chemokines in chronic progressive neurological multiple sclerosis
TRANSCRIPT

Chemokines in chronic progressive neurological
multiple sclerosis
Peter A Calabresi*,1, Roland Martin2 and Steven Jacobson2
1Department of Clinical Neuroscience, Rhode Island Hospital-Brown University, Providence, Rhode Island 020903, USA;and 2Neuroimmunology Branch of the National Institute of Neurological Disorders and Stroke, National Institutes ofHealth, Bethesda, Maryland 20892, USA
It is hypothesized that in MS and HTLV-1, chemokine and chemokine receptorexpression are important mechanisms by which T cells migrate to sites ofin¯ammation. Preliminary evidence supports the roles of several chemokines,including MIP 1b, in mediating the enhanced migration capacity of MS derivedPBLs. In addition, the ligand CCR-5 seems to be up regulated on PBLs from some
between cytokine phenotype and chemokine receptor pro®le. The chemokinesand chemokine receptor family are likely to be important molecules in chronicprogressive neurological diseases, in which immune cells invade the centralnervous system.
Keywords: multiple sclerosis; HTLV-1; chemokine receptors; migration
Introduction
Chemokines are 8 ± 12 kd chemoattractant cytokinescharacterized by a four cysteine motif (Luster, 1998;Ransohoff et al, 1996; Schall, 1994; Taub, 1996).The presence or absence of an intervening aminoacid(s) (X) between the N terminal cysteines de®nesthe CXC/CX3C and CC families, respectively. Athird family, C chemokines, lacks an N terminalcysteine but is otherwise homologous. It wasoriginally thought that the CXC chemokinesmediated neutrophil migration and CC chemokinesmediated mononuclear cell migration, but now asubclass of CXC chemokines that lacks the se-quence gluatamic acid ± leucine ± arginine near theN terminal are potent chemotactic agents foractivated T cells. Chemokines enhance chemoat-traction and migration by several pathways (Taub,1996). Chemokines bind to chemokine receptors,which are members of the seven transmembranereceptor family and are coupled to G proteins.Chemokine signaling induces a conformationalchange in integrin molecules and enhances their
binding to the cell adhesion molecules ICAM andVCAM. In addition, chemokines may have directeffects on T cell activation and release of matrixmetalloproteinases that digest the extracellularmatrix underlying cerebral endothelium. The che-mokines and chemokine receptor family are there-fore likely to be important molecules in chronicprogressive neurological diseases in which immunecells invade the central nervous system.
Multiple sclerosis is a chronic, predominantlydemyelinating, disease of the central nervoussystem, characterized by perivenular in¯ammatorycell in®ltrates (Martin and McFarland, 1995; Raine,1994). Although the pathogenesis of the disease isunknown, a number of mechanisms have beensuggested, including: (1) the disease is associatedwith autoreactive T cells directed against myelinpeptides or (2) the disease is secondary to aninappropriate immune response to microbial anti-gens, predominantly viral. The concepts are notmutually exclusive, since an immune responsedirected against a virus may result in inappropriateimmune reactivity through several mechanismsincluding persistent viral infection with: (1) mole-cular mimicry between viral epitopes and myelin orother CNS constituents; (2) in¯ammatory cytokinemediated bystander damage, or (3) viral infection ofoligodendrocytes that leads to impaired myelin
*Correspondence: PA Calabresi, Departement of Neurology, Rho-de Island Hospital-Brown University, 110 Lockwood St., Suite 3-42, Providence, Rhode Island 02903, USAReceived 4 September 1998; revised 21 September 1998; accepted25 September 1998
Journal of NeuroVirology (1999) 5, 102 ± 108
ã
MS patients. Analysis of T cell clones does not reveal a de®nite correlation
http://www.jneurovirology.com
diseases: HTLV-1 associated myelopathy and
1999 Journal of NeuroVirology, Inc.

maintenance and an indirect immune response todamaged tissues.
Several lines of evidence support the autoim-mune theory including: (1) immunogenetic predis-position conveyed by certain HLA DR molecules;(2) a higher prevalence in women (3 : 1) as is seen inother autoimmune diseases; (3) a greater frequencyof autoreactive T cells in MS patients' peripheralblood that do not require a second or costimulatorysignal, which suggests they have been primed invivo (Lovett-Racke et al, 1998), and (4) the clinicalresponse in some patients to immunosuppressivetherapies (Lucchinetti and Rodriguez, 1997; Martinand McFarland, 1995). Proponents of the autoim-mune hypothesis also suggest that intercurrent viralor bacterial infections may activate autoreactive Tcells, which may be associated with diseaseexacerbations.
