comparative physiolog ofy the vertebrate …jeb.biologists.org/content/jexbio/40/3/421.full.pdf ·...
TRANSCRIPT

J. Exp. Biol. (1963), 40, 421-436yVith 10 text-figuresPrinted in Great Britain
COMPARATIVE PHYSIOLOGY OF THE VERTEBRATEAUTONOMIC NERVOUS SYSTEM
II. INNERVATION OF THE URINARY BLADDER OF THE RINGTAILPOSSUM (PSEUDOCHEIRUS PEREGRINUS)
BY G. BURNSTOCK AND G. CAMPBELL
Department of Zoology, University of Melbourne, Victoria, Australia
{Received 30 January 1963)
INTRODUCTION
The work reported in this paper is one of a series of comparative physiologicalstudies, which is designed to clarify the pattern of evolution of the vertebrate auto-nomic nervous system. There have been many investigations of the anatomy andphysiology of the autonomic nervous system of placental mammals. (See reviews byLangley (1921), McSwiney (1931), Gruber (1933) and Hillarp (i960).) Less attentionhas been given to the physiology of the autonomic nervous systems of lower verte-brates (Nicol, 1952). The autonomic nervous system of marsupial mammals has beenparticularly neglected.
The present work is concerned with the innervation of the bladder of a commonAustralian marsupial, the ringtail possum (Pseudocheirus peregrinus). The results arediscussed in relation to the innervation of the bladder in other vertebrate groups.A brief preliminary account of this work has been presented previously (Campbell &Burnstock, 1962).
METHOD
Specimens of the ringtail possum were caught in the vicinity of Melbourne, and were usedfor experiments within 2 weeks of capture. General histological studies were made onbladders fixed in Bouin's solution and stained with haematoxylin and eosin or with Masson'striple stain. Nerve fibres were stained by inflating the bladder with 0-05 % methylene-bluesolution at pH 6. The bladders were then fixed in 8 % ammonium molybdate and mounted inflattened strips.
In preparing the bladder for physiological experiments the animal was killed and openedon the ventral mid-line and the pubic symphysis was split. The ureters and, in the male, thevasa deferentia were severed close to the bladder wall. The vesical blood vessels and attachednerves were freed from the mesentery and sectioned near their origins from the common iliacs.The urethral blood vessels, together with the pelvic nerves, were tied in situ with a loop ofcotton, and then sectioned near the origin of the vessels from the common iliacs. The urethrawas then severed near the orifice and removed with the bladder and the nerve supply stillattached.
The bladder was tied to a Perspex holder with a cotton looped through the base of thebladder fundus. The holder was then immersed in modified Krebs's solution (Biilbring,1953) in an organ bath. A gas mixture containing 95 % Oa+ 5 % CO2 was bubbled through thesolution in the bath. The bath temperature was maintained at 30 ± 0-5° C. The nerves to bestimulated were looped over two platinum electrodes embedded in the Perspex holder. Inmost experiments both sets of nerves were stimulated, but in some experiments the nerve
S7-2

422 G . BURNSTOCK AND G. CAMPBELL
supplies were stimulated separately. A Grass stimulator was used to deliver square-wavepulses of variable frequency, duration and strength. Recording was either isometric (with aGrass force-displacement transducer recording with a Grass polygraph) or isotonic (with afrontal-writing lever recording on a smoked kymograph drum).
RESULTS
Morphology of the bladder
The bladder of the ringtail opossum lies above the pelvic cavity, and is borne on along urethra (Fig. i). The ureters and the vasa deferentia or the lateral vaginae enterthe bladder at the urethro-vesical junction. An artery and vein, arising from the
Ureter
Bladder
Urethra
Vu deferent
Dorsal aorta
Common lilacarteryBladder arteryand veslcal nerveUrethra! arteryPelvic nerve
Caudal artery
Fig. i. Diagram of the anatomy of the bladder of a male possum. There is a dual innervation ofthe bladder: anterior (vesical) nerves running intimately with the bladder arteries and posteriorpelvic nerves which run parallel to, but separate from, the urethral arteries.
common iliacs, pass up each antero-lateral face of the bladder, supplying the bulk ofthe bladder fundus. The urethra is also supplied with blood vessels from the iliacs,branches of these vessels extending on to the bladder neck and the lower part of thefundus. For convenience these vessels have been termed 'bladder' and 'urethral'respectively.
The bladder vessels are accompanied by nerves to the bladder fundus ('vesicalnerves'). Lying just caudal to the urethral vessels are the pelvic nerves which alsosupply the bladder with fibres. The pelvic nerves appear to arise from the second andthird sacral nerves, but it is not clear whether the vesical nerves are branches of thepelvic nerves or whether they have an independent origin. Ganglia can be demon-strated along the trunks of the vesical nerves and a large accumulation of ganglia is alsofound in the bladder neck.
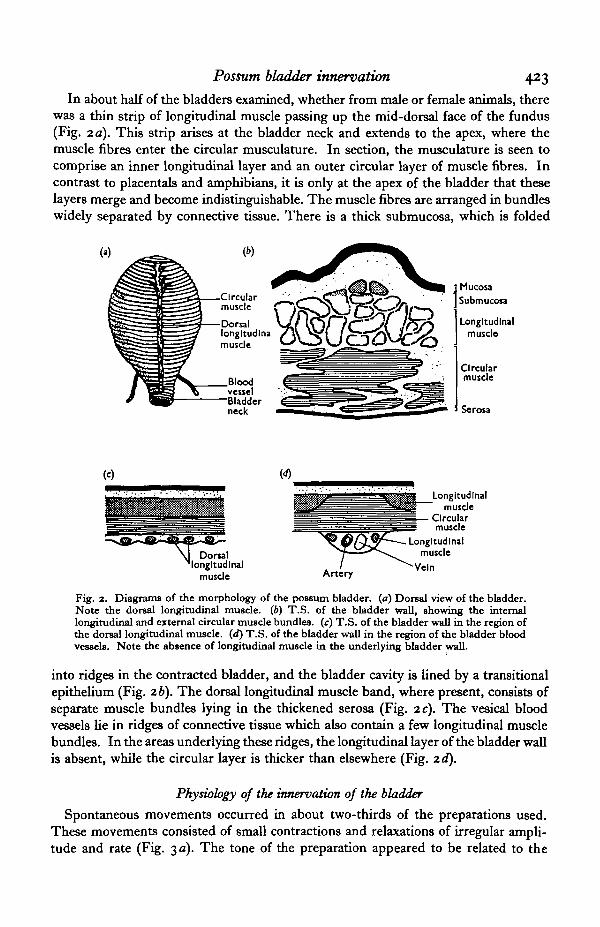
Possum bladder innervation 423
In about half of the bladders examined, whether from male or female animals, therewas a thin strip of longitudinal muscle passing up the mid-dorsal face of the fundus(Fig. 2 a). This strip arises at the bladder neck and extends to the apex, where themuscle fibres enter the circular musculature. In section, the musculature is seen tocomprise an inner longitudinal layer and an outer circular layer of muscle fibres. Incontrast to placentals and amphibians, it is only at the apex of the bladder that theselayers merge and become indistinguishable. The muscle fibres are arranged in bundleswidely separated by connective tissue. There is a thick submucosa, which is folded
Circularmuscle
Dorsallongltudlmmuscle
BloodvesselBladderneck
MucosaSubmucosa
Longitudinalmuscle
Circularmuscle
Serosa
Dorsallongitudinal
muscle
Longitudinalmuscle
Circularmuscle
Artery
Longitudinalmuscle
Vein
Fig. 2. Diagrams of the morphology of the possum bladder, (a) Dorsal view of the bladder.Note the dorsal longitudinal muscle. (6) T.S. of the bladder wall, showing the internallongitudinal and external circular muscle bundles, (c) T.S. of the bladder wall in the region ofthe dorsal longitudinal muscle, (d) T.S. of the bladder wall in the region of the bladder bloodvessels. Note the absence of longitudinal muscle in the underlying bladder wall.
