chironomidae (diptera) species distribution related to
TRANSCRIPT

Western North American Naturalist Western North American Naturalist
Volume 60 Number 1 Article 4
1-20-2000
Chironomidae (Diptera) species distribution related to Chironomidae (Diptera) species distribution related to
environmental characteristics of the metal-polluted Arkasas River, environmental characteristics of the metal-polluted Arkasas River,
Colorado Colorado
L. P. Ruse Environment Agency (Thames Region), Reading, England
S. J. Herrmann University of Southern Colorado, Pueblo, Colorado
J. E. Sublette Tucson, Arizona
Follow this and additional works at: https://scholarsarchive.byu.edu/wnan
Recommended Citation Recommended Citation Ruse, L. P.; Herrmann, S. J.; and Sublette, J. E. (2000) "Chironomidae (Diptera) species distribution related to environmental characteristics of the metal-polluted Arkasas River, Colorado," Western North American Naturalist: Vol. 60 : No. 1 , Article 4. Available at: https://scholarsarchive.byu.edu/wnan/vol60/iss1/4
This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Western North American Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected].

The Arkansas River in Colorado has beenpolluted by heavy metals since mining beganin 1859. Remedial action on the most affectedsites started in 1991. There have been manydescriptive and experimental studies of pollu-tion effects on benthic macroinvertebratesinhabiting the first 30 km of the river byresearchers of the Bureau of Reclamation andColorado State University (e.g., Roline andBoehmke 1981, Roline 1988, Kiffney andClements 1993, Clements 1994, Clements andKiffney 1994). Typically, invertebrates weresampled using mesh sizes of 500 µm or greaterand Chironomidae (non-biting midges) werenever identified beyond the subfamily level.Armitage and Blackburn (1985) demonstratedthat specific identification of Chironomidaedistinguished varying degrees of metal pollu-tion as efficiently as using all macroinverte-brate data with chironomids identified only tosubfamily. Clements (1994) has accepted thatresearch on metal tolerances of orthocladiinespecies (a subfamily of Chironomidae) is nec-essary for the Arkansas River. The collectionand specific identification of Chironomidae
can be made easier and more efficient by sam-pling pupal exuviae, compared with larvae (Fer-rington et al. 1991). Although exuviae willremain afloat for 2–3 d after adult emergence,they do not drift far before entrapment at rivermargins or midstream obstacles (McGill 1980,Ruse 1995a). Exuvial collections should there-fore be representative of local adult emergence,integrated over a few days before sampling.
In 1983 a major surge of metal sludge in theUpper Arkansas River affected sites 220 kmdownstream (Kimball et al. 1995). Emergingadult chironomids, and later pupal exuviae,were collected from sites along this length ofthe Arkansas River during 1984–85 to investi-gate the effects of metal pollution on speciesspatial distribution. At that time many individ-uals could not be identified to species, particu-larly pupal exuviae. Associations between lar-vae, pupae, and adults from rivers in Coloradoand neighboring states have since enabled spe-cific identification (Sublette et al. 1998). Thishas led to a retrospective investigation of therelationship between species distribution andavailable environmental data using statistical
Western North American Naturalist 60(1), pp. 34–56
CHIRONOMIDAE (DIPTERA) SPECIES DISTRIBUTION RELATED TO ENVIRONMENTAL CHARACTERISTICS OF THE METAL-POLLUTED ARKANSAS RIVER, COLORADO
L.P. Ruse1, S.J. Herrmann2, and J.E. Sublette3
ABSTRACT.—Mining in the Upper Arkansas catchment has polluted the river with heavy metals for 140 yr. Pupal andadult chironomid species distribution and sedimentary metal concentrations are provided for 22 stations along 259 km ofmain river during 1984–85. Complete species identification was achieved only recently. This has produced an unprece-dented record of chironomid species distribution for a comparable length of river in the USA. Chemically or physicallyperturbed sites had poor species richness compared with the next site downstream, suggesting that larvae may driftthrough unfavorable habitats to benign ones. Using canonical correspondence analysis, we found species composition tobe most strongly related to variables expressing the longitudinal axis of the river (distance/altitude, temperature, latitude),while toxicity to zinc was a significant secondary correlate. These river-related environmental variables accounted for agreater proportion of pupal species variation than for adults. This was considered to result from a proportion of adultsemerging from habitats beyond the main river. Multivariate analysis identified metal-tolerant and -intolerant species.Generic data revealed the same major trends but indicator taxa were lost. The study provides a disturbed-state referencefor monitoring effects of remedial actions begun in 1991, and for comparisons with other Colorado rivers.
Key words: Chironomidae, heavy metals, multivariate analysis, pupal exuviae, adults, spatial distribution, sediments,species richness.
1Environment Agency (Thames Region), Fobney Mead, Rose Kiln Lane, Reading RG2 0SF, England.2University of Southern Colorado, 2200 Bonforte Boulevard, Pueblo, CO 81001, USA.33550 North Winslow Drive, Tucson, AZ 85750, USA.
34

packages that were not available during thesurvey period. This study also differed fromother research on the Arkansas River by relat-ing invertebrate distribution to sedimentaryconcentrations of heavy metals rather thanwater measurements. Kiffney and Clements(1993) found that suspended metal concentra-tions in the Arkansas River underestimatedavailability of metals to benthic macroinverte-brates. Bioaccumulated metal concentrationswere better related to those measured in sedi-mentary minerals and periphyton. This surveyprovides the only reference for measuring theeffect of subsequent remedial actions on thechironomid assemblage of the Arkansas Riverand relating their distribution to sedimentarymetal concentrations during a period of severepollution.
METHODS
Study Sites
Twenty-two sites were chosen along 259km of the East Fork (EF) and Arkansas River(AR) between Climax and Pueblo, east of theContinental Divide in central Colorado (Fig.1). We adopted sites EF1 downstream to AR9from those surveyed by the Bureau of Recla-mation and reported by Roline (1988). Otherbiological surveys of the Upper Arkansascatchment have adopted the same site codes,but since these may refer to different loca-tions, care should be taken when cross-refer-encing with previous publications.
Metal-rich water enters East Fork betweenEF1 and EF2 via Leadville Drain, but thegreatest source of metals to the catchmentcomes from California Gulch between AR2and AR3 (Kimball et al. 1995). This surveyoccurred between 2 major metal sludge surgesinto California Gulch on 23 February 1983 and22 October 1985. Water diverted from thewestern slopes of the Continental Divide sup-plements flows from Turquoise Lake and TwinLakes, entering the Arkansas River above AR4and AR9, respectively. Iowa Gulch, and dif-fuse sources of metals between AR4 and AR8,carried discharge from an active mine duringthe study period. Mining affects other tribu-taries to the river downstream of AR8, butconcentrations of metals are much lower thanthose found upstream. The Arkansas Riverwas impounded above AR19 by the PuebloDam in 1974. Sediment analysis of Pueblo
Reservoir reveals that a substantial metal loadis transported there from the Leadville area,particularly due to resuspension of river sedi-ments by snowmelt runoff (Kimball et al. 1995).The U.S. Environmental Protection Agency(EPA) declared the California Gulch catch-ment and the Arkansas River from above AR2to below AR3 a Superfund site in 1983. Newwater treatment plants on the Leadville Drainand California Gulch were in operation byJune 1992, and the last major mining opera-tion in Leadville ceased in January 1999.
Biological Data
We collected adult Chironomidae at eachsite monthly from May 1984 until September1985 using sweep net, beating sheet, water-skimming, hand-picking and ultraviolet lighttraps. Adults were dissected in absoluteethanol. Body parts, except for wings and 1 setof legs, were cleared in potassium hydroxideand then all parts slide-mounted in Euparal.Adult Plecoptera and Trichoptera were alsocollected and are reported in the followingpaper (Ruse and Herrmann 2000).
We sampled chironomid pupal exuviae usingthe “Thienemann net technique” (Thiene-mann 1910): a 200-µm-mesh net attached to acircular frame on a pole is used to collect float-ing debris accumulating behind obstacles atriver margins. This method supplementedadult collections during a 3-month visitingscholarship by the senior author. Each site wassampled in July, August, and September 1985.The broad emergence period by many tem-perate, lotic species of Chironomidae shouldensure that a large proportion of species pre-sent over the whole year are represented bythis frequency of sampling (Ruse and Wilson1984, Ruse 1995b). Samples were refloated,agitated, and randomly subsampled by sieve.All chironomid pupal exuviae were removedfrom a subsample and sufficient subsampleswere sorted to obtain about 200 exuviae, whenpossible. Exuviae were mounted on glass micro-scope slides in Euparal or retained in vials of70% ethanol. Initially identified to genericlevel, the material remained in excellent con-dition until 12 yr later when it became possi-ble to determine species. Specific identifica-tion was achieved by comparing exuviae withthose obtained from adult rearings of larvaeand pupae collected subsequently from theArkansas River and neighboring catchments in
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 35

Colorado and New Mexico. The associatedmaterial is held by author JES. Unassociatedpupal species are designated by the suffix n-P.
Environmental Data
At each site water temperature was recordedonce during each monthly visit to collect adultinsects. The 3 dominant superficial substra-
tum types, among 5 size classes, were assessedvisually. Latitude, longitude, altitude, slope, anddistance downstream from EF1 were obtainedfrom maps.
We determined metals from 2 samples ofsubmerged fine sand taken at each site during18–19 October following the 2nd metal sludgesurge into California Gulch. These data still
36 WESTERN NORTH AMERICAN NATURALIST [Volume 60
Fig. 1. Upper Arkansas River sampling points.

