conserved role for autophagy in rho1-mediated cortical...
TRANSCRIPT

Conserved role for autophagy in Rho1-mediatedcortical remodeling and blood cell recruitmentPavan Kadandalea, Joshua D. Stenderb, Christopher K. Glassb, and Amy A. Kigera,1
aSection of Cell & Developmental Biology and bDepartments of Cellular and Molecular Medicine and Medicine, University of California, San Diego, CA 92093
Edited by Kathryn V. Anderson, Sloan-Kettering Institute, New York, NY, and approved May 3, 2010 (received for review December 15, 2009)
Dynamic regulation of cell shape underlies many developmentaland immune functions. Cortical remodeling is achieved under thecentral control of Rho GTPase pathways that modulate anexquisite balance in the dynamic assembly and disassembly ofthe cytoskeleton and focal adhesions. Macroautophagy (autoph-agy), associated with bulk cytoplasmic remodeling through lyso-somal degradation, has clearly defined roles in cell survival anddeath. Moreover, it is becoming apparent that proteins, organ-elles, and pathogens can be targeted for autophagic clearanceby selective mechanisms, although the extent and roles of suchdegradation are unclear. Here we report a conserved role forautophagy specifically in the cortical remodeling of Drosophilablood cells (hemocytes) and mouse macrophages. Continuousautophagy was required for integrin-mediated hemocyte spread-ing and Rho1-induced cell protrusions. Consequently, hemocytesdisrupted for autophagy were impaired in their recruitment toepidermal wounds. Cell spreading required ref(2)P, the Drosophilap62 multiadaptor, implicating selective autophagy as a novelmechanism for modulating cortical dynamics. These results illumi-nate a specific and conserved role for autophagy as a regulatorymechanism for cortical remodeling, with implications for immunecell function.
selective autophagy | cell spreading | Drosophila | hemocyte | macrophage
Blood cells are subject to high demands for environmentallyresponsive cell shape changes for both developmental and
immune functions that involve cell spreading, migration, andphagocytosis. Drosophila blood cells, called hemocytes, are anevolutionary equivalent of mammalian macrophages in terms ofontogeny, gene expression, and immune roles in pathogen clear-ance. Hemocytes that arise in the embryo persist into larval stagesin free circulation with known surveillance functions and havebeen studied for in vivo requirements for cell morphology andmigration (1–3).A theme in the remodeling of cell shape involves the re-
organization of cortical cytoskeleton and membrane. Members ofthe Rho GTPase family control the organization and stability offilamentous actin (F-actin) in cell protrusion and membrane dy-namics (4). Cell protrusion formation and spreading also are asso-ciated with integrin–matrix interactions for surface attachmentand focal adhesion function (5). Importantly, both of these exam-ples require multipronged, dynamically regulated cellular pathwaysfor proper cortical remodeling; for instance, Rho GTPase pathwaycomponents are subject to regulation via mechanisms that changeprotein activity throughmembrane trafficking (6), protein complexformation (7), ubiquitination-mediated protein stability (8), or re-versible protein phosphorylation (9).The process of autophagy, or “self-eating” through cytoplas-
mic turnover in the lysosome, is a mechanism of cellular recy-cling and remodeling involved in development, homeostasis, anddisease. Autophagy is best understood as a nonselective responseto starvation in which the recycling of bulk cytoplasm serves asa pro-survival or pro-death mechanism (10). However, recentwork indicates that specific protein aggregates, organelles, andpathogens can be selectively targeted for autophagic degrada-tion. The p62 multiadaptor binds both to ubiquitinated proteins
and to Atg8/LC3, a protein central to autophagosome formation,and thus is proposed to act as a receptor for selective autophagicclearance (11–13). Consistent with this, a loss of function of ref(2)P,the Drosophila homolog of p62, was found to result in accumula-tion of ubiquitinated proteins and disruption of neuronal function(14). However, the extent of roles and identity of targets for se-lective autophagy are largely unknown.We identified Atg1 in an RNAi screen for kinase functions re-
quired for cell shape change in a Drosophila hemocyte-derived cellline. Previous studies independently discovered the Atg1 Ser/Thrkinase with conserved roles in axonal outgrowth (15–17) and as anessential component for autophagosome formation (18, 19). Herewe explore the apparent dual role for Atg1 in the regulation ofmorphogenesis and autophagy, and evaluate the significance ofautophagy in hemocyte cortical behavior and in vivo functions. Weshow that basal autophagy is not essential for larval hemocytesurvival, but is required to promote hemocyte cell spreading andextension of Rho1-induced protrusions. Importantly, autophagy isrequired for blood cell recruitment to larval wound sites and forcell spreading of mouse macrophages, suggesting conserved rolesfor autophagy in controlling blood cell shape and function. Ourwork points to p62-selective autophagy as an additionalmechanismregulating cortical dynamics.
ResultsAtg1 Is Required Cell-Autonomously for Hemocyte Cell Spreading.Hemocytes dissected out of Drosophila larvae displayed a cell-spreading response visualized by GFP in live cells (Fig. 1 A andE; 93% cell spread). Starting as round cells, hemocytes flat-tened over 30 min, while extending spiky radial protrusionscontaining both bundled F-actin and microtubules (Fig. 1B). Incontrast, hemocytes from Atg1Δ3D mutant larvae (18) remainedround (Fig. 1C and E; 10% cell spread) and appeared to lack F-actin protrusions (Fig. 1D). To further test the requirement forAtg1 function in hemocytes, we used the Pxn-GAL4 (hemo-cytes) or Cg-GAL4 drivers (hemocytes and fat body). We foundthat hemocyte-directed expression of a wild-type Atg1 cDNAcould rescue the effects of the Atg1Δ3D mutation on cellspreading (Fig. S1A), whereas the defect in cell spreading wassimilarly observed with hemocyte-targeted Atg1 RNAi (20)(Fig. S1 B and C). Taken together, these findings indicatea specific, cell-autonomous requirement for Atg1 function forhemocyte spreading.
Autophagy Is Required for Initiation and Maintenance of CellSpreading. If the role for Atg1 in cell spreading reflects a rolefor autophagy in cellular morphogenesis, then blocking autoph-
Author contributions: P.K. and A.A.K. designed research; P.K. performed research; J.D.S.and C.K.G. contributed new reagents/analytic tools; P.K. analyzed data; and A.A.K. andP.K. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.0914168107/-/DCSupplemental.
10502–10507 | PNAS | June 8, 2010 | vol. 107 | no. 23 www.pnas.org/cgi/doi/10.1073/pnas.0914168107

agy in hemocytes by other means also should result in round cellsthat fail to spread. To test this, we used 3-methyladenine (3MA),an inhibitor of autophagy (21). Wild-type hemocytes treated with3MA did not spread (2% cell spread), similar to Atg1 mutanthemocytes (Figs. 1E and 2A). In addition, hemocyte-targetedRNAi (22) depletion of other known autophagy-related genes(Atg4, Atg6, Atg7, Atg8a and Atg9) that function directly inautophagosome formation (23) abolished the ability of hemo-cytes to spread and extend F-actin protrusions (5–8% cellspread; Fig. 1E and Fig. S1 D and E). Thus, the Atg1 functionrequired for hemocyte spreading reflects a general requirementfor autophagy in cortical remodeling.We used 3MA to temporally distinguish whether autophagy is
required for the initiation or maintenance of hemocyte spread-ing. wild-type hemocytes remained spread when washed withPBS (Fig. 2C), but failed to remain spread once treated with3MA, instead displaying retracted cell protrusions and a morerounded shape (Fig. 2B′ and C; 12% cells spread). The auto-phagy dependence was reversible; with hemocytes spreading af-ter 3MA was removed (Fig. 2B′′; 93% cell spread). These find-ings indicate that continuous autophagy is required for initiationand maintenance of hemocyte spreading.
