characterization of the homologs of a diagnostically
TRANSCRIPT

CHARACTERIZATION OF THE HOMOLOGS OF A
DIAGNOSTICALLY SIGNIFICANT BRUGIA
MALAYI GENE (Bm17DIII) IN
WUCHERERIA BANCROFTI, LOA LOA AND
ONCHOCERCA VOLVULUS
ROS AZEANA BINTI ABDUL AZIZ
UNIVERSITI SAINS MALAYSIA
2006

CHARACTERIZATION OF THE HOMOLOGS OF A
DIAGNOSTICALLY SIGNIFICANT BRUGIA MALAYI GENE
(Bm17DIII) IN WUCHERERIA BANCROFTI,
LOA LOA AND ONCHOCERCA VOLVULUS
by
ROS AZEANA BINTI ABDUL AZIZ
Thesis submitted in fulfillment of the requirements
for the degree of
Master of Science
June 2006

DEDICATIONS
My father, mother, brothers and sisters

ACKNOWLEDGEMENTS
First, I would like to express my utmost appreciation to my supervisor, Professor (Dr.)
Rahmah Noordin for her constant guidance, critical discussion, concern and patience
throughout the study period.
Gratitude is extended to all the staff and students of the School of Medical Sciences,
School of Health Science, Institute for Research in Molecular Medicine (INFORMM),
Institute of Post-Graduate Studies, Pusat Bahasa and library for the advice, moral
support and technical assistance.
I really appreciate the cooperation and advice that I received from the following
teachers (and the students) namely Prof. Asma Ismail, Prof. Mohd Zaki Salleh, Assoc.
Prof. Rosli Ismail, Prof. Zainul Fadziruddin, and Cikgu Mahani. Thank to Dr. Peter
Fischer (Benhard NOcht Institute for Tropical Medicine, Hamburg, Germany) and Dr.
Dr. B. Ravindran, (Division of Immunology, Regional Medical Research, Indian Council
of Medical Research, Bhubaneswar, India), Prof. J. Ashraful Haq (Department of
Microbiology, Bangladesh Institute of Research & Rehabilitation in Diabetes,
Endocrine & Metabolic Disorders, Daka, Bangladesh) and Filarial Genome Research
Center, Smith College, Northampton, MA, USA for providing the parasite materials,
sera samples and helpful comments.
Personally I would like to thank Dr. Lim Boon Huat for his exhaustive effort, assistance,
moral support and technical advice in helping me finishing up this study. I am thankful
to my dear friends for the guidance and knowledge sharing especially on the molecular

aspects of this thesis namely kak Noor A’Shikin, kak Rohana, kak Azni, kak Suharni,
Hafiz, Muhammad Sarhan, Atif Ali, kak Kiren, kak Aziah, kak Halisa, Roziawati,
Norhayati, Norhaida and also my grateful to the following sisters for their assistance
namely kak Suriati, kak Ida, kak Suzanne, kak Mah and kak Anum.
I would like to express my personal gratitude to my parents, Abdul Aziz Ismail and
Rosni Muhammad for their eternal love and inspiration. Special thanks to Yusshalizaa
Harun for his true love, understanding, great patience and unconditional support.
The financial support from Prof. (Dr.) Rahmah Noordin’s research grants. There were
Malaysian Government’s IRPA grant, No. 06-02-05-1007PR0016/06-05 and European
Commission (EC) grant, No. ICA4-CT-2001-10081.

TABLE OF CONTENTS Contents Page TITLE i DEDICATIONS ii ACKNOWLEDGEMENTS iii TABLE OF CONTENTS v LIST OF ABBREVIATIONS ix LIST OF TABLES x LIST OF FIGURES xi ABSTRAK xv ABSTRACT xviii CHAPTER ONE INTRODUCTION
1.1 Human Filariasis
1
1.2 Lymphatic Filariasis
3
1.3 Wuchereria bancrofti 5 1.3.1 Pathology and Clinical Manifestation 7 1.3.2 Diagnosis of Bancroftian Filariasis 9 1.3.2.1 Direct physical examination and survey 9 1.3.2.2 Thick blood film (TBF) 9 1.3.2.3 Concentration techniques 10 1.3.2.4 Ultrasonography 10 1.3.2.5 Polymerase chain reaction 11 1.3.2.6 Detection of circulating antigen 11 1.3.2.7 Detection of anti filarial IgG4 antibody 12 1.3.3 Treatment and prognosis 13
1.3.4 Global Programme to Eliminate Lymphatic Filariasis (GPELF) 15 1.3.5 Lymphatic Filariasis in the Malaysian Context
16
1.4 Onchocerciasis 19 1.4.1 Onchocerca volvulus 21 1.4.2 Life cycle of Onchocerca volvulus 21 1.4.3 Pathology and clinical disease 22 1.4.4 Diagnosis of onchocerciasis 25 1.4.4.1 Histopathological methods 25 1.4.4.2 Mazzotti test 26 1.4.4.3 Serodiagnostic assays 26 1.4.4.5 Polymerase chain reaction 27
1.4.5 Treatment of onchocerciasis
28
1.5 Loiasis 29 1.5.1 Life cycle of Loa loa 31 1.5.2 Pathology and clinical disease of L. loa 33

1.5.2.1 Diagnosis of loiasis 33 1.5.2.2 Serological test 34 1.5.2.3 Parasitological assessment 34 1.5.2.4 Polymerase chain reaction 34 1.5.2.5 Direct examination based on clinical signs 35
1.5.3 Treatment of loiasis 35 1.5.4 Prevention and control programs for loiasis
36
1.6 Statement of the problem
36
1.7 Rationale of the study
39
1.8 Objectives of the research project 40 CHAPTER TWO MATERIALS AND METHODS
2.1 General overview
41
2.2 Web-based homology search using multiple DNA and protein databases
44
2.3 BRUGIArapid™ procedure
45
2.4 Sources of parasite DNA
47
2.4.1 W. bancrofti lyophilized mf samples 47 2.4.2 Whole blood samples
47
2.5 Sources of sera samples for immunoassays
48
2.6 cDNA libraries 48 2.6.1 Amplification of cDNA libraries 49 2.6.2 Determination of best titers of cDNA libraries
52
2.7 DNA extraction methods 55 2.7.1 Extraction of DNA from lyophilized mf 55 2.7.2 Extraction of DNA from blood sample 57 2.7.3 Extraction of DNA from lambda (�) phage of amplified cDNA library
58
2.7.4 Preparation of FTA® disc for PCR analysis
59
2.8 Determination of yield and quality of the extracted DNA
59
2.9 Polymerase chain reaction (PCR) 59 2.9.1 Optimization of PCR parameters 67
2.9.1.1 Quality check on W. bancrofti genomic DNA for use in PCR amplification
67
2.9.1.2 Optimization of template quantity 68 2.9.1.3 Optimization of the annealing temperature 68 2.9.1.4 Optimization of MgCl2 concentration 69 2.9.1.5 Selection of suspension medium (SM buffer) 70

2.9.2 Analysis of the intron site 70 2.9.2.1 Analysis of the intron of Wb17DIII gene sequence
71
2.10 Agarose gel electrophoresis (AGE) 73 2.10.1 Agarose gel preparation 73 2.10.2 Loading and running the gel
73
2.11 Southern blot analysis 74 2.12 Chemiluminescences (ECL) technique for development of Western blots 76 2.13 Purification of PCR product 78
2.13.1 Dissolving the gel slice 78 2.13.2 DNA purification
78
2.14 TOPO cloning and DNA sequence analysis 79 2.14.1 Preparation of competent cells 79
2.14.2 TOPO-cloning 80
2.14.3 PCR screening 81 2.14.4 Plasmid preparation 81 2.14.5 Sequencing and DNA analysis
82
2.15 Preparation of glycerol cell stocks
83
2.16 Sub-cloning into bacterial expression vector 83 2.16.1 pPROEX TMHT/ Ov17DIII 83 2.16.2 Restriction digestion by EcoR1 84 2.16.3 Rapid ligation and transformation
85
2.17 Protein expression and purification 87 2.17.1 Time-course analysis of protein expression 87 2.17.2 Sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE)
87
2.17.3 Larger scale protein expression 88 2.17.4 Purification of recombinant protein 89 2.17.5 Determination of total protein concentration
89
2.18 Western blot analysis
90
2.19 Enzyme-linked immunosorbent assay (ELISA) and its evaluation 93 CHAPTER THREE RESULTS
3.1 Web-based search for homologs of Bm17DIII gene in multiple DNA and
protein databases of W. bancrofti, L. loa and O. volvulus
95
3.2 Determination of the best titers of the cDNA libraries
99
3.3 Optimization of PCR for amplification of the homolog of Bm17DIII gene 101

