallas EG, Moore JP, Koff WC. 2008. Nonhuman primate models and thefailure of the Merck HIV-1 vaccine in humans. Nat. Med. 14:617–21
58. Valentine LE, Watkins DI. 2008. Relevance of studying T cell responses in SIV-infected rhesusmacaques. Trends Microbiol. 16:605–11
59. Hellmann I, Lim SY, Gelman RS, Letvin NL. 2011. Association of activating KIR copy number variationof NK cells with containment of SIV replication in rhesus monkeys. PLoS Pathog. 7:e1002436
60. Bimber B, O’Connor DH. 2008. KIRigami: the case for studying NK cell receptors in SIV+ macaques.Immunol. Res. 40:235–43
61. Colantonio AD, Bimber BN, Neidermyer WJ Jr, Reeves RK, Alter G, et al. 2011. KIR polymor-phisms modulate peptide-dependent binding to an MHC class I ligand with a Bw6 motif. PLoS Pathog.7:e1001316
62. Rosner C, Kruse PH, Hermes M, Otto N, Walter L. 2011. Rhesus macaque inhibitory and activatingKIR3D interact with Mamu-A-encoded ligands. J. Immunol. 186:2156–63
63. Webster RL, Johnson RP. 2005. Delineation of multiple subpopulations of natural killer cells in rhesusmacaques. Immunology 115:206–14
64. Schmitz JE, Kuroda MJ, Santra S, Sasseville VG, Simon MA, et al. 1999. Control of viremia in simianimmunodeficiency virus infection by CD8+ lymphocytes. Science 283:857–60
65. Jin X, Bauer DE, Tuttleton SE, Lewin S, Gettie A, et al. 1999. Dramatic rise in plasma viremia afterCD8+ T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 189:991–98
66. Choi EI, Reimann KA, Letvin NL. 2008. In vivo natural killer cell depletion during primary simianimmunodeficiency virus infection in rhesus monkeys. J. Virol. 82:6758–61
67. Sun Y, Asmal M, Lane S, Permar SR, Schmidt SD, et al. 2011. Antibody-dependent cell-mediatedcytotoxicity in simian immunodeficiency virus-infected rhesus monkeys. J. Virol. 85:6906–12
68. Asmal M, Sun Y, Lane S, Yeh W, Schmidt SD, et al. 2011. Antibody-dependent cell-mediated viralinhibition emerges after simian immunodeficiency virus SIVmac251 infection of rhesus monkeys coin-cident with gp140-binding antibodies and is effective against neutralization-resistant viruses. J. Virol.85:5465–75
69. Flores-Villanueva PO, Yunis EJ, Delgado JC, Vittinghoff E, Buchbinder S, et al. 2001. Control of HIV-1viremia and protection from AIDS are associated with HLA-Bw4 homozygosity. Proc. Natl. Acad. Sci.USA 98:5140–45
70. Martin MP, Gao X, Lee JH, Nelson GW, Detels R, et al. 2002. Epistatic interaction between KIR3DS1and HLA-B delays the progression to AIDS. Nat. Genet. 31:429–34
71. Martin MP, Qi Y, Gao X, Yamada E, Martin JN, et al. 2007. Innate partnership of HLA-B and KIR3DL1subtypes against HIV-1. Nat. Genet. 39:733–40
72. Boulet S, Kleyman M, Kim JY, Kamya P, Sharafi S, et al. 2008. A combined genotype of KIR3DL1 highexpressing alleles and HLA-B∗57 is associated with a reduced risk of HIV infection. AIDS 22:1487–91
www.annualreviews.org • Control of Human Viral Infections by NK Cells 187
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
73. Boulet S, Sharafi S, Simic N, Bruneau J, Routy JP, et al. 2008. Increased proportion of KIR3DS1homozygotes in HIV-exposed uninfected individuals. AIDS 22:595–99
74. Boulet S, Song R, Kamya P, Bruneau J, Shoukry NH, et al. 2010. HIV protective KIR3DL1 and HLA-B genotypes influence NK cell function following stimulation with HLA-devoid cells. J. Immunol.184:2057–64
75. Kamya P, Boulet S, Tsoukas CM, Routy JP, Thomas R, et al. 2011. Receptor-ligand requirementsfor increased NK cell polyfunctional potential in slow progressors infected with HIV-1 coexpressingKIR3DL1∗h/∗y and HLA-B∗57. J. Virol. 85:5949–60
76. Parsons MS, Boulet S, Song R, Bruneau J, Shoukry NH, et al. 2010. Mind the gap: lack of associa-tion between KIR3DL1∗004/HLA-Bw4-induced natural killer cell function and protection from HIVinfection. J. Infect. Dis. 202(Suppl. 3):S356–60
77. Long BR, Ndhlovu LC, Oksenberg JR, Lanier LL, Hecht FM, et al. 2008. Conferral of enhancednatural killer cell function by KIR3DS1 in early human immunodeficiency virus type 1 infection. J.Virol. 82:4785–92
78. Ravet S, Scott-Algara D, Bonnet E, Tran HK, Tran T, et al. 2007. Distinctive NK-cell receptor reper-toires sustain high-level constitutive NK-cell activation in HIV-exposed uninfected individuals. Blood109:4296–305
79. Pelak K, Need AC, Fellay J, Shianna KV, Feng S, et al. 2011. Copy number variation of KIR genesinfluences HIV-1 control. PLoS Biol. 9:e1001208
80. Fellay J, Shianna KV, Ge D, Colombo S, Ledergerber B, et al. 2007. A whole-genome association studyof major determinants for host control of HIV-1. Science 317:944–47