Autophagy Is Blocked in Atg1Δ3D Hemocytes. If autophagy is re-quired for hemocyte spreading, then we would expect autopha-gosomes to be normally present in wild-type but absent inAtg1Δ3D mutant hemocytes. Using transmission electron mi-croscopy (TEM), we identified double-membrane–bound struc-tures typical of autophagosomes in wild-type cells (1.8 per cell;Fig. 3A and Fig. S2 A and B). In contrast, the Atg1Δ3D mutanthemocytes were mostly devoid of double-membrane–boundstructures (0.5/cell; Fig. 3A and Fig. S2A). Conversely, blockageof autophagy is predicted to result in an accumulation of proteinsnormally targeted for autophagic degradation. Atg1Δ3D hemo-cytes exhibited an increased number of puncta containing Ref(2)
P (5.6 vs. 2.4 puncta/cell) (14, 24), the Drosophila homolog of p62(11–13) (Fig. 3 B and C). The lack of double-membrane struc-tures and the accumulation of Ref(2)P demonstrate that Atg1Δ3D
disrupts autophagosome formation and autophagic clearancein hemocytes.
Disruption of Autophagy Has No Effect on Larval Hemocyte Number,Adhesion, or Survival. Cell morphology is responsive to changes incell homeostasis, such as cell survival, proliferation, and adhe-sion. There was no significant difference in the total cell counts,
Fig. 1. Autophagy is required for hemocyte spreading. (A) Control hemo-cytes spread with radial protrusions; GFP. (B) Protrusions rich in F-actin andmicrotubules. (C and D) Atg1Δ3D/ Atg1Δ3D hemocytes did not spread or ex-tend protrusions. (A and C) Cg-GAL4 UAS-GFP. (B and D) F-actin (magenta)and microtubules (green). (E) Percentage of round or spread hemocytes; Cg-GAL4 for RNAi. (Scale bar: 5 μm.)
Fig. 2. Autophagy is required for initiation and maintenance of cell spread-ing. (A) Hemocytes did not spread when blocked for autophagy with additionof 3MA for 1 h; Cg-GAL4 UAS-GFP. (B–B′′) Example of a single hemocyte im-aged over time; Pxn-GAL4 UAS-GFP. Hemocyte spread in PBS (B), retractedprotrusions and rounded after 30 min +3MA (B′), and respread 15 min afterwashout (B′′). (C) All hemocytes were identified as round or spread (mean ±SEM), following 30 min in PBS or +3MA and a 30-min wash with replacementmedium, as shown. (Scale bar: 5 μm.)
Fig. 3. Autophagy is blocked in Atg1Δ3D mutant hemocytes. Autophago-some formation and autophagy function are disrupted in Atg1Δ3D mutanthemocytes. (A) Number of double membrane-bound structures per cell inTEM micrographs, 12 hemocytes each condition (bar, mean ± SEM). (B)Number of Ref(2)P puncta per hemocyte (mean ± SEM). (C) Accumulation ofRef(2)P (green, arrowheads), indicating blockage of autophagic clearance inAtg1Δ3D mutant hemocytes. F-actin is shown in magenta. (Scale bar: 5 μm.)
Kadandale et al. PNAS | June 8, 2010 | vol. 107 | no. 23 | 10503
CELL
BIOLO
GY

suggesting homeostasis of cell proliferation and survival ofAtg1Δ3D mutant hemocytes. Normal cell numbers were present inthe two distinct hemocyte populations in the circulating hemo-lymph, which is easily spilled on dissection (Fig. S2C), and theadherent sessile hemocytes attached to the body wall (Fig. S2D)(25), also indicating that cell adhesion was unaffected. We foundno difference in the percentage of dead or dying cells detected bypropidium iodide staining (Fig. S2E).Autophagy provides a means for cells to generate energy when
nutrient-deprived, raising the possibility that failure of cellspreading might be due to decreased amino acids or ATP pro-duction from blockage of autophagy. We found the same resultsfor control and mutant hemocytes regardless of whether assayswere done in buffer or serum-containing medium. The additionof methyl pyruvate, which can be directly incorporated into theKrebs cycle and has been used to overcome the energy-depletingeffects of autophagy inhibition (26), did not restore the hemocytespreading of Atg1 mutants (Fig. S2F). These findings suggest thatindirect effects of cell homeostasis or bioenergetics are not re-sponsible for the autophagy-dependent role in cell spreading,further implicating a specific role in cortical remodeling.
Autophagy Is Required for Protrusion Extension but Not CorticalDynamics. Cell spreading involves coordinated events in cyto-skeletal remodeling for the removal of inhibitory cortical tensionand generation of dynamic protrusive forces. A cortical ring ofF-actin was present in the autophagy-inhibited hemocytes, sim-ilar to unspread wild-type hemocytes immediately after dissec-tion. To address dynamics at the cell surface, we performed time-lapse microscopy imaging of GFP-labeled hemocytes. We foundthat the hemocyte cortex was dynamic with membrane rufflingunder both control and Atg1Δ3D mutant conditions (Fig. 4 andMovie S1), suggesting that autophagy is not essential for totalcytoskeletal dynamics or initiation of protrusions. Although theAtg1Δ3D mutant hemocytes could extend and retract short pro-trusions, the cells did not extend the longer protrusions at thecover glass surface that were characteristic of wild-type hemo-cytes (Movie S1). The live cell imaging suggests that autophagymight contribute to the cell protrusion attachment or extensionthat occurs along with cell flattening.
Autophagy and Integrin Share Functions in Cell Spreading. Cellprotrusion formation and spreading are associated with a focaladhesion function for surface attachment, specifically throughintegrin–matrix interactions (5). Drosophila βPS1-integrin, en-coded by myospheroid (mys), has been shown to be required forthe spreading of a Drosophila hemocyte-derived cell line (27).Similar to the autophagy requirement, we found that mys func-
tion was required for hemocyte cell spreading (Fig. 5B), but notfor association of sessile hemocytes with the body wall (Fig.S3A). Given the shared defect in cell spreading, we investigatedβPS1-integrin distribution. In wild-type hemocytes, we found βPS1-integrin along the surface of round cells before spreading and infoci that colocalized with F-actin in spread cells, presumably in-dicating focal adhesions (Fig. S3B). We found that βPS1-integrinalso localized to the plasma membrane in Atg1Δ3D mutant hemo-cytes and, in rare instances of spread hemocytes with this genotype,in a punctate pattern (Fig. S3C), suggesting that autophagy is notrequired for βPS1-integrin cell surface localization.Integrin-independent mechanisms in cell flattening and spread-
ing can be evoked through lectin-mediated surface attachment toCon A (28). Hemocytes disrupted for eithermys integrin-mediatedadhesion (Fig. 5F) orAtg1-mediated autophagy (Fig. 5H) were ableto flatten and spread with an increased cell footprint on Con A–coated cover glass, signifying that an autophagy-dependent functionfor cell spreading was not a broad essential requirement for cellattachment or flattening. However, unlike wild-type andmysRNAihemocytes, which extended spiky, filopodial-like protrusions duringthe first hour after plating onConA (Fig. 5E and F), theAtg1RNAior mutant hemocytes exhibited a discoid shape with lamellipodiarather than the normal protrusions (Fig. 5H). Taken together, thesefindings indicate that both autophagy and integrin functions inhemocytes are required for integrin-dependent cell spreading, butwith a distinct role for autophagy essential to hemocyte protrusionextension. This suggests that autophagy might have two separateroles or, alternatively, that autophagy plays a primary role in cellprotrusion that affects integrin-dependent attachment in spreading.
