chapter 2 properties of nucleic acids. 2.1 nucleic acid structure (dna & rna) : bases ...
TRANSCRIPT
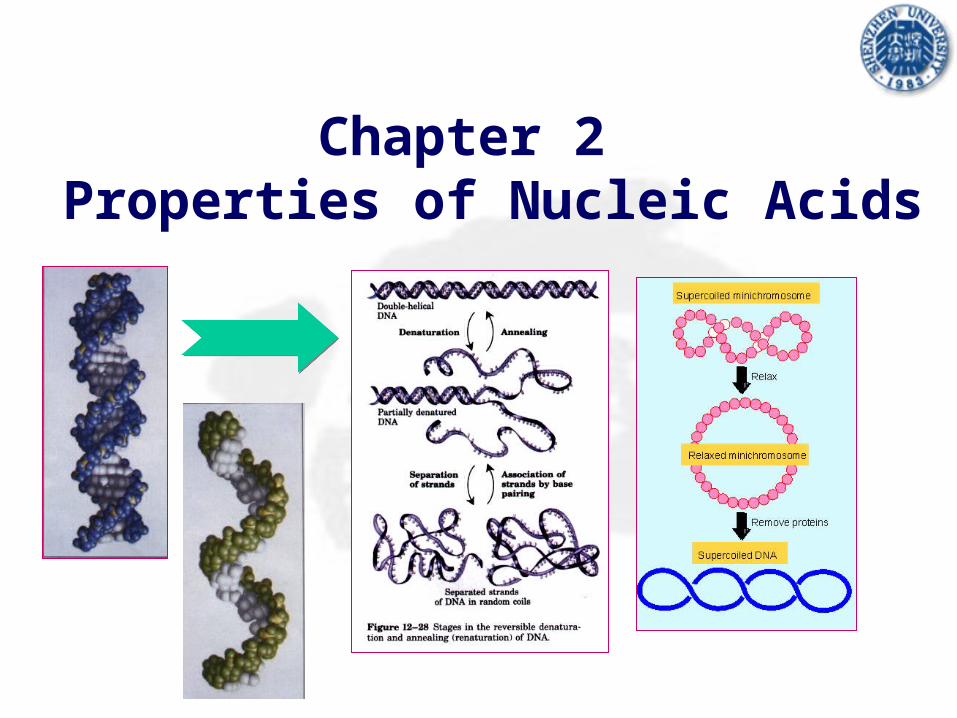
Chapter 2 Properties of Nucleic Acids

2.1 Nucleic Acid Structure (DNA & RNA): basesnucleosides nucleotide phosphodiester bonds primary sequence structure, modified nucleic acids
2.2 Chemical and Physical Properties of Nucleic Acids (DNA & RNA): stability (force), chemical properties (acid, alkali, chemical denaturation), physical properties (viscosity, buoyant density)
2.3 Spectroscopic and Thermal Properties of Nucleic Acids (DNA & RNA): UV absorption, hyperchromocity, quantitation and purity
2.4 DNA Supercoiling (DNA): closed circular molecule, supercoiling & energy, topoisomer & topoisomerase

Bicyclic Purines:
Thymine (T) is a 5-methyluracil (U)
2.1 Nucleic Acid Structure (DNA & RNA): Bases
Monocyclic pyrimidine:
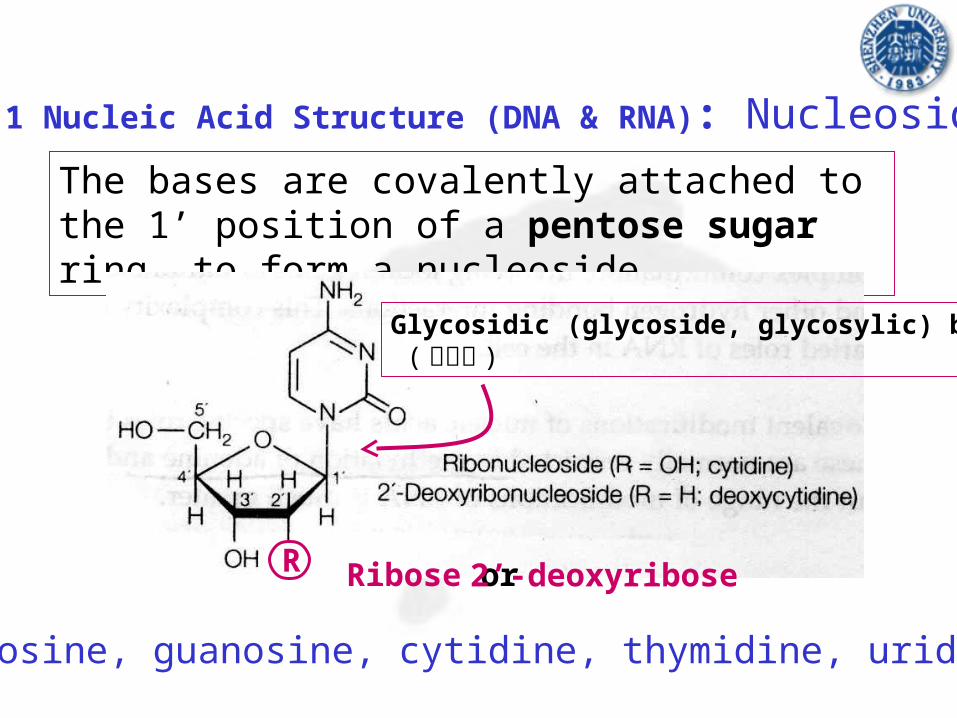
The bases are covalently attached to the 1’ position of a pentose sugar ring, to form a nucleoside
Adenosine, guanosine, cytidine, thymidine, uridine
Glycosidic (glycoside, glycosylic) bond ( 糖苷键 )
R Ribose or 2’-deoxyribose
2.1 Nucleic Acid Structure (DNA & RNA): Nucleosides
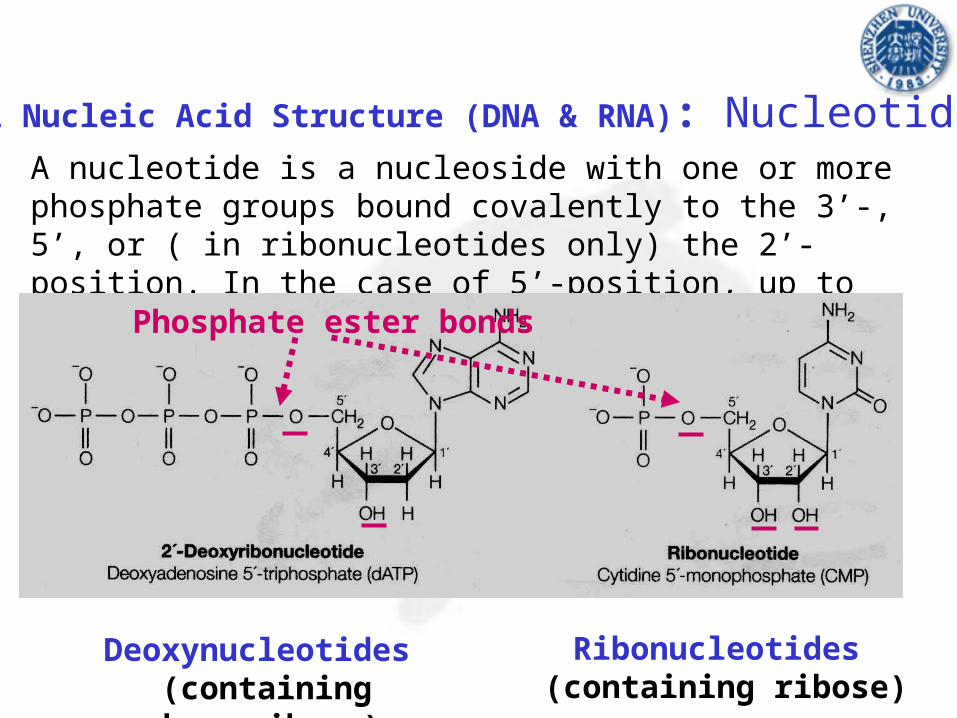
A nucleotide is a nucleoside with one or more phosphate groups bound covalently to the 3’-, 5’, or ( in ribonucleotides only) the 2’-position. In the case of 5’-position, up to three phosphates may be attached.
Deoxynucleotides (containing deoxyribose)
Ribonucleotides (containing ribose)
Phosphate ester bonds
2.1 Nucleic Acid Structure (DNA & RNA): Nucleotides
BASES NUCLEOSIDES
NUCLEOTIDES
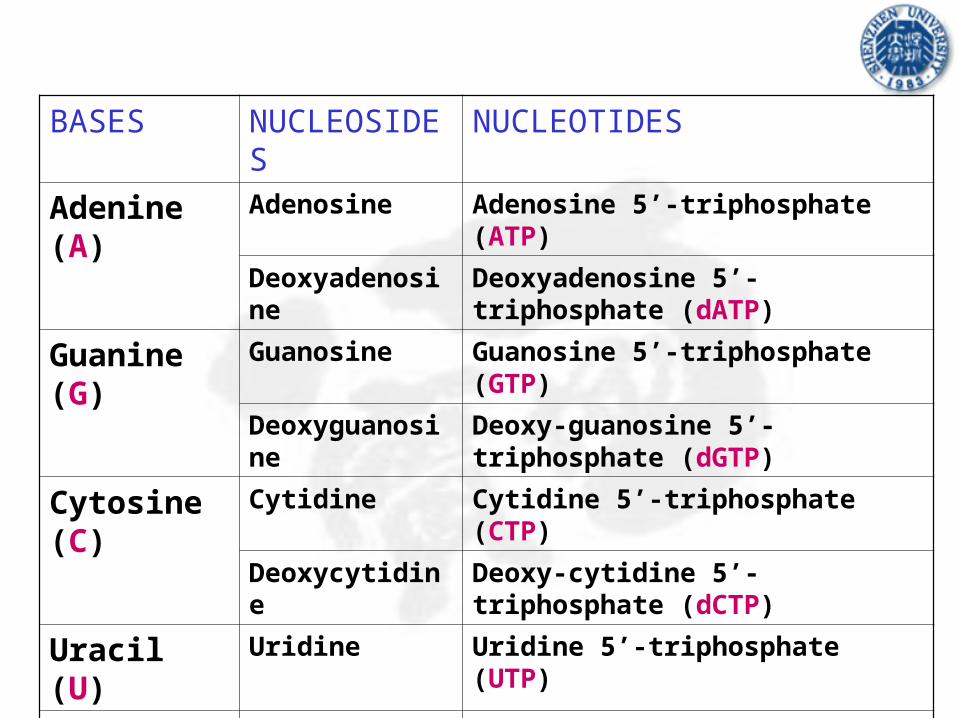
Adenine (A) Adenosine Adenosine 5’-triphosphate (ATP)
Deoxyadenosine Deoxyadenosine 5’-triphosphate (dATP)
Guanine (G) Guanosine Guanosine 5’-triphosphate (GTP)
Deoxyguanosine Deoxy-guanosine 5’-triphosphate (dGTP)
Cytosine (C) Cytidine Cytidine 5’-triphosphate (CTP)
Deoxycytidine Deoxy-cytidine 5’-triphosphate (dCTP)
Uracil (U) Uridine Uridine 5’-triphosphate (UTP)
Thymine (T) Thymidine/
Deoxythymidie
Thymidine/deoxythymidie
5’-triphosphate (dTTP)