3.3.1 Quality check on W. bancrofti genomic DNA for use in PCR amplification
101
3.3.2 Optimizations of annealing temperature, MgCl2 concentration and template quantity
104
3.3.3 Selection of suspension medium (SM) for λ phage amplification
108
3.3.4 Amplification of Bm17DIII homologs in genomic DNA and cDNA libraries
111
3.4 Southern blot analysis on cDNA libraries of W. bancrofti 113 3.5 TOPO-TA cloning and sequencing analysis 116
3.5.1 Sequencing analysis of pTOPO ⁄ Wb17DIII 119 3.5.2 Sequencing analysis of Ov17DIII/TOPO or Ll17DIII/TOPO
119
3.6 Analysis of the intron sites of Bm17DIII and Wb17DIII 122 3.6.1 Analysis of the intron site of B. malayi genomic DNA 122 3.6.2 Analysis of the intron site of W. bancrofti genomic DNA
124
3.7 Map of pPROEXTMHTa ⁄ Ov17DIII plasmid
133
3.8 Subcloning Into Bacterial Expression Vector 137
3.9 Analysis of Ov-BmR1 by SDS-PAGE and Western blot analysis
141
3.10 Antibody reactivities to B. malayi BmR1 antigen and its homologs in O. volvulus (Ov-BmR1)
146
CHAPTER FOUR DISCUSSION
150
4.1 Web-based search for homologs of Bm17DIII in W. bancrofti, O. volvulus and L. loa
154
4.2 Laboratory-based studies to isolate homologs of Bm17DIII in W. bancrofti, O. volvulus and L. loa
156
4.3 Expression of Ov17DIII or Ll17DIII and purification of Ov-BmR1
161
4.4 Antibody reactivity to BmR1 and its homologs in O. volvulus (Ov-BmR1 )
165
4.5 Conclusion 167 REFERENCES
169
APPENDIX
184
LIST OF PUBLICATIONS 192

LIST OF ABBREVIATIONS
No.
Abbreviations Full words
1 BLAST Basic Local Alignment Search Tool
2 BR BRUGIArapid
3 conc Concentration
4 COV Cut off value
5 ddH20 Double distilled water
6 dH20 Deionised water
7 g Gram
8 g Gradient
9 hr Hour
10 L1 Larva stage 1
11 L2 Larva stage 2
12 L3 Larva stage 3
13 L4 Larva stage 4
14 LF Lymphatic filariasis
15 mf Microfilaria
16 min Minute
17 NC Negative control
18 nr Non-redundant
19 OD Optical density
20 p Page
21 PC Positive control
22 pfu Plaque forming unit
23 rpm Revolution per minute
24 RT Room temperature
25 sec Second
26 T Time
27 � Lambda
28 + Positive
29 - Negative/ minus
30 � Micro
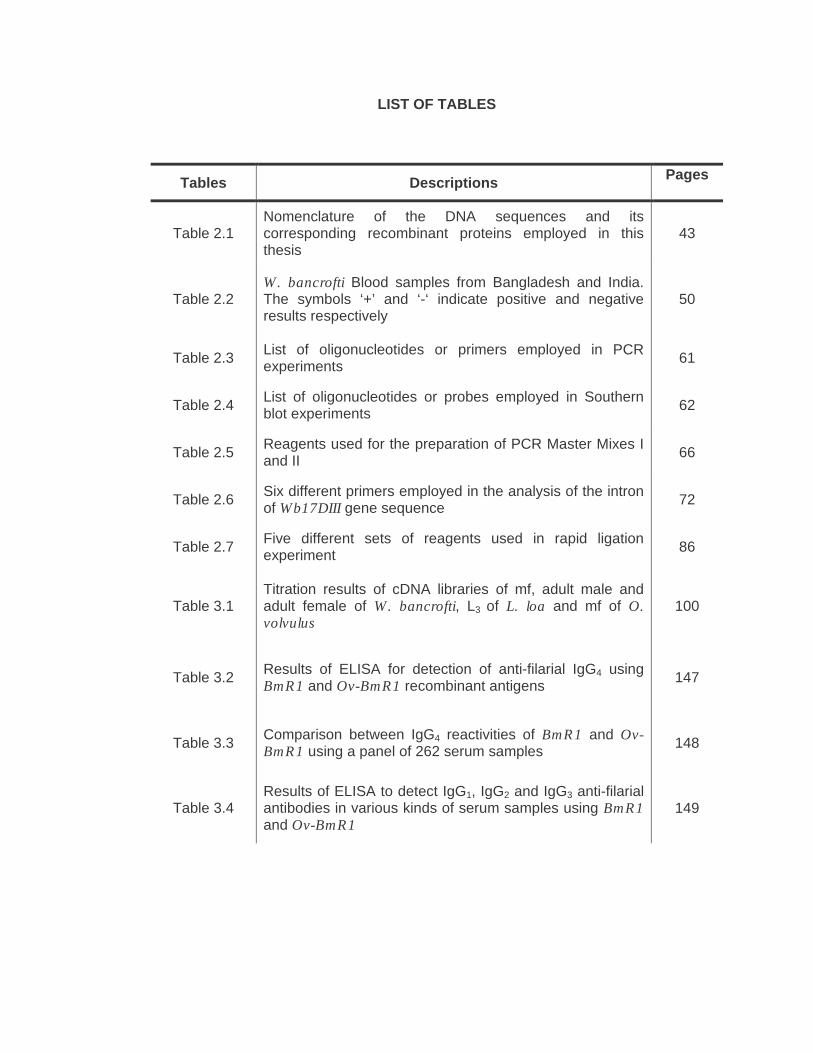
LIST OF TABLES
Tables Descriptions Pages
Table 2.1 Nomenclature of the DNA sequences and its corresponding recombinant proteins employed in this thesis
43
Table 2.2 W. bancrofti Blood samples from Bangladesh and India. The symbols ‘+’ and ‘-‘ indicate positive and negative results respectively
50
Table 2.3 List of oligonucleotides or primers employed in PCR experiments 61
Table 2.4 List of oligonucleotides or probes employed in Southern blot experiments 62
Table 2.5 Reagents used for the preparation of PCR Master Mixes I and II 66
Table 2.6 Six different primers employed in the analysis of the intron of Wb17DIII gene sequence 72
Table 2.7 Five different sets of reagents used in rapid ligation experiment 86
Table 3.1 Titration results of cDNA libraries of mf, adult male and adult female of W. bancrofti, L3 of L. loa and mf of O. volvulus
100
Table 3.2 Results of ELISA for detection of anti-filarial IgG4 using BmR1 and Ov-BmR1 recombinant antigens 147
Table 3.3 Comparison between IgG4 reactivities of BmR1 and Ov-BmR1 using a panel of 262 serum samples 148
Table 3.4 Results of ELISA to detect IgG1, IgG2 and IgG3 anti-filarial antibodies in various kinds of serum samples using BmR1 and Ov-BmR1
149

LIST OF FIGURES
Figures Descriptions Pages
Figure 1.1 A map showing filariasis endemic countries 2
Figure 1.2 The schematic life cycle of W. bancrofti 6
Figure 1.3 Elephantiasis of the legs due to the lymphatic 8
Figure 1.4 Two signs of onchocerciasis 20
Figure 1.5 The incubation phase of filarial parasite which affective from the point of infection to the time of appearance of first mf and resulted in eye lesions, altered pigmentation and loss of elasticity of skin
24
Figure 1.6 Both figures show the signs of loiasis; (a) African eye worm and (b) Calabar swellings on the hands 30
Figure 1.7 The schematic life cycle of L. loa 32
Figure 2.1 Bm17DIII gene sequence and its translated amino acid sequence obtained from the GenBank accession number AF225296 and GenBank protein I.D AAQ13914
42
Figure 2.2 BRUGIArapid™ dipsticks used for detection of Brugia infection in serum, plasma or whole blood samples 46
Figure 2.3 Amplified λ plaques growth on LB agar with a layer of top agarose. 51
Figure 2.4 Serial dilutions and plating of cDNA library. Each tube contains 90 µl SM buffer 54
Figure 2.5 Extraction of DNA from lyophilized mf using Genispin™ Tissue DNA Kit 56
Figure 2.6 Map of Bm17DIII gene showing the locations of each primer and probe and the sites of intron and exon 63
Figure 2.7 Capillary transfer of DNA from agarose gel 77
Figure 2.8 The materials arranged in a sandwich form used in the semi dry electrotransfer 92
Figure 2.9 Specific detection of immunoblotted proteins using antibodies 92