Autophagy Is Required for Extension of Rho1-Induced Cell Protrusions.The hemocyte-targeted expression of a dominant negative Rho1N19
allele or disruption of the Rho1 effectors dia and rok resulted inhemocytes that maintained a round cell shape (Fig. S3 D–F).Normal Rho1 pathway function was essential for hemocytespreading on both cover glass and (unlike autophagy) Con A sub-strates (Fig. 5G). In contrast, hemocytes expressing the constitu-tively active Rho1V14 allele resulted in enhanced cell spreading,with cells exhibiting longer and more prominent F-actin cell pro-trusions (Fig. 5C and J; 2.3± 0.1 μm for control vs. 5.5± 0.3 μm forRho1V14). Although hemocytes could spread, the extensive pro-trusions induced by the Rho1V14 allele were suppressed by simul-taneous inhibition of autophagy with theAtg1Δ3Dmutation (Fig. 5Dand J; 1.0 ± 0.1 μm). This finding demonstrates that autophagy canmodulate Rho1 pathway functions and implicates a specific effectof autophagy at the level of cortical protrusion and remodeling.
Ref(2)P Function Reveals a Role for p62-Selective Autophagy inHemocyte Spreading. Autophagosome formation requires com-mon assembly factors and the addition of new membrane, re-gardless of the encapsulated contents (23), which could indirectlydisrupt cortical remodeling through effects on intracellularmembrane dynamics. Alternatively, autophagy possibly could playa role in cortical remodeling through the sequestration and/orlysosomal degradation of a specific factor that normally eitherantagonizes protrusions or promotes a round cell shape. p62 hasbeen proposed to serve as an adaptor for delivery of ubiquitinatedcargo to the autophagosome in selective forms of auto-phagy (11, 29).We found that hemocyte spreading was dependenton ref(2)P encoding Drosophila p62 (Fig. 5I′) (30). This points toselective autophagy of an ubiquitinated substrate as a likely targetof an autophagy-dependent mechanism for cortical remodeling.
Autophagy Is Required for Hemocyte Recruitment to Wound Sites.Wounding of the larval epidermis leads to melanized clot for-mation and recruitment of hemocytes from circulation (31). At 6 hafter puncture wounding, microscopy imaging through the cuticleof live wild-type larvae revealed GFP-positive hemocytes accu-
Fig. 4. Hemocyte cortical dynamics occur but are altered in absence ofautophagy. The cell cortex was dynamic in both GFP-labeled wild-type(Upper) and Atg1Δ3D mutant hemocytes (Lower; Movie S1), with stuntedprotrusions observed in the absence of autophagy. Frames every 40 s fromtime-lapse microscopy imaging. (Scale bar: 5 μm.)
10504 | www.pnas.org/cgi/doi/10.1073/pnas.0914168107 Kadandale et al.

mulating around and within the clot (50.5 cells; Fig. 6 A, A′, andD). In contrast, fewer hemocytes surrounded the wound site inAtg1Δ3D mutant larvae (22.2 cells) and in larvae with hemocyte-targeted Atg1 RNAi (18.2 cells) (Fig. 6 B–D). This finding dem-onstrates that autophagy in hemocytes is important for physio-logically relevant blood cell behavior within the intact animal.
A Role of Autophagy in Regulated Cell Shape Changes in PrimaryMouse Macrophages. To address whether autophagy is broadly re-quired for blood cell remodeling, we tested the effects of disruptionof autophagy on cell shape changes in mammalian macrophages.Primary mouse macrophages plated on glass demonstrated cellspreading with dramatic cell elongation (Fig. 7 A and D; 2.8 major:minor axis); however, macrophages disrupted for autophagy withsiRNA depletion ofULK1 (Atg1) orBeclin-1 (Atg6) did not undergo
extensive cell spreading or elongation and remained predominantlycircular in shape (Fig. 7 B and D; major:minor axis, 1.9 for ULK1siRNA and 2.2 for BECN1 siRNA; P < 0.005). We found similarcell spreading and elongation defects with 3MA disruption of auto-phagy in two different activated mammalian macrophage cell lines(Fig. S4). These data suggest a broad role for autophagy in regulatinghemocyte and macrophage cell shape changes, with significance forconserved immune cell functions.
DiscussionWe have identified a continuous role for autophagy in blood cellcortical remodeling, with involvement in the extension of cellprotrusions during initiation andmaintenance of integrin-mediatedcell spreading. The cortical regulation of actin-based membrane
Fig. 5. Role for Ref(2)P-selective autophagy in integrin-dependent spreading and Rho1-induced cell protrusions. F-actin in hemocytes plated on cover glass(A–D, I and I′) or Con A–coated cover glass (E–H); Pxn-GAL4. Hemocytes with mys βPS1-integrin RNAi failed to spread on cover glass (B), but spread withprotrusions on con A (F); in contrast, Atg1Δ3D mutant hemocytes lacked protrusions on con A (H). Cells expressing dominant-negative Rho1N19 remained roundon cover glass (Fig. S3D) and on con A (G). (C) Hemocyte expression of constitutively active RhoV14 resulted in longer cell protrusions. (D) Blocking autophagysuppressed RhoV14-induced cell protrusions. (I and I′) Hemocyte spreading is p62-dependent. ref(2)Pod1/+ (I) and mutant hemocytes (I′) on cover glass. (J)Protrusion length (μm, mean ± SEM). (Scale bar: 5 μm; 2.5 μm in zoomed crops of cell edges).
Fig. 6. Autophagy is required for hemocyte recruitment to larval wound sites. Whole larva indicates approximate position of wound sites (arrow) and regionof low magnification images (white box). Recruitment of GFP-positive hemocytes 6 h. after larval wounding in control (A and A′), Atg1Δ3D mutant (B and B′),and hemocyte-targeted Atg1 RNAi larvae (C and C′), with Pxn-GAL4 GFP expression. White outlines depict the regions (Upper) shown at higher magnificationbelow, and the sites of melanized wounds (Lower). (D) Mean number of hemocytes (± SEM) within two wound diameters per larva. (Scale bar: 100 μm.)
Kadandale et al. PNAS | June 8, 2010 | vol. 107 | no. 23 | 10505
CELL
BIOLO
GY

protrusions, such as lamellipodia or filopodia, and cell–matrixattachments are linked through shared pathways (5), and both playmajor roles in cell migration and probing cell contacts relevant toimmune cell function. The cell mechanics of cortical remodelinginvolve cyclic processes for protrusive force and contractions, aswell as focal adhesion assembly and disassembly. Responsive mo-lecular mechanisms must dynamically monitor and balance com-peting activities through macromolecular turnover and rapid re-novation of cellular architecture. Examples of this are seen atmultiple levels withinRhoGTPase pathways that alter localization,activity, and/or destruction of key regulators that control microfil-ament network organization and dynamics. We propose proteinsequestration in autophagosomes and autophagic clearance as ad-ditional mechanisms through which cellular remodeling for dy-namic cell states can be achieved.Previous studies of autophagy have focused on its roles in cell
growth, survival, and death as responses to starvation and stress(10). We found that basal autophagy does not appear to affectcell survival, numbers, differentiation, bioenergetics, or adhesion oflarval hemocytes. Autophagy is known to encapsulate bulk cyto-plasmic contents and to selectively target specific protein aggregatesand organelles. The regulation and targets for selective autophagyare not well understood, however. An emergingmechanism appearsto involve parallels to the ubiquitin-proteosome system (29), inwhich p62 is envisioned to serve as a bridge for the delivery ofubiquitinated cargo to autophagosomes. Interestingly, p62 also hasbeen implicated in growth factor–induced neurite outgrowth withpossible roles in recruitment of ULK1 to cortical growth receptors(16). This lends further support to the idea that autophagy mightplay a role in neurite outgrowth (15–17) and synaptic developmentat theDrosophila neuromuscular junction (32). One known target ofselective autophagy is in the cell renovation that follows the mor-phogenetic event of cytokinesis, whereby an ubiquitinated midbodyring complex is disposed of through p62-mediated autophagicclearance (33). Given the prevalence and possibilities for ubiquitinprotein modifications, ubiquitination could play roles in both pro-teosomal and selective autophagic turnover (29).