Phosphodiester bond
Primary sequence:
5’end: not always has attached phosphate groups3’ end: free hydroxyl (-OH) group
2.1 Nucleic Acid Structure (DNA &
RNA): Phosphodiester bonds & primary sequence

2.1 Nucleic Acid Structure : DNA double helix
•Watson and Crick , 1953
•The genetic material of all
organisms except for some
viruses
•The foundation of the
molecular biology

•Two separate strands Antiparellel (5’3’ direction)Complementary (sequence)Base pairing: hydrogen bonding that holds two strands together
Essential for replicating DNA and transcribing RNA
5’
3’
3’
5’
• Sugar-phosphate backbones (negatively charged): outside• Planner bases (stack one above the other): inside
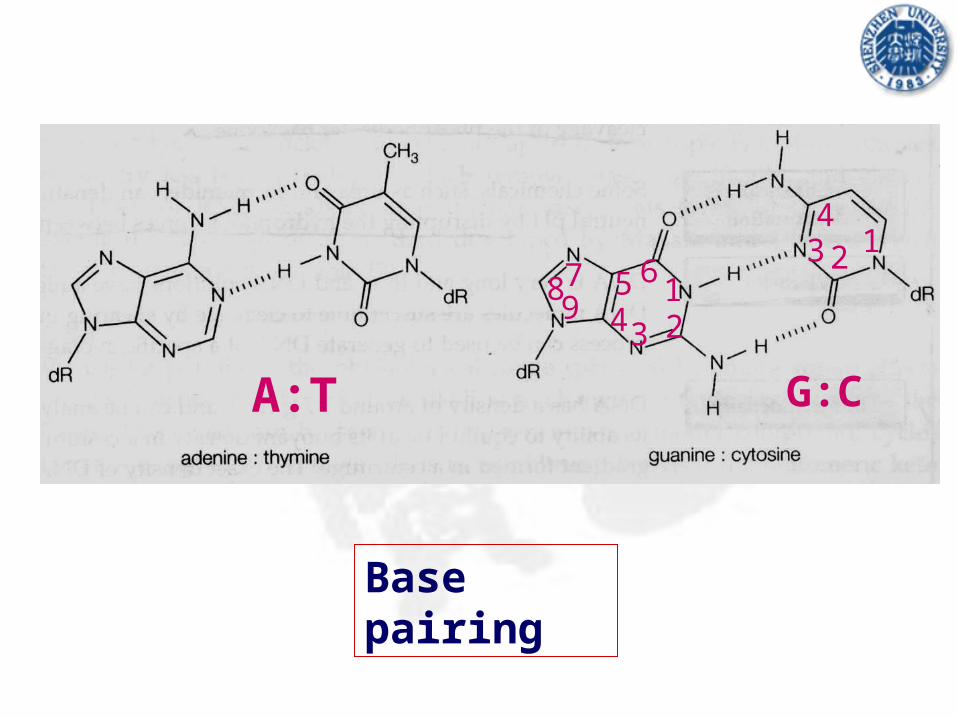
Base pairing
A:T G:C
1234
897 65
4 3 21
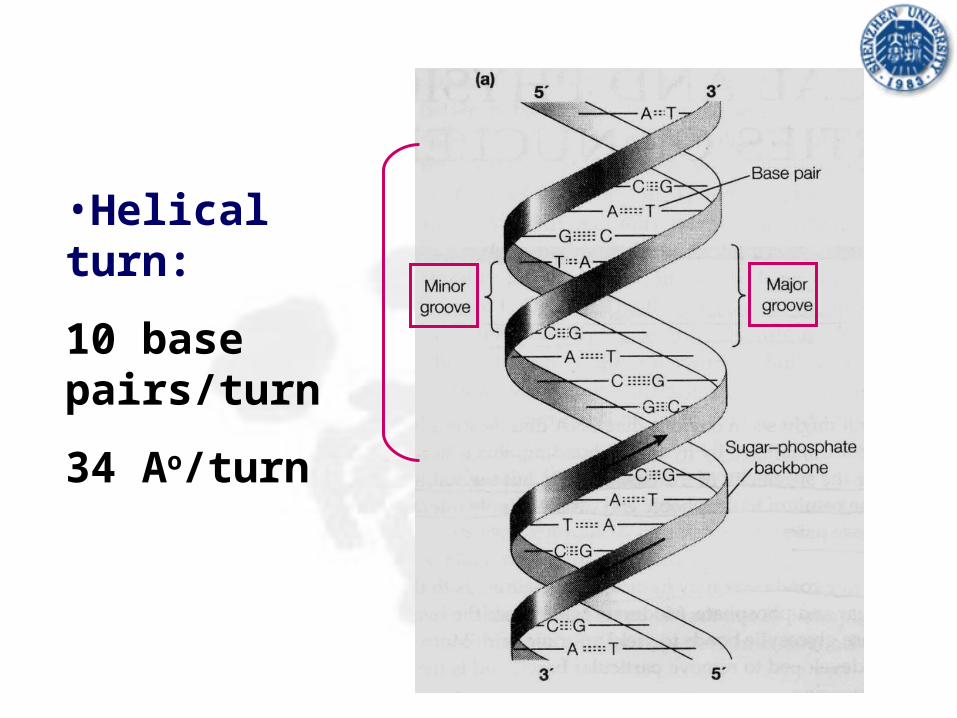
•Helical turn:
10 base pairs/turn
34 Ao/turn

