characterization of newcastle disease viruses in wild 2...
TRANSCRIPT

1
Characterization of Newcastle disease viruses in wild 1
and domestic birds in Luxembourg from 2006 to 2008 2
Running title: Characterization of NDV in Luxembourg 3
4
Chantal J. Snoeck1, Marianna Marinelli1*, Emilie Charpentier1, Aurélie Sausy1, Tom 5
Conzemius2, Serge Losch3, Claude P. Muller1# 6
7
1Institute of Immunology, Centre de Recherche Public - Santé /Laboratoire National de 8
Santé, Luxembourg, Luxembourg 9
2Lëtzebuerger Natur- a Vulleschutzliga, Kockelscheuer, Luxembourg 10
3Laboratoire de Médecine Vétérinaire de l’Etat, Administration des Services Vétérinaires, 11
Ministère de l'Agriculture, de la Viticulture et du Développement rural, Luxembourg, 12
Luxembourg 13
14
#Corresponding author: Claude P. Muller, Institute of Immunology, Centre de Recherche 15
Public - Santé /Laboratoire National de Santé, 20A rue Auguste Lumière, L-1950 16
Luxembourg. Tel: +352 49 06 04 220. Fax: +352 49 06 86. E-mail address: 17
19
*Current affiliation: Novartis Vaccines and Diagnostics srl Via Fiorentina 1, 53100 Siena, 20
Italy 21
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.02437-12 AEM Accepts, published online ahead of print on 16 November 2012
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2
Abstract 22
Newcastle disease virus (NDV) is one of the most important viral diseases of birds. Wild 23
birds constitute a natural reservoir of low virulent viruses while poultry are the main 24
reservoir of virulent strains. Exchange of virus between both reservoirs represents a risk 25
for both bird populations. Samples from wild and domestic birds collected between 2006 26
and 2010 in Luxembourg were analyzed for NDV. Three similar avirulent genotype I 27
strains were found in ducks during consecutive years, suggesting that the virus may have 28
survived and spread locally. However, separate introductions cannot be excluded because 29
no recent complete F gene sequences of genotype I from other European countries are 30
available. Detection of vaccine-like strains in wild waterbirds suggested the spread of 31
vaccine strains, despite the non-vaccination policy in Luxembourg. Among domestic 32
birds, only one chicken was positive for a genotype II strain differing from the LaSota 33
vaccine and exhibiting a so far unrecognized fusion protein cleavage site of predicted 34
lowvirulence. Three genotype VI strains from pigeons were the only virulent strains 35
found. The circulation of NDV in wild and free-ranging domestic birds warrants a 36
continuing surveillance because of increased concern that low virulent wild bird viruses 37
could become more virulent in domestic populations. 38
39
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

3
Introduction 40
Newcastle disease, caused by virulent strains of the Newcastle disease virus, is the most 41
important viral disease in poultry together with highly pathogenic avian influenza. 42
Newcastle disease virus (NDV), or avian paramyxovirus type-1, is a negative single-43
stranded RNA virus belonging to the Avulavirus genus in the Paramyxoviridae family. 44
Based on clinical signs in chickens, several pathotypes have been defined (23). The 45
sequence of the fusion (F) protein cleavage site is considered a major virulence 46
determinant and it appears that a minimum of three basic amino acids between residues 47
112 and 116, followed by a phenylalanine at residue 117 are required for virulence (23). 48
However, other factors are also involved in the virulent phenotype as indicated by strains 49
found in pigeons around the world and in healthy migratory ducks in Alaska that have a 50
virulent cleavage site motif but are not always virulent for chicken in standard 51
pathogenicity tests (9, 27). 52
Besides its phenotypic heterogeneity, NDV is also genetically diverse and several 53
phylogenetic classes and genotypes (10) or lineages (2) are recognized and its diversity is 54
still unfolding (10, 26). Class I strains and genotypes I and X of class II are mainly 55
constituted by avirulent strains. Genotype II contains a broad spectrum of strains but 56
nowadays, mainly avirulent strains are found. The other class II genotypes (III to IX and 57
XI to XV) contain almost exclusively viruses with a virulent cleavage site. 58
Wild birds constitute a natural reservoir of viruses of low virulence for chickens. In 59
particular waterbirds may play an important role in NDV epidemiology due to shedding 60
of viral particles into the aquatic environment more favorable for virus stability, and their 61
potential for long distance dissemination by migration (14). Although no direct 62
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

4
epidemiological link was found, successions of outbreaks in the UK, Sweden, Denmark 63
and Finland between 1996 and 2005 were all related at least on the basis of strain 64
similarity, suggesting multiple introductions of viruses from the same pool by wild birds 65
(5). Moreover, isolation of similar viruses in wild birds (a goosander in Finland, a 66
cormorant in Denmark) together with the proximity to water of a significant number of 67
the affected flocks was suggestive of a wild bird reservoir at least of genotype XIII 68
strains in Western and Northern Europe (4). Another more common threat for poultry 69
arises from pigeon paramyxovirus type-1 (PPMV-1) strains, variants of NDV grouping 70
within genotype VI. PPMV-1 strains exhibit a broad range of pathogenicity for poultry 71
(9) and pathogenicity may increase after serial passages in chickens (9, 20). On several 72
occasions, PPMV-1 viruses caused outbreaks in chickens (7, 12, 34). 73
During the last decade, virulent viruses from genotypes VI, VII and XIII have been 74
detected in wild and domestic birds in several European countries (4), and avirulent 75
strains of class I and class II (genotypes I and II) have also been reported sporadically 76
(http://ec.europa.eu/food/animal/diseases/controlmeasures/avian/crls_proceedings_en.ht77
m). However, in Europe, only few sequences from wild birds are available, despite their 78
importance for epidemiological surveillance. In this retrospective study, we examined 79
stored samples collected in the framework of avian influenza surveillance in Luxembourg 80
to investigate the presence of avirulent and virulent NDV in wild and domestic birds. All 81
viruses found had a predicted low virulence, except for three PPMV-1 strains. 82
Material and Methods 83
Sample collection. Between January 2006 and July 2010, pooled tracheal and cloacal 84
swabs (n=576), cloacal swabs (n=196), tracheal swabs (n=22) or fresh feces (n=337) 85
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