The role of autophagy in blood cell spreading could work at thelevel of facilitating removal of cortical tension or promoting ex-tension of protrusions, which might be coordinated with substrateattachment. We envision that autophagy could serve as a rapidmeans to sequester or dampen specific signaling activities in re-sponse to a changing environment or cues. This is analogous toother posttranslational mechanisms that use ubiquitination orphosphorylation. The shared role for ref(2)P suggests a model in-volving p62-mediated autophagosome sequestration or autophagicdegradation of a potentially ubiquitinated substrate. We identifiedtheRho1 pathway as a potential point of intersection. Interestingly,a growing list of Rho pathway components with roles relevant tocontrol of cell protrusion and spreadinghavebeen found toundergoubiquitination and changes in protein stability (34–36), and recentconnections suggest a potential reciprocal regulatory relationshipbetween the Rho and autophagic pathways (37, 38). Componentsthat are involved in changing levels of focal adhesion activity andthat correspond to a rounding-up of cells also have been identifiedas being under ubiquitin regulation (39, 40). Connections also havebeen made linking focal adhesion components directly to the pro-cess of autophagy (20, 41, 42), as well as to cell spreading. Con-ceptually, numerous candidate targets of selective autophagyremain to be investigated.Autophagy has known functions in immune cells in the moun-
ting of antimicrobial responses through its roles in antigen pre-sentation and the targeted destruction of intracellular pathogens(43). Our present findings demonstrate that autophagy also con-trols immune cell remodeling, which has significance in immunecell surveillance. We found that hemocytes that were disruptedfor autophagy and the ability to undergo cell spreading ex vivoalso showed decreased recruitment to wound sites in the animals.Consistent with this finding, wild-type hemocytes recruited tolarval wound sites exhibited cell spreading, extension of cellprocesses, and more persistent attachment, in contrast to thesessile hemocyte population with rounded-up morphology (31).We have shown that mouse macrophages subject to blockage ofautophagy exhibited defects in induced cell shape changes, withsimilar morphological phenotypes to those of effects on signalingthrough integrin and Rho pathways also important for macro-phage migration in inflammatory infiltration (44). The require-ment for continuous autophagy in hemocyte spreading also mightbe relevant in other contexts that require a switch to modulatecell morphology, such as rounding-up and respreading duringmitosis and events in metastasis.
Materials and MethodsFor additional information, see SI Materials and Methods.
Hemocyte Isolation. Wandering third instar larvae were used for all experi-ments. To isolate hemocytes, larvae were rinsed briefly in 70% ethanol andthen in PBS, and placed in 100 μL of PBS or medium on a cover glass (FisherScientific). The cuticle was gently ripped open with fine forceps, and re-leased hemocytes were allowed to settle and spread at 25 °C for 30 min.
Hemocyte Treatments. Studies were done using 10 μM 3MA (Sigma-Aldrich),400 μM methyl pyruvate (Sigma-Aldrich), or 1 μg/mL of propidium iodide(Sigma-Aldrich) in PBS. Cell counts were determined using a hemocytometer.
Wounding Procedure and Live in Vivo Imaging. Wandering third instar larvaewere immobilized ventral side down on double-sided tape. Larvae werewounded with a pulled injection needle in the middle of the A5 segment. Adrop of water was used to recover wounded larvae, then incubated on grape-agar plates at 25 °C for 6 h. The larvae were remounted on a glass slide withdouble-sided tape, and GFP-labeled hemocytes around the melanizedwound site were visualized by wide-field fluorescence on a Leica DM1600with 5× (NA 0.15) and 20× (NA 0.5) objectives.
Live Imaging of Hemocyte Dynamics. Hemocytes expressing eGFP driven by Cg-GAL4 or Pxn-GAL4were dissected as above into 100-μL completemedium andallowed to settle on the cover glass for 30 min at 25 °C before imaging. For
Fig. 7. Autophagy plays a role in mouse macrophage spreading and elon-gation. (A–C) F-actin–stained primary mouse macrophages following siRNA.(A) Control siRNA macrophages elongated on glass. (B and C) RNAi depletionof ULK1 (B and B′) or BECN1 (C and C′) inhibited macrophage spreading andelongation. (D) Cell shape as ratio of major:minor cell axis lengths (1 =round; mean ± SEM) for >94 cells. (Scale bar: 25 μm.)
10506 | www.pnas.org/cgi/doi/10.1073/pnas.0914168107 Kadandale et al.

time-lapse recordings, images were captured every 5 s for a total of 180 s onan Olympus FV1000 point scanning microscope.
Primary Macrophage Isolation, siRNA, and Spreading Assay. Primary macro-phages were elicited by i.p. injection of thioglycollate, as described previously(45). For siRNA knockdown, 2.5 × 106 cells plated per 35-mm dish weretransfected with 30 nM Dharmacon siGENOME SMARTpool siRNAs (control,#D-001206-13-05; ULK-1, #M040155-00-0005; and Beclin1, #M055895-01-0005). The DeliverX siRNA Transfection Kit (Panomics) was used in accordancewith the manufacturer’s instructions, with modifications. After incubationwith the transfection mix, cells were added to 1.5 mL of OptiMEM Reduced-Serum medium (Invitrogen), incubated at 37 °C for 60 h, and then harvestedfor cell-spreading assays or protein isolation for Western blot analyses (SIMaterials and Methods). For the macrophage-spreading assay, 5 × 105 cellsafter siRNA transfection were seeded on 12-mm circular cover glass (FisherScientific), incubated at 37 °C for 3 h, and then fixed and stained for F-actin.
Microscopy Image Analysis. To quantitate cell spreading, images of cell fieldswere manually counted for spread and round cells. To quantitate the sessilehemocytes, CellProfiler was used to automatically identify hemocytes.Lengths of cell protrusions were manually traced and recorded using Volocity
(PerkinElmer). The 20× images of wounded larvae were analyzed in Pho-toshop (Adobe Systems) by manually counting hemocytes within two-wounddiameters. All data were exported to Microsoft Excel, and means, SEs, andStudent t test values were determined.
ACKNOWLEDGMENTS. We thank T. Neufeld (University of Minnesota),D. Contamine and S. Gaumer (University Versailles-St. Quentin), I. Andó(Hungary Academy of Sciences), G. Chen (Institute of Biological Chemistry,Taipei), M. Galko (University of Texas, M.D. Anderson Cancer Center), theBloomington Drosophila Stock Center, the Vienna Drosophila RNAi Center,and Developmental Studies Hybridoma Bank for reagents; P. Wiesner andY. Miller (University of California, San Diego) for providing mmLDL andJ774A.1 cells; K. Lau and T. Nguyen for technical support; and members ofthe A.A.K. laboratory for comments. We are grateful to T. Meerloo for tech-nical assistance with electron microscopy preparation and M. Farquhar foraccess to the CMM Electron Microscope Facility (National Institutes ofHealth Grant DK17780). Imaging was conducted in part in the University ofCalifornia San Diego Neuroscience Microscopy Shared Facility (National Insti-tutes of Health Grant P30 NS047101). This work was supported by funds fromthe Human Frontier Science Program and The David & Lucille Packard Foun-dation (to A.A.K.), NIH LIPID MAPs consortium Grant GM069338 (to C.K.G.)and Ruth L. Kirschstein National Research Service Award NIH/NCI T32CA009523 (to J.D.S.).