Several forms of DNA…
• Two types of DNATwo types of DNAright-handed helix(right-handed helix( 右手螺旋右手螺旋 ) ) :: A-DNA and A-DNA and
B-DNAB-DNAleft-handed helix (left-handed helix ( 左手螺旋左手螺旋 ) ) :: Z-DNAZ-DNA• DNADNA 的水溶液通常为的水溶液通常为 B-DNAB-DNA ,另外 ,另外 A-TA-T 丰富的丰富的
DNADNA 片段常呈现片段常呈现 B-DNAB-DNA ;;• DNADNA 的双链中一条被相应的双链中一条被相应 RNARNA 链所替换,就会形链所替换,就会形
成成 A-DNAA-DNA 。如,在杂交分子或。如,在杂交分子或 DNADNA 处于转录状态处于转录状态时;时;
• BB 型 型 DNADNA 中的多聚中的多聚 G-CG-C 区易形成左手螺旋区易形成左手螺旋 DNA,DNA,即即 ZZ 型型 DNA DNA


3-14
双螺旋 碱基倾角 碱基距离 螺旋直径 每轮碱基数 螺旋方向
B-DNA 1 0.34 2.37 10.4 右手
A-DNA 19 0.23 2.55 11 右手
Z-DNA 9 0.38 1.84 12 左手

1 在正常细胞的水环境中 DNA 以 B-DNA 为基本构象,脱水或盐浓度升高时,转变为 A-DNA 。
2 细胞内阳离子较多时交替的 GC 区段一般处于 B-DNA ,在嘧啶被甲基化后,转向 Z-DNA ,转录活性降低。
3 某些 Z-DNA 结合蛋白能作为一种特异识别信号,使 B-
DNA 转变为 Z-DNA 。
4 在转录区域 B-DNA 与 RNA 形成 A 型结构。
总之, DNA 的结构是处在动态的变化中的,与其功能相适应。
DNA 结构的动态性与精细结构

3-16
(1) T.S DNA 的发现与证实 1953 年以前 Pauling (Chemist)
提出 T.S DNA 存在的可能性 1953 年 Watson & Crick 提出 D.S DNA model
证明沿大沟存在多余的氢键给体与受体
潜在的专一与 DNA ( 蛋白质 ) 结合的能力
形成 T.S DNA 可能性
三股螺旋 DNA (Trpile Helix DNA, T.S
DNA)

3-17
1957 年 Davis , Felsenfeld , Rich 发现
poly(U) + poly(U) + poly(A) T.S RNA
T.S DNA 的概念
1966 年 Miller & Sobell
实现 RNA + D.S DNA
Triple poly Nt
as Repressor 关闭基因
但由于 D.S DNA 的提出而被忽视
但因证明 LacI 产物为
Repressor 而被忽视
1963 年 Hoogsteen 提出了 DNA 三螺旋结构理论

3-18
1987 年 Mirkin . S . M Nature 330 (495)
证明 plasmid DNA 在 pH= 4.3 的溶液中 ,
有 T.S DNA的存在
● 1987 年 Dervan . Moser Science 238 (645)
合成 S.S DNA + D.S DNA → T.S DNA
• 实现 DNA 的定点切割• 研究 X-ray photograph• 核磁共振 → 结构功能
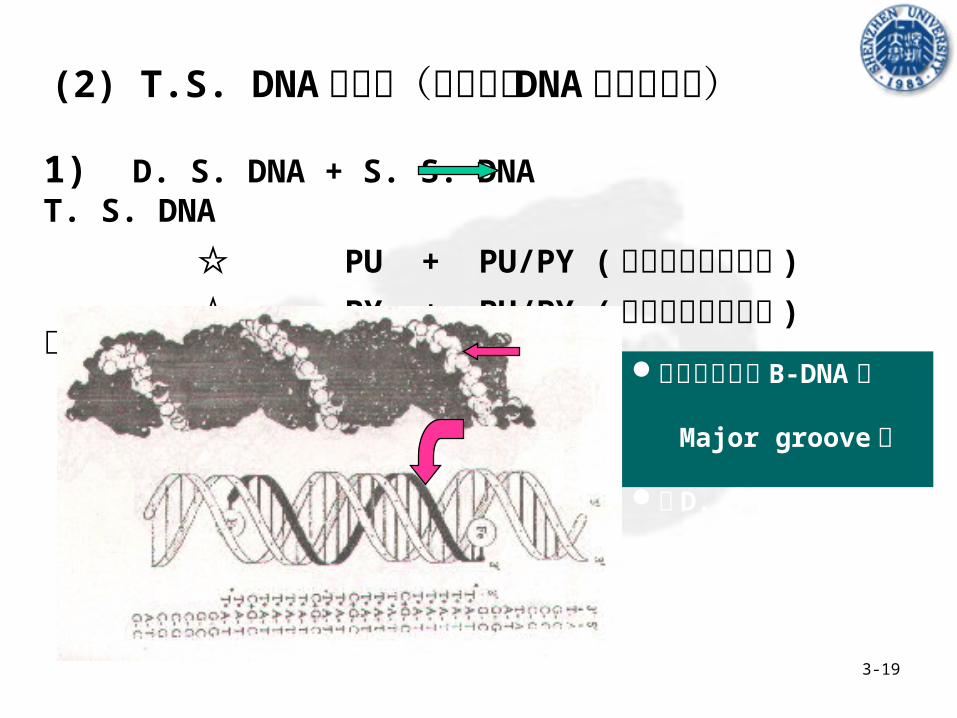
3-19
1) D. S. DNA + S. S. DNA T. S. DNA
☆ PU + PU/PY (偏碱性介质中稳定 )
☆ PY + PU/PY (偏酸性介质中稳定 ) 常见类型
第三条链位于 B-DNA的 Major groove 中
与 D. S. DNA 一起旋转
(2) T.S. DNA 的类型(分子内的 DNA 三螺旋结构)

3-20
PolyT/A TTTTTTTTTTT
AAAAAAAAAA
PolyT/A TTTTTTTTTTT
AAAAAAAAAA
TTTTTTTTTTT
AAAAAAAAAA
TTTTTTTTTTT
AAAAAAAAAA
2 ) D.S. DNA + D.S.DNA
T.S. DNA + S.S. DNA

3-21
a )分子内同源回文( Homologous palindromic sequence ) in a D. S. DNA
(mirror image structure)Mirkin (1987) pGG332 plasmid DNA
-------TTCCCTCTTTCCC------CCCTTTCTCCCTT----
-------AAGGGAGAAAGGG-----GGGAAAGAGGGAA----
--------TTCCCTCTTTCCC--------
--------AAGGGAGAAAGGG-------
-------TTCCCTCTTTCCC---------
--------AAGGGAGAAAGGG--------
Nodule DNA Hinged DNA
free DNA