5
were collected during active and passive surveillance for avian influenza virus. The 86
majority of the samples originated from wild birds (n= 1003), but a smaller number of 87
domestic birds was also sampled, including chickens (n=120), turkey (n=1), quail (n=1), 88
peafowl (n=1) and pheasants (n=2, Table 2). Samples from healthy passerines were 89
mainly collected with mist nests during migration surveys at three locations: Nospelt, 90
Ubersyren and Schifflange. Wild waterfowl, primarily targeted for avian influenza 91
surveillance, were mainly sampled along the Moselle River in Remich and Wasserbillig. 92
Some samples from injured or sick animals, especially from birds of prey, were also 93
collected at a wildlife shelter, while exotic species (n=3) were sampled at a zoological 94
park and at the international airport. The other bird species were sampled throughout the 95
country but with a bias towards the southern part of the country (Fig. 1). 96
All swab samples were immediately placed in the field in 500 µl of virus transport 97
medium (PBS pH 7.0 with 2000 U penicillin/ml, 200 mg streptomycin/ml, 2000 U 98
polymyxin B/ml, 250 mg gentamicin/ml, 60 mg ofloxacin/ml, 200 mg 99
sulfamethoxazole/ml and 2.5 mg amphotericin B/ml). All samples were kept refrigerated 100
until delivery to the laboratory. Approximately 100 mg of fecal samples were 101
homogenized in 500 μl of virus transport medium upon arrival at the laboratory. 786 102
samples were processed immediately and cDNA was kept at -20°C, while 345 samples 103
were stored at -80°C before being processed. The majority of the samples (86%, 104
968/1131) were stored at -80°C within 48h after collection. 105
Nucleic acid extraction, PCR, and sequencing. RNA was extracted from 140 μl of 106
medium using QIAamp Viral RNA Mini Kit (Qiagen, Venlo, The Netherlands) or from 107
50 μl using MagMAXTM-96 AI/ND Viral RNA Isolation Kit (Life Technologies, 108
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

6
Merelbeke, Belgium) with KingFisher (Thermo Fisher, Waltham, Massachusetts, USA). 109
Screening for class I strains was only performed on the 345 samples collected after June 110
2007 (according to Kim et al. (19)). Class II strain detection by nested PCR (16, 26) and 111
sequencing of positive PCR products (26) were carried out as described previously. 112
Whenever enough material was available, the entire F gene was amplified using several 113
primer combinations in (semi-)nested PCR formats (Table 1). 114
Statistical analyses. Statistical analyses to assess whether differences in sampling or 115
sample processing had an effect on the outcome of the detection tests were performed 116
using the Chi-square test with Yates correction in SigmaPlot software. 117
Sequence analyses. Kimura distances were calculated according to the Kimura 2-118
parameter model on partial (240 nt) or complete (1662 nt) F gene sequences. The 119
sequence lengths used for distance calculations are mentioned between brackets in the 120
Results section. Phylogenetic relationships were inferred by comparing the 121
Luxembourgish strains with all NDV sequences available on NCBI (downloaded in 122
October 2012) after dataset curation. Datasets were aligned using ClustalW (30). Trees 123
were calculated with the Neighbour-Joining method, using the Kimura 2-parameter 124
model and 1000 bootstrap replicates as implemented in MEGA v5.03 software (28). 125
Representative strains were selected based on these preliminary analyses and are 126
displayed in Fig. 2 and 3. The classification nomenclature was used according to Diel et 127
al. (10) and the nucleotide numbering of F gene sequence according to Kho et al. (16). 128
Nucleotide sequence accession numbers. Sequences were submitted to GenBank under 129
accession numbers HE972209 to HE972217. The following strain nomenclature was 130
used: host/country/strain number/year. 131
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

