characterization of chromosome aberrations - cancer research
TRANSCRIPT

[CANCER RESEARCH 47, 5162-5170, October 1, 1987]
Characterization of Chromosome Aberrations Induced by Incubation at aRestrictive Temperature in the Mouse Temperature-sensitive MutanttsFT20 Strain Containing Heat-labile DNA Polymerase a1
Toshihiko Eld,2 Takemi Enomoto, Yasufumi Murakami,3 Fumio Hanaoka, and Masa-atsu Yamada4
Department of Physiological Chemistry, Faculty of Pharmaceutical Sciences, University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113, Japan
ABSTRACT
tsFT20 cells derived from a mouse mammary carcinoma cell line,FM3A, which has temperature-sensitive DNA polymerase a activity (Y.Murakami, (I. Yasuda, H. Miyazawa, F. Hanaoka, and M. Yamada,Proc. Nati. Acad. Sci. USA, 82:1761-1765,1985) were rapidly committed to death after temperature upshift to 39°C.tsFT20 cells synchronized
in S phase were more sensitive to the restrictive temperature thanexponentially growing cells. In order to gain insight into the processesfrom the interruption of DNA synthesis to cell death, we analyzedchromosome aberrations induced in tsFT20 cells which had been incubated for 2 or 4 h at the restrictive temperature and then cultured at thepermissive temperature. The majority of metaphase cells showed extensive chromosome aberrations such as chromatid gaps, breaks, and exchanges; chromosome pulverizations; their mixed types; and ring chromosomes. Analyses with the use of cell synchronization and autoradiog-raphy revealed that chromosome aberrations were induced only in thecells which synthesized DNA during incubation at 39°C.We classified
the chromosome aberrations into five types: gap or break type; exchangetype; pulverization type; complex type; and ring type. The temporal orderof the appearance of these types of chromosome aberrations was foundto be the above described order. It was further found that cycloheximidedramatically repressed the induction of chromosome aberrations, andmetaphases with many chromosome aberrations exhibited a large numberof sister chromatid exchanges. These results indicate that abnormalcessation of DNA replication in tsFT20 cells at the restrictive temperature due to the inactivation of DNA polymerase a results in cell deathvia induction of double-strand breaks which lead to chromosome aberrations as well as sister chromatid exchanges.
INTRODUCTION
During recent years, it has become well known that besidesdirect DNA lesions induced by X-rays or chemicals, the indirectDNA lesions induced by imbalance of DNA precursor metabolism also cause extensive chromosome instability in eukaryoticcells. In addition, it is known that when de novo synthesis ofthymidylate is blocked, growing cells die. This phenomenon iscalled thymineless death and has been initially found in pro-karyotic cells (1). Thymineless death has been studied in eukaryotic cells treated with drugs that block thymidylate metabolism(2-6) and auxotrophic thy"5 mutants (7-9). Recent studies
using thy mutants have shown that thymidine starvation inmammalian cells induces extensive chromosome aberrationsand results in thymineless death (10, 11). On the other hand,little is known of what happens in cells in which enzymes
Received 3/3/87; revised 6/19/87; accepted 6/24/87.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1This work was supported in part by Grants-in-Aid for Scientific Researchand for Cancer Research from the Ministry of Education, Science and Culture,Japan, and by the Naito Foundation.
2To whom requests for reprints should be addressed.3 Present address: Riken Gene Bank, The Institute of Physical and Chemical
Research.4 Present address: Hatano Research Institute, Food and Drug Safety Center.5The abbreviations used are: thy", thymidylate synthase-negative; CMF-PBS,
calcium- and magnesium-free phosphate-buffered saline; FdUrd, 5-fluoro-2'-deoxyuridine; SCE, sister chromatid exchange; ts. temperature-sensitive.
directly participating in DNA replication are inhibited becauseof the lack of specific inhibitors for DNA replication enzymesexcept for aphidicolin and the unavailability of DNA ts mutantshaving ts enzymes that participate in DNA replication.
In recent years, we have tried to isolate ts mutants related toDNA replication in order to get a tool for the analysis of themolecular mechanism of DNA replication in mammalian cells,and we have isolated several such mutants (12-14). One ofthese ts mutants, tsFT20, was found to have heat-labile DNApolymerase «activity (12). Recently, we have succeeded inproving that the DNA polymerase a molecule of tsFT20 cellsitself is heat-labile by using an ¡mmunoaffinity-purified enzyme.6 tsFT20 cells are typical DNA ts mutants, which showrapid decrease in DNA-synthesizing ability after temperatureupshift and are arrested in the S phase. It was found that therapid decrease in DNA-synthesizing ability correlated well withthe decrease in the intracellular level of DNA polymerase aactivity (15) and was due to the decrease in the frequency ofreplicón initiation (16). In addition, it has been observed thatts cells are rapidly committed to death after exposure to therestrictive temperature.
In this study we have examined chromosome aberrationsinduced in tsFT20 cells in order to gain insight into the processes from the interruption of DNA replication, due to theinactivation of the DNA polymerase a molecule, to cell death.
MATERIALS AND METHODS
Cell Culture. tsFT20 cells (12) and FM3A clone 28 cells (wild-type)which were originally established from a spontaneous mammary carcinoma in a C3H/He mouse (17) were incubated at 33°C(permissivetemperature) or at 39°C(restrictive temperature) in suspension culture
in RPMI 1640 (Flow Laboratories, England) supplemented with 10%calf serum (Flow Laboratories, North Ryde, Australia). Cells used inthese experiments were free of Mycoplasma contamination.
