combined heat shock protein 90 and ribosomal rna sequence phylogeny supports multiple replacements...
TRANSCRIPT

Combined Heat Shock Protein 90 and Ribosomal RNA Sequence PhylogenySupports Multiple Replacements of Dinoflagellate Plastids
KAMRAN SHALCHIAN-TABRIZI,a,b MARIANNE A. MINGE,a THOMAS CAVALIER-SMITH,b JOACHIM M. NEDREKLEPP,a
DAG KLAVENESSc and KJETILL S. JAKOBSENa
aDepartment of Biology, Centre for Ecological and Evolutionary Synthesis (CEES), University of Oslo, N-0316 Oslo, Norway andbDepartment of Zoology, University of Oxford, South Park Road, Oxford, OX1 3PS, United Kingdom
cDepartment of Biology, Program for Plankton Biology, University of Oslo N-0316036, Norway
ABSTRACT. Dinoflagellates harbour diverse plastids obtained from several algal groups, including haptophytes, diatoms, cryptophytes,and prasinophytes. Their major plastid type with the accessory pigment peridinin is found in the vast majority of photosynthetic species.Some species of dinoflagellates have other aberrantly pigmented plastids. We sequenced the nuclear small subunit (SSU) ribosomal RNA(rRNA) gene of the ‘‘green’’ dinoflagellate Gymnodinium chlorophorum and show that it is sister to Lepidodinium viride, indicating thattheir common ancestor obtained the prasinophyte (or other green alga) plastid in one event. As the placement of dinoflagellate species thatacquired green algal or haptophyte plastids is unclear from small and large subunit (LSU) rRNA trees, we tested the usefulness of the heatshock protein (Hsp) 90 gene for dinoflagellate phylogeny by sequencing it from four species with aberrant plastids (G. chlorophorum,Karlodinium micrum, Karenia brevis, and Karenia mikimotoi) plus Alexandrium tamarense, and constructing phylogenetic trees forHsp90 and rRNAs, separately and together. Analyses of the Hsp90 and concatenated data suggest an ancestral origin of the peridinin-containing plastid, and two independent replacements of the peridinin plastid soon after the early radiation of the dinoflagellates. Thus, theHsp90 gene seems to be a promising phylogenetic marker for dinoflagellate phylogeny.
Key Words. Bayes factor, covarion, dinoflagellate evolution, Hsp90, phylogeny, plastid replacement, rRNA.
I T is widely accepted that chloroplasts evolved from endosym-biotic cyanobacteria in a single evolutionary event (Cavalier-
Smith 2000). Such plastids are called primary plastids and arefound only in red algae, Viridiplantae and glaucophytes (together,the Plantae). All other algae gained their plastids from otherplastid-containing eukaryotes through secondary and tertiaryendosymbioses (Cavalier-Smith 2003; Palmer and Delwiche1998; Tengs et al. 2000).
Dinoflagellates harbour an exceptional diversity of plastidsobtained from several eukaryotic algal groups (Bj�rnland 1990;Chesnick et al. 1997; Delwiche 1999; Gibbs 1981; Jeffrey,Sielicki, and Haxo 1975; Tengs et al. 2000). Furthermore, abouthalf their species lack plastids, whereas an increasing number ofspecies has been reported as mixotrophic (Stoecker 1999). Dino-flagellates are thus an ideal group for investigating plastid evolu-tion (Schnepf 1993). The most abundant type of dinoflagellateplastid, often called the typical plastid, uniquely uses the xantho-phyll peridinin as the predominant accessory pigment and is en-veloped by three membranes (Bj�rnland and Liaaen-Jensen 1989;Whatley 1993).
Aberrantly pigmented dinoflagellates (i.e. using non-peridininpigments) often harbour plastids which are periodically lost andgained during the life of the host (Chesnick, Morden, and Schmieg1996; Dodge and Crawford 1969; Janson 2004; Takishita et al.2002). A few species, such as Dinophysis spp., Kryptoperidiniumfoliaceum, and Durinskia baltica (synonym Peridinium balticum),harbour reduced photosynthetic endosymbionts that are kept forlonger periods, but are probably not fully integrated organelles;the endosymbionts of the latter two species are least reduced, re-taining mitochondria, functional nuclei, and almost identical genesequences to free-living diatom populations (Chesnick, Morden,and Schmieg 1996; Dodge and Crawford 1969; Janson 2004;Kempton et al. 2002; McEwan and Keeling 2004; Takishita et al.2002). At present, only two examples of aberrantly pigmentedplastids are regarded as fully integrated with the dinoflagellatehost (Cavalier-Smith 2003). One of these, comprising severalKarlodinium, Karenia, and Takayama species (de Salas et al.
2003; Haywood et al. 2004), acquired their plastids fromhaptophytes, which are the only known stable and fully integrat-ed plastids gained through tertiary endosymbiosis: plastids trans-ferred serially from a cyanobacterium to a red alga, then to ahaptophyte and finally into a dinoflagellate (Bj�rnland 1990;Takishita, Nakano, and Uchida 2000; Tengs et al. 2000). The oth-er known type of stable dinoflagellate non-peridinin plastid is inGymnodinium chlorophorum and Lepidodinium viride. Pigmen-tation data suggest that this plastid probably is derived from aprasinophyte, although no sequence analysis has yet supportedthis hypothesis (Elbrachter and Schnepf 1996; Watanabe et al.1990, 1991). Ultrastructural similarity of the latter two greendinoflagellate host cells, such as the shape of the apical groove,suggest that they are closely related (Elbrachter and Schnepf1996; Honsell and Talarico 2004), but it has been unclear wheth-er their plastids were established in a common ancestor or wereacquired independently. We have therefore sequenced the nuclearsmall subunit (SSU) ribosomal RNA (rRNA) gene of G.chlorophorum and shown for the first time that it is closely relat-ed to Lepidodinium, implying that the prasinophyte plastid wasprobably acquired once only by their common ancestor.