served to characterize the relative contamina-tion of sites by metals emanating from Lead-ville mines. A 25-mm-diameter PVC pipe wasinserted to a depth of 15 cm. Sediments weredried at 70°C for 48 h and ground with a mor-tar and pestle until they passed through a 250-µm-mesh sieve. Metals were extractedfrom triplicate subsamples of approximately500 ± 0.1 mg using a sequence of hot diges-tions and evaporations with nitric and hydro-chloric acids (Caravajal et al. 1983). A reagentblank was prepared before and after each setof 6 sediment digestions for a site and takenthrough the same protocol prior to metalsdetermination. Determination of lead (Pb), iron(Fe), manganese (Mn), zinc (Zn), and copper(Cu) by flame atomic absorption spectrometryfollowed the methods of Mahan et al. (1987).Cadmium (Cd) was measured by electrother-mal atomization atomic absorption spectrome-try (Sandoval et al. 1992). The mean concen-tration of 6 samples from each site was used insubsequent data analysis.
Data Analysis
Species abundances for samples from thesame site were combined for both pupal andadult data sets so they could be related toenvironmental characteristics recorded on onlya single occasion. Spatial variation in thesedata sets was directly compared with environ-mental variation using canonical correspon-dence analysis (CCA; Ter Braak and Prentice1988). CCA selected the linear combination ofenvironmental variables achieving the maxi-mum separation of species by multiple regres-sion along the 1st axis. Subsequent axes wereextracted from the residual variation to maxi-mize dispersion of species, provided theywere uncorrelated to previous axes. Signifi-cance of the regression between biological andenvironmental data was tested against the pos-sibility of a random association by comparingthe F-ratio with 99 unrestricted Monte Carlopermutations of these data (Ter Braak 1990). Aprobability of ≤0.05 was considered signifi-cant. Forward stepwise regression was used toobjectively select variables, one at a time,according to the amount of biological variationeach explained. Selection stopped when therewas no significant increase in explained varia-tion, tested against Monte Carlo permutations.
Before analysis, we converted chironomidspecies abundances to percentages of the total
number of individuals identified from a site.Species recorded at 1 site only were omittedfrom CCA in case of spurious association witha coincidental extreme environmental mea-surement; their distributions are recorded inthe Appendix. An ordinal value representingrelative variation in substratum between siteswas obtained by assuming a mean particle sizefor each of 3 categories: boulder/bedrock (215mm), rubble/gravel (9.5 mm), and sand (0.25mm). The dominant substratum was assumedto cover 50% of the site, and the next 2 re-corded substrata were assumed to cover 30%and 20% of the site, respectively. Mean parti-cle size at each site was calculated from thesum of products of size times proportionalcoverage. To account for the ameliorating effectof increased hardness on metal toxicity tobiota, we calculated EPA hardness-based waterquality criterion for Zn (Clements and Kiffney1995). Water hardness was not measured dur-ing this survey, but data were available forsites EF1 to AR9 (Roline and Boehmke 1981,Clements and Kiffney 1995) and for inlet andoutlet flows of Pueblo Reservoir (Herrmannand Mahan 1977). The presence of carbonaterocks between AR10 and AR12 and river-exposed deposits of calcium and magnesiumnear AR16 (Kimball et al. 1995) was also takeninto account when estimating water hardness.For each site, we divided the observed sedi-mentary Zn-loading by the criterion value forassumed water hardness. Resultant ratioswere classified into an ordinal scale of toxicityto Zn: <2.0 = 1, 2.0–9.9 = 2, 10.0–19.9 = 3,20.0–39.9 = 4, >39.9 = 5. These broad bandsreduced the effect of imprecise hardness esti-mates. Environmental data were not trans-formed for CCA; measurements of tempera-ture, slope, Zn toxicity, total Mn, and total Fewere normally distributed. Latitude and longi-tude values were decimalized and only themaximum water temperature recorded at eachsite was used. Environmental data were stan-dardized to have a mean of zero and unit vari-ance to remove arbitrary variation in units ofmeasurement. CCA species scores wereweighted mean sample scores (CANOCO ver-sion 3.1 scaling + 2). The analysis was there-fore sensitive to relative variation betweensites, and it was not necessary to have precisedata on particle size or water hardness torelate these characteristics to trends in speciesdistribution.
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 37

Direct statistical comparisons of pupal andadult species proportions were made using aχ2 test of independence (Sokal and Rohlf 1981).The null hypothesis was that proportions ofeach species collected were independent ofsampling method, aquatic netting, or aerialnetting. Pupal species unassociated with rearedadults were excluded, as were species with ex-pected counts <5 in both data sets.
RESULTS
Environmental Data
The obtuse-angled line of the main riverprevented latitude or longitude having thesimple linear relationship with distance thataltitude had (Table 1). The river gradient wasreduced at the last 3 sites, but the trend wasvariable along most of the watercourse. Meanparticle size at the first 11 sites was oftensmaller than at downstream sites. Site AR10was characterized by a steep gradient and tor-rential flow over a substratum dominated bybedrock, boulder, and rubble. Maximum re-corded temperatures increased downstream toAR7 but were suppressed below the Twin Lakesconfluence until AR13. Hypolimnion flowsfrom Pueblo Reservoir lowered temperatureat AR19. Sedimentary total Cu was the onlymetal to reach a peak at AR3, below CaliforniaGulch, while the next most Cu-contaminatedsites were AR5 and AR7. Zn toxicity, total Zn,Mn, and Cd peaked at AR5, AR7, or AR8, allreduced-gradient sites compared with AR3,AR4, and AR6. Concentrations of sedimentaryFe below California Gulch remained highthroughout the river, except at AR12 andAR19, peaking at AR11.
Pupal Exuviae
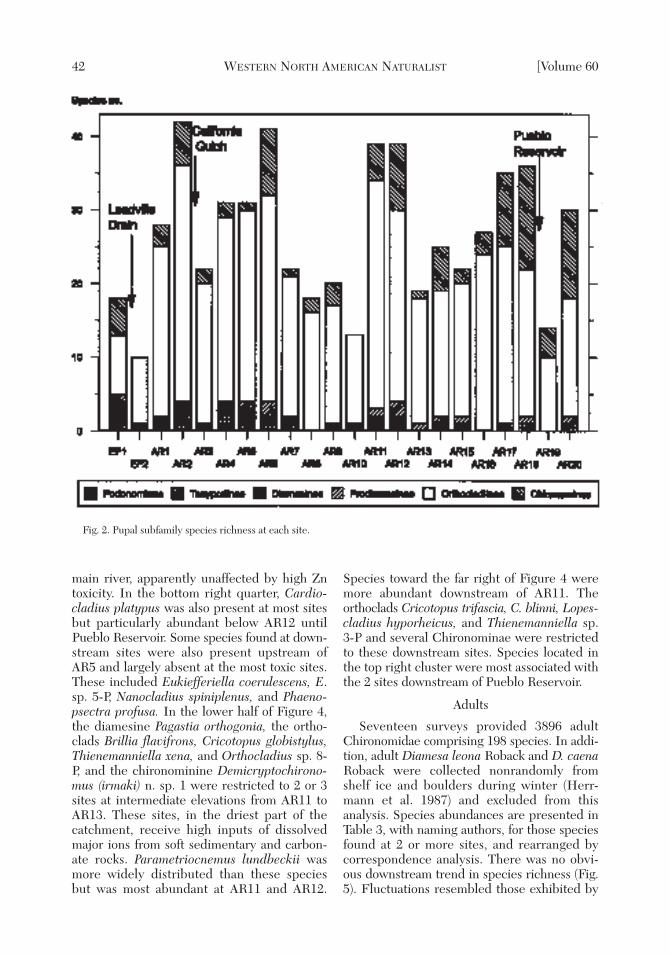
A total of 10,120 chironomid pupal exuviaewere identified to 127 species from 22 sites.Species abundances are presented in Table 2,with authors’ names, for species collected from2 or more sites. Species and sites in Table 2are arranged according to the 1st axis of a cor-respondence analysis (Ter Braak and Prentice1988) so that downstream turnover in speciescomposition can be assessed. Species richnesswas lowest at EF2, below Leadville Drain, thenincreased downstream to the richest site atAR2, above the confluence with CaliforniaGulch (Fig. 2). Species richness was poor atAR3, recovered at the 3 sites below Turquoise
Lake confluence, and then declined until AR10.Species numbers were high at AR11–AR12and depleted below Pueblo Reservoir atAR19. Orthocladiinae was the dominant sub-family throughout the survey. There were noobvious downstream trends in total or subfam-ily species richness except for the absence ofDiamesinae below AR12. Classifying pupalexuviae according to presumed feeding modesof their associated larvae (Table 2) revealed adominance by algal grazers at all sites (Fig. 3).Predators increased from AR13 until PuebloReservoir. Detritivores were present in lowproportions except at AR10. Filterers appearedfrom AR16 to AR18.
ORDINATION.—Stepwise regression selecteddistance downstream, maximum temperature,latitude, and Zn toxicity as significantly corre-lated with variation in species compositionamong sites. Altitude was also significant buthighly correlated with distance and was ex-cluded to prevent multicollinearity (variationinflation factor = 189; Ter Braak 1990). The 4selected variables explained 43.4% of biologi-cal variation in CCA. The species-environ-ment relationship was significantly differentfrom random for the first 2 CCA axes (P =0.01), accounting for 32.9% of all biologicalvariation and 75.7% of explained variation.
Species turnover among samples wasstrongly related to change along the longitudi-nal axis of the river. Dominance of the 1stCCA axis compared with the 2nd resulted inan archlike configuration of sites in Figure 4.Gradient lengths for the first 2 unconstrained(biological data alone) axes were 6.24 and 2.82s units, respectively. Detrending or reductionof environmental variables did not remove thearching trend, and separation into 2 data setswas impractical for the small number of sam-ples. The 1st CCA axis was most significantlyrelated to downstream distance (canonical co-efficient t-value 5.42, interset correlation 0.97).The 2nd axis was principally related to varia-tions in maximum temperature (t-value 6.05,correlation –0.46) and Zn toxicity (t-value 2.77,correlation –0.29), resulting in lateral spread-ing of samples upstream of AR9, at AR13, andbelow Pueblo Reservoir. Sites EF2 and AR3,downstream of the most significant metalinputs of Leadville Drain and California Gulch,respectively, were closely associated. SitesAR5–AR8 had the highest Zn toxicity ratiosand similar species composition, although AR5
38 WESTERN NORTH AMERICAN NATURALIST [Volume 60

2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 39
TAB
LE
1. E
nvir
onm
enta
l dat
a; m
ean
tota
l met
al c
once
ntra
tions
are
µg
g–1
dry
wei
ght
Site
Lat
itude
Lon
gitu
deM
ax.
Alti
tude
Slop
eD
ist.
Zin
cTo
tal
Tota
lTo
tal
Tota
lTo
tal
Tota
lM
ean
(Deg
.)(D
eg.)
tem
p.(m
)(%
)(k
m)
tox.
C
uZ
nPb
Mn
FeC
dpa
rtic
le(°
C)
size
(mm
)
EF
139
.28
106.
2213
.030
421.
10.
001
13.9
132
19.0
374
8981
0.40
2.9
EF
239
.27
106.
3313
.629
691.
16.
352
10.5
935
88.9
824
8014
0.84
4.6
AR
139
.25
106.
3214
.529
441.
67.
872
6.0
548
69.8
824
6376
0.90
4.6
AR
239
.23
106.
3514
.229
051.
411
.11
24.
832
041
.060
257
730.
574.
6A
R3
39.2
210
6.35
14.4
2899
1.4
11.1
83
157.
023
7477
9.0
825
3040
02.
974.
6A
R4
39.2
010
6.35
16.1
2865
1.0
14.4
83
39.7
917
267.
073
012
570
1.37
4.6
AR
539
.17
106.
3317
.428
350.
520
.49
580
.028
3686
5.0
780
3100
03.
504.
6A
R6
39.1
310
6.32
18.1
2795
1.7
22.8
64
46.6
1679
451.
011
4915
760
2.70
4.6
AR
739
.12
106.
3018
.627
710.
825
.91
572
.630
3876
3.0
1017
3010
04.
234.
6A
R8
39.0
810
6.28
17.8
2748
0.7
29.0
84
47.6
2392
582.
014
7418
570
1.65
24.0
AR
939
.07
106.
2816
.027
430.
430
.35
216
.550
817
6.0
368
1236
00.
834.
6A
R10
38.9
710
6.20
15.5
2573
1.1
45.8
52
14.0
420
161.
459
011
350
0.73
115.
0A
R11
38.7
810
6.08
16.5
2338
0.9
71.7
52
21.7
401
112.
747
332
170
0.58
21.9
AR
1238
.53
106.
0217
.521
430.
610
4.77
26.
813
539
.012
369
100.
5221
.9A
R13
38.4
310
5.82
19.4
2033
0.5
129.
282
15.5
263
59.6
256
2074
00.
8221
.9A
R14
38.4
010
5.58
19.2
1879
0.6
156.
592
17.6
269
36.3
448
2737
00.
5021
.9A
R15
38.4
710
5.40
19.9
1746
0.6
177.
802
19.0
309
59.1
438
3007
00.
7321
.9A
R16
38.4
310
5.25
21.5
1618
0.7
195.
452
18.0
332
39.5
399
2372
00.
3521
.9A
R17
38.3
110
5.00
21.5
1535
0.4
217.
801
9.8
129
12.9
229
1869
00.
383.
2A
R18
38.2
610
4.92
21.7
1497
0.3
228.
981
11.0
198
7.8
290
1241
00.
783.
2A
R19
38.1
910
4.70
19.6
1444
0.2
253.
741
4.8
281.
084
6810
0.25
45.3
AR
2038
.19
104.
6721
.014
310.
325
8.56
16.
368
10.2
143
1882
00.
084.
6