Epidemiological studies suggest there may be acritical environmental trigger that predisposespeople to getting MS who spend their childhoodyears in temperate zones, especially areas around458 latitude (Hogancamp et al, 1997). In addition,studies of monozygotic twins have demonstrated a30 ± 40% concordance rate, which again arguesthat there must be an environmental factor thatprovokes the process in those individuals who areimmunogenetically predisposed (McFarland et al,1985). Numerous viruses have been implicated inthe pathogenesis of MS over the years. Reports ofelevated titers of EBV, measles virus, caninedistemper virus, HTLV-1 and recently HHV-6have at times raised hopes that one or more ofthese viruses may be implicated in the pathogen-esis of the disease (Lucchinetti and Rodriguez,1997; Soldan et al, 1997). HTLV-1 has since beendemonstrated to be the etiologic agent for TropicalSpastic Paraparesis-HTLV-1 associated myelopa-thy (HAM-TSP), a chronic progressive neurologi-cal disorder with clinical features similar to someforms of MS (Hollberg and Ha¯er, 1993; Nakagawaet al, 1995). With respect to the other viruses,some critics argue that there is non-speci®callyenhanced immune reactivity in MS that accountsfor elevated anti-viral antibody titers. Recently,our lab has found evidence in some MS patientsfor an IgM response to HHV-6, which is the causeof Roseola and a ubiquitous childhood infection(Caserta and Breese Hall, 1993). In addition, weare able to amplify HHV-6 DNA from the serum ofapproximately one-third of MS patients (Soldan etal, 1997). Others have found evidence for HHV-6expression in the oligodendrocytes of MS patients(Challoner, 1995). Whether these data are anartifact of in¯amed tissue and enhanced immu-noreactivity is unclear, but the role of herpesviruses remains of particular interest because oftheir propensity to encode chemokines andchemokine receptors (Meini et al, 1996; Smith etal, 1996).
Viruses and chemokines
Many of the large double stranded DNA viruses arenow recognized to contain genes that encodeproteins, which allow the virus to evade the hostimmune response. Viral homologues of IL-10, IFN-gand TNF receptor, IFN responsive elements (IRE),apoptosis inhibiting proteins (BCL-2) and chemo-kines and chemokine receptors have been identi®ed(Smith et al, 1996). HHV-6 has regions in itsgenome which encode for two chemokines and achemokine receptor (Gompels et al, 1995). HHV-8also has high homology for chemokines and hasrecently been implicated in the pathogenesis ofKaposi's sarcoma and certain CNS lymphomas(Ensoli and Sturzl, 1998). CMV also has regions ofhomology with chemokines and chemokine recep-tors. The chemoattractant molecules may be animportant mechanism by which lymphotropicherpes viruses evade the initial immune responseand propagate themselves to immunologicallyprivileged tissues. According to this hypothesis,viral infected lymphocytes may express chemokinesor chemokine receptors, which allow them topreferentially migrate to the brain. Viral infectionof lymphocytes may occur as the primary eventwith subsequent migration to CNS tissues, or thelymphocytic in¯ammatory reaction may occursecondarily in response to a primary CNS infection.Astrocytes and endothelial cells can producechemokines during in¯ammatory situations or inresponse to infection and may signal in¯ammatorycells (Ransohoff et al, 1996; Ebnet et al, 1997; Guptaet al, 1998).
The mechanisms by which HIV-1 infection cancause CNS pathology are beyond the scope of thisarticle, but may be determined to some extent bychemokine or chemokine receptor expression. It isclear that the chemokine receptors CCR-5 andCXCR-4 are necessary cofactors for the infection ofT cells by the macrophage tropic and lymphotropicstrains respectively of the virus, and that enhancedexpression of certain chemokines may be protectiveby causing receptor down regulation (Dragic et al,1996; Kelly et al, 1998). The CCR-5 mutation thatprotects against HIV-1 infection in a minority ofCaucasians does not seem to offer any protectionagainst MS (Bennetts et al, 1997).
HTLV-1
HTLV-1 infection is endemic in certain regions ofthe Caribbean and only a minority of the infectedpeople manifest medical illness (Hollberg andHa¯er, 1993; Nakagawa et al, 1995). Disorders thathave been de®nitively associated with HTLV-1infection include adult T cell leukemia (ATL) andHAM-TSP, a chronic progressive myelopathy withfeatures similar to primary progressive MS.