Assay for Colony-forming Ability. Exponentially growing tsFT20cells and wild-type cells or tsFT20 cells synchronized at Gi-S boundary(approximately 1.5 x IO6) were inoculated in glass tubes (1.5 x 10.5
cm) containing 5 ml of the growth medium. The cells were incubatedat 39°Cfor indicated periods in a water bath and then collected bycentrifugation at 1400 x g for 5 min at 4°C.After a washing with ice-
cold CMF-PBS, the cells were suspended in and diluted with the growthmedium to a cell concentration of 100 and 300/ml. One volume of thediluted cell suspension was mixed with 2 volumes of 0.5% agar solution(agar Noble; Difco Laboratories, Detroit, MI) dissolved in RPMI 1640containing 10% calf serum and antibiotics (100 //g/nil streptomycinsulfate and 100 units/ml penicillin G potassium), and 3 ml of themixture were poured on 4 ml of 0.5% underlayer agar in a glass dish(60 mm in diameter). After incubation at 33°Cfor 2 weeks in CO2
incubator, the number of colonies in the soft agar was counted (4dishes/point).
Cell Synchronization. Exponentially growing cells were inoculatedinto the growth medium containing 10% dialyzed calf serum and theantibiotics at a concentration of 1.5 x IO5 cells/ml. The cells wereincubated at 33°Cfor 1 day and FdUrd was added at a final concentra-
" R. Takayama, S. Tada, F. Hanaoka, and M. Ui, unpublished data.
5162
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
tion of 3 x 10~8 M. The cells were incubated at 33°Cfor 14 h to be
arrested at d-S boundary. The cells were released from the FdUrdblock by two washings with ice-cold CMF-PBS and suspended in warmgrowth medium containing the antibiotics. Over 80% of the populationthus synchronized was arrested at the d-S boundary and most of theremainder was arrested in S phase according to cytofluorometric analysis (15).
Assays for Chromosome Aberrations and Mitotic Index. The cells (3x IO5) from exponentially growing cultures or synchronized cultures
were inoculated into a glass dish (diameter 30 mm) with 2 ml of thegrowth medium. After incubation for indicated periods, Colcemid wasadded to the culture at a final concentration of 0.1 ¿ig/ml,and the cellswere incubated at 33 °Cfor indicated periods in a CC>2incubator. The
cells were chilled in an ice-water bath and transferred into a glass tube.The cells were collected by centrifugation and washed with ice-coldCMF-PBS. The pellet was resuspended in 1 ml of 0.5% sodium citrateand incubated at 37°Cfor 5 min with shaking, and then 5 ml of the
fixative (methanol:acetic acid, 3:1) were added. After standing at roomtemperature for 5 min, the suspension was centrifuged, and the pelletwas washed once with the fixative. The resultant pellet was suspendedin a small volume of the fixative. The suspension was dropped on aglass slide and air dried. Cells fixed on the glass were stained with 3%Giemsa solution for 15 min. Mitotic index was determined by examining about 500 cells. As for chromosome aberrations, about 200metaphases were examined at x 400. Chromosome aberrations areclassified as follows. "Gap or break type" is defined as the aberration
exhibiting a chromosome or chromatid gap or break observed in ametaphase which contains less than six aberrations. "Exchange type"
is defined as the aberration exhibiting a chromatid interchange or achromosome/chromatid interchange observed in a metaphase whichcontains less than six aberrations. "Complex type" is an aberration
consisting of more than six aberrations within a metaphase includingchromatid gaps, breaks, exchange types, and the fragmented chromosomes as shown in Fig. 2, b-d. "Pulverization type" is represented bypulverized chromosomes as shown in Fig. le. "Ring type" is the
aberration exhibiting a centric ring or a ring derived from a chromo-some-chromatid interchange. The frequencies of gap or break type,exchange type, and ring type are expressed as the number of sitesexhibiting aberrations per 100 metaphases by examining about 200metaphases. The frequencies of complex type and pulverization typeare expressed as the number of metaphases exhibiting the aberrationper 100 metaphases by examining about 200 metaphases.
Autoradiographic Analysis. Exponentially growing cells (5 x 10s)were inoculated into a culture flask (25 cm2) containing 5 ml of the
growth medium supplemented with 10% dialyzed calf serum and theantibiotics. The cells were incubated at 33"C for 2 days and thenincubated at 39°Cfor 4 h. The cells were pulse-labeled with 4 ^Ci/ml[3H]thymidine (20 Ci/mmol; Amersham, United Kingdom) for 10 min.
The labeled cells were collected by centrifugation and washed twicewith ice-cold CMF-PBS. The cells were suspended at a concentrationof 1.5 x 10' cells/ml in the growth medium containing 1 x IO"5 Mthymidine and 0. l j/g/ml Colcemid. After incubation at 33°Cfor 24 h,
the cells were fixed on a glass slide and processed to autoradiographyas described previously (16). The existence of silver grains and chromosome aberrations was examined with about 300 metaphases at x400.