into ridges in the contracted bladder, and the bladder cavity is lined by a transitionalepithelium (Fig. 2b). The dorsal longitudinal muscle band, where present, consists ofseparate muscle bundles lying in the thickened serosa (Fig. 2 c). The vesical bloodvessels lie in ridges of connective tissue which also contain a few longitudinal musclebundles. In the areas underlying these ridges, the longitudinal layer of the bladder wallis absent, while the circular layer is thicker than elsewhere (Fig. 2d).
Physiology of the innervation of the bladder
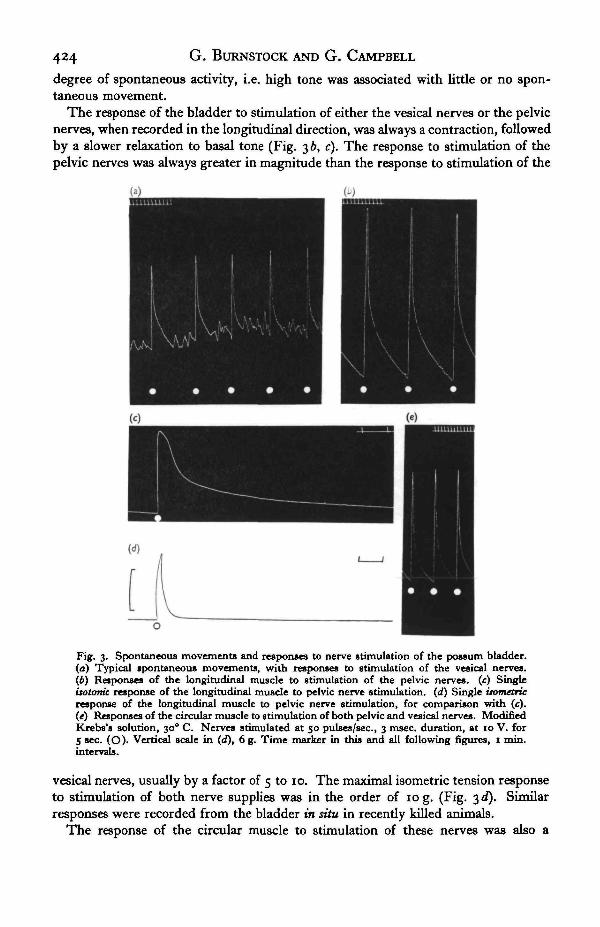
Spontaneous movements occurred in about two-thirds of the preparations used.These movements consisted of small contractions and relaxations of irregular ampli-tude and rate (Fig. 3 a). The tone of the preparation appeared to be related to the
424 G. BURNSTOCK AND G. CAMPBELL
degree of spontaneous activity, i.e. high tone was associated with little or no spon-taneous movement.
The response of the bladder to stimulation of either the vesical nerve3 or the pelvicnerves, when recorded in the longitudinal direction, was always a contraction, followedby a slower relaxation to basal tone (Fig. 3 b, c). The response to stimulation of thepelvic nerves was always greater in magnitude than the response to stimulation of the
Fig. 3. Spontaneous movements and responses to nerve stimulation of the possum bladder.(a) Typical spontaneous movements, with responses to stimulation of the vesical nerves,(ft) Responses of the longitudinal muscle to stimulation of the pelvic nerves, (c) SingleUotonit response of the longitudinal muscle to pelvic nerve stimulation, (d) Single isometricresponse of the longitudinal muscle to pelvic nerve stimulation, for comparison with (c).(e) Responses of the circular muscle to stimulation of both pelvic and vesical nerves. ModifiedKrebs's solution, 30° C. Nerves stimulated at 50 pulses/sec., 3 msec, duration, at 10 V. for5 sec. (O). Vertical scale in (d), 6 g. Time marker in this and all following figures, 1 min.intervals.
vesical nerves, usually by a factor of 5 to 10. The maximal isometric tension responseto stimulation of both nerve supplies was in the order of 10g. (Fig. 3d). Similarresponses were recorded from the bladder in situ in recently killed animals.
The response of the circular muscle to stimulation of these nerves was also a
Possum bladder innervation 425
Rntraction (Fig. 3 e). In some experiments the sympathetic chains were stimulatedat various points in situ, but no effect on the bladder wa9 ever seen.
Variations in tone of the preparation did not cause qualitative changes in theresponse to nerve stimulation. At high tone, however, the size of the response wassmaller than at low tone.