Chemokines in HTLV-1 and MSPA Calabresi et al
103

ATL is characterized by massive monoclonalexpansion and subsequent in®ltration of infectedcirculating CD4+ lymphocytes into several tissuesand secondary lymphoid organs. The extravasationof malignant cells into tissues involves the sameadhesive pathways responsible for migration ofin¯ammatory cells. ATL cells spontaneously adhereto endothelial cells, and this appears to be mediatedby enhanced expression of the chemokines MIP-1 aand MIP-1 b (Tanaka et al, 1998). Chemokines bindto seven transmembrane G-protein-coupled recep-tors, which causes a conformational change in theintegrin LFA-1 and appears to enhance ®rmadhesion to the ligand ICAM.
HAM-TSP is a neurological manifestatioan ofHTLV-1 infection. Patients develop a slowly pro-gressive paraparesis with minimal sensory involve-ment. Several in¯ammatory mediators includingTNF, adhesion molecules, and MCP-1 are expressedby in®ltrating cells (Fox et al, 1996; Umehara et al,1996). HAM-TSP patients have very high frequen-cies of HTLV-1 speci®c CD8+ CTL and these havebeen shown to produce numerous in¯ammatorymediators including MIP-1 a and MIP-1 b (Biddisonet al, 1997a). Also, human T cell lines infected withHTLV-1 express the chemokines SDF, RANTES,and MIP-1 a and MIP-1 b. Expression of the HTLV-1viral protein Tax induces the expression of thesechemokine genes as well as other in¯ammatorycytokines (Baba et al, 1996; Arai et al, 1998; Mendezet al, 1997). Therefore, it seems likely that the samemechanisms involved in leukocyte recruitment toin¯ammatory sites are involved in mediating tissuedamage by HTLV-1 infected T cells.
It is also possible that HTLV-1 can infect CNScells directly. HTLV-1 DNA has been detected by insitu hybridization in cells that appear to beastrocytes (Lehky et al, 1995). Astrocytes are animportant source of in¯ammatory mediators in-cluding chemokines (Ransohoff et al, 1996). There-fore, HTLV-1 infection within the CNS may inducerecruitment of antigen speci®c T cells throughchemokine signaling pathways. This recruitmentprocess may be very similar to what has alreadybeen elegantly shown to occur in ATL.
EAE
EAE is an experimental model of immune mediateddemyelination, which has some features in commonwith MS (Martin and McFarland, 1995). The diseasecan be induced by inoculation of myelin reactive Tcells that migrate to the CNS and mediate in¯am-matory destruction of myelin. Recent studies havedemonstrated the expression of many chemokinesincluding; RANTES, MIP-1 a, MIP-1 b, IP-10, MCP-1and 3, KC, TCA3 in conjunction with in¯ammatorycell in®ltrates, but before clinical signs of disease,which suggests these molecules play a critical role
in the immune ampli®cation process that leads todemyelination (Berman et al, 1996; Glabinski et al,1997; Godiska et al, 1995). In®ltrating T cells seemto be primarily responsible for production ofRANTES and MIP-1 a, MIP-1 b, whereas MCP-1,IP-10, and KC expression is con®ned to astrocytes(Glabinski et al, 1995, 1997). The precise roles ofthese molecules is currently being better de®nedand is discussed elsewhere.
Multiple sclerosis
As discussed previously, the etiology of MSremains unknown. The hypothesis that a microbialantigen triggers an aberrant immune response incertain immunogenetically predisposed patientsremains attractive. However, it is possible that thedisease is a result of a purely autoimmune process(Lucchinetti and Rodriguez, 1997). Regardless, thepathology demonstrates perivenular in¯ammatorycell in®ltrates, and there seems to be a de®niteclinical response to immunomodulating therapies.Therefore, it is likely that in MS chemokine andchemokine receptor expression are important me-chanisms by which antigen driven T cells migrateto sites of in¯ammation (Merrill and Benveniste,1996; Merrill and Murphy, 1997; Ransohoff et al,1996). An analysis of chemokines within MS brainscon®rmed expression of RANTES, MIP-1 a, MIP-1 b,and MCP-1. RANTES appeared to be restricted toperivascular sites, while MIP-1 a, MIP-1 b, andMCP-1 were also expressed by glial cells within andaround plaques (Simpson et al, 1998). Both CD8+and CD4+ myelin reactive T cell clones derivedfrom peripheral blood may express chemokinesincluding MIP-1 a, MIP-1 b, IP-10, and IL-16, andthe matrix metalloproteinases MMP-9 and MMP-2(Biddison et al, 1997b, 1998 and unpublishedobservations). PBMC's derived from MS patientshave increased adherence in binding assays andincreased migration across a ®bronectin matrix inresponse to the chemokines RANTES, MIP-1 a, andMCP-1 (Stuve et al, 1997). Interferon beta, which istherapeutic in MS inhibits migration and may do sopartly by abrogating MCP-1 induced production ofthe matrix degrading enzyme MMP-9 (Leppert et al,1995; Stuve et al, 1996).