Assay for SCEs. Exponentially growing cells (2.5 x IO5)were inoculated into a culture flask (25 cm2) containing 5 ml of the growth
medium supplemented with 10% dialyzed calf serum and the antibiotics. 5-Bromo-2'-deoxyuridine (Sigma Chemical Co., St. Louis, MO)
was added to the culture at a concentration of 5 Mg/ml, and then thecells were incubated at 33°Cfor 18 h (approximately 1.2 cell cyclesunder these conditions) in the dark. The cells were incubated at 39°Cfor 4 h and further incubated at 33°Cfor 20 h. Colcemid was added at
a concentration of 0.08 Mg/ml to the culture, and the cells wereincubated at 33°Cfor 6 h. The cells were collected and fixed on a glassslide as described under "Assays for Chromosome Aberrations andMitotic Index." Sister chromatids were differentially stained by a
modified method of Perry and Wolff (18) as follows. The glass wasdipped in 30 /¿g/mlHoechst 33258 solution at room temperature for
15 min, washed with water, and soaked in Mcllvaine's buffer (196 mM
disodium hydrogen phosphate-2 HIMcitric acid, pH 8.25). The glassslide was covered with a cover glass and sealed with colorless nail polishto hold the Mcllvaine's buffer between the glass slide and the coverglass.
The glass slide was exposed to UV at a 5-cm distance from a germicida!lamp (Toshiba GL15) for 10 min on a hot plate set at 60"( , washed
with water, and stained with 2% Giemsa solution. The stained metaphases were examined at x 1000.
RESULTS
Decrease in Colony-forming Ability during Incubation at theRestrictive Temperature. As reported previously (12, 15, 16),tsFT20 cells have heat-labile DNA polymerase a activity. Asshown in Fig. la, DNA polymerase a activity in tsFT20 cellsdecreased immediately after temperature upshift in contrast tothe activity in wild-type cells. Fig. \b shows the colony-formingability of tsFT20 cells after the temperature upshift. Exponentially growing tsFT20 cells lost colony-forming ability rapidlyafter incubation at 39°C.When tsFT20 cells were synchronized
with FdUrd and shifted to the restrictive temperature after therelease from the FdUrd block, the decrease in colony-formingability of the synchronized cells became much more prominentthan that of exponentially growing cells. Only 8% of colony-forming ability was obtained with the synchronized cells afterincubation at 39°Cfor 8 h. Little decrease in colony-forming
ability was observed with synchronized wild-type cells.Chromosome Aberrations Induced in tsFT20 Cells at the Re
strictive Temperature. We next studied the effect of the exposureof tsFT20 cells to the restrictive temperature from the chromosome-morphological point of view, especially chromosomeaberrations. Fig. 2 shows typical metaphase figures containingchromosome aberrations obtained from the cells which havebeen incubated at 39°Cfor 4 h and then incubated at 33°Cfor
24 h in the presence of Colcemid. Various types of chromosomeaberrations were observed: simple chromatid-type chromosomeaberrations such as chromatid gaps and breaks (Fig. la; ArrowsE-H and J) and chromatid interchanges (Arrows A-D and /);"complex-type" chromosome aberrations characterized by
many chromatid gaps, breaks, and exchanges (Fig. 2, b-d); and
~ loo
so
012« 8Incubation period al 39 *C (h)
0124 eIncubation period at 39'C (h)
Fig. 1. (a) Changes in the level of intracellular ONA polymerase a activity.Exponentially growing tsFT20 cells (•)and wild-type cells (O) were shifted up to39'C and cultured for the indicated periods. The preparation of crude cell extracts
and assay for DNA polymerase a were performed as described previously (12).(b) Changes in colony-forming ability of exponentially growing tsFT20 cells,synchronized tsFT20 cells, and synchronized wild-type cells. Cells were synchronized at the d-S boundary with FdUrd as described under "Materials andMethods." After the release from the FdUrd block, the cells were incubated at39"C for the indicated periods and assayed colony-forming ability as describedunder "Materials and Methods." Bars, SE. Synchronized tsFT20 cells (•),exponentially growing tsFT20 cells (•),wild-type cells (O).
5163
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
F
a
Fig. 2. Chromosome aberrations induced in tsFT20 cells by incubation at the restrictive temperature. Exponentially growing tsFT20 cells were incubated at 39'Cfor 4 h and then incubated at 33'C for 24 h in the presence of 0.1 <¿g/mlColcemid. (a) Slightly damaged metaphase figure containing chromatid gaps or breaks(Arrows ill, and J) and chromatid interchanges (Arrows A-D, and /); (¿>)metaphase figure exhibiting slight complex-type chromosome aberrations, in which severalchromatid interchanges and breaks are observed; (c) and (</) metaphase figures exhibiting typical complex-type chromosome aberrations; (e) metaphase figureexhibiting pulverization-type chromosome aberrations. Bars, 10 t/m.
5164
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
fsfÃÕL.*̂AM-- «-̂^?.'£)& "*¿W^ t * ¿e.A -'%^>^
:^ - ^» \• ** ^i. x. V
'W»
Fig. 3. Relationship between the induction of chromosome aberrations and DNA replication. Exponentially growing tsFT20 cells were incubated at 39'C for 4 hand pulse-labeled with ['Hjthymidine for 10 min. After being washed, the cells were cultured at 33*C for 24 h in the presence of Colcemid and fixed. Metaphasescontaining normal chromosomes and pulverization-type chromosomes and that exhibiting complex-type chromosome aberrations, which were stained with Giemsa,are shown in (<•)and (¡I).The samples were destained with isopropyl alcohol and processed for autoradiography. (a) and (h) are autoradiograms of the samemetaphases of (c) and (</), respectively. Hun, 10 »mi.
chromosome pulverization forming pulverized chromosomefragments (Fig. 2e). The complex-type chromosome aberrationand chromosome pulverization characterized the chromosomeaberration induced in tsFT20 cells by exposing the cells to therestrictive temperature, and such aberrations were induced atvery low frequency in wild-type cells and the revenants derivedfrom tsFT20 cells (data not shown).