7
Results 132
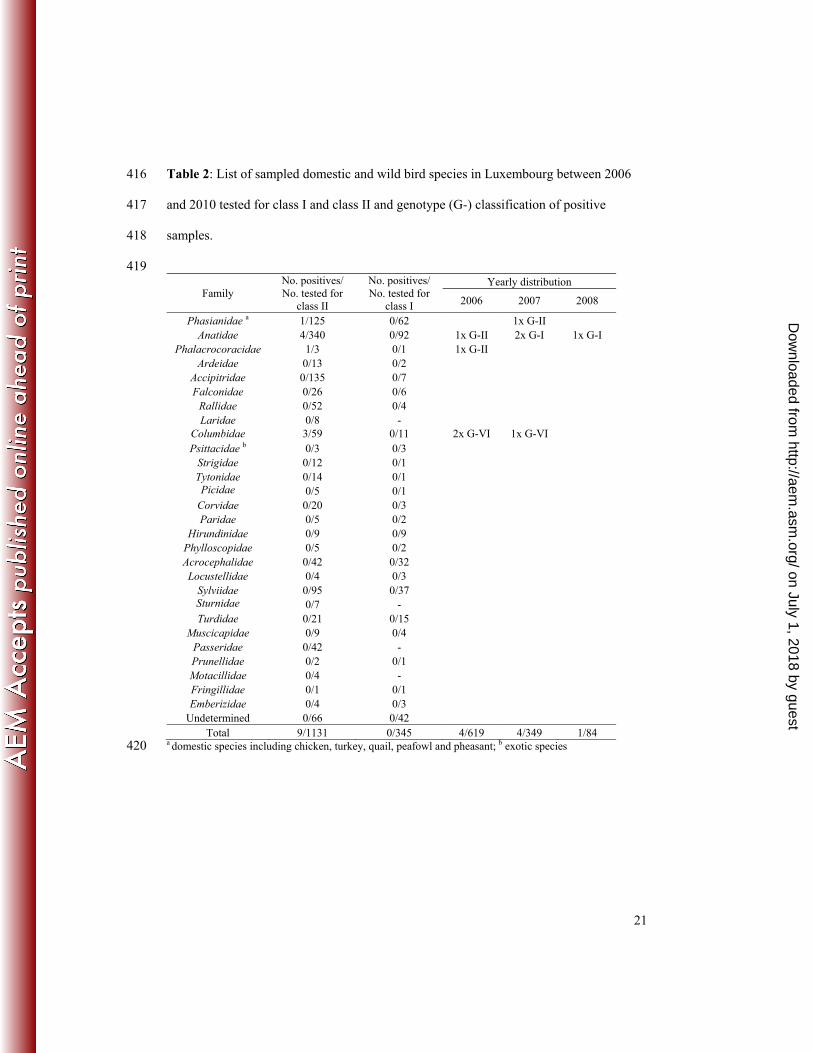
None of the 345 samples tested for class I strains was positive for these strains. A total of 133
9 samples out of 1131 were positive in the PCR detecting class II NDV strains (Table 2), 134
corresponding to an overall prevalence of 0.8% during the 2006-2010 period (4/619 in 135
2006, 4/349 in 2007, 1/84 in 2008, 0/67 in 2009 and 0/12 in 2010). However, prevalences 136
may be somewhat underestimated since virus isolation and some RT-PCR protocols may 137
be more sensitive. Statistical analyses revealed that the type of samples, the time between 138
collection and storage (categorized as less or more than 48h) or the type of material used 139
in the PCR (cDNA stored at -20°C or freshly prepared from original samples stored at -140
80°C) had no significant effect on the number of positive samples per group (p-141
values=0.816, 0.847, 0.583 respectively). The apparent absence of NDV in 2009-2010 as 142
well as in the northern part of the country (Fig. 1) was most probably due to a suboptimal 143
surveillance effort rather than a disappearance of NDV in Luxembourg or the existence 144
of regional differences. 145
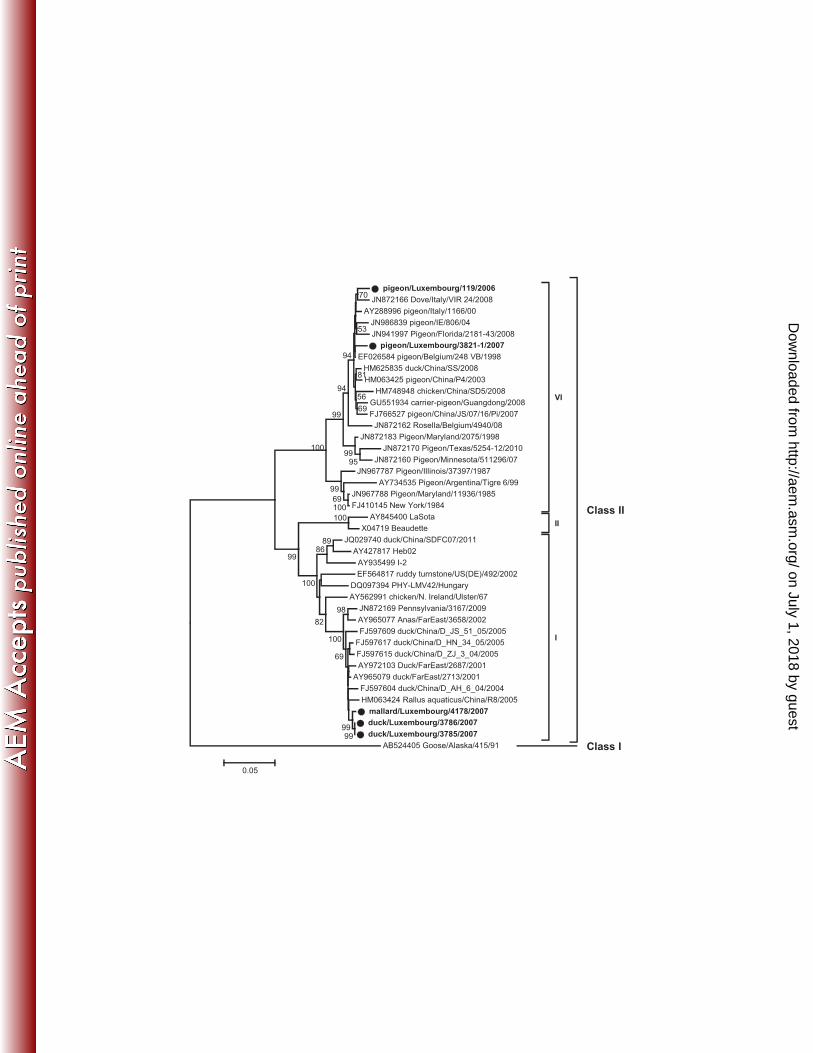
Phylogenetic analyses of partial (9 strains; Fig. 2) and complete (5 strains; Fig. 3) F gene 146
sequences revealed an equal distribution of the samples in three class II genotypes. 147
Genotype I. Three similar strains (Kimura distances from 0 to 0.5%, 1662 nt) from 148
ducks, including one Mallard Anas platyrhynchos, clustered in genotype I. They clustered 149
together with strains from waterfowls from Finland, the Far East and China (Fig. 2 and 3) 150
and were most closely related to each other based on complete F gene sequences (Fig. 3). 151
They all had a Kimura distance of 0.4% (240 nt) to mallard/Finland/9360/2010. The 152
deduced amino acid sequence of the F protein cleavage site 112GKQGR*L117 was typical 153
of avirulent genotype I strains. 154
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