81. Pereyra F, Jia X, McLaren PJ, Telenti A, de Bakker PI, et al. 2010. The major genetic determinants ofHIV-1 control affect HLA class I peptide presentation. Science 330:1551–57
82. Kulkarni S, Savan R, Qi Y, Gao X, Yuki Y, et al. 2011. Differential microRNA regulation of HLA-Cexpression and its association with HIV control. Nature 472:495–98
83. Kiepiela P, Leslie AJ, Honeyborne I, Ramduth D, Thobakgale C, et al. 2004. Dominant influence ofHLA-B in mediating the potential co-evolution of HIV and HLA. Nature 432:769–75
84. Cohen GB, Gandhi RT, Davis DM, Mandelboim O, Chen BK, et al. 1999. The selective downregulationof class I major histocompatibility complex proteins by HIV-1 protects HIV-infected cells from NK cells.Immunity 10:661–71
85. Hansasuta P, Dong T, Thananchai H, Weekes M, Willberg C, et al. 2004. Recognition of HLA-A3 andHLA-A11 by KIR3DL2 is peptide-specific. Eur. J. Immunol. 34:1673–79
86. Alter G, Rihn S, Walter K, Nolting A, Martin M, et al. 2009. HLA class I subtype-dependent expansionof KIR3DS1+ and KIR3DL1+ NK cells during acute human immunodeficiency virus type 1 infection.J. Virol. 83:6798–805
87. Alter G, Teigen N, Ahern R, Streeck H, Meier A, et al. 2007. Evolution of innate and adaptive effectorcell functions during acute HIV-1 infection. J. Infect. Dis. 195:1452–60
88. Elliott JM, Yokoyama WM. 2011. Unifying concepts of MHC-dependent natural killer cell education.Trends Immunol. 32:364–72
89. Elliott JM, Wahle JA, Yokoyama WM. 2010. MHC class I-deficient natural killer cells acquire a licensedphenotype after transfer into an MHC class I-sufficient environment. J. Exp. Med. 207:2073–79
90. Kim S, Sunwoo JB, Yang L, Choi T, Song YJ, et al. 2008. HLA alleles determine differences in humannatural killer cell responsiveness and potency. Proc. Natl. Acad. Sci. USA 105:3053–58
91. Kim S, Poursine-Laurent J, Truscott SM, Lybarger L, Song YJ, et al. 2005. Licensing of natural killercells by host major histocompatibility complex class I molecules. Nature 436:709–13
92. Alter G, Martin MP, Teigen N, Carr WH, Suscovich TJ, et al. 2007. Differential natural killer cell-mediated inhibition of HIV-1 replication based on distinct KIR/HLA subtypes. J. Exp. Med. 204:3027–36
93. Carr WH, Rosen DB, Arase H, Nixon DF, Michaelsson J, Lanier LL. 2007. Cutting edge: KIR3DS1, agene implicated in resistance to progression to AIDS, encodes a DAP12-associated receptor expressedon NK cells that triggers NK cell activation. J. Immunol. 178:647–51
94. Gardiner CM, Guethlein LA, Shilling HG, Pando M, Carr WH, et al. 2001. Different NK cell surfacephenotypes defined by the DX9 antibody are due to KIR3DL1 gene polymorphism. J. Immunol. 166:2992–3001
188 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
95. O’Connor GM, Yamada E, Rampersaud A, Thomas R, Carrington M, McVicar DW. 2011. Analysisof binding of KIR3DS1∗014 to HLA suggests distinct evolutionary history of KIR3DS1. J. Immunol.187:2162–71
96. Fauriat C, Ivarsson MA, Ljunggren HG, Malmberg KJ, Michaelsson J. 2010. Education of human naturalkiller cells by activating killer cell immunoglobulin-like receptors. Blood 115:1166–74
97. Hollenbach JA, Meenagh A, Sleator C, Alaez C, Bengoche M, et al. 2010. Report from the killerimmunoglobulin-like receptor (KIR) anthropology component of the 15th International Histocom-patibility Workshop: worldwide variation in the KIR loci and further evidence for the co-evolution ofKIR and HLA. Tissue Antigens 76:9–17
98. Peruzzi M, Wagtmann N, Long EO. 1996. A p70 killer cell inhibitory receptor specific for severalHLA-B allotypes discriminates among peptides bound to HLA-B∗2705. J. Exp. Med. 184:1585–90
99. Rajagopalan S, Long EO. 1997. The direct binding of a p58 killer cell inhibitory receptor to humanhistocompatibility leukocyte antigen (HLA)-Cw4 exhibits peptide selectivity. J. Exp. Med. 185:1523–28
100. Altfeld M, Goulder P. 2007. ‘Unleashed’ natural killers hinder HIV. Nat. Genet. 39:708–10101. Boyington JC, Motyka SA, Schuck P, Brooks AG, Sun PD. 2000. Crystal structure of an NK cell
immunoglobulin-like receptor in complex with its class I MHC ligand. Nature 405:537–43102. Fadda L, O’Connor GM, Kumar S, Piechocka-Trocha A, Gardiner CM, et al. 2011. Common HIV-1
peptide variants mediate differential binding of KIR3DL1 to HLA-Bw4 molecules. J. Virol. 85:5970–74103. Brackenridge S, Evans EJ, Toebes M, Goonetilleke N, Liu MK, et al. 2011. An early HIV mutation
within an HLA-B∗57-restricted T cell epitope abrogates binding to the killer inhibitory receptor 3DL1.J. Virol. 85:5415–22
