context-specific saccadic adaptation in monkeys
TRANSCRIPT

Vision Research 50 (2010) 2403–2410
Contents lists available at ScienceDirect
Vision Research
journal homepage: www.elsevier .com/locate /v isres
Context-specific saccadic adaptation in monkeys
Jing Tian a,⇑, David S. Zee a,b,c,d
a Department of Neurology, The Johns Hopkins University School of Medicine, Baltimore, MD 21287, USAb Department of Ophthalmology, The Johns Hopkins University School of Medicine, Baltimore, MD 21287, USAc Department of Otolaryngology-Head and Neck Surgery, The Johns Hopkins University School of Medicine, Baltimore, MD 21287, USAd Department of Neuroscience, The Johns Hopkins University School of Medicine, Baltimore, MD 21287, USA
a r t i c l e i n f o
Article history:Received 17 February 2010Received in revised form 27 July 2010
Keywords:SaccadeAdaptationContextMotor learningMonkey
0042-6989/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.visres.2010.09.014
⇑ Corresponding author. Address: Department of NHospital, 600 N. Wolfe Street, Pathology 2-210, Baltim614 1746.
E-mail address: [email protected] (J. Tian).
a b s t r a c t
When environmental or sensory conditions change suddenly, the brain must be capable of learning dif-ferent behavioral modes to produce accurate movements under multiple circumstances. A form of thisdual-state adaptation known as ‘‘context-specific adaptation” has been widely investigated using the sac-cade gain adaptation paradigm in humans. In this study, we asked whether or not context-specific adap-tation of saccade gain exists in monkeys and if so to explore its properties. Here, vertical eye position wasused as a context cue for adaptation of horizontal saccade gain. We asked for a gain-increase in one con-text and gain-decrease in another context, and then determined if a change in the context would invokeswitching between the adapted states. After training, our monkeys developed context-specific adapta-tion: in most cases gain-decrease adaptation could be induced, but there was little or no gain-increaseadaptation. This context-specific adaptation developed gradually and switching of gains was evidenton the first saccades with each change in context. Along with these results, the retention of an adaptationaftereffect overnight indicates that contextual-specific adaptation in monkeys is not a strategy, butinvolves a true adaptive process of reorganization in the brain. We suggest that context-specific adapta-tion in monkeys could be an important tool to provide insights into the mechanisms of saccade adapta-tion that occurs during the more natural circumstances of daily life.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Motor learning as it is usually studied in the laboratory occursin a constrained, artificial environment in which the stimulusand desired response are rigidly controlled. Indeed, such experi-ments allow one to hone in on fundamental, relatively low-levelmechanisms that underlie adaptive motor control. On the otherhand, the learning that occurs during the more natural circum-stances of daily life must be optimized for a complex sensory(and motor) environment in which the particular ‘contextual’ cir-cumstances in which the learning takes place dictate the necessarytype of response. In other words, because environmental or sen-sory conditions can change suddenly, the brain must be capableof learning different behavioral modes to produce accurate move-ments under multiple circumstances. For example, pointing move-ments can be adapted in a dual-state manner in the presence andabsence of a prism-induced perturbation (Mcgonigle & Flook,1978; Welch, Bridgeman, Anand, & Browman, 1993). This form of
ll rights reserved.
eurology, The Johns Hopkinsore, MD 21287. Fax: +1 410
dual-state adaptation is termed ‘‘context-specific adaptation”,and can be defined as the ability to learn and store simultaneouslytwo different adapted responses of a motor task, and to invokethe appropriate adapted states with the associate context cue(e.g., Shelhamer, Aboukhalil, & Clendaniel, 2005; Shelhamer &Clendaniel, 2002a). Once learned such contextual cues can beextremely powerful; witness the inappropriate postural responseand near fall that occurs when one takes a first step upon a brokenescalator (Bronstein, Bunday, & Reynolds, 2009; Reynolds &Bronstein, 2003).
Saccades are an ideal model to study motor adaptation becauseit is relatively easy both to artificially manipulate the stimuli thatdrive saccades, and to measure the consequent change in the mo-tor response (Hopp & Fuchs, 2004; Tian, Ethier, Shadmehr, Fujita, &Zee, 2009). Adaptation of saccade gain in both humans and mon-keys has been studied extensively with a conventional double-stepparadigm (McLaughlin, 1967). Adaptation is usually more rapidand robust for gain-decrease than increase and is slower in mon-keys than in humans (Deubel, Wolf, & Hauske, 1986; Straube,Fuchs, Usher, & Robinson, 1997). Much evidence indicates thatthe dorsal vermis (lobules VI–VII), the underlying posterior fasti-gial nuclei and perhaps the adjacent cerebellar hemispheres arecrucial areas for saccadic adaptation (Alahyane et al., 2008; Barashet al., 1999; Catz, Dicke, & Thier, 2008; Choi, Kim, Cho, & Kim, 2008;

2404 J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410
Optican & Robinson, 1980; Robinson, Fuchs, & Noto, 2002;Soetedjo, Kojima, & Fuchs, 2008; Takagi, Zee, & Tamargo, 1998).
In humans, it has been well established that saccades can bemade to have two different gains in response to the same targetdisplacement: which gain can depend on various contextual cues:e.g., target distance (Chaturvedi & vanGisbergen, 1997), horizontaland vertical eye position in the orbit (Alahyane & Pelisson, 2004;Shelhamer & Clendaniel, 2002a; Shelhamer et al., 2005), head ori-entation (Shelhamer & Clendaniel, 2002a, 2002b), and the visualproperties of the target (Herman, Harwood, & Wallman, 2009).However, little is known about the neural mechanisms responsiblefor context-dependent adaptation of saccade gain, and especiallyits anatomical substrate.
While much has been learned about saccade gain adaptationusing experimental studies in monkeys, and especially its neuro-physiological underpinnings, context-specific saccade adaptationof the type described above has not been studied in monkeys. Mon-keys have been shown to develop different gains depending uponsaccade amplitude (Watanabe, Noto, & Fuchs, 2000) and the degreeof disconjugate saccade adaptation that occurs to wearing a prismin front of one eye can be made to depend upon orbital position(Oohira & Zee, 1992). While such adaptive capabilities can beimportant, for example, in adaptation to a paresis of a single eyemuscle, it is not the ‘higher-level’ type of adaptation that we pro-pose to study here. Furthermore, context-specific adaptation inmonkeys could be an important tool to provide insights into howsaccade adaptation occurs in the more natural behaviors within acomplex sensory environment. Thus, we asked here whether ornot context-specific adaptation of saccade gain exists in monkeysand if so to explore some of its properties. In this study we usethe conventional saccade gain adaptation paradigm, in which hor-izontal saccades of equal amplitude are alternately subjected to dif-ferent adaptive demands (gain-increase and gain-decrease) in twodifferent vertical eye positions (context) during the same adapta-tion session.