8
Genotype II. Three samples from a chicken, a Great Cormorant Phalocrocorax carbo 155
and a duck clustered in genotype II, together with the commonly used vaccines LaSota 156
and B1. Kimura distances (240 nt) to the LaSota and B1 vaccines ranged from 0 157
(duck/Luxembourg/26/06) to 1.3% (chicken/Luxembourg/2871-18/07). Both 158
duck/Luxembourg/26/06 and Great Cormorant/Luxembourg/2547/2006 exhibited a 159
cleavage site typical of avirulent genotype II strains, 112GRQGR*L117, while the 160
chicken/Luxembourg/2871-18/07 strain encoded 112GGQGR*L117 due to a non-161
synonymous A to G substitution at nucleotide position 380. 162
Genotype VI. The last three sequences from pigeons Columba livia var. domestica 163
clustered in genotype VI, together with recent isolates mainly found in Columbiformes 164
worldwide. Kimura distances ranged from 0.4% between pigeon/Luxembourg/2657-2/06 165
and pigeon/Luxembourg/119/06 (240 nt) to 2.6% between pigeon/Luxembourg/119/06 166
and pigeon/Luxembourg/3821-1/07 (1662 nt). All genotype VI strains encoded for 167
virulent fusion cleavage site motifs 112RRQKR*F117, as defined by World Organization 168
for Animal Health standard (23), and were similar to those of other previously described 169
PPMV-1 strains. 170
Discussion 171
There is increasing evidence that wild waterbirds are natural carriers of avirulent class I 172
and class II genotype I and X strains (13, 18, 22, 24, 36). It was thus not surprising to find 173
five out of nine positive samples in waterbirds, including three avirulent genotype I 174
strains in ducks in our study. Based on the full F gene sequences (Fig. 3), these genotype 175
I strains formed a monophyletic cluster, which may suggest that they evolved from a 176
recent common ancestor and resulted from a single introduction event in Luxembourg. In 177
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

9
this scenario, the detection of similar strains in 2007 and 2008 would indicate that 178
avirulent viruses could be maintained in the local bird population throughout the year. 179
However, their relationship with NDV isolates recently identified in migratory species in 180
Finland (21, 22) could not be further clarified on the basis of the short Finnish sequences. 181
Wild Anatidae, such as Eurasian Teals, Mallards and Northern Shoveler, have different 182
migratory routes, which mainly depend on weather conditions and the availability of food 183
resources. These three species, among others, are also commonly observed at migratory 184
stopovers in Luxembourg. Similarly, the three ducks sampled in August 2007 and in 185
October 2008 in Luxembourg could have arrived in Luxembourg shortly before being 186
sampled. It is therefore possible that genotype I strains have been introduced by 187
migratory species on a single or several occasions. Complete F gene sequences from 188
other European countries, as well as more detailed information on the bird species 189
sampled, would be required to further address this question. 190
The strains of genotype VI found in three Luxembourg pigeons probably originate from 191
separate introduction events, as they do not share a direct common ancestor and are 192
interspersed with strains found in other countries (Fig. 2 and 3). While PPMV-1 strains 193
initially circulated mainly in racing and show pigeons, they are now considered enzootic 194
in feral pigeons and doves in countries such as Germany and Italy (29, 34). Cases in 195
pigeons have been detected almost every year between 2000 and 2009 in the 196
neighbouring countries (4) and dissemination to Luxembourg is not unexpected. 197
While genotypes I and X (class II) and class I strains are often detected in waterfowl, 198
vaccine-like strains of genotype II are mainly found in poultry and are usually associated 199
with the use of live vaccines (26, 35, 37). In this respect, our finding of a vaccine-like 200
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

10
strain in a Great Cormorant and a duck in Luxembourg is somewhat unusual. The 201
detection of vaccine strains in non-vaccinated flocks suggested that lentogenic vaccine 202
strains can spread at least within poultry (33). In addition, wild type virus transmission 203
between wild and domestic birds were already suspected to be at the origin of the 204
similarity of strains found in wild birds and domestic birds in live bird markets (17, 18) 205
or in flocks with possible contacts with wild birds (14), and of the spill-back of virulent 206
strains into wild birds (31, 37). Therefore it seems reasonable to expect that vaccine 207
strains may also be exchanged between poultry and wild birds. Similar cases of strains 208
close to LaSota or B1 vaccines in wild birds have been reported elsewhere, including 209
Asia (China, India and Malaysia), Argentina or France (Fig. 2). Although NDV 210
vaccination is not allowed in Luxembourg, vaccinated animals are also sometimes 211
imported (Losch S., pers comm.). Also bridge species such as sparrows that live in close 212
proximity with domestic birds (37), in particular in regions where backyard chickens are 213
commonly reared, or food/water contamination (3, 7) may have contributed to the 214
transmission of vaccine-like strains to waterbirds in Luxembourg. On the other hand, 215
wild birds may have been infected in other European countries which sometimes allow 216
vaccination. Since Great Cormorants tend to migrate late during the season, it is difficult 217
to known whether the bird infected with a genotype II strain sampled in April spent the 218
winter locally, or was sampled during its northwards migration. Unfortunately, the 219
species of the other bird infected with a genotype II strain was not known. 220
The only strain found in a domestic bird belonged to genotype II, but did not seem to be 221
directly related to a vaccine strain because of three mutations in a short region of the F 222
gene leading to two amino acid substitutions, one being in the cleavage site. To our 223
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

11
knowledge, this particular cleavage site sequence has not been reported before, but this 224
strain is probably not virulent for chickens as it contains only one basic amino acid 225
between residues 112 and 116 and a leucine at residue 117. Unfortunately no further 226
information about potential clinical signs in the flock was available. 227
No virulent NDV strains were found in wild or domestic birds in Luxembourg, except for 228
the three PPMV-1 strains. Although the latter are normally found in pigeons, PPMV-1 229
transmission to poultry was reported on a few occasions in Europe during the past decade 230
(4). Even if most cases occurred in small backyard flocks with low biosecurity, the 231
circulation of PPMV-1 in pigeons represents a potential threat for the poultry industry. 232
The presence of avirulent strains in wild birds may also be a risk for poultry. Although 233
excessively rare in the field, virulent strains may develop from low virulent strains after 234
mutation, as was postulated for outbreaks that occurred in Ireland (6) and Australia (11). 235
This was also demonstrated by serial experimental passages in chickens (25), which may 236
have given minor populations of virulent NDV in field isolates a selective advantage 237
(15). All these scenarios highlight the importance of virological surveillance and 238
preventive measures to reduce the intermingling of wild and domestic birds. 239
In conclusion, we found avirulent genotype I strains in waterfowls in Luxembourg 240
similar to those circulating in wild migratory birds in Finland, suggesting that these 241
viruses represent typical avirulent strains found in European wild birds and that migratory 242
birds may contribute to their spread. Detection of vaccine-like strains in wild waterbirds 243
suggests the spread of vaccine strains, despite the non-vaccination policy in Luxembourg. 244
Although the three PPMV-1 strains from pigeons were the only virulent strains found in 245
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