104. Thananchai H, Gillespie G, Martin MP, Bashirova A, Yawata N, et al. 2007. Cutting edge: allele-specificand peptide-dependent interactions between KIR3DL1 and HLA-A and HLA-B. J. Immunol. 178:33–37
105. Thananchai H, Makadzange T, Maenaka K, Kuroki K, Peng Y, et al. 2009. Reciprocal recognition ofan HLA-Cw4-restricted HIV-1 gp120 epitope by CD8+ T cells and NK cells. AIDS 23:189–93
106. Fadda L, Korner C, Kumar S, van Teijlingen NH, Piechocka-Trocha A, et al. 2012. HLA-Cw∗0102-restricted HIV-1 p24 epitope variants can modulate the binding of the inhibitory KIR2DL2 receptorand primary NK cell function. PLoS Pathog. 8:e1002805
107. Alter G, Heckerman D, Schneidewind A, Fadda L, Kadie CM, et al. 2011. HIV-1 adaptation to NK-cell-mediated immune pressure. Nature 476:96–100
108. Cerboni C, Neri F, Casartelli N, Zingoni A, Cosman D, et al. 2007. Human immunodeficiency virus1 Nef protein downmodulates the ligands of the activating receptor NKG2D and inhibits natural killercell-mediated cytotoxicity. J. Gen. Virol. 88:242–50
109. Jost S, Altfeld M. 2012. Evasion from NK cell-mediated immune responses by HIV-1. Microbes Infect.14:904–15
110. Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M. 1989. Isolation of a cDNA clonederived from a blood-borne non-A, non-B viral hepatitis genome. Science 244:359–62
111. Lavanchy D. 2009. The global burden of hepatitis C. Liver Int. 29(Suppl. 1):74–81112. Day CL, Lauer GM, Robbins GK, McGovern B, Wurcel AG, et al. 2002. Broad specificity of virus-
specific CD4+ T-helper-cell responses in resolved hepatitis C virus infection. J. Virol. 76:12584–95113. Lauer GM, Ouchi K, Chung RT, Nguyen TN, Day CL, et al. 2002. Comprehensive analysis of CD8+-
T-cell responses against hepatitis C virus reveals multiple unpredicted specificities. J. Virol. 76:6104–13114. Grakoui A, Shoukry NH, Woollard DJ, Han JH, Hanson HL, et al. 2003. HCV persistence and immune
evasion in the absence of memory T cell help. Science 302:659–62115. Ulsenheimer A, Gerlach JT, Gruener NH, Jung MC, Schirren CA, et al. 2003. Detection of functionally
altered hepatitis C virus-specific CD4 T cells in acute and chronic hepatitis C. Hepatology 37:1189–98116. Welsch C, Jesudian A, Zeuzem S, Jacobson I. 2012. New direct-acting antiviral agents for the treatment
of hepatitis C virus infection and perspectives. Gut 61(Suppl. 1):i36–46117. Guidotti LG, Chisari FV. 2006. Immunobiology and pathogenesis of viral hepatitis. Annu. Rev. Pathol.
Mech. Dis. 1:23–61118. Marcello T, Grakoui A, Barba-Spaeth G, Machlin ES, Kotenko SV, et al. 2006. Interferons α and
λ inhibit hepatitis C virus replication with distinct signal transduction and gene regulation kinetics.Gastroenterology 131:1887–98
www.annualreviews.org • Control of Human Viral Infections by NK Cells 189
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
119. Grebely J, Petoumenos K, Hellard M, Matthews GV, Suppiah V, et al. 2010. Potential role forinterleukin-28B genotype in treatment decision-making in recent hepatitis C virus infection. Hepatology52:1216–24
120. Tillmann HL, Thompson AJ, Patel K, Wiese M, Tenckhoff H, et al. 2010. A polymorphism nearIL28B is associated with spontaneous clearance of acute hepatitis C virus and jaundice. Gastroenterology139:1586–92
121. Thomas DL, Thio CL, Martin MP, Qi Y, Ge D, et al. 2009. Genetic variation in IL28B and spontaneousclearance of hepatitis C virus. Nature 461:798–801
122. Rauch A, Kutalik Z, Descombes P, Cai T, Di Iulio J, et al. 2010. Genetic variation in IL28B is associ-ated with chronic hepatitis C and treatment failure: a genome-wide association study. Gastroenterology138:1338–45
123. Doherty DG, O’Farrelly C. 2000. Innate and adaptive lymphoid cells in the human liver. Immunol. Rev.174:5–20
124. Lassen MG, Lukens JR, Dolina JS, Brown MG, Hahn YS. 2010. Intrahepatic IL-10 maintainsNKG2A+Ly49− liver NK cells in a functionally hyporesponsive state. J. Immunol. 184:2693–701
125. Shi FD, Ljunggren HG, La Cava A, Van Kaer L. 2011. Organ-specific features of natural killer cells.Nat. Rev. Immunol. 11:658–71
126. Khakoo SI, Thio CL, Martin MP, Brooks CR, Gao X, et al. 2004. HLA and NK cell inhibitory receptorgenes in resolving hepatitis C virus infection. Science 305:872–74
127. Knapp S, Warshow U, Hegazy D, Brackenbury L, Guha IN, et al. 2010. Consistent beneficial effectsof killer cell immunoglobulin-like receptor 2DL3 and group 1 human leukocyte antigen-C followingexposure to hepatitis C virus. Hepatology 51:1168–75
128. Romero V, Azocar J, Zuniga J, Clavijo OP, Terreros D, et al. 2008. Interaction of NK inhibitory receptorgenes with HLA-C and MHC class II alleles in hepatitis C virus infection outcome. Mol. Immunol.45:2429–36