2. Methods
2.1. General experimental procedures
Experiments were conducted on three male rhesus monkeys(M1, M2 and M3) weighing from 4 to 6 kg. All experimental andsurgical procedures, including anesthesia and post-operative anal-gesia, were performed according to a protocol that was approvedby the Institutional Animal Care and Use Committee of the JohnsHopkins University and following the NIH Guidelines. The animalswere first trained to come out of their cages and sit comfortably ina primate chair. For head immobilization during experiments, aplastic head plate was attached to the skull with dental acrylic.In single procedures for each eye, dual scleral coils made of stain-less steel wire were implanted binocularly in M1 and in the left eyein M2, as has been previously described (Tian, Zee, & Walker,2007). Dual resonant ‘‘leadless” coils, small multiple-turn loop offine wire connected to a capacitor (Roberts, Shelhamer, & Wong,2008), were implanted in the right eye in M2 and M3. Animalswere trained to fixate and follow a small visual target for a liquidreward.
Both M1 and M2 underwent a left trochlear nerve section andM2 had an ipsilateral trigeminal nerve (V1 and V2) section as wellas part of a different study. The lesion produced a vertical misalign-ment of the eyes: the paretic eye was higher than the normal eye,and this misalignment was greatest in down gaze when the pareticeye was adducted. Experiments reported here were conducted inM1 about 14 months after the superior oblique palsy surgery andin M2 both before and 5 months after the surgery. For the adapta-
tion data collected after the animal had the trochlear nerve section,the normal eye was always the viewing eye (the paretic eye waspatched) during the recording session and we show the data fromthe normal eye. The vertical misalignment was stable in both mon-keys (6.7 ± 0.3� in M1, 5.7 ± 0.3� in M2). For M2, there was no dif-ference in the performance of gain adaptation of horizontalsaccades before and after trochlear nerve section. Accordingly weare confident that the vertical strabismus produced by the troch-lear nerve section did not alter performance on the horizontal sac-cade adaptation paradigm. Thus all the results in a givenadaptation paradigm are pooled.
2.2. Eye movement measurements
During each experimental session, the animals were seated in aprimate chair with head fixed. Eye movements were measuredwith the magnetic field search coil method in M1 and in the lefteye of M2. The coil frame consisting of three orthogonal magneticfields was attached to the top of the primate chair, such that thehead was centered within it. Signals from each coil were demodu-lated by frequency detectors, sampled at 1000 Hz, and stored oncomputer for later analysis. For the right eye of M2 and M3, eyemovements were measured with a ‘‘leadless” search coil system(Roberts et al., 2008), which uses a resonant coil and no connectinglead wire. A transmitter emits a stream of pulses to the eye coil,and a receiver then detects the resonant oscillations emitted fromthe eye coil. The relative intensity of the signal as received by setsof orthogonal receiver coils determined the orientation of the eyecoil. The leadless coil system has a noise level of 0.05� peak to peak,and provides a sample rate up to 1000 Hz.
The coil signal was calibrated by requiring the animal to fix suc-cessively on targets at 5� intervals over a range of ±20� horizontaland vertical. Calibration data were obtained with one eye viewingand the other occluded. Visual targets were back-projected using amirror-controlled laser beam onto a translucent screen (72� � 72�)located 1 m in front of the animal. The room was dimly illuminated.
2.3. Experimental paradigms
A standard double-step paradigm was used to induce an increaseor a decrease in saccade amplitude (McLaughlin, 1967). As monkeysmade saccades in response to 15� horizontal target movements, wedetected each saccade as the eye crossed a virtual 2� window placedaround the starting position. The occurrence of a saccade triggered atarget step either 5� away from (gain-increase) or 5� closer to (gain-decrease), the starting position of the saccade. This created a visualerror at the end of the movement as if the saccade had been toosmall or too large, requiring the animal to make a corrective saccadeto fixate the target. This procedure, when repeated over severalhundred times, gradually increased or decreased the saccade ampli-tude. We adapted saccades to both leftward and rightward targetmovements. Adapting saccades in one direction does not influencesaccades in the opposite direction (Deubel et al., 1986; Miller,Anstis, & Templeton, 1981; Straube et al., 1997), so we treatedsaccades in the two directions independently.
2.3.1. Contextual adaptationIn this paradigm, adaptation to backward and forward target
steps were elicited concurrently and associated with different ver-tical eye positions. For the gain-increase trials, an initial target waspresented at left 10� (�10�). After the monkey fixated the target for500–1000 ms, the target jumped to +5�. During the saccade, thistarget was displaced to +10� and maintained for 500 ms. This loca-tion then became the fixation point for the next trial and the samesequence was presented in the opposite direction. For the gain-decrease trials, the initial fixation target was presented at �5�.

Fig. 1. An example of the time course of context-specific adaptation from M1. Theamplitude of the leftward saccade for gain-increase and gain-decrease trials isplotted as a function of saccade numbers. � Indicates the saccade amplitudes forgain-increase trials with eye up 10�, and s for gain-decrease trials with eyes down10�. Solid curves are exponential fits. Test trials before and after adaptation are setoff by dotted vertical lines.
J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410 2405
The first target step was to +10�, and the second target step was to+5�. This location then became the fixation point for the next trialin the opposite direction. Gain-increase adaptation was carried outwith the eyes directed up 10� above straight-ahead gaze, and gain-decrease adaptation with the eyes directed down 10�. These twocontext/adaptation conditions were alternately presented �30times each, with 30 adaptation trials in each block (�1800 trials to-tal). The intertrial time was 750 ms. Between each change in con-text/adaptation condition, the animal received 20 s of a rest toavoid fatigue. This protocol was also repeated with the conditionsreversed. A total of 23 experiments were performed with the threemonkeys.
2.3.2. Control adaptationTo determine the effectiveness of context-specific adaptation, a
control adaptation paradigm was performed in M3. A single type ofadaptation (gain-increase or gain-decrease) was elicited with theeyes either looking up 10� or looking down 10�. This experimentwas designed to look for generalization of adaptation in one con-text to the second context.