was closer to sites downstream of outflowsfrom Turquoise Lake and Twin Lakes (AR4,AR9, and AR10).
Species toward the top left of Figure 4 weremost abundant at, or restricted to, upstreamsites. Diplocladius cultriger was present belowCalifornia Gulch but absent from the mostcontaminated sites. Several Orthocladius species
were associated with high sedimentary metal-loadings. Krenosmittia camptophleps, whichlives among coarse gravel, was found aboveand below California Gulch but was absent atsites with the highest sedimentary Zn-load-ings. Other species with an upstream distribu-tion and which may be sensitive to high sedi-mentary Zn concentrations were Eukiefferiella
40 WESTERN NORTH AMERICAN NATURALIST [Volume 60
TABLE 2. Proportions of pupal exuviae species at each site: 1 = 0.1–4.9%; 2 = 5.0–9.9%; 3 = 10.0–19.9%; 4 = 20.0–39.9%;5 = 40.0+%. G = Grazer, D = Detritivore, P = Predator, F = Filterer.
TrophicCode Species name group Site
E E A A A A A A A A A A A A A A A A1 1 1 1 1 1 1
1 2 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6
PROC_SUB Procladius subletti Roback P 1 – – 1 – – – 1 – – – – – – – – – –THIE_FUS Thienemannimyia fusciceps (Edwards) P 1 – – 1 – 1 – – – – – – – – – – – –DIAM_HET Diamesa heteropus (Coquillet) G – – – 1 1 2 1 1 – – – – – – – – – –POTT_MON Potthastia montium (Edwards) D 1 – 1 – – – – – – – – – – – – – – –PAGA_PAR Pagastia partica (Roback) D 2 1 1 1 – 1 1 – 1 – 1 – 1 1 – – – –HYDR_FUS Hydrobaenus fuscistylus (Goetghebuer) G 4 5 – 1 2 1 1 1 1 1 – – 1 1 – – – –HYDR_PIL Hydrobaenus pilipes (Malloch) G – – – 1 – – – 1 – – – – – – – – – –DIPL_CUL Diplocladius cultriger Kieffer D – – – 1 1 1 – 1 – – – – – – – – – –EUKI_ILK Eukiefferiella ilkleyensis (Edwards) G – 1 1 2 1 1 1 1 – 1 – 1 1 1 – 1 – –EUKI_2-P Eukiefferiella sp. 2-P G – 1 1 1 2 1 1 1 – – – 1 – 1 – – – –EUKI_n9 Eukiefferiella n. sp. 9 G 1 1 – 3 2 1 1 – – – – – 1 – – 1 – 1ORTH_DUB Orthocladius dubitatus Johannsen G – – – 1 – – 1 – – – – – – – – – – –ORTH_LUT Orthocladius luteipes Goetghebuer G – – 1 1 1 1 – 1 – 1 – – – – – – – –ORTH_APP Orthocladius appersoni Soponis G – – – 1 – – 1 – – – – – – – – – – –ORTH_5-P Orthocladius sp. 5-P G – – – 1 – – – – 1 – – – – – – – – –ORTH_NIG Orthocladius nigritus Malloch G – 3 – 1 1 1 1 – 1 1 – – 1 1 – – – –ORTH_OBU Orthocladius obumbratus Johannsen G – – – 1 1 1 – 1 1 – – – – – – – – –PARA_n3 Paratrichocladius n. sp. 3 G – – 1 – – – – – – – – – 1 – – – – –PSEC_SPI Psectrocladius spinifer (Johannsen) G – – 1 1 – – – 1 – – – – – – – – – –RHEO_EMI Rheocricotopus eminelobus Sæther G – 3 1 1 3 1 1 1 1 1 – 1 1 – – – 1 –TVET_PAU Tvetenia paucunca (Sæther) G – – 4 4 4 1 1 1 1 1 1 – 1 1 – – – 1CORY_LOB Corynoneura lobata Edwards G – – 1 1 – – – 1 – – – – – – – – – –CORY_5-P Corynoneura sp. 5-P G – – – – – 1 – – – – – – 1 – – – – –KREN_CAM Krenosmittia camptophleps (Edwards) G – 1 1 1 1 1 – – – – – – 1 – – – – –THIE_5-P Thienemanniella sp. 5-P G 1 – 1 1 – 1 – – – – – – – – – – – –POLY_n1 Polypedilum n. sp. 1 D 1 – 1 1 1 1 1 – 1 1 – 1 – – – – – –TANY_8-P Tanytarsus sp. 8-P D 1 – 1 – – – – – – – – – – – – – – –TANY_n5 Tanytarsus n. sp. 5 D – – 1 1 – – – – – – – – 1 – – 1 – –BRUN_EUM Brundiniella eumorpha (Sublette) P – – – – – – 1 – 1 – – – – – – – – –CRIC_BIF Cricotopus bifurcatus Cranston & Oliver G – – – – – – – 1 1 – – – – – – – – –CRIC_n18 Cricotopus n. sp. 18 G – – 1 1 – 1 1 2 2 1 1 – 2 1 1 – – –CRIC_19P Cricotopus sp. 19-P G – – – – – – 1 1 – – – – – – – – – –HETE_MAE Heterotrissocladius maeaeri Brundin D – – – – – – 1 1 – – 1 – 1 – – – – –ORTH_FRI Orthocladius frigidus (Zetterstedt) G 4 3 1 1 1 5 4 4 4 5 4 1 1 – – 1 – –KREN_HAL Krenosmittia halvorseni (Cranston
& Oliver) G – – – – – – 1 1 2 1 1 1 – – – – – –SERG_ALB Sergentia albescens (Townes) P 1 – – 1 – – – 2 – – – – – – – – – –CRIC_TRE Cricotopus tremulus (Linnaeus) G – – 1 1 1 1 1 1 1 – 1 2 1 1 1 1 1 1CRIC_SLO Cricotopus slossonae Malloch G – – 2 1 1 – 1 1 1 1 2 1 2 1 2 1 1 1ORTH_RVA Orthocladius rivicola Kieffer G 1 1 4 2 2 4 4 3 3 2 3 4 1 4 3 4 4 4ORTH_MAL Orthocladius mallochi Kieffer G 1 – – 1 1 1 3 3 3 2 2 1 1 2 2 3 1 1ORTH_10P Orthocladius sp. 10-P G – – – 1 – – 1 – – – – – – – – – – 1THIE_1-P Thienemanniella sp. 1-P G – – 2 1 2 1 – 1 – – – – – 1 – – – 1POLY_ALB Polypedilum albicorne (Meigen) D – – – 1 – – – 1 – – – – – 1 – – – –MICR_n6 Micropsectra n. sp. 6 D – – – 1 1 1 – – – 1 1 – – 1 – 1 – –
A A A A1 1 1 27 8 9 0
– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –1 – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –1 – – –– – – –– – – –– – – –
– – – –– – – –– 1 – –1 – – –4 2 – 11 1 – 1– – – –1 1 – –– – – –– – – 1