Results and Discussion
Our studies con®rm modestly enhanced migrationof MS patient PBLs as compared to controls,towards the chemokine MIP-1b (Figure 1a). Thereis minimal to no spontaneous migration from PBLs,but the addition of IL-2 and chemokines results inenhanced migration (Figure 1b). However, there issigni®cant variability from one individual to an-other, which suggests that the expression of the
Chemokines in HTLV-1 and MSPA Calabresi et al
104
chemokine receptors must also be divergent. Thishas also been observed at the clonal level, and evenwithin clones derived from the same patient (Figure1c). Therefore, we undertook to determine whetherchemokine receptor expression is selectively upregulated on MS PBLs or T cell clones with speci®ccytokine expression patterns.
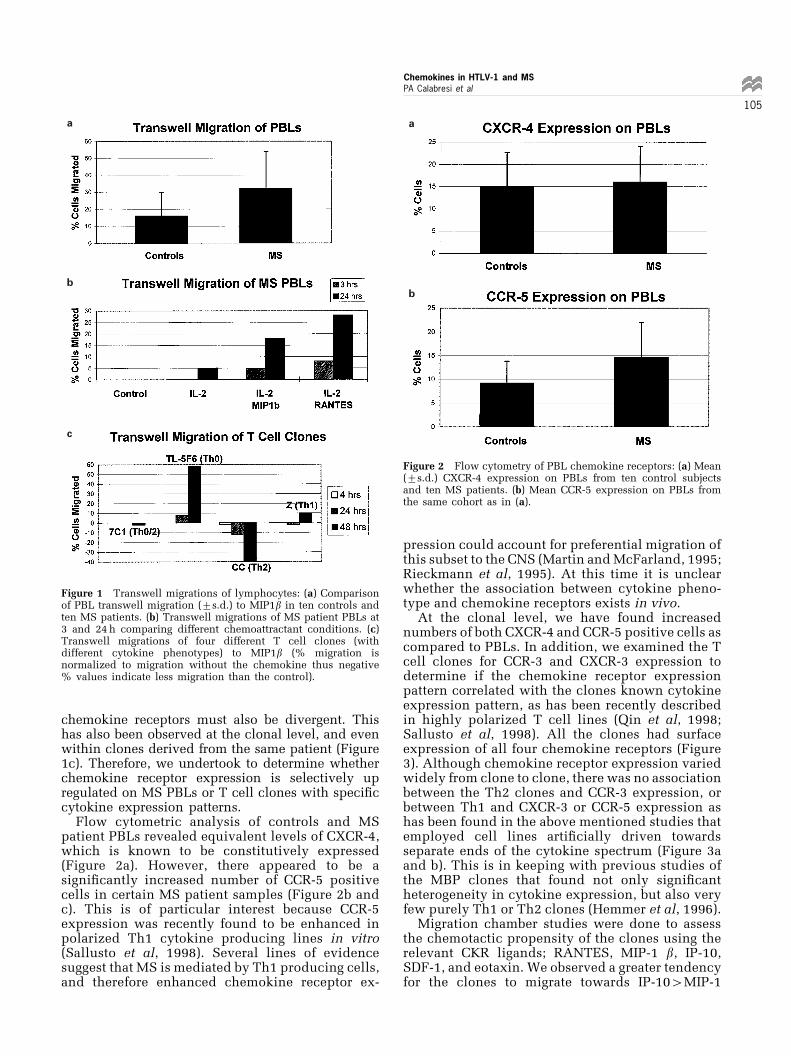
Flow cytometric analysis of controls and MSpatient PBLs revealed equivalent levels of CXCR-4,which is known to be constitutively expressed(Figure 2a). However, there appeared to be asigni®cantly increased number of CCR-5 positivecells in certain MS patient samples (Figure 2b andc). This is of particular interest because CCR-5expression was recently found to be enhanced inpolarized Th1 cytokine producing lines in vitro(Sallusto et al, 1998). Several lines of evidencesuggest that MS is mediated by Th1 producing cells,and therefore enhanced chemokine receptor ex-
pression could account for preferential migration ofthis subset to the CNS (Martin and McFarland, 1995;Rieckmann et al, 1995). At this time it is unclearwhether the association between cytokine pheno-type and chemokine receptors exists in vivo.