Figure 3.1 Result of similarity search from nematode database showing the O. volvulus DNA sequence which is similar to the sequence of Bm17DIII gene
96
Figure 3.2 Result of similarity search from nematode database showing the O. volvulus DNA sequence which is similar to the sequence of Bm17DIII gene
97
Figure 3.3a Result of the amplified SspI repeat regions of W. bancrofti genomic DNA extracted from blood samples 102
Figure 3.3b SspI repeat sequence (size of 195 bp) obtained from W. bancrofti lyophilized mf DNA 103
Figure 3.4 PCR amplification results of W. bancrofti infected blood samples applied on FTA cards using NV1 and NV2 primers 103
Figure 3.5 Optimization result of annealing temperature using RT57 W. bancrofti lyophilized mf sample 105
Figure 3.6 Result of optimization of the annealing temperature using variable sources cDNA libraries namely W. bancrofti mf cDNA library, O. volvulus mf cDNA library and L. loa L3 cDNA library
105
Figure 3.7a Optimization result of MgCl2 concentration performed on RT57 W. bancrofti genomic DNA sample 106
Figure 3.7b Optimization result of MgCl2 concentration performed on W. bancrofti mf cDNA libraries 106
Figure 3.7c Optimization result of MgCl2 concentration performed on O. volvulus mf cDNA libraries 107
Figure 3.8 Optimization result of template quantity performed on RT57 W. bancrofti genomic DNA sample 107
Figure 3.9 A gel photo of various stages of W. bancrofti mf cDNA library collected in Medium A (SM buffer with gelatin) 109
Figure 3.10 A gel photo of various stages of W. bancrofti mf cDNA library collected in either Medium B (SM buffer with no gelatin) or Medium C (nuclease-free water)
110
Figure 3.11a PCR amplification results of homolog of Bm17DIII gene in cDNA and DNA of W. bancrofti performed at PCR optimal conditions 112
Figure 3.11b PCR amplification results of homolog of Bm17DIII gene in cDNA libraries of O. volvulus mf and L. loa L3 at PCR optimal conditions 112

Figure 3.12 PCR product of W. bancrofti mf cDNA libraries employed using RNF and RNR primers 114
Figure 3.13 Result of Southern blotting performed on PCR product of W. bancrofti mf cDNA library probed with Bm4c 114
Figure 3.14 Result of Southern blotting performed on PCR product of W. bancrofti mf cDNA library probed with Wb2 115
Figure 3.15 Result of Southern blotting performed on PCR product of W. bancrofti mf cDNA library probed with Bm1c 115
Figure 3.16a Purified pTOPO ⁄ Wb17DIII recombinant plasmids prepared for sequencing analysis 117
Figure 3.16b Purified pTOPO ⁄ Ov17DIII and pTOPO ⁄ Ll17DIII recombinant plasmids prepared for sequencing analysis 117
Figure 3.17 DNA sequence map of pTOPO ⁄ Wb17DIII, pTOPO ⁄ Ov17DIII or pTOPO ⁄ Ll17DIII plasmid. Restriction sites are labeled to indicate the actual cleavage sites
118
Figure 3.18 The DNA and amino acid sequences of BmR1 and homologs in W. bancrofti, O. volvulus and L. loa 121
Figure 3.19a PCR amplification results on B. malayi adult worms DNA using RNF and RNR primers 123
Figure 3.19b PCR amplification results on B. malayi adult worms DNA using RNF and RNR primers 123
Figure 3.20 PCR amplification results of W. bancrofti infected blood samples applied on FTA cards using RNF and RNR primers 126
Figure 3.21 PCR products of four W. bancrofti DNA samples using RIR and RIF primers 127
Figure 3.22 DNA sequence of PCR product of W. bancrofti genomic DNA (RT57) amplified using RIF and RIR primers 128
Figure 3.23 Blastn alignment result between Wb17DIII intron sequence (RT57) and a B. malayi mf cDNA library (AI043417) 129
Figure 3.24a PCR amplification results of RT21 W. bancrofti genomic DNA using different sets of primers to search for the Wb17DIII intron site 130
Figure 3.24b PCR amplification results of RT22 and RT82 W. bancrofti genomic DNA samples using different sets of primers 130

Figure 3.25 PCR amplification results of W. bancrofti infected blood samples applied on FTA cards using RNF and RNR2 primers 131
Figure 3.26 PCR amplification results of W. bancrofti DNA samples using RNF2 and RNR primers 132
Figure 3.27 Map of pPROEXTMHTa ⁄ Ov17DIII/ or pPROEXTMHTa ⁄ Ll17DIII/ recombinant plasmid 134
Figure 3.28 Sequence map of pPROEX HTa ⁄ Ov17DIII/ or pPROEX HTa ⁄ Ll17DIII recombinant plasmid 136
Figure 3.29a Purified DNA of pPROEX HTa before and after cutting with EcoRI in a single digestion experiment 138
Figure 3.29b Purified digested DNA of pPROEX HTa 138
Figure 3.30a Purified digested DNA of Ov17DIII before and after cutting with EcoRI in a single digestion experiment 139
Figure 3.30b Purified digested DNA of Ov17DIII 139
Figure 3.31 Ligated plasmid of pPROEX HTa to the 630 bp DNA insert of Ov17DIII 140
Figure 3.32 Analysis of time point samples of protein from non-induced and induced recombinant bacterial clones (Ov-BmR1) 142
Figure 3.33 Protein profiles of purified recombinant antigens of BmR1 and its homolog (Ov-BmR1) 143
Figure 3.34 Immunorecognition of Ov-BmR1 recombinant antigen by human sera 144
Figure 3.35 Nitrocelluose strips showing the immunorecognition of Ov-BmR1 recombinant antigen detected by anti-His (H3) probe 145

PENCIRIAN HOMOLOG BAGI GEN BRUGIA MALAYI (Bm17DIII) YANG
SIGNIFIKAN SECARA DIAGNOSTIK DI DALAM WUCHERERIA BANCROFTI, LOA
LOA
DAN ONCHOCERCA VOLVULUS
ABSTRAK
Satu ujian pantas yang dikenali sebagai BRUGIArapid (BR) yang mengesan antibodi
IgG4 terhadap antigen rekombinan B. malayi (BmR1) adalah sangat berguna dalam
pemetaan dan pengawasan kawasan yang endemik bagi filariasis brugia. Kajian
penilaian lapangan yang telah dilakukan menunjukkan bahawa BmR1 adalah sangat
sensitif dan spesifik dalam mengesan individu bermikrofilaria. Corak reaktiviti IgG4
terhadap BmR1 daripada sampel individu yang terjangkit dengan W. bancrofti adalah
berbeza-beza mengikut kawasan, manakala reaktiviti BmR1 terhadap sampel serum
daripada individu yang dijangkiti O. volvulus dan L. loa adalah sangat minima.
Memandangkan antigen rekombinan BmR1 adalah signifikan secara diagnostik, maka
adalah penting untuk mencirikan BmR1 secara lebih mendalam. Perbezaan reaktiviti
BmR1 terhadap sampel serum daripada pesakit yang dijangkiti parasit filarial yang lain
(selain B. malayi & B. timori) mencetuskan persoalan samada antigen tersebut
mempunyai gen homolog di dalam W. bancrofti (Wb-BmR1), O. volvulus (Ov-BmR1)
dan L. loa (Ll-BmR1). Sehubungan dengan itu, dalam kajian ini, jujukan DNA dan/atau
cDNA bagi W. bancrofti, O. volvulus dan L. loa dikenalpasti, proteinnya diekspresi dan
reaktiviti terhadap sampel serum pesakit diuji.