2.3.3. Test sessionsTest trials in each context state (vertical position) were per-
formed before (pretest, 15 trials in each direction) and after (post-test, 20 trials in each direction, tested twice) adaptation. Thesewere similar to the adaptation trials, except that there was no sec-ond target jump: the target did not reappear after the first targetjump. There was thus no visual feedback as to the accuracy ofthe saccade, providing an open-loop measure of saccade gain.The fixation target for the next trial appeared 1000 ms later.
2.4. Data analysis
Data were analyzed offline with custom software developed inMATLAB™ (The Mathworks, Natick, MA). An interactive programwas used to mark automatically the onset and the end of saccadesusing velocity criteria. The onset of the saccade was defined whenthe eye velocity first exceeded 30�/s and the end of the saccadewhen the eye velocity first dropped below 30�/s. The correctnessof these markings was verified by the experimenter. Targeting sac-cades elicited by an initial target step were selected for analysis.Trials were rejected if the latency of saccade was less than 60 ms,or if the trial was contaminated by blinks. These criteria excludedapproximately 5% of all saccades during contextual adaptation inM1, 7% in M3 and 11% in M2, who was a less motivated worker.The average latency over all the adaptation experiments was179 ± 23 ms in M1, 236 ± 29 ms in M2 and 244 ± 41 ms in M3.When we applied a latency criterion of 100 ms to assess the extentof predictive saccades, the number of excluded trials increased by4% in M1 and there was no difference in M2 and M3.
The horizontal amplitude of saccade was calculated as the dif-ference between initial and final eye positions. The percent ampli-tude change of the saccade between the posttest and the pretestsessions was calculated as follows: [(posttest mean ampli-tude � pretest mean amplitude)/pretest mean amplitude] � 100.To assess the time course of contextual adaptation, the first sac-cade upon each change in context was identified. This saccadegives an indication of whether or not gain switching indeed occursimmediately upon a change in context cue, before any adaptationtrials are performed. A linear regression line was fit separately tothe gain-increase and gain-decrease trials. The rate of adaptationwas estimated by the slope of regression line. We presented the re-sults of both rightward and leftward saccades in normal monkeys(M2 before the surgery and M3), but only leftward saccades in M1and M2 after the surgery, since there were more adaptation trialsavailable for analysis in the leftward direction than the rightward.
3. Results
3.1. General features
In general, we found some degree of context-specific adaptationwhen in one context the target steps backward during saccadesand in the other context the target steps forward. Fig. 1 shows anexample of the time course of adaptation from monkey 1 (M1),who was trained with gain-decrease adaptation with eyes downand gain-increase adaptation with eyes up. The amplitude of theleftward saccade for gain-increase and gain-decrease trials is plot-ted as a function of the saccade number. The saccades have an ini-tial amplitude of approximately 15�, the size of the first targetdisplacement. Little adaptation is evident during the first set ofgain-increase trials and the subsequent set of gain-decrease trials.As the session progresses (the two contexts alternate), gain-decrease adaptation becomes more clear, meanwhile gain-increaseadaptation still shows a decline but at a much slower rate. At theend of this process, the saccades in the gain-decrease trials havebecome significantly smaller; the saccades in the gain-increasetrials have also become smaller but by much less than the gain-decrease trials. Note as was typical for all monkeys that thesaccade amplitude within each individual context setting in theadaptation process was variable but, on average, it changed mono-tonically over the entire adaptation process in each context asshown by the exponential fit.
3.2. Aftereffect of adaptation
The aftereffect of adaptation was measured by comparing, be-fore and after training, the amplitude of saccades directed towardstargets that had been extinguished once the saccade began. Fig. 2summarizes the changes of saccade amplitude in all 23 experi-ments performed with the three monkeys. The aftereffect of train-ing was variable across experiments as seen in Fig. 2 (each linerepresents the results from different days). Significant changeswere found for gain-decrease adaptation in 17 experiments andfor gain-increase in 5 experiments. On average, mean saccadeamplitude was reduced by 4.3 ± 2.3% (means ± SD) in M1,5.9 ± 2.3% in M2 and 4.5 ± 1.5% in M3 after gain-decrease adapta-tion, which is consistent with the expected effect in the appropri-ate context (gaze directed down as well as up). However, theadaptation was less effective in the gain-increase context (gaze di-rected up as well as down except for gaze down in M3), as meansaccade amplitude slightly decreased by 0.8 ± 1.8% in M1, in-creased by 1.8 ± 3.5% in M2 and 3.0 ± 4.0% in M3. Nevertheless, in
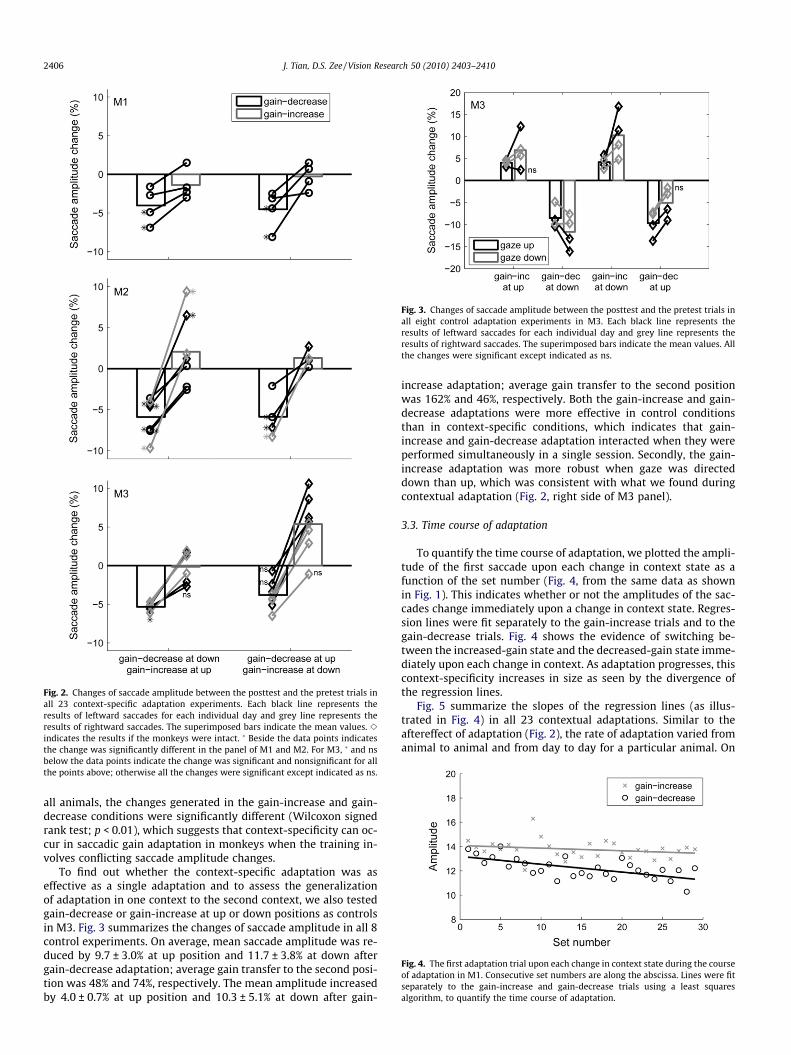
Fig. 2. Changes of saccade amplitude between the posttest and the pretest trials inall 23 context-specific adaptation experiments. Each black line represents theresults of leftward saccades for each individual day and grey line represents theresults of rightward saccades. The superimposed bars indicate the mean values. e
indicates the results if the monkeys were intact. � Beside the data points indicatesthe change was significantly different in the panel of M1 and M2. For M3, � and nsbelow the data points indicate the change was significant and nonsignificant for allthe points above; otherwise all the changes were significant except indicated as ns.