3-22
三链螺旋
双链螺旋
Hinged DNAHinged DNA
a b
b) 分子间 D. S. DNA + S. S. DNA T. S. DNA
形成结DNA ( Nodule DNA ) or 铰链DNA ( Hinged DNA)
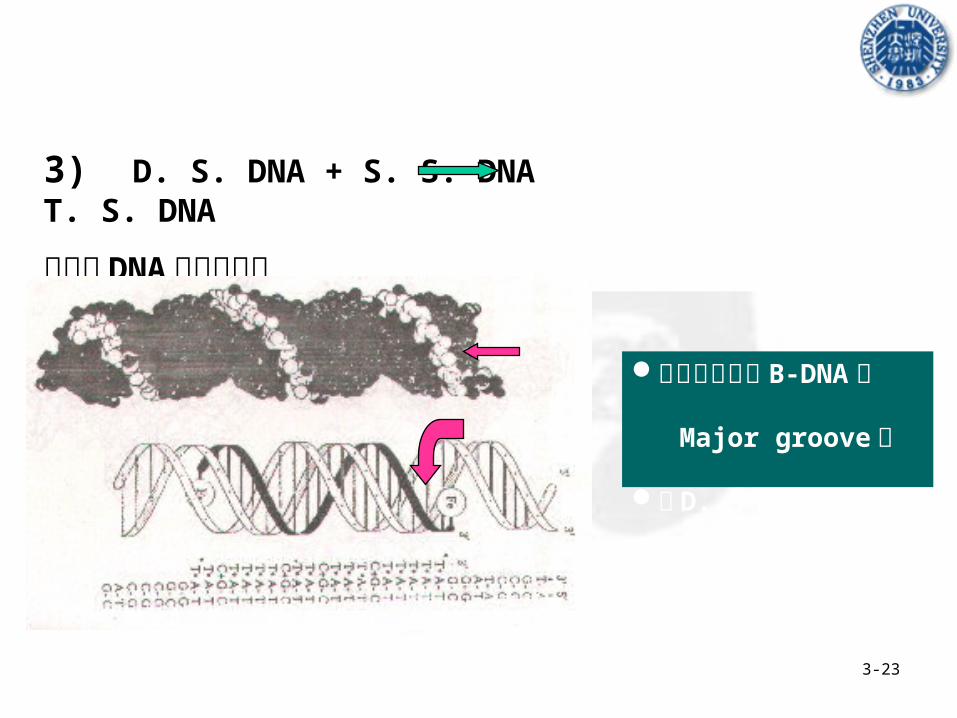
3-23
3) D. S. DNA + S. S. DNA T. S. DNA
平行的 DNA三螺旋结构
第三条链位于 B-DNA的 Major groove 中
与 D. S. DNA 一起旋转

3-24
4 ) T. S. DNA 的连接键 a. Watson bonding A = T G ≡ C (D. S. DNA)
H+
b. Hoogsteen bonding
G = C+ (pH 小于 7)
G = C+ A = A (同向平行,反向平行) T = A G = G ( 同向平行,反向平行)
c. 第三链 质子化 第二链的 purine 6‘,7’ 与第三链的碱基形成 H 键。

3-25
Py Pu * Py Py Pu * Py+
Py Pu * Pu
3-26
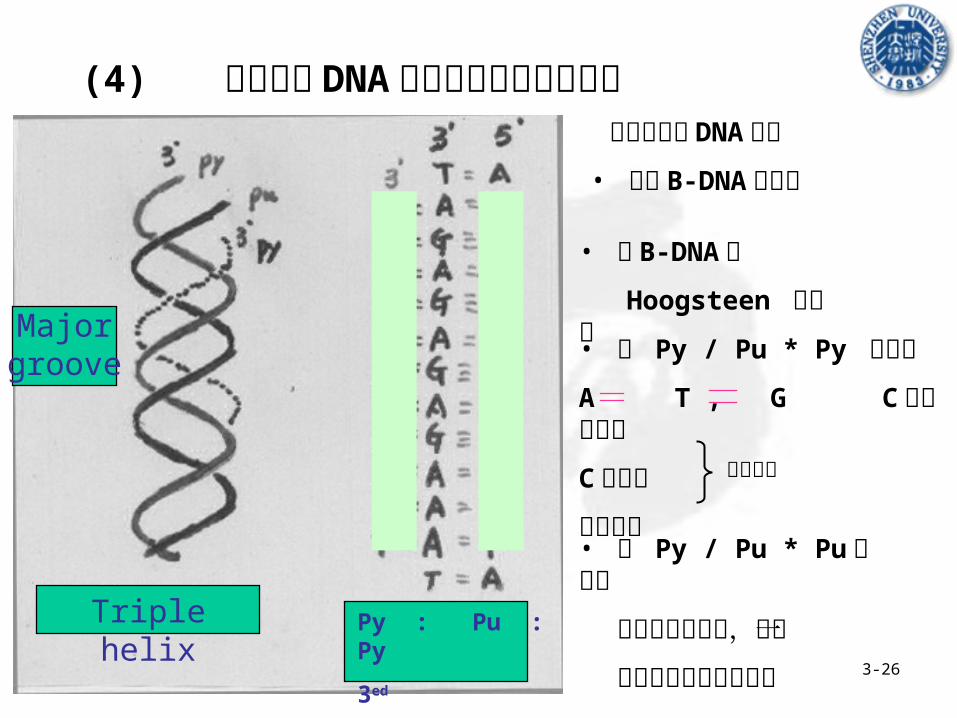
(4) 三股螺旋 DNA 形成的条件及结构特点 第三条单链 DNA 分子
• 位于 B-DNA 大沟内
• 与 B-DNA 以
Hoogsteen 键连接
• 在 Py / Pu * Pu 结构中
有多种配对方式,不一
定需要序列的镜向结构Triple helix
Major groove
Py : Pu : Py
3ed
• 在 Py / Pu * Py 结构中
A T , G C 两氢键配对
C 质子化
镜相结构必需条件

3-27
无论何种形式的三螺旋 DNA, 第二股中间
链必须是 Purine 链 第三股链至少长于 8 dNt
真核生物基因组内 , 约 1% 左右的序列为
大于 100 bp 的 homologous cluster (重复序
列与调控序列 ) , T. S. DNA多存在于其中。

(5) T. S. DNA 可能的功能
a) T. S. DNA 可阻止调节蛋白与 DNA 结合 , 关闭基因转录过程。
b) T. S. DNA 与基因重组交换有关。
c) 加入第三条 S. S. DNA 作为分子剪刀(molecular scissors), 定点切割 DNA 分子。
d) 加入反义的第三条链 (anti-sence polydNt) 终止基因的表达。
3-29
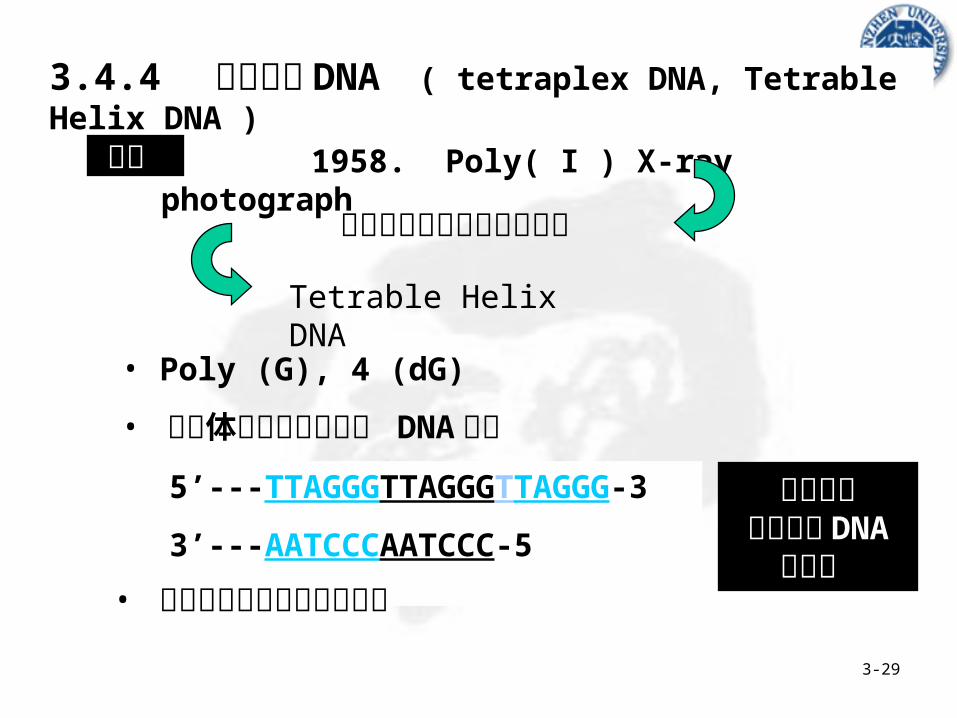
3.4.4 四股螺旋 DNA ( tetraplex DNA, Tetrable Helix DNA )
发现 1958. Poly( I ) X-ray photograph
碱基形成环状氢键连接结构
Tetrable Helix DNA
均有形成四股螺旋 DNA
的可能
5’---TTAGGGTTAGGGTTAGGG-3’
3’---AATCCCAATCCC-5’
• Poly (G), 4 (dG)
• 染色体端粒高度重复的 DNA 序列
• 着丝点附近的高度重复序列
3-30
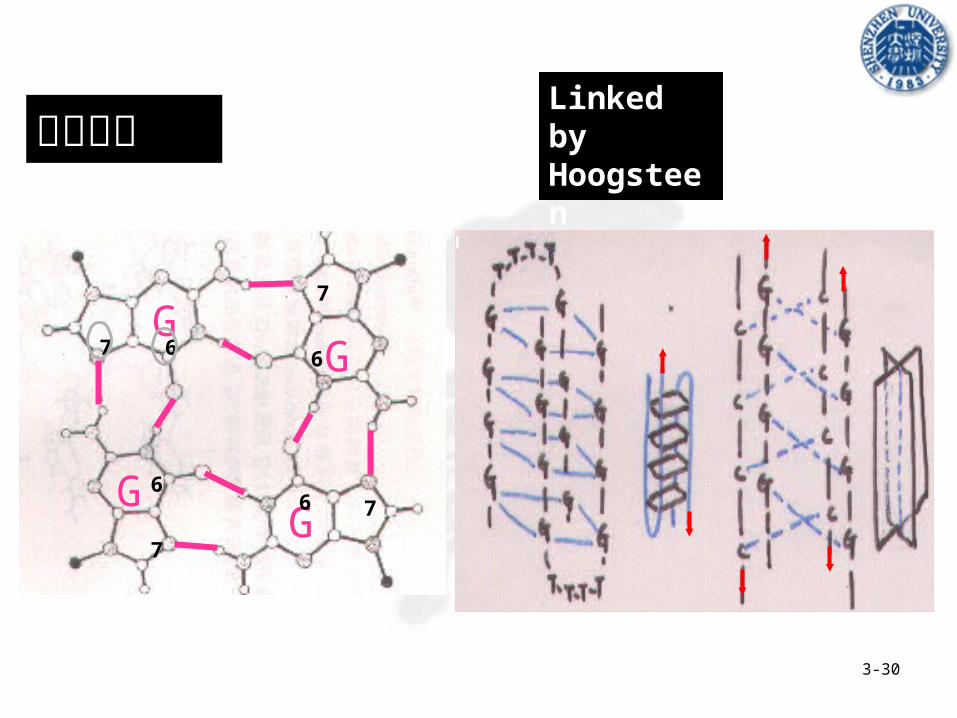
结构特点Linked by Hoogsteen Bonding
7
G
G
G
G
67
67
7
6
6

