characteristics of the iitterspbcific...
TRANSCRIPT

JORDANKA GEORGIEVA-TODOR,OVA
Institute of Genetics, Sofia, Bulgaria
INTRODUCTION
The hexaploid species of Helianthus are ofinterest in the hybridization of cultivated sun-flower as source of genes determining impor-tant biological features such as resistance, the,content of certain valuable fatty acids, proteinetc. On the other hand, the cytogenetic studyof hybrids may contribute to explain theorigin of the hexapioid species.
The object of such hybridization has so farbeen the hexaploid species H. tuberosus andH. rigidus, this species manifesting completeincompatibility with H. annuus (G e o r g i e v a-T o d o r o v a, 1976). In this connection it isinteresting to study the possibility of hybri-dization of cultivated sunflower with H. resi-nosus.
This paper presents the first results of thecytogenetic study of hybrids between the twospecies.
MATER,IALS AND METHODS
The mother plants used were from our cyto-plasmic male sterile form (Georgieva-To d o r o v a, 1976). The pollinator was a popu-lation of H. resinosus, and in some cases polli-nation was carried out by a mixture of pollenfrom H. resinosuslpollen from species withrlifferent levels of ploidy : H. nuttallii, H. argi-rophyllus (2n:34), H. rigidus (2n:102) etc.
The cytogenetic study of hybrids embracesthe characterization of their most importantmorphological characters, pollen viability, seedset as well as meiosis in the initiai spec,iesand hybrids (degree of conjugation on thebasis of the type of the chromosome associa-tion in diakinesis, the character of the indi-vidual meiotic phases, etc.).
CYTOGENETICALCHARACTERISTICS
OF THE IITTERSPBCIFIC HYBRIDSHelianthus annuus L. x
Helianthus resinosus Small
RESULTS AND DISCUSSION
H. resinosus (2n: 102) is a perennial speciesincluded in section Gigantei. The species iswell differentiated, with strictly manifestedroproductive barriers, despite the faet that itis to be encountered in the same areas às manyother wild species of Heli.anthus. Under ourconditions, develops quite well. The roots arelong, some upwaid, becoming woody ; rhiso-mes short, thick to slender ; stems stout, 1-3m tall (H e i s e r, et al., 1967). .The leavesmostly alternate or at least alternate above,from narrowly lanceolate to broadly ovate ;disks 2-3 cm in diameter. H. resinosus is resis-tant to Plasn"Loper& halstedi,i., Pucciniu heli.anthi,etc. The data obtained from a study of the ga-mete compatibility and of the possibilities ofobtaining developed seeds in Fs (Table 1) in-dicate that the number of seeds in one inflo-rescence is low ; it usually varies between 3and 100/s when only H. resinosus participatesas pollinator and between 10 and 200/6 whenpollination is done with mixed pollen.