Fig. 3. Changes of saccade amplitude between the posttest and the pretest trials inall eight control adaptation experiments in M3. Each black line represents theresults of leftward saccades for each individual day and grey line represents theresults of rightward saccades. The superimposed bars indicate the mean values. Allthe changes were significant except indicated as ns.
Fig. 4. The first adaptation trial upon each change in context state during the courseof adaptation in M1. Consecutive set numbers are along the abscissa. Lines were fitseparately to the gain-increase and gain-decrease trials using a least squaresalgorithm, to quantify the time course of adaptation.
2406 J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410
all animals, the changes generated in the gain-increase and gain-decrease conditions were significantly different (Wilcoxon signedrank test; p < 0.01), which suggests that context-specificity can oc-cur in saccadic gain adaptation in monkeys when the training in-volves conflicting saccade amplitude changes.
To find out whether the context-specific adaptation was aseffective as a single adaptation and to assess the generalizationof adaptation in one context to the second context, we also testedgain-decrease or gain-increase at up or down positions as controlsin M3. Fig. 3 summarizes the changes of saccade amplitude in all 8control experiments. On average, mean saccade amplitude was re-duced by 9.7 ± 3.0% at up position and 11.7 ± 3.8% at down aftergain-decrease adaptation; average gain transfer to the second posi-tion was 48% and 74%, respectively. The mean amplitude increasedby 4.0 ± 0.7% at up position and 10.3 ± 5.1% at down after gain-
increase adaptation; average gain transfer to the second positionwas 162% and 46%, respectively. Both the gain-increase and gain-decrease adaptations were more effective in control conditionsthan in context-specific conditions, which indicates that gain-increase and gain-decrease adaptation interacted when they wereperformed simultaneously in a single session. Secondly, the gain-increase adaptation was more robust when gaze was directeddown than up, which was consistent with what we found duringcontextual adaptation (Fig. 2, right side of M3 panel).
3.3. Time course of adaptation
To quantify the time course of adaptation, we plotted the ampli-tude of the first saccade upon each change in context state as afunction of the set number (Fig. 4, from the same data as shownin Fig. 1). This indicates whether or not the amplitudes of the sac-cades change immediately upon a change in context state. Regres-sion lines were fit separately to the gain-increase trials and to thegain-decrease trials. Fig. 4 shows the evidence of switching be-tween the increased-gain state and the decreased-gain state imme-diately upon each change in context. As adaptation progresses, thiscontext-specificity increases in size as seen by the divergence ofthe regression lines.
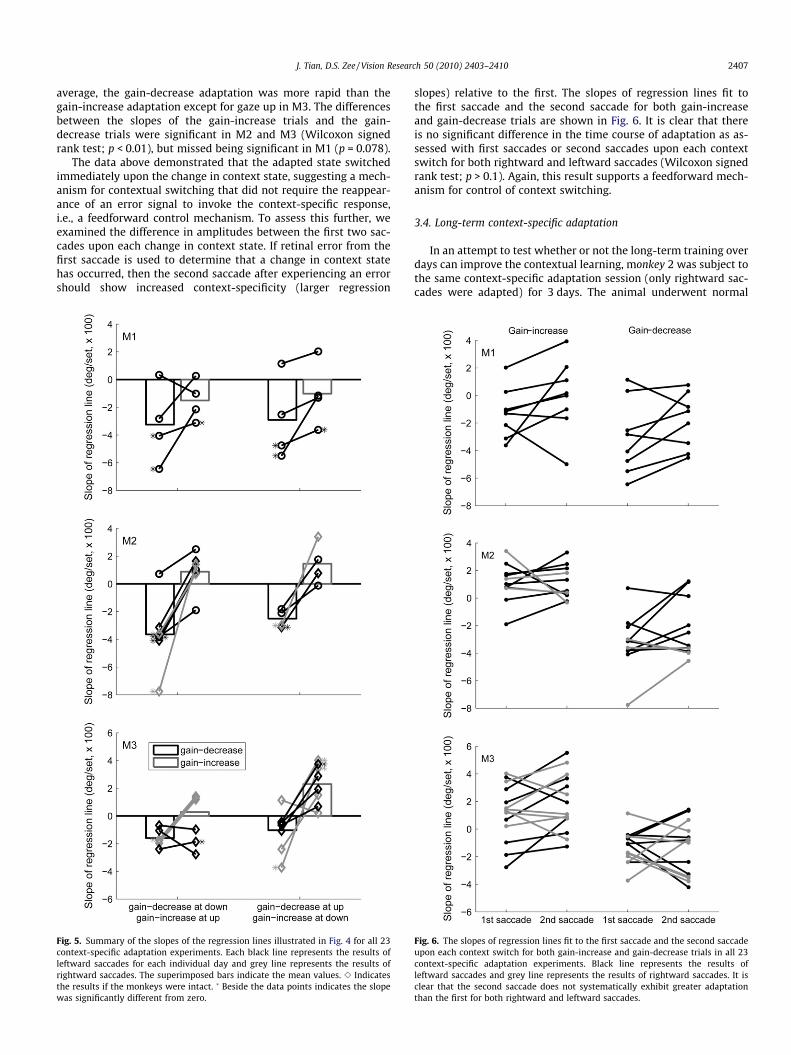
Fig. 5 summarize the slopes of the regression lines (as illus-trated in Fig. 4) in all 23 contextual adaptations. Similar to theaftereffect of adaptation (Fig. 2), the rate of adaptation varied fromanimal to animal and from day to day for a particular animal. On
J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410 2407
average, the gain-decrease adaptation was more rapid than thegain-increase adaptation except for gaze up in M3. The differencesbetween the slopes of the gain-increase trials and the gain-decrease trials were significant in M2 and M3 (Wilcoxon signedrank test; p < 0.01), but missed being significant in M1 (p = 0.078).