Analysis of Relationship between Induction of ChromosomeAberrations and Position of Cells in the Cell Cycle. In order todetermine the relationship between the induction of chromosome aberrations and DNA replication, tsFT20 cells were pulse-labeled for 10 min with [3H]thymidine after incubation at the
restrictive temperature for 4 h and then incubated at the permissive temperature for 32 h in the presence of Colcemid.
Fig. 3 shows metaphase figures exhibiting chromosome aberrations (c, pulverization type; d, exchange type) and autoradiograms of the same metaphases containing silver grains (a,lì).From the results summarized in Table 1, it is revealed thatall metaphases exhibiting chromosome aberrations contain silver grains, suggesting that chromosome aberrations are inducedonly in the cells which were in the S phase when they wereincubated at the restrictive temperature. On the other hand, the
Table 1 Relationship between DNA synthesis and the induction of chromosomeaberrations in lsFT20 cells by incubation at the restrictive temperature
Exponentially growing tsFT20 cells were pulse-labeled with [3H]thymidine for10 min after incubation at 39'C for 4 h. After washing, mitotic cells wereaccumulated at 33*C for 32 h in the presence of Colcemid. They were processedas described under "Materials and Methods."
Chromosome aberration (+)Grain (+)Chromosome aberration (+)Grain (—)Chromosome aberration (—)Grain (+)Chromosome aberration (—)Grain (-)Sample
1103
0
92
55Cell
no.Sample
2118
0
116
95Sample
3103
0
91
67
metaphases containing silver grains do not always show chromosome aberrations.
To analyze in detail susceptible points to induce chromosomeaberrations in the cell cycle, we have performed experimentsthe scheme of which is shown in Fig. 4. At the indicated timesafter release from synchronization, cells were shifted to 39°Cfor 2 h and then incubated at 33°Cfor 24 or 32 h in the presence
5165
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
of Colcemid. As shown in Fig. 5, the cells which were shiftedto 39°Cduring S phase exhibited various types of chromosome
aberrations at high frequencies (Fig. 5; a, gap or break type; b,exchange type; c, pulverization type; d, complex type). Theclassification of chromosome aberrations is described under"Materials and Methods." The exchange type consisted of
2h39 «C
33 «C
K h
FdUrd
Xh
release
colcemid
-»32h
sample fixationFig. 4. Scheme for analysis of the relationship between the induction of
chromosome aberrations and the position of cells in the cell cycle.
¿n^FinriH r-, nnrppn-,Pn„
n nI 3 4 S 6
n_ n 1 HO 1 Z 3 i 5 E B 10 12
• S ..-02-M-».—G1-»
Time otter release from synchronization ( h )
Fig. S. Histograms indicating the frequencies of various types of chromosomeaberrations induced at various phases in the cell cycle. Experiments were performed as described in Fig. 4. The histograms indicate the frequencies of gap- orbreak-type (a), exchange-type (b), pulverization-type (c), and complex-type (d)chromosome aberrations observed in mitotic cells which had been incubated at33°Cfor 24 h (solid column) or for 32 h (broken column) after temperature
upshift for 2 h at the indicated times. The phases of cell cycle in which themajority of the synchronized cell population exist are shown under these columnsby arrows. The classification of chromosome aberration types is described under"Materials and Methods." Lower columns, tsFT20 cells; upper columns, wild-type
cells.
2 h or 4 h39'C
33 *C
K h
sample fixation
it inn itt I l t t t t î
colcemid( 2h or Ah interval )
FäUrd releaseFig. 6. Scheme for analysis of the temporal order of the appearance of various
types of chromosome aberrations.
20
10
0
20
10
0
20
10
0
40
20
20
32 40 16 24 32 40
Time after release from synchronization ( h )
Fig. 7. Histograms indicating the temporal order of the appearance of varioustypes of chromosome aberrations in tsFT20 cells. Experiments were performedas described in Fig. 6. Shadowed regions, period of temperature upshift (2 and 410-(a) and (g), Mitotic index; (b) and (A), gap or break type; (c) and (i), exchangetype; (</) and ( ;'), complex type; (c) and (A), pulverization type; (/) and (/), ring
type. The classification of the chromosome aberration types is described under"Materials and Methods."
mainly interchange-type chromosome aberrations, especiallychromatid interchanges and a small number of chromosome/chromatid interchanges.
The cells shifted to the restrictive temperature during non-Sphase and wild-type cells showed few chromosome aberrations.It was observed that the metaphases exhibiting heavy chromosome aberrations, complex type and pulverization type, increased as the cells were incubated for longer period at 33°C
after temperature upshift.Temporal Order of the Appearance of Various Types of Chro
mosome Aberrations. In order to analyze the temporal order ofthe appearance of various types of chromosome aberrations,the experiment designed as shown in Fig. 6 was performed.The cells synchronized by the treatment of FdUrd were shiftedto 39°Cfor 2 or 4 h after the release from the FdUrd block and
then harvested at 2- or 4-h intervals to measure mitotic indexand the frequency of each type of chromosome aberration. Theresults obtained with tsFT20 cells are shown in Fig. 7.