Table 7
Crossability of Helianthus annu,lts (2n:34) withHeli"anthus resinosus (2n:102)
Number ofseeds perinflores-
cence(%)
0-3
withH. re-sino-szs+pollen
ofwildspe-cles
Number of seedsper inflorescence
(%)
,-,0
I
,,-*
It is known that crossability is improvedwhen pollen from species of the same generais added to the male parent (Georgieva,1976). The number of seeds in F6 of ttre crossH. annuusxH. resinosus increased following
23
the addition of polien from the species H' nut-tatti.i (2n:34), H' argophgllus (2n: 34) andH. rigidus (2n- 102). It is difficrlt to us toexptain the reason of improving qle seed sett-ing in F0 (in certain cases -
200/6 when hy-bridization was carried out with mixed pollen)'It may be assumed that pollen of differentspecieÀ contains different enzymes' some of*tti"tt are favourable to the germination ofpollen from H. resinosus, as w-ell as to thegrowth of the pollen tubes and fertilization ofà htger number of egg-cells. It should be men-tioneâ in this respect that there exists mar-kedly expressed interspecific incompatibilitybetween H. annuus and the species referred toabove (G e o r g i ev a -T o d o r ov a, 1976)'
The results of the development of the F1
hybrid plants are presented in Table 2, whichrÈow. that hybrids were obtained in almostall progenies,- with the exception of one,but none of them have shown complete germi-nation of the seeds. For instance in progeny57 /82 from 550 sown seeds germinated 86
i.e.15.640/s, and only 8.73010 of the total numberdeveloped completely. In some progenies like56/82 neither of the seeds germinated' Thisshows, as we have established in other hybri-dcombinations, that the interspecific incompati-bility in the crossing qf gglgtically distantfornis may be manifested at different stages ofontogenesis
Table 2
Besults of the investigations of the seed germination ofH. snnuus X H. tesino'sus and the developmcnt of
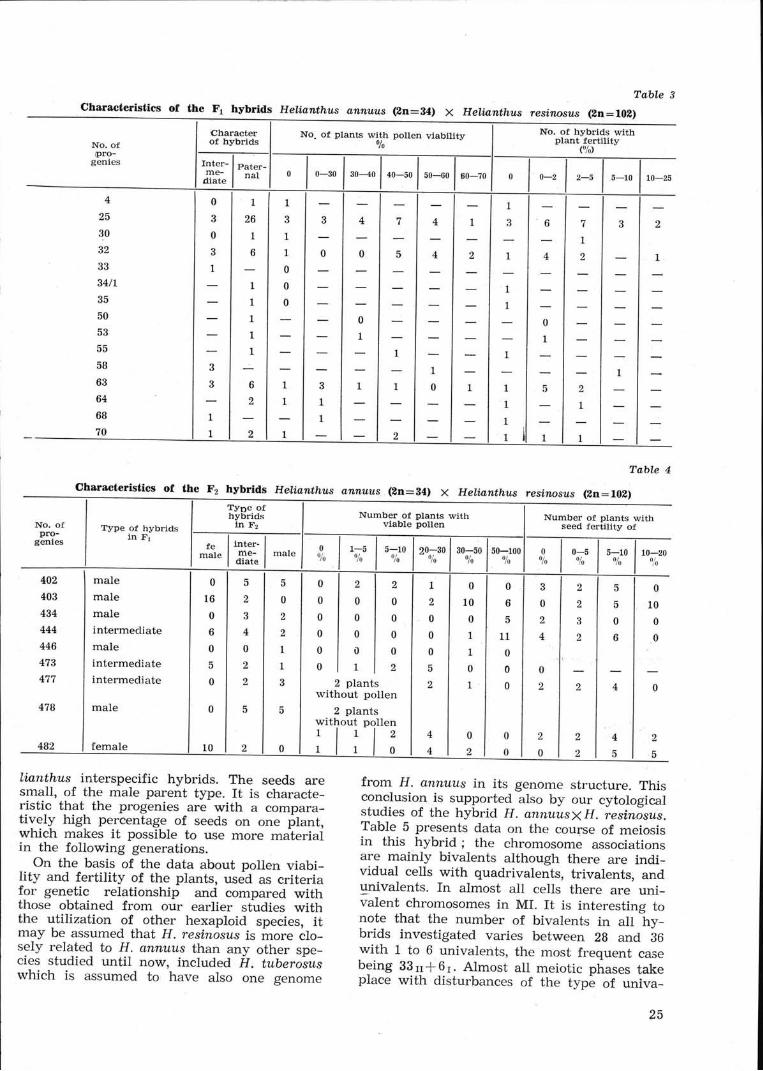
The F1 hybrids proved to be phenotypicaiiyinteresting (Table 3). It shouid be pointed outthat the first hybrids from this combinationwere of an intermediary type, though pheno-typically much more similar to the cultivatedparent with shorter stems lightly ramified'iipening 10-15 days earlier, but with reducedviability of pollen and lower plant fertility'They were iesistant to Plasmoçtara halsteliiunder field conditions.
