chronic caffeine consumption increases the number of brain adenosine receptors
TRANSCRIPT

Life Sciences, Vol. 32, pp. 1135-1142 Pergamon Press Printed in the U.S.A.
CHRONIC CAFFEINE CONSUMPTION INCREASES THE NUMBER OF BRAIN ADENOSINE RECEPTORS
Jean-Philippe Boulenger 1, J i t~ndra Patel 2, Rober~ M. Post 1, Alexandra M. Parma-, Paul J. Marangos-
~Biological Psychiatry Branch, NIMH, Bethesda, Maryland 20205 Laboratory of Clinical Science, NIMH, Bethesda, Maryland 20205
(Received in final form November 22, 1983)
Summary
Caffeine, a potent central stimulant, is known to com- petitively inhibit the specific binding of both adenosine and benzodiazepine receptor ligands to brain membranes in vitro. In mice receiving a diet containing non-toxic doses of caffeine (200 or 400 mg/kg diet) for periods up to 40 days~ a dose-~elated increase in the number of binding sites for [ H]-CHA and [ HI DPX was observed in whole brain membranes without modifications of the receptors' a~finity. Furthermore, a trans- itory increase in the number of [ H]-DZP binding sites was observed. These preliminary data seem to confirm the involvement of the adenosine receptors in the mode of action of caffeine and may be relevant to the development of both tolerance and dependence to some of the central effects of this compound.
Caffeine, a methylxanthine (MX) exhibiting a variety of stimulant effects upon the central nervous system (CNS), is one of the most widely used psychotropic substances (i). Chronic caffeine consumption is extremely common, yet surprisingly little attention has been paid to the long-term biochemical effects of its administration. Recent animal studies have suggested that some of the acute stimulant effects of caffeine are related to its ability to block central ~denosine receptors (2,3). However, caffeine also competitively inhibits [ H]-diazepam (DZP) binding (4), an effect which may be related to its convulsant effects when administered at high doses (5). Although caffeine's affinity is lower for benzodiazepine (BDZ) receptors than for adenosine receptors (2,6), the interaction of caffeine with BDZ receptors may still be related to some of its effects, since only a small percentage of these receptors need to be occupied to manifest various pharmacological
effects (7).
In order to investigate the neurochemical changes relevant to the mode of action of caffeine, we studied the effects of chronic caffeine consumption upon central adenosine and BDZ receptors in mice. We report here preliminary evidence that oral administration of non-toxic doses of caffeine for periods up to 40 days induces an increase in the number of adenosine receptor sites in
Correspondence and Reprint Requests to : Dr. Paul J. Marangos
0024-3205/83/101135-08503.00/0

1136 Chronic Caffeine and Adenosine Receptors Vol. 32, No. 10, 1983
mouse whole brain membranes, as well as a transitory increase in the number of BDZ receptors. An increase in the number of brain adenosine receptors has also been recently reported after chronic administration of theophylline to rats (8). The receptor modifications may be relevant to the development of both tolerance and dependence to some of the central effects of caffeine as described in both animals (9-12) and humans (13-16).
Methods
Three groups of NIH general purpose mice (National Institutes of Health), weighing 15-20 g each, received pellets of a standard semi-purified diet (American Institute of Nutrition <76: the first group was given a caffeine- free diet; the second received a diet supplemented with 200 mg of caffeine (ICN Nutritional Biochemicals) per kg of diet; the third was given a diet supplemented with 400 mg of caffeine per kg of diet.