12
Luxembourg, the presence of NDV in wild and free-ranging domestic birds justifies the 246
need for continuous surveillance in wild and domestic birds. 247
Acknowledgments 248
The authors wish to thank Dr. A. Reye and F. Leenen for technical assistance using 249
ArcGIS and SigmaPlot software, as well as S. Farinelle for sharing her ornithological 250
knowledge. They gratefully acknowledge the Administration des Services Vétérinaries 251
and the Lëtzebuerger Natur- a Vulleschutzliga for their expertise in sample collection. 252
The authors wish to thank Dr. Claudio Afonso, Southeast Poultry Research Laboratory, 253
USDA, for providing positive controls. 254
The authors acknowledge the Ministry of Health, the Ministry of Research and the Centre 255
de Recherche Public-Santé for their generous financial and moral support. C.J. Snoeck 256
was supported by an AFR fellowship TR_PHD BFR08-095 from the Fonds National de 257
la Recherche, Luxembourg. 258
References 259
1. Abolnik C, Horner RF, Bisschop SP, Parker ME, Romito M, Viljoen GJ. 260
2004. A phylogenetic study of South African Newcastle disease virus strains 261
isolated between 1990 and 2002 suggests epidemiological origins in the Far East. 262
Arch Virol 149:603-619. 263
2. Aldous EW, Mynn JK, Banks J, Alexander DJ. 2003. A molecular 264
epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) 265
isolates by phylogenetic analysis of a partial nucleotide sequence of the fusion 266
protein gene. Avian Pathol 32:239-256. 267
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

13
3. Alexander DJ. 1995. The epidemiology and control of avian influenza and 268
Newcastle disease. J Comp Pathol 112:105-126. 269
4. Alexander DJ. 2011. Newcastle disease in the European Union 2000 to 2009. 270
Avian Pathol 40:547-558. 271
5. Alexander DJ, Banks J, Collins MS, Manvell RJ, Frost KM, Speidel EC, 272
Aldous EW. 1999. Antigenic and genetic characterisation of Newcastle disease 273
viruses isolated from outbreaks in domestic fowl and turkeys in Great Britain 274
during 1997. Vet Rec 145:417-421. 275
6. Alexander DJ, Campbell G, Manvell RJ, Collins MS, Parsons G, McNulty 276
MS. 1992. Characterisation of an antigenically unusual virus responsible for two 277
outbreaks of Newcastle disease in the Republic of Ireland in 1990. Vet Rec 278
130:65-68. 279
7. Alexander DJ, Parsons G, Marshall R. 1984. Infection of fowls with Newcastle 280
disease virus by food contaminated with pigeon faeces. Vet Rec 115:601-602. 281
8. Cattoli G, Fusaro A, Monne I, Molia S, Le Menach A, Maregeya B, Nchare 282
A, Bangana I, Maina AG, Koffi JN, Thiam H, Bezeid OE, Salviato A, Nisi R, 283
Terregino C, Capua I. 2010. Emergence of a new genetic lineage of Newcastle 284
disease virus in West and Central Africa--implications for diagnosis and control. 285
Vet Microbiol 142:168-176. 286
9. Collins MS, Strong I, Alexander DJ. 1994. Evaluation of the molecular basis of 287
pathogenicity of the variant Newcastle disease viruses termed "pigeon PMV-1 288
viruses". Arch Virol 134:403-411. 289
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

14
10. Diel DG, da Silva LH, Liu H, Wang Z, Miller PJ, Afonso CL. 2012. Genetic 290
diversity of avian paramyxovirus type 1: Proposal for a unified nomenclature and 291
classification system of Newcastle disease virus genotypes. Infect Genet Evol 292
12:1770-1779. 293
11. Gould AR, Kattenbelt JA, Selleck P, Hansson E, Della-Porta A, Westbury 294
HA. 2001. Virulent Newcastle disease in Australia: molecular epidemiological 295
analysis of viruses isolated prior to and during the outbreaks of 1998-2000. Virus 296
Res 77:51-60. 297
12. Irvine RM, Aldous EW, Manvell RJ, Cox WJ, Ceeraz V, Fuller CM, Wood 298
AM, Milne JC, Wilson M, Hepple RG, Hurst A, Sharpe CE, Alexander DJ, 299
Brown IH. 2009. Outbreak of Newcastle disease due to pigeon paramyxovirus 300
type 1 in grey partridges (Perdix perdix) in Scotland in October 2006. Vet Rec 301
165:531-535. 302
13. Jindal N, Chander Y, Chockalingam AK, de Abin M, Redig PT, Goyal SM. 303
2009. Phylogenetic analysis of Newcastle disease viruses isolated from waterfowl 304
in the upper midwest region of the United States. Virol J 6:191. 305
14. Jørgensen PH, Handberg KJ, Ahrens P, Therkildsen OR, Manvell RJ, 306
Alexander DJ. 2004. Strains of avian paramyxovirus type 1 of low pathogenicity 307
for chickens isolated from poultry and wild birds in Denmark. Vet Rec 154:497-308
500. 309
15. Kattenbelt JA, Stevens MP, Selleck PW, Gould AR. 2010. Analysis of 310
Newcastle disease virus quasispecies and factors affecting the emergence of 311
virulent virus. Arch Virol 155:1607-1615. 312
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