Another key question concerning the evolution of theseaberrantly pigmented plastids is whether they were establishedin heterotrophic dinoflagellate lineages, or if the peridinin-containing plastid is ancestral and was replaced by the otherplastids (Hoek, Mann, and Jahns 1995; Leander and Keeling2003; Saldarriaga et al. 2001; Tengs et al. 2000). Molecularphylogenies have been used to resolve the evolution of the dino-flagellates and to address the position of the different plastid typesin the host phylogeny (Cavalier-Smith and Chao 2004; Edvardsenet al. 2003; Saldarriaga et al. 2001; Tengs et al. 2000). However,the nuclear small and large subunit rRNA genes (SSU and LSU,respectively) have incongruent tree topologies and weak statisticalsupport for the vast majority of the internal, basal branches(Cavalier-Smith and Chao 2004; Daugbjerg et al. 2000; Jakobsenet al. 2002; Murray et al. 2005; Saldarriaga et al. 2001; Saunderset al. 1997; Tengs et al. 2000). It is therefore difficult to excludeany hypothesis about the distribution of the different plastidsamong dinoflagellates. Recently, another gene, the 90-kDa cyto-solic heat shock protein (Hsp90), appears to be useful for phylo-genies of a wide range of eukaryotes, including alveolates anddeep branching dinoflagellates (Fast et al. 2002; Leander
Corresponding Author: Kjetill S. Jakobsen, Department of Biology,Centre for Ecological and Evolutionary Synthesis (CEES) University ofOslo, N-0316 Oslo, Norway—Telephone number: 147 22854602; FAXnumber: 147 22854726; e-mail: [email protected]
217
J. Eukaryot. Microbiol., 53(3), 2006 pp. 217–224r 2006 The Author(s)Journal compilation r 2006 by the International Society of ProtistologistsDOI: 10.1111/j.1550-7408.2006.00098.x

and Keeling 2004; Simpson, Lukes, and Roger 2002; Stechmannand Cavalier-Smith 2003). Hsp90 trees usually have relativelyhomogenous branch-lengths, in particular for species with unusu-ally rapidly evolving rRNA (Oxyrrhis marina) and deeply diverg-ing alveolates (Perkinsus marinus; Fast et al. 2002; Leander andKeeling 2004), and could be important for reducing long-branchproblems recently seen in dinoflagellate mitochondrial cyto-chrome b phylogenies (Zhang, Bhattacharya, and Lin 2005).As only a few Hsp90 sequences of dinoflagellate have beensequenced, we have extended the data by sequencing cytosolicHsp90 from G. chlorophorum, three species with haptophyte-derived plastids (Karlodinium micrum, Karenia brevis, andKarenia mikimotoi) and another with peridinin (Alexandriumtamarense). All trees reconstructed from the Hsp90 and concate-nated Hsp901SSU1LSU sequences were congruent, supportingtwo separate replacements of plastids in dinoflagellates, eithersubstituting the ancestral peridinin plastid in photosynthetic spe-cies or being established in heterotrophic species that already hadlost the peridinin plastids.
MATERIALS AND METHODS
Algal cultures and DNA isolation. The G. chlorophorum cul-ture was obtained from the Scandinavian Culture Centre for Algaeand Protozoa. DNA was isolated using DNA DIRECT (DynalBiotech AS, Oslo, Norway) as described previously (Rudi et al.1997) or the Bugs’n Beads ver U kit (Genpoint AS, Oslo, Norway)following the manufacturer’s instructions.
PCR and sequencing. The nuclear SSU rRNA gene from G.chlorophorum was amplified using a combination of dinoflagel-late-specific and universal primers. The 30-end was gained witha dinoflagellate matching primer 475FChl1: 50-CAT CCA TGTCTT GTA ATT G-30 as described previously (Oldach et al. 2000)and the universal primer B (Medlin et al. 1988).Subsequently, the 50-end was amplified using primer 668R50-CAA GAA GAT GCC AAG GCC GA-30 with primer A (Med-lin et al. 1988). Amplification of Hsp90 was carried out insemi-nested PCR applying degenerate outer primers OuterF:50-CAGCTGATGTCCCTGATCATYAAYACNTTYTA-30 andOuterR: 50-TCGAGGGAGAGRCCNARCTTRATCAT-3 0 (Sim-pson, Lukes, and Roger 2002), and in the subsequent stepusing InnerR:- 50-CGCCTTCATDATNCKYTCCATTRTTNGC-30 (Keeling and Leander 2003). The temperature cycles used in thereactions were: 15 min or 4 min at 95 1C; 35–45 cycles of 15 s at95 1C, 20 s at 48–52 1C, and 1.5 min at 72 1C; and 10 min at 72 1C.An annealing temperature of 48 1C was used in the initial ampli-fication of the Hsp90 gene, raised to 52 1C in the nested PCR. Allamplifications used a Perkin-Elmer thermocycler (GeneAmp PCRSystem 2400, Perkin-Elmer, (Wellesley, MA, USA)) and 1mL oftemplate (of DNA-extraction or a 1:10 dilution of the initial PCRproduct), 0.125–0.25 mM of each primer, 0.2 mM dNTP, 1� PCRbuffer, 0.25mg bovine serum albumin, and 1.5 U of HotStar Taq(Qiagen, Crawley, UK) or 1 U of DynaZyme polymerase (Finn-zymes Oy, Espoo, Finland) in a total 25-mL vol. Purified PCR-prod-ucts were directly sequenced or ligated into the pCR-2.1 TOPOvector (Invitrogen, Carlsbad, CA) before sequencing with ABI BIG-DYETM Terminator Cycle Sequencing Ready Reaction Kit onABI3700 or ABI3730 (Applied Biosystems, Foster City, CA).
Sequence alignment, model testing, and phylogeneticanalyses. Sequences of nuclear SSU rRNA, LSU rRNA, andHsp90 (Table 1) were aligned with other dinoflagellate data us-ing CLUSTALX (Thompson et al. 1997) and subsequently editedmanually (the Hsp90 sequence from A. carterae assembled fromcDNA library sequences was kindly provided by Tsvetan Bach-varoff and Chuck Delwiche). In concatenated Hsp901SSU1LSUsequences, the missing LSU sequences were filled with question
marks (for alignments, contact KST). Model comparisons for allnuclear SSU rRNA, LSU rRNA, and Hsp90 sequences werecarried out in a Bayesian framework in which the Bayes factorwas used as criterion for choosing the best-fitting model. TheBayes factors were estimated from the harmonic mean of thelikelihood values sampled after the burn-in period from the Mark-ov chain Monte-Carlo (MCMC) chains implemented in MrBayesv3.0 (Ronquist and Huelsenbeck 2003). Two types of rate modelswere tested for the DNA and protein sequences, which allow var-iable substitution rates across sites (assuming a G distributionof rates and one category of invariable sites) and variable ratesbetween sequences using switch-rate parameters (COV),allowing sites to change between invariable and variable statesaccording to the covarion hypothesis (Huelsenbeck 2002; Miya-moto and Fitch 1995). The stationary substitution model for therDNA sequences was set to a general time reversible (GTR) mod-el, whereas the stationary model for Hsp90 protein was chosenapplying Bayesian information criterion (BIC) in ProtTest v1.2(Abascal, Zardoya, and Posada 2005). Inferences for combinedsequence alignments were carried out by partitioning the data andapplying the best-fitting model for each gene. The following pa-rameters were unlinked in the Bayesian inferences: g shape,branch lengths, switching rate for the covarion model, substitu-tion rates of the GTR model, and the character state frequencies.The latter two parameters were used for rDNA sequences only.