n. sp. 9 and Tanytarsus n. sp. 5. Toward thebottom left of Figure 4 are species found atsites with highest potential Zn toxicity such asKrenosmittia halvorseni, in contrast to its con-gener. Brundiniella eumorpha may haveoccurred at the most Zn-toxic sites due to thepresence of numerous small springs. Hydro-baenus pilipes is known to emerge in early
spring (Tokeshi 1995) but was collected inAugust at AR2 and AR6. Orthocladius frigiduswas found at all sites upstream of AR12 andwas most abundant in East Fork and from AR4to AR9. Orthocladius mallochi, O. rivicola,Cricotopus slossonae, C. tremulus, and Eukief-feriella claripennis were the most widespreadand evenly distributed species throughout the
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 41
TABLE 2. Continued
TrophicCode Species name group Site
E E A A A A A A A A A A A A A A A A1 1 1 1 1 1 1
1 2 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6
BORE_LUR Boreoheptagyia lurida (Garrett) G – – – – – – – – – – – 2 – – – – 1 –MONO_1-P Monodiamesa sp. 1-P D – – – – – – 1 1 – – – – – 1 – – – –EUKI_CLA Eukiefferiella claripennis (Lundbeck) G – – 1 1 1 1 2 2 3 3 3 4 3 3 2 1 1 1STEN_2-P Stenochironomus sp. 2-P D – – – – – – – 1 – – – – – – – – – –PAGA_ORT Pagastia orthogonia Oliver D – – – – – – – – – – – – 1 1 – – – –BRIL_FLA Brillia flavifrons Johannsen G – – – – – – – – – – – – 1 – 1 – – –CRIC_BIC Cricotopus bicinctus (Meigen) G – – 1 – – – 1 – 1 – – – – – – – – –CRIC_GLO Cricotopus globistylus Roback G – – – – – – – – – – – – 1 1 – – – –THIE_XEN Thienemanniella xena (Roback) G – – – – – – – – – – – – 1 – 1 – – –EUKI_1-P Eukiefferiella sp. 1-P G – – – – – – – 1 1 – – – 1 – 1 1 – 1ORTH_RUB Orthocladius rubicundus (Meigen) G – – – – 1 – – 1 – – 1 – 3 1 1 2 1 1ORTH_8-P Orthocladius sp. 8-P G – – – – – – – – – – – – 1 1 – 1 – –DEMI_n1 Demicryptochironomus (irmaki) n. sp. 1 P – – – – – – – – – – – – 1 1 – 1 – –ODON_FER Odontomesa ferringtoni Sæther D 1 – – – – – – 1 – – – – 1 1 1 1 1 –CARD_PLA Cardiocladius platypus (Coquillett) P – – 1 1 – 1 1 1 1 1 1 2 1 2 4 4 4 4CRIC_HER Cricotopus herrmanni Sublette G – – – 1 – 1 – 1 1 – – – 1 2 1 – 1 1CRIC_INF Cricotopus infuscatus (Malloch) G – – – – – – – – – 1 1 – – – – – – 1EUKI_5-P Eukiefferiella sp. 5-P G – – – 1 – – – – – – – – – – – 1 1 –NANO_SPI Nanocladius spiniplenus Sæther G – – 1 – – 1 1 – – – – – – – – – 1 1PARA_LUN Parametriocnemus lundbeckii
(Johannsen) G – – – – – – 1 1 – – 1 1 4 4 1 2 3 3PHAE_PRO Phaenopsectra profusa (Townes) D 1 – – – – – – – – – – – 1 1 – 1 – –POLY_LAE Polypedilum laetum (Meigen) D – – – – – – – 1 1 – 1 – – 1 1 1 1 1PENT_INC Pentaneura inconspicua (Malloch) P – – – – – – – – – – – – – – – – – –CRIC_ANN Cricotopus annulator Goetghebuer G – – – – – – – – – 1 1 – 1 1 1 1 3 2CRIC_TFA Cricotopus trifascia Edwards G – – – – – – – – – – – – – 1 – – – –CRIC_BLI Cricotopus blinni Sublette G – – – – – – – – – – – – – – 1 – 1 1EUKI_4-P Eukiefferiella sp. 4-P G – – – – – – – – – – – – – – – – 1 –EUKI_COE Eukiefferiella coerulescens (Kieffer) G – – – 1 – 1 – – – – – – – 1 2 1 1 1RHEOCRn1 Rheocricotopus n. sp. 1
(nr. chalybeatus) G – – – – – – – – – – – – – – – – – –TVET_VIT Tvetenia vitraces (Sæther) G – – – – – – – – – – 1 – – 1 – 1 1 1HELE_1-P Heleniella sp. 1-P G – – – – – – – – – – – – – – 1 – – 1LOPE_HYP Lopescladius hyporheicus Coffman
& Roback D – – – – – – – – – – – – – – – – 1 3THIE_3-P Thienemanniella sp. 3-P G – – – – – – – – – – – – – – – – – 2CHIR_DEC Chironomus decorus Johannsen D – – – – – – – 1 – – – – – – – 1 1 –CYPH_GIB Cyphomella gibbera Sæther D – – – – – – – – – – – – – 1 – – – –DICR_FUM Dicrotendipes fumidus (Johannsen) D – – – – – – – – – – – – – 1 – – – –MICR_PES Microtendipes sp. D – – – – – – – – – – – – – – – – – –POLY_PAR Polypedilum parascalaenum Beck D – – – – – – – – – – – – – – – – – –SAET_n1 Saetheria n. sp. 1 D – – – – – – – – – – – – – – – – – –PSEU_PSE Pseudochironomus pseudoviridis
(Malloch) D – – – – – – – – – – – – – – – – – –CLAD_2-P Cladotanytarsus sp. 2-P D – – – – – – – – – – – – – – – – – –RHEO_n4 Rheotanytarsus n. sp. 4 F – – – – – – – – – – – – – – – – – 1
A A A A1 1 1 27 8 9 0
– – – –– – – –1 1 1 –– – – 1– – – –– – – –1 1 1 1– – – –– – – –1 – – –– 1 – –– – – –– – – –– 1 – 13 3 1 11 1 4 11 1 1 1– 1 – 11 – – 1
1 1 – 11 1 1 11 1 – 11 – – 13 3 1 31 1 3 31 1 5 41 – – –1 1 1 1
1 1 – –1 1 – 1– – – –
1 1 – 14 3 – –1 1 – 1– 1 – 1– – 1 1– 1 – 11 1 – 1– 1 – 1
– – 1 1– 1 1 11 2 – –
main river, apparently unaffected by high Zntoxicity. In the bottom right quarter, Cardio-cladius platypus was also present at most sitesbut particularly abundant below AR12 untilPueblo Reservoir. Some species found at down-stream sites were also present upstream ofAR5 and largely absent at the most toxic sites.These included Eukiefferiella coerulescens, E.sp. 5-P, Nanocladius spiniplenus, and Phaeno-psectra profusa. In the lower half of Figure 4,the diamesine Pagastia orthogonia, the ortho-clads Brillia flavifrons, Cricotopus globistylus,Thienemanniella xena, and Orthocladius sp. 8-P, and the chironominine Demicryptochirono-mus (irmaki) n. sp. 1 were restricted to 2 or 3sites at intermediate elevations from AR11 toAR13. These sites, in the driest part of thecatchment, receive high inputs of dissolvedmajor ions from soft sedimentary and carbon-ate rocks. Parametriocnemus lundbeckii wasmore widely distributed than these speciesbut was most abundant at AR11 and AR12.
Species toward the far right of Figure 4 weremore abundant downstream of AR11. Theorthoclads Cricotopus trifascia, C. blinni, Lopes-cladius hyporheicus, and Thienemanniella sp.3-P and several Chironominae were restrictedto these downstream sites. Species located inthe top right cluster were most associated withthe 2 sites downstream of Pueblo Reservoir.
Adults
Seventeen surveys provided 3896 adultChironomidae comprising 198 species. In addi-tion, adult Diamesa leona Roback and D. caenaRoback were collected nonrandomly fromshelf ice and boulders during winter (Herr-mann et al. 1987) and excluded from thisanalysis. Species abundances are presented inTable 3, with naming authors, for those speciesfound at 2 or more sites, and rearranged bycorrespondence analysis. There was no obvi-ous downstream trend in species richness (Fig.5). Fluctuations resembled those exhibited by
42 WESTERN NORTH AMERICAN NATURALIST [Volume 60
Fig. 2. Pupal subfamily species richness at each site.

pupal data except at sites AR5, AR7, andAR20. Species richness fell downstream ofIowa Gulch at AR5 and increased at the next 2sites. Both Leadville Drain and CaliforniaGulch preceded falls in species richness whilethe poorest site was AR10. Species richnessdeclined after Pueblo Reservoir, contrastingthe recovery exhibited by pupae from AR20.Adult data confirmed the dominance by Ortho-cladiinae among pupal exuviae althoughspecies of Chironominae were relatively moreabundant. Adult Diamesinae were found at allsites except AR18 (if D. leona is included),while Tanypodinae and Podonominae werealso more widely collected compared withpupal data.
The relative abundance of Chironominae isreflected in the increased importance of detri-tivores in Figure 6 compared with pupal data.Grazers were dominant at all sites except AR5,
which was dominated by Chironominae, andat AR18 where they were the rarest trophicgroup. Grazers and detritivores were co-domi-nant at AR12. Filterers were an importantcomponent of the chironomid assemblage atAR18 but, as with pupal data, were absentbelow Pueblo Reservoir.
ORDINATION.—Latitude, Zn toxicity, andparticle size were the only significant vari-ables selected, explaining 22.3% of biologicalvariability. Total Fe was interchangeable withZn toxicity, but the latter was used to maintaincomparability with pupal data. Only the 1stCCA axis was significant (P = 0.01), explaining9.8% of all biological variation and 43.8% ofthe species-environment relationship. Length-wise variation, best explained by latitude, wasagain the dominant influence along the pri-mary axis (t-value 19.2, correlation –0.94). Zntoxicity (t-value 2.3, correlation –0.44) and
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 43
Fig. 3. Proportions of pupae classified by trophic group at each site.

particle size (t-value 6.3, correlation 0.11) werealso significantly related to biological variationalong the first axis.
There was no arch effect in Figure 7 be-cause the first 2 axes were of similar impor-tance (4.41 and 3.56 s units). A north–to–southdistribution of sites occurred along the 1staxis, with lateral spreading of closely situatedupstream sites. Sites with the highest Zn toxi-city were positioned together in the top left ofFigure 7, while the least toxic sites wereplaced diagonally opposite. Sites AR12–AR16and AR19 had relatively large mean substra-
tum particle sizes and site EF1 had the small-est. Site AR10 had the largest particle size, butits position reflects the greater importance oflatitude and Zn toxicity. The association be-tween Krenosmittia halvorseni and the mostZn-toxic sites revealed by pupal data was sup-ported by adult collections. Also in the top leftof Figure 7, two cold-water adapted species,Paracladius alpicola and Cladopelma viridula,as well as Orthocladius subletti and Polypedi-lum trigonus were all present at AR7 (high Zntoxicity) and AR11. Adult Micropsectra nigrip-ila were collected from East Fork downstream
44 WESTERN NORTH AMERICAN NATURALIST [Volume 60
Fig. 4. CCA ordination of pupal data. Arrows indicate importance and direction of maximum change in species com-position among samples as the variable increases. Open circles used for sites, points for species. Species codes fromTable 2.

to AR16, dominating collections from AR5,whereas pupal exuviae were found only atAR11. Gymnometriocnemus brumalis is proba-bly terrestrial; it was absent from pupal collec-tions but adults were collected from AR4,AR5, and AR12, between 2000 and 3000 m.Adults of Cricotopus coronatus were found atsites with high Zn toxicity or at intermediatealtitude. Both adult and pupal collections ofOrthocladius frigidus and O. nigritus indicatedthat these were montane species tolerant ofZn concentrations downstream of CaliforniaGulch.
Among downstream-distributed specieslocated toward the lower right of Figure 7were a few species that also occurred upstreamof AR4. Procladius subletti and Limnophyes n.sp. 3 were collected at EF1 and AR2, respec-tively, were absent at the most Zn-toxic sites,and were present in the vicinity of PuebloReservoir. Pupal exuviae of P. subletti, how-ever, were collected at AR6. Adult Cricotopus
bicinctus and Parametriocnemus lundbeckiiwere both widely distributed except at Zn-toxic sites; however, their pupal exuviae werefound at toxic sites. Adult and pupal C. infus-catus had a downstream distribution but toler-ated metals at AR8. Smittia n. sp. 3, Polypedilumdigitifer, and Micropsectra logani (pupae atAR6) were collected from the first 4 sitesabove California Gulch and then disappeareduntil AR17, or further downstream.
Independence of Sampling Method
In a test for association between pupal andadult data, χ2 = 5908.5, significantly (P < 0.001)exceeding the critical χ2
.05[65] of 106.0 forassociated data. Species most affected by themethod of sampling were Micropsectra nigrip-ila (pupae fewer than expected, adults greater),Rheotanytarsus n. sp. 1 (pupae greater, adultsfewer), Orthocladius rivicola (adults fewer), O.obumbratus (adults greater), Diamesa heteropus
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 45
Fig. 5. Adult subfamily species richness at each site.