15
16. Kho CL, Mohd-Azmi ML, Arshad SS, Yusoff K. 2000. Performance of an RT-313
nested PCR ELISA for detection of Newcastle disease virus. J Virol Methods 314
86:71-83. 315
17. Kim BY, Lee DH, Kim MS, Jang JH, Lee YN, Park JK, Yuk SS, Lee JB, 316
Park SY, Choi IS, Song CS. 2012. Exchange of Newcastle disease viruses in 317
Korea: The relatedness of isolates between wild birds, live bird markets, poultry 318
farms and neighboring countries. Infect Genet Evol 12:478-482. 319
18. Kim LM, King DJ, Curry PE, Suarez DL, Swayne DE, Stallknecht DE, 320
Slemons RD, Pedersen JC, Senne DA, Winker K, Afonso CL. 2007. 321
Phylogenetic diversity among low-virulence newcastle disease viruses from 322
waterfowl and shorebirds and comparison of genotype distributions to those of 323
poultry-origin isolates. J Virol 81:12641-12653. 324
19. Kim LM, Suarez DL, Afonso CL. 2008. Detection of a broad range of class I 325
and II Newcastle disease viruses using a multiplex real-time reverse transcription 326
polymerase chain reaction assay. J Vet Diagn Invest 20:414-425. 327
20. Kommers GD, King DJ, Seal BS, Brown CC. 2001. Virulence of pigeon-origin 328
Newcastle disease virus isolates for domestic chickens. Avian Dis 45:906-921. 329
21. Lindh E, Ek-Kommonen C, Vaananen VM, Alasaari J, Vaheri A, Vapalahti 330
O, Huovilainen A. 2012. Molecular epidemiology of outbreak-associated and 331
wild waterfowl- derived Newcastle disease virus strains in Finland, including a 332
novel class I genotype. J Clin Microbiol. 333
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

16
22. Lindh E, Huovilainen A, Ratti O, Ek-Kommonen C, Sironen T, Huhtamo E, 334
Poysa H, Vaheri A, Vapalahti O. 2008. Orthomyxo-, paramyxo- and flavivirus 335
infections in wild waterfowl in Finland. Virol J 5:35. 336
23. OIE. 2012. Chapter 2.3.14. - Newcastle disease. In OIE (ed.), Manual of 337
Diagnostic Tests and Vaccines for Terrestrial Animals, 7th ed, 338
http://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.03.14_NEWCA339
STLE_DIS.pdf. 340
24. Ruenphet S, Jahangir A, Shoham D, Morikawa K, Miyoshi Y, Hanawa E, 341
Okamura M, Nakamura M, Takehara K. 2011. Surveillance and 342
characterization of Newcastle disease viruses isolated from northern pintail (Anas 343
acuta) in Japan during 2006-09. Avian Dis 55:230-235. 344
25. Shengqing Y, Kishida N, Ito H, Kida H, Otsuki K, Kawaoka Y, Ito T. 2002. 345
Generation of velogenic Newcastle disease viruses from a nonpathogenic 346
waterfowl isolate by passaging in chickens. Virology 301:206-211. 347
26. Snoeck CJ, Ducatez MF, Owoade AA, Faleke OO, Alkali BR, Tahita MC, 348
Tarnagda Z, Ouedraogo JB, Maikano I, Mbah PO, Kremer JR, Muller CP. 349
2009. Newcastle disease virus in West Africa: new virulent strains identified in 350
non-commercial farms. Arch Virol 154:47-54. 351
27. Takakuwa H, Ito T, Takada A, Okazaki K, Kida H. 1998. Potentially virulent 352
Newcastle disease viruses are maintained in migratory waterfowl populations. Jpn 353
J Vet Res 45:207-215. 354
28. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. 355
MEGA5: molecular evolutionary genetics analysis using maximum likelihood, 356
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

17
evolutionary distance, and maximum parsimony methods. Mol Biol Evol 357
28:2731-2739. 358
29. Terregino C, Cattoli G, Grossele B, Bertoli E, Tisato E, Capua I. 2003. 359
Characterization of Newcastle disease virus isolates obtained from Eurasian 360
collared doves (Streptopelia decaocto) in Italy. Avian Pathol 32:63-68. 361
30. Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTAL W: improving the 362
sensitivity of progressive multiple sequence alignment through sequence 363
weighting, position-specific gap penalties and weight matrix choice. Nucleic 364
Acids Res 22:4673-4680. 365
31. Vidanovic D, Sekler M, Asanin R, Milic N, Nisavic J, Petrovic T, Savic V. 366
2011. Characterization of velogenic Newcastle disease viruses isolated from dead 367
wild birds in Serbia during 2007. J Wildl Dis 47:433-441. 368
32. Wang Z, Liu H, Xu J, Bao J, Zheng D, Sun C, Wei R, Song C, Chen J. 2006. 369
Genotyping of Newcastle disease viruses isolated from 2002 to 2004 in China. 370
Ann N Y Acad Sci 1081:228-239. 371
33. Wehmann O, Herczeg J, Tanyi J, Nagy E, Lomniczi B. 1999. Lentogenic field 372
isolates of Newcastle disease virus isolated in Canada and Hungary are identical 373
with the vaccine type used in the region. Avian Pathol 28:6-12. 374
34. Werner O, Romer-Oberdorfer A, Kollner B, Manvell RJ, Alexander DJ. 375
1999. Characterization of avian paramyxovirus type 1 strains isolated in Germany 376
during 1992 to 1996. Avian Pathol 28:79-88. 377
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

18
35. Wu S, Wang W, Yao C, Wang X, Hu S, Cao J, Wu Y, Liu W, Liu X. 2011. 378
Genetic diversity of Newcastle disease viruses isolated from domestic poultry 379
species in Eastern China during 2005-2008. Arch Virol 156:253-261. 380
36. Zanetti F, Berinstein A, Pereda A, Taboga O, Carrillo E. 2005. Molecular 381
characterization and phylogenetic analysis of Newcastle disease virus isolates 382
from healthy wild birds. Avian Dis 49:546-550. 383
37. Zhu W, Dong J, Xie Z, Liu Q, Khan MI. 2010. Phylogenetic and pathogenic 384
analysis of Newcastle disease virus isolated from house sparrow (Passer 385
domesticus) living around poultry farm in southern China. Virus Genes 40:231-386
235. 387
388
389
390
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