After 12, 26 and 40 days of continuous diet, 8 animals in each group were sacrificed. The whole brain of each animal was quickly removed and homogenized for i0 sec at speed 5, in 20 volumes of 50 mM Tris-HCl buffer (pH 7.4), with a Brinkmann polytron. The homogenate was centrifuged at 30,000 x g for 20 min. The resulting membrane pellet was resuspended in 20 volumes of the same buffer and incubated at room temperature for 30 min with 2 units/ml of calf intestinal mucosa adenosine deaminase (Sigma) to remove endogenous adenosine (16). The membrane solution was centrifuged a second time at 30,000 x g for 30 min, resuspended in 20 volumes of buffer and finally stored at -70°C. Because adenosine itself has proven an unsatisfactory ligand for adenosine r~cept~rs (18,19), binding assays were done with stable3adenosine analogues, [~H]-N v- cyclohexyladenosine (CHA) and 1,3,diethyl-8-[ H]-ph~nylxanthine (DPX), which label central adenosine receptors (17,20). Using [~H]-DZP as ligand, binding a~says for BDZ receptors were done. The same membranes were used to measure [ H]-DZP binding after they were centrifuged and resuspended twice more, as described above, in order to remove residual GABA (21). Binding was determined for each animal individually at single ligand concentration by assays routinely performed in triplicate by incubating 0.3-0.6 mg of membrane suspension in 50 ram Tris HCI buffer (pH 7.4) containing one concentration of each tritiated ligand in a final volume of 0.5 ml. The incubation time and temperature and the a~ount of ligand used were: a) 2 hours at room temperature with 3.5 nM for [~H]-~HA (11.5 ci/mmol, New England Nuclear); b) one hour at 4°C with 3.75 nM [~H]-DPX (12.4 ci/mmol, New England Nuclear); c) 15 min at 4°C with 1.25 nM [~H]-DZP (80 ci/rmmol, New England Nuclear). For each sample, non- specific binding was determined after addition of 0.025 ml of, respectively: unlabeled CHA (imM), 2-chloro-adenosine (imM) and DZP (0.1mM)° Incubations were terminated by filtration using 4x3 ml ice-cold 50 mM Tris HCI. The filtration was done through ~atman GF/B filters. The air dried filters were then counted in i0 ml Aquasol (New England Nuclear). For Scatchard analysis the membrane suspensions of the animals belonging to the same group, i.e., control, caffeine 200 mg/kg diet or caffeine 400 mg/kg diet, were pooled. Binding assays were routinely performed in triplicate with a concentration range of the same tritiated ligands in a final volume of 0.5 ml as indicated previously. The protein measurements were performed according to Lowry (22).
Results
Preliminary studies were performed where caffeine at a dose of 20 mg/kg was injected into mice (i.p.) twice daily. In these studies, significant increases in adenosine receptors were observed only after two ~eeks of injections, while non-significant increases were observed in [~H] DZP binding. A preliminary study using caffeine supplemented diets at a level of 200 mg/kg
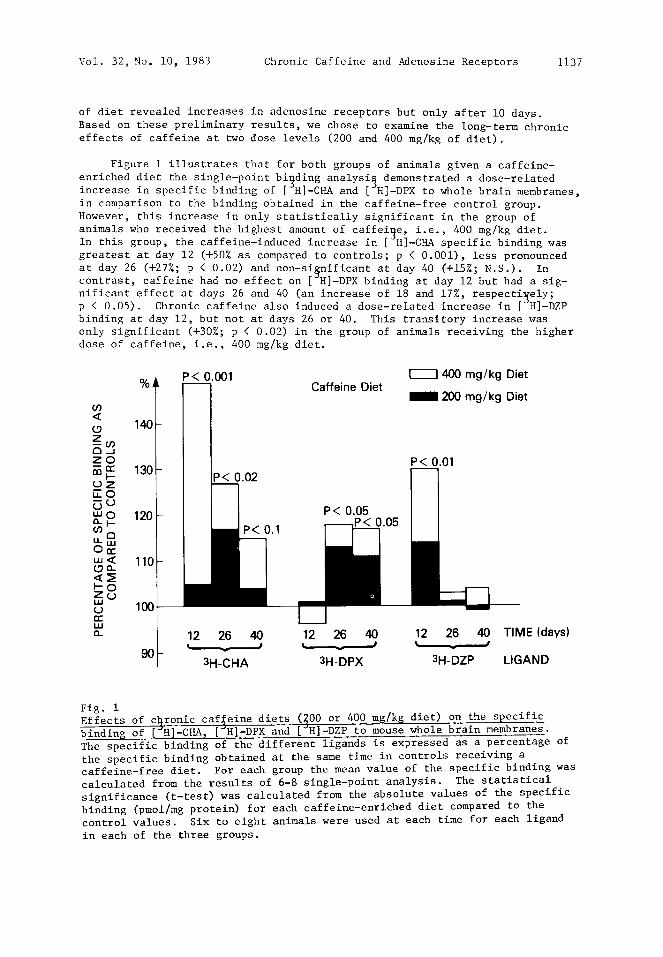
Vol. 32, No. i0, 1983 Chronic Caffeine and Adenosine Receptors 1137
of diet revealed increases in adenosine receptors but only after i0 days. Based on these preliminary results, we chose to examine the long-term chronic effects of caffeine at two dose levels (200 and 400 mg/kg of diet).