129. Vidal-Castineira JR, Lopez-Vazquez A, Diaz-Pena R, Alonso-Arias R, Martinez-Borra J, et al. 2010.Effect of killer immunoglobulin-like receptors in the response to combined treatment in patients withchronic hepatitis C virus infection. J. Virol. 84:475–81
130. Marangon AV, Silva GF, de Moraes CF, Grotto RM, Pardini MI, et al. 2011. KIR genes and theirhuman leukocyte antigen ligands in the progression to cirrhosis in patients with chronic hepatitis C.Hum. Immunol. 72:1074–78
131. Alter G, Jost S, Rihn S, Reyor L, Nolan B, et al. 2010. Reduced frequencies of NKp30+NKp46+,CD161+, and NKG2D+ NK cells in acute HCV infection may predict viral clearance. J. Hepatol.55:278–88
132. Yawata M, Yawata N, Draghi M, Partheniou F, Little AM, Parham P. 2008. MHC class I-specificinhibitory receptors and their ligands structure diverse human NK-cell repertoires toward a balance ofmissing self-response. Blood 112:2369–80
133. Rauch A, Laird R, McKinnon E, Telenti A, Furrer H, et al. 2007. Influence of inhibitory killerimmunoglobulin-like receptors and their HLA-C ligands on resolving hepatitis C virus infection. TissueAntigens 69(Suppl. 1):237–40
134. Amadei B, Urbani S, Cazaly A, Fisicaro P, Zerbini A, et al. 2010. Activation of natural killer cells duringacute infection with hepatitis C virus. Gastroenterology 138:1536–45
135. Moesta AK, Norman PJ, Yawata M, Yawata N, Gleimer M, Parham P. 2008. Synergistic polymorphismat two positions distal to the ligand-binding site makes KIR2DL2 a stronger receptor for HLA-C thanKIR2DL3. J. Immunol. 180:3969–79
136. Fadda L, Borhis G, Ahmed P, Cheent K, Pageon SV, et al. 2010. Peptide antagonism as a mechanismfor NK cell activation. Proc. Natl. Acad. Sci. USA 107:10160–65
137. Paladino N, Flores AC, Marcos CY, Fainboim H, Theiler G, et al. 2007. Increased frequencies ofactivating natural killer receptors are associated with liver injury in individuals who do not eliminatehepatitis C virus. Tissue Antigens 69(Suppl. 1):109–11
138. Lopez-Vazquez A, Rodrigo L, Martinez-Borra J, Perez R, Rodriguez M, et al. 2005. Protective effectof the HLA-Bw4I80 epitope and the killer cell immunoglobulin-like receptor 3DS1 gene against the
190 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
development of hepatocellular carcinoma in patients with hepatitis C virus infection. J. Infect. Dis.192:162–65
139. Zuniga J, Romero V, Azocar J, Terreros D, Vargas-Rojas MI, et al. 2009. Protective KIR-HLA interac-tions for HCV infection in intravenous drug users. Mol. Immunol. 46:2723–77
140. Katz G, Gazit R, Arnon TI, Gonen-Gross T, Tarcic G, et al. 2004. MHC class I-independent recognitionof NK-activating receptor KIR2DS4. J. Immunol. 173:1819–25
141. Carneiro VL, Lemaire DC, Bendicho MT, Souza SL, Cavalcante LN, et al. 2010. Natural killer cellreceptor and HLA-C gene polymorphisms among patients with hepatitis C: a comparison betweensustained virological responders and non-responders. Liver Int. 30:567–73
142. Beziat V, Dalgard O, Asselah T, Halfon P, Bedossa P, et al. 2012. CMV drives clonal expansion ofNKG2C+ NK cells expressing self-specific KIRs in chronic hepatitis patients. Eur. J. Immunol. 42:447–57
143. Bukh J. 2012. Animal models for the study of hepatitis C virus infection and related liver disease.Gastroenterology 142:1279–87
144. Wang SH, Huang CX, Ye L, Wang X, Song L, et al. 2008. Natural killer cells suppress full cycle HCVinfection of human hepatocytes. J. Viral Hepat. 15:855–64
145. Tseng CT, Klimpel GR. 2002. Binding of the hepatitis C virus envelope protein E2 to CD81 inhibitsnatural killer cell functions. J. Exp. Med. 195:43–49
146. Crotta S, Brazzoli M, Piccioli D, Valiante NM, Wack A. 2010. Hepatitis C virions subvert natural killercell activation to generate a cytokine environment permissive for infection. J. Hepatol. 52:183–90
147. Crotta S, Stilla A, Wack A, D’Andrea A, Nuti S, et al. 2002. Inhibition of natural killer cells throughengagement of CD81 by the major hepatitis C virus envelope protein. J. Exp. Med. 195:35–41
148. Yoon JC, Shiina M, Ahlenstiel G, Rehermann B. 2009. Natural killer cell function is intact after directexposure to infectious hepatitis C virions. Hepatology 49:12–21
149. Herzer K, Falk CS, Encke J, Eichhorst ST, Ulsenheimer A, et al. 2003. Upregulation of major histo-compatibility complex class I on liver cells by hepatitis C virus core protein via p53 and TAP1 impairsnatural killer cell cytotoxicity. J. Virol. 77:8299–309