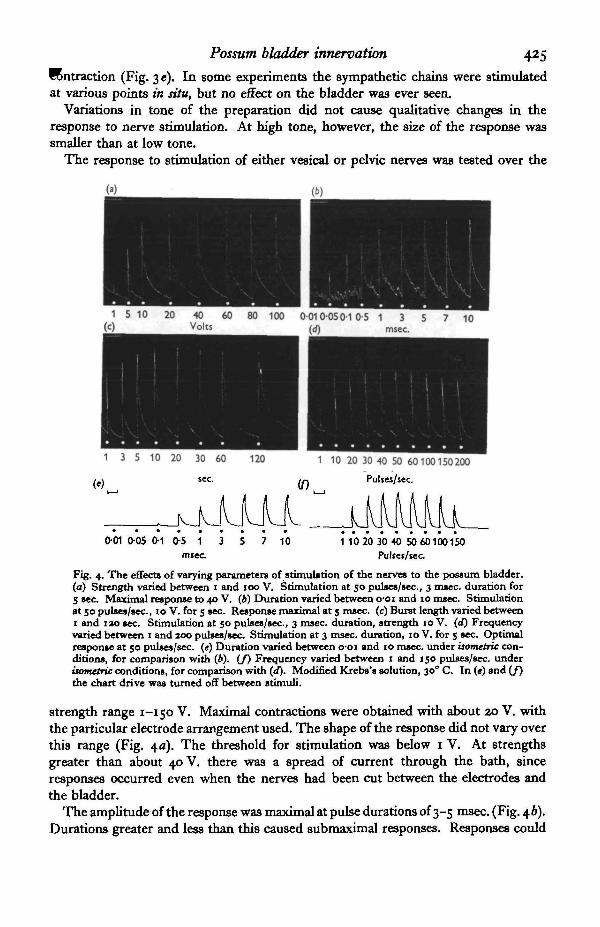
The response to stimulation of either vesical or pelvic nerves was tested over the
1 5 10 20 40 60 80 100 0010-05 0-1 OS 1 3 5V o l t s (d) msec.
10
1 10 20 30 40 50 60100150200
0-01 0-05 0 - 1 0 - 5 1 3
msec.7 10 110 20 30 40 50 60100150
Pulsei/iec
Fig. 4. The effects of varying parameters of stimulation of the nerves to the possum bladder,(a) Strength varied between i and IOO V. Stimulation at 50 pulses/sec., 3 msec, duration for5 sec Maximal response to 40 V. (6) Duration varied between o-oi and 10 msec. Stimulationat 50 pulses/sec., 10 V. for 5 sec. Response maximal at 5 msec, (c) Burst length varied between1 and iao sec. Stimulation at 50 pulses/sec., 3 msec, duration, strength 10 V. (d) Frequencyvaried between 1 and 200 pulses/sec. Stimulation at 3 msec, duration, 10 V. for 5 sec. Optimalresponse at 50 pulses/sec. («) Duration varied between o-oi and 10 msec, under isortietric con-ditions, for comparison with (b). (/) Frequency varied between 1 and 150 pulses/sec, underisometric conditions, for comparison with (d). Modified Krebs's solution, 300 C. In («) and (/)the chart drive was turned off between stimuli.
strength range 1-150 V. Maximal contractions were obtained with about 20 V. withthe particular electrode arrangement used. The shape of the response did not vary overthis range (Fig. 4 a). The threshold for stimulation was below 1 V. At strengthsgreater than about 40 V. there was a spread of current through the bath, sinceresponses occurred even when the nerves had been cut between the electrodes andthe bladder.
The amplitude of the response was maximal at pulse durations of 3-5 msec. (Fig. \b).Durations greater and less than this caused submaximal responses. Responses could
426 G. BURNSTOCK AND G. CAMPBELL
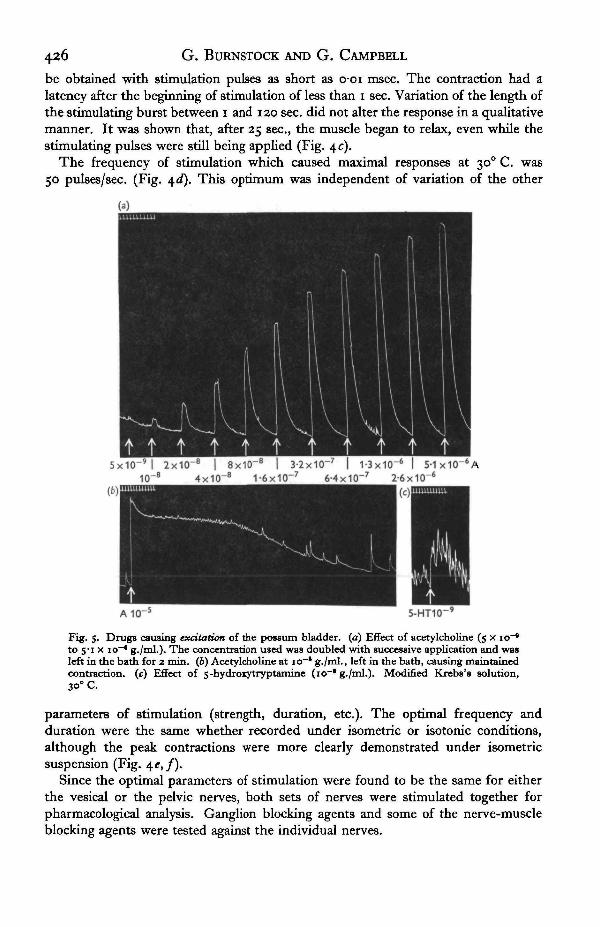
be obtained with stimulation pulses as short as o-oi msec. The contraction had alatency after the beginning of stimulation of less than i sec. Variation of the length ofthe stimulating burst between i and 120 sec. did not alter the response in a qualitativemanner. It was shown that, after 25 sec., the muscle began to relax, even while thestimulating pulses were still being applied (Fig. 4 c).
The frequency of stimulation which caused maximal responses at 300 C. was50 pulses/sec. (Fig. \d). This optimum was independent of variation of the other
5x10"'I 2x10"8 | 8x1(T8 I 3-2x1(T7 I 1-3x10"6 I 5-1 x10"6/1(T8 4x1(r8 1-6x10~7 6-4x10~7 26x10"6
A 10 5-HT10"
Fig. 5. Drugs causing excitation of the possum bladder, (a) Effect of acetylcholine (5 x io~*to 5"i x io~* g./ml.). The concentration used was doubled with successive application and wasleft in the bath for 2 min. (6) Acetylcholine at io~* g./ml., left in the bath, causing maintainedcontraction, (c) Effect of 5-hydroxytryptamine (io~f g./ml.). Modified Krebs's solution,30° C.
parameters of stimulation (strength, duration, etc.). The optimal frequency andduration were the same whether recorded under isometric or isotonic conditions,although the peak contractions were more clearly demonstrated under isometricsuspension (Fig. \e,f).
Since the optimal parameters of stimulation were found to be the same for eitherthe vesical or the pelvic nerves, both sets of nerves were stimulated together forpharmacological analysis. Ganglion blocking agents and some of the nerve-muscleblocking agents were tested against the individual nerves.