The data above demonstrated that the adapted state switchedimmediately upon the change in context state, suggesting a mech-anism for contextual switching that did not require the reappear-ance of an error signal to invoke the context-specific response,i.e., a feedforward control mechanism. To assess this further, weexamined the difference in amplitudes between the first two sac-cades upon each change in context state. If retinal error from thefirst saccade is used to determine that a change in context statehas occurred, then the second saccade after experiencing an errorshould show increased context-specificity (larger regression
Fig. 5. Summary of the slopes of the regression lines illustrated in Fig. 4 for all 23context-specific adaptation experiments. Each black line represents the results ofleftward saccades for each individual day and grey line represents the results ofrightward saccades. The superimposed bars indicate the mean values. e Indicatesthe results if the monkeys were intact. � Beside the data points indicates the slopewas significantly different from zero.
slopes) relative to the first. The slopes of regression lines fit tothe first saccade and the second saccade for both gain-increaseand gain-decrease trials are shown in Fig. 6. It is clear that thereis no significant difference in the time course of adaptation as as-sessed with first saccades or second saccades upon each contextswitch for both rightward and leftward saccades (Wilcoxon signedrank test; p > 0.1). Again, this result supports a feedforward mech-anism for control of context switching.
3.4. Long-term context-specific adaptation
In an attempt to test whether or not the long-term training overdays can improve the contextual learning, monkey 2 was subject tothe same context-specific adaptation session (only rightward sac-cades were adapted) for 3 days. The animal underwent normal
Fig. 6. The slopes of regression lines fit to the first saccade and the second saccadeupon each context switch for both gain-increase and gain-decrease trials in all 23context-specific adaptation experiments. Black line represents the results ofleftward saccades and grey line represents the results of rightward saccades. It isclear that the second saccade does not systematically exhibit greater adaptationthan the first for both rightward and leftward saccades.
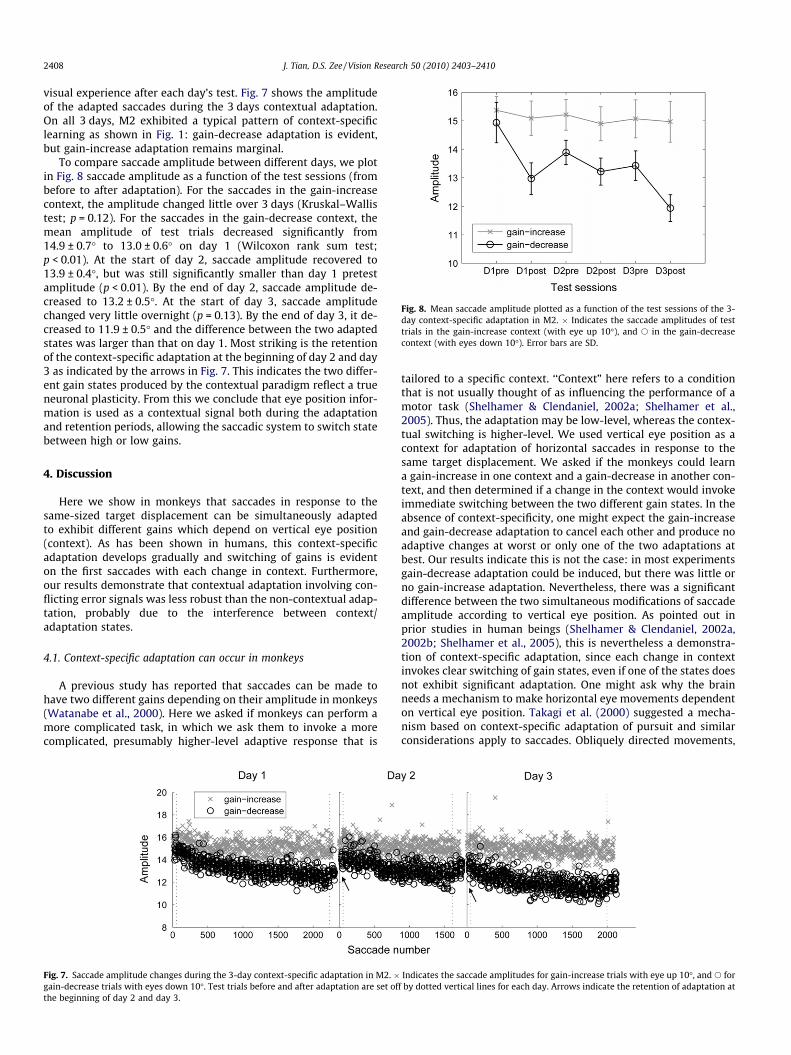
Fig. 8. Mean saccade amplitude plotted as a function of the test sessions of the 3-day context-specific adaptation in M2. � Indicates the saccade amplitudes of testtrials in the gain-increase context (with eye up 10�), and s in the gain-decreasecontext (with eyes down 10�). Error bars are SD.
2408 J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410
visual experience after each day’s test. Fig. 7 shows the amplitudeof the adapted saccades during the 3 days contextual adaptation.On all 3 days, M2 exhibited a typical pattern of context-specificlearning as shown in Fig. 1: gain-decrease adaptation is evident,but gain-increase adaptation remains marginal.
To compare saccade amplitude between different days, we plotin Fig. 8 saccade amplitude as a function of the test sessions (frombefore to after adaptation). For the saccades in the gain-increasecontext, the amplitude changed little over 3 days (Kruskal–Wallistest; p = 0.12). For the saccades in the gain-decrease context, themean amplitude of test trials decreased significantly from14.9 ± 0.7� to 13.0 ± 0.6� on day 1 (Wilcoxon rank sum test;p < 0.01). At the start of day 2, saccade amplitude recovered to13.9 ± 0.4�, but was still significantly smaller than day 1 pretestamplitude (p < 0.01). By the end of day 2, saccade amplitude de-creased to 13.2 ± 0.5�. At the start of day 3, saccade amplitudechanged very little overnight (p = 0.13). By the end of day 3, it de-creased to 11.9 ± 0.5� and the difference between the two adaptedstates was larger than that on day 1. Most striking is the retentionof the context-specific adaptation at the beginning of day 2 and day3 as indicated by the arrows in Fig. 7. This indicates the two differ-ent gain states produced by the contextual paradigm reflect a trueneuronal plasticity. From this we conclude that eye position infor-mation is used as a contextual signal both during the adaptationand retention periods, allowing the saccadic system to switch statebetween high or low gains.