150. Nattermann J, Nischalke HD, Hofmeister V, Ahlenstiel G, Zimmermann H, et al. 2005. The HLA-A2restricted T cell epitope HCV core 35-44 stabilizes HLA-E expression and inhibits cytolysis mediatedby natural killer cells. Am. J. Pathol. 166:443–53
151. Wen C, He X, Ma H, Hou N, Wei C, et al. 2008. Hepatitis C virus infection downregulates the ligandsof the activating receptor NKG2D. Cell. Mol. Immunol. 5:475–78
152. Mondelli MU, Varchetta S, Oliviero B. 2010. Natural killer cells in viral hepatitis: facts and controversies.Eur. J. Clin. Investig. 40:851–63
153. Par G, Rukavina D, Podack ER, Horanyi M, Szekeres-Bartho J, et al. 2002. Decrease in CD3-negative-CD8dim+ and Vδ2/Vγ9 TcR+ peripheral blood lymphocyte counts, low perforin expression and theimpairment of natural killer cell activity is associated with chronic hepatitis C virus infection. J. Hepatol.37:514–22
154. Bonavita MS, Franco A, Paroli M, Santilio I, Benvenuto R, et al. 1993. Normalization of depressednatural killer activity after interferon-α therapy is associated with a low frequency of relapse in patientswith chronic hepatitis C. Int. J. Tissue React. 15:11–16
155. Corado J, Toro F, Rivera H, Bianco NE, Deibis L, De Sanctis JB. 1997. Impairment of natural killer(NK) cytotoxic activity in hepatitis C virus (HCV) infection. Clin. Exp. Immunol. 109:451–57
156. Morishima C, Paschal DM, Wang CC, Yoshihara CS, Wood BL, et al. 2006. Decreased NK cell fre-quency in chronic hepatitis C does not affect ex vivo cytolytic killing. Hepatology 43:573–80
157. Ahlenstiel G, Titerence RH, Koh C, Edlich B, Feld JJ, et al. 2010. Natural killer cells are polarized towardcytotoxicity in chronic hepatitis C in an interferon-alfa-dependent manner. Gastroenterology 138:325–35
158. De Maria A, Fogli M, Mazza S, Basso M, Picciotto A, et al. 2007. Increased natural cytotoxicity receptorexpression and relevant IL-10 production in NK cells from chronically infected viremic HCV patients.Eur. J. Immunol. 37:445–55
159. Oliviero B, Varchetta S, Paudice E, Michelone G, Zaramella M, et al. 2009. Natural killer cell functionaldichotomy in chronic hepatitis B and chronic hepatitis C virus infections. Gastroenterology 137:1151–60
www.annualreviews.org • Control of Human Viral Infections by NK Cells 191
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
160. Duesberg U, Schneiders AM, Flieger D, Inchauspe G, Sauerbruch T, Spengler U. 2001. Natural cyto-toxicity and antibody-dependent cellular cytotoxicity (ADCC) is not impaired in patients suffering fromchronic hepatitis C. J. Hepatol. 35:650–57
161. Golden-Mason L, Madrigal-Estebas L, McGrath E, Conroy MJ, Ryan EJ, et al. 2008. Altered naturalkiller cell subset distributions in resolved and persistent hepatitis C virus infection following single sourceexposure. Gut 57:1121–28
162. Guidotti LG, Chisari FV. 2001. Noncytolytic control of viral infections by the innate and adaptiveimmune response. Annu. Rev. Immunol. 19:65–91
163. Pelletier S, Drouin C, Bedard N, Khakoo SI, Bruneau J, Shoukry NH. 2010. Increased degranulationof natural killer cells during acute HCV correlates with the magnitude of virus-specific T cell responses.J. Hepatol. 53:805–16
164. Cheent K, Khakoo SI. 2011. Natural killer cells and hepatitis C: action and reaction. Gut 60:268–78165. Bonorino P, Ramzan M, Camous X, Dufeu-Duchesne T, Thelu MA, et al. 2009. Fine characterization
of intrahepatic NK cells expressing natural killer receptors in chronic hepatitis B and C. J. Hepatol.51:458–67
166. Harrison RJ, Ettorre A, Little AM, Khakoo SI. 2010. Association of NKG2A with treatment for chronichepatitis C virus infection. Clin. Exp. Immunol. 161:306–14
167. Jinushi M, Takehara T, Tatsumi T, Kanto T, Miyagi T, et al. 2004. Negative regulation of NK cellactivities by inhibitory receptor CD94/NKG2A leads to altered NK cell-induced modulation of dendriticcell functions in chronic hepatitis C virus infection. J. Immunol. 173:6072–81
168. Golden-Mason L, Kelly AM, Doherty DG, Traynor O, McEntee G, et al. 2004. Hepatic interleuklin15 (IL-15) expression: implications for local NK/NKT cell homeostasis and development. Clin. Exp.Immunol. 138:94–101
169. Nattermann J, Feldmann G, Ahlenstiel G, Langhans B, Sauerbruch T, Spengler U. 2006. Surface ex-pression and cytolytic function of natural killer cell receptors is altered in chronic hepatitis C. Gut55:869–77
170. Gunturi A, Berg RE, Forman J. 2004. The role of CD94/NKG2 in innate and adaptive immunity.Immunol. Res. 30:29–34
171. Kramer B, Korner C, Kebschull M, Glassner A, Eisenhardt M, et al. 2012. Natural killer p46High