Pencarian homolog berasaskan web bagi Bm17DIII di dalam W. bancrofti, O. volvulus
dan L. loa menggunakan beberapa pangkalan data DNA melalui server BLASTN
menunjukkan tiada sebarang persamaan dengan jujukan W. bancrofti dan L. loa,
manakala bagi O. volvulus, terdapat persamaan dengan dua jujukan L3 dengan nilai
E-value yang rendah. Pencarian homolog melalui BLASTP (pangkalan data protein)
menunjukkan protein BmR1 tidak mempunyai persamaan dengan jujukan protein lain.
Kaedah PCR digunakan untuk pemencilan dan identifikasi jujukan cDNA daripada
perpustakaan cDNA dan/atau DNA genomik daripada W. bancrofti, O. volvulus dan L.
loa menggunakan primer RNR dan RNF. Produk PCR bersaiz 618 bp (iaitu Wb-BmR1,
Ov-BmR1 dan Ll-BmR1) kemudiannya diklon ke dalam vector TOPO, dijujuk dan
dianalisis menggunakan program Vector NTI dan server BLAST. Wb17DIII didapati
100% menyerupai jujukan BmR1, manakala Ov17DIII dan Ll17DIII didapati mirip
terhadap satu sama lain dan berkongsi homologi sebanyak 99.7% dengan Bm17DIII.
Oleh itu keputusan pencarian gen homolog Ov17DIII melalui web tidak selaras dengan
keputusan yang diperolehi dalam makmal. Kajian ini juga menunjukkan bahawa
keseluruhan gen Wb17DIII tidak mengandungi sebarang intron; dengan itu berbeza
daripada gen Bm17DIII yang mempunyai satu intron dan dua ekson.
Gen rekombinan Ov17DIII or Ll17DIII kemudiannya diekspresi di dalam pPROEXTM
HTa/TOP10F. Molekul Ov-BmR1 or Ll-BmR1 didapati bersaiz ~25 kDa dan analisis
secara blot Western menunjukkan ia reaktif terhadap sampel serum daripada pesakit
L. loa dan O. volvulus yang bermikrofilaria dan tidak reaktif dengan sampel serum
daripada penderma darah yang sihat. Dengan menggunakan kaedah IgG4-ELISA,
corak pengenalan antibodi IgG4 dalam semua sampel serum didapati sama terhadap

BmR1 dan Ov-BmR1 or Ll-BmR1. ini termasuklah reaktiviti antibodi IgG4 yang lemah
yang dipamerkan oleh sampel serum daripada pesakit yang dijangkiti O. volvulus dan
L. loa. Kajian tentang reaktiviti terhadap subkelas IgG yang lain menunjukkan bahawa
sampel serum daripada pesakit yang dijangkiti O. volvulus dan L. loa memberikan
keputusan positif (bila diuji dengan Ov-BmR1 or Ll-BmR1 atau BmR1) hanya dengan
IgG1 dan tidak dengan subkelas IgG2 atau IgG3. Begitu juga reaktiviti BmR1 terhadap
sampel serum daripada individu yang terjangkit dengan B. malayi dan W. bancrofti
(jangkitan aktif dan kronik) turut memberi reaktiviti positif terhadap IgG1 dan negatif
terhadap subkelas antibodi IgG2 atau IgG3. Namun begitu, sampel serum daripada
individu normal dan yang dijangkiti cacing bawaan tanah juga menunjukkan corak
reaktiviti yang serupa (iaitu positif dengan IgG1 dan negatif dengan IgG2 & IgG3).
Kajian ini menunjukkan bahawa homolog bagi antigen rekombinan BmR1 wujud di
dalam W. bancrofti, O. volvulus dan L. loa dengan konservasi yang tinggi.
Pengecaman antigen (BmR1, Wb-BmR1, dan Ov-BmR1 or Ll-BmR1) oleh sampel
serum pesakit adalah sama terhadap IgG1, IgG2 atau IgG3, tetapi berbeza bagi
antibodi IgG4. Kesimpulannya ialah, walaupun antigen BmR1 adalah sesuai digunakan
untuk pengesanan antibodi IgG4 terhadap jangkitan filariasis brugia, bagaimanapun,
protein homolognya (Wb-BmR1, Ov-BmR1 dan Ll-BmR1) tidak sesuai digunakan bagi
tujuan pengesanan jangkitan penyakit filaria yang lain.

CHARACTERIZATION OF THE HOMOLOGS OF A DIAGNOSTICALLY
SIGNIFICANT BRUGIA MALAYI GENE (Bm17DIII) IN WUCHERERIA BANCROFTI,
LOA LOA
AND ONCHOCERCA VOLVULUS
ABSTRACT
An antibody-detection rapid test, BRUGIArapid, that detects IgG4 antibodies reactive to
a recombinant B. malayi antigen (BmR1), is a promising tool for mapping and
monitoring the areas where brugian filariasis is endemic. Field trials have revealed that
BmR1 is highly sensitive and specific in detecting microfilariaemic individuals. In sera
of individuals infected with Wuchereria bancrofti the IgG4 reactivity to BmR1 is
variable, and cross-reactivity of sera from individuals infected with O. volvulus or L. loa
was observed only in single cases.
Due to its diagnostic significance, it is therefore important to characterize the BmR1
antigen more closely. The varying degree of BmR1 recognition in other filarial
infections (other than B. malayi & B. timori) raises the question whether the
homologous antigen is also present in W. bancrofti (Wb-BmR1), O. volvulus (Ov-
BmR1) and L. loa (Ll-BmR1). In this study, the respective cDNA sequences were
identified, the protein expressed and the antibody reactivities of patients’ sera to the
homologous recombinant antigens was studied.
Web-based homology searches for homologs of Bm17DIII in W. bancrofti, O. volvulus
and L. loa via BLASTN server of several DNA databases resulted in no similarity to
any sequence of W. bancrofti and L. loa, while for O. volvulus, there were two L3

sequences which had a low E-value. Homology searches via BLASTP (protein
databases) revealed that BmR1 protein did not have any similarity with other protein
sequence.
PCR was used to isolate the cDNA sequences from cDNA libraries and/or genomic
DNA of W. bancrofti, O. volvulus and L. loa based on RNR & RNF primers. The 618 bp
PCR products (namely Wb17DIII, Ov17DIII and Ll17DIII) was then cloned into TOPO
vector, sequenced and analysed using Vector NTI software and BLAST server.
Wb17DIII was found to be 100% identical to Bm17DIII, while Ov17DIII and Ll17DIII
were found to be identical to each other and shared 99.7% homology with Bm17DIII.
Thus the results of the web-based search for Ov17DIII were not in agreement with the
laboratory results. This study also revealed that, unlike the complete Bm17DIII gene
which contains an intron (and two exons), the complete Wb17DIII gene did not
possess any intron.
The Ov17DIII or Ll17DIII recombinant gene was then expressed in pPROEXTM
HTa/TOP10F. The MW of Ov-BmR1 or Ll-BmR1 was ~25 kDa and analysis by
Western blot showed reactivity with sera from L. loa and O. volvolus mf+ patients and
no reactivity with serum from healthy blood donor. By employing IgG4-ELISA, the
pattern of IgG4 recognition of all serum samples to Ov-BmR1 or Ll-BmR1 and BmR1
was found to be identical. This included weak IgG4 reactivities demonstrated by sera
from L. loa- and O. volvulus-infected patients. With respect to reactivities to other IgG
subclasses, sera from O. volvulus- and L. loa- infected patients showed positive
reactions (when tested with Ov-BmR1 or Ll-BmR1 or BmR1) only with IgG1; and no
reactivity was observed with IgG2 or with IgG3. Similarly, sera from individuals infected

with B. malayi or W. bancrofti (active and chronically-infected patients) were positive
with BmR1 only for IgG1 and were negative when tested with IgG2 and IgG3
subclasses. However, it is also noted that sera from non-endemic normals and soil-
transmitted helminth infections also showed similar reactivities i.e. IgG1 positive and
IgG2 and IgG3 negative.
This study demonstrated that Bm17DII gene and its homologs in W. bancrofti, O.
volvulus and L. loa are highly conserved. Recognition of the recombinant gene
products (BmR1 or Wb-BmR1 and Ov-BmR1 or Ll-BmR1) by patients’ sera are similar
with regard to IgG1, IgG2 and IgG3, but different for IgG4 antibodies. Thus this study
demonstrated that although IgG4 antibodies to BmR1 are a good infection marker for
brugian filariasis, its homologs are not of diagnostic value.