1. Paladi M, Tepass U (2004) Function of Rho GTPases in embryonic blood cell migrationin Drosophila. J Cell Sci 117:6313–6326.
2. Stramer B, et al. (2005) Live imaging of wound inflammation in Drosophila embryosreveals key roles for small GTPases during in vivo cell migration. J Cell Biol 168:567–573.
3. Williams MJ, Habayeb MS, Hultmark D (2007) Reciprocal regulation of Rac1 and Rho1in Drosophila circulating immune surveillance cells. J Cell Sci 120:502–511.
4. Ridley AJ (2006) Rho GTPases and actin dynamics in membrane protrusions and vesicletrafficking. Trends Cell Biol 16:522–529.
5. DeMali KA, Burridge K (2003) Coupling membrane protrusion and cell adhesion. J CellSci 116:2389–2397.
6. Palamidessi A, et al. (2008) Endocytic trafficking of Rac is required for the spatialrestriction of signaling in cell migration. Cell 134:135–147.
7. Kunda P, Craig G, Dominguez V, Baum B (2003) Abi, Sra1, and Kette control thestability and localization of SCAR/WAVE to regulate the formation of actin-basedprotrusions. Curr Biol 13:1867–1875.
8. Chen Y, et al. (2009) Cullin mediates degradation of RhoA through evolutionarilyconserved BTB adaptors to control actin cytoskeleton structure and cell movement.Mol Cell 35:841–855.
9. Niwa R, Nagata-Ohashi K, Takeichi M, Mizuno K, Uemura T (2002) Control of actinreorganization by Slingshot, a family of phosphatases that dephosphorylate ADF/cofilin. Cell 108:233–246.
10. He C, Klionsky DJ (2009) Regulation mechanisms and signaling pathways ofautophagy. Annu Rev Genet 43:67–93.
11. Bjørkøy G, et al. (2005) p62/SQSTM1 forms protein aggregates degraded byautophagy and has a protective effect on huntingtin-induced cell death. J Cell Biol171:603–614.
12. Komatsu M, et al. (2007) Homeostatic levels of p62 control cytoplasmic inclusion bodyformation in autophagy-deficient mice. Cell 131:1149–1163.
13. Pankiv S, et al. (2007) p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradationof ubiquitinated protein aggregates by autophagy. J Biol Chem 282:24131–24145.
14. Nezis IP, et al. (2008) Ref(2)P, the Drosophila melanogaster homologue of mammalianp62, is required for the formation of protein aggregates in adult brain. J Cell Biol 180:1065–1071.
15. Ogura K, et al. (1994) Caenorhabditis elegans unc-51 gene required for axonalelongation encodes a novel serine/threonine kinase. Genes Dev 8:2389–2400.
16. Zhou X, et al. (2007) Unc-51–like kinase 1/2–mediated endocytic processes regulatefilopodia extension and branching of sensory axons. Proc Natl Acad Sci USA 104:5842–5847.
17. Tomoda T, Bhatt RS, Kuroyanagi H, Shirasawa T, Hatten ME (1999) A mouse serine/threonine kinase homologous to C. elegans UNC51 functions in parallel fiberformation of cerebellar granule neurons. Neuron 24:833–846.
18. Scott RC, Schuldiner O, Neufeld TP (2004) Role and regulation of starvation-inducedautophagy in the Drosophila fat body. Dev Cell 7:167–178.
19. Matsuura A, Tsukada M, Wada Y, Ohsumi Y (1997) Apg1p, a novel protein kinaserequired for the autophagic process in Saccharomyces cerevisiae. Gene 192:245–250.
20. Chen GC, et al. (2007) Genetic interactions between Drosophila melanogaster Atg1and paxillin reveal a role for paxillin in autophagosome formation. Autophagy 4:37–45.
21. Seglen PO, Gordon PB (1982) 3-Methyladenine: Specific inhibitor of autophagic/lysosomal protein degradation in isolated rat hepatocytes. Proc Natl Acad Sci USA 79:1889–1892.
22. Dietzl G, et al. (2007) A genome-wide transgenic RNAi library for conditional gene
inactivation in Drosophila. Nature 448:151–156.23. Xie Z, Klionsky DJ (2007) Autophagosome formation: Core machinery and adaptations.
Nat Cell Biol 9:1102–1109.24. Wyers F, Dru P, Simonet B, Contamine D (1993) Immunological cross-reactions and
interactions between the Drosophila melanogaster ref(2)P protein and sigma
rhabdovirus proteins. J Virol 67:3208–3216.25. Lanot R, Zachary D, Holder F, Meister M (2001) Postembryonic hematopoiesis in
Drosophila. Dev Biol 230:243–257.26. Lum JJ, et al. (2005) Growth factor regulation of autophagy and cell survival in the
absence of apoptosis. Cell 120:237–248.27. Kiger AA, et al. (2003) A functional genomic analysis of cell morphology using RNA
interference. J Biol 2: article 27.28. Jani K, Schöck F (2007) Zasp is required for the assembly of functional integrin
adhesion sites. J Cell Biol 179:1583–1597.29. Kirkin V, McEwan DG, Novak I, Dikic I (2009) A role for ubiquitin in selective
autophagy. Mol Cell 34:259–269.30. Gay P, Contamine D (1993) Study of the ref(2)P locus of Drosophila melanogaster, II:
Genetic studies of the 37DF region. Mol Gen Genet 239:361–370.31. Babcock DT, et al. (2008) Circulating blood cells function as a surveillance system for
damaged tissue in Drosophila larvae. Proc Natl Acad Sci USA 105:10017–10022.32. Shen W, Ganetzky B (2009) Autophagy promotes synapse development in Drosophila.
J Cell Biol 187:71–79.33. Pohl C, Jentsch S (2009) Midbody ring disposal by autophagy is a post-abscission event
of cytokinesis. Nat Cell Biol 11:65–70.34. Wang HR, et al. (2003) Regulation of cell polarity and protrusion formation by
targeting RhoA for degradation. Science 302:1775–1779.35. DeWard AD, Alberts AS (2009) Ubiquitin-mediated degradation of the formin mDia2
upon completion of cell division. J Biol Chem 284:20061–20069.36. Yoo Y, Ho HJ, Wang C, Guan JL (2010) Tyrosine phosphorylation of cofilin at Y68 by v-
Src leads to its degradation through the ubiquitin-proteasome pathway. Oncogene
29:263–272.37. Baisamy L, Cavin S, Jurisch N, Diviani D (2009) The ubiquitin-like protein LC3 regulates
the Rho-GEF activity of AKAP-Lbc. J Biol Chem 284:28232–28242.38. Bauer PO, et al. (2009) Inhibition of Rho kinases enhances the degradation of mutant
huntingtin. J Biol Chem 284:13153–13164.39. Dao VT, Dupuy AG, Gavet O, Caron E, de Gunzburg J (2009) Dynamic changes in Rap1
activity are required for cell retraction and spreading during mitosis. J Cell Sci 122:
2996–3004.40. Shao Y, Elly C, Liu YC (2003) Negative regulation of Rap1 activation by the Cbl E3
ubiquitin ligase. EMBO Rep 4:425–431.41. Ueda H, Abbi S, Zheng C, Guan JL (2000) Suppression of Pyk2 kinase and cellular
activities by FIP200. J Cell Biol 149:423–430.42. Hara T, et al. (2008) FIP200, a ULK-interacting protein, is required for autophagosome
formation in mammalian cells. J Cell Biol 181:497–510.43. Virgin HW, Levine B (2009) Autophagy genes in immunity. Nat Immunol 10:461–470.44. Okigaki M, et al. (2003) Pyk2 regulates multiple signaling events crucial for
macrophage morphology and migration. Proc Natl Acad Sci USA 100:10740–10745.45. Pascual G, et al. (2005) A SUMOylation-dependent pathway mediates transrepression
of inflammatory response genes by PPAR-gamma. Nature 437:759–763.