3-31
G-quartet formation
G-quartets in Vivo ? Telomeres Immunoglobulin switch regions HIV1 RNA Fragile X repeat Ribosomal DNA

3-32
G-rich oligonucleotides or GROs : A new Cancer treatment

3-33
3-34
5‘
3‘
T
T T
GG
G
G
G
G
G
G GG
G
GGG
G
G
G
G
T
TT
T
TT
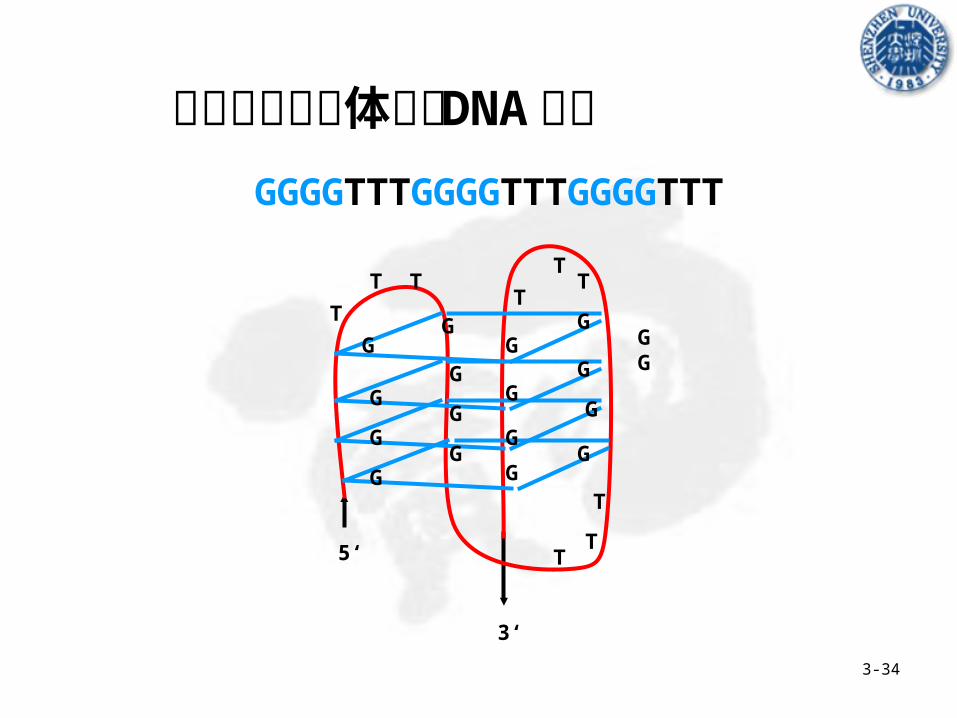
GGGGTTTGGGGTTTGGGGTTT
真核生物染色体端粒 DNA 结构
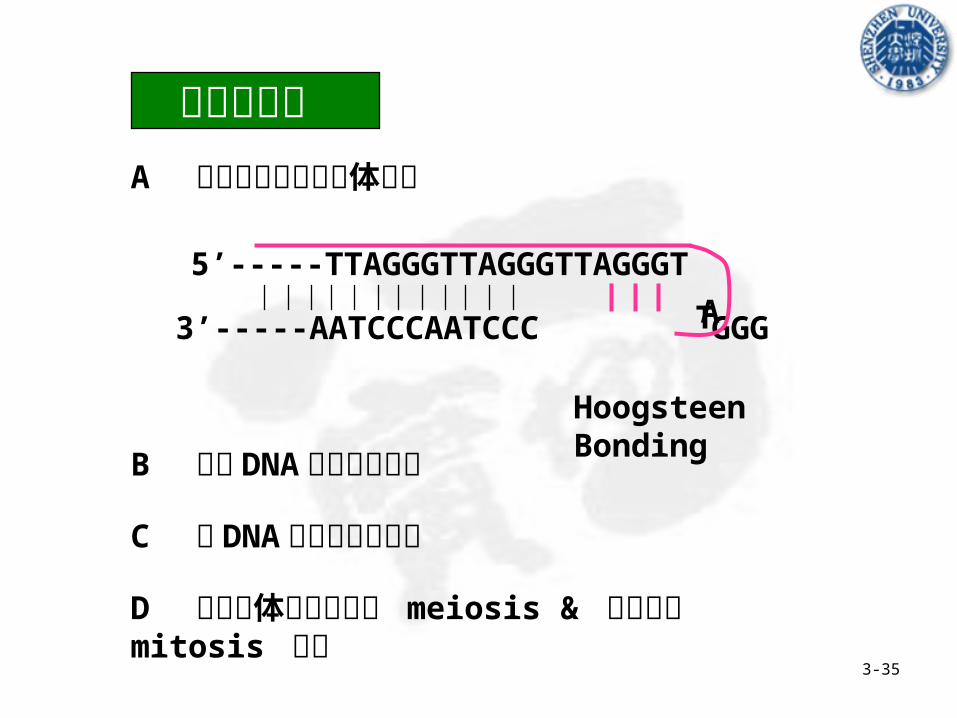
3-35
可能的功能 A 稳定真核生物染色体结构
B 保证 DNA 末端准确复制
C 与 DNA 分子的组装有关
D 与染色体的减数分裂 meiosis & 有丝分裂 mitosis 有关
Hoogsteen Bonding
5’-----TTAGGGTTAGGGTTAGGGT
3’-----AATCCCAATCCC GGG
TA

1. Single stranded nucleic acid2. Secondary structure are formed
some time3. Globular tertiary structure are
important for many functional RNAs, such as tRNA, rRNA and ribozyme RNA
Forces for secondary and tertiary structure: intramolecular hydrogen bonding and base stacking.
2.1 Nucleic Acid Structure : RNA structure

Ribozyme RNA
Tertiary structure
tRNA Secondary structure

Conformational variability of RNA is important for the much more diverse roles of RNA in the cell, when compared to DNA.
Structure and Function correspondence of protein and nucleic acids
Protein Nucleic Acids
Fibrous protein Globular protein
Helical DNA
Globular RNA
Structural proteins
•Enzymes,
•antibodies,
•receptors etc
Genetic information maintenance
•Ribozymes
•Transfer RNA (tRNA)
•Signal recognition etc.