At the clonal level, we have found increasednumbers of both CXCR-4 and CCR-5 positive cells ascompared to PBLs. In addition, we examined the Tcell clones for CCR-3 and CXCR-3 expression todetermine if the chemokine receptor expressionpattern correlated with the clones known cytokineexpression pattern, as has been recently describedin highly polarized T cell lines (Qin et al, 1998;Sallusto et al, 1998). All the clones had surfaceexpression of all four chemokine receptors (Figure3). Although chemokine receptor expression variedwidely from clone to clone, there was no associationbetween the Th2 clones and CCR-3 expression, orbetween Th1 and CXCR-3 or CCR-5 expression ashas been found in the above mentioned studies thatemployed cell lines arti®cially driven towardsseparate ends of the cytokine spectrum (Figure 3aand b). This is in keeping with previous studies ofthe MBP clones that found not only signi®cantheterogeneity in cytokine expression, but also veryfew purely Th1 or Th2 clones (Hemmer et al, 1996).
Migration chamber studies were done to assessthe chemotactic propensity of the clones using therelevant CKR ligands; RANTES, MIP-1 b, IP-10,SDF-1, and eotaxin. We observed a greater tendencyfor the clones to migrate towards IP-104MIP-1
a
b
c
Figure 1 Transwell migrations of lymphocytes: (a) Comparisonof PBL transwell migration (+s.d.) to MIP1b in ten controls andten MS patients. (b) Transwell migrations of MS patient PBLs at3 and 24 h comparing different chemoattractant conditions. (c)Transwell migrations of four different T cell clones (withdifferent cytokine phenotypes) to MIP1b (% migration isnormalized to migration without the chemokine thus negative% values indicate less migration than the control).
a
b
Figure 2 Flow cytometry of PBL chemokine receptors: (a) Mean(+s.d.) CXCR-4 expression on PBLs from ten control subjectsand ten MS patients. (b) Mean CCR-5 expression on PBLs fromthe same cohort as in (a).
Chemokines in HTLV-1 and MSPA Calabresi et al
105

b4eotaxin (Figure 4). This again did not correlatewell with cytokine or CKR expression, although ®tswith a slight TH1 predominance. The discrepancybetween the CKR surface expression and functionalcapacity to migrate suggests that alternative me-chanisms such as binding avidity and metallopro-teinase production are involved in migration acrossthe coated ®lters.
Materials and methods
CellsTen patients with clinically de®nite MS and tennormal controls were studied. PBMCs were isolatedfrom heparinized whole blood (Calabresi et al,1997) and allowed to adhere for 2 h to tissue culturewells. Four MBP reactive T cell clones with knowncytokine phenotypes were also studied (Hemmer etal, 1996).
MigrationTo better de®ne the role of chemokines in MS, wehave employed Transwell migration chambers
(Costar) to study chemotaxis of MS patient PBLs aswell as MBP reactive T cell clones. Brie¯y, 56105
lymphocytes were placed in the upper chamber of aTranswell insert in IMDM supplemented with 10%human AB serum (Sigma) and 10 u/ml IL-2.Transmigration across a 5 mm pore polycarbonate®lter coated with Matrigel (Becton-Dickinson) wasassessed at the indicated time points by countingthe cells from the lower chamber on a hemacyt-ometer.
Flow cytometry26105 PBLs/T cell clones were placed on ice for60 min in FACS buffer containing phosphatebuffered saline (PBS) and 0.1% bovine serumalbumin (BSA) and incubated with PE conjugatedantibody to CD3 (Sigma) and monoclonal anti-bodies to human CCR5, CXCR-4, CXCR-3, CCR-3,and CCR-2 (R&D Systems and kind gift of WalterNewman, LeukoSite). A secondary FITC labeledantibody was incubated for 30 min. Appropriateisotype controls were also employed. The cellswere washed and analyzed immediately on aFACSCAN (Becton-Dickinson, San Jose, CA,USA). Data are presented as % of cells stainingpositive for both CD3 and the respective chemokinereceptor after subtracting out background fromsubtype control antibodies.
Conclusion
These data, in conjunction with previous studiesthat demonstrated chemokine production by MBPand PLP clones, indicate that chemokine-chemo-kine receptor signaling is likely an importantmechanism by which antigen speci®c lymphocytesmigrate to sites of in¯ammation and recruit
a
b
Figure 3 Flow cytometry of T cell clone chemokine receptors:(a) Mean chemokine receptor expression for clone 7H11 (Th2cytokine phenotype (high IL-4 low IFN-g by ELISA of condi-tioned medium)) and clone MS-1 (Th-1 cytokine phenotype). (b)Mean chemokine receptor expression for clone TL-5F6 (Th0cytokine phenotype (moderate IL-4 and high IFN-g by ELISA ofconditioned medium)) and clone 7C1 (Th-0/2 high IL-4 andmoderate IFN-g cytokine phenotype).
Figure 4 Cell clone migrations: percent cells migrated tochemokines divided by spontaneous migration on X axis,chemokines used for chemotactic gradient on Y axis, and T cellclones migrated on Z-axis.