Phylogenetic analyses of single gene and concatenated nuclearSSU rRNA, LSU rRNA, and Hsp90 sequences were performedusing MrBayes v3.0 (Ronquist and Huelsenbeck 2003). The Gdistribution of site rates was approximated using four rate cate-gories with equal probability. Other prior settings were set todefault values. The MCMC chains lasted for 2,000,000–10,000,000generations, and trees were saved each 100 generations. Afterburn-in, which was set to 300,000–5,000,000 generations basedon visual inspection of the stationary phase of the MCMC chains,the remaining trees were used for calculating the consensus tree(shown in all figures) and posterior probability values. Conver-gence of the MCMC chains was tested running the Bayesian in-ference at least twice from different random starting trees. As treetopologies, mean likelihood scores, and the posterior probabilityvalues showed almost identical results after burn-in from theindependent runs, we assumed that the MCMC had lasted longenough to converge.
Maximum likelihood (ML) bootstrap analysis on the Hsp90data was performed using a rtREV1G1I substitution modelimplemented in PHYML v.2.4.4 (Guindon and Gascuel 2003).In addition, bootstrap analyses of the SSU and LSU data wereperformed by PAUP� (Swofford 1998) using LogDet neighborjoining distances, and ML criteria applying a GTR1G1Ievolutionary model, one heuristic search per pseudoreplicateand NNI branch swapping. All bootstrap values were estimatedwith 100 pseudoreplicates.
RESULTS
Model comparison. Model comparison in a Bayesian frame-work favoured different models for the Hsp90, SSU rRNA, andLSU rRNA genes. For the SSU rRNA gene, the covarion modelGTR1G1COV received significantly higher harmonic meanlikelihood values than competing models, whereas for the LSUrRNA and Hsp90 data the various rate models were estimated tobe almost equally fit. We interpreted the differences in likelihoodscores for the various evolutionary models in accordance with theguidelines provided by Newton and Raftery (1994) and Kass andRaftery (1995). To avoid over-parameterisation only the G distri-butions of site rates were applied in inferences of the latter twogenes. In addition, using the ProtTest program, the best-fitting
218 J. EUKARYOT. MICROBIOL., VOL. 53, NO. 3, MAY– JUNE 2006
stationary substitution model for the Hsp90 protein sequence wasestimated to be the rtREV model.
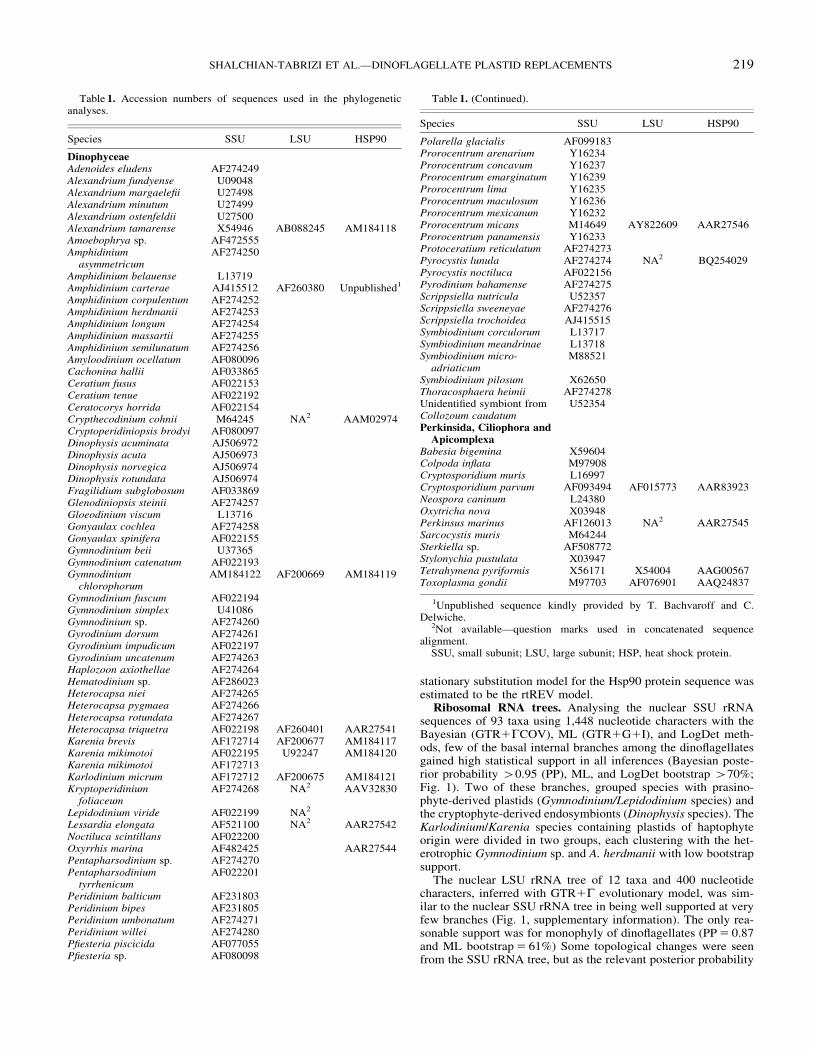
Ribosomal RNA trees. Analysing the nuclear SSU rRNAsequences of 93 taxa using 1,448 nucleotide characters with theBayesian (GTR1GCOV), ML (GTR1G1I), and LogDet meth-ods, few of the basal internal branches among the dinoflagellatesgained high statistical support in all inferences (Bayesian poste-rior probability 40.95 (PP), ML, and LogDet bootstrap 470%;Fig. 1). Two of these branches, grouped species with prasino-phyte-derived plastids (Gymnodinium/Lepidodinium species) andthe cryptophyte-derived endosymbionts (Dinophysis species). TheKarlodinium/Karenia species containing plastids of haptophyteorigin were divided in two groups, each clustering with the het-erotrophic Gymnodinium sp. and A. herdmanii with low bootstrapsupport.
The nuclear LSU rRNA tree of 12 taxa and 400 nucleotidecharacters, inferred with GTR1G evolutionary model, was sim-ilar to the nuclear SSU rRNA tree in being well supported at veryfew branches (Fig. 1, supplementary information). The only rea-sonable support was for monophyly of dinoflagellates (PP 5 0.87and ML bootstrap 5 61%) Some topological changes were seenfrom the SSU rRNA tree, but as the relevant posterior probability
Table 1. Accession numbers of sequences used in the phylogeneticanalyses.