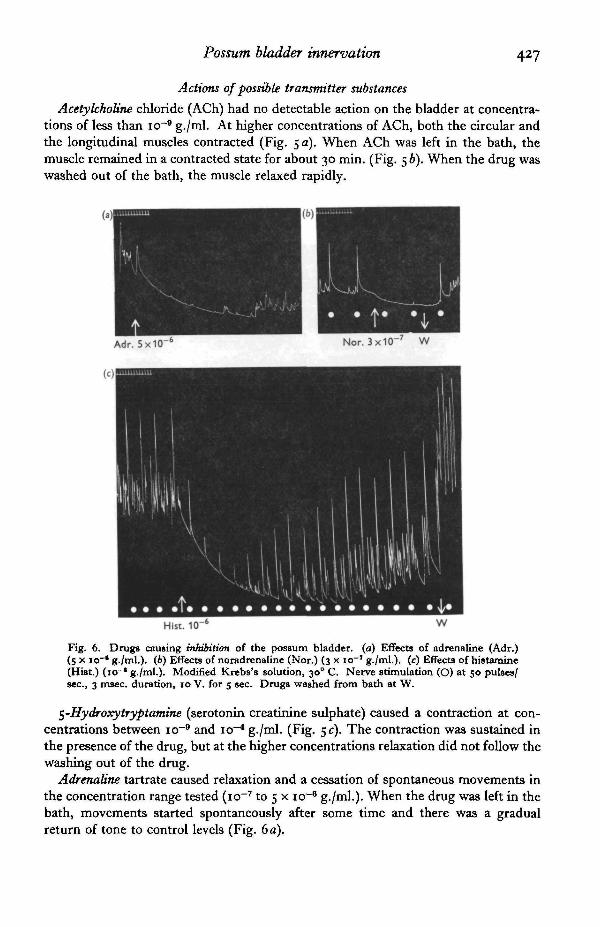
Possum bladder innervation 427
Actions of possible transmitter substances
Acetylcholine chloride (ACh) had no detectable action on the bladder at concentra-tions of less than io~9 g./ml. At higher concentrations of ACh, both the circular andthe longitudinal muscles contracted (Fig. 5 a). When ACh was left in the bath, themuscle remained in a contracted state for about 30 min. (Fig. 56). When the drug waswashed out of the bath, the muscle relaxed rapidly.
Hist. 10
Fig. 6. Drugs causing inhibition of the possum bladder, (a) Effects of adrenaline (Adr.)(5 x io"' g./ml.). (6) Effects of noradrenaline (Nor.) (3 x io~7 g./ml.). (c) Effects of histamine(Hist.) (io~* g./ml.). Modified Krebs's solution, 300 C. Nerve stimulation (O) at 50 pulses/sec., 3 msec, duration, 10 V. for 5 sec. Drugs washed from bath at W.
5-Hydroxytryptatnine (serotonin creatinine sulphate) caused a contraction at con-centrations between io~9 and icr4 g./ml. (Fig. 5 c). The contraction was sustained inthe presence of the drug, but at the higher concentrations relaxation did not follow thewashing out of the drug.
Adrenaline tartrate caused relaxation and a cessation of spontaneous movements inthe concentration range tested (icr7 to 5 x icr6 g./ml.). When the drug was left in thebath, movements started spontaneously after some time and there was a gradualreturn of tone to control levels (Fig. 6a).
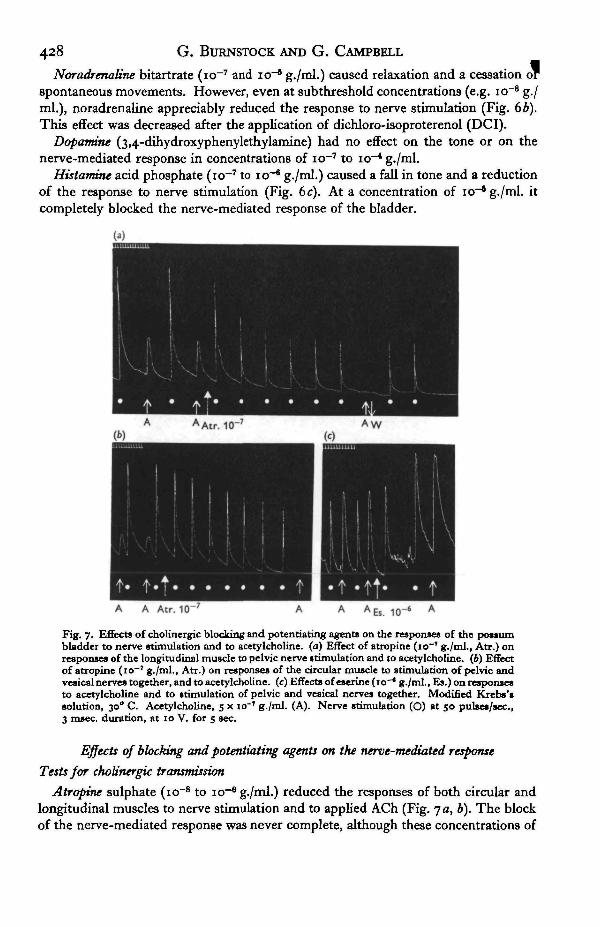
428 G. BlJRNSTOCK AND G. CAMPBELL
Noradrenatine bitartrate (io~7 and io"6 g./ml.) caused relaxation and a cessation orspontaneous movements. However, even at subthreshold concentrations (e.g. io~* g./ml.), noradrenaline appreciably reduced the response to nerve stimulation (Fig. 6b).This effect was decreased after the application of dichloro-isoproterenol (DCI).
Dopamne (3,4-dihydroxyphenylethylamine) had no effect on the tone or on thenerve-mediated response in concentrations of io~7 to io~* g./ml.
Histamme acid phosphate (io~7 to icr6 g./ml.) caused a fall in tone and a reductionof the response to nerve stimulation (Fig. 6c). At a concentration of io"8 g./ml. itcompletely blocked the nerve-mediated response of the bladder.
10-Fig. 7. Effects of cholinergic blocking and potentiating agents on the responses of the possumbladder to nerve stimulation and to acetylcholine. (a) Effect of atropine (io~T g./ml., Atr.) onresponses of the longitudinal muscle to pelvic nerve stimulation and to acetylcholine. (6) Effectof atropine (io~' g./ml., Atr.) on responses of the circular muscle to stimulation of pelvic andvesical nerves together, and to acetylcholine. (c) Effects of eserine (io~* g./ml., Es.) on responsesto acetylcholine and to stimulation of pelvic and vesical nerves together. Modified Krebs'ssolution, 300 C. Acetylcholine, 5 x io~' g./ml. (A). Nerve stimulation (O) at 50 pulses/sec.,3 msec, duration, at 10 V. for 5 sec.