1
CHAPTER ONE
INTRODUCTION
1.1 Human Filariasis
Filariasis is caused by roundworms that inhabit the lymphatic and subcutaneous
tissues and cause some of the most debilitating diseases, including elephantiasis and
river blindness (WHO, 2002a,b; 2003). Human filarial infections are endemic in tropical
regions (Figure 1.1) and affect an estimated 200 million people, and exposing another
billion to the risk of infection (Ottesen et al., 1997). The parasites are transmitted by
blood-feeding insects, which act as vectors. Filarial nematodes enter the human body
at their third larval stage by escaping from the mouthparts of their vector arthropod as
they bend during biting and enter through the bite wound in the skin.
Eight main species infect human namely Wuchereria bancrofti, Brugia malayi and
Brugia timori which causes lymphatic filariasis; Loa loa which causes loaisis;
Onchocerca volvulus which causes onchocerciasis; and three species of Mansonella.
Filariasis is rarely fatal, it is the second leading cause of permanent and long-term
disability in the world. The World Health Organization (WHO) has named filariasis one
of only six “potentially eradicable” infectious diseases and has embarked upon a 20-
year campaign to eradicate the disease. The immunologic hallmark of infections by
filarial parasites is induction of allergic type responses (Bundy et al., 1991, King, 2000).
Typically this produces peripheral eosinophilia and elevated levels of polyclonal and
parasite specific IgE. Filarial specific IgG subclasses are also present, with IgG4
subclass most prominently elevated (Ottesen et al., 1985).

2
Figure 1.1 A map showing filariasis endemic countries (WHO, 1997).
1.2 Lymphatic Filariasis

3
Lymphatic filariasis (LF) also known as elephantiasis is a disabling and disfiguring
infection caused by W. bancrofti, B. malayi or B. timori (Michael and Bundy, 1997). It is
estimated that 120 million people or 2% of the world’s population are infected in around
80 countries throughout the tropics and subtropics (WHO, 2000; WHO, 2002;
Molyneux, 2003).
It is a major cause of acute and chronic morbidity affecting persons of all ages and
both sexes (Ottesen, et al., 1997) and prevails in those of low socioeconomic level
(Dreyer, et al., 2000). LF is a major burden on health and hospital resources
(Gyapong, et al., 1996) includes high medical expenses, loss of productivity,
diminished social function and reduced quality of life (Evans, et al., 1993).
Approximately 44 million people demonstrate signs of elephantiasis, lymphoedema
and genital pathology (Ottesen, et al., 1997). Persons with chronic manifestations of
the disease are often unable to work or marry, become dependent for care and
financial support and consequently lead to lost self-confidence. Thus, LF has been
identified by the World Health Organization (WHO) as the second leading causes of
permanent and long-term disability world-wide (WHO, 1997).
These filarial infections affect individuals from all age groups and races. Men are more
susceptible compared to woman and children (Kazura, 1999). This phenomenon may
be explained by the greater exposure of men to mosquito vectors at work. In endemic
areas, children are usually exposed to this infection early in their life and as many as
one third of the children were infected before the age of five (Witt & Ottesen, 2001).
The total global burden of LF is not known and mapping of its endemicity and
prevalence is on-going. LF is the most prevalent filarial infection of humans, most of
which is caused by W. bancrofti, while the closely related species, B. malayi and B.
timori cause the remaining infections (Taylor, 2003). Of the three parasites causing LF,

4
W. bancrofti accounts for over 90% of the global burden (WHO, 2002a,b). It is endemic
in India, Africa, South America, Indonesia, Burma, Vietnam, Egypt and China. B.
malayi is limited in distribution to Asia (India, China, Indonesia, Philippines, Thailand,
Vietnam and Malaysia) and B. timori to a few islands in Indonesia (WHO, 2003).
Basically, three groups of people will be found in filarial-endemic areas (Ottesen, 1992,
1993; Ottesen, and Campbell, 1994). There are those who are exposed to the infection
but display no evidence of disease, these are the so-called as endemic normals; the
second group are those who are clinically asymptomatic but demonstrate presence of
the larvae (asymptomatic microfilaraemics). The third groups are those with chronic
disease such as chronic lymphoedema, hydrocele and elephantiasis. Infected people
suffer episodes of acute filarial disease such as acute filarial lymphangitis. This is
believed to be partly due to the result of an immunological reaction to dead or dying
adult worms which have either been killed by the immune system or by chemotherapy
(Dreyer et al., 1999a, b). The compromised lymphatic function coupled with
accumulation of protein-rich fluid in the tissue predisposes the limbs to bacterial
secondary infection, and is an important risk factor for development of elephantiasis
(Dissanayake et al., 1995)
Lymphatic filariasis can be transmitted by more than 70 species and subspecies of
mosquitoes. However, the principal mosquito species that are efficient vectors are
found in the following four genuses: Anopheles (W. bancrofti, B. malayi and B. timori),
Aedes (W. bancrofti and B. malayi), Culex (W. bancrofti) and Mansonia (W. bancrofti
and B. malayi) (Scott, 2000). Rural-urban migration and uncontrolled urbanization often
lead to over burdening of sewerage and waste-water systems. The resulting pools of
stagnant, polluted water provide an ideal breeding ground for Culex quinquefasciatus,
a major vector of filariasis (Ottesen, et al., 1997; Mak, 1987). In contrast to

5
Anophelines, Culicines can efficiently transmit filariasis in situations where the
microfilaria (mf) density is low (Webber, 1991).
1.3 Wuchereria bancrofti
W. bancrofti is a filarial nematode which lives in lymphatic channels and lymph nodes
of humans. The adult worm is elongated and slender (30 to 100 mm long by 100 to 300
µm wide); and males are about half the size of females. The width of the microfilaria
(mf) is the diameter of a red blood cell and the length is 250 to 300 µm. Adults produce
mf measuring 244 to 296 µm by 7.5 to 10 µm, which are sheathed and have nocturnal
periodicity, except for the South Pacific strain which have the absence of marked
periodicity (Cross, 2003). W. bancrofti belongs to the class of Secernentea, subclass of
Spiruria, a family of Filariidae and super family of Filarioidea.
Elephantiasis has been written about since the time of the early Greeks and Romans.
The larval mf was first seen in hydrocele fluid by the French surgeon Jean-Nicolas
Demarquay in 1863 and in urine by Otto Henry Wucherer in Brazil in 1866. The adult
worm was described by Joseph Bancroft in 1876 and named Filarial bancrofti in his
honor by the British helminthologist, Thomas Spencer Cobbold. The discovery of the
life cycle by Patrick Manson in 1877 is regarded as one of the most significant
discoveries in tropical medicine (Cross, 2003).
The life cycle of W. bancrofti is shown in figure 1.2. When an infected mosquito bites a
person who has lymphatic filariasis, the mf circulating in the person's blood enter and
infect the mosquito. The mf passes from the mosquito through skin, and travel to lymph
vessels. In the human lymph vessels they grow into adults and these adult worm lives
for about 5–10 years. When matured, the adult worms mate and release millions of mf
into their host blood system; these in turn are picked up by mosquitoes during their
blood meal.

6
Figure 1.2 The schematic life cycle of W. bancrofti (CDC Image Library, 2004).

7
1.3.1 Pathology and Clinical Manifestation
LF can present a wide variety of clinical manifestations, ranging from apparently
asymptomatic cases to severe disfigurement of the limbs and genitalia. Following
infection with third stage larvae there is usually a period of vigorous immune response
to the invading larvae (Dreyer et al., 2000). The incubation phase from the point of
infection to the time of appearance of first mf seems to be symptomless, but in some
cases transient lymphatic inflammation occurred. Then the second stage, acute or
inflammatory phase follows. This is the time when females reach maturity and releases
the mf causing intense lymphatic inflammation, chills, fever, swollen lymph nodes,
hydrocele and lymphadenitis to the infected human (Dreyer et al., 2000).
Lymphoedema is the worst clinical manifestations of lymphatic filariasis. An important
contributor to this condition is the repeated adenolymphangitis and cellulitis caused by
secondary bacterial infections (Dissanayake et al., 1995). This can result in gross
enlargement of the affected limbs and with W. bancrofti infections, these enlargements
are usually unilateral (Figure 1.3 a, b, c, d). Besides being physically handicapped,
patients’ abilities to carry out their daily activities are greatly restricted. There are four
different stages of lymphoedema. In the initial stage, the lymphoedema can
spontaneously be reversible through elevation and resulted in no skin changes. In the
final stage, the lymphoedema is non-reversible, skin thickens and accompanied by
warts or papilloma (WHO, 1992).
The mf in the blood and lungs can also cause an IgE-mediated allergic response which
can result in asthma-like symptoms (tropical pulmonary eosinophilia or TPE).