Almost all hybrid progenies in 1982 and 1983were phenotyficaliy entirely of the type ofH. resinosu.s. Àmong the numerous interspecifichybrids of Heliuntlnls, which we have obtained,rtttil no*, such a high percentage of hybridsmanifesting the type of the wild parent(S0.33%) hàs not been obtained in any of thecombinàtions in which the cultivated specieswas used as mother. Since there is no infor-mation in the literature about hybrids obtainedbetween these species, it is not possible to ex-plain the reason for the stronger phenotypemanifestation of the wild parent. However asimilar phenomenon, though not so marked,has been observed in the hybridization ofH. annuus with another hexaploid species(H. tuberosus) and with the tetraploid speciesH. clecapetalus and H.hirsutus. It is interestingto note that this phenomenon is observed inthe combination in which the wild parent ishighly polyploid and participate as father. Toall piobability, the number of the conjugatedhomologous chromosomes from the two typesis siqnificant at the recombination of the ge-netic material of the higher-ploid parent withthat of the lower-ploid one, something whichis reflected also in the phenotype of the hy-brids. The viability of pollen was strongly le-ducecl in all hybrid plants, and 14.4rf0 of thehybrids had totally sterile pollen. The highestis tfre percentage of plants with pollen viabi-hty belween 40 and 500/0. The fertility of thehybrids is likewise low but these hybrids areoi highe. fertility, compared with the hybridsobtained by us between other species.
The hybrids keep their phenotype in F2 (theplants fiom the wild parent prevail over theôthers). In Table 4 we show some data for thephenotype variety of the hybrids in F2. Thetable gives an idea of the type of inheritanceas well as of the fertility of the plants. Thephenotype segregation of different progenies arenot the same. The parent type dominatès inF2 in most of them. In some progenies as foreiample in 403/83 from the F1 plants of thepaternal type in F2 we have observed plarrtsàf mother type (from 18 plants
- 16 of mothertype and 2 of intermediate type) or in the F1
progenies of the intermediate type in F2, therehave been observed plants of the three types:mother, intermediate and paternal. Pollen via-bility of hybrids varies in F2, but as a rule'the plants with higher pollen viability domi-nate. The fertiiity is reduced as in other He-
YearNo. ofproge-
niesSeedssown
Hybrid PlantsdeveloPed
----|-No. I oilr
143I
6
14.60
13.79
2t.8250.0018.75
29.7r12.50
52.6316.67
40.000
8.7320.00
4.557.50
16.67
37.93A 1n
100.00
25.82
1981 65
65a65b2532
48
49
50tl52
53
54
55
56
tr58
59
60
65
66
68
69
?0
500
2
t375B
275B
32
138
B
19
t25
I550
10
22
120
29
24
2
rB2
1461
6
29.20
50.0085.?1
28.6050.0085.71
20
10
61
5a
43
I10
4
2
0
B6
2
611
I
11A
2
4B
20a
60
4
6.11
1
10
2
0
4B
2
I
o
^
11I
2
47
14.60
L7.2:l22.18
62.5021.8831.1612.50
52.63
35.3340.00
0
15.64
20.0027.279.t7
29.17
37.93
16.67
100.00
26.:ti
.'t ll4a
Characteristics of the F1 hybrids Helianthus &nnul.ts (2n=34; Hel;i,anthus resinosus
Cha,raeteristics of the Fz hybriils Heli,antltus artnurls (2n=34; Hetia,nthws resitwsus
Table 3
(2n:102)
Table 4
(Zn:102)
lianthus interspecific hybrids. The seeds aresmall, of the male parent type. It is characte-risti_c t-hat the progenies are with a compara-tively high percentage of seeds on one plant,which makes it possible to use more materialin the following generations.