Chemokines in HTLV-1 and MSPA Calabresi et al
106

secondary immune cells. It remains unclearwhether one component of this signaling pathwayis preferentially up regulated in MS, either by avirus or as part of a Th1 immune bias. It is alsouncertain what role astrocytes or endothelial cellsmight play in the recruitment process, as these cellscan express chemokines and chemokine receptors.
It is likely that chemokines and chemokinereceptors also are important in the pathogenesis ofHAM/TSP. The role of chemokines in the HTLV-1hematological disease, ATL, has already beendemonstrated, and recent data suggest the HTLV-1Tax can induce chemokine production. It is unclearwhether immune cells, astrocytes, or both areinfected, and if the virus does indeed initiate animmune response that ultimately results in tissuedamage.
Our understanding of chemokines and chemo-kine receptors in immune processes has grownexponentially. Recent data suggest that thesemolecules also likely have signi®cant roles in non-immune cells, both to signal the immune system toinjury and as a part of neuronal migration duringembryogenesis and possibly tissue repair (Zou et al,1998). It is likely that our appreciation of the diverseroles of the chemokines in diseases of the nervoussystem will undergo a similar expansion.
Acknowledgements
This study was supported in part by a grant fromthe National MS Society: PP0600.
References
Arai M, Ohashi T, Tuskahara T, Murakami T, Hori T,Uchiyama T, Yamamoto N, Kannagi M, Fujii M(1998). Human T-cell leukemia virus type 1 TAxprotein induces the expression of lymphocyte che-moattractant SDF-1/PBSF. Virology 241: 298 ± 303.
Baba M, Imai T, Yoshida T, Yoshie O (1996). Constitu-tive expression of various chemokine genes in humanTh-0 cell lines infected with human T-cell leukemiavirus type 1: role of the viral transactivator Tax. Int JCancer 66: 124 ± 129.
Bennetts BH, Teutsch SM, Buhler MM, Heard RN,Stewart GJ (1997). The CCR-5 deletion mutation failsto protect against multiple sclerosis. Human Immunol58: 52 ± 59.
Berman J, Guida M, Warren J, Amat J, Bronsan C (1996).Localization of monocyte chemoattractant peptide-1expression in the central nervous system in experi-mental autoimmune encephalomyelitis and trauma inthe rat. J Immunol 156: 3017 ± 3023.
Biddison W, Cruikshank W, Center D, Pelfrey C, Taub D,Turner R (1998). CD8+ Myelin Peptide-Speci®c T cellscan chemoattract CD4+ Myelin Peptide-Speci®c Tcells: Importance of IFN-Inducible Protein 10. The JImmunol 160: 444 ± 448.
Biddison WE, Kubota R, Kawanishi T, Taub DD,Cruikshank WW, Center DM, Connor EW, Utz U,Jacobson S (1997a). Human T cell leukemia virus typeI (HTLV-I)-speci®c CD8+ CTL clones from patientswith HTLV-I-associated neurologic disease secreteproin¯ammatory cytokines, chemokines, and matrixmetalloproteinase. J Immunol 159: 2018 ± 2025.
Biddison WE, Taub DD, Cruikshank WW, Center DM,Connor EW, Honma K (1997b). Chemokine and matrixmetalloproteinase secretion by myelin proteolipidprotein-speci®c CD8+ T cells. The J Immunol 158:3046 ± 3053.
Calabresi PA, Pelfrey CM, Tranquill LR, McFarland HF(1997). VLA-4 expression on peripheral blood lym-phocytes is downregulated after treatment of multiplesclerosis with interferon beta. Neurology 49: 1111 ±1116.
Caserta MT, Breese Hall C (1993). Human Herpes Virus-6. Ann Rev Med 44: 377 ± 383.
Challoner P, Smith KT, Parker JD, MacLeod DL, CoulterSN, Rose TM, Schulz J, Bennett JL, Garber RL, ChangM, Schad PA, Stewart PM, Nowinski RC, Brown JP,Burmer GC (1995). Plaque associated expression ofhuman herpes virus 6 in multiple sclerosis. Proc NatlAcad Sci 92: 7440 ± 7444.
Dragic T, Litwin V, Allaway GP, Martin SR, Huang Y,Nagashima KA, Cayanan C, Maddon PJ, Koup RA,Moore JP, Paxton WA (1996). HIV-1 entry into CD4+cells is mediated by the chemokine receptor CC-CKR-5. Nature 381: 667 ± 673.