Species SSU LSU HSP90
DinophyceaeAdenoides eludens AF274249Alexandrium fundyense U09048Alexandrium margaelefii U27498Alexandrium minutum U27499Alexandrium ostenfeldii U27500Alexandrium tamarense X54946 AB088245 AM184118Amoebophrya sp. AF472555Amphidinium
asymmetricumAF274250
Amphidinium belauense L13719Amphidinium carterae AJ415512 AF260380 Unpublished1
Amphidinium corpulentum AF274252Amphidinium herdmanii AF274253Amphidinium longum AF274254Amphidinium massartii AF274255Amphidinium semilunatum AF274256Amyloodinium ocellatum AF080096Cachonina hallii AF033865Ceratium fusus AF022153Ceratium tenue AF022192Ceratocorys horrida AF022154Crypthecodinium cohnii M64245 NA2 AAM02974Cryptoperidiniopsis brodyi AF080097Dinophysis acuminata AJ506972Dinophysis acuta AJ506973Dinophysis norvegica AJ506974Dinophysis rotundata AJ506974Fragilidium subglobosum AF033869Glenodiniopsis steinii AF274257Gloeodinium viscum L13716Gonyaulax cochlea AF274258Gonyaulax spinifera AF022155Gymnodinium beii U37365Gymnodinium catenatum AF022193Gymnodinium
chlorophorumAM184122 AF200669 AM184119
Gymnodinium fuscum AF022194Gymnodinium simplex U41086Gymnodinium sp. AF274260Gyrodinium dorsum AF274261Gyrodinium impudicum AF022197Gyrodinium uncatenum AF274263Haplozoon axiothellae AF274264Hematodinium sp. AF286023Heterocapsa niei AF274265Heterocapsa pygmaea AF274266Heterocapsa rotundata AF274267Heterocapsa triquetra AF022198 AF260401 AAR27541Karenia brevis AF172714 AF200677 AM184117Karenia mikimotoi AF022195 U92247 AM184120Karenia mikimotoi AF172713Karlodinium micrum AF172712 AF200675 AM184121Kryptoperidinium
foliaceumAF274268 NA2 AAV32830
Lepidodinium viride AF022199 NA2
Lessardia elongata AF521100 NA2 AAR27542Noctiluca scintillans AF022200Oxyrrhis marina AF482425 AAR27544Pentapharsodinium sp. AF274270Pentapharsodinium
tyrrhenicumAF022201
Peridinium balticum AF231803Peridinium bipes AF231805Peridinium umbonatum AF274271Peridinium willei AF274280Pfiesteria piscicida AF077055Pfiesteria sp. AF080098
Table 1. (Continued).
Species SSU LSU HSP90
Polarella glacialis AF099183Prorocentrum arenarium Y16234Prorocentrum concavum Y16237Prorocentrum emarginatum Y16239Prorocentrum lima Y16235Prorocentrum maculosum Y16236Prorocentrum mexicanum Y16232Prorocentrum micans M14649 AY822609 AAR27546Prorocentrum panamensis Y16233Protoceratium reticulatum AF274273Pyrocystis lunula AF274274 NA2 BQ254029Pyrocystis noctiluca AF022156Pyrodinium bahamense AF274275Scrippsiella nutricula U52357Scrippsiella sweeneyae AF274276Scrippsiella trochoidea AJ415515Symbiodinium corculorum L13717Symbiodinium meandrinae L13718Symbiodinium micro-
adriaticumM88521
Symbiodinium pilosum X62650Thoracosphaera heimii AF274278Unidentified symbiont fromCollozoum caudatum
U52354
Perkinsida, Ciliophora andApicomplexa
Babesia bigemina X59604Colpoda inflata M97908Cryptosporidium muris L16997Cryptosporidium parvum AF093494 AF015773 AAR83923Neospora caninum L24380Oxytricha nova X03948Perkinsus marinus AF126013 NA2 AAR27545Sarcocystis muris M64244Sterkiella sp. AF508772Stylonychia pustulata X03947Tetrahymena pyriformis X56171 X54004 AAG00567Toxoplasma gondii M97703 AF076901 AAQ24837
1Unpublished sequence kindly provided by T. Bachvaroff and C.Delwiche.
2Not available—question marks used in concatenated sequencealignment.
SSU, small subunit; LSU, large subunit; HSP, heat shock protein.
219SHALCHIAN-TABRIZI ET AL.—DINOFLAGELLATE PLASTID REPLACEMENTS

0.1
Alexandrium tamarenseAlexandrium fundyense
Alexandrium minutumAlexandrium ostenfeldii
Alexandrium margalefeiiPyrocystis lunula
Pyrocystis noctilucaFragilidium subglobosum
Pyrodinium bahamenseProtoceratium reticulatum
Ceratocorys horrida
Gonyaulax spiniferaCeratium fusus
Ceratium tenueDinophysis acuminataDinophysis norvegicaDinophysis acuta
Dinophysis rotundataAmphidinium massartiiAmphidinium carteraeAmphidinium belauense
Amphidinium asymmetricumCrypthecodinium cohnii
Gyrodinium uncatenumGyrodinium dorsum
Glenodiniopsis steiniiSymbiodinium meandrinaeSymbiodinium corculorumSymbiodinium pilosumSymbiodinium microadriaticum
Gymnodinium simplexGymnodinium beiiPolarella glacialis
Amphidinium corpulentumGleodinium viscum
Prorocentrum limaProrocentrum arenariumProrocentrum maculosum
Prorocentrum concavumUnidentified symbiont from Collozoum caudatumScrippsiella nutricula
Kryptoperidinium foliaceumPeridinium balticum
Peridinium umbonatumPentapharsodinium sp.Pentapharsodinium tyrrhenicum
Prorocentrum mexicanumProrocentrum micans
Prorocentrum emarginatumProrocentrum panamensis
Amphidinium longumPeridinium willeiPeridinium bipes
Adenoides eludensPfiesteria piscicida
Pfiesteria sp.Cryptoperidiniopsis brodyi
Amyloodinium ocellatumThoracosphaera heimiiGyrodinium impudicum
Gymnodinium catenatumLepidodinium viride
Gymnodinium chlorophorumGymnodinium fuscum
Scrippsiella sweeneyaeScrippsiella trochoideaHeterocapsa nieiCachonina halliiHeterocapsa pygmaeaHeterocapsa rotundataHeterocapsa triquetra
Haplozoon axiothellaeAmphidinium semilunatum
Hematodinium sp.Amoebophrya sp.