19
Figure legends 391
FIG 1 Geographic distribution of collected samples by municipalities in Luxembourg. 392
The shading corresponds to the number of samples collected per municipalities. The 393
numbers indicate the origin of the following isolates: 1) duck/Luxembourg/26/2006 and 394
pigeon/Luxembourg/119/2006; 2) great cormorant/Luxembourg/2547/2006; 3) 395
chicken/Luxembourg/2871-18/2007; 4) duck/Luxembourg/3785/2007 and 396
duck/Luxembourg/3786/2007; 5) pigeon/Luxembourg/3821-1/2007; 6) 397
mallard/Luxembourg/4178/2008. The strain pigeon/Luxembourg/2657-2/2006 originated 398
from an animal rescued at the animal wildlife shelter in Dudelange (7). Template map © 399
Origine Cadastre: Droits Reserves a l’Etat de Grand-Duche de Luxembourg (2012). 400
401
FIG 2 Phylogenetic analysis of partial F gene sequences based on nucleotides 332–571. 402
Sequences generated in this study are indicated by the symbols ● (strains presented in 403
Fig. 1 and Fig. 2) and ■ (strains presented in Fig. 1 only). Previously published sequences 404
are indicated with their accession numbers. Only bootstrap values ≥ 50% are shown. The 405
scale corresponds to number of base substitutions per site. 406
407
FIG 3 Phylogenetic analysis of complete F gene sequences (1662 nt). Symbols are as in 408
Fig. 1. Only bootstrap values ≥ 50% are shown. The scale corresponds to number of base 409
substitutions per site.410
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

20
Tables 411
Table 1: Sequences of primers used to amplify and sequence partial or full F genes. 412
413 Primer Orientation Sequence (5’ – 3’) Localization Reference FOP1 Forward TACACCTCATCCCAGACAGGGTC F gene (16) FOP2 Reverse AGGCAGGGGAAGTGATTTGTGGC F gene (16) FIP1 Forward TACTTTGCTCACCCCCCTT F gene (16) FIP2 Reverse CATCTTCCCAACTGCCACT F gene (16) M610 Forward CTGTACAATCTTGCGCTCAATGTC M gene (1)
P1 Forward ATGGGCYCCAGAYCTTCTAC F gene (32) F581 Reverse CTGCCACTGCTAGTTGTGATAATCC F gene (1)
F-4639f Forward TGAYGGCAGGCCTCTT F gene this study F-4932f Forward CAACCGCTGCACAGATAA F gene this study F-4954f Forward AGCTGCGGCYCTRATACAA F gene this study F-5042f Forward GAGGTCACYGACGGATTAT F gene this study F-5488f Forward TCAGCACTTGTCCCAAA F gene this study
F-1258-R Reverse ACATTGCATGAWTGTCTRTC F gene (8) F-5566r Reverse CAGTATGAGGTGTCAAGTT F gene this study F-5749f Forward AGACCCTCCAGGYATCA F gene this study F-5888f Forward GGCTCAGTGGGGAAT F gene this study F-6086f Forward GGTACACTTAGCCTGRTHTT F gene this study F-6146r Reverse CTTYTGTTGCGCCTTT F gene this study
F-7979-R Reverse AGRGCCACYTGCTTRTATA HN gene (8)
414 415
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
21
Table 2: List of sampled domestic and wild bird species in Luxembourg between 2006 416
and 2010 tested for class I and class II and genotype (G-) classification of positive 417
samples. 418
419
Family No. positives/ No. tested for
class II
No. positives/ No. tested for
class I
Yearly distribution
2006 2007 2008
Phasianidae a 1/125 0/62 1x G-II Anatidae 4/340 0/92 1x G-II 2x G-I 1x G-I
Phalacrocoracidae 1/3 0/1 1x G-II Ardeidae 0/13 0/2
Accipitridae 0/135 0/7 Falconidae 0/26 0/6
Rallidae 0/52 0/4 Laridae 0/8 -
Columbidae 3/59 0/11 2x G-VI 1x G-VI Psittacidae b 0/3 0/3
Strigidae 0/12 0/1 Tytonidae 0/14 0/1 Picidae 0/5 0/1
Corvidae 0/20 0/3 Paridae 0/5 0/2
Hirundinidae 0/9 0/9 Phylloscopidae 0/5 0/2 Acrocephalidae 0/42 0/32 Locustellidae 0/4 0/3
Sylviidae 0/95 0/37 Sturnidae 0/7 - Turdidae 0/21 0/15
Muscicapidae 0/9 0/4 Passeridae 0/42 - Prunellidae 0/2 0/1 Motacillidae 0/4 - Fringillidae 0/1 0/1 Emberizidae 0/4 0/3
Undetermined 0/66 0/42 Total 9/1131 0/345 4/619 4/349 1/84
a domestic species including chicken, turkey, quail, peafowl and pheasant; b exotic species 420
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

0 10 205 Kilometers
0
1 to 10
11 to 50
> 50
1
2
3
7
46
5
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