Chromosome aberrations appeared in the cells exposed tothe restrictive temperature for 2 h as the following order: gapor break type (mode time, 8-10 h); exchange type (14-16 h);pulverization type (18-20 h); complex type (20-22 h); and ringtype (38-40 h). Double minute-like chromosomes were often
5166
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
OCO»Dercu
30
20
IO
O
20
IO
O
20
IO
40
20
40
Time after release from synchronization
( h )Fig. 8. Histograms indicating the temporal order of the appearance of various
types of chromosome aberrations in wild-type cells. Experiments were performedas described in Fig. 6. The shadowed regions indicate the period of temperatureupshift (2 h). (a) Mitotic index; (A) gap or break type; (c) exchange type; (d)complex type. The classification of the chromosome aberration types is describedunder "Materials and Methods."
observed in the cells containing ring-type chromosome aberrations (data not shown).
In the cells incubated for 4 h at the restrictive temperature,the temporal order of the appearance of various types of chromosome aberrations was the same as that of 2-h-exposed cellsbut it was observed that appearance of heavy aberrations suchas complex type and pulverization type was retarded as compared to that of 2-h-exposed cells. The time courses of theappearance of chromosome aberrations in wild-type cells areshown in Fig. 8. Slight aberrations, gap or break type (Fig. 8A)
Xa/TJc
uo
¡I
uC
3CT
a>
ov>ook_.co
(b)
16 32
Time after release from synchronization
( h )Fig. 9. Schematic illustration of the fluctuation pattern of mitotic index and
various types of chromosome aberrations. The curves were drawn according tothe results shown in Fig. 7 (a-f). (a) Mitotic index; (b) chromosome aberrations;Curve A, gap or break type; Curve B, exchange type; Curve C, complex type;Curve D, pulverization type; Curve E, ring type.
and exchange type (Fig. 8c), were detected at a considerablefrequency but few complex-type aberrations (Fig. Sd) and nopulverization-type aberrations were observed in these cells. Theaberrations observed in the wild-type cells may be mainly dueto thymidylate stress by the treatment of FdUrd as describedpreviously (6).
For easy understanding of the temporal order of the appearance of various types of chromosome aberrations, Fig. 9 showsthe fluctuation pattern of mitotic index and various types ofchromosome aberrations, which was made on the basis of theresults shown in Fig. 7.
Effect of Cycloheximide on the Induction of ChromosomeAberrations in tsFT20 Cells. It has been observed that additionof cycloheximide prevents cells from thymineless death anddecreases the frequency of chromosome aberrations induced bythymidylate stress (10, 19). Therefore, we examined the effectof cycloheximide on the induction of chromosome aberrationsin tsFT20 cells exposed to the restrictive temperature.
tsFT20 cells were incubated at 39°Cfor 4 h in the presence
or absence of cycloheximide, and mitotic cells were accumulatedby an incubation at 33°Cfor 32 h in the presence of Colcemid.
As shown in Table 2, the induction of heavy chromosomeaberrations (complex-type) was repressed dramatically by theaddition of cycloheximide. It was found that the addition ofcycloheximide alone had little effect on the induction of chromosome aberrations as reported previously (10) (data notshown).
Induction of Sister Chromaticl Exchanges in tsFT20 CellsExposed to the Restrictive Temperature. From many studies onSCE, it has been suggested that SCE is a specific phenomenonrelated to DNA replication (20-23). Therefore, the induction
5167
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
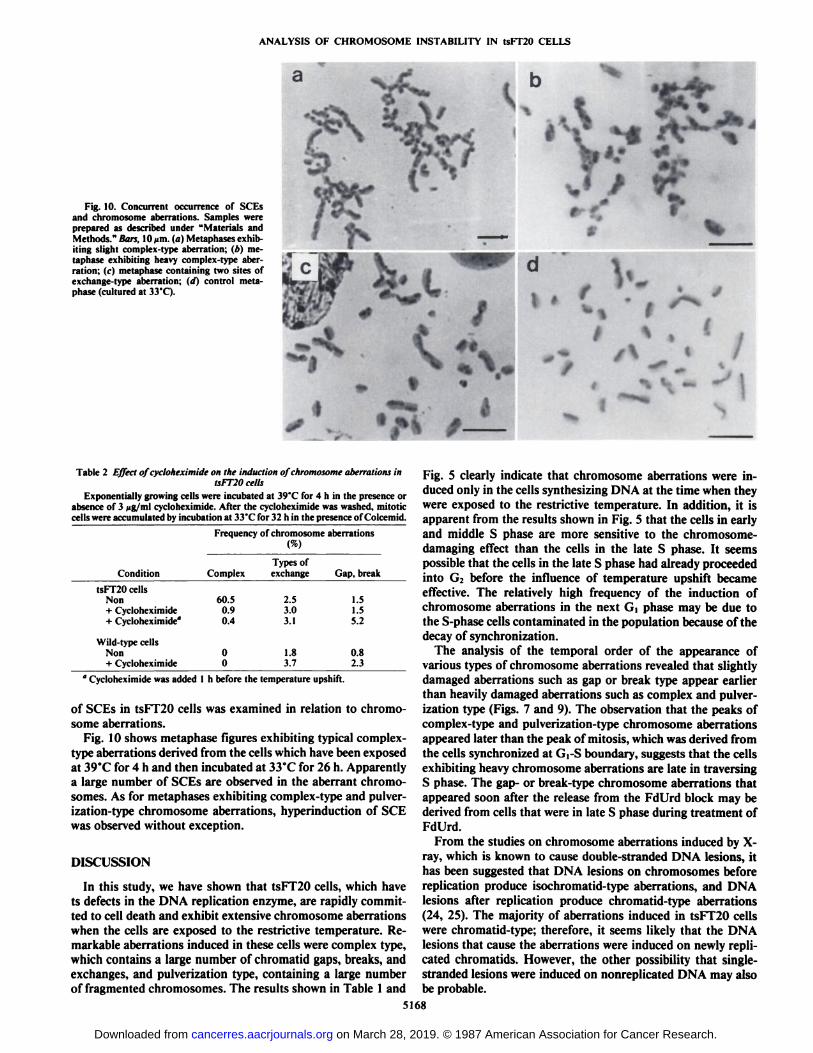
ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
Fig. 10. Concurrent occurrence of SCEsand chromosome aberrations. Samples wereprepared as described under "Materials andMethods." Bars, 10ptn.(a) Metaphases exhibiting slight complex-type aberration; (/>) me-taphase exhibiting heavy complex-type aberration; IM metaphase containing two sites ofexchange-type aberration; (¡I)control metaphase (cultured at 33*C).