Effects of blocking and potentiating agents on the nerve-mediated response
Tests for cholinergic transmission
Atropine sulphate (io"8 to IO"6 g./ml.) reduced the responses of both circular andlongitudinal muscles to nerve stimulation and to applied ACh (Fig. ja, b). The blockof the nerve-mediated response was never complete, although these concentrations of
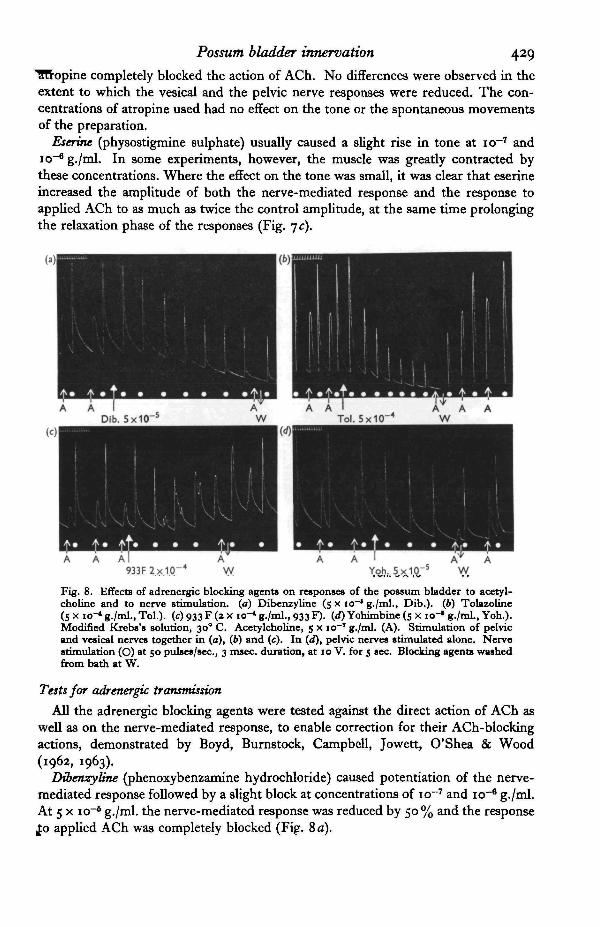
Possum bladder irmervation 429IHTopine completely blocked the action of ACh. No differences were observed in theextent to which the vesical and the pelvic nerve responses were reduced. The con-centrations of atropine used had no effect on the tone or the spontaneous movementsof the preparation.
Eserine (physostigmine sulphate) usually caused a slight rise in tone at io~7 andio"6 g./ml. In some experiments, however, the muscle was greatly contracted bythese concentrations. Where the effect on the tone was small, it was clear that eserineincreased the amplitude of both the nerve-mediated response and the response toapplied ACh to as much as twice the control amplitude, at the same time prolongingthe relaxation phase of the responses (Fig. jc).
93_3_F2x10 Yoh,. 5 xJ.IT W
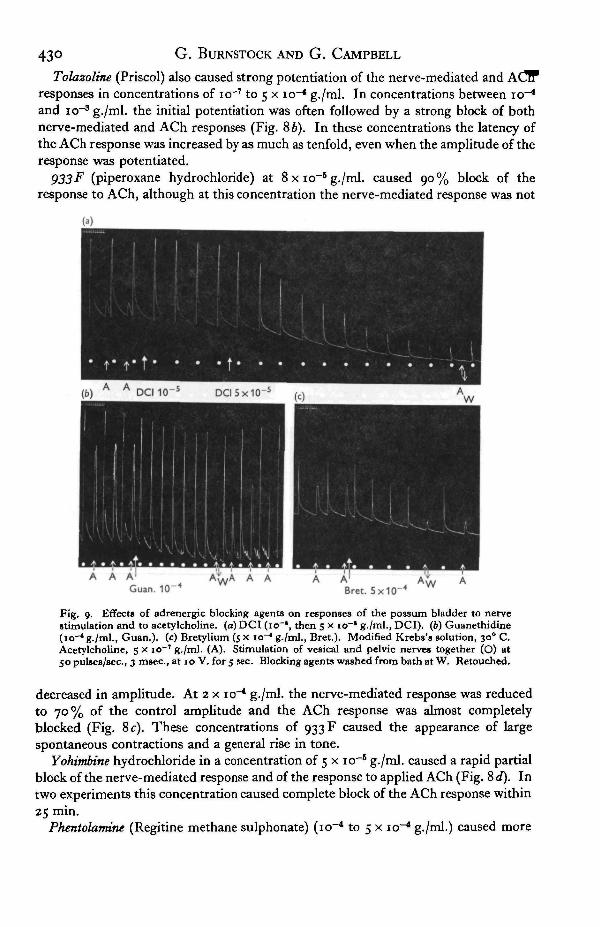
Fig. 8. EflfectB of adrenergic blocking agents on responses of the possum bladder to acetyl-choline and to nerve stimulation, (a) Dibenzyline (5 x io~* g./ml., Dib.). (6) Tolazoline(5 x 10-* g./ml., Tol.). (c) 933 F (a x 10-* g./ml., 933 F). (</) Yohimbine (5 x io"1 g./mL, Yoh.).Modified Krebs's solution, 300 C. Acetylcholine, 5 x io~T g./ml. (A). Stimulation of pelvicand vesical nerves together in (a), (6) and (c). In (d), pelvic nerves stimulated alone. Nervestimulation (O) at 50 pul«es/sec., 3 msec, duration, at 10 V. for 5 sec. Blocking agents washedfrom bath at W.
Tests for adrenergic transmission
All the adrenergic blocking agents were tested against the direct action of ACh aswell as on the nerve-mediated response, to enable correction for their ACh-blockingactions, demonstrated by Boyd, Burnstock, Campbell, Jowett, O'Shea & Wood(1962, 1963).
Dibenzyline (phenoxybenzamine hydrochloride) caused potentiation of the nerve-mediated response followed by a slight block at concentrations of io~7 and io~* g./ml.At 5 x io~* g./ml. the nerve-mediated response was reduced by 50 % and the response£0 applied ACh was completely blocked (Fig. 8 a).
43° G. BURNSTOCK AND G. CAMPBELL
Tolazoline (Priscol) also caused strong potentiation of the nerve-mediated and ACTFresponses in concentrations of io~7 to 5 x io~* g./ml. In concentrations between io^1
and icr3 g./ml. the initial potentiation was often followed by a strong block of bothnerve-mediated and ACh responses (Fig. 86). In these concentrations the latency ofthe ACh response was increased by as much as tenfold, even when the amplitude of theresponse was potentiated.