Figure 1 illustrates that for both groups of animals given a caffeine- enriched diet the single-point bi~ding analysi~ demonstrated a dose-related increase in specific binding of [~H]-CHA and [~H]-DPX to whole brain membranes, in comparison to the binding obtained in the caffeine-free control group. However, this increase in only statistically significant in the group of animals who received the highest amount of caffeine, i.e., 400 mg/kg diet. In this group, the caffeine-induced increase in [ H]-CHA specific binding was greatest at day 12 (+50% as compared to controls; p < 0.001), less pronounced at day 26 (+27%; p < 0.02) and non-si§nificant at day 40 (+15%; N.S.). In contrast, caffeine had no effect on [ H]-DPX binding at day 12 but had a sig- nificant effect at days 26 and 40 (an increase of 18 and 17%, respectively; p < 0.05). Chronic caffeine also induced a dose-related increase in [~H]-DZP binding at day 12, but not at days 26 or 40. This transitory increase was only significant (+30%; p < 0.02) in the group of animals receiving the higher dose of caffeine, i.e., 400 mg/kg diet.
Oo <
(.9 z - - 0 0 t ~ . j Z O
( J Z ,7O
w O a . t - ( /3
u J < Lga-
~ O Z u i i i (J t ~ uJ 0_
,
140
130
120
P< 0.001
110
P< 0.02
) . 1
Caffeine Diet
P< 0.05 - .05
[ - - -7 400 rng/kg Diet
1 200 rng/kg Diet
P< 0.01
100 U
I 12 26 40 12 26 40 12 26 40 TIME (days) 9 0 ~ v J ~ v / i v b
3H_CH A 3H_DPX 3H-DZP LIGAND
Fig. i Effects of c~ronic caffeine diets (~00 or 400 mg/kg diet) on the specific binding of [~H]-CHA, [~H]-DPX and [ H]-DZP to mouse whole brain membranes. The specific binding of the different ligands is expressed as a percentage of the specific binding obtained at the same time in controls receiving a caffeine-free diet. For each group the mean value of the specific binding was calculated from the results of 6-8 single-point analysis. The statistical significance (t-test) was calculated from the absolute values of the specific binding (pmol/mg protein) for each caffeine-enriched diet compared to the control values. Six to eight animals were used at each time for each ligand
in each of the three groups.