Table 2 Effect of cycloheximide on the induction of chromosome aberrations intsFT20 cells
Exponentially growing cells were incubated at 39'C for 4 h in the presence or
absence of 3 jig/ml cycloheximide. After the cycloheximide was washed, mitoticcells were accumulated by incubation at 33'C for 32 h in the presence of Colcemid.
Frequency of chromosome aberrations
ConditiontsFT20
cellsNon+
Cycloheximide+Cycloheximide"Wild-type
cellsNon+
CycloheximideComplex60.50.90.400Types
ofexchange2.53.03.11.83.7Gap,break1.51.55.20.82.3
* Cycloheximide was added l h before the temperature upshift.
of SCEs in tsFT20 cells was examined in relation to chromosome aberrations.
Fig. 10 shows metaphase figures exhibiting typical complex-type aberrations derived from the cells which have been exposedat 39°Cfor 4 h and then incubated at 33°Cfor 26 h. Apparently
a large number of SCEs are observed in the aberrant chromosomes. As for metaphases exhibiting complex-type and pulverization-type chromosome aberrations, hyperinduction of SCEwas observed without exception.
DISCUSSION
In this study, we have shown that tsFT20 cells, which havets defects in the DNA replication enzyme, are rapidly committed to cell death and exhibit extensive chromosome aberrationswhen the cells are exposed to the restrictive temperature. Remarkable aberrations induced in these cells were complex type,which contains a large number of chromai id gaps, breaks, andexchanges, and pulverization type, containing a large numberof fragmented chromosomes. The results shown in Table 1 and
Fig. 5 clearly indicate that chromosome aberrations were induced only in the cells synthesizing DNA at the time when theywere exposed to the restrictive temperature. In addition, it isapparent from the results shown in Fig. 5 that the cells in earlyand middle S phase are more sensitive to the chromosome-damaging effect than the cells in the late S phase. It seemspossible that the cells in the late S phase had already proceededinto G2 before the influence of temperature upshift becameeffective. The relatively high frequency of the induction ofchromosome aberrations in the next d phase may be due tothe S-phase cells contaminated in the population because of thedecay of synchronization.
The analysis of the temporal order of the appearance ofvarious types of chromosome aberrations revealed that slightlydamaged aberrations such as gap or break type appear earlierthan heavily damaged aberrations such as complex and pulverization type (Figs. 7 and 9). The observation that the peaks ofcomplex-type and pulverization-type chromosome aberrationsappeared later than the peak of mitosis, which was derived fromthe cells synchronized at d-S boundary, suggests that the cellsexhibiting heavy chromosome aberrations are late in traversingS phase. The gap- or break-type chromosome aberrations thatappeared soon after the release from the FdUrd block may bederived from cells that were in late S phase during treatment ofFdUrd.
From the studies on chromosome aberrations induced by X-ray, which is known to cause double-stranded DNA lesions, ithas been suggested that DNA lesions on chromosomes beforereplication produce isochromatid-type aberrations, and DNAlesions after replication produce chromatid-type aberrations(24, 25). The majority of aberrations induced in tsFT20 cellswere chromatid-type; therefore, it seems likely that the DNAlesions that cause the aberrations were induced on newly replicated enromadas. However, the other possibility that single-stranded lesions were induced on nonreplicated DNA may alsobe probable.
5168
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
Ring-type chromosome aberrations were often observed inthe metaphases containing double minute chromosomes whichbelong to isochromatid-type aberrations (not shown). Themechanism of formation of the pulverized chromosomes is notclear at present; however, it must be noted that the morphologyof the pulverized chromosomes is very similar to that of premature condensed chromosomes of late S-phase cells (26).