933F (piperoxane hydrochloride) at 8 x io~B g./ml. caused 90% block of theresponse to ACh, although at this concentration the nerve-mediated response was not
Bret. 5x10"
Fig. 9. Effects of adrenergic blocking agents on responses of the possum bladder to nervestimulation and to acetylcholine. (a) DCI (io~*, then 5 x io~' g./ml., DCI). (b) Guanethidine(io~* g./ml., Guan.). (c) Bretylium (5 x io~* g./ml., Bret.). Modified Krebs's solution, 300 C.Acetylcholine, 5 x io~? g./ml. (A). Stimulation of vesical and pelvic nerves together (O) at50 pulses/sec, 3 msec., at 10 V. for 5 sec. Blocking agents washed from bath at W. Retouched.
decreased in amplitude. At 2 x io~* g./ml. the nerve-mediated response was reducedto 70% of the control amplitude and the ACh response was almost completelyblocked (Fig. 8c). These concentrations of 933 F caused the appearance of largespontaneous contractions and a general rise in tone.
Yohimbine hydrochloride in a concentration of 5 x io~B g./ml. caused a rapid partialblock of the nerve-mediated response and of the response to applied ACh (Fig. 8 d). Intwo experiments this concentration caused complete block of the ACh response within25 min.
Phentolanrine (Regitine methane sulphonate) (io"4 to 5 x io~* g./ml.) caused more

Possum bladder innervation 431than 50% reduction of the nerve-mediated response and completely blocked theresponse to ACh.
Ergotamine tartrate (2 x io~5 g./ml.) caused a slow partial block of both nerve-mediated and ACh responses. DCI caused a stronger block of the nerve-mediatedresponse and an almost complete block of the ACh response at concentrations of io~5
and 5 x 1 o""5 g./ml. (Fig. 9 a).Guanethidine (5 x io~5 g./ml.) rarely had any effect on the nerve-mediated response,
but at io"4 g./ml. it reduced the response to ACh without affecting the nerve-mediatedresponse (Fig. 96).
A A A Mec. 5x10-5
Fig. 10. Effects of ganglionic blocking agents on responses of the possum bladder to nervestimulation and to acetylcholine. (a) Hexamethonium (5 x io~* g./ml., C6). (b) Pentolinium(io~5 g./ml., Pent.), (c) Mecamylamine (5 x io"6 g./ml., Mec). Modified Krebs's solution,300 C. Acetylcholine 5 x io~' g./ml. (A). Stimulation of the pelvic nerves alone (O) at50 pulses/sec, 3 msec, at 10 V. for 5 sec. Blocking agents washed from bath at W.
Bretylium tosylate (Darenthin) (icr4 g./ml.) caused a slight potentiation of bothnerve-mediated and ACh responses, but at 5 x io~* g./ml. it caused a partial block ofthe nerve-mediated response and a stronger block of the ACh response (Fig. 9 c).

432 G. BURNSTOCK AND G. CAMPBELL
Tests for ganglia
Hexameihonium bromide (5 x io""5 to 5 x icr4 g./ml.) caused potentiation of theresponse to ACh, at the same time reducing the response to either pelvic or vesicalnerve stimulation by 10-20% (Fig. 10a). In some cases these concentrations causedpotentiation of the nerve-mediated response. At io"3 g./ml. the nerve-mediatedresponse was reduced by more than 20%, but the response to applied ACh was alsoreduced.
Pentotinium tartrate, at io"6 to io"6 g./ml., had no effect on the response to appliedACh, but at 10-4 g./ml. reduced this response to some extent. At io"8 g./ml. theresponse to stimulation of either the vesical or the pelvic nerves was reduced by about20-25 % (Fig. 10ft).
Mecamylamine hydrochloride potentiated the response to ACh in concentrationsbetween io"6 and 5x10-° g./ml., while at icr* g./ml. (and, in some cases, at 5 x io~* g./ml.) it caused large spontaneous contractions which obscured the effect of the drug onthe ACh responses. At 5 x io~* g./ml. this drug blocked the vesical and pelvic nerve-mediated responses by 30-40% (Fig. 10c). In contrast with pentolinium and hexa-methonium, it was difficult to decrease the block after mecamylamine by washing thedrug out of the bath.
DISCUSSION
The nerve fibres supplying the bladder of the possum are excitatory. These fibresappear to be cholinergic. A large proportion of the nerve fibres reaching the bladdervia either the pelvic nerves or the vesical nerves appear to be postganglionic. There is asyet no evidence for the presence of adrenergic fibres. Supporting evidence for theseconclusions is as follows:
(a) The nerve-mediated response is always a contraction. An inhibitory responsecould not be revealed by any variation of the parameters of the stimulating pulses, norby treatment with any of the drugs used. Stimulation of the sympathetic chains in situhad no effect on the bladder.
(6) The nerve-mediated response is mimicked by the action of ACh, but not by theactions of adrenaline, noradrenaline or dopamine.
(c) The nerve-mediated response is reduced by atropine and potentiated byeserine, as is the response to ACh.
(d) The adrenergic blocking agents tested affect the nerve-mediated response onlywhen they also affect the ACh response in a similar manner.
(e) Ganglia have been demonstrated along the trunks of the vesical nerves andaggregated at the base of the bladder.
(/) Ganglion blocking agents, at concentrations which do not reduce the response toACh, block the response to stimulation of either pelvic or vesical nerves by up to 40 %.
(g) There has been no anatomical demonstration of any nerve supply apart fromthose fibres which were stimulated in the above experiments.
The bladders of placental mammals, reptiles and amphibians are all outgrowths ofthe alimentary canal (Nelsen, 1953). Some parts of the bladder neck of placentals andreptiles are derived from the mesodermal tissues of the ureters. Buchanan & Fraser(1918) demonstrated a similar dual mode of origin for the marsupial bladder. In

Possum bladder innervation 433
amphibians it seems unlikely that there should be any contribution of ureteral tissues tothe bladder, since the ureters do not at any stage enter the bladder directly. In teleostand ganoid fish, however, the bladders are simply the swollen ends of the ureters.There is no bladder as such in elasmobranchs and birds.
