chemical nature of in vivo dna base damage in hydrogen peroxide-treated mammalian cells
TRANSCRIPT

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 285, No. 2, March, pp. 388-390, 1991
COMMUNICATION
Chemical Nature of in Viva DNA Base Damage in Hydrogen Peroxide- Treated Mammalian Cells
Miral Dizdaroglu, * Zeena Nackerdien,**t Bing-Chun Chao,$ Ewa Gajewski,* and Govind Rao$*’ *Center for Chemical Technology, National Institute of Standards and Technology, Gaithersburg, Maryland 20899; iDepartment of Radiotherapy, University of Stellenbosch, Tygerberg, South Africa; and $Chemical and Biochemical Engineering Program and Medical Biotechnology Center of the Maryland Biotechnology Institute, Uniuersity of Maryland Baltimore County, Baltimore, Maryland 21228
Received September 17,1990, and in revised form November 13,199O
Hydrogen peroxide is generated in mammalian cells by normal metabolism or by treatment with external agents. Treatment of mammalian cells with this oxidizing agent results in DNA damage. Little is known about the chemical nature of hydrogen peroxide-mediated DNA damage in mammalian cells. Here we report on the chemical characterization of in vivo base damage to nu- clear DNA in mammalian cells caused by exposure to H202. Chromatin was isolated from cells and analyzed by gas chromatography/mass spectrometry with se- lected-ion monitoring. Ten DNA base products were identified and quantitated. Modified bases identified were typical hydroxyl radical-induced products of DNA bases. Results indicate involvement of hydroxyl radicals in the mechanism of nuclear DNA damage in mammalian cells caused by HzOz. Q 1991 Academic Press, Inc.
Free radicals generated in uiuo have been implicated in a number of human diseases ( 1,2). Superoxide radical and Hz02 are produced in mammalian cells by normal aerobic metabolism or by treatment with external agents (e.g., redox-cycling drugs) ( 1, 3, 4). Toxicity of these species is thought to arise by their metal ion-dependent conversion into hydroxyl radicals (5, 6). Free radicals generated in uivo may damage biomolecules, in- cluding DNA. Thus free radicals are mutagenic and may act as promoters of carcinogenesis (7, 8). Treatment of mammalian cells in culture with Hz02 has been shown to result in DNA damage, mutagenesis, sister chromatid exchange, and chromo- some aberrations (9-11) . However, little is known about the chemical nature of H 2O ,-induced DNA damage in mammalian cells. Understanding of biological consequences and cellular re- pair of the DNA damage in living cells depends on the knowledge of the chemical nature of DNA lesions. HPLC assays are avail-
’ To whom correspondence should be addressed.
388
able for detection of a limited number of free radical-modified bases in DNA ( 12-14). When using these assays, the identifi- cation of an analyte is done by its coelution with the authentic compound without providing any specific evidence of the struc- ture of the analyte. On the other hand, a superior approach is available through the use of gas chromatography/mass spec- trometry with selected-ion monitoring (GC/MS-SIM) ,2 which provides precise identification with structural evidence and quantitation of a large number of free radical-induced lesions in DNA or in chromatin (15-17). Most contemporary mea- surements of chemical modifications to DNA bases have been made on isolated DNA and do not portray a realistic picture of actual damage occurring at the chromatin level. The GC/MS- SIM technique also provides the ability to measure DNA base damage directly in isolated chromatin ( 16,17). The significance of this ability is that it eliminates the inaccuracies associated with extraction of DNA from damaged cells (18). We have used this technique to chemically characterize DNA base damage in the chromatin of H202-treated mammalian cells.
MATERIALS AND METHODS
Cell culture and treatment of cells. The cells used were SP2/0-derived murine hybridomas. Cells were grown in a 5% CO2 atmosphere at 37°C. When cells reached a density of 8.2 X 10’ per milliliter, they were split into six 50-ml parts and transferred into T-125 tissue culture flasks. Cell viability as determined by trypan blue exclusion was 96%. Hz02 was added to the flasks such that two contained 2 mM Hz02 each and two contained 20 mM H202 each. One pair of T-flasks without H202 served as control. All flasks were placed in the incubator at 37°C for 1 h, sampled for cell viability, and then simultaneously placed into an ice water bath. In all cases, cell viability was essentially unchanged. After 10 min, cells were harvested at -2O’C at 1OOOg. The cells were washed by resuspending in phosphate buffer saline containing 0.001% sodium azide and spun down again. Subsequently, chromatin was isolated and characterized as described previously ( 16). One nanomole of 6-azathy-
‘Abbreviation used: GC/MS-SIM, gas chromatography/mass spec- trometry with selected-ion monitoring.