8
(a) (b) (c) (d)
Figure 1.3 Elephantiasis of the legs due to the lymphatic filariasis, (a) Elephantiasis
of the legs due to lymphatic filariasis; (b) Elephantiasis of the leg in a
young mother (c) Comparison of a normal and diseased leg of a young
woman with elephantiasis; (d) Close-up of the legs of a 64-year-old coir
factory worker with elephantiasis. She has been infected for 45 years
(TDR Image Library, 2005).

9
1.3.2 Diagnosis of Bancroftian Filariasis
LF is diagnosed by a combination of the appropriate epidemiological history, physical
findings and laboratory tests. Accurate and early diagnosis of filarial infection, followed
by appropriate treatment can prevent sufferings due to the irreversible chronic
elephantiasis. Signs of chronics filariasis are easily recognized, especially among
individuals living in the endemic areas. However detection of adenolymphangitis
(filarial fever) or TPE among suspected patients may not be as easy. Diagnosis is not
only important to the infected individuals but is also a useful screening tool for the
mapping of the endemic areas which is the first important step towards realizing the
success of the lymphatic filariasis elimination program (WHO, 1997; 2002a,b).
1.3.2.1 Direct physical examination and survey
Information for the distribution of infection and disease can be assessed by rapid
assessment through questionnaire among local residents. In areas where
questionnaires cannot be administered or patients are not knowledgeable about the
disease prevalence, physical examinations can be used as a rapid assessment tool
(WHO, 2000) to look for lymphoedema of the limbs or hydrocele in males.
1.3.2.2 Thick blood film (TBF)
The accurate diagnosis of active infection can be made essentially only by detecting mf
in the blood of infected individuals (Weil, 1987). The simplest method is a thick blood
film of capillary blood stained with Giemsa stain (Khamboonruang et al., 1987). For
epidemiological screening, 60 µl of finger-prick blood can be dried on a slide, stained
and examined under a microscope. Disadvantages of thick blood films include the
need to collect blood at night since the mf in peripheral blood peak between 10 p.m.
and 2 a.m. The sensitivity of this method is relatively low, leading to misdiagnoses of
people with low-density infection, amicrofilaraemic stage of infection and single sex

10
infection. Consequently this allows them to progress to irreversible major lymphatic
damages (Braga et al., 2003).
1.3.2.3 Concentration techniques
Use of concentration techniques increases the sensitivity but amicrofilaraemic cases
will still not be detected. An old but still widely used method is that Knott (1935). One
milliliter of blood is added to 9 ml of a 2% formalin solution in water. After red cell lysis
is complete, the mixture is centrifuged and the deposit is examined for mf. The
theoretical detection limit is one mf per ml. The problem is when blood is processed
from individuals with a large amount of plasma gamma globulin. The formalin
precipitates the protein and makes the examination of the deposit difficult. The Knott’s
method has been improved by Melrose et al. (2000) who add a small amount of Triton
100-X to the diluents which dissolves most of the proteingenous deposit and enhances
the visibility of the mf. Another technique is membrane filter technique in which 1-5 ml
of blood which has been diluted in water is passed through a filter fitted with a
polycarbonate membrane which traps the mf. The membrane is removed and the mf is
stained and counted (Moulia-Pelat et al., 1992; McMahon et al., 1979).
1.3.2.4 Ultrasonography
High-frequency ultrasonography can directly visualize adult worms in the dilated
lymphatics. It has being used to detect adult worms in the scrotum and breast (Amaral
et al., 1994; Dreyer et al., 1996a,b; 1999c) and has detected viable worms in children
(Dreyer et al., 1999a).

11
1.3.2.5 Polymerase chain reaction
PCR methods have been successfully used for the detection of parasite-specific DNA
of W. bancrofti in blood, plasma, paraffin-embedded tissues sections (McCarthy et al.,
1996) and sputum (Abbasi et al., 1996, 1999). PCR assays offer rapidity, higher
sensitivity and specificity over conventional dissection and microscopic technique but
they are relatively expensive (Zhong et al., 1996). This is mainly due to the
involvement of costly chemicals and lengthy procedures of DNA extractions and
facilities not often available in filarial-endemic areas. A highly sensitive and specific
PCR assay, based on Ssp I repeats sequence, has been developed for detecting W.
bancrofti in human blood and vectors (Zhong et al., 1996). This Ssp I PCR assay was
found to be highly species specific, as it did not detect the DNA of a closely related
filarial parasite, B. malayi and also proved to be highly sensitive as it did detect as little
as 0.04 pg of W. bancrofti DNA (Hoti et al., 2001). Therefore, it has potential
application in rapid assessment of transmission of filariasis.
1.3.2.6 Detection of circulating antigen
Two different groups of inventors independently produced monoclonal antibodies,
Og4C3 and AD12, which recognized a protein moiety of a major phosphocholine-
containing circulating antigen of W. bancrofti (More and Copeman, 1990; Weil, 1987).
These antigen detection assays were found to be more convenient than the
microfilarial detection technique since night blood sampling could be avoided as
circulating antigens were present even in daytime blood samples (Pani et al., 2000)
collected on filter paper (Itoh et al., 1998).
The murine monoclonal antibody, Og4C3 directed against antigen of Onchocerca
gibsoni was used successfully as the detection-antibody in a sandwich ELISA for the
detection of circulating antigen of W. bancrofti. However, this assay was not effective

12
in detecting antigens of B. malayi, B. timori, O. volvulus or Loa loa (More and
Copeman, 1990).
Another commercially available antigen detection assay used the AD12 monoclonal
antibody in a rapid-format card test for the detection of bancroftian filariasis. The test is
a rapid ICT technique using specific monoclonal and polyclonal antibodies which
recognized the filarial antigen in the blood of infected humans. This kit (ICT Filariasis)
that was initially manufactured by ICT Diagnostics, Balgowlah, New South Wales,
Australia is currently marketed by Binax NowRICT, Portland, USA. It utilizes capillary or
venous blood collected either the night or day and is very easy to handle, very fast to
perform, can be used in the field by people with a minimum amount of training (Weil et
al., 1997). It has previously been shown to be highly sensitive for infections with W.
bancrofti and highly specific with respect to other filarial parasites including O. volvulus,
B. malayi, L. loa and Mansonella Streptocerca (Weil et al., 1997). Its reported
sensitivity and specificity rates were 96-100% and 100% respectively. Thus, this test is
useful for mapping of endemic areas in control programs for bancroftian filariasis.
1.3.2.7 Detection of anti filarial IgG4 antibody
New and more sensitive and specific assays for diagnosis of LF have been developed
(Dissanayake et al., 1994; Chandrashekar et al., 1994). One study on bancroftian
filariasis reported that IgG4 antibodies reacted well with recombinant W. bancrofti SXP-
1 antigen (Engelbrecht et al., 2003). A sensitivity of 100% was obtained in patients with
patent W. bancrofti infections using the Wb-SXP-1 antigen in IgG4-ELISA (Rao et al.,
2000). This recombinant antigen has now being developed into a rapid test for the
identification of total IgG antibodies to Wb-SXP-1. The test is a flow-through
immunofiltration test that employed colloidal gold-protein A as the antibody capture
reagent (Baskar et al., 2004). Another two recombinant-antigens, namely Bm14 and
BmR1, have been developed into IgG4 antibody detection tests and have been shown