Kadandale et al. PNAS | June 8, 2010 | vol. 107 | no. 23 | 10507
CELL
BIOLO
GY

Supporting InformationKadandale et al. 10.1073/pnas.0914168107SI TextTo address whether autophagy is broadly required for blood cellremodeling, we tested the effects of autophagy disruption onmammalian macrophage cell shape changes that coincide withactivation. RAW 264.7 macrophages underwent enhanced cellspreading and elongation on growth in low-glucose media (Fig. S4A and B; 1.7 major:minor axis); however, macrophages disruptedfor autophagy with 3MA failed to undergo low glucose–inducedelongation and remained predominantly round (Fig. S4 C and D;1.4 axis ratio; P < 0.001). Similarly, in the J774A.1 mouse macro-phage cell line, cellular elongation from a round to a bipolar cellshape occurred in response to treatment with aminimally oxidizedform of LDL (mmLDL) relevant to the early events leading toatherosclerosis (1) (Fig. S4 E and F; 1.8 axis ratio). J774A.1 cellsdisrupted for autophagy with 3MA failed to elongate when treatedwith mmLDL and instead remained round or extended severalmultipolar protrusions (Fig. S4G andH; 1.5 axis ratio; P < 0.001).These findings suggest a broad role for autophagy in regulatinghemocyte and macrophage cell shape changes.
SI Materials and MethodsFly Strains. The following genotypes were used: (i) Atg1Δ3D/TM6BTb Hu, provided by T. Neufeld (University of Minnesota) (2), andCg-GAL4, UAS-EGFP; Atg1Δ3D/TM6C Sb Tb, w; Pxn-GAL4,Atg1Δ3D/TM6B Tb Hu, w; UAS-EGFP; Pxn-GAL4, Atg1Δ3D/TM6BTb Hu and w; UAS-mCherry:Atg1; Atg1Δ3D/TM6B Tb Hu; (ii)UAS-dAtg1-IR, provided by G. Chen (Institute of BiologicalChemistry, Taipei) (3); (iii) w1118; UAS-IR-Atg4V107317, w1118;UAS-IR-Atg6V22122, w1118; UAS-IR-Atg7V27432, w1118; UAS-IR-At-g8aV43097, w1118; UAS-IR-Atg9V10045, w1118; UAS-IR-mysV103704
and w1118; UAS-IR diaV103914/CyO, received from the ViennaDrosophila RNAi Center (4); (iv) w1118; Df(3L)ED4486, P{w[+mW.Scer\FRT.hs3]=3′.RS5+3.3′}ED4486/TM6C, cu1 Sb1, w;Cg-GAL4, P{w[+mC]=UAS-2xEGFP}AH2, w*; P{w[+mC]=UAS-Rho1.V14}2.1, w*; P{w[+mC]=UAS-Rho1.N19}1.3, andw*; P{w[+mC]=UAS-rok.CAT-KG}3, received from Bloo-mington Drosophila Stock Center; (v) ref(2)P[od1]/CyO, Act-GFPand ref(2)P[od3]/CyO, Act-GFP, rebalanced from stocks providedfrom D. Contamine (University Versailles-St. Quentin) (5); and(vi) w; Pxn-GAL48.1.1, UAS-GFP, provided by M. Galko (Uni-versity of Texas, M.D. Anderson Cancer Center).
Construction ofmCherry:Atg1 Flies.Atg1 cDNA was amplified fromclone LP23904 (from Drosophila Genomics Resource Center)with primers CACCATGAATATTGTCGGGGAATATGAA-TACAGC and AGCATTCTCATCGGTCATGTATAT, clonedinto pENTR/D-TOPO Gateway entry vector (Invitrogen), andthen recombined into the pTW1129 destination vector (providedby Terence Murphy; Drosophila Gateway Vector Collection) togenerate pUAST-mCherry:Atg1. The DNA construct was in-jected into flies (BestGene) to obtain the corresponding trans-genic stocks.
Hemocyte Isolation and Enrichment for Molecular Analysis. Thirdinstar larvae were scooped out of the culture vials and washed with70% ethanol and then with PBS, with ∼150 mg larval wet weightused per preparation. The larvae were added to a cell strainerwith a 70-μm nylon mesh placed in a shallow collecting dish andcrushed with the back of an Eppendorf tube. The released he-mocytes within the crude preparation were filtered through the70-μm filter on the cell strainer into a 50-mL tube, eliminatinglarge tissues. The crushed larvae were washed three times with
PBS (pH 7.4). The hemocytes were centrifuged at 8 × g for 5 min,and the supernatant containing the hemocytes was carefully col-lected in a fresh tube and centrifuged at 805 × g for 10 min. Thesupernatant was discarded, and the collected hemocytes were usedfor subsequent steps. This procedure resulted in a highly purifiedhemocyte preparation, as indicated by (i) visual examination ofa preparation from larvae with hemocyte-specific Pxn-GAL4,UAS-GFP expression showing that ∼70–80% of intact cells wereGFP-positive, and (ii) RT-PCR showing little or no contamina-tion of a target RNA knocked down only in hemocytes.
RT-PCR in Hemocytes. Hemocytes were collected as describedabove. RNA was extracted using TRIzol reagent (Invitrogen),following the manufacturer’s instructions. First-strand cDNAsynthesis was achieved using the Invitrogen SuperScript III First-Strand synthesis kit. Gene-specific primers were then used tocheck for expression of target genes.
Immunofluorescence Cell Staining. Hemocytes were dissected into100 μL of PBS, allowed to attach to a glass coverslip for 30–60min at 25 °C, then fixed with 3.7% formaldehyde in PBS. Cellswere permeabilized with 0.1% Triton X-100 in PBS, blockedwith PBSTB (3% BSA + 0.1% Triton X-100 in PBS), andstained with primary antibodies rabbit anti-Ref(2)P (1:1,000,provided by D. Contamine and S. Gaumer) (5), mouse anti-β-PS1 CF.6G11 (1:50, obtained from the Developmental StudiesHybridoma Bank), or anti-P1 (1:100, provided by I. Andó) (6)overnight at 4 °C. Cells were washed three times with PBS, in-cubated with secondary antibodies or stain for F-actin [PBSTB,Alexa Fluor 546 phalloidin, and Alexa Fluor 488 goat anti-mouseor anti-rabbit (Invitrogen)] at 4 °C overnight, washed three timeswith PBS, incubated with DAPI for 10 min, washed three moretimes, and mounted in FluorSave (Calbiochem).
Electron Microscopy. Hemocytes were dissected out of wanderingthird-instar larvae of the appropriate genotype as describedabove, and then concentrated by centrifugation for fixation.Samples were immersed in modified Karnovsky’s fixative [1.5%glutaraldehyde, 3% paraformaldehyde, and 5% sucrose in 0.1 Mcacodylate buffer (pH 7.4)] for 8 h, postfixed in 1% osmiumtetroxide in 0.1 M cacodylate buffer for 1 h, and then stained enbloc in 1% uranyl acetate for 1 h. The samples were then de-hydrated in ethanol, embedded in epoxy resin, sectioned at 60–70 nm, and picked up on Formvar- and carbon-coated coppergrids. The grids were stained with uranyl acetate and lead nitrate,viewed using a Philips CM-10 transmission electron microscope,and photographed using a Gatan digital camera.