0003-9861/91 $3.00 Copyright 0 1991 by Academic Press, Inc.
All rights of reproduction in any form reserved.
DNA BASE DAMAGE IN HYDROGEN PEROXIDE-TREATED MAMMALIAN CELLS 389
5500
t
11 5000 9
3
I b i Oh..-- I.. .,-
4.0 5.0 6.0 7.0 s.O T.0 10.0 11.0 12.8 13.0 T,me <min.)
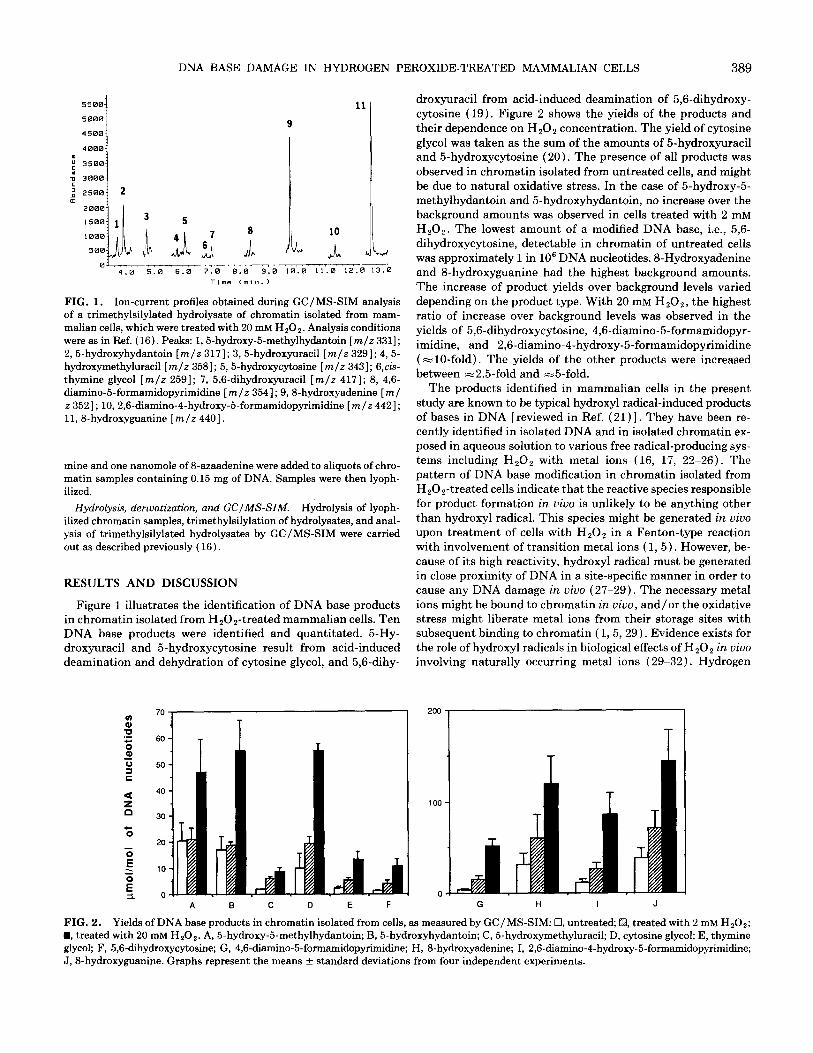
FIG. 1. Ion-current profiles obtained during GC/MS-SIM analysis of a trimethylsilylated hydrolysate of chromatin isolated from mam- malian cells, which were treated with 20 mM HzO,. Analysis conditions were as in Ref. (16). Peaks: 1,5-hydroxy-5methylhydantoin [m/z 3311; 2,5-hydroxyhydantoin [m/z 3171; 3,5-hydroxyuracil [m/z 3291; 4,5- hydroxymethyluracil [m/z 3581; 5, 5-hydroxycytosine [m/z 3431; 6,cis- thymine glycol [m/z 2591; 7, 5,6-dihydroxyuracil [m/z 4171; 8, 4,6- diamino-5-formamidopyrimidine [m/z 3541; 9,8hydroxyadenine [ml z 3521; 10,2,6-diamino-4-hydroxy&formamidopyrimidine [m/z 4421; 11, 8hydroxyguanine [m/z 4401.
mine and one nanomole of 8-azaadenine were added to aliquots of chro- matin samples containing 0.15 mg of DNA. Samples were then lyoph- ilized.