4. Discussion
Here we show in monkeys that saccades in response to thesame-sized target displacement can be simultaneously adaptedto exhibit different gains which depend on vertical eye position(context). As has been shown in humans, this context-specificadaptation develops gradually and switching of gains is evidenton the first saccades with each change in context. Furthermore,our results demonstrate that contextual adaptation involving con-flicting error signals was less robust than the non-contextual adap-tation, probably due to the interference between context/adaptation states.
4.1. Context-specific adaptation can occur in monkeys
A previous study has reported that saccades can be made tohave two different gains depending on their amplitude in monkeys(Watanabe et al., 2000). Here we asked if monkeys can perform amore complicated task, in which we ask them to invoke a morecomplicated, presumably higher-level adaptive response that is
Fig. 7. Saccade amplitude changes during the 3-day context-specific adaptation in M2. �gain-decrease trials with eyes down 10�. Test trials before and after adaptation are set ofthe beginning of day 2 and day 3.
tailored to a specific context. ‘‘Context” here refers to a conditionthat is not usually thought of as influencing the performance of amotor task (Shelhamer & Clendaniel, 2002a; Shelhamer et al.,2005). Thus, the adaptation may be low-level, whereas the contex-tual switching is higher-level. We used vertical eye position as acontext for adaptation of horizontal saccades in response to thesame target displacement. We asked if the monkeys could learna gain-increase in one context and a gain-decrease in another con-text, and then determined if a change in the context would invokeimmediate switching between the two different gain states. In theabsence of context-specificity, one might expect the gain-increaseand gain-decrease adaptation to cancel each other and produce noadaptive changes at worst or only one of the two adaptations atbest. Our results indicate this is not the case: in most experimentsgain-decrease adaptation could be induced, but there was little orno gain-increase adaptation. Nevertheless, there was a significantdifference between the two simultaneous modifications of saccadeamplitude according to vertical eye position. As pointed out inprior studies in human beings (Shelhamer & Clendaniel, 2002a,2002b; Shelhamer et al., 2005), this is nevertheless a demonstra-tion of context-specific adaptation, since each change in contextinvokes clear switching of gain states, even if one of the states doesnot exhibit significant adaptation. One might ask why the brainneeds a mechanism to make horizontal eye movements dependenton vertical eye position. Takagi et al. (2000) suggested a mecha-nism based on context-specific adaptation of pursuit and similarconsiderations apply to saccades. Obliquely directed movements,
Indicates the saccade amplitudes for gain-increase trials with eye up 10�, and s forf by dotted vertical lines for each day. Arrows indicate the retention of adaptation at

J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410 2409
which occur often naturally, demand different horizontal innerva-tions depending on the vertical position of the eye because of thechange in pulling directions of the horizontal muscles with verticaleye position. Accordingly vertical eye position could be an effectivecontextual cue for the appropriate horizontal innervations neededto make an accurate saccade.
One can ask if the vertical strabismus produced by the trochlearnerve section (part of a different study) affected the contextualadaptation in our monkeys, since two of the three contextual adap-tation experiments performed before M2 had the nerve sectionappeared to have a greater aftereffect than those after, and theadaptation in M1 was even weaker. Taking the results togetherin all three monkeys, however, we think it unlikely that the strabis-mus had a major effect on the contextual adaptation in the twooperated monkeys. The variability of adaptation itself, from animalto animal and from day to day for a specific animal makes it diffi-cult to make a firm conclusion about the strabismus. Furthermore,the amount of aftereffect after conventional adaptation was similarbefore and after the nerve section in M2 (data not shown). Accord-ingly we are confident that the vertical strabismus did not alter theperformance of the horizontal saccade adaptation per se and, espe-cially considering that the monkey was always tested with only theintact eye viewing.
4.2. Comparison to studies with humans
In humans, two previous studies have investigated the context-specificity of saccade adaptation using vertical eye position as acontext (Alahyane & Pelisson, 2004; Shelhamer & Clendaniel,2002a). Our findings in monkeys are most similar to those ofShelhamer and Clendaniel (2002a), who found there was consider-able undesired transfer from gain-decrease adaptation to gain-increaseadaptation, with the result that there was little gain-increase adap-tation. However, Alahyane and Pelisson (2004) reported that sig-nificant adaptation can be induced not only in the gain-decreasecontext but also in the gain-increase context. One possible reasonfor this difference, as has been suggested (Alahyane & Pelisson,2004), is that the two vertical eye positions where the conflictingadaptations were tested were farther away from each other in theirstudy (up 12.5� and down 25�) than in ours and Shelhamer andClendaniel’s experiments (up 10� and down 10�). The closer thetwo contexts are, the less effective it might be to prevent transferof adaptation from gain-decrease condition to the gain-increasecondition. A similar spatial extent effect is seen, for example, indisconjugate saccade adaptation in monkeys in response to wear-ing a laterally-displacing hemifield prism in front of one eye(Oohira & Zee, 1992).
The efficacy of different context cues (sensory and motor) hasbeen assessed in prior studies (Shelhamer & Clendaniel, 2002a,2002b). Eye position in the orbit is an example of a motor contextcue: the motor commands sent to the muscles to generate a partic-ular movement are different in the two context states. Head roll-tilt is an example of a sensory cue as head tilt is sensed by the oto-liths and proprioception but saccades are always same in the head.It has been demonstrated that the motor context (change in sac-cade direction) is relatively stronger than the sensory context(head roll-tilt) (Shelhamer & Clendaniel, 2002b), and horizontaleye position was more effective than vertical eye position for dem-onstrating context-specific adaptation of horizontal saccades(Shelhamer & Clendaniel, 2002a). Taken together, these resultssuggest that a context cue is more effective when it is immediatelyrelevant to the movement being adapted since naturally occurringadaptation is more commonly a motor repair than a sensoryremapping mechanism. However, this notion has been challengedby a recent study of Herman et al. (2009), who reported that avisual context (stable/flicking target) can serve as an effective
contextual cue for saccade adaptation, although an earlier attemptwith target color had been unsuccessful (Deubel, 1995). It may bethat the ability to modify saccade gains based on the target visualproperties reflects a much higher-level and more sophisticatedcognitive type of adaptation.