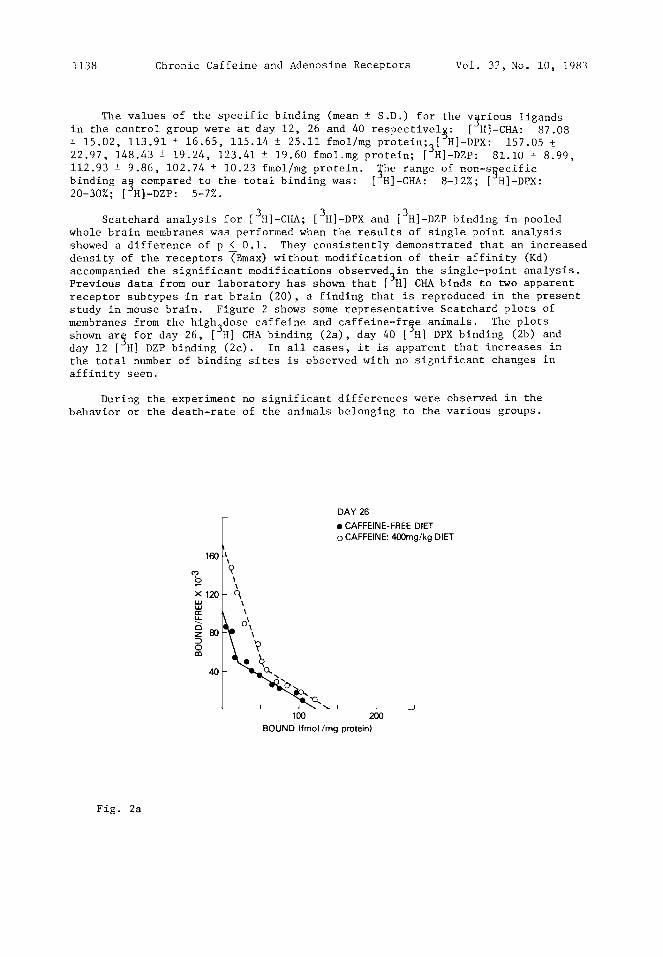
1138 Chronic Caffeine and Adenosine Receptors Vol. 32, No. i0, 1983
The values of the specific binding (mean ± S.D.) for the v~rious ligands in the control group were at day 12, 26 and 40 respectivel~: [ H]-CHA: 87.08 ± 15.02, 113.91 ± 16.65, 115.14 ± 25.11 fmol/mg protein;R[~H]-DPX: 157.05 ± 22.97, 148.43 ± 19.24, 123.41 ± 19.60 fmol.mg protein; [~H]-DZP: 81.10 ± 8.99, 112.93 ± 9.86, 102.74 ± 10.23 fmol/mg protein. The range of non-s~ecific binding a~ compared to the total binding was: [JH]-CHA: 8-12%; [~H]-DPX: 20-30%; [~H]-DZP: 5-7%.
Scatchard analysis for [3H]-CHA; [3H]-DPX and [3H]-DZP binding in pooled whole brain membranes was performed when the results of single point analysis showed a difference of p < 0.i. They consistently demonstrated that an increased density of the receptors ~Bmax) without modification of their affinity (Kd) accompanied the significant modifications observed3in the single-point analysis. Previous data from our laboratory has shown that [ H] CHA binds to two apparent receptor subtypes in rat brain (20), a finding that is reproduced in the present study in mouse brain. Figure 2 shows some representative Scatchard plots of membranes from the high^dose caffeine and caffeine-free animals. The plots shown ar~ for day 26, [~H] CHA binding (2a), day 40 [JH] DPX binding (2b) and day 12 [ H] DZP binding (2c). In all cases, it is apparent that increases in the total number of binding sites is observed with no significant changes in affinity seen.