expression defines a natural killer cell subset that is potentially involved in control of hepatitis C virusreplication and modulation of liver fibrosis. Hepatology 56:1201–13
172. Golden-Mason L, Cox AL, Randall JA, Cheng L, Rosen HR. 2010. Increased natural killer cell cyto-toxicity and NKp30 expression protects against hepatitis C virus infection in high-risk individuals andinhibits replication in vitro. Hepatology 52:1581–89
173. Morens DM, Taubenberger JK. 2011. Pandemic influenza: certain uncertainties. Rev. Med. Virol. 21:262–84
174. World Health Organization. 2009. Influenza (seasonal). Factsheet No. 211, April, World Health Org.,Geneva. http://www.who.int/mediacentre/factsheets/fs211/en/
175. Thompson WW, Shay DK, Weintraub E, Brammer L, Bridges CB, et al. 2004. Influenza-associatedhospitalizations in the United States. JAMA 292:1333–40
176. Thompson WW, Shay DK, Weintraub E, Brammer L, Cox N, et al. 2003. Mortality associated withinfluenza and respiratory syncytial virus in the United States. JAMA 289:179–86
177. Thompson WW, Weintraub E, Dhankhar P, Cheng PY, Brammer L, et al. 2009. Estimates of USinfluenza-associated deaths made using four different methods. Influenza Other Respir. Viruses 3:37–49
178. Doherty PC, Topham DJ, Tripp RA, Cardin RD, Brooks JW, Stevenson PG. 1997. Effector CD4+ andCD8+ T-cell mechanisms in the control of respiratory virus infections. Immunol. Rev. 159:105–17
179. Thomas PG, Keating R, Hulse-Post DJ, Doherty PC. 2006. Cell-mediated protection in influenzainfection. Emerg. Infect. Dis. 12:48–54
180. Ennis FA, Meager A, Beare AS, Qi YH, Riley D, et al. 1981. Interferon induction and increased naturalkiller-cell activity in influenza infections in man. Lancet 2:891–93
181. Gregoire C, Chasson L, Luci C, Tomasello E, Geissmann F, et al. 2007. The trafficking of natural killercells. Immunol. Rev. 220:169–82
192 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
182. Reynolds CW, Timonen T, Herberman RB. 1981. Natural killer (NK) cell activity in the rat. I. Isolationand characterization of the effector cells. J. Immunol. 127:282–87
183. Stein-Streilein J, Bennett M, Mann D, Kumar V. 1983. Natural killer cells in mouse lung: surfacephenotype, target preference, and response to local influenza virus infection. J. Immunol. 131:2699–704
184. Nogusa S, Ritz BW, Kassim SH, Jennings SR, Gardner EM. 2008. Characterization of age-relatedchanges in natural killer cells during primary influenza infection in mice. Mech. Ageing Dev. 129:223–30
185. Stein-Streilein J, Guffee J. 1986. In vivo treatment of mice and hamsters with antibodies to asialo GM1increases morbidity and mortality to pulmonary influenza infection. J. Immunol. 136:1435–41
186. Stein-Streilein J, Guffee J, Fan W. 1988. Locally and systemically derived natural killer cells participatein defense against intranasally inoculated influenza virus. Reg. Immunol. 1:100–5
187. Liu B, Mori I, Hossain MJ, Dong L, Takeda K, Kimura Y. 2004. Interleukin-18 improves the earlydefence system against influenza virus infection by augmenting natural killer cell-mediated cytotoxicity.J. Gen. Virol. 85:423–28
188. Dong L, Mori I, Hossain MJ, Kimura Y. 2000. The senescence-accelerated mouse shows aging-relateddefects in cellular but not humoral immunity against influenza virus infection. J. Infect. Dis. 182:391–96
189. Abdul-Careem MF, Mian MF, Yue G, Gillgrass A, Chenoweth MJ, et al. 2012. Critical role of naturalkiller cells in lung immunopathology during influenza infection in mice. J. Infect. Dis. 206:167–77
190. Verbist KC, Rose DL, Cole CJ, Field MB, Klonowski KD. 2012. IL-15 participates in the respiratoryinnate immune response to influenza virus infection. PLoS ONE 7:e37539
191. Gazit R, Gruda R, Elboim M, Arnon TI, Katz G, et al. 2006. Lethal influenza infection in the absenceof the natural killer cell receptor gene Ncr1. Nat. Immunol. 7:517–23
192. Glasner A, Zurunic A, Meningher T, Lenac Rovis T, Tsukerman P, et al. 2012. Elucidating the mech-anisms of influenza virus recognition by Ncr1. PLoS ONE 7:e36837
193. Achdout H, Meningher T, Hirsh S, Glasner A, Bar-On Y, et al. 2010. Killing of avian and swine influenzavirus by natural killer cells. J. Virol. 84:3993–4001
194. Narni-Mancinelli E, Jaeger BN, Bernat C, Fenis A, Kung S, et al. 2012. Tuning of natural killer cellreactivity by NKp46 and Helios calibrates T cell responses. Science 335:344–48
195. Hashimoto G, Wright PF, Karzon DT. 1983. Antibody-dependent cell-mediated cytotoxicity againstinfluenza virus-infected cells. J. Infect. Dis. 148:785–94