RNA 结构特点及与 DNA 的区别• 碱基组成不同;• RNA 分子中有许多稀有碱基;• RNA 分子中的戊糖是 D -核糖,而 DNA 不是;• RNA 分子是多聚核苷酸单链;• RNA 分子中的碱基不严格遵守 Chargaff 法则;• RNA 分子在碱性溶液中敏感,易水解;• RNA 分子内只有部分双链区域;• RNA 分子是遗传信息的传递体;• 某些 RNA 病毒,是以 RNA 分子作为遗传信息的载体
;• 核酶 RNA 分子具有催化功能;

细胞内 RNA 的分布• 成熟的 RNA 主要分布在细胞质中。• 主要分为三大类:转运 RNA ( tRNA );信
使 RNA ( mRNA );核蛋白体RNA ( rRNA )。
• 细胞核内的 RNA ( nRNA ),另一部分是核内小 RNA ( snRNA )核不均一RNA ( hnRNA ),转移-信使RNA ( tmRNA ) siRNA , micRNA 。

细胞中 RNA 的分布细胞内总RNA
非编码 RNA占总量的 94%
编码 RNA占总量的 4%
hnRNA
前 rRNA tmRNA snRNA snoRNA scRNA非编码rRNA 前 tRNA
rRNA tRNA mRNA
所有生物 仅真核生物 仅细菌

RNA 的分类概述1 、 mRNA 的结构• mRNA 存在于细胞质,总量不到细胞总 RNA
的 5 %;• 真核细胞 mRNA 是单顺反子 (monocistron) ;• 原核细胞是多顺反子 (poly-cistron) ;
m7G5’ppp5’Nm(Nm)
AUG
UGAUAAUAG AAUAAA
(A)nAAOH3’poly A3’ 非编码区5’ 非编码区5’ 帽
子编码区

● In Prokaryote mRNA
5’-end; 300±Nt leading seq. (A/G-------------AUG)
S.D seq---------------------AUG
poly-cistron
9Nt betterrich A,U,
Shine-Dalgarno seq. (S.D seq) GGAGG
→ G mut. translation go down

● In Eukaryote mono-cistron
5’ m7Gppp--- -----CCACC-----A-3---A1U2G3G4—
leading seq.
核糖体小亚基扫描AUG 的信号序列
至关准确翻译
But mRNA of chloroplast shows similarities to prokaryote
type1; S-D seq. with greater secondary structure in leading Seq.
type2; rich AU with little secondary structure in leading Seq.
poly-cistron

● 微小 RNA, 沉降系数 4 - 4.5s, (74-95 Nt)● tRNA phe, 77Nt 构成的三叶草型 (1964 Holly R.)
● 5 arms & 4 loops
● Nt 多被甲基化修饰 , (含有稀有碱基组成的核苷,如:假尿嘧啶核苷(),二氢尿苷( D ),肌苷( I )等)
2 tRNA 的结构
● tRNA 由 74 - 95 个核苷酸组成,通常为 76 个;
● tRNA 含量较多,约占总细胞总 RNA 的 15 %;

• 1 、含有稀有碱基和稀有核苷酸,达核苷酸总量的 5 - 20 %;
• 2 、 3’ 端含有一个 CCA 序列,是所有 tRNA 接受氨基酰化的位置,它是 tRNA 合成后加上的;
• 3 、所有的 tRNA 分子形成三叶草的二级结构和 L
型构象;• 4 、 tRNA 分子有 5 个臂和 4 个环;
tRNA 的二级结构特征

5 副密码子 Paracodon
由若干Nt 组成,存在于 tRNA 不定位置上
与 AARS侧链基团的分子发生特异的“契合”
成为 tRNA 准确负载氨基酸的机制之一
6 tRNA 的” L” 三维结构与功能
“L” 构型的结构力
二级结构中双链区的碱基堆积力和氢键
二级结构中非双链区在“ L” 结构中,形成氢键结合

---aa accept arm ;
loading aa at 3’ end
---DHU loop;
contact with AARS
---anti-codon loop;
34th is wobble base
---TΨC loop;
contact with 5s rRNA
---extra loop;
classification marker ?
I type ; 3-5 Nt 3/4 tRNA
II type ; 13-21 Nt

“L”结构域的功能---aa accept arm 位于“ L” 的一端,契合于核糖体的肽基 转移酶结合位点 P A, 以利肽键的形成。---anti-codon arm 位于” L” 另一端,与结合在核糖体 小亚基上的 codon of mRNA 配对。
A Paa’-tRNAaa

DHU loopextra loop
TC loop
Anti-co
don a
rm
Amino acid arm
--- TΨC loop & DHU loop
位于“ L” 两臂的交界处,
利于“ L” 结构的稳定
---“L” 结构中碱基堆积力大
使其结构趋于稳定
wobble base
位于“ L” 结构末端
堆积力小
自由度大
使碱基配对摇摆
“L”结构域的功能

3. rRNA 的结构• rRNA 是所有 RNA 中含量最多的一类,占细胞总 RNA 的
80 %以上;与蛋白质结合存在。• 核糖体由 RNA 和蛋白组成,真核细胞中 1 : 1 ,原核细胞
中 2 : 1 。• rRNA 中修饰碱基比 tRNA 少,甲基化约占 2 %。不同生物
来源的 rRNA (类 16s , 23s )一级结构的某些区域具有高度的序列同源性。
• rRNA 分子内有大量的茎环结构,使其具有多种构象,二级结构十分复杂。
• rRNA 的结构在蛋白质合成中,随着 mRNA 和 tRNA 的结合及亚基蛋白分子的组装将发生改变, rRNA 的二级结构始终处于动态的变化中。
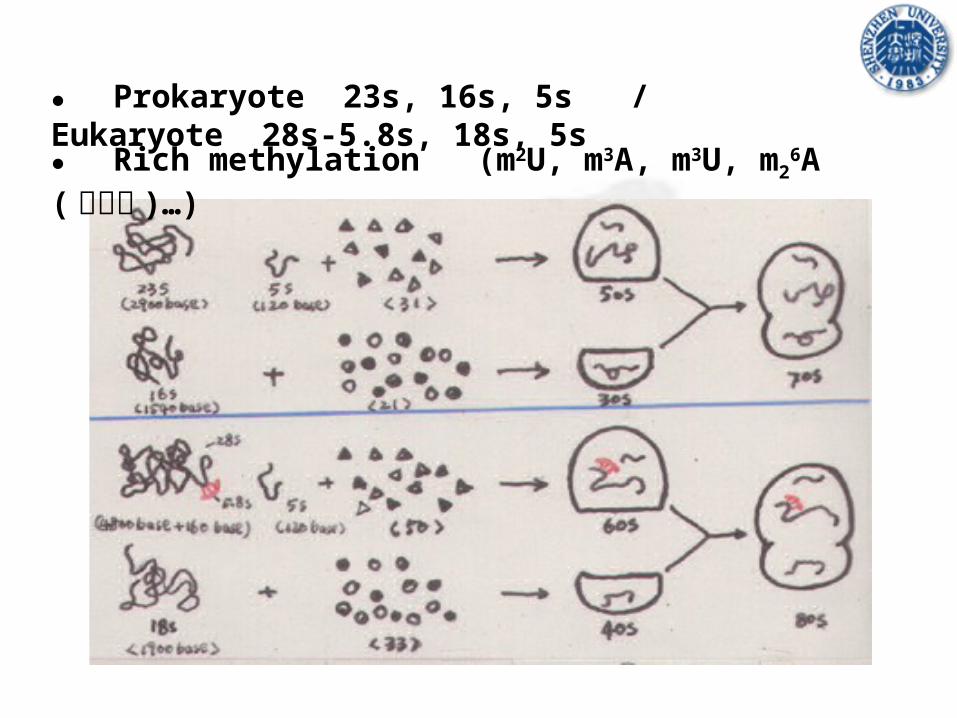
● Prokaryote 23s, 16s, 5s / Eukaryote 28s-5.8s, 18s, 5s
● Rich methylation (m2U, m3A, m3U, m26A( 二甲基 )…)
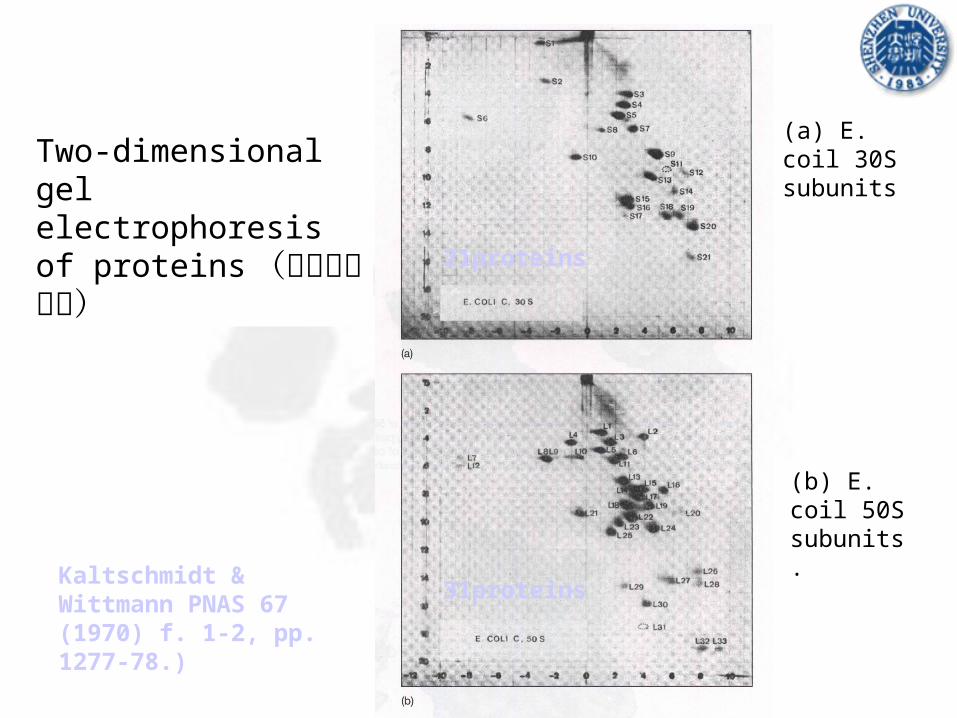
Two-dimensional gel electrophoresis of proteins (蛋白二维电泳)
(a) E. coil 30S subunits
(b) E. coil 50S subunits.
Kaltschmidt & Wittmann PNAS 67 (1970) f. 1-2, pp. 1277-78.)
21proteins
31proteins