In placental mammals the pelvic nerves to the bladder are excitatory and cholinergic(Elliott, 1907; Henderson & Roepke, 1934; Ursillo & Clark, 1956; Gyermek, 1961).The cat and the dog are the only placentals in which the sympathetic influence on thebladder has been studied in detail. Many workers have described an excitation followedby inhibition resulting from stimulation of the hypogastric nerve in these animals(Mellanby & Pratt, 1939, 1940; Kuntz & Saccomanno, 1944). The results reported byEdge (1955) suggest that both the contraction and the inhibition of the cat bladderresulting from hypogastric stimulation are mediated by adrenergic nerves. Elliott(1907) studied a series of placentals and concluded that there may be excitatory orinhibitory effects or no effects at all of the hypogastric nerve on the bladder muscula-ture of different species, even within one family, but this has not been confirmed.Similarly, the excitatory nerves to the toad bladder are cholinergic (Burnstock, O'Shea& Wood, 1963). On the other hand, the lizard bladder appears to be innervated by amixture of excitatory cholinergic and both excitatory and inhibitory adrenergic fibres(Burnstock, Wood & O'Shea, 1961). The innervation of the teleost bladder is excita-tory, but the nature of the transmitter used is unknown (Young, 1936).
Thus it would appear that excitatory cholinergic nerve fibres to the urinary bladderare present throughout the vertebrates. On the other hand, adrenergic nerve fibres,where present, exert inhibitory, and possibly also excitatory, influences on the bladderfundus. There is as yet no indication of any evolutionary trend in the presence orabsence of adrenergic innervation.
During the course of this work, several features of general pharmacological interesthave been noted.
Many of the drugs which reduce the nerve-mediated responses of the possumbladder block the ACh response to a greater extent. This effect is particularly markedwith atropine, and seems to be similar to the atropine-resistance found in manyplacental mammal preparations, and in particular in thebladder (Ursillo, 1961). Ursillo& Clark (1956) showed that the time taken for atropine to cause 50% block ofthe nerve-mediated response of the rabbit bladder strip/pelvic nerve preparation waseight times longer than the time taken to block the ACh response to the same extent.In common with the placental bladder, the marsupial bladder cannot be completelyblocked with atropine. In fact, Henderson & Roepke (1934) took this as evidence for anon-cholinergic process in transmission, but Gyermek (1961) found no evidence tosupport this view.
The anti-ACh action of the pre- and post-synaptic adrenergic blocking agents usedhas also been noted in preparations from other animals, and has been made the subjectof a more detailed study (Boyd et al. 1962, 1963). The apparent paradox of the possumbladder being blocked by both cholinergic and adrenergic blocking agents is thusexplained by the atropine-like action of the anti-adrenergic agents. Similarly, thepotentiation of ACh and nerve responses by some of these agents is explicable in termsof their anti-cholinesterase actions, already described for some of these drugs by Boyd,Chang & Rand (i960).

434 G. BURNSTOCK AND G. CAMPBELL
The relaxation of the possum bladder by histamine (io~7 to io~* g./ml.) is also o¥pharmacological interest, since very few visceral smooth muscles have been shown tobe relaxed by histamine. Gyermek (1961) has shown that the bladders of cats and dogsare contracted in vivo by histamine. The inhibition of the nerve response by histamineand also by adrenaline and noradrenaline is most probably due to a general depressionof the excitability of the muscle.
When ganglion-blocking agents are used in pharmacological analysis it is necessaryto check the action of the blocking agents against the response to directly appliedterminal transmitter. If the concentration of blocking agent used reduces the responseto the terminal transmitter, the amount of block of the nerve response is not a reliableindication of the degree of ganglionic blockade, e.g. on the possum bladder, hexa-methonium and pentolinium cannot be used at concentrations greater than 5 x io~*and io"8 g./ml. respectively. Similarly, the degree of ganglionic blockade is probablymore than would be indicated by the depression of the response if the direct responseof the muscle to applied transmitter is potentiated by the ganglionic blocking agent,e.g. hexamethonium at concentrations less than 5 x io~* g./ml. and mecamylamine atconcentrations between io"8 and 5 x io~* g./ml. potentiate the response to ACh. Anadditional problem is presented by the action of mecamylamine (io~* g./ml.) inincreasing the spontaneous movements to such an extent that responses can no longerbe distinguished clearly. The potentiation of excitatory responses and increase inexcitability produced by hexamethonium and mecamylamine on the possum bladderis not easily explained. Zauder (1954) and Biilbring (1955) reported similar effects ofhexamethonium on placental mammal preparations, and suggested that they were dueto a general increase in excitability of the muscles. However, Sjostrand (1962) showedthat, although hexamethonium potentiates the excitatory responses of the guinea-pigvas deferens to noradrenaline, it reduces responses to ACh. Another explanationmight be that these agents have an anti-cholinesterase action, but Paton & Zaimis(1949) have reported that hexamethonium has little, if any, anti-cholinesteraseproperty.
The ACh-blocking action of hexamethonium and pentolinium on the possumbladder is similar to the atropine-like actions of these drugs on some placental mammalpreparations (Feldberg, 1951; Mason & Wien, 1955). In contrast, Stone, Torchiana,Navarro & Beyer (1956) reported that mecamylamine had no atropine-like action at2 x io~* g./ml., a result which was also true for the possum bladder.
If the amount of blockade of the nerve response produced by these three ganglionicblocking agents is taken as a measure of potency, the drugs may be arranged in thefollowing order of increasing potency: hexamethonium:pentolinium:mecamylamine.However, in the light of the above discussion, the factor of specificity should also beincluded in any true index of potency.
It would seem that, in the absence of any truly specific blocking agents, the use ofanalytical pharmacology for the determination of the transmitter substance involved inan autonomic nerve/smooth muscle preparation is a difficult and, in some cases, incon-clusive process. It is advisable to use such an analysis in combination with a quantita-tive biochemical assay of the perfusates from organs after stimulation of the nervesupply.

Possum bladder innervation 435
SUMMARY
1. A histological study of the structure of the urinary bladder of the ringtailpossum has been made. The innervation of the bladder has been studied in vitro,using the technique of analytical pharmacology.
2. The bladder has well-defined inner longitudinal and outer circular muscle layers.Nerves supplying the bladder are found both in the pelvic nerves and in the vesicalnerves which run with the vascular supply of the bladder fundus. Ganglia have beendemonstrated along the trunks of the vesical nerves and also aggregated at the bladderneck.
3. The response of the bladder to stimulation of either nerve supply in situ or invitro is always a simultaneous contraction of both longitudinal and circular muscles.Inhibitory responses to nerve stimulation have never been observed. The optimalfrequency for stimulation of these nerves at 300 C. is 50 pulses/sec.