Ebnet K, Brown K, Siebenlist U, Simon M, Shaw S(1997). Borrelia burgdorferi activates nuclear factor-kband is a potent inducer of chemokine and adhesionmolecule gene expression in endothelial cells and®broblasts. J Immunol 158: 3285 ± 3292.
Ensoli B, Sturzl M (1998). Kaposi's Sarcoma: A result ofthe Interplay among In¯ammatory Cytokines, Angio-genic Factors and Viral Agents. Cytokine and GrowthFactors Rev 9: 63 ± 83.
Fox RJ, Levin MC, Jacobson S (1996). TNF alphaexpression in the spinal cord of HAM/TSP patients.J Neurovirol 2: 323 ± 329.
Glabinski A, Tani M, Strieter R, Tuohy V, Ransohoff R(1997). Synchronous synthesis of alpha and betachemokines by cells of diverse lineage in the centralnervous system of mice with relapses of chronicexperimental autoimmune encephalomyelitis. Am JPathol 150: 617 ± 630.
Glabinski AR, Tani M, Tuohy VH, Tuthill RJ, RansohoffRM (1995). Central nervous system chemokine mRNAaccumulation follows initial leukocyte entry at theonset of acute murine experimental autoimmuneencephalomyelitis. Brain Behav Immunol 9: 315 ± 330.
Godiska R, Chantry D, Dietsch GN, Gray PW (1995).Chemokine expression in murine experimental aller-gic encephalomyelitis. J Neuroimmunol 58: 167 ± 176.
Gompels U, Nicholas J, Lawrence G et al (1995). TheDNA sequence of human herpes virus-6: structure,coding content, and genome evolution. Virology 209:29 ± 51.
Chemokines in HTLV-1 and MSPA Calabresi et al
107

Gupta S, Lysko P, Pillarisetti K, Ohlstein E, Stadel J(1998). Chemokine Receptors in Human EndothelialCells. J Biol Chem 273: 4282 ± 4287.
Hemmer B, Vergelli M, Calabresi PA, Huang T, McFar-land H, Martin R (1996). Cytokine phenotype ofhuman autoreactive T cell clones speci®c for theimmunodominant myelin protein peptide. J NeurolSci 45: 852 ± 862.
Hogancamp W, Rodriguez M, Weinshenker BG (1997).The Epidemiology of Multiple Sclerosis. Mayo ClinProc 72: 871 ± 878.
Hollberg P, Ha¯er DA (1993). Pathogenesis of diseaseinduced by HTLV-1 infection. N Engl J Med 328:1173 ± 1182.
Kelly MD, Hassan MN, Adams SL, Cunningham AL,Lloyd AR (1998). Cutting Edge: Dichotomus Effects ofb-Chemokines on HIV Replication in Monocytes andMonocyte-Derived Macrophages. J Immunol 160:3091 ± 3095.
Lehky TJ, Fox CH, Levin MC, Flerlage N, Izumo S, SatoE, Raine CS, Osame M, Jacobson S (1995). Detectionof HTLV-1Tax RNA in the CNS of HAM/TSP patientsby in situ hybridization. Ann Neurol 47: 167 ± 175.
Leppert D, Waubant E, Galardy R, Bunnett NW, HauserSL (1995). T cell gelatinases mediate basementmembrane transmigration in vitro. J Immunol 154:4379 ± 4389.
Lovett-Racke AE, Trotter JL, Lauber J, Perrin PJ, June CH,Racke MK (1998). Decreased dependence of myelinbasic protein-reactive T cells on CD28-mediatedcostimulation in multiple sclerosis patients: a markerof activated/memory T cells. J Clin Inv 101: 725 ± 730.
Lucchinetti CF, Rodriguez M (1997). The controversysurrounding the pathogenesis of the multiple sclerosislesion. Mayo Clin Proc 72: 665 ± 678.
Luster AD (1998). Chemokines - Chemotactic CytokinesThat Mediate In¯ammation. N Engl J Med 338: 436 ±445.
Martin R, McFarland HF (1995). Immunological aspectsof experimental allergic encephalomyelitis and multi-ple sclerosis. Crit Rev Clin Lab Sci 32: 121 ± 182.
McFarland HF, Greenstein F, McFarlin DE, et al (1985).Family and twin studies in multiple sclerosis. AnnNY Acad Sci 436: 118 ± 124.
Meini E, Fickenscher H, Fleckenstein B (1996). Chemo-kine receptors and chemokine-inducing molecules oflymphotrophic herpes viruses. Immunol Today 17:198.
Mendez E, Kawanashi T, Clemens K, Siomo H, CalabresiPA, Brady J, Jacobson J (1997). Astrocyte Speci®cExpression of Human T-Cell Lymphotrophic Type I(HTLV-1) Tax: Induction of TNF alpha and Suscept-ibility to Lysis by CD8+ HTLV-1 Speci®c Cytotoxic TCells. J Virology 71: 9143 ± 9149.