On the basis of the data about pollen viabi-lity and fertility of the plants, used as criteriafor genetic relationship and compared withthose obtained from our earlier studies withthe utilization of other hexaploid species, itmay be assumed that H. resinàsus is more clo-sely related to H. annuzs than any other spe-cies studied until now, included F/. tuberoiuswhich is assumed to have also one genome
from I{. annuus in its genome structure. Thisconolusion is supported also by our cytologicalstudies of the hybrid H. annuusyH. resînosus.Table 5 presents data on the course of meiosisin this hybrid ; the chromosome associationsare mainly bivalents although there are indi_vidual cells with quadrivalents, trivalents, and.univalents. fn almost all cells there are uni_valent chromosomes in MI. It is interesting tonote that the number of bivalents in all hy-brids investigated varies between 2g and J6with L to 6 univalents, the most frequent casebeing 331*61. Almost all meiotic phases takeplace with disturbances of the typà of univa-
Charâeterof hybrids
Inter- | pater_lle- | natûrâte I
No. oflpro-
genies
No of plants wittl pollen viabilityo/o
No. of hybrids withplant fertility
(!ol
0 0-30 3o-4ol4o-5ol50-60 60-70 1 0 l0-2 2-5 5-10 10-25
4 l0zs ls30 lo32 l:33 lr34/L | _35 l-50 l-53 l-55 l-5B ls63 le64
I68 lr?0 11
I26
1
6
I
t
1
6
,
2
1
3
1
I0
0
0
1
I
3
0
3
1
t
; l;;l;-l:
t-0l'l I'l-tllt-
- t;
4
4
t
0
I
2
2
I
I
3
r
2
1
Tir:' | '"o',î'*,uoto"genres | -
TyDe ofhybridsln F2
Number ol plants withviable pouen
Number of plants withseed fertility of
female
inter-me-diâte
male 0o/o
1-5a/o
5-10o/o
t0-30- o/o30-50
o/o50-100
u/o0
o/o0-5
Yo
5-10o/o
10-20%
402 | male403 | male434 | male444 | intermediate446 | male473 | intermediate477
| intermediate
478 I
*r"
482 | tu-rte
0
16
0
6
0
D
0
0
10
D
t3
4
0
2
2
5
2
5
0
2
2
1
1
3
5
0
0
0
0l0l0l0l
witt2
with1lI1l
2lilul0lûl.lrl
plantr.out pc
plantrout po1l
1l
2
0
0
0
0
2
lleni
Ilen2
0
1
2
0
0
0
5
2
1
0
10
0
I1
0
0
2
0
6
D
t1
0
0
0
0
0
3
0
4
0
2
2
0
2
t
2
2
2
2
5
D
0
6
^
4
5
0
10
0
0
0
2
5
25

'Table 5
Helianthus resirwsus (2n:102)Meiosis in F1 hybritls Helianthus annuus (2n:34) X
Metaphase I Anaphase I
No. of plânts
25/82-2
25/82-8
25/82-14
Diakinæis chromosomeconfigurâtions
Telophase II I
--'-- t-l Pollen
totâl I distur- lviabilityno. of I bances I e/o)cells | % I
127 64.50
totalno. ofcells
distur-bances
(%)
I'ertilityolo
0.'70
t2.97
15.50
35II +5r34rr +4r33rr +6r33rr +6r33rr +6 r28rr +llv +3r35rr +1r32II +lrrr +5r33rr +2r
32u +4r31II +3r32il +4r36rr
32rr +1nr +5r32rr +5r26rr +5r28rv +3r31rr +1rv +2r31r
31u +2r29rr +1ry +4r
lents in metaphase I, lagging chromosomes, andchromosomes bridges in anaphases I and II'Iagging chromosomes in telophase II etc. Inter-esiing too, is the alreqdy mentioned fact thatthere is a larger percentage of a higher numberof bivalents, this being an indication of theexistence of homology between the chromo-somes of H. resi,nosus and those of H. annuus.
CONCLUSION
Hybridization between H. annuus (2n: 34)and H. resi.nosus (2n:102) is successful. Thehybrids, obtained for the first time, indicatethat it is possible te use this species with aview to transferring to sunflower the geneticmaterial conditioning the resistance to diseases.The cytological study has made it possible todrâw certain conclusions about the homologyof the chromosomes of the two species.
REFERENCES
Georgieva-Todorova J., 1974, Proceed'ings oIth,e 6th Inter. Suntl. Conf., Bucharest - Roma-nia, p. 343-349.
Georgieva-Todorova J., 1976, Mezhduoi.douiotnosherna o roil' Helianthus, M' Izd: BAN.