Macrophage Culture and Assays. J774A.1 cells grown in standardDMEM were treated for 3 h with 50 μg/mL of mmLDL [providedby Y. Miller and prepared as described previously (1)], in eitherthe absence or presence of 10 mM 3MA (Sigma-Aldrich). Thencells were fixed and stained for F-actin. RAW 264.7 cells weregrown in DMEM supplemented with 4.5 g/L of glucose (highglucose). To induce elongation, cells were shifted to mediumwith 1 g/L of glucose (low glucose) in either the absence orpresence of 1 mM 3MA, and incubated overnight.
Western Blot Analysis. A total of 2 × 106 primary macrophages fromeach siRNA condition were lysed in 150 μLof IP buffer [25mMTris(pH 7.4), 1 mM EDTA (pH 8.0), 0.1 mM EGTA (pH 8.0), 5 mMMgCl2, 150 mMNaCl, 2 mM sodium orthovanadate, 10% glycerol,1% IGEPAL-630/Nonidet P-40, 1 mM DTT, and 1× protease in-
Kadandale et al. www.pnas.org/cgi/content/short/0914168107 1 of 5
hibitor mixture (Sigma-Aldrich)] for 20 min at 4 °C. Lysates wereclearedby centrifugation at 12,000×g for 10minat 4 °C.A total of 20μL of lysate was used in each lane of a NuPAGENovex 4–12%Bis-Tris Gel (Invitrogen). Gels were transferred to an ImmobilonP-PVDF transfer membrane (Millipore) and stained with anti-
tubulin (Sigma-Aldrich; #T9026, 1:1,000), anti-ULK1 (Sigma-Aldrich;#A7481, 1:1,000), or anti-Beclin1 (Sigma-Aldrich;#B6061,1:2,500) antibodies. HRP-conjugated anti-rabbit or anti-mouse (In-vitrogen) were used for detection with the SuperSignal West FemtoMaximum Sensitivity Kit (Pierce).
1. Miller YI, et al. (2003) Minimally modified LDL binds to CD14, induces macrophagespreading via TLR4/MD-2, and inhibits phagocytosis of apoptotic cells. J Biol Chem 278:1561–1568.
2. Scott RC, Schuldiner O, Neufeld TP (2004) Role and regulation of starvation-inducedautophagy in the Drosophila fat body. Dev Cell 7:167–178.
3. Chen GC, et al. (2007) Genetic interactions between Drosophila melanogaster Atg1and paxillin reveal a role for paxillin in autophagosome formation. Autophagy 4:37–45.
4. Dietzl G, et al. (2007) A genome-wide transgenic RNAi library for conditional geneinactivation in Drosophila. Nature 448:151–156.
5. Wyers F, Petitjean AM, Dru P, Gay P, Contamine D (1995) Localization of domainswithin the Drosophila Ref(2)P protein involved in the intracellular control of sigmarhabdovirus multiplication. J Virol 69:4463–4470.
6. Kurucz E, et al. (2007) Definition of Drosophila hemocyte subsets by cell-type–specificantigens. Acta Biol Hung 58(Suppl):95–111.
Cg>GFP, >IR-Atg1
B
D
E
C
+ - + - + - + -RT
ControlAtg1 ControlAtg1RNAi
ActinAtg1Primer
A
Pxn>GFP/mCh:Atg1+;Atg1D3D
Pxn>GFP;Atg1D3D
GFP GFP GFP
Pxn>GFP;Atg1D3D/+
Atg6 Actin
lacZ Atg6
Primer
RNAi
Atg7 Actin
lacZ Atg7
Atg8a Actin
lacZ Atg8a
F-Actin
Cg>GFP
Cg>GFP, >IR-Atg4 Cg>GFP, >IR-Atg6
Cg>GFP, >IR-Atg7 Cg>GFP, >IR-Atg8a Cg>GFP, >IR-Atg9
Cg>GFP
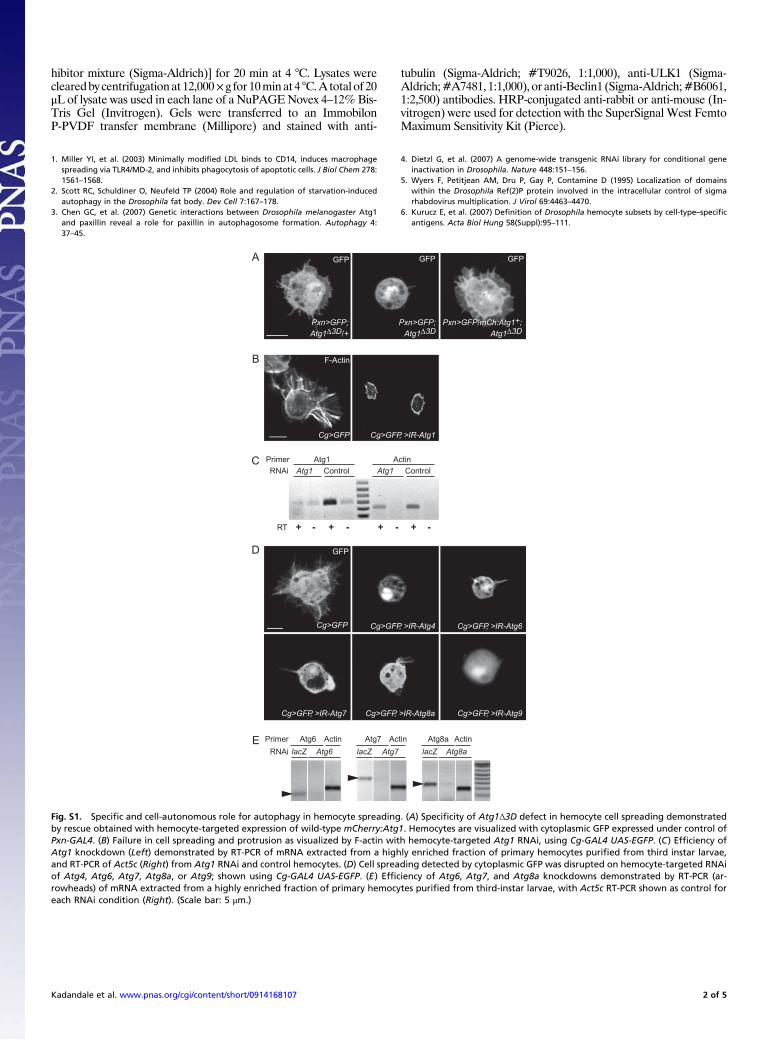
GFP
Fig. S1. Specific and cell-autonomous role for autophagy in hemocyte spreading. (A) Specificity of Atg1Δ3D defect in hemocyte cell spreading demonstratedby rescue obtained with hemocyte-targeted expression of wild-type mCherry:Atg1. Hemocytes are visualized with cytoplasmic GFP expressed under control ofPxn-GAL4. (B) Failure in cell spreading and protrusion as visualized by F-actin with hemocyte-targeted Atg1 RNAi, using Cg-GAL4 UAS-EGFP. (C) Efficiency ofAtg1 knockdown (Left) demonstrated by RT-PCR of mRNA extracted from a highly enriched fraction of primary hemocytes purified from third instar larvae,and RT-PCR of Act5c (Right) from Atg1 RNAi and control hemocytes. (D) Cell spreading detected by cytoplasmic GFP was disrupted on hemocyte-targeted RNAiof Atg4, Atg6, Atg7, Atg8a, or Atg9; shown using Cg-GAL4 UAS-EGFP. (E) Efficiency of Atg6, Atg7, and Atg8a knockdowns demonstrated by RT-PCR (ar-rowheads) of mRNA extracted from a highly enriched fraction of primary hemocytes purified from third-instar larvae, with Act5c RT-PCR shown as control foreach RNAi condition (Right). (Scale bar: 5 μm.)