Another difference among these studies is the trial structure ofcontextual paradigm: two different adaptations in two contextshas been alternated randomly across successive trials (Alahyane& Pelisson, 2004; Deubel, 1995), in blocks of fixed length(Shelhamer & Clendaniel, 2002a, 2002b), or short blocks of irregu-lar length (Herman et al., 2009). Although random context presen-tation may slow contextual learning since half of the adaptationtrials would be preceded by trials with different or oppositeerror signals, one study has shown robust adaptation in humans(Alahyane & Pelisson, 2004). In monkeys, we chose to alternatethe gain-increase and gain-decrease adaptations in blocks (30trials), because adaptation in monkeys is much slower than inhumans and also in this way undesired saccades when switchingthe contexts (between two vertical eye positions) can be avoided.With this protocol, our monkeys exhibit context-dependent learn-ing though it is not complete. Further study will be required todetermine how the trial structure affects the extent and timecourse of context-specific adaptation.
4.3. Context-specific adaptation process is a true neuronal plasticity
One might question whether our monkeys used a cognitivestrategy to alter their saccades in different contexts since the targetlocations were predictable. Several lines of evidence argue againstthe idea that their contextual adaptation was strategic. First, theadaptation developed gradually over thousands of trials and gener-ally gain-decrease adaptation was more developed than gain-increase adaptation. This is also the case with conventional saccadeadaptation and supports the idea that our data reflect true adapta-tion, not a cognitive strategy. Second, as adaptation proceeds,switching of adapted gains upon each change in context occurredimmediately. Thus, an initial saccade was not necessary to provideretinal error feedback in order to trigger saccades of appropriategain. Third, contextual adaptation was weakened by the interfer-ence between the gain-increase and gain-decrease adaptations,suggesting that volitional control of this process is unlikely. Finally,there was some residual amplitude change the day after an exper-iment, despite the fact that monkey had normal visual experiencein between. If contextual adaptation were strategic, we would notexpect the strong retention after the animal had spent 24 h in theirnatural visual environment. Similar retention of gain change in-duced by conventional saccade adaptation has also been found inhumans (Alahyane & Pelisson, 2005) and in monkeys (Noto,Watanabe, & Fuchs, 1999). It may appear surprising that the adap-tation can be retained over days, despite the subjects having re-ceived visual feedback from many saccades made during theirdaily activities. One possible explanation is that the amplitude ofsaccades generated naturally is different from that of adapted sac-cades during the experiments. It has been reported that most nat-urally occurring saccades in humans are less than 15� (Bahill,Adler, & Stark, 1975). Another possible factor would be that inter-nally triggered saccades have little effect on de-adaptation of visu-ally triggered saccades performed in the laboratory. Takentogether, these results suggest that contextual-specific adaptationcannot be controlled voluntarily, but involves a true adaptive pro-cess of reorganization in the brain.
4.4. Neuronal substrate of context-specific adaptation
The neuronal substrates of conventional (non-contextual) sacc-adic adaptation have been extensively investigated. A variety of

2410 J. Tian, D.S. Zee / Vision Research 50 (2010) 2403–2410
neurophysiological studies demonstrated that the cerebellum iscritically involved in saccadic adaptation (Barash et al., 1999; Catzet al., 2008; Optican & Robinson, 1980; Robinson et al., 2002;Soetedjo et al., 2008; Takagi et al., 1998). However, much less isknown about the neuronal site of context-specific adaptation. Ina monkey reaching study with prism adaptation, it has been shownthat pointing responses can be differentially adapted for eachviewing eye. This capability was lost after focal lesions of the dorsalvermal and paravermal cerebellar cortex, but in the same animalsnon-contextual adaptation to a prism was spared, suggesting thatthese regions of cerebellum are required in context-dependent mo-tor learning (Lewis & Tamargo, 2001). Based on these results, thecerebellum is a possible candidate area for context-specific saccad-ic adaptation.
Recently, Lee and Schweighofer (2009) proposed that a parallelmodel with a fast and a slow process can explain the dual or multi-ple motor adaptations. In their model, the fast process containsonly a single state and the slow process contains multiple statesswitched via context cues. In the case of conventional saccadicadaptation, a similar two-timescale process has been observed ina gain adaptation paradigm (Ethier, Zee, & Shadmehr, 2008), andin a cross-axis adaptation paradigm (Chen-Harris, Joiner, Ethier,Zee, & Shadmehr, 2008). In human patients with cerebellar degen-eration including the vermis and hemispheres, a recent study dem-onstrated that the fast process of adaptation was absent, but theslow process was less impaired (Xu-Wilson, Chen-Harris, Zee, &Shadmehr, 2009). This suggest that the damage to the cerebellarcortex mostly affects the fast process of adaptation, and supportsthe idea that the two processes derive from different neural mech-anisms and possibly have different anatomical substrates. Simi-larly, in a monkey lesion study, Barash et al. (1999) proposedthat a fast and a slow process that adjust the saccade gain dependon the cerebellar cortex and nuclei separately. Context-specificadaptation involves recruitment of different adaptation modulesfor different contexts and increases the complexity of its underly-ing neural circuitry. The precise role played by separate cerebellarloci, or other parts of the brain, in contextual adaptation, especiallyin learning, storage and retrieval of different adapted states, needto be elucidated. To this important and topical question abouthow the brain learns new behaviors, the monkey model of contex-tually driven saccade adaptation can provide critical information.
Acknowledgments
This study was supported by the National Institutes of Health(R01-EY01849); the Albert Pennick fund; Leon Levy Foundation;and by the Arnold-Chiari Foundation. Dr. Tian is a Paul and BettyCinquegrana Scholar. C. Bridges, A. Lasker, and D. Roberts providedtechnical support. The authors are grateful to M. Shelhamer for acritical review and discussion of this manuscript.
References
Alahyane, N., Fonteille, V., Urquizar, C., Salemme, R., Nighoghossian, N., Pelisson, D.,et al. (2008). Separate neural substrates in the human cerebellum for sensory-motor adaptation of reactive and of scanning voluntary saccades. Cerebellum, 7,595–601.
Alahyane, N., & Pelisson, D. (2004). Eye position specificity of saccadic adaptation.Investigative Ophthalmology & Visual Science, 45, 123–130.
Alahyane, N., & Pelisson, D. (2005). Long-lasting modifications of saccadic eyemovements following adaptation induced in the double-step target paradigm.Learning Memory, 12, 433–443.
Bahill, A. T., Adler, D., & Stark, L. (1975). Most naturally occurring human saccadeshave magnitudes of 15 degrees or less. Investigative Ophthalmology, 14,468–469.
Barash, S., Melikyan, A., Sivakov, A., Zhang, M., Glickstein, M., & Thier, P. (1999).Saccadic dysmetria and adaptation after lesions of the cerebellar cortex. Journalof Neuroscience, 19, 10931–10939.