Noctiluca scintillansPerkinsus marinus
Oxytricha novaStylonychia pustulataSterkiella sp.Colpoda inflata
Babesia bigeminaCryptosporidium murisNeospora caninumToxoplasma gondiiSarcocystis muris
Gonyaulaxcochlea
0.9888-
1.0 98 90
CILIOPHORA
APICOMPLEXA
PERKINSIDA
DINOPHYCEAE
Ha
Di
Pr
Cr
Karenia mikimotoiKarenia brevisKarenia mikimotoi
Amphidinium herdmanii
Gymnodinium sp.Karlodinium micrum
Fig. 1. Small subunit rRNA tree of dinoflagellates and alveolate outgroup species, inferred with Bayesian inferences applying the GTR1G1COVmodel. Thick branches denote branches with posterior probability values 40.95 and bootstrap support gained from LogDet distance and maximumlikelihood analysis 470%. Species with different plastid types are marked according to their origin or pigmentation by the abbreviations: Pr,prasinophyte-derived; Ha, haptophyte-derived; Di, diatom-derived. New sequence is marked in bold.
220 J. EUKARYOT. MICROBIOL., VOL. 53, NO. 3, MAY– JUNE 2006

values were even lower, none of the differences are consideredmeaningful.
Heat shock protein 90 tree. The Hsp90 tree of 17 taxa using523 amino-acid characters was inferred using the rtREV1G ev-olutionary model (Fig. 2). The inferred tree supported dinoflagel-lates as a group with the exclusion of Apicomplexa, ciliates,Perkinsida, and O. marina with PP 5 1.00, and ML 5 99%. AllGymnodiniales were placed in different branches at the base of thearmoured dinoflagellates, separated from Prorocentrales, Peridini-ales, and Gonyaulacales (PP 5 0.99 and ML 5 59%). The per-idinin-containing plastid of A. carterae was placed as the mostbasal branch (PP 5 0.97 and ML o50%), whereas speciescontaining haptophyte- and prasinophyte-derived plastids alsobranched relatively early in the dinoflagellate tree. In addition,the diatom-containing K. foliaceum was placed together with theheterotrophic C. cohnii (PP 5 1.00 and ML 5 57%). Comparedwith the nuclear SSU and LSU rRNA trees, the Hsp90 tree showedseveral topological differences and received relatively much high-er posterior probability values. ML bootstrap analysis generatedhighly congruent tree topology, but only a few of the nodes werehighly supported, such as the clustering of species with the ha-ptophyte-derived plastids (ML 5 99%). Altogether, the bootstrapanalysis does not contradict the Bayesian tree, but does not sup-port it substantially either.
The usefulness of the Hsp90 and nuclear SSU rRNA sequencesfor dinoflagellate phylogeny was compared by generating another,smaller SSU rRNA tree equivalent to the taxon sampling in theHsp90 data (i.e. 17 taxa and 1,448 nucleotide characters; Fig. 2 insupplementary information). Comparison showed considerablyhigher posterior probability values for almost all deep nodes inthe dinoflagellate Hsp90 tree.
Concatenated Hsp901SSU1LSU tree. The tree generatedfrom concatenated Hsp901SSU1LSU sequences, which consist-ed of 17 taxa and 2,398 mixed nucleotide and amino-acid
characters, was inferred using separate models for the Hsp90and the rRNA partitions in agreement with the estimated best fit-ting models (i.e. Hsp90: rtREV1G, SSU rRNA: GTR1G1COV,LSU rRNA: GTR1G). The phylogeny (Fig. 3) showed almost thesame branching pattern as the Hsp90 tree (Fig. 2). The only top-ological change was the placement of L. elongata (Peridiniales),moderately supported as a sister to one of the gonyaulacaleanclades (PP 5 0.75). The posterior probability values were quitesimilar to the Hsp90 tree, but a few noticeable branches wereslightly better supported, such as the Gonyaulacales/Peridinalesbranch (PP 5 1.00).
DISCUSSION
Haptophyte and prasinophyte plastids were each acquiredonly once. Previous works showed that the basal branching orderof dinoflagellate phylogeny is difficult to resolve using rRNAgenes (Cavalier-Smith and Chao 2004; Grzebyk, Sako, andBerland 1998; Murray et al. 2005; Saldarriaga et al. 2001; Saun-ders et al. 1997; Tengs et al. 2000). One problem may be related tocovarion substitution patterns in the SSU and LSU sequences,which have recently been identified in various SSU sequences(Galtier 2001; Huelsenbeck 2002; Lopez, Forterre, and Philippe1999). Estimation of the model fit in a Bayesian framework for thenuclear SSU rRNA gene showed that the COV model was almost83 likelihood scores or a Bayes factor of 166 better than for thebest model without the COV parameters. Having in mind that aBayes factor larger than 10 is usually regarded as decisive evi-dence for the better model (Kass and Raftery 1995), it clearly fitsthe dinoflagellate SSU rRNA sequences better than other testedmodels. However, using the COV substitution model, the result-ing SSU rRNA tree was only supported at the terminal branches,suggesting that SSU rRNA genes contain insufficient phylogenet-ic information to resolve dinoflagellate phylogeny, likely caused
Fig. 3. Tree inferred from combined sequences of heat shock protein(Hsp) 90, small subunit (SSU) and large subunit (LSU) rRNA. The datawere partitioned in agreement with the gene lengths and the best fittingmodel for each partition was used for the protein and DNA coding regionsas follows: Hsp90—rtREV1G; SSU rRNA—GTR1G1COV; and LSUrRNA—GTR1G. The tree shows similar topology and posterior proba-bilities for the nodes as the Hsp90 tree (cf. Fig. 2).
Fig. 2. Dinoflagellate heat shock protein (Hsp) 90 tree inferred withthe Bayesian Markov Chain Monte Carlo chain method using thertREV1G substitution model. Numbers at internal branches indicate theBayesian posterior probability values and maximum likelihood values450%. Abbreviations indicating the plastid origins: Pe, peridinin-containing; Pr, prasinophyte-derived; Ha, haptophyte-derived; Di, diatom-derived. New sequences for this report are marked in bold.
221SHALCHIAN-TABRIZI ET AL.—DINOFLAGELLATE PLASTID REPLACEMENTS

by rapid early divergence of the group (Saldarriaga et al. 2004).Nevertheless, the nuclear SSU rRNA tree strongly groups the twogreen-coloured L. viride and G. chlorophorum species within agroup bearing peridinin-containing plastids, suggesting that theirprasinophyte-derived plastids were established in a photosyntheticcommon ancestor by replacement of the peridinin-containingplastid. Alternatively, the plastid could have been gained by aheterotrophic species that had already lost its peridinin plastid.The monophyly of dinoflagellates with haptophyte-derived plas-tids was not strongly supported in SSU trees, being clustered todifferent heterotrophic species. However, these three species re-ceived strong support in Hsp90 and combined DNA and proteintrees. The nuclear SSU rRNA data alone may indicate polyphyle-tic origin of their plastids, but as statistical supports for the asso-ciated nodes are low and inferences of plastid phylogenies alwaysplace these species as a single group within the haptophytes(Shalchian-Tabrizi et al. 2006; Tengs et al. 2000; Yoon et al.2005), it is more likely caused by insufficient signal in the nuclearSSU rRNA data for this branch. Therefore, probably only twoendosymbiotic events gave rise to fully integrated and aberrantlypigmented plastids of dinoflagellates. The Hsp90 and concatenat-ed analyses placed K. foliaceum as sister to C. cohnii. Other spe-cies that harbour endosymbionts belong to unrelated genera (seeFensome et al. 1993; Saldarriaga et al. 2001), such as Dinophysis,Amphidinium, Amphisolenia, and Noctiluca, suggesting that fac-ultative endosymbioses are widely distributed among dinoflagel-lates.