The precise mechanism by which DNA lesions are inducedin tsFT20 cells is unknown. The observations that a largenumber of SCEs were induced in the cells with chromosomeaberrations (Fig. 10) and that cycloheximide inhibited the induction of chromosome aberrations (Table 2) may provide aclue to consider the mechanism. It is known that SCEs areformed via DNA double-strand breaks. One can speculate thatthe mechanism which induces DNA strand breaks in tsFT20cells is as follows. Upon temperature upshift, although DNAreplication once started can proceed normally within a replicón,the initiation of DNA replication at replicónorigins is inhibiteddue to the inactivation of DNA polymerase a as reportedpreviously (16). Under these conditions, replication forks maybe arrested near or at the termination points of active repliconsor replicónclusters adjacent to inactive nonreplicated replicons,and subsequent persistence of single-stranded gaps is susceptible to endonuclease attack. Furthermore, replicónorigins whichare activated to initiate replication but cannot initiate due tolimiting supply of functional DNA polymerase a may becomethe target of endonuclease attack. The inhibiting effect ofcycloheximide on the induction of chromosome aberrations canbe explained by the decrease in activated replicons because theinitiation of DNA replication at replicón origins is suggestedto be dependent on protein(s) which turn over very rapidly (27,28). However, it also may be possible that the abnormal cessation of DNA replication induces an enzyme or a protein involved in DNA strand breaks and that cycloheximide blocksthe synthesis of such an inducible protein.
Several studies on chromosomal instability have been performed with mammalian cell mutants. Besides tsF'l 20 cells,
only a small number of mutants with defects in DNA polymerase a activity have been reported (13, 29-31), and one ofthe mutants, aph'-4 strain isolated from Chinese hamster V79cells, which contains aphidicolin-resistant DNA polymerase a,has been shown to exhibit chromosome instability as well ashypermutability and UV sensitivity. Hori et al. (11) have reported that extensive chromosome aberrations were induced bythymidylate stress in thy" mutants isolated from FM3A cells.
Under slight thymidylate stress, the induction of somatic mutations at two genetic loci (the 6-thioguanine-resistant locusand the ouabain-resistant locus) was observed in the thy" mu
tant (32). In addition, it is well known that the interruption ofDNA synthesis by the treatment of inhibitors for DNA precursor synthesis induces chromosome instability and sometimesresults in gene amplification to produce drug-resistant mutants(33). However, in our tests, the induction of mutation in tsFT20cells by incubation at 39°Cwas not observed at least at the
ouabain locus.As to production of SCEs, several models have been pro
posed. Painter (23) proposed a model based on the idea thatdouble-strand breaks are generated at junctions between a completely replicated replicón cluster and a partially replicatedreplicón cluster. This model seems to fit the case of tsFT20cells. Therefore, tsFT20 cells may become a good tool for theanalysis of the mechanism of formation of SCEs as well as theanalysis of the molecular mechanism of eukaryotic DNA replication.
ACKNOWLEDGMENTS
We thank Y. Eguchi for his technical assistance.
REFERENCES
1. Cohen, S. S., and Barner, H. D. Studies on unbalanced growth in Escherichiacoli. Proc. Nati. Acad. Sci. USA, 40: 885-893, 1954.
2. Barclay, B. J., Kunz, B. A., Little, J. G., and Haynes, R. H. Genetic andbiochemical consequences of thymidylate stress. Can. J. Biochem., 60: 172-194, 1982.
3. Borsa, J., and Whitmore, G. J. Cell killing studies on the mode of action ofmethotrexate on L-cells in vitro. Cancer Res., 29: 737-744, 1969.
4. Hrynick, W. M., and Berlino, J. R. Growth rate and cell kill. Ann. NY Acad.Sci., 186: 330-342, 1971.
5. Reuckert, R. R., and Mueller, G. C. Studies on unbalanced growth in tissueculture. I. Induction and consequences of thymidine deficiency. Cancer Res.,20:1584-1591,1960.
6. Taylor, J. H., Haut, W. F., and Tung, J. Effect of fluorodeoxyuridine onDNA replication, chromosome breakage and reunion. Proc. Nati. Acad. Sci.USA, 4«:190-198, 1962.
7. Ayusawa, D., Koyama, H., Iwata, K.. and Seno, T. Single-step selection ofmouse FM3A cell mutants defective in thymidylate synthase. Somatic CellGenet., 6: 261-270, 1980.
8. Ayusawa, D., Koyama, H., Iwata, K., and Seno, T. Selection of mammalianthymidine auxotrophic cell mutants defective in thymidylate synthase bytheir reduced sensitivity to methotrexate. Somatic Cell Genet., 7: 523-534,1981.
9. Little, J. G., and Haynes, R. H. Isolation and characterization of yeastmutants auxotrophic for 2-deoxythymidine-5'-monophosphate. Mol. Gen.Genet., J68: 141-151, 1979.
10. Ayusawa, D., Shimizu, K., Koyama, H., Takeishi, K., and Seno, T. Accumulation of DNA strand breaks during thymineless death in thymidylatesynthase-negative mutants of mouse FM3 A cells. J. Biol. Chem., 258:12448-12454, 1983.
11. Hori, T., Ayusawa, D., Shimizu, K., Koyama, H., and Seno, T. Chromosomebreakage induced by thymidylate stress in thymidylate synthase-negativemutants of mouse FM3A cells. Cancer Res., 44: 703-709, 1984.