4. The bladder is contracted by ACh and 5-hydroxytryptamine, but is relaxed byadrenaline, noradrenaline and histamine.
5. The response to nerve stimulation is reduced by atropine and potentiated byeserine. Adrenergic blocking agents do not affect the nerve-mediated response unlessthey also affect the response to applied ACh in a similar manner.
6. Ganglionic blocking agents, in concentrations which do not reduce the responseto ACh, cause up to a 40 % reduction of the response to stimulation of either thevesical or the pelvic nerves.
7. It is concluded that the nerve fibres supplying the possum bladder are cholin-ergic, perhaps 40 % of them being stimulated pre-ganglionically.
8. The evolutionary significance of these observations is discussed.9. Some points of pharmacological interest have been discussed in relation to drug
actions on placental mammal preparations.
This work was supported by a P.H.S. research grant (B-2902) from the NationalInstitute of Neurological Diseases and Blindness, Public Health Service (U.S.A.) andby the National Health and Medical Research Council (Aust.).
We would like to express our thanks to the following pharmaceutical firms for theirkind donations of drugs: CIBA, Basle; May and Baker, Dagenham; Merck, Sharp andDohme; Smith, Kline and French; Wellcome Research Laboratories. We would alsolike to thank Mr D. Rogers for his valuable assistance with histological material.
REFERENCES
BOYD, H., BURNSTOCK, G., CAMPBELL, G., JOWETT, A., O 'SHEA, J. & WOOD, M. (1962). An investiga-tion of the use of adrenergic blocking agents as analytical tools. Aust. J. Sci. 25, 107.
BOYD, H., BURNSTOCK, G., CAMPBELL, G., JOWETT, A., O 'SHEA, J. & WOOD, M. (1963). The cholinergicblocking action of adrenergic blocking agents in pharmacological analysis of autonomic innervation.Brit. J. Pharmacol, (in the Press).
BOYD, H., CHANG, V. & RAND, M. J. (i960). The anticholinesterase activity of some anti-adrenalineagents. Brit. J. Pharmacol. 15, 525-31.
BUCHANAN, G. & FRASER, E. A. (1918). The development of the urinogenital system in the Marsupialia,with specific reference to Trichosurus vulpccula. J. Anat., Land., 53, 35-96.
BOLBRING, E. (1953). Measurements of oxygen consumption in smooth muscle. J. Physiol. 12a, 111—34.BOLBRING, E. (1955). Correlations between membrane potential, spike discharge and tension in
smooth muscle. J. Physiol. ia8, 200-21.28 Exp. BioL 40, 3

436 G. BURNSTOCK AND G. CAMPBELL
BURNSTOCK, G., O'SHEA, J. & WOOD, M. (1963). Comparative physiology of the vertebrate autonormc"nervous system. I. Innervation of the urinary bladder of the toad (Bufo marimu). J. Exp. Biol. 40,403-19-
BURNSTOCK, G., WOOD, M. & O'SHEA, J. (1961). Studies in the comparative physiology of the vertebrateautonomic nervous system. Aust. J. Set. 24, 192.
CAMPBELL, G. & BURNSTOCK, G. (1962). The innervation of the marsupial bladder. Auit. J. Set. 25, 108.EDGE, N. D. (I 955). A contribution to the innervation of the urinary bladder of the cat. J. Pkyriol. 127,
53-68-ELLIOTT, T. R. (1907). The innervation of the bladder and urethra. J. Pkytiol. 35, 367-445.FBLDBBRG, W. (1951). Effects of ganglionic blocking substances on the small intestine. J. Pkytiol. 113,
8GRUBBR, C. M. (1933). The autonomic innervation of the genito-urinary system. Phytiol. Rev. 13, 497-
609.GYERMEE, L. (1961). Cholinergic stimulation and blockade on urinary bladder. Amer. J. Pkytiol. 201,
HENDERSON, V. E. & ROEPKE, M. H. (1934). The role of acetylcholine in bladder contractile mechanismsand in parasympathetic ganglia. J. Pharmacol. 51, 97— m .
HILLARP, N. A. (i960). In Handbook of Pkytiology-Neurophytiology, Vol. 2, pp. 979-1006. Washington:American Physiological Society.
KUNTZ, A. & SACCOMANNO, G. (1944). Sympathetic innervation of the detrusor muscles. J. Urol. 51,535-43-
LANOLEY, J. N. (1931). The Autonomic Nervous System, Part 1. Cambridge: Heffer and Sons.MCSWTNEY, B. A. (1931). Innervation of the stomach. Pkytiol. Rev. 11, 478-514.MASON, D. F. J. & WIEN, R. (1955). The actions of heterocyclic bisquaternary compounds, especially of
a pvrrolidinhim series. Brit.J. Pharmacol. 10, 124-32.MKLLANBY, J. & PRATT, C. L. G. (1939). The reactions of the urinary bladder of the cat under conditions
of constant pressure. Proc. Roy. Soc. B, 127, 307-22.MELLANBY, J. & PRATT, C. L. G. (1940). The reactions of the urinary bladder of the cat under conditions
of constant volume. Proc. Roy. Soc. B, 128, 186-201.NELSEN, O. E. (1953). Comparative Embryology of the Vertebrates, pp. 787—8. London: Constable.NICOL, J. A. C. (1952). Autonomic nervous systems in lower chordates. Biol. Rev. 27, 1-49.PATON, W. D. M. & ZAIMIS, E. J. (1949). The pharmacological actions of polymethylene bistrimethyl-
ammonium salts. Brit. J. Pharmacol. 4, 381-400.SJOSTRAND, N. O. (1962). Inhibition by ganglion blocking agents of the motor response of the isolated
guinea-pig vas deferens to hypogastric nerve stimulation. Ada pkytiol. tcand. 54, 306-15.STONE, C. A., TORCHIANA, M. L., NAVARRO, A. tc BEYER, K. H. (1956). Ganglionic blocking properties
of 3-methylamino-isocamphane hydrochloride (Mecamylamine); a secondary amine. J. Pharmacol.117, 169-83.
URSILLO, R. C. (1961). Investigation of certain aspects of atropine-resistant nerve effects. J. Pharmacol.131, 231-6.
URSIIXO, R. C. & CLARK, B. B. (1956). The action of atropine on the urinary bladder of the dog and onthe isolated nerve-bladder strip preparation of the rabbit. J. Pharmacol. 118, 338-47.
YOUNG, J. Z. (1936). The innervation and reaction to drugs of the viscera of teleostean fish. Proc. Roy.Soc. B, 120, 303-18.
ZAUDER, H. L. (1954). Potentiation of acetylcholine and histamine by heramethonium. Fed. Proc. 13,420.