Hydrolysis, deriuatizatzim, and GC/MS-SIM. Hydrolysis of lyoph- ilized chromatin samples, trimethylsilylation of hydrolysates, and anal- ysis of trimethylsilylated hydrolysates by GC/MS-SIM were carried out as described previously ( 16).
RESULTS AND DISCUSSION
Figure 1 illustrates the identification of DNA base products in chromatin isolated from HzOz-treated mammalian cells. Ten DNA base products were identified and quantitated. 5-Hy- droxyuracil and 5hydroxycytosine result from acid-induced deamination and dehydration of cytosine glycol, and 5,6-dihy-
droxyuracil from acid-induced deamination of 5,6-dihydroxy- cytosine (19). Figure 2 shows the yields of the products and their dependence on HzOz concentration. The yield of cytosine glycol was taken as the sum of the amounts of 5-hydroxyuracil and 5-hydroxycytosine (20). The presence of all products was observed in chromatin isolated from untreated cells, and might be due to natural oxidative stress. In the case of 5-hydroxy-5- methylhydantoin and 5-hydroxyhydantoin, no increase over the background amounts was observed in cells treated with 2 mM HzOz. The lowest amount of a modified DNA base, i.e., 5,6- dihydroxycytosine, detectable in chromatin of untreated cells was approximately 1 in lo6 DNA nucleotides. SHydroxyadenine and S-hydroxyguanine had the highest background amounts. The increase of product yields over background levels varied depending on the product type. With 20 mM H,Oz, the highest ratio of increase over background levels was observed in the yields of 5,6-dihydroxycytosine, 4,6-diamino-5-formamidopyr- imidine, and 2,6-diamino-4-hydroxy-5-formamidopyrimidine (=lO-fold). The yields of the other products were increased between =2.5-fold and -5-fold.
The products identified in mammalian cells in the present study are known to be typical hydroxyl radical-induced products of bases in DNA [reviewed in Ref. (21) 1. They have been re- cently identified in isolated DNA and in isolated chromatin ex- posed in aqueous solution to various free radical-producing sys- tems including HzOz with metal ions (16, 17, 22-26). The pattern of DNA base modification in chromatin isolated from Hz0 z-treated cells indicate that the reactive species responsible for product formation in uiuo is unlikely to be anything other than hydroxyl radical. This species might be generated in uiuo upon treatment of cells with H202 in a Fenton-type reaction with involvement of transition metal ions ( 1, 5). However, be- cause of its high reactivity, hydroxyl radical must be generated in close proximity of DNA in a site-specific manner in order to cause any DNA damage in uiuo (27-29). The necessary metal ions might be bound to chromatin in uiuo, and/or the oxidative stress might liberate metal ions from their storage sites with subsequent binding to chromatin ( 1,5,29). Evidence exists for the role of hydroxyl radicals in biological effects of HzOz in uiuo involving naturally occurring metal ions (29-32). Hydrogen
100 -
01
FIG. 2. Yields of DNA base products in chromatin isolated from cells, as measured by GC/MS-SIM: 0, untreated; q treated with 2 mM H,O,; n , treated with 20 mM HZ02. A, 5-hydroxy-5-methylhydantoin; B, 5-hydroxyhydantoin; C, 5-hydroxymethyluracil; D, cytosine glycol; E, thymine glycol; F, 5,6dihydroxycytosine; G, 4,6-diamino-5-formamidopyrimidine; H, 8-hydroxyadenine; I, 2,6-diamino-4-hydroxy-5-formamidopyrimidine; J, 8-hydroxyguanine. Graphs represent the means + standard deviations from four independent experiments.

390 DIZDAROGLU ET AL.
peroxide alone does not cause formation of the DNA products indentified in the present study ( 17, 22-24, 26).
In conclusion, modified DNA bases in chromatin of HzOz- treated mammalian cells were chemically characterized and quantitated for the first time. The pattern of base modification indicates the involvement of hydroxyl radicals in the mechanism underlying DNA base damage in H x0 ,-treated mammalian cells. We have demonstrated the power of the GC/MS-SIM technique for the sensitive and quantitative detection of a large number of DNA base modifications occurring in uivo by exposure of mammalian cells to HzOz. The approach used opens up several avenues not only for investigation of the implications of free radical-mediated DNA damage for a number of pathological conditions, but also in toxicology/mutagenesis studies.
ACKNOWLEDGMENTS
This work has been supported in part by the Office of Health and Environmental Research, Office of Energy Research, U.S. Department of Energy, Washington, DC. G.R. acknowledges support from the Na- tional Science Foundation (EET 8808775). Z.N. acknowledges support from the South African Medical Council.