● 5s RNA 与 TΨC loop of tRNA部分同源,并可配对
● In Prok. 5s RNA 121nt 与 23s rRNA 组成大亚基
3’-end of 16s rRNA rich CCU conservative seq.complementary with
5’ leading seq. of mRNAShine-Dilgarno seq. of rich AGG
● In Prok. tRNA 核糖体 rRNA , mRNA 之间的相互关系如何?
● In Prok. 16s RNA 1542nt 与大亚基组成核糖体

● 23s rRNA
--- 2904 nt 6 domains
--- 有的与对抗生素的抗性有关
---2660±Nt region α-I loop(alpha Sarcine )
binding with complex of aa-tRNAaa~(EF)-Tu~GTP
G2661 C , aa-tRNAaa into A site go down
---G2252, G2253 双突变为 C, 将对转肽酶的活性产生抑制

● In Euk.
3’-end of 18s rRNA 与原核生物高度相似 ,
但无与 S.D.seq.互补的保守序列
在 mRNA 的 AUG上游存在 CCACC 核糖体 scanning seq 成为核糖体识别第一个 AUG 的信号
AMEAMECCUGCGGUUGGAUGACCUCCUU
AMEAMECCUGCGGAAGGAUGAUUA
16SBacterial
18srRNAMammalian
高度相似
Shine-Dalgarno seq. (S.D seq) GGAGG

● In Euk.
大亚基由 28s rRNA 4718nt , 5.8s rRNA
158nt 5s rRNA 120nt 与 45 种蛋白构成;
小亚基由 18s rRNA 1874nt rRNA 与 30 种蛋白构成。
特点:修饰碱基比 tRNA 少;
二级结构复杂;
5sRNA 一级结构序列比其它 rRNA 保守性高 ;
rRNA 的构象处于不断的动态变化中;

● More hairpin structue

• 真核细胞核内特有 snRNA, 存在于细胞核内或核质及核仁中,在基因转录加工过程中具有重要作用;
• snRNA 含有 70 - 300nt ,不是任何 RNA前体;
• snRNA 有 20 多种其中 13 中富含 U ,占分子内的 35 %。 U1 , U2 , U3 在真核细胞转录中对 hnRNA 的加工是必需的;
• snRNA 通常是与蛋白质结合形成核蛋白体( snRNP )复合物发挥作用;
4. snRNA

• 核仁小分子 RNA广泛分布于酵母和真核细胞的核仁区;
• 哺乳动物细胞的 snoRNA 以 U命名,酵母的以 snR命名
• 根据结构比较,将其分为两类:① boxC/D 型: 5’端含有 boxC(UGAUGA) 序列, 3’ 端有 boxD (CUGA), 在 boxD上游有 8 - 14nt 与 rRNA 内的保守核心序列互补;② boxH/ACA 型:其 3’ 端第三个核苷酸上游有一个保守的 ACA 或类似序列AGA , AUA 。整个分子形成两个发卡结构中间是保守的 boxH(ANANNA) 的铰链区。
5. snoRNA ( small nucleolar RNA)

• 功能:1. 作为 rRNA前体加工复合体的重要组分
,参与 rRNA前体的加工。2.参与 rRNA2’-O- 核糖的甲基化修饰。3.参与部分 snRNA tRNA 的甲基化修饰。4.boxH/ACA 型能指导 rRNA 中的嘧啶向假
尿嘧啶的转换。
• 近年来发现的一类非编码 RNA 。• 不具有 ORF 。• 功能:主要参与胚胎发育、肿瘤形成和抑
制、细胞生长和分化、染色体失活等。
6. 非编码 RNA

C1 Nucleic Acid Structure : Modified Nucleic Acids
Modifications correspond to numbers of specific roles. We will discuss them in some related topics. For example, methylation of A and C to can avoid restriction digestion of endogenous DNA sequence
C. Properties of nucleic acids

2.2 Chemical and Physical Properties of Nucleic Acids
1. Stability of Nucleic Acids
2. Effect of Acid &
applications
3. Effect of alkali &
applications
4. Chemical denaturation
5. Viscosity & applications
6. Buorant density &
application
Chemical properties
Physical properties

Stability of Nucleic Acids1. Hydrogen bonding • Does not normally contribute the stability of nucleic acids
or protein• Contributes to specific structures of these macromolecules.
For example, -helix, -sheet, DNA double helix, RNA secondary structure
2. Stacking interaction/hydrophobic interaction between aromatic base pairs/bases contribute to the stability of nucleic acids.• It is energetically favorable for the hydrophobic bases to exclude waters and stack on top of each other • This stacking is maximized in double-stranded DNA
Fig

Effect of Acid
Strong acid + high temperature: completely hydrolyzed to bases, riboses/deoxyrobose, and phosphate
pH 3-4 : apurinic nucleic acids (嘌呤核酸)[glycosylic bonds attaching purine (A and G) bases to the ribose ring are broken ], can be generated by formic acid (甲酸)

Effect of Alkali & Application
keto form enolate form keto formenolate form
Base pairing is not stable anymore because of the change of tautomeric states of the bases, resulting in DNA denaturation
1. High pH (> 7-8) has subtle (small) effects on DNA structure2. High pH changes the tautomeric (互变异构 )state of the bases

RNA hydrolyzes at higher pH because of 2’-OH groups in RNA
RNA is unstable at higher pH
OH free 5’-OH
2’, 3’-cyclic phosphodiester
alkali
C. Properties of nucleic acids