VI
II
X
I
Class I
JX844035 pigeon/Finland/17557/2008pigeon/Luxembourg/2657-2/2006
JN986839 pigeon/IE/806/04AJ306304 pigeon/France/99299/1999
JN941997 Pigeon/Florida/2181-43/2008pigeon/Luxembourg/119/2006
GU002430 pigeon/Slovenia/SLO-263/2004AJ306305 pigeon/France/99106/1999AY288996 pigeon/Italy/1166/00GU002440 blackbird/Slovenia/SLO-12/2006HM748948 chicken/China/SD5/2008
JN872162 Rosella/Belgium/4940/08FJ766527 pigeon/China/JS/07/16/Pi/2007
HM063425 pigeon/China/P4/2003EF026584 pigeon/Belgium/248 VB/1998
AY208697 pigeon/China/HLJ-4/2002GU002428 pigeon/Slovenia/SLO-17/2004
pigeon/Luxembourg/3821-1/2007HM625835 duck/China/SS/2008
GU551934 carrier-pigeon/Guangdong/2008JX844036 pigeon/Finland/15475/2009
JN872160 Pigeon/Minnesota/511296/07AY034799 pigeon/Finland/Fin-96c/1996
JN872183 Pigeon/Maryland/2075/1998EU477192 Eurasian collared dove/US/TX4156/2005JN872170 Pigeon/Texas/5254-12/2010
JN638235 dove/Italy/11RS100_104VIR/2011AY734535 Pigeon/Argentina/Tigre 6/99
JN967787 Pigeon/Illinois/37397/1987JN967788 Pigeon/Maryland/11936/1985FJ410145 New York/1984
AB465606 Japan/Ibaraki/85FJ772475 chicken/Niger/2602-605/2008 --- XIV
JN627508 goose/China/GD450/2011 --- XIIAF458011 chicken/China/XJ-2-97/1997 --- XVAF358786 fowl/Taiwan/TW/2000/2000 --- VII
AY865652 Sterna/Astr/2755/2001 --- XIIIAF048763 chicken/Malaysia/AF2240/1960 --- VIII
FJ705462 cormorant/US(WI)/18719-03/2003 --- VFJ436302 chicken/China/F48E8/1946 --- IX
HQ266604 chicken/Madagascar/MG_MEOLA_08/2008 --- XIEF201805 India/Mukteswar --- III
AY741404 Herts/33 --- IVJX482549 seafowl/China/H5/2011
chicken/Luxembourg/2871-18/2007JX482548 seafowl/China/H1/2011DQ227250 penguin/Beijing/QE01/1999AY359876 parrot/India/NDVCUL97JN872150 B1AJ415880 pigeon/France/99143/1999AY845400 LaSotaFJ938171 sparrow/Guangxi/NN10/2007AY727883 flamingo/Argentina/88T.00/2000
great cormorant/Luxembourg/2547/2006duck/Luxembourg/26/2006
GQ901891 parrot/Malaysia/MB061/06/2006AY390314 pigeon/China/PK9910
HM001217 penguin/China/QBJ-00/2000AY130863 yellow nape parrot/U.S.(PA)/22027/96
JN942033 Avian/Chile/37646/1996FJ939313 Chicken/Egypt/1/2005
X04719 BeaudetteGQ288390 mallard/US(MN)/00-32/2000AY727881 duck/Argentina/32C/T.98/1998
AY562991 chicken/N. Ireland/Ulster/1967M24693 QueenslandV4/66
AY427817 Heb02AY935499 I-2
DQ097394 PHY-LMV42/HungaryEF564817 ruddy turnstone/US(DE)/492/2002
AY965077 Anas/FarEast/3658/2002FJ597613 duck/China/D_ZJ_1_04/2004JN872169 Pennsylvania/3167/2009FJ597609 duck/China/D_JS_51_05/2005JX844031 pheasant/Finland/7775/2003
JX844039 mallard/Finland/8803/2009FJ597606 duck/China/D_JS_40_05/2005
HM063424 Rallus aquaticus/China/R8/2005JX844041 teal/Finland/10666/2009
FJ597617 duck/China/D_HN_34_05/2005FJ597615 duck/China/D_ZJ_3_04/2005FJ597604 duck/China/D_AH_6_04/2004
JX844048 mallard/Finland/8768/2010EU256542 Muskovy duck/Ukraine/14/2005
AY972103 Duck/FarEast/2687/2001JX844053 mallard/Finland/9149/2010
AY972101 Anas/FarEast/3652/2002JX844040 Eurasian wigeon/Finland/9067/2009
AY965079 duck/FarEast/2713/2001EU493450 teal/Finland/12074/2006
JX844043 Eurasian_wigeon/Finland/8625/2010JX844058 mallard/Finland/10541/2010
mallard/Luxembourg/4178/2008duck/Luxembourg/3786/2007duck/Luxembourg/3785/2007
JX844056 mallard/Finland/9360/2010EU493453 teal/Finland/12136/2006
AB524405 Goose/Alaska/415/91
98
87
99
61
69
51
6053
58
71
69
8499
54
5283
62
89
7063
67
75
92
64
60
51
86
0.05
Class II
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
pigeon/Luxembourg/119/2006
JN872166 Dove/Italy/VIR 24/2008
AY288996 pigeon/Italy/1166/00
JN986839 pigeon/IE/806/04
JN941997 Pigeon/Florida/2181-43/2008
pigeon/Luxembourg/3821-1/2007
EF026584 pigeon/Belgium/248 VB/1998
HM625835 duck/China/SS/2008
HM063425 pigeon/China/P4/2003
HM748948 chicken/China/SD5/2008
GU551934 carrier-pigeon/Guangdong/2008
FJ766527 pigeon/China/JS/07/16/Pi/2007
JN872162 Rosella/Belgium/4940/08
JN872183 Pigeon/Maryland/2075/1998
JN872170 Pigeon/Texas/5254-12/2010
JN872160 Pigeon/Minnesota/511296/07
JN967787 Pigeon/Illinois/37397/1987
AY734535 Pigeon/Argentina/Tigre 6/99
JN967788 Pigeon/Maryland/11936/1985
FJ410145 New York/1984
AY845400 LaSota
X04719 Beaudette
JQ029740 duck/China/SDFC07/2011
AY427817 Heb02
AY935499 I-2
EF564817 ruddy turnstone/US(DE)/492/2002
DQ097394 PHY-LMV42/Hungary
AY562991 chicken/N. Ireland/Ulster/67
JN872169 Pennsylvania/3167/2009
AY965077 Anas/FarEast/3658/2002
FJ597609 duck/China/D_JS_51_05/2005
FJ597617 duck/China/D_HN_34_05/2005
FJ597615 duck/China/D_ZJ_3_04/2005
AY972103 Duck/FarEast/2687/2001
AY965079 duck/FarEast/2713/2001
FJ597604 duck/China/D_AH_6_04/2004
HM063424 Rallus aquaticus/China/R8/2005
mallard/Luxembourg/4178/2007
duck/Luxembourg/3786/2007
duck/Luxembourg/3785/2007
AB524405 Goose/Alaska/415/91
100
9999
98
69
100
8986
82
100
99
10069
99
100
9599
99
94
70
69
81
56
53
94
0.05
I
Class I
Class II
VI
II
on July 1, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from