46 WESTERN NORTH AMERICAN NATURALIST [Volume 60
TABLE 3. Proportions of adult species collected at each site (see Table 2 for explanation).
TrophicCode Species name group Site
A A A A A A A A A A A A A A A E E A1 2 1 1 1 1 1 1 1 19 0 7 8 3 4 5 6 5 6 7 8 9 1 2 1 2 1
LARS_PLA Larsia planensis (Johannsen) P – – – – – – – – 1 – – – – – 1 – – –PARO_KIE Parochlus kiefferi (Garrett) P – – – – – – – – – – – – – – – 1 1 –DIAM_DAV Diamesa davisi Edwards G – – – – – – – – – 1 – – 2 – – – – –DIAM_SPI Diamesa spinacies Sæther G – – – – – – – – – – – – 2 – – – – –PAGA_ORT Pagastia orthogonia Oliver D – – – – – – – – 1 – – – – – 1 – – –PAGA_PAR Pagastia partica (Roback) D – – – – – – – – – – – – 1 1 1 – – –ODON_FER Odontomesa ferringtoni Sæther D – – – – – – – – – – 1 – – – 1 – – –HYDR_FUS Hydrobaenus fuscistylus (Goetghebuer) G – – – – – – – – – – – – – 1 1 1 – –ACRI_NIT Acricotopus nitidellus (Malloch) D – – – – – – – – – 2 – – 1 2 – – – –BRIL_FLA Brillia flavifrons Johannsen G – – – – – – – – – – – – – – – 2 – 3CRIC_BIF Cricotopus bifurcatus Cranston & Oliv. G – – – – – – – – 1 1 – – – – – – – 2CRIC_TIB Cricotopus tibialis (Meigen) G – – – – – – – – – – – 1 2 – – – – –CRIC_GLO Cricotopus globistylus Roback G – – – – – – – – – – – – 1 – 1 – – –EUKI_n4 Eukiefferiella n. sp. 4 G – – – – – – – – – – – – – 1 – – – 1ORTH_FRI Orthocladius frigidus (Zetterstedt) G – – – – – – 1 – 1 2 1 4 2 1 – 3 4 1ORTH_SUB Orthocladius subletti Soponis G – – – – – – – – – – 1 1 – 1 – – – –ORTH_WIE Orthocladius wiensi Sæther G – – – – – – – – 1 – – – – – – – – –PARA_ALP Paracladius alpicola (Zetterstedt) G – – – – – – – – – – 1 – – 1 – – – –PARA_n3 Paracladius n. sp. 3 G – – – – – – – – – – – – – – 1 – – 1PSEC_SPI Psectrocladius spinifer (Johannsen) G – – – – – – – – – – 1 – – – – 1 – –RHEOCRn1 Rheocricotopus n. sp. 1 (nr. chalybeatus) G – – – – – – – – – 1 – 1 1 1 – 1 – –RHEO_EMI Rheocricotopus eminelobus Sæther G – – – – – – – – 1 – – – – 1 – 1 2 1TOKU_ROW Tokunagaia rowensis (Sæther) D – – – – – – – – – – – – – – 1 1 – 3TVET_PAU Tvetenia paucunca (Sæther) G – – – – – – – – – – – – – – – 1 2 1LIMN_ELT Limnophyes eltoni (Edwards) G – – – – – – – – – – – – – – – 1 2 –LIMN_NAT Limnophyes natalensis (Kieffer) G – – – – – – – – – – – – – – – – – –GYMN_BRU Gymnometriocnemus brumalis (Edwards) G – – – – – – – – 2 – – – – – 1 – – –KREN_n1 Krenosmittia n. sp. 1 G – – – – – – – – – – – – – – – – 1 –KREN_HAL Krenosmittia halvorseni (Cranston
& Oliver) G – – – – – – – – – 1 1 1 – – – – – –LIMN_n1 Limnophyes n. sp. 1 G – – – – – – – – – 1 – – 1 – – – 4 1LIMN_n2 Limnophyes n. sp. 2 G – – – – – – – – – – – – – – – 1 – –METR_BRU Metriocnemus brusti Sæther G – – – – – – – – – – – 1 – – – – – 1LIMN_n4 Limnophyes n. sp. 4 G – – – – – – – – – – – – – – – – 1 1PARAPSEU Paraphaenocladius pseudirritus nearticus
Saether & Wang D – – – – – – – – – – – 1 – – – – – –PARAPNAS Paraphaenocladius nasthecus Sæther D – – – – – – – – – – 1 – – – – – – –SMIT_ATE Smittia aterrima (Meigen) G – – – – – – – – – – – – 1 – 1 – – –SMIT_n1 Smittia n. sp. 1 G – – – – – – – – – – – – – 1 – – – –THIE_ELA Thienemaniella spp. G – – – – – – – – – – – – – – 1 – – 1CHIR_RIP Chironomus riparius Meigen D – – – – – – – – – – – 1 1 – – – – –CLAD_VIA Cladopelma viridula (Linnaeus) D – – – – – – – – – – 1 – – 1 – – – –DICR_NER Dicrotendipes nervosus (Staeger) D – – – – – – – – – – 1 – – – – – – –PARA_NIX Paracladopelma nixe (Townes) P – – – – – – – – – – – – 1 1 – – – –POLY_ALB Polypedilum albicorne (Meigen) D – – – – – – – – 1 – 1 1 – – – 2 1 1POLY_TRI Polypedilum trigonus Townes D – – – – – – – – – – 1 – – 1 – – – –TANY Tn2 Tanytarsus n. sp. 2 D – – – – – – – – – – 1 – – – – 2 – –CRIC_COR Cricotopus coronatus Hirvenoja G – – 1 – 1 – – – 1 3 3 3 2 2 1 – – –CRIC_SLO Cricotopus slossonae Malloch G – – 1 – – 1 – – 1 – – 1 1 1 1 – 1 1CRIC_SYL Cricotopus sylvestris (Fabricius) G – – – – 1 – – – – – 1 – 1 1 1 – – –EUKI_n9 Eukiefferiella n. sp. 9 G 1 – – – – – – – – – – 1 1 1 – – – 1ORTH_NIG Orthocladius nigritus Malloch G – – – – – 1 – – 1 – – 1 – – – – – –LIMN_ASQ Limnophyes asquamatus Andersen G – – – – – 1 – – – – – 1 1 – – 1 – –PSEU_FOR Pseudosmittia forcipata (Goetghebuer) G – – – – – 1 – – – – – 1 – 2 – – – –
A A A A1
2 3 4 0
– – – –1 – – –– – 3 4– 2 3 –– – – –– – 1 –– – 1 –– – 1 –– – – –1 – 1 –– 1 – –– – – –– – – –– – – –– – 3 –– – – –2 – – –– – – –– – – –– – 1 –1 – – –1 1 – –– – – –1 – – –2 – – –1 1 1 –– – 1 –– 1 – –
– – – –3 1 – –2 – 1 –– 1 – –1 – – –
1 3 – –– 1 1 –– – – –– – – 21 – – –– 1 2 –– – – –– – 1 –– – – –1 – – –– – – –1 – – –– – – –1 – 1 –– – – –1 – – –– – 2 –1 – 1 –– 1 1 –
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 47
TABLE 3. Continued.
TrophicCode Species name group Site
A A A A A A A A A A A A A A A E E A1 2 1 1 1 1 1 1 1 19 0 7 8 3 4 5 6 5 6 7 8 9 1 2 1 2 1
SERG_ALB Sergentia albescens (Townes) P – – – – – 1 – – 1 1 1 – – – 1 – – –MICR_POL Micropsectra polita (Malloch) D – – – – – – – 2 – – 4 1 – – 2 – – –PARA_SMI Paramerina smithae (Sublette) P – – – 1 – – – – – 1 – – – – – – – –DIAM_HET Diamesa heteropus (Coquillet) G 4 1 – – 1 – – – 2 4 – – 2 – – – – –PSEU_PER Pseudodiamesa pertinax (Garrett) D – – – – 1 – – – – – – – – 1 1 – – –EUKI_CLA Eukiefferiella claripennis (Lundbeck) G 1 1 1 – 1 1 – 1 1 1 1 2 2 1 2 – 2 2ORTH_MAL Orthocladius mallochi Kieffer G 4 1 – – 4 2 – – 1 2 3 1 3 1 1 1 – 2CHAE_n1 Chaetocladius n. sp. 1 G – – – – 1 – – – 1 – – – – – – – – –SMIT_n3 Smittia n. sp. 3 G – – 1 – – – – – – – – – – – – 1 1 –MICR_NIG Micropsectra nigripila (Johannsen) D 1 – – – 1 – 1 3 4 2 – – – 2 – 2 – –MICR_LOG Micropsectra logani (Johannsen) D – 1 – – – – – – – – – – – – – 1 – 1CRIC_TRE Cricotopus tremulus (Linnaeus) G – – – 1 1 1 – – – – – – 1 1 1 – 1 1CHIR_MAT Chironomus maturus Johannsen D – – – 1 1 – – – – – – – – 1 1 – – 3BORE_LUR Boreoheptygia lurida (Garrett) G – – – – – – 1 4 – – – 1 2 1 – – – 1POLY_n1 Polypedilum n. sp. 1 D – – 1 – 1 1 – 1 – – 1 1 – – – 1 – 1ORTH_RVA Orthocladius rivicola Kieffer G 1 1 1 1 3 1 3 1 1 1 1 1 1 1 1 1 1 1PSIL_n1 Psilometriocnemus n. sp. 1 G – – – – – 1 – – – – – – – – – – – –PHAE_PRO Phaenopsectra profusa (Townes) D – 1 1 – – 1 – 1 – – 1 1 1 – – – – 1PROC_CUL Procladius culiciformis (Linnaeus) P 1 – – 1 – – – – – – – – – 1 – – – –PROC_FRE Procladius freemani Sublette P – – – 1 – – – – – 1 – – – – – – – –PROC_SUB Procladius subletti Roback P – – – 2 – – – – – – – – – 1 – 1 – –DIAM_ANC Diamesa ancysta (Roback) G – – – – – – 4 1 – 1 1 1 – – – – – –HYDR_PIL Hydrobaenus pilipes (Malloch) G 1 1 – – – – – – – – 2 – – – – – – –CARD_PLA Cardiocladius platypus (Coquillet) P – – 1 1 1 3 1 1 – 2 – 1 – 1 – 1 – –CRIC_BIC Cricotopus bicinctus (Meigen) G 1 – 1 1 – – – 1 – – – – – 1 – 1 1 –CRIC_HER Cricotopus herrmanni Sublette G – – 4 1 3 3 3 3 – – – 1 – 2 2 1 – –CRIC_INF Cricotopus infuscatus (Malloch) G 1 1 3 1 – – – – – – – 1 1 – – – – –PARA_CNV Paracladius conversus (Walker) G 1 – 1 1 2 1 – – – – 1 – – 1 1 – – –PSEC_BMS Psectrocladius barbimanus (Edwards) D – – – – – 1 – – – – – – – 1 – – – –TVET_VIT Tvetenia vitraces (Sæther) G – – – 1 – 1 – 1 – – – – 1 – – – – 1LIMN_n3 Limnophyes n. sp. 3 D – – 1 – – – – – – – – – – – – – – –PARA_LUN Parametriocnemus lundbeckii
(Johannsen) G 1 – 1 1 – 1 1 – – – – – 1 1 – 1 1 –CHIR_DEC Chironomus decorus Johannsen D – – 1 2 1 1 – – – 1 1 – – 1 1 1 – –CHIR_ATR Chironomus atrella (Townes) D – – – – – 1 – – – – – – – 1 – – – –CYPH_COR Cyphomella cornea Sæther D – 1 – – 1 – – – – – – – – 1 – – – –DICR_FUM Dicrotendipes fumidus (Johannsen) D 1 2 – – 1 – – – – – – – – – 4 – – –POLY_DIG Polypedilum digitifer Townes D 1 – – 1 – – – – – – – – – – – 1 – –PSEU_RIC Pseudochironomus richardsoni (Malloch) D – – – 1 – – – 1 – – – – – 1 – – – –PROC_BEL Procladius bellus (Loew) P 1 – – 1 – – – 1 – – – – – – – – – –ABLA_MAL Ablabesmyia mallochi (Walley) P 1 – – 1 – – – – – – – – – – – – – –CRIC_ANN Cricotopus annulator Goetghebuer G – – 3 2 3 4 4 3 – – – – 1 1 – – – –CRIC_TFA Cricotopus trifascia Edwards G 2 1 2 1 – – – – – – – – – – – – – –CRIC_BLI Cricotopus blinni Sublette G 3 4 3 1 – – – – – 1 – – – – – 2 – –EUKI_COE Eukiefferiella coerulescens (Keiffer) G 1 2 – – – – – 1 – – – – – – – – – –ORTH_TRI Orthocladius trigonolabis Edwards G 1 1 1 – – – 1 1 – – – – – – – – –ORTH_OBU Orthocladius obumbratus Johannsen G 4 1 – – – – – 1 – – – – – – – – – –PARA.RUV Paratrichocladius rufiventris (Meigen) G – – – – 1 1 – – – – – – – – – – – –POLY_LAE Polypedilum laetum (Meigen) D – – 1 1 1 1 – 1 – – – – – – 1 – – –POLY_SUL Polypedilum sulaceps Townes D – – 1 1 1 – – – – – – 1 – – – – – –POLY_SCA Polypedilum scalaenum (Schrank) D – – 1 3 1 1 – 1 – – – – – – – 1 – –STIC_MAR Stictochironomus marmoreus (Townes) D – – 1 1 – – – – – – – – – – – – – –CLAD_n1 Cladotanytarsus n. sp. 1 D – 1 – 1 – – – – – – – – – – – – – –RHEOTAn1 Rheotanytarsus n. sp. 1 F – – 1 4 – – – 2 – – – – – – – – – –
A A A A1
2 3 4 0
1 – – –– – – –1 – – –1 – 1 –– – – –1 – 1 21 – 1 –– – 1 –– – – –1 1 1 –– – – –– – – 2– – – –1 – 1 41 – 1 –1 1 1 –– – 1 –– – – –– – – –– – – –– – – –1 – – –– – – –– 2 – –– – – –– – – –– – – –– – – –– – – –– – – –1 – – –
– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – – –– – 1 –– – – –– – – –– – 1 –