Kadandale et al. www.pnas.org/cgi/content/short/0914168107 2 of 5
1 µm1 µm
Atg1D3D/+ Atg1D3DA
C
B
D
E F + methyl pyruvate
% P
I-positiv
e c
ells
# s
essile
hem
ocyte
s
per
segm
ent
350
300
250
200
150
100
50
0
5 5
Atg1D3D-/+ -/-
Atg1D3D-/+ -/-
# h
em
ocyte
s
per
larv
a (
x1000) 6
5
4
3
2
1
0
5 5
-/+-/-
1 hour 6 hours
Atg1D3D70
60
50
40
30
20
10
0Cg>GFP;
Atg1D3D/+Cg>GFP;Atg1D3D
GFP
0.1 µm
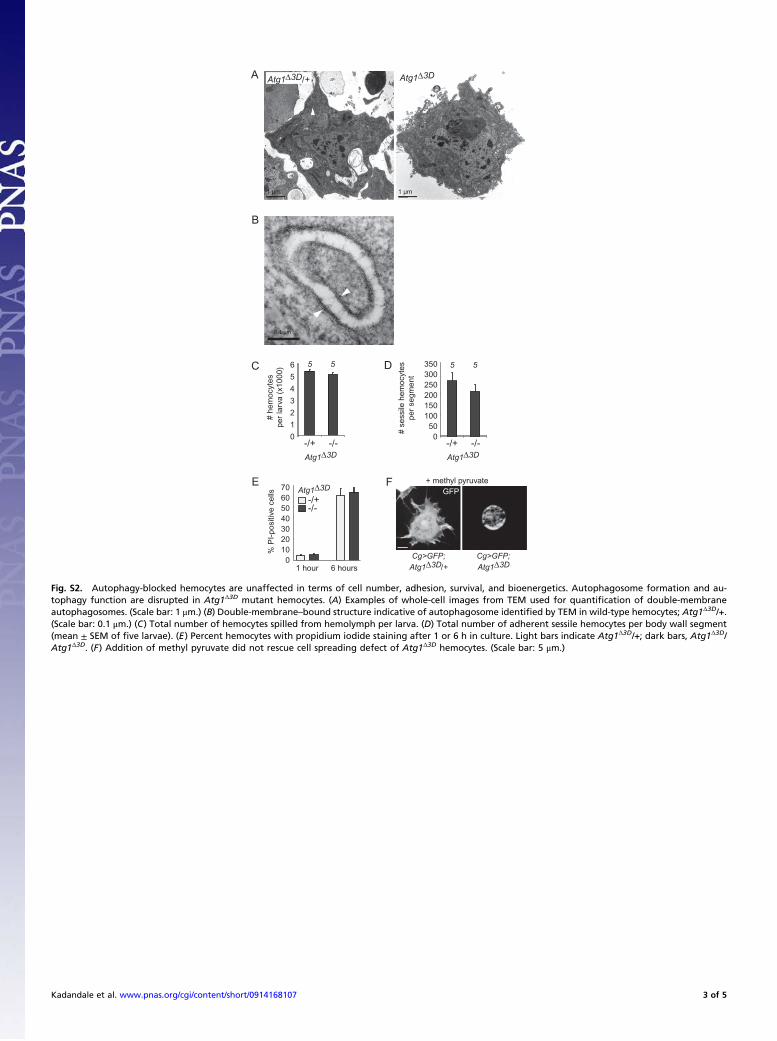
Fig. S2. Autophagy-blocked hemocytes are unaffected in terms of cell number, adhesion, survival, and bioenergetics. Autophagosome formation and au-tophagy function are disrupted in Atg1Δ3D mutant hemocytes. (A) Examples of whole-cell images from TEM used for quantification of double-membraneautophagosomes. (Scale bar: 1 μm.) (B) Double-membrane–bound structure indicative of autophagosome identified by TEM in wild-type hemocytes; Atg1Δ3D/+.(Scale bar: 0.1 μm.) (C) Total number of hemocytes spilled from hemolymph per larva. (D) Total number of adherent sessile hemocytes per body wall segment(mean ± SEM of five larvae). (E) Percent hemocytes with propidium iodide staining after 1 or 6 h in culture. Light bars indicate Atg1Δ3D/+; dark bars, Atg1Δ3D/Atg1Δ3D. (F) Addition of methyl pyruvate did not rescue cell spreading defect of Atg1Δ3D hemocytes. (Scale bar: 5 μm.)
Kadandale et al. www.pnas.org/cgi/content/short/0914168107 3 of 5

ControlControl
β-PS1DAPIB
C
A
120
20
Con
trol
60
80
40
100
# s
essile
hem
ocyte
s p
er
segm
ent
Atg
1 R
NAi
mys RNAi
D
Cg>Rho1N19 Cg>rokCat-KGCg>IR-dia
F-Actin E F
Atg1 Atg1Pxn-GAL4
3D3DΔ Δ
Fig. S3. Role of autophagy in hemocyte spreading shared with integrin and Rho1 functions. (A) Mean number (± SEM) of GFP-positive sessile hemocytes persegment imaged through the cuticle of live larvae from control and animals with Pxn-GAL4 expression of Atg1 or mys RNAi hairpins. (B and C) PS1 integrinprotein (green) was localized to the plasma membrane in few examples of round, unspread hemocytes from control and in most Atg1Δ3D mutant hemocytes(Left). PS1 integrin was in foci at the cell surface and along the periphery in most control hemocytes, and in rare examples of spread Atg1Δ3D mutant hemocytes(Right). DNA is shown in magenta. (D and F) Inhibition or knockdown of Rho1 pathway components disrupted hemocyte spreading, with Cg-GAL4 expressionof Rho1N19 (D), dia RNAi (E), or rokCat-KG (F). (Scale bars: 5 μm; 2.5 μm in the zoomed views of cell edges shown in D–F.)
AD
Hi Glc Low Glc
+3MA
F-actin DAPI
RA
W 2
64.7
cells
E
HMedia Media + mmLDL
+3MA
F-actin DAPI
J774A
.1 c
ells
1.6
1.7
1.8
1.9 p < 0.001
+m
mLD
L
Majo
r:M
inor
axis
1.5
1.6
1.7
1.8 p < 0.001
Low
Glc
Majo
r:M
inor
axis
+3MAControl
+3MAControl
B
F
C
G
Fig. S4. Autophagy plays a role in mouse macrophages for induced cell shape changes. (A and B) Mouse RAW 264.7 cells elongated on a switch from high-glucose to low-glucose media. (C) Addition of 3MA prevented cell elongation, and cells remained round in low glucose. (D) Cell shape as ratio of major:minorcell axis lengths (1 = round) for macrophages in low glucose; values are mean ± SEM for >500 cells. (E and F) J774A.1 mouse macrophages elongated inresponse to minimally modified LDL (mmLDL). (G) Addition of 3MA inhibited mmLDL-treated cell elongation, and cells became multipolar or remained round.(H) Ratio of major:minor cell axis lengths (1 = round) for macrophages in mmLDL; mean +SEM for > 500 cells. (Scale bar: 5 μm.)
Kadandale et al. www.pnas.org/cgi/content/short/0914168107 4 of 5

Movie S1. The cell cortex is highly dynamic in both wild-type and Atg1Δ3D mutant hemocytes. In autophagy-blocked hemocytes, only short protrusions withlack of cell spreading were observed.
Movie S1
Kadandale et al. www.pnas.org/cgi/content/short/0914168107 5 of 5