During the experiment no significant differences were observed in the behavior or the death-rate of the animals belonging to the various groups.
f X 120
,,=
z 80
o 133
40
DAY 26
• CAFFEINE-FREE DIET o CAFFEINE: 400mg/kg DIET
o~
i
100 200
BOUND (fmol/mg protein)
Fig. 2a

Vol. 32, No. i0, 1983 Chronic Caffeine and Adenosine Receptors 1139
200
X ,,=,
100
o o3
DAY 12 • CAFFEINE-FREE DIET O CAFFEINE: 400mg/kg DIET
\\o \ \
"~ ~",,, " ~ . o',\
i I \ l IN J 200 400
BOUND (fmol/mg protein)
Fig. 2b
DAY 40 F CAFFEINE-FREE DIET
80L o CAFFEINE: 400mg/kg DIET
/ o }
X 60~-. O
4o- ~
20
100 2(10 300 41]0
BOUND (fmol/mg protein)
Fig. 2c Fig. 2 Scatchard analysis of [3H]-CHA (2a), [~-DPX (2b) and [3H]-DZP (2c) bindin$ to whole brain membranes in mice chronically receiving either a caffeine-free diet or a caffeine-enriched diet (400 mg caffeine/kgodiet). The kinetic parameters of the plots shown are, respectively, 2a) [JH]-CHA, day 26. Controls: Kd = 0.73 and 3.96 nM, Bmax = 40 and 85 fmol/mg protein, (total = 125); caffeine-dietR = Kd = 0.85 and 3.93 riM, Bmax = 73 and 72 fmol/mg
protein, (total = 145); 2b [~H]-DPX, day 40. Control: Kd 5.59 nM, Bmax = 286 f~ol/mg protein; caffeine-diet: Kd = 6.60 nM, Bmax = 417 fmol/mg protein; 2c) [ H]-DZP, day 12. Control: Kd = 2.06 nM, Bmax = 284 fmol/mg protein; caffeine diet: Kd = 2.14 nM, Bmax = 413 fmol/mg protein.

1140 Chronic Caffeine and Adenosine Receptors Vol. 32, No. i0, 1983
Discussion
Caffeine, a potent CNS stimulant, is widely consumed in the United States where its daily intake is approximately 200 mg per adult (i). Chronic caffeine consumption is extremely common and may induce dependence and tolerance, especially on those consuming doses higher than 500 mg/day (14). However, little attention has been paid to the long-term biochemical effects of its administration in animals. In our experiment, mice were given chronic diets enriched with either 200 or 400 mg of caffeine/kg diet. Assuming that the mice were eating 5g of food per day, a rough estimate of their caffeine con- sumption would be, respectively, 50 and i00 mg/kg/day. Such a dose, however, was administered over a 24 hour period, a factor which needs to be taken into account when interpreting the results of this study. Similar doses adminis- tered all at once are known to induce behavioral stimulant effects in mice (23,24). Furthermore, chronic oral doses up to 300 mg/kg/day administered in the diet do not have any toxic effect on mice (25), a fact explaining the absence of differences in lethality observed between our different groups of animals. ~#hen correction is made for surface area (i), these doses would be equivalent to the daily consumption of 4 or 8 cups of coffee containing i00 mg of caffeine each in humans.
In this report, we describe preliminary evidence that chronic con- sumption of moderate to high non-toxic doses of caffeine is able to induce a dose-related increase in the number of both central adenosine and benzodiaze- pine receptors. Recent experiments have implied that a variety of the central stimulant effects of acute caffeine administration were related to a blockade of adenosine receptors (2,3) and that MX were more potent at competing for adenosine receptors than in mediating any other biochemical effect (26). Thus, a prolonged blockade of these receptors is likely to induce a long-lasting decrease in adenosine interaction with its receptors and a compensatory in- crease in the receptors' number or sensitivity (27). Such a phenomenon has already been documented for dopaminergic receptors when submitted to a chronic blockade by neuroleptic drugs (28). Preliminary results suggest that in vivo, an increased sensitivity of the peripheral adenosine receptors may develop during chronic oral caffeine consumption in rats, as demonstrated by the greater blood pressure drop induced by the same adenosine levels after with- drawal than during caffeine administration (9). Chronic administration of another MX, theophylline, induces an increase in the number of [~H]-CHA binding sites in rat cortex (8).