(adults greater), and Polypedilum scalaenum(adults greater). Species sampled equally wellas pupae and adults (combined χ2 < 1.6) werePagastia partica, Cricotopus herrmanni, Tveteniavitraces, Cricotopus blinni, and Phaenopsectraprofusa.
Effect of Classification Level
Generic adult data were ordinated to inves-tigate the influence of taxonomic level becauseof the large number of species in this data set.Stepwise regression selected maximum watertemperature, total Cu, and mean particle size,explaining 25.1% of generic adult chironomidvariability. The first 2 axes were significant(both P = 0.04), together explaining 18.3% ofbiological variation. The primary axis was sig-nificantly explained by temperature (t-value6.93), while all 3 variables significantly ex-plained the 2nd axis, particle size being theleast important. Despite the overlap of tem-perature and particle size vectors in Figure 8,
the 2 variables were independent (Pearson correlation –0.18, r.05[20] = 0.42) and all vari-ance inflation factors were below 1.1. Siteswere approximately ordered from warmest tocoolest along the diagonal of the temperaturevector in Figure 8. Almost at right angles wasa gradient of metal contamination; AR3 hadalmost twice the Cu concentration of the nextmost contaminated samples from AR5 (Table1). Except for AR3, sites were closer to theorigin of Figure 8 than they were in a speciesCCA. No genera were solely associated withAR3; the closest genera were Paraphaenocla-dius (2 species used for adult CCA), Metrioc-nemus (1 sp.), and Krenosmittia (2 sp.). Thesegenera were found at several upstream sitesbut particularly the most metal-contaminated(AR3–AR8). In the lower half of Figure 8,Parametriocnemus (1 sp.) exhibited metal intol-erance revealed by species CCA, as did Tvete-nia (2 sp.). Responses of other adult species,
48 WESTERN NORTH AMERICAN NATURALIST [Volume 60
Fig. 6. Proportions of adults classified by trophic group at each site.

previously highlighted as metal-intolerant,have been lost among the conflicting trends oftheir congeners within species-rich generasuch as Procladius (4 sp.), Cricotopus (13 sp.),and Polypedilum (7 sp.). Orthocladius (8 sp.),Chironomus (4 sp.), Eukiefferiella (4 sp.), andDiamesa (4 sp.) were also central to the ordi-nation because of counterbalancing speciesdistributions. Limnophyes (7 sp.) was associatedwith low-temperature sites, as only 2 speciesappeared downstream of AR9, and in smallproportions. Micropsectra was associated withmetal-impacted sites due to the distribution ofM. nigripila and M. polita and despite occur-rences of M. logani.
DISCUSSION
Comparisons of Pupal and Adult Data
An unprecedented description of chirono-mid species distribution has been provided for259 km of a major U.S. river. Proportionalspecies abundances across the 22 ArkansasRiver sites were not equally represented bysamples of pupal exuviae and adults. Greaterproportions of adult detritivores indicated thatsources of associated larvae may haveincluded lentic, semi-terrestrial, and terres-trial habitats beyond the Arkansas River. Theabsence of small-bodied Corynoneura and
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 49
Fig. 7. CCA ordination of specific adult data. Explanation as for Figure 4, species codes from Table 3.

Thienemanniella adults indicated that aerial nets were ineffective at catching these midges.The large proportion of predators amongpupal data from sites AR13–AR18 was due torheophilic Cardiocladius platypus, which mayhave been underrepresented in adult collec-tions. Assuming adult data included individu-als from external sources, this would explainwhy river-related environmental variablesaccounted for less biological variation thanthat achieved with pupal data. Despite dis-crepancies in expected numbers of species,there were similarities in species distributionbetween the 2 life stages. Examples cited wereKrenosmittia halvorseni, Orthocladius nigritus,O. frigidus, and Cricotopus infuscatus. Bothpupal and adult collections revealed the pres-ence of filterers upstream of Pueblo Reservoirand their absence downstream. Herrmann andMahan (1977) found that turbidity at the outletwas typically lower than in the reservoir, or atthe inlet, during the first 2 yr of its existence.
Site AR18 was observed to have faster currentthan sites below the reservoir. Species sam-pled equally well as adults and pupae mayhave had broad emergence patterns, beingmultivoltine or asynchronous. Cool-adaptedDiamesa heteropus, as well as Orthocladiusobumbratus, were underrepresented as pupaebecause their main emergence period hadpassed before pupal exuviae were collected.Adults of O. obumbratus were collected fromAR16–AR20 while pupal exuviae were obtainedfrom cooler stations at AR2–AR7. Micropsec-tra nigripila, the most abundant adult species,and Polypedilum scalaenum were also betterrepresented in adult collections. Both speciesprefer lentic habitats and may have originatedfrom extraneous sources. Rheophilic Rheotany-tarsus n. sp. 1 and Orthocladius rivicola werethe most abundant pupal species and wereunderrepresented in adult collections, proba-bly because they were “diluted” by speciesfrom other sources.
50 WESTERN NORTH AMERICAN NATURALIST [Volume 60
Fig. 8. CCA ordination of generic adult data. Explanation as for Figure 4.

Species Richness
Collections of pupal exuviae typically revealgreater species richness than direct samplingof stream habitats for larvae (Ferrington et al.1991, Ruse 1995a). The present study obtainedgreater species richness from adult collections.This could be explained partly by adults origi-nating from extrinsic habitats. Additionally, 17months of adult sampling would increase thenumber of species obtained compared with 3months of pupal sampling. The pupal total of127 species compares favorably with speciestotals for other montane or subalpine streamspresented in a review by Lindegaard andBrodersen (1995), which gave an average mon-tane species total of 71 (range 26–144). Thetotal of 200 adult species was not comparablewith surveys of larvae or pupal exuviae becauseof their uncertain origin. Both pupal and adultdata exhibited a decline in species richness atthe 1st site below Leadville Drain and againbelow California Gulch, the major sources ofmetal pollution. Sites with the highest sedi-mentary concentrations of Zn, Pb, Mn, and Cd(AR5, AR7, AR8) had about average speciesrichness. Other research on the effects ofmetal-polluted mine drainage on chironomidshas demonstrated a reduction in species rich-ness (Winner et al. 1980, Armitage and Black-burn 1985, Yasuno et al. 1985, Wilson 1988).Conversely, Cranston et al. (1997) demonstratedan increase in chironomid species richnessbelow a mine adit, which they attributed to agreater pool of tolerant species in Australiacompared with northern, temperate regions.Neither pupal nor adult data conformed to thedownstream trend of increasing species rich-ness found by Ward (1986) in a neighboringcatchment. Pupal and adult data sets revealeda low number of species from site AR10, whichhad the coarsest substratum and a strong cur-rent. Clements and Kiffney (1994) reported areduced macroinvertebrate species richness ata site approximately 10 km downstream of oursite AR10. The next site downstream, AR11,had the highest number of adult species andthe 3rd highest number of pupal species. Lar-vae of species avoiding sites with metal inputs(EF2, AR3) or with high physical stress (AR10)may have drifted through to the next site,increasing its species richness. The effect isless dramatic below California Gulch becauseof high sedimentary metal concentrations fur-ther downstream. Williams (1989), who pump-
sampled with 50-µm-mesh nets, has suggestedthat Chironomidae actively redistribute them-selves and colonize preferred habitats throughdrifting, particularly as 1st or 2nd instars. Thisbehavior would explain the contrast in speciesrichness between sites EF2 and AR1, AR3 andAR4/6, and AR10 and AR11.
Species Distribution and the Effect of Metals
Environmental measurements most corre-lated with a successive downstream turnoverin species composition (distance/altitude, lati-tude, and temperature) were aligned with theprimary CCA axis of both data sets. Pupal databest reflected a smooth downstream gradientin species turnover. In a neighboring river,Ward (1986) classified 4 zones of species assem-blage related to altitudes between 3414 m and1544 m, although chironomid taxa showedmuch greater overlap than did Plecoptera andTrichoptera. A longitudinal zonation amongChironomidae was suggested by Ward andWilliams (1986) when Chironomini replacedOrthocladiinae in a 36-km-long Canadian river.In the Arkansas River pupal Chironominae in-creased from AR17 downstream, except belowthe reservoir outlet, but there was no evidencefor altitudinal zonation rather than succession.The most abrupt changes were anthropogenic:mining, regulation, and impoundment. In thepupal CCA, localized effects of metal pollutionwithin a 20-km reach were overwhelmed byeffects of downstream succession along 259km of the river. The importance of altitudeand latitude to macroinvertebrate speciesstructure, mediated through their effect ontemperature, has been demonstrated locallyby Ward (1986) and globally by Jacobsen et al.(1997). Latitude was strongly related to dis-tance but, because it changed most betweensites EF1 and AR12, it also had a correlationwith chironomid species variability amongmetal-polluted sites. Longitude varied mostbetween sites AR13 and AR20, where therewas relatively less species variability; conse-quently, it was never selected by forwardregression after latitude had been chosen. In astudy of 6 Colorado streams, including theArkansas River, Clements and Kiffney (1995)found that altitudinal variation confoundedthe effects of metal on benthic macroinverte-brates. Using CCA, we noted that metal pollu-tion still had a significant explanatory value in
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 51