Haptophyte and prasinophyte plastids replaced peridinin-containing ones. Compared with the SSU rRNA tree, the top-ologies of the Hsp90 and Hsp901SSU1LSU trees are highlycongruent and supported by substantially higher posterior proba-bilities. This demonstrates that the dinoflagellate Hsp90 sequenc-es contain stronger phylogenetic signal than the rDNA sequencesand therefore they are likely more suitable for inferences of thedinoflagellate phylogeny. As trees derived from the Hsp90 and theconcatenated sequences both place the peridinin-containing A.carterae as the most basal dinoflagellate (apart from O. marina),our results suggest that the peridinin-containing plastid wasestablished before the aberrantly pigmented plastid types (see al-so J�rgensen, Murray, and Daugberg 2004) and replaced by otherplastids in at least two independent endosymbiotic events. Fur-thermore, the branching patterns may indicate that both the ha-ptophyte- and prasinophyte-derived plastids were establishedrelatively early in the radiation of the dinoflagellates, presumablyafter the earliest divergence of peridinin-containing species, but alarger taxon sampling will be needed to test this hypothesis. Somecaution should also be associated with these conclusions due tothe weak bootstrap values for the associated deep nodes. Partic-ularly, the data do not completely rule out the possibility of theKarlodinium and Karenia clade (i.e. haptophyte-derived plastids)being the deepest branch of plastid-containing dinoflagellates.Thus, there exists the possibility of independent acquisitions ofhaptophyte-derived and peridinin-containing plastids. Oxyrrhismarina is placed as sister species to the dinoflagellates, confirm-ing previous Hsp90 trees with fewer dinoflagellate taxa. There-fore, previous contradictory phylogenies of O. marina in nuclearSSU rRNA and Hsp90 trees were probably not caused by inap-propriate Hsp90 taxon sampling (Cavalier-Smith and Chao 2004),but rather caused by rapid evolution of the SSU rRNA gene in O.marina. The inferred Hsp90- and three-gene trees indicate that allGymnodiniales diverged early in the evolution of dinoflagellatesand subsequently gave rise to lineages with the characteristic platearmour (Fensome et al. 1993), including the plate-forming Pro-rocentrales (superficially covered by two plates), the Gonyaulac-ales, and Peridiniales (multiple plates arranged in series of plates).This conflicts with the tentative conclusion reached from nuclear
SSU rRNA trees, from which it was suggested that Gymnodialesare polyphyletically derived from armoured dinoflagellates bymultiple losses of plates (Cavalier-Smith and Chao 2004). How-ever, although Gonyaulacales and Peridiniales were inferred to beclosely related, the placements of C. cohnii (Gonyaulacales) andL. elongata (Peridiniales) obscure the demarcation between thetwo orders (Fensome et al. 1993), and lead one to question wheth-er the Hsp90 tree is as reliable as its high posterior probabilityvalues implies. Interestingly, recent cytochrome b trees alsogrouped Gonyaulacales and Peridiniales together, and clusteredGymnodiales and Prorocentrales as two monophyletic groups, butapparently the placement of C. cohnii was prone to long-branchattraction to some outgroup species (Zhang, Bhattacharya, andLin 2005). Despite some congruence between Hsp90 and cyto-chrome b trees, differences in branching order among the clades,such as the placement of Prorocentrales, and the unclear place-ment of some species like C. cohnii, suggest that a broader sam-pling of taxa and sequences from a greater variety of genes areessential to resolve dinoflagellate phylogeny reliably, and to fur-ther address the different hypotheses about the evolution ofplastids in this group.
ACKNOWLEDGMENTS
We thank Torstein Tengs and Knut Rudi for critically readingan earlier draft of this manuscript, and Andreas Botnen for assist-ing the computer analyses. We also thank Geir Storvik for dis-cussion on statistical methods. All analyses were performed at theBioportal, University of Oslo (http://www.bioportal.uio.no).T. Bachvaroff and C. Delwiche kindly provided Hsp90 sequenc-es from the A. carterae cDNA library. This work was supportedby grants from the Norwegian Research Council to K. S. J.(no 118894/431), D. K. (121187/720), and grants from NERC toT. C. S.
LITERATURE CITED
Abascal, F., Zardoya, R. & Posada, D. 2005. Prottest: selection of best-fitmodels of protein evolution. Bioinformatics, 21:2104–2105.
Bj�rnland, T. 1990. Carotenoid structures and lower plant phylogeny.In: Krinsky, N. I., Mathews-Roth, M. M. & Taylor, R. F. (ed.),Carotenoids: Chemistry and Biology. Plenum Press, New York.p. 21–37.
Bj�rnland, T. & Liaaen-Jensen, S. 1989. Distribution patterns ofcarotenoids in relation to chromophyte phylogeny and systematics. In:Green, J. C., Leadbeater, B. S. C. & Diver, W. L. (ed.), The Chromo-phyte Algae: Problems and Perspectives. Published for the SystematicsAssociation by Clarendon Press, Oxford. p. 37–60.
Cavalier-Smith, T. 2000. Membrane heredity and early chloroplast evo-lution. Trends Plant. Sci., 5:174–182.
Cavalier-Smith, T. 2003. Genomic reduction and evolution of novelgenetic membranes and protein-targeting machinery in eukaryote-eukaryote chimaeras (meta-algae). Philos. Trans. Roy. Soc. London BBiol. Sci., 358:109–133 (discussion p. 133–134).
Cavalier-Smith, T. & Chao, E. E. 2004. Protalveolate phylogeny andsystematics and the origins of Sporozoa and dinoflagellates (phylumMyzozoa nom. nov.). Eur. J. Protistol., 40:185–212.
Chesnick, J. M., Morden, C. W. & Schmieg, A. M. 1996. Identity of theendosymbiont of Peridinium foliaceum (Pyrrophyta): analysis of therbcLS operon. J. Phycol., 32:850–857.