In our experiment the d~screpancy existing in the time-course and extent of the binding increase of [~H]-CHA and [~H]-DPX may indicate that these ligands bind to different sub-categories of adenosine receptors as already suggested by various ~nvestigators (17,19,30). Furthermore, caffeine's p~tency to displace [ H]-CHA binding is usually higher than that to displace [~H]-DPX binding (7,17), a result which may contribute to the differences observed between the two ligands used in our study. Modifications of adenosine release, as those demonstrated after acute caffeine administration (31), may explain the limited extent of the receptor changes observed here, together with the possible presence of residual caffeine. Further experiments are thus needed in order to determine the exact mechanism of this adenosine binding increase by chronic caffeine consumption and its precise localization.
In our experiment, chronic caffeine consumption was also able to induce a transitory increase in the number of brain [~H]-DZP receptors without modification of their affinity. Caffeine, similar to other MX, is known to antagonize several pharmacological effects of BDZ (5,32) and to inhibit BDZ binding in vitro (4) at concentrations (i0- M) at least one order of magnitude

Vol. 32, No. i0, 1983 Chronic Caffeine and Adenosine Receptors 1141
higher than those inhibiting adenosine receptor binding (10-5M) (2,6). However, we demonstrate here that in vivo the relative potency of caffeine as an effector of the adenosine and BDZ receptors is apparently closer_than that ~ocumented by in vitro studies, since a marked increase of both [3H]-DZP and [ H]-CHA binding wa-sso-bserved at day 12.
Recent experiments comparing caffeine- and pentylenetetrazole (PTZ)- induced seizures and their antagonism by inosine and BDZ suggest that these compounds may both act at the level of the BDZ receptor (5). Importantly, repeated administrations of PTZ, which also competitively inhibits DZP binding in vivo (5), induce a significant increase in mouse forebrain [~H]-DZP binding without modification of its affinity for the receptor (33). Although the prolonged blockade of B~Z receptors by caffeine is likely to play a major role in the increased [JH]-DZP binding, modifications of the concentrations of their putative endogenous ligands by caffeine cannot be ruled out. Among others, purines have been implicated as being possible low affinity ligands or modulators for the BDZ receptors (34) and some of their derivatives, like EMD 28422, are known to induce an increase in the number of BDZ receptor sites without modification of their affinity (35).
The increase in number of both central adenosine and BDZ receptors docu- mented in this study may be relevant to understand the mechanisms underlying the development of both tolerance to (10-12,15) and dependence on (9,14,16) the effects of chronically administered caffeine.
References
i. M. GILBERT, Research Advances in Alcohol and Drug Problems, Vol. 3, pp. 49-176, J. Wiley, New York (1976).
2. S.H. SNYDER, J.J. KATIMS, Z. ANNAU, R.F. BRUNS and J.W. DALY, Proc. Natl. Acad. Sc., U.S.A. 78 3260-3264 (1981).
3. F.L. ELRIDGE, D.E.-MILHORN and T.G. WALDROP, Fed. Proc. __41 1690 (1982). 4. p.J. MARANGOS, S.M. PAUL, A.M. PARMA, F.K. GOODWIN, K. SYAPIN and
P. SKOLNICK, Life Sci. 24 851-858 (1979). 5. p.J. MARANGOS, A.M. MARTI--NO, S.M. PAUL and P. SKOLNICK, Psychopharma-
cology 72 269-273 (1981). 6. J.P. BOUL---ENGER, J. PATEL and P.J. MARANGOS, Neurosci. Lett. 30 161-166
(1982). 7. S.M. PAUL, P. SYAPIN, B. PAUGH, V. MONCADA and P. SKOLNICK, Nature 281
688-689 (1979). 8. T.F. MURRAY, Europ. J. Pharmacol. __82 113-114 (1982). 9. R.W. VON BORSTEL, Fed. Proc. 41 1669 (1982).
i0. J.M. CARNEY, Br. J. Pharmacol.---75 451-454 (1982). ii. F.A. HOLLOWAY, H.E. MODROW and D.--C. BIRD, Soc. Neurosci. Abstr. ii 924
(1981). 12. M.L. WAYNER, F.B. JOLICOEUR, D.B. RONDEU and F.C. BARONE, Pharmacol.