Georgieva R., 19?6, M. Izd. BAN.Heiser C. 8., Smith D. M., Clevanger S.,
M a r t i n rvV. C., 1969, ?he North Arnericon Sun'' ' llowers (Helianthus), Mem. Torrey Club, 22'
26
CARACTÉRISTIQUES CYTOGÉNÉTIQUES DES' HYBRIDES INTERSPÉCIFIQUES HELIANTHUS
AIV]VUUS IL.XHELIAN"HUS RESINOSUS SMALL
Résumé
On a étudié et décrit I espèce sâuvage H' resinosus(2n:102). Celle-ci est une source de résistance à I'at-taque de plusieurs pathogènes du tournesol, parmi le-s-quèh plostnopara halstectrii et Puccinia helianth'i. I-enombre des semences hybrides (Fo) a considérable-ment augmenté quand le pollen de cette espèle a- étémélangéâvec le pollen d'autres espèces de Helianth'us'
L'incompâtibilité interspécifique entre If. annltrus et'H. resinosus s'est manifestée lors de différents stadesde I'ontogénèse. Les hybrides F1 ont été phénotypique-ment différents, plus de 800/6 des cas étaient sem-blables èt H. resinôsus, surtout quand ceci a été utilisécomme géniteur mâle.
La viabilité du pollen des plantes F1 a été trèsréduite, et une stérilité totale du pollen a été enre-gistrée chez L4,4ofn des plantes. Quoique la fertilitéàes plantes F1 a été basse, elle est bien plus élevéeque èelle des hybrides résultés du croisement d'autreseipèces de Helianthu.s. Les types parentaux ont pré-dominé dans la génération F2.
Conformément aux données sur la viabilité du pol-len et Ia fertitité des plantes, on peut affirmer queH. resinosus est plus apparenté à H' amnu,us que d'au-tres espèces hexàploides, comme H. tuberosus. La mé-iose dôs hybrides H. annuusxli. resirwsus a indiquéI'existence de l'homologie d'une partie des chromo-somes de l'espèce hexaploide et 'ceux de H. anrwuss
L'hybridation entre les deux espèces étant relative-ment facile à réaliser, il est possible de transférer dumatériel génétique important, de I'espèce sauvage àcelle cultivée.
The numberof non included
univalents in MI
2, s, 4,15, 6. 7

CARACTERISTICAS CITOGENETICASDE IJOS HIBRIDOS INTERSPECIFICOS
HELIANTHUS âNNUUS L.XHELIANTHU SRESINOSUS
Res{tmen
Estâ descrita la especie H. resinosus (2n - l0Z).Ésta es una fuente de resistencia al ataque de variasenfermedades de girasol, entre las cuales plasrnoparaholsteilii y Puccinia helianthi,. El nrimero de semillashfbridas (Fo) aunient6 considerablemente al mezclardel polen de esta especie con polen de otras especiesde Helianthus.
La incompatibilidad interespecifica entre H. annuusy H. resinosus se manifestô en diferentes estadios deontogenésis. Los hibridos F1 fueron diferentes feno_tipican:rente, màs del B0 por ciento de los casos se
parecieron al H. resi,nosus, sobre todo .cuando éste seempleô como pariente paterno.La viabilidad del ..polen de las plantas F1, fue muyreducrda, y en 14,4u/s de los {ndividuos se ha seùa-Iado una esterilidad total ÈrI polen. A pesar de quela,fertilidad de las plantas F1 fïe Uaja, éità es muchomâs alta que la de los hiËridos iesultados por eI
cruce- de -otras especies_de Heliamth,us. Los tipôs pa-rentales dominaron en la generaciôn F2.
Partiendo de los datos referentes â la viabilidaddel polen y la fertilidad de ilas plantas ,se puede afir_mar que H. resinosus estâ ernparentadâ mâs conH_. ennuus que otras especies îrexaplôiàes, comoH, tuberosus. La rneiosis de los hibridàs H.'onnuu"xH. resinosus indicô la existencia de la homôlogia entreuna parte de los cromosomas de la especié hexa_ploida y los de Il. annulr*
La hibridacidn entre las dos especies siendo rela_tivamente fâcil de realizarse, es posible transferarmater?l ge!étic,o importante desde -la
especie salvajehasta la cultivada.