Bronstein, A. M., Bunday, K. L., & Reynolds, R. (2009). What the ‘‘broken escalator”phenomenon teaches us about balance. Basic and Clinical Aspects of Vertigo andDizziness, 1164, 82–88.
Catz, N., Dicke, P. W., & Thier, P. (2008). Cerebellar-dependent motor learning isbased on pruning a Purkinje cell population response. Proceedings of the NationalAcademy of Sciences USA, 105, 7309–7314.
Chaturvedi, V., & vanGisbergen, J. A. M. (1997). Specificity of saccadic adaptation inthree-dimensional space. Vision Research, 37, 1367–1382.
Chen-Harris, H., Joiner, W. M., Ethier, V., Zee, D. S., & Shadmehr, R. (2008). Adaptivecontrol of saccades via internal feedback. Journal of Neuroscience, 28,2804–2813.
Choi, K. D., Kim, H. J., Cho, B. M., & Kim, J. S. (2008). Saccadic adaptation in lateralmedullary and cerebellar infarction. Experimental Brain Research, 188, 475–482.
Deubel, H. (1995). Is saccade adaptation context-specific? In J. M. Findlay, R. Walker,& R. W. Kentridge (Eds.), Eye movement research: Mechanisms, processes andapplications (pp. 177–187). Amsterdam: Elsevier Science.
Deubel, H., Wolf, W., & Hauske, G. (1986). Adaptive gain control of saccadic eyemovements. Human Neurobiology, 5, 245–253.
Ethier, V., Zee, D. S., & Shadmehr, R. (2008). Spontaneous recovery of motor memoryduring saccade adaptation. Journal of Neurophysiology, 99, 2577–2583.
Herman, J. P., Harwood, M. R., & Wallman, J. (2009). Saccade adaptation specific tovisual context. Journal of Neurophysiology, 101, 1713–1721.
Hopp, J., & Fuchs, A. E. (2004). The characteristics and neuronal substrate of saccadiceye movement plasticity. Progress in Neurobiology, 72, 27–53.
Lee, J. Y., & Schweighofer, N. (2009). Dual adaptation supports a parallel architectureof motor memory. Journal of Neuroscience, 29, 10396–10404.
Lewis, R. F., & Tamargo, R. J. (2001). Cerebellar lesions impair context-dependentadaptation of reaching movements in primates. Experimental Brain Research,138, 263–267.
Mcgonigle, B. O., & Flook, J. (1978). Long-term retention of single and multistateprismatic adaptation by humans. Nature, 272, 364–366.
McLaughlin, S. (1967). Parametric adjustment in saccadic eye movements.Perception & Psychophysics, 2, 359–362.
Miller, J. M., Anstis, T., & Templeton, W. B. (1981). Saccadic plasticity: Parametricadaptive control by retinal feedback. Journal of Experimental Psychology: HumanPerception and Performance, 7, 356–366.
Noto, C. T., Watanabe, S., & Fuchs, A. F. (1999). Characteristics of simian adaptationfields produced by behavioral changes in saccade size and direction. Journal ofNeurophysiology, 81, 2798–2813.
Oohira, A., & Zee, D. S. (1992). Disconjugate ocular motor adaptation in rhesusmonkey. Vision Research, 32, 489–497.
Optican, L. M., & Robinson, D. A. (1980). Cerebellar-dependent adaptive control ofprimate saccadic system. Journal of Neurophysiology, 44, 1058–1076.
Reynolds, R. F., & Bronstein, A. M. (2003). The broken escalator phenomenon –Aftereffect of walking onto a moving platform. Experimental Brain Research, 151,301–308.
Roberts, D. C., Shelhamer, M., & Wong, A. (2008). A new ‘‘wireless” search-coilsystem. In Proceedings of the 2008 symposium on eye tracking research &applications, pp. 197–204.
Robinson, F. R., Fuchs, A. F., & Noto, C. T. (2002). Cerebellar influences on saccadeplasticity. Annals of the New York Academy of Sciences, 956, 155–163.
Shelhamer, M., Aboukhalil, A., & Clendaniel, R. (2005). Context-specific adaptationof saccade gain is enhanced with rest intervals between changes in contextstate. Annals of the New York Academy of Sciences, 1039, 166–175.
Shelhamer, M., & Clendaniel, R. A. (2002a). Context-specific adaptation of saccadegain. Experimental Brain Research, 146, 441–450.
Shelhamer, M., & Clendaniel, R. (2002b). Sensory, motor, and combined contexts forcontext-specific adaptation of saccade gain in humans. Neuroscience Letters, 332,200–204.
Soetedjo, R., Kojima, Y., & Fuchs, A. F. (2008). Complex spike activity in theoculomotor vermis of the cerebellum: A vectorial error signal for saccade motorlearning? Journal of Neurophysiology, 100, 1949–1966.
Straube, A., Fuchs, A. F., Usher, S., & Robinson, F. R. (1997). Characteristics ofsaccadic gain adaptation in rhesus macaques. Journal of Neurophysiology, 77,874–895.
Takagi, M., Abe, H., Hasegawa, T., Usui, T., Hasebe, H., Miki, A., et al. (2000). Context-specific adaptation of pursuit initiation in humans. Investigative Ophthalmology& Visual Science, 41, 3763–3769.
Takagi, M., Zee, D. S., & Tamargo, R. (1998). Effects of lesions of the oculomotorvermis on eye movements in primate: Saccades. Journal of Neurophysiology, 80,1911–1930.
Tian, J., Ethier, V., Shadmehr, R., Fujita, M., & Zee, D. S. (2009). Some perspectives onsaccade adaptation. Annals of the New York Academy of Sciences, 1164, 166–172.
Tian, J., Zee, D. S., & Walker, M. F. (2007). Rotational and translational optokineticnystagmus have different kinematics. Vision Research, 47, 1003–1010.
Watanabe, S., Noto, C. T., & Fuchs, A. F. (2000). Flexibility of saccade adaptation inthe monkey: Different gain states for saccades in the same direction.Experimental Brain Research, 130, 169–176.
Welch, R. B., Bridgeman, B., Anand, S., & Browman, K. E. (1993). Alternating prismexposure causes dual adaptation and generalization to a novel displacement.Perception & Psychophysics, 54, 195–204.
Xu-Wilson, M., Chen-Harris, H., Zee, D. S., & Shadmehr, R. (2009). Cerebellarcontributions to adaptive control of saccades in humans. Journal of Neuroscience,29, 12930–12939.