13
to be sensitive and specific for determining LF infection/exposure. The Bm14 antigen
was reported to be equally sensitive for Wuchereria and Brugia infection/exposure
(Ramzy et al., 1995; Weil et al., 1999). This antigen has some cross-reactivity with sera
from patients with other filarial infections (loiasis and onchocerciasis), but not with sera
from people with non-filarial nematode infections (Ramzy et al, 1995). Field studies in
Egypt showed that prevalence rates of antibody to Bm14 prior to initiation of MDA were
much higher than antigen or mf prevalence rates in young children (Weil et al, 1999).
The BmR1 antigen which performed well in the detection of brugia infections, has
limited sensitivity in detecting W. bancrofti infection (Lammie et al, 2004).
1.3.3 Treatment and prognosis
Individuals found to be mf-positive or filarial antigen-positive during the initial
assessment period, monitoring, or on voluntary examination should be treated with
diethylcarbamazine (DEC). DEC has been used to treat filariasis since 1947 (Santiago-
Stevenson et al., 1947) and still is the most widely used anti-filarial. Many countries
use a 12-day course (W. bancrofti) or a 6-day course (B. malayi) of 6 mg/kg/day of
DEC (WHO, 1992a,b). In some of the control programs in the Pacific and Papua New
Guinea, colour-coded DEC tablets are used without weighing the patient. A 300mg
tablet is given to adults and a 150mg tablet to children. DEC is very effective in killing
microfilaria but only partly effective against adult filarial parasites. This is evidenced by
the work by Weil et al. (1988) who demonstrated that filarial antigenaemia persists for
up to 12 months after DEC therapy. Figueredo-Silva et al. (1996) removed nodules
after DEC treatment and found that all contained degenerating adult worms, thus
proving that DEC has a limited amount of macrofilaricidal activity.
A study showed that single dose albendazole (400mg) has similar efficacy in the
clearance of mf as that of DEC (6 mg/kg) or the co-administration of the two drugs, i.e.

14
albendazole (400mg) plus DEC (6 mg/kg) (Pani et al., 2002). This showed that
albendazole can be used in mass single dose administration for the control of LF.
In areas co-endemic with onchocerciasis or loiasis, ivermectin has been used widely
as microfilaricidal in LF cases to avoid the potentially severe allergic reaction with
DEC. Ivermectin is a highly effective and generally well-tolerated drug for the treatment
of LF (Brown et al., 2000) and a series of single dose ranging studies from 20 to
200µg/kg for the treatment of bancroftian filariasis was effective in decreasing blood
microfilaria density (Cartel et al., 1990a,b,c,d; Kumaraswami et al., 1988). Ivermectin
with higher doses resulting in more sustained clearance of mf (Kar et al., 1993a, b;
Kumaraswami et al., 1988). It is a very effective microfilaricide but how effective it is
against adult worm is a contentious issue.
Simple hygiene measures, supplemented with antibiotics or antibacterial cream helps
prevent damage tissues from worsening, stop secondary bacterial infections and help
to reduce the limb enlargement caused by repeated filarial and bacterial infections
(Ottesen, 1997). Effective hygiene measures include regular twice-daily washing of the
affected parts with soap and water; raising the affected limb at night; keeping the nails
clean; wearing shoes; and using local antiseptics or antibiotic creams to treat small
wounds.
Anti filarial drugs alone will not be able to revert the fibrotic changes of the skin and
connective tissue involved (Kazura, 1999). Now expertise is available for surgical
therapy of genital manifestations of filariasis. Generally, the surgery involves removal
of the excess fibrotic tissue. The most common surgery for hydrocele is complete
excision of the sac. However, its effect is short lasting as the edema will recur. As an
alternative, micro vascular surgery can be performed (Kazura, 1999) which involves
anastomosis of the lymph vessels with the nearby veins (WHO, 2002a,b).

15
1.3.4 Global Programme to Eliminate Lymphatic Filariasis (GPELF)
The goal of the Global Programme to Eliminate Lymphatic Filariasis (GPELF) is
defined as “The Elimination of Lymphatic Filariasis as a Public Health Problem by
2020” (WHO, 1997). The 50th World Health Assembly in 1997 had passed a resolution
identifying the elimination of LF as a public health problem, a priority. This includes
some strategic plans which are identified to have four major elements (WHO, 1999a,b)
namely the interruption of transmission; the prevention of disability; the provision of
additional technical support; and the implementation of operational research (up-
scaling program). Interruption in transmission can be achieved in reducing and
eliminating the reservoir of mf through treating the human population and by reducing
contact between humans and mosquito vectors (Chaubal et al., 2003).
The traditional method i.e. thick blood smear examination which was used before, have
a sensitivity of only 25-40% (WHO, 1998). If it is the sole diagnostic method used in
post intervention surveillance, many false negatives will occur; hence a low incidence
will not actually indicate successful interruption of transmission. Therefore, there is a
clear need for specific, sensitive and field applicable tests as to ensure the success of
this global elimination program.
Mass drug administration (MDA) of at-risk population (Freeman et al., 2001) is being
used to interrupt transmission. This is based on the evidence of the effectiveness of a
single dose of DEC (6mg/kg) in the clearance of mf and sustaining this over a period of
at least one year (Farid et al., 2003). Thus this reduces the number of mf in the blood
to levels below which the mosquito vectors can no longer transmit infection. These
comprise two approaches. First is the once-yearly treatment with single dose of two
drugs given together i.e. albendazole plus either ivermectin or DEC for 4-6 years

16
(Molyneux, 2003). The second approach is the use of DEC-fortified table or cooking
salt for 1-2 years (Lymphatic Filariasis Support Center, 2004).
GlaxoSmithKline has committed to provide the entire supply of albendazole, while
Merck & Co. Inc., committed to supply ivermectin in those African countries with
onchocerciasis and loaisis (WHO, 1997). By the end of 2001, nearly 26 million people
in over 22 countries had been administered the combination drugs in MDA campaigns
(WHO, 2002a,b). Vector control has been used as an important adjunct in the LF
program in certain areas. The strategy to prevent disability is designed to encourage
home-based self-care, i.e. regular skin care, exercise and appropriate footwear, and by
trying to develop facilities in the health care system for disability control (WHO, 1997).
The control of lymphoedema management is the prevention of acute ADL attacks
through basic hygiene by using soap and water, and prevention and treatment of small
skin lesions with application of topical antifungal or antibiotic cream (WHO, 2000). This
basic hygiene can stop the acute attacks and improve the patient’s condition.
1.3.5 Lymphatic Filariasis in the Malaysian Context
The Filariasis Control Program in Malaysia was established more than 30 years ago;
however the disease is still a public health problem in rural areas, especially in some
areas in Sarawak, Pahang, Terengganu, Johor, Perak, Sabah, Kedah and Kelantan.
More than 85% of the annual incidence rate of lymphatic filariasis in Malaysia is due to
B. malayi (Ministry of Health, 1990-1999). The annual incidence of chronic
elephantiasis is around 5 to 10, and about 2.9 million people are at risk of acquiring
this disfiguring disease (Che Abdullah, 2004). In early 1960, the Vector Borne Disease
Control Unit (RKPBV) of Ministry of Health initiated filariasis surveillance to detect and
treat every filariasis cases. Later, three approaches were implemented in Malaysia,
namely, mass treatment and treatment of index cases; eradication and control of the

17
vectors; and avoiding human-mosquito contact (Noorhayati, 1999). In June 2003,
Malaysia started the National Program for Elimination of Lymphatic Filariasis and the
aim is to eliminate lymphatic filariasis as a public health problem by 2013 (Che
Abdullah, 2004).
The mosquito vectors belonging to the Anopheles and Mansonia genera are involved
in the transmission of filariasis in Malaysia, the latter being the more important vector.
Anopheles donaldi was found to be infected with infective larvae of B. malayi
(Vythilingam et al., 1996) where the peak biting time was around 11 pm to 12 am.
Monkeys and domestic cats are the reservoir hosts for the subperiodic strain of B.
malayi (Marzhuki et al., 1993). The most common monkeys in Malaysia are the
macaques (Macaca spp.) and leaf monkeys (Presbytis spp.) [WHO, 1984].
The historical record of filariasis in Malaysia can be categorized under three phases
(Lim, 2005). The first phase (1908-1952) mainly focused on the statistics of microfilarial
carriers which had been identified in Hospital Kuala Lumpur and Raub. The second
phase (1953-1961) dealt with the mapping of endemic foci in East Pahang,
Terengganu and Kelantan. At this phase it was found that Culex quinquefasciatus was
implicated for spreading bancroftian filariasis. Successful mass chemotherapheutic
trials were conducted in two villages of Pahang i.e. treatment regime of weekly and
monthly doses of diethylcarbamazine (DEC) at 4-6 mg/kg body weight for 6 weeks and
6 months respectively. The activities of the third phase (1962-onwards) was mainly
towards identifying the most suitable method for filariasis control.
In Malaysia mf densities in many infected individuals are too low to be detected by the
traditional method of thick blood smear examination. In addition, thick blood smears do