12. Murakami, Y., Yasuda, H., Miyazawa, H., Hanaoka, F., and Yamada, M.Characterization of a temperature-sensitive mutant of mouse FM3A cellsdefective in DNA replication. Proc. Nati. Acad. Sci. USA, 82: 1761-1765,1985.
13. Tsai. Y.-.I., Hanaoka, F., Nakano, M. M., and Yamada, M. A mammalianDNA" mutant decreasing nuclear DNA polymerase a activity at nonpermis
sive temperature. Biochem. Biophys. Res. Commun., 91:1190-1195, 1979.14. Yasuda, H., Matsumoto, Y., Mita, S., Marunouchi, T., and Yamada, M. A
mouse temperature-sensitive mutant defective in HI histone phosphorylationis defective in deoxyribonucleic acid synthesis and chromosome condensation. Biochemistry, 20:4414-4419, 1981.
15. Murakami, Y., Eki, T., Miyazawa, H., Enomoto, T., Hanaoka, F., andYamada, M. Further characterization of a murine temperature-sensitivemutant, tsFT20 strain, containing heat-labile DNA polymerase a activity.Exp. Cell Res., 163: 135-142, 1986.
16. Eki, T., Murakami, Y., Enomoto, T., Hanaoka, F., and Yamada, M. Characterization of DNA replication at a restrictive temperature in a mouse DNAtemperature-sensitive mutant, tsFT20 strain, containing heat-labile DNApolymerase a activity. J. Biol. Chem., 261:8888-8893, 1986.
17. Nakano, N. Establishment of cell lines in vitro from a mammary ascitestumor of mouse and biological properties of the established lines in a serum-containing medium. Tohoku J. Exp. Med., 88: 69-84, 1966.
18. Perry, P., and Wolff, S. New Giemsa method for the differential staining ofsister chromatids. Nature (Lond.), 251: 156-158, 1974.
19. Hori, T., Ayusawa, D., and Seno, T. Thymidylate stress and sister chromâtidexchanges. In: R. R. Tice and A. Hollaender (eds.), Sister Chromatid Exchanges, pp. 149-159. New York: Plenum Publishing Corporation, 1984.
20. Cleaver, J. E. Correlation between sister chromatid exchange frequencies andreplicón sizes. A model for the mechanism of SCE production. Exp. CellRes., 136: 27-30, 1981.
21. Ishii, Y., and Bender, M. Effects of inhibitors of DNA synthesis on spontaneous and ultraviolet light-induced sister-chromatid exchanges in Chinesehamster cells. Mutât.Res., 79: 19-32, 1980.
22. Kato, H. Mechanisms for sister chromatid exchanges and their relation tothe production of chromosome aberrations. Chromosoma (Beri.), 59: 179-191, 1977.
23. Painter, R. B. A replication model for sister-chromatid exchange. Mutât.Res., 70:337-341, 1980.
24. Evans, H. J. Chromosome aberrations induced by ionizing radiations. Int.Rev. Cytol., 13:221-321, 1962.
25. Savage, J. R. K. Classification and relationships of induced chromosomalstructural changes. J. Med. Genet., 13: 103-122, 1975.
26. Hirschberg, J„and Marcus, M. Isolation by a replica-plating technique ofChinese hamster temperature-sensitive cell cycle mutants. J. Cell. Physiol.,113:159-166,1982.
27. Fujiwara, Y. Effect of cycloheximide on regulatory protein for initiatingDNA replication at the nuclear membrane. Cancer Res., 32: 2089-2095,1972.
5169
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

ANALYSIS OF CHROMOSOME INSTABILITY IN tsFT20 CELLS
28. Hori, T., and Lark, K. G. Effect of puromycin on DNA replication in Chinesehamster cells. J. Mol. Biol., 77: 391-404, 1973.
29. Liu, P. K., Chang, C.-C, Trosko, J. E., Dube, D. K., Martin, G. M., andLoeb, L. A. Mammalian mutator mutant with an aphidicolin-resistant DNApolymerase a. Proc. Nati. Acad. Sci. LISA, 80: 797-801, 1983.
30. Nishimura, M., Yasuda, H.. Ikegami, S., Ohashi, M., and Yamada, M.Aphidicolin resistant mutant of which DNA polymerase a is induced by thisdrug. Biochem. Biophys. Res. Commun., 91: 939-945. 1979.
31. Sugino, A., and Nakayama, K. DNA polymerase a mutants from a Drosophilamelanogaster cell line. Proc. Nati. Acad. Sci. USA, 70: 245-248, 1980.
32. Koyama, H., Ayusawa. D., Tsuji, M., and Seno. T. Thymineless death andmutation induction in cultured mouse FM3A cell mutants deficient in thy-midylate synthase. Mutât.Res., 105:433-438, 1982.
33. Shimke, R. T. Gene amplification in cultured animal cells. Cell, 37: 705-713, 1984.
5170
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

1987;47:5162-5170. Cancer Res Toshihiko Eki, Takemi Enomoto, Yasufumi Murakami, et al.
αHeat-labile DNA Polymerase Temperature-sensitive Mutant tsFT20 Strain ContainingIncubation at a Restrictive Temperature in the Mouse Characterization of Chromosome Aberrations Induced by
Updated version
http://cancerres.aacrjournals.org/content/47/19/5162
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/47/19/5162To request permission to re-use all or part of this article, use this link
on March 28, 2019. © 1987 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from