our study, even when generic-level adult datawere considered. Herrmann and Mahan (1977)found that metal-enriched water was reachingPueblo Reservoir, and subsequent research byKimball et al. (1995) confirmed that metalinputs, and their transportation, extend through-out 250 km of river. Sites AR3 and AR5–AR8were extreme examples of metal pollution,whereas concentrations of sedimentary Zn atremaining sites were still high downstream toPueblo Reservoir. The work of Kiffney andClements (1993) revealed that macroinverte-brates bioaccumulated more Zn and Cd at siteAR5 than at AR3 while the reverse was usuallytrue for Cu. These results are in accord withdistributions of chironomid species reportedhere.
Metal-tolerant assemblages of chironomidspecies below California Gulch are evidentfrom Tables 2 and 3. Individual species werehighlighted for their tolerance or intolerance,some of which have been connected previ-ously with metal impacts by other researchers.In the English Pennines, Wilson (1988) founda high proportion of Krenosmittia camptoplepsbelow a Zn-polluted mine adit although thespecies was absent from a neighboring river ofthe same catchment which was also Zn pol-luted. Wilson suspected that metal pollutionalone was not determining species distribu-tion. In the Arkansas River this species wasreplaced by its congener K. halvorseni at siteswith the highest sedimentary Zn-loadings. Inthe same catchment studied by Wilson, Ortho-cladius frigidus was found by Armitage andBlackburn (1985) in moderately Zn-pollutedsites (0.77–1.68 mg L–1) but was absent athigher concentrations (2.08–7.6 mg L–1). O.frigidus reached its highest proportions atsites AR4 and AR8; these sites have recordedsuspended Zn concentrations within the mod-erate range (Roline and Boehmke 1981, Kim-ball et al. 1995) but could be exposed to higherconcentrations in spring (Clements 1994). Thestudy of Elam’s Run in Ohio by Winner et al.(1980) provided evidence of metal tolerance forseveral Arkansas River species that inhabitedsites AR3–AR8: Orthocladius dubitatus, O.obumbratus, Cricotopus bicinctus, C. infuscatus,Diplocladius cultriger, and Larsia planensis(adult). Waterhouse and Farrell (1985) drewattention to C. bicinctus being succeeded byC. infuscatus along a gradient of decliningmetal pollution in Elam’s Run. C. bicinctus
was a minor component of the Arkansas Riverchironomid assemblage, even at the most metal-polluted sites. C. bicinctus did appear belowLeadville Drain at EF2 (adults) and belowCalifornia Gulch at AR5 and AR7 (pupae),while C. infuscatus did not appear until AR8with a predominantly downstream distribution(pupae and adults). C. slossonae was absentfrom the 2 most metal-polluted sites on Elam’srun, but was present at all the most pollutedArkansas River sites. Eukiefferiella claripenniswas not found in Elam’s Run, but its presenceat Zn-polluted sites was recorded by the 2English studies mentioned (Armitage andBlackburn 1985, Wilson 1988) and was tolerantof severely Cu-contaminated (>50 µg L–1)streams in southwest England (Gower et al.1994). E. claripennis, distributed extensivelyalong the Arkansas River, was subdominant toOrthocladius species within pupal collectionsat the most metal-polluted sites.
Species indicated as intolerant of severeheavy-metal pollution included some newspecies: Eukiefferiella n. sp. 9, E. sp. 5-P, Limno-phyes n. sp. 3, and Tanytarsus n. sp. 5. E. co-erulescens avoided the most toxic sites and wasalso reported by Wilson (1988) to be absent atZn-polluted sites. Specific comparison of metaltolerance, especially across widely separatedriver systems, has its limitations. Postma et al.(1995) have demonstrated that chironomid pop-ulations from metal-polluted rivers can exhibitless sensitivity to some metals compared withconspecifics derived from unpolluted sites.They suggest this has a genetic basis.
Future Study
This study of the Arkansas River during1984–85 provides a reference for assessingchanges that have occurred since remediationwork began in 1991. Now that Leadville mineshave ceased operating, subsequent monitoringof chironomid species distribution would recordhow the Arkansas River responds. Biomonitor-ing using generic-level data would save time,provided there was no significant loss of infor-mation. Generic data reduced the amount ofunexplained species variation that probablyarose from the uncertain origin of the rareradult species. There was more homogeneity ofgeneric assemblages between sites, althoughsensitivity to Cu pollution, or perhaps sus-pended metals, was greater than with specific
52 WESTERN NORTH AMERICAN NATURALIST [Volume 60

data. Generic data revealed the same 2 majorgradients, of longitudinal variation and metalcontamination, identified by specific adult andpupal data. Multivariate analysis of 10 benthicmacroinvertebrate data sets by Bowman andBailey (1997) led them to suggest that if trade-offs were necessary to investigate communityvariation, it would be better to sacrifice taxo-nomic resolution than quantitative data. Ananalysis of specific- and generic-level chirono-mid data along a metal-pollution gradient byWaterhouse and Farrell (1985) revealed goodagreement when using nonspecific diversityindices, but important information was lost ifindicators within species-rich genera wererelied upon. The importance of specific identi-fication of chironomid indicators of metal pol-lution was stressed by Gower et al. (1994)using CCA, although this was addressed toresearchers relying on subfamily chironomiddata. The metal-related distribution of severalspecies belonging to the genera Cricotopus,Orthocladius, and Eukiefferiella would havebeen lost if identification of Arkansas Riverpupae and adults had been generic only. Evenamong 2 species of Krenosmittia, pupal datarevealed a distinct difference in metal-relateddistribution. Generic data would be adequatefor a large-scale description of environmentalinfluences but would have diminished valuewhen monitoring recovery of individual sites.
ACKNOWLEDGMENTS
LPR was in receipt of a Winston ChurchillTravelling Fellowship in 1985, and his subse-quent work was supported by the U.K. Envi-ronment Agency. SJH and JES received fund-ing from the U.S. Environmental ProtectionAgency through the Colorado Department ofHealth (Contract C379551). We are indebtedto Mary Sublette for management of type spec-imens and data tabulation, and to Kent Mahanfor sediment chemistries. The views expressedare the authors’ and do not necessarily repre-sent those of their respective agencies.
LITERATURE CITED
ARMITAGE, P.D., AND J.H. BLACKBURN. 1985. Chironomi-dae in a Pennine stream system receiving minedrainage and organic enrichment. Hydrobiologia121:165–172.
BOWMAN, M.F., AND R.C. BAILEY. 1997. Does taxonomicresolution affect the multivariate description of thestructure of freshwater benthic macroinvertebrate
communities? Canadian Journal of Fisheries andAquatic Sciences 54:1802–1807.
CARAVAJAL, G.S., K.I. MAHAN, D. GOFORTH, AND D.E.LEYDEN. 1983. Evaluation of methods based on acidextraction and atomic absorption spectrometry formulti-element determinations in river sediments.Analytica Chimica Acta 147:133–150.
CLEMENTS, W.H. 1994. Benthic invertebrate communityresponses to heavy metals in the Upper ArkansasRiver Basin, Colorado. Journal of the North Ameri-can Benthological Society 13:30–44.
CLEMENTS, W.H., AND P.M. KIFFNEY. 1994. Integratedlaboratory and field approach for assessing impactsof heavy metals at the Arkansas River, Colorado.Environmental Toxicology and Chemistry 13:397–404.
______. 1995. The influence of elevation on benthic com-munity responses to heavy metals in Rocky Mountainstreams. Canadian Journal of Fisheries and AquaticSciences 52:1966–1977.
CRANSTON, P.S., P.D. COOPER, R.A. HARDWICK, C.L.HUMPHREY, AND P.L. DOSTINE. 1997. Tropical acidstreams—the chironomid (Diptera) response innorthern Australia. Freshwater Biology 37:473–483.
FERRINGTON, L.C., M.A. BLACKWOOD, C.A. WRIGHT, N.H.CRISP, J.L. KAVANAUGH, AND F.T. SCHMIDT. 1991. Aprotocol for using surface-floating pupal exuviae ofChironomidae for rapid bioassessment of changingwater quality. Pages 181–190 in Sediment and streamwater quality in a changing environment: trends andexplanation. IAHS Publication 203.
GOWER, A.M., G. MYERS, M. KENT, AND M.E. FOULKES.1994. Relationships between macroinvertebratecommunities and environmental variables in metal-contaminated streams in south-west England. Fresh-water Biology 32:199–221.
HERRMANN, S.J., AND K.I. MAHAN. 1977. Effects of im-poundment on water and sediment in the ArkansasRiver at Pueblo Reservoir. Bureau of ReclamationReport REC-ERC-76-19.
HERRMANN, S.J., J.E. SUBLETTE, AND M. SUBLETTE. 1987.Midwinter emergence of Diamesa leona Roback inthe Upper Arkansas River, Colorado, with notes onother diamesines (Diptera: Chironomidae). Entomo-logica Scandinavica Supplement 29:309–322.
JACOBSEN, D., R. SCHULTZ, AND A. ENCALADA. 1997. Struc-ture and diversity of stream invertebrate assem-blages: the influence of temperature with altitudeand latitude. Freshwater Biology 38:247–261.
KIFFNEY, P.M., AND W.H. CLEMENTS. 1993. Bioaccumula-tion of heavy metals by benthic invertebrates at theArkansas River, Colorado. Environmental Toxicologyand Chemistry 12:1507–1517.
KIMBALL, B.A., E. CALLENDER, AND E.V. AXTMANN. 1995.Effects of colloids on metal transport in a riverreceiving acid mine drainage, Upper Arkansas River,Colorado, USA. Applied Geochemistry 10:285–306.
LINDEGAARD, C., AND K.P. BRODERSEN. 1995. Distributionof Chironomidae (Diptera) in the river continuum.Pages 257–271 in P. Cranston, editor, Chironomids:from genes to ecosystems. CSIRO, Melbourne, Aus-tralia.
MAHAN, K.I., T.A. FODERARO, T.L. GARZA, R.M. MARTINEZ,G.A. MARONEY, M.R. TRIVISONNO, AND E.M. WILL-GING. 1987. Microwave digestion techniques in thesequential extraction of calcium, iron, chromium,maganese, lead and zinc in sediments. AnalyticalChemistry 59:938–945.
2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 53