Merrill JE, Benveniste EN (1996). Cytokines in in¯am-matory brain lesions: helpful and harmful. TrendsNeurosci 19: 331 ± 338.
Merrill JE, Murphy SP (1997). In¯ammatory events at theblood brain barrier: regulation of adhesion molecules,cytokines, and chemokines by reactive nitrogen andoxygen species. Brain Behav Immunol 11: 245 ± 263.
Nakagawa M, Izumo S, Ijichi S, Kubota H, Arimura K,Kawabata M, Osame M (1995). HTLV-1 associatedmyelopathy: analysis of 213 patients based on clinicalfeatures and laboratory ®ndings. J Neurovirol 1: 50 ±61.
Qin S, Rottman JB, Myers P, Kassam N, Weinblatt M,Loetscher M, Koch AE, Moser B, Mackay CR (1998).The Chemokine Receptors CXCR3 and CCR5 MarkSubsets of T Cells Associated with Certain In¯amma-tory Reactions. J Clin Invest 101: 746 ± 754.
Raine CS (1994). Multiple Sclerosis: Immune systemmolecule expression in the central nervous system. JNeuropathol Exp Neurol 53: 328 ± 337.
Ransohoff RM, Glabinski A, Tani M (1996). Chemokinesin immune-mediated in¯ammation of the centralnervous system. Cytokine & Growth Factor Rev 7:35 ± 46.
Rieckmann P, Albrecht M, Kitze B, Weber T, Tumani H,Broocks A, Luer W, Helwig A, Poser S (1995). Tumornecrosis factor-a mRNA expression in patients withrelapsing-remitting multiple sclerosis is associatedwith disease activity. Ann Neurol 37: 82 ± 88.
Sallusto F, Lenig D, Mckay CR, Lanzavecchia A (1998).Flexible Programs of Chemokine Receptor Expressionon Human Polarized T Helper 1 and 2 Lymphocytes. JExp Med 187: 875 ± 883.
Schall T (1994). The Chemokines. The Cytokine Hand-book. London, Academic Press: 419 ± 460.
Simpson JE, Newcombe J, Cuzner ML, Woodroofe MN(1998). Expression of monocyte chemoattractant pro-tein-1 and other beta-chemokines by resident glia andin¯ammatory cells in multiple sclerosis lesions. JNeuroimmunol 84: 238 ± 249.
Smith GL (1996). Virus proteins that bind cytokines,chemokines or interferons. Curr Opin Immunol 8:467 ± 471.
Soldan S, Bert R, Salem N, Secchiero P, Calabresi PA,Maloni H, McFarland HF, Lin HC, Patnaik M,Jacobson S (1997). Association of human herpes virus6 (HHV-6) with multiple sclerosis: Increased IgMresponse to HHV-6 early antigen and detection ofserum HHV-6 DNA. Nature Med 3: 1394 ± 1397.
Stuve O, Chabot S, Jung SS, Williams G, Wee Yong V(1997). Chemokine-enhanced migration of humanperipheral blood mononuclear cells is antagonizedby interferon beta-1b through an effect on matrixmetalloproteinase-9. J Neuroimmunol 80: 38 ± 46.
Stuve O, Dooley NP, Uhm JH, Antel JP, Francis GS,Williams G, Wee Yong V (1996). Interferon b-1bdecreases the migration of T lymphocytes in vitro:Effects on matrix metalloproteinase-9. Ann Neurol 40:853 ± 863.
Tanaka Y, Mine S, Figdor C, Wake A, Hirano H, TsudadaJ, Aso M, Fujii K, Saito K, Van Kooyk Y, Eto S (1998).Constitutive chemokine production results in activa-tion of leukocyte function-associated antigen-1 onadult T-cell leukemia cells. Blood 91: 3909 ± 3919.
Taub D (1996). Chemokine-Leukocyte Interactions. Cyto-kine and Growth Factor Rev 7: 355 ± 376.
Umehara F, Izumo S, Takeya M, Takahashi K, Sato E,Osame M (1996). Expression of adhesion moleculesand monocyte chemoattractant protein-1 (MCP-1) inthe spinal cord lesions in HTLV-I-associated myelo-pathy. Acta Neuropathol (Berl) 91: 343 ± 350.
Zou Y, Kottmann A, Kuroda M, Taniuchi I, Littman D(1998). Function of the chemokine receptor CXCR4 inhaematopoiesis and in cerebellar development. Nature393: 595 ± 599.
Chemokines in HTLV-1 and MSPA Calabresi et al
108