196. Jegerlehner A, Schmitz N, Storni T, Bachmann MF. 2004. Influenza A vaccine based on the extracellulardomain of M2: weak protection mediated via antibody-dependent NK cell activity. J. Immunol. 172:5598–605
197. Ahlenstiel G, Martin MP, Gao X, Carrington M, Rehermann B. 2008. Distinct KIR/HLA compoundgenotypes affect the kinetics of human antiviral natural killer cell responses. J. Clin. Investig. 118:1017–26
198. La D, Czarnecki C, El-Gabalawy H, Kumar A, Meyers AF, et al. 2011. Enrichment of variations inKIR3DL1/S1 and KIR2DL2/L3 among H1N1/09 ICU patients: an exploratory study. PLoS ONE6:e29200
199. Aranda-Romo S, Garcia-Sepulveda CA, Comas-Garcia A, Lovato-Salas F, Salgado-Bustamante M, et al.2012. Killer-cell immunoglobulin-like receptors (KIR) in severe A (H1N1) 2009 influenza infections.Immunogenetics 64:653–62
200. Arnon TI, Achdout H, Lieberman N, Gazit R, Gonen-Gross T, et al. 2004. The mechanisms controllingthe recognition of tumor- and virus-infected cells by NKp46. Blood 103:664–72
201. Draghi M, Pashine A, Sanjanwala B, Gendzekhadze K, Cantoni C, et al. 2007. NKp46 and NKG2Drecognition of infected dendritic cells is necessary for NK cell activation in the human response toinfluenza infection. J. Immunol. 178:2688–98
202. Du N, Zhou J, Lin X, Zhang Y, Yang X, et al. 2010. Differential activation of NK cells by influenza Apseudotype H5N1 and 1918 and 2009 pandemic H1N1 viruses. J. Virol. 84:7822–31
203. Jost S, Reardon J, Peterson E, Poole D, Bosch R, et al. 2010. Expansion of 2B4+ NK cells and decreasein NKp46+ NK cells in response to influenza. Immunology 132:516–26
204. Owen RE, Yamada E, Thompson CI, Phillipson LJ, Thompson C, et al. 2007. Alterations in receptorbinding properties of recent human influenza H3N2 viruses are associated with reduced natural killercell lysis of infected cells. J. Virol. 81:11170–78
www.annualreviews.org • Control of Human Viral Infections by NK Cells 193
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31CH07-Altfeld ARI 14 February 2013 21:16
205. Zhang M, Gaschen B, Blay W, Foley B, Haigwood N, et al. 2004. Tracking global patterns of N-linkedglycosylation site variation in highly variable viral glycoproteins: HIV, SIV, and HCV envelopes andinfluenza hemagglutinin. Glycobiology 14:1229–46
206. Achdout H, Arnon TI, Markel G, Gonen-Gross T, Katz G, et al. 2003. Enhanced recognition of humanNK receptors after influenza virus infection. J. Immunol. 171:915–23
207. Achdout H, Manaster I, Mandelboim O. 2008. Influenza virus infection augments NK cell inhibitionthrough reorganization of major histocompatibility complex class I proteins. J. Virol. 82:8030–37
208. He XS, Holmes TH, Zhang C, Mahmood K, Kemble GW, et al. 2006. Cellular immune responses inchildren and adults receiving inactivated or live attenuated influenza vaccines. J. Virol. 80:11756–66
209. Long BR, Michaelsson J, Loo CP, Ballan WM, Vu BA, et al. 2008. Elevated frequency of gammainterferon-producing NK cells in healthy adults vaccinated against influenza virus. Clin. Vaccine Immunol.15:120–30
210. Denney L, Aitken C, Li CK, Wilson-Davies E, Kok WL, et al. 2010. Reduction of natural killer butnot effector CD8 T lymphocytes in three consecutive cases of severe/lethal H1N1/09 influenza A virusinfection. PLoS ONE 5:e10675
211. Heltzer ML, Coffin SE, Maurer K, Bagashev A, Zhang Z, et al. 2009. Immune dysregulation in severeinfluenza. J. Leukoc. Biol. 85:1036–43
212. Jost S, Quillay H, Reardon J, Peterson E, Simmons RP, et al. 2011. Changes in cytokine levels and NKcell activation associated with influenza. PLoS ONE 6:e25060
213. Guo X, Chen Y, Li X, Kong H, Yang S, et al. 2011. Dynamic variations in the peripheral blood lymphocytesubgroups of patients with 2009 pandemic H1N1 swine-origin influenza A virus infection. Virol. J. 8:215
214. Fox A, Le NM, Horby P, van Doorn HR, Nguyen VT, et al. 2012. Severe pandemic H1N1 2009 infectionis associated with transient NK and T deficiency and aberrant CD8 responses. PLoS ONE 7:e31535
215. Guo H, Kumar P, Moran TM, Garcia-Sastre A, Zhou Y, Malarkannan S. 2009. The functional impair-ment of natural killer cells during influenza virus infection. Immunol. Cell Biol. 87:579–89
216. Mao H, Tu W, Qin G, Law HK, Sia SF, et al. 2009. Influenza virus directly infects human natural killercells and induces cell apoptosis. J. Virol. 83:9215–22
194 Jost · Altfeld
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31-Frontmatter ARI 27 February 2013 7:16
Annual Review ofImmunology
Volume 31, 2013
ContentsYears in Cologne
Klaus Rajewsky � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
The Biology of Recent Thymic EmigrantsPamela J. Fink � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �31