MCGILL, J.D. 1980. The distribution of Chironomidaethroughout the River Chew drainage system, Avon,England. Doctoral thesis, University of Bristol, Eng-land.
POSTMA, J.F., M. KYED, AND W. ADMIRAAL. 1995. Site spe-cific differentiation in metal tolerance in the midgeChironomus riparius (Diptera, Chironomidae). Hydro-biologia 315:159–165.
ROLINE, R.A. 1988. The effects of heavy metals pollutionof the Upper Arkansas River on the distribution ofaquatic macroinvertebrates. Hydrobiologia 160:3–8.
ROLINE, R.A., AND J.R. BOEHMKE. 1981. Heavy metalspollution of the Upper Arkansas River, Colorado,and its effects on the distribution of the aquaticmacrofauna. Bureau of Reclamation Report REC-ERC-81-15.
RUSE, L.P. 1995a. Chironomid community structurededuced from larvae and pupal exuviae of a chalkstream. Hydrobiologia 315:135–142.
______. 1995b. Chironomid emergence from an Englishchalk stream during a three year study. Archiv fürHydrobiologie 133:223–244.
RUSE, L.P., AND S.J. HERRMANN. 2000. Plecoptera and Tri-choptera species distribution related to environmen-tal characteristics of the metal-polluted ArkansasRiver, Colorado. Western North American Naturalist60:57–65.
RUSE, L.P., AND R.S. WILSON. 1984. The monitoring ofriver water quality within the Great Ouse basin usingthe chironomid exuvial analysis technique. WaterPollution Control 83:116–135.
SANDOVAL, L., J.C. HERRAEZ, G. STEADMAN, AND K.I.MAHAN. 1992. Determination of lead and cadmiumin sediment slurries by ETA-AAS: a comparison ofmethods for the preparation and analysis of sedi-ment slurries. Mikrochimica Acta 108:19–27.
SOKAL, R.R., AND F.J. ROHLF. 1981. Biometry. Freeman,New York.
SUBLETTE, J.E., L.E. STEVENS, AND J.P. SHANNON. 1998.Chironomidae (Diptera) of the Colorado River, GrandCanyon, Arizona, USA, Ι: systematics and ecology.Great Basin Naturalist 58:97–146.
THIENEMANN, A. 1910. Das Sammeln von Puppenhäutender Chironomiden. Archiv für Hydrobiolgie 6:213–214.
TER BRAAK, C.J.F. 1990. Update notes: CANOCO version3.1. Agricultural Mathematics Group, Wageningen,The Netherlands.
TER BRAAK, C.J.F., AND I.C. PRENTICE. 1988. A theory ofgradient analysis. Advances in Ecological Research18:271–317.
TOKESHI, M. 1995. Life cycles and population dynamics.Pages 225–268 in P. Armitage, P.S. Cranston, andL.C.V. Pinder, editors, The Chironomidae: biologyand ecology of non-biting midges. Chapman and Hall,London.
WARD, A.F., AND D.D. WILLIAMS. 1986. Longitudinal zona-tion and food of larval chironomids (Insecta: Diptera)along the course of a river in temperate Canada.Holarctic Ecology 9:48–57.
WARD, J.V. 1986. Altitudinal zonation in a Rocky Mountainstream. Archiv für Hydrobiologie Supplement 74:133–199.
WATERHOUSE, J.C., AND M.P. FARRELL. 1985. Identifyingpollution related changes in chironomid communi-ties as a function of taxonomic rank. Canadian Jour-nal of Fisheries and Aquatic Sciences 42:406–413.
WILLIAMS, C.J. 1989. Downstream drift of the larvae ofChironomidae (Diptera) in the River Chew, S.W.England. Hydrobiologia 183:59–72.
WILSON, R.S. 1988. A survey of the zinc-polluted RiverNent (Cumbria) and the East and West Allen (North-umberland), England, using chironomid pupal exu-viae. Spixiana Supplement 14:167–174.
WINNER, R.W., M.W. BOESEL, AND M.P. FARRELL. 1980.Insect community structure as an index of heavy-metal pollution in lotic ecosystems. Canadian Jour-nal of Fisheries and Aquatic Sciences 37:647–655.
YASUNO, M., S. HATAKEYAMA, AND Y. SUGAYA. 1985. Char-acteristic distribution of chironomids in the riverspolluted with heavy metals. Verhandlung der Inter-nationalen Vereinigung für Limnologie 22:2371–2377.
Received 28 September 1998Accepted 8 February 1999
54 WESTERN NORTH AMERICAN NATURALIST [Volume 60

2000] CHIRONOMID DISTRIBUTION IN THE ARKANSAS RIVER 55
APPENDIX. Species found at only 1 site, either as pupal exuviae or adults. Listed in alphabetical order within tribes.
Species name Pupa/Adult Site
Derotanypus alaskensis (Malloch) A AR3Psectrotanypus dyari (Coquillet) A AR7Radotanypus submarginella (Sublette) A AR11Ablabesmyia basalis (Walley) A AR7Ablabesmyia monilis (Linneaus) A AR11Ablabesmyia sp. A AR2Conchapelopia pallens (Coquillet) P AR18Pentaneura inconspicua (Malloch) A AR18Telopelopia okoboji (Walley) A AR18Thienemannimyia barberi (Coquillet) A AR18Thienemannimyia senata (Walley) A AR18Zavrelimyia sp. 1-P P AR4Procladius prolongatus Roback A AR11Procladius ruris Roback A AR7Tanypus neopunctipennis Sublette A AR18Tanypus nubifer Coquillet A AR18Tanypus stellatus Coquillet A AR18Diamesa garretti Sublette & Sublette A AR12Prodiamesa olivacea (Meigen) A AR2Cardiocladius n. sp. 2 A AR14Cricotopus intersectus (Staeger) A AR19Cricotopus lestralis (Edwards) A AR6Cricotopus sylvestris (Fabricius) P AR12Cricotopus tricinctus (Meigen) A AR5Cricotopus trifasciatus (Panzer) A AR5Cricotopus vierriensis Goetghebuer P AR12Cricotopus n. sp. 18 A AR8Cricotopus sp. 14-P P AR4Cricotopus sp. 15-P P AR2Cricotopus sp. 18-P P AR12Cricotopus sp. 20-P P AR11Cricotopus sp. 21-P P AR20Eukiefferiella brevineris (Malloch) A AR4Eukiefferiella n. sp. 4 P AR11Eukiefferiella n. sp. 8 A AR9Eukiefferiella sp. 10-P P AR17Heterotrissocladius sp. A AR7Nanocladius anderseni Saether A AR17Nanocladius distinctus (Malloch) A AR17Nanocladius rectinervis (Kieffer) A AR15Orthocladius anteilis (Roback) A AR15Orthocladius appersoni Soponis A AR15Orthocladius carlatus (Roback) A AR11Orthocladius dorenus (Roback) A AR1Orthocladius holsatus Goet A AR2Orthocladius nanseni Kieffer P AR11Orthocladius trigonolabis Edwards P AR5Orthocladius sp. 13-P P AR19Paracladius conversus (Walker) P EF1Paratrichocladius skirwithensis (Edwards) A EF1Psectrocladius vernalis (Malloch) A AR16Rheocricotopus chapmani (Edwards) A AR11Metriocnemus n. sp. 2 A AR6Metriocnemus n. sp. 5 A AR11Limnophyes hastulatus Saether A AR2Corynoneura sp. 2-P P AR1Lopescladius hyporheicus Coffman & Roback A AR16Parakiefferiella subaterrima (Malloch) P/A EF1/AR20Paraphaenocladius exagitans (Johannsen) A AR11Paraphaenocladius tonsuratus Saether & Wang A AR5Smittia polaris (Kieffer) A AR8Smittia n. sp. 2 A EF1Rheosmittia sp. 1-P P AR1

56 WESTERN NORTH AMERICAN NATURALIST [Volume 60
Thienemanniella similis (Malloch) P AR1Thienemanniella xena (Roback) A AR18Thienemanniella n. sp. 2 P/A AR16/AR17Thienemanniella sp. 6-P P AR11Chironomus stigmaterus Say A AR20Chironomus n. sp. 5 A AR20Chironomus n. sp. 8 A AR12Cladopelma sp. 4-P P AR6Cryptochironomus fulvus (Johannsen) A AR18Cryptochironomus sp. P AR17Cryptotendipes casuaria (Townes) A AR11Cryptotendipes sp. 2-P P EF1Cyphomella gibbera Saether A AR18Demicryptochironomus (irmaki) n. sp. 1 A AR18Dicrotendipes crypticus Epler A AR18Dicrotendipes lobiger (Kieffer) A AR2Dicrotendipes modestus (Say) A AR18Glyptotendipes sp. A AR19Microtendipes caelum Townes A AR11Nilothauma babiyi (Rempel) A AR14Parachironomus abortivus (Malloch) A AR18Parachironomus arcuatus (Goetghebuer) A AR18Parachironomus directus (Dendy & Sublette) A AR19Parachironomus tenuicaudatus (Malloch) A AR19Paracladopelma undine (Townes) A AR11Paracladopelma n. sp. 2 P AR17Paracladopelma sp. 4-P P AR6Paratendipes fuscitibia Sublette A AR7Paratendipes subequalis (Malloch) A AR6Paratendipes thermophilus Townes P AR17Polypedilum artifer (Curran) A EF1Polypedilum fuscipenne (Meigen) A AR12Polypedilum illinoense (Malloch) P/A AR18/AR18Polypedilum pedatum Townes A AR12Polypedilum scalaenum (Schrank) P AR16Polypedilum sp. 2-P P AR18Polypedilum sp. 8-P P AR18Polypedilum sp. 9-P P AR17Stictochironomus varius (Townes) A AR19Pseudochironomus rex Hauber A AR12Robackia claviger (Townes) P/A AR17/AR18Stictochironomus annulicrus (Townes) A AR2Stictochironomus n. sp. 1 P/A AR18/AR18Pseudochironomus pseudoviridis (Malloch) A AR18Cladotanytarsus n. sp. 2 A AR6Cladotanytarsus n. sp. 3 A AR2Cladotanytarsus sp. 3-P P AR2Micropsectra logani (Johannsen) P AR6Micropsectra nigripila (Johannsen) P AR11Micropsectra n. sp. 3 A AR4Micropsectra n. sp. 5 A AR2Micropsectra n. sp. 6 A EF1Paratanytarsus dubius (Malloch) A AR12Paratanytarsus similatus (Malloch) A AR11Paratanytarsus tenuis (Meigen) A AR11Paratanytarsus n. sp. 1 A AR7Stempellinella sp. 1-P P AR12Sublettea coffmani (Roback) A AR1Tanytarsus bathophilus Kieffer A AR11Tanytarsus fimbriatus Reiss & Fittkau A AR11Tanytarsus pallidicornis (Walker) A AR12Tanytarsus n. sp. 1 A AR20Tanytarsus n. sp. 6 P AR12Tanytarsus n. sp. 13 A AR7Tanytarsus sp. 2-P P AR6