Biochem. Behav. 5 343-348 (1976). 13. T. COLTON, R.E. ~OSSELIN and R.P. SMITH, Clin. Pharmacol Ther. 9 31-39
(1968). 14. J.F. GREDEN, Substance Abuse. Clinical Problems and Perspectives,
J.H. Lowinson and P. Ruiz (eds.), pp. 274-286, Williams and Wilkens,
Baltimore (1981). 15. D. ROBERTSON, D. WADE, R. WORKMAN, R.L. WOOSLEY and J.A. OATES. J. Clin.
Invest. 67 1111-1117 (1981). 16. B.C. WHIT--E, C.A. LINCOLN, N.W. PEARCE, R. REEB and C. VAIDA, Science 209
1547-1548 (1980). 17. R.F. BRUNS, J.W. DALY and S.H. SNYDER, Proc. Natl. Acad. Sci., U.S.A. 77
5547-5551 (1980).

1142 Chronic Caffeine and Adenosine Receptors Vol. 32, Noo i0, 1983
18. M.E. NEWMAN, R. DeLUCIA, J. PATEL and H. MclLWAIN, Biochem. Soc. Transl. 8 141-142 (1980).
19. U. SCHWABE, H. KIFFE, C. PUCIISTEIN and T. TROST, Naunyn. Schmiedebergs Arch. Pharmacol. 310 59-67 (1979).
20. J. PATEL, P.J. MAPJ~NGOS, J. STIVERS and F.K. GOODWIN, Brain Res. 237 203-214 (1982).
21. P.J. MARANGOS and A.M. MARTINO, Molec. Pharmacol. 20 16-21 (1981). 22. O.H. LOWEY, N.J. ROSEBRAUGH, L. FARR and R.J. RANDALL, J. Biol. Chem. 193
265-275 (1951). 23. J.R. BAISSIER and P. SIMON, Arch. Int. Pharmacodyn. Ther. 158 212-221
(1965). 24. J.R. BAISSIER and P. SIMON, Arch. Int. Pharmacodyn. Ther. 166 362-369
(1964). 25. S.S. EPSTEIN, W. BASO, E. ARNOLD and Y. BISHOP, Food Cosmet. Toxicol. 8
381-401 (1970). 26. J.W. DALY, R.F. BRUNS and S.H. SNYDER, Life Sci. 28 2083-2097 (1981). 27. D.H. OVERSTREET and H.I. YAMAMURA, Life Sci. 25 1865-1878 (1979). 28. D.R. BURT, I. CREESE and S. SNYDER, Science 196 326-328 (1977). 29. T.V. DUNWIDDIE and T. WORTH, J. Pharmacol. Exp. Ther. 220 70-76 (1982). 30. K.M.M. MURPHY and S.H. SNYDER, Soc. Neurosci. Abstr. ii 425 (1981). 31. T.W. STONE, C. HOLLINS and H. LLOYD, Brain Res. 207 421-431 (1981). 32. P. POLC, E.P. BONETTI, L. PIERI, R. CUMIN, R.M. ANGIOL, H. MOHLER and
W.E. HAEFELY, Life Sci 28 2265-2275 (1981). 33. P.J. SYAPIN and D.W. RICKMAN, Eur. J. Pharmacol. 72 117-120 (1981). 34. P.J. MARANGOS, S.M. PAUL, F.K. GOODWIN and P. SKOLNICK, Life Sci. 25
1093-1102 (1979). 35. P. SKOLNICK, K.L. LOCK, S.M. PAUL, P.J. MARNAGOS, R. JONES and K. IRMSHER,
Eur. J. Pharmacol. 67 179-186 (1980).