Immunogenic Cell Death in Cancer TherapyGuido Kroemer, Lorenzo Galluzzi, Oliver Kepp, and Laurence Zitvogel � � � � � � � � � � � � � � � � �51
Recognition of Bacteria by InflammasomesJakob von Moltke, Janelle S. Ayres, Eric M. Kofoed, Joseph Chavarrıa-Smith,
and Russell E. Vance � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �73
The Immunology of FibrosisGeorg Wick, Cecilia Grundtman, Christina Mayerl, Thomas-Florian Wimpissinger,
Johann Feichtinger, Bettina Zelger, Roswitha Sgonc, and Dolores Wolfram � � � � � � � � � 107
Memory T Cell Subsets, Migration Patterns, and Tissue ResidenceScott N. Mueller, Thomas Gebhardt, Francis R. Carbone, and William R. Heath � � � � � 137
Control of Human Viral Infections by Natural Killer CellsStephanie Jost and Marcus Altfeld � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 163
Functional T Cell Immunodeficiencies (with T Cells Present)Luigi D. Notarangelo � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 195
Controlling Natural Killer Cell Responses: Integration of Signals forActivation and InhibitionEric O. Long, Hun Sik Kim, Dongfang Liu, Mary E. Peterson,
and Sumati Rajagopalan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 227
Metabolic Regulation of T LymphocytesNancie J. MacIver, Ryan D. Michalek, and Jeffrey C. Rathmell � � � � � � � � � � � � � � � � � � � � � � � � 259
Mesenchymal Stem Cell: Keystone of the Hematopoietic Stem Cell Nicheand a Stepping-Stone for Regenerative MedicinePaul S. Frenette, Sandra Pinho, Daniel Lucas, and Christoph Scheiermann � � � � � � � � � � � 285
Interleukin-4- and Interleukin-13-Mediated Alternatively ActivatedMacrophages: Roles in Homeostasis and DiseaseSteven J. Van Dyken and Richard M. Locksley � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 317
Brain-Reactive Antibodies and DiseaseB. Diamond, G. Honig, S. Mader, L. Brimberg, and B.T. Volpe � � � � � � � � � � � � � � � � � � � � � � � 345
v
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.

IY31-Frontmatter ARI 27 February 2013 7:16
Immunology of the Maternal-Fetal InterfaceAdrian Erlebacher � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 387
Regulation of Ligands for the NKG2D Activating ReceptorDavid H. Raulet, Stephan Gasser, Benjamin G. Gowen, Weiwen Deng,
and Heiyoun Jung � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 413
Pathways of Antigen ProcessingJanice S. Blum, Pamela A. Wearsch, and Peter Cresswell � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 443
The Immune Response in TuberculosisAnne O’Garra, Paul S. Redford, Finlay W. McNab, and Chloe I. Bloom,
Robert J. Wilkinson, and Matthew P.R. Berry � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 475
The Adaptable Major Histocompatibility Complex (MHC) Fold: Structureand Function of Nonclassical and MHC Class I–Like MoleculesErin J. Adams and Adrienne M. Luoma � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 529
The Dendritic Cell Lineage: Ontogeny and Function of Dendritic Cells andTheir Subsets in the Steady State and the Inflamed SettingMiriam Merad, Priyanka Sathe, Julie Helft, Jennifer Miller, and Arthur Mortha � � � 563
T Cell–Mediated Host Immune Defenses in the LungKong Chen and Jay K. Kolls � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 605
Human Hemato-Lymphoid System Mice: Current Use and Future Potentialfor MedicineAnthony Rongvaux, Hitoshi Takizawa, Till Strowig, Tim Willinger,
Elizabeth E. Eynon, Richard A. Flavell, and Markus G. Manz � � � � � � � � � � � � � � � � � � � � � � 635
Signaling by the Phosphoinositide 3-Kinase Family in Immune CellsKlaus Okkenhaug � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 675
Broadly Neutralizing Antiviral AntibodiesDavide Corti and Antonio Lanzavecchia � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 705
Molecular Control of Steady-State Dendritic Cell Maturation and ImmuneHomeostasisGianna Elena Hammer and Averil Ma � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 743
Indexes
Cumulative Index of Contributing Authors, Volumes 21–31 � � � � � � � � � � � � � � � � � � � � � � � � � � � 793
Cumulative Index of Articles Titles, Volumes 21–31 � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 800
Errata
An online log of corrections to Annual Review of Immunology articles may be found athttp://immunol.annualreviews.org/errata.shtml
vi Contents
Ann
u. R
ev. I
mm
unol
. 201
3.31
:163
-194
. Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
WIB
6063
- M
ediz
inis
che
Hoc
hsch
ule
Han
nove
r (M
HH
) on
09/
25/1
3. F
or p
erso
nal u
se o
nly.