Certain commercial equipment or materials are identified in this paper in order to specify adequately the experimental procedure. Such iden- tification does not imply recommendation or endorsement by the Na- tional Institute of Standards and Technology, nor does it imply that the materials or equipment identified are necessarily the best available for the purpose.
REFERENCES
1. Halliwell, B., and Gutteridge, J. M. C. (1990) in Methods in En- zymology (Packer, L., and Glazer, A. N., Eds.), Vol. 186, pp. l-85, Academic Press, San Diego.
2. Cross, C. (1987) Ann& Internal Med. 107,526-545. 3. Chance, B., Sies, H., and Boveris, A. (1979) Physiol. Rev. 59,527-
605. 4. Fridovich, I. (1986) Arch. Biochem. Biophys. 247, l-11.
5. Halliwell, B., and Gutteridge, J. M. C. (1988) ZSZ Atlas Sci. B&hem. 1,48-52.
6. Nassi-Calo, L., Mello Filho, A. C., and Meneghini, R. (1989) Car- cimgenesis 10,1055-1057.
7. Cerutti, P. A. (1985) Science 227, 375-381.
8. Weitzman, S. A., Weitberg, A., B., Clark, E. P., and Stossel, T. P. (1985) Science 227, 1231-1233.
9.
10. 11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
Aruoma, 0. I., Halliwell, B., and Dizdaroglu, M. (1989) J. Biol. Chem. 264,13,024-13,028.
Aruoma, 0. I., Ha&well, B., Gajewski, E., and Dizdaroglu, M. (1989) J. Biol. Chem. 264,20,509-20,512. Blakely, W. F., Fuciarelli, A. F., Wegher, B. J., and Dizdaroglu, M. (1990) Radiot. Res. 121, 338-343. Dizdaroglu, M., Aruoma, 0. I., and Halliwell, B. (1990) Biochemistry 29,8447-8451.
26.
27.
Aruoma, 0. I., Halliwell, B., Gajewski, E., and Dizdaroglu, M. (1991) Biochem. J., in press.
Ward, J. F., Blakely, W. F., and Joner, E. I. (1985) Radiat. Res. 103,383-392.
28. Goldstein, S., and Czapski, G. (1986) J. Free Rod. Biol. Med. 2, 3- 11.
29. 30.
Halliwell, B. ( 1987) FASEB J. 1,35&364. Mello Filho, A. C., Hoffman, R. E., and Meneghini, R. (1984) Biochem. J. 218,273-275.
31. Imlay, J. A., and Linn, S. (1988) Science 240,1302-1309.
32. Meneghini, G. (1988) Mutat. Res. 195, 215-230.
Hoffmann, M. E., and Meneghini, R. ( 1979) Photo&em. Photobiol. 30,151-155.
Speit, G. (1986) Mutat. Res. 174, 21-26. Moraes, E. C., Keyse, S. M., and Tyrrell, R. M. (1990) Carcinogenesis 11,283-293. Frenkel, K., Cummings, A., Solomon, J., Cadet, J., Steinberg, J. J., and Teebor, G. W. (1985) Biochemistry 24,4527-4533.
Floyd, R. A., Watson, J. J., Wong, P. K., Altmiller, D. H., and Rich- ard, R. C. (1986) Free Rad. Res. Commun. 1,163-172. Kasai, H., Crain, P. F., Kuchino, Y., Nishimura, S., Ootsuyama, A., and Tanooka, H. (1986) Carcinogenesis 7,1849-1851.
Dizdaroglu, M., and Gajewski, E. ( 1990) in Methods in Enzymology (Packer, L., and Glazer, A. N., Eds.), Vol. 186, pp. 530-544, Academic Press, San Diego.
Gajewski, E., Rao, G., Nackerdien, Z., and Dizdaroglu, M. (1990) Biochemistry 29,7876-7882.
Dizdaroglu, M., Rao, G., Halliwell, B., and Gajewski, E. (1991) Arch. Biochem. Biophys. 285, 317-324.
Mee, L. K., and Adelstein, S. J. ( 1979) Znt. J. R&at. Biol. 36,359- 366. Dizdaroglu, M., Holwitt, E., Hagan, P. A., and Blakely, W. F. (1986) Biochem. J. 236,531-536.
Fuciarelli, A. F., Wegher, B. J., Gajewski, E., Dizdaroglu, M., and Blakely, W. F. (1989) Radiat. Res. 119,219-231. von Sonntag, C. ( 1987) The Chemical Basis of Radiation Biology, Taylor & Fancis, London.