Chesnick, J. M., Kooistra, W. H., Wellbrock, U. & Medlin, L. K. 1997.Ribosomal RNA analysis indicates a benthic pennate diatom ancestryfor the endosymbionts of the dinoflagellates Peridinium foliaceum andPeridinium balticum (Pyrrhophyta). J. Eukaryot. Microbiol., 44:314–320.
Daugbjerg, N., Hansen, G., Larsen, J. & Moestrup, Ø. 2000. Phylogeny ofsome of the major genera of dinoflagellates based on ultrastructure andpartial LSU rDNA sequence data, including the erection of three newgenera of unarmoured dinoflagellates. Phycologia, 39:302–317.
222 J. EUKARYOT. MICROBIOL., VOL. 53, NO. 3, MAY– JUNE 2006

Delwiche, C. F. 1999. Tracing the thread of plastid diversity through thetapestry of life. Am. Nat., 154(Suppl.):S164–S177.
Dodge, J. D. & Crawford, R. M. 1969. Observations on the fine structureof the eyespot and associated organelles in the dinoflagellate Glen-odinium foliaceum. J. Cell Sci., 5:479–493.
Edvardsen, B., Shalchian-Tabrizi, K., Jakobsen, K. S., Medlin, L. K.,Dahl, E., Brubak, S. & Paasche, E. 2003. Genetic variability andmolecular phylogeny of Dinophysis species (Dinophyceae) from Nor-wegian waters inferred from single cell analyses of rDNA. J. Phycol.,39:395–408.
Elbrachter, M. & Schnepf, E. 1996. Gymnodinium chlorophorum, anew, green, bloom-forming dinoflagellate (Gymnodiniales, Din-ophyceae) with a vestigal prasinophyte endosymbiont. Phycologia, 35:381–393.
Fast, N. M., Xue, L., Bingham, S. & Keeling, P. J. 2002. Re-examiningalveolate evolution using multiple protein molecular phylogenies.J. Eukaryot. Microbiol., 49:30–37.
Fensome, R. A., Taylor, F. J. R., Norris, G., Sarjeant, W. A. S., Wharton,D. I. & Williams, G. L. 1993. A classification of living and fossil dino-flagellates. Micropaleontology, Special Publication No. 7. p. 1–351.
Gibbs, S. P. 1981. The chloroplasts of some algal groups may haveevolved from endosymbiontic algae. Ann. N. Y. Acad. Sci., 361:193–297.
Grzebyk, D., Sako, Y. & Berland, B. 1998. Phylogenetic analysis of ninespecies of Prorocentrum (Dinophyceae) inferred from 18S ribosomalDNA sequences, morphological comparisons, and description of Pro-rocentrum panamensis sp. nov. J. Phycol., 34:1055–1068.
Guindon, S. & Gascuel, O. 2003. A simple, fast and accurate method toestimate large phylogenies by maximum-likelihood. Syst. Biol., 52:696–704.
Haywood, A. J., Steidinger, K. A., Truby, E. W., Bergquist, P. R.,Bergquist, P. L., Adamson, J. & MacKenzie, L. 2004. Comparativemorphology and molecular phylogenetic analysis of three new speciesof the genus Karenia (Dinophyceae) from New Zealand. J. Phycol.,40:165–179.
Hoek, C. v. d., Mann, D. G. & Jahns, H. M. 1995. Algae: An Introductionto Phycology. Cambridge University Press, Cambridge.
Honsell, G. & Talarico, L. 2004. Gymnodinium chlorophorum (Din-ophyceae) in the Adriatic Sea: electron microscopical observations.Bot. Marina, 47:152–166.
Huelsenbeck, J. P. 2002. Testing a covariotide model of DNA substitution.Mol. Biol. Evol., 19:698–707.
Jakobsen, K. S., Tengs, T., Vatne, A., Bowers, H. A., Oldach, D. W.,Burkholder, J. M., Glasgow, H. B. Jr., Rublee, P. A. & Klaveness, D.2002. Discovery of the toxic dinoflagellate Pfiesteria in northern Euro-pean waters. Proc. Roy Soc. B., 22:211–214.
Janson, S. 2004. Molecular evidence that plastids in the toxin-producingdinoflagellate genus Dinophysis originate from the free-living crypto-phyte Teleaulax amphioxeia. Environ. Microbiol., 6:1102–1106.
Jeffrey, S. W., Sielicki, M. & Haxo, F. T. 1975. Chloroplast pigment pat-terns in dinoflagellates. J. Phycol., 21:50–56.
J�rgensen, M. J., Murray, S. & Daugbjerg, N. 2004. Amphidinium revis-ited. I. Redefinition of Amphidinium (Dinophyceae) based on cladisticand molecular phylogenetic analysis. J. Phycol., 40:351–365.
Kass, R. E. & Raftery, A. E. 1995. Bayes factors. J. Am. Stat. Assoc.,90:773–795.
Keeling, P. J. & Leander, B. S. 2003. Characterisation of a non-canonicalgenetic code in the oxymonad Streblomastix strix. J. Mol. Biol.,326:1337–1349.
Kempton, J. W., Wolny, J., Tengs, T., Rizzo, P., Morris, R., Tunnell, J.,Scott, P., Steidinger, K., Hymele, S. N. & Lewitus, A. L. 2002.Kryptoperidinium foliaceum blooms in South Carolina: a multi-analyt-ical approach to identification. Harmful Algae, 1:383–392.
Leander, B. S. & Keeling, P. J. 2003. Morphostasis in alveolate evolution.Trends Ecol. Evol., 18:395–402.
Leander, B. S. & Keeling, P. J. 2004. Early evolutionary history of dino-flagellates and apicomplexans (Alveolata) as inferred from Hsp90 andactin phylogenies. J. Phycol., 40:341–350.
Lopez, P., Forterre, P. & Philippe, H. 1999. The root of the tree of life inthe light of the covarion model. J. Mol. Evol., 49:496–508.
McEwan, M. L. & Keeling, P. J. 2004. Hsp90, tubulin and actin are re-tained in the tertiary endosymbiont genome of Kryptoperidinium folia-ceum. Eukaryot. Microbiol., 51:651–659.
Medlin, L., Elwood, H. J., Stickel, S. & Sogin, M. L. 1988. The charac-terisation of enzymatically amplified eukaryotic 16S-like rRNA-codingregions. Gene, 71:491–499.
Miyamoto, M. M. & Fitch, W. M. 1995. Testing the covarion hypothesis ofmolecular evolution. Mol. Biol. Evol., 12:503–513.
Murray, S., J�rgensen, M. F., Ho, S. Y. W., Patterson, D. J. & Jermiin, L.S. 2005. Improving the analysis of dinoflagellate phylogeny based onrDNA. Protist, 156:269–286.
Newton, M. A. & Raftery, A. E. 1994. Approximate Bayesian inference bythe weighted likelihood bootstrap (with discussion). J. Roy. Stat. Soc.,56:3–48.