18
not allow for the detection of individuals harbouring nonfertile worms, pre-patent
infections and single sex infections (Turner et al., 1992).
In the official report of the district of Pasir Mas in the state of Kelantan, one case in
2428 samples (0.04%) from the subdistrict of Gual Periok was detected in 1997; and in
2000, two cases were detected in 803 samples (0.25%) from the subdistricts of Gual
Periok and Rantau Panjang (Pasir Mas Health Office 1997–2001). However, no cases
were reported in 1998 (1385 samples), 1999 (985 samples) and 2001 (1100 samples).
These figures probably do not reflect the true prevalence of the infection as it is based
on the insensitive thick blood smear examination. Recently, Rahmah et al (2003a)
conducted a filariasis survey on 5138 pupils schools located in the subdistricts (mukim)
of Pasir Mas bordering Thailand border. Out of 2439 boys and 2699 girls screened.
Brugia malayi infection was detected in 18 children, giving an overall prevalence of
0.35% (18 of 5138). The investigators employed a recombinant antigen (BmR1) based
ELISA (Brugia-Elisa) that has been shown to highly specific and sensitive for detection
of brugian filariasis (Supali et al., 2004; Rahmah et al., 2003b). In 2001, Lim et al had
collected a total of 1,134 finger-pricked blood samples from residents of Setiu,
Terengganu and the findings showed that 0.26% (3/1,134) were positive by thick blood
smear examination, while 2.47% (28/1,134) were positive using Brugia-Elisa. In
another study conducted by Jamail et al (2005) among residents of seven endemic
districts in the state of Sarawak, the overall prevalence of brugian filariasis as
determined by a rapid test was 9.4% while that determined by microscopy was 0.90%
thus the dipstick detected about 10 times more cases than microscopy. The test
employed was BRUGIArapid™ dipstick test, which is also based on the BmR1
recombinant antigen. Equal percentages of adults and children were found to be
positive by the dipstick whereas microscopy showed that the number of infected
children was seven times less than infected adults. Thus the results of the above
studies showed that the use of insensitive microscopic examination leads to many

19
untreated cases of infected people, which will become reservoir for the transmission of
the infection. Due to its superior sensitivity, BRUGIArapid™ is being employed by the
Ministry of Health Malaysia to assist in the National Lymphatic Filariasis Elimination
Program.
1.4 Onchocerciasis
Onchocerciasis or river blindness is a major public-health and socio-economic problem
in many rural areas (Shah et al., 1987). It is caused by the filarial worm Onchocerca
volvulus, vectored by the black fly, Simulium spp. (Cox, 2002; Greene, 1992) including
S. damnosum (Shah et al., 1987). It infects about 20 million people and is endemic in
28 countries in Africa, 6 countries in the Americas and in Yemen (WHO, 2002a,b).
Onchocerciasis is one of the leading causes of infectious blindness worldwide (Klotz et
al., 2000; Duke, 1990). It has caused visual impairment in 500,000 and blindness in
270,000 people, rendering onchocerciasis the second most frequent cause of
preventable blindness in sub-Saharan Africa (WHO, 1995). Rarely life-threatening, the
disease cause chronic suffering and severe disability (Figure 1.5).
The most important signs are blindness and chronic skin disease such as scaly, itchy
and unusual nodular skin (Cox, 2002; Pogonka et al., 1997). Onchocerciasis used to
be the major cause of blindness throughout sub-Saharan Africa, often affecting more
than 50% of the inhabitants of towns and villages in endemic areas (Greene, 1992). In
some small communities in Africa and Central America, most of the people of middle
age and over are blind.

20
(a) (b) (c)
Figure 1.4 Two signs of onchocerciasis; (a) and (b) a 60-year-old male farmer
scratching his legs which show the tell-tale depigmentation (leopard
skin), the consequence of years of fenzied scratching. Having the
incessant itching chronic skin disease, and (c) close up picture of an
eye damaged (river blindness) as a result of infection with O. volvulus
(TDR Image Library, 2005).

21
1.4.1 Onchocerca volvulus
Infection with O. volvulus is initiated by the feeding of an infected black fly on a human.
There are no reservoir hosts for this parasite, but some experimental infections have
been established in non-human models such as cattle (Yien-Ming and Bianco, 1995),
chimpanzees and mice (Abraham et al., 2001). It is a member of class of Secernentea,
subclass of Spiruria, order Spirurida, belonged to the superfamily of Filarioidea and a
family member of Onchocercidae.
The morphology is similar to that of W. bancrofti. They are slender and blunt at both
ends. Lips and a buccal capsule are absent, and 2 circles of four papillae each
surround the mouth. Males are 19 to 42 cm long by 130 to 210 µm wide and the
females are 33.5 to 50 cm long by 270 to 400 µm wide, while the mf are 300mm in
length and 0.8mm in diameter, unsheathed with sharply pointed, curved tails.
1.4.2 Life cycle of Onchocerca volvulus
Human onchocerciasis is caused by the filarial parasite O. volvulus whose life cycle
occurs in two different hosts namely Simulium black flies and humans (Figure 1.6).
Simulium vectors breed in fast flowing rivers. A female worm may produce 1000 mf per
day which is shed in the tissues and blood of their human host (Klotz et al., 2000). The
life cycle begins when a parasitized female black fly takes a blood-meal. The host’s
skin is stretched by the fly’s apical teeth and sliced by its mandibles. The larvae from
the fly then move to the subcutaneous tissues (molt to L3, the infective stage for
human) where they migrate, form and lodge in nodules, and slowly mature into adult
worms (L4) in the subcutaneous tissue for years (Klotz et al., 2000). The adult worm
locates to a single niche in the subcutaneous tissue. New worms form new nodules or
find existing nodules and cluster together. After mating, eggs from inside the female
worm develop into mf and leave the worm one by one. The mf migrate throughout this
tissue, inducing injury to a variety of anatomical sites contiguous with that tissue or

22
where they die after several years (Klotz et al., 2000). When female black flies take a
blood meal they ingest the mf that will then undergo transitions to L2 stage in the fly.
1.4.3 Pathology and clinical disease
Unlike other filarial infections, the problems of onchocerciasis are caused by mf rather
than adult worms. An early sign of infection with Onchocerca is the raised nodules that
can be seen under the skin. These are most often seen in areas over a bony
prominence and may develop into a firm, non-tender nodule called as onchocercomata
which contained adult worms (Klotz et al., 2000). It has been suggested that this
phenomenon occurs because the larvae are immobilized in these locations long
enough for them to be trapped by the body’s cellular defense mechanisms. This
migration lead to intense pruritus manifesting as dermatitis, whereby the skin may
become thickened, edematous, wrinkled and depigmented (Klotz et al., 2000).
The mf can be found free in the fluid within the nodules and in the dermal layers of the
skin, spreading away from the nodules containing the adults. Mf also can be found in
the blood and eye during heavy infection (Klotz et al., 2000). Mf can be killed only
during a limited period of their development, after which the larvae become resistant to
attack by the immune response (Abraham et al., 2001). Reactions to dead mf around
these nodules can lead to several unpleasant conditions, including serious visual
impairment and blindness, skin rashes, lesions, itching and depigmentation of the skin,
lymphadenitis (lead to hanging groin) and general debilitation (Klotz et al., 2000). In the
skin, there is destruction of the elastic tissues and the formation of redundant folds.
Dead mf in the eye leads to an inflammatory immune response and the eventual
formation of secondary cataracts and ocular lesions. Because of this, heavy infections
often lead to progressive blindness. Much of the pathology associated with

23
onchocerciasis takes place during the immune responses against the mf that are found
in the skin and ocular tissues (Kazura et al., 1993).
The establishment of a chronic filarial infection in humans is accompanied by
characteristic cell-mediated and humoral immune responses. The antifilarial humoral
immune response is characterized by high levels of immunoglobulin E (IgE)
(Kurniawan et al., 1993), eosinophilia (Gbakima et al., 1996). In areas of endemicity,
most filarial infections are initiated during early childhood (Gbakima, 1996). The
precise pathogenesis of onchocerciasis lesions is still unknown.

24
Figure 1.5 The incubation phase of filarial parasite which affective from the point of
infection to the time of appearance of first mf and resulted in eye
lesions, altered pigmentation and loss of elasticity of skin (Cross, 1992).