Oldach, D. W., Delwiche, C. F., Jakobsen, K. S., Tengs, T., Brown, E. G.,Kempton, J. W., Schaefer, E. F., Bowers, H. A., Glasgow, H. B. Jr.,Burkholder, J. M., Steidinger, K. A. & Rublee, P. A. 2000. Heterodu-plex mobility assay-guided sequence discovery: elucidation of the smallsubunit (18S) rDNA sequences of Pfiesteria piscicida and related dino-flagellates from complex algal culture and environmental sample DNApools. Proc. Natl. Acad. Sci. USA, 97:4303–4308.
Palmer, J. D. & Delwiche, C. F. 1998. The origin and evolution of plastidsand their genomes. In: Soltis, D. E., Soltis, P. S. & Doyle, J. J. (ed.),Molecular Systematics of Plants II: DNA Sequencing. Chapman &Hall, New York.
Ronquist, F. & Huelsenbeck, J. P. 2003. MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinformatics, 19:1572–1574.
Rudi, K., Kroken, M., Dahlberg, O. J., Deggerdal, A., Jakobsen, K. S. &Larsen, F. 1997. Rapid, universal method to isolate PCR-ready DNAusing magnetic beads. BioTechniques, 22:506–511.
de Salas, M. F., Bolch, C. J. S., Botes, L., Nash, G., Wright, S. W. &Hallegraeff, G. M. 2003. Takayama gen. nov. (Gymnodiniales, Din-ophyceae), a new genus of unarmored dinoflagellates with sigmoid api-cal grooves, including the description of two new species. J. Phycol.,39:1233–1246.
Saldarriaga, J. F., Taylor, F. J. R., Cavalier-Smith, T., Menden-Deuer, S. &Keeling, P. J. 2004. Molecular data and the evolutionary history ofdinoflagellates. Eur. J. Protistol., 40:85–111.
Saldarriaga, J. F., Taylor, F. J., Keeling, P. J. & Cavalier-Smith, T. 2001.Dinoflagellate nuclear SSU rRNA phylogeny suggests multiple plastidlosses and replacements. J. Mol. Evol., 53:204–213.
Saunders, G. W., Hill, D. R. A., Sexton, J. P. & Andersen, R. A. 1997.Small-subunit ribosomal RNA sequences from selected dinoflagellates:testing classical evolutionary hypotheses with molecular systematicmethods. Plant Syst. Evol., 11(Suppl.):237–259.
Schnepf, E. 1993. From prey via endosymbiont to plastid: comparativestudies in dinoflagellates. In: Lewin, R. A. (ed.), Origins of Plastids,Symbiogenesis, Prochlorophytes, and the Origins of Chloroplasts.Chapman & Hall, New York. p. 53–77.
Shalchian-Tabrizi, K., Skanseng, M., Ronquist, F., Klaveness, D., Bach-varoff, T. R., Delwiche, C. F., Botnen, A., Tengs, T. & Jakobsen, K. S.2006. Covarion structure in rhodophyte-derived second-hand plastidgenes: implications for addressing the origin and evolution of dinoflag-ellate plastids. Mol. Biol. Evol. (in press)
Simpson, A. G., Lukes, J. & Roger, A. J. 2002. The evolutionary historyof kinetoplastids and their kinetoplasts. Mol. Biol. Evol., 19:2071–2083.
Stechmann, A. & Cavalier-Smith, T. 2003. Phylogenetic analysis ofeukaryotes using heat-shock protein Hsp90. J. Mol. Evol., 57:408–419.
Stoecker, D. K. 1999. Mixotrophy among dinoflagellates. J. Eukaryot.Microbiol., 46:397–401.
Swofford, D. L. 1998. PAUP: Phylogenetic Analysis Using Parsimony(� and Other Methods). Sinauer Associates, Inc., Sunderland, MA.
Takishita, K., Nakano, K. & Uchida, A. 2000. Origin of the plastid in theanomalously pigmented dinoflagellate Gymnodinium mikimotoi (Gym-nodiniales, Dinophyta) as inferred from phylogenetic analysis based onthe gene encoding the large subunit of form I-type RuBisCO. Phycol.Res., 48:85–89.
Takishita, K., Koike, K., Maruyama, T. & Ogata, T. 2002. Molecular ev-idence for plastid robbery (kleptoplastidy) in Dinophysis, a dinoflagel-late causing diarrhetic shellfish poisoning. Protist, 153:293–302.
Tengs, T., Dahlberg, O. J., Shalchian-Tabrizi, K., Klaveness, D., Rudi, K.,Delwiche, C. F. & Jakobsen, K. S. 2000. Phylogenetic analyses indi-cate that the 190Hexanoyloxy-fucoxanthin-containing dinoflagellates
223SHALCHIAN-TABRIZI ET AL.—DINOFLAGELLATE PLASTID REPLACEMENTS

have tertiary plastids of haptophyte origin. Mol. Biol. Evol., 17:718–729.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F. & Higgins,D. G. 1997. The CLUSTAL_X windows interface: flexible strategiesfor multiple sequence alignment aided by quality analysis tools. Nucl.Acids Res., 25:4876–4882.
Watanabe, M. M., Suda, S., Inouye, I., Sawaguchi, T. & Chihara, M. 1990.Lepidodinium viride, new genus, new species (Gymnodiniales, Din-ophyta), a green dinoflagellate with a chlorophyll A-containing andchlorophyll B-containing endosymbiont. J. Phycol., 26:741–751.
Watanabe, M. M., Sasa, T., Suda, S., Inouye, I. & Takaichi, S. 1991. Majorcaretenoid composition of an endosymbiont in a green dinoflagellate,Lepidodinium viride. J. Phycol., 27(Suppl.):75.
Whatley, J. M. 1993. Chloroplast ultrastructure. In: Berner, T. (ed.),Ultrastructure of Microalgae. CRC Press, Boca Raton.
Yoon, H. S., Hackett, J. D., Van Dolah, F. M., Nosenko, T., Lidie, L.& Bhattacharya, D. 2005. Tertiary endosymbiosis driven genome
evolution in dinoflagellate algae. Mol. Biol. Evol., 22:1299–1308.
Zhang, H., Bhattacharya, D. & Lin, S. 2005. Phylogeny of dinoflagellatesbased on mitochondrial cytochrome b and nuclear small subunit rDNAsequence comparisons. J. Phycol., 41:411–420.
Supplementary Material
The following supplementary material is available for thisarticle online:
Fig. S1Fig. S2
Received: 04/15/05, 11/02/05, 02/11/06; accepted: 02/12/06
224 J. EUKARYOT. MICROBIOL., VOL. 53, NO. 3, MAY– JUNE 2006