Chemical Denaturation
Urea (H2NCONH2) ( 尿素) : denaturing PAGE
Formamide (HCONH2) (甲酰胺) : Northern blot
Disrupting the hydrogen bonding of the bulk water solution
Hydrophobic effect (aromatic bases) is reduced
Denaturation of strands in double helical structure

Viscosity (粘性)
Reasons for the DNA high viscosity 1. High axial ratio2. Relatively stiff
Applications:Long DNA molecules can easily be shortened by shearing force. Remember to avoid shearing problem when isolating very large DNA molecule.
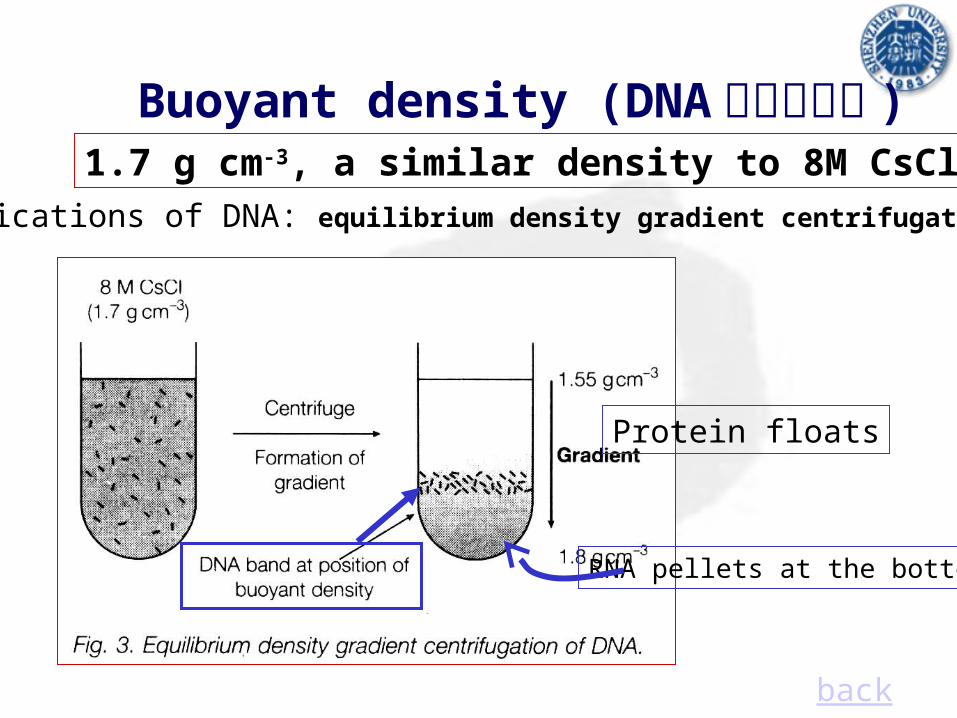
Buoyant density (DNA 的浮力密度 )1.7 g cm-3, a similar density to 8M CsCl
Purifications of DNA: equilibrium density gradient centrifugation
back
RNA pellets at the bottom
Protein floats

1. UV absorption: • nucleic acids absorb UV light due to the aromatic bases• The wavelength of maximum absorption by both DNA and
RNA is 260 nm (max = 260 nm)• Applications: detection, quantitation, assessment of purity
(A260/280)
2. Hypochromicity (减色效应) : caused by the fixing of the bases in a hydrophobic environment by stacking, which makes these bases less accessible to UV absorption. dsDNA, ssDNA/RNA, nucleotide
2.3 Spectroscopic and Thermal Properties of Nucleic Acids

3. Quantitation of nucleic acidsExtinction coefficients: 1 mg/ml dsDNA has an A260 of 20
ssDNA and RNA, 25The values for ssDNA and RNA are approximate(1) The values are the sum of absorbance contributed by the
different bases ( : purines > pyrimidines)(2) The absorbance values also depend on the amount of secondary
structures due to hypochromicity.
4. Purity of DNA A260/280:
dsDNA--1.8
pure RNA--2.0
protein--0.5

back
5. Thermal denaturation/melting: heating leads to the destruction of double-stranded hydrogen-bonded regions of DNA and RNA.
RNA: the absorbance increases gradually and irregularlyDNA: the absorbance increases cooperatively.
melting temperature (Tm): the temperature at which 40% increase in absorbance is achieved.

3-75
1.0
1.37OD
℃
Concentration 50μg/ml
Opetical Density
D.S DNA A260 = 1
S.S DNA A260 = 1.37
dNTPs A260 = 1.60
Tm (melting temperature)
= midpoint of the temperature range over which DNA is denatured

6. Renaturation:Rapid cooling: only allow the formation of local base paring. Absorbance is slightly decreasedSlow cooling: whole complementation of dsDNA. Absorbance decreases greatly and cooperatively.
Annealing: base paring of short regions of complementarity
within or between DNA strands. (example: annealing step in PCR
reaction)
Hybridization: renaturation of complementary sequences
between different nucleic acid molecules.
(examples: Northern or Southern hybridization)
Fig. 2.

3-77
3.7.2.1 DNA 复性的必要条件:
①、一定的离子强度
Na+ 可以中和磷酸基团的负电荷产生的排 斥力
②、较高的温度
维持合适的温度是必须的,温度太低,易产生错配,太高难以形成氢键。

3-78
3.7.2.2 影响 DNA 复性过程的因素 :
① 阳离子浓度 0.15 ~ 0.5 M Na+ 可消除 polydNt 间的静电斥力
② 复性反应的温度 Tm - 25 (60-65 ) ℃ ℃ 以消除 S.S. DNA 分子内的部分二级结构
③ S.S. DNA 分子的长度S.S. DNA愈长→ 分子扩散愈慢→ 复性愈慢S.S. DNA愈短→ 分子扩散愈快→ 复性愈快

3-79
影响 DNA 复性过程的因素 :
⑤ DNA 分子中 , dNt 的排列状况 (随机排列 , 重复排列 )
④ S.S, DNA 的初始浓度 C0
同一种 DNA 分子浓度越高,复性越快。
DNA 的复杂性,简单分子易复性,复杂分子复性难
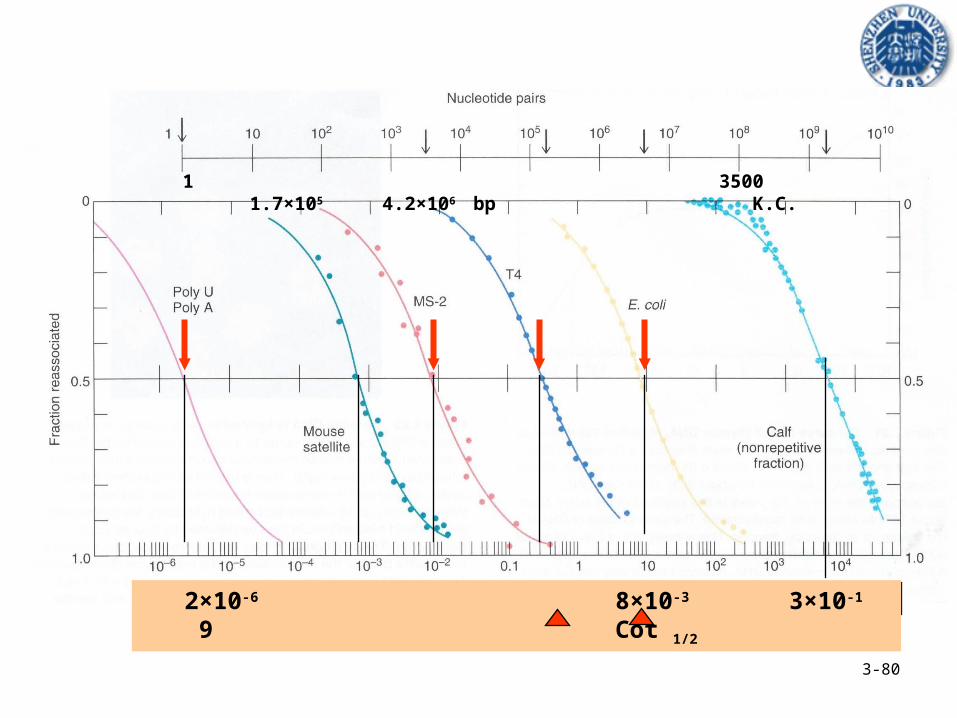
3-80
1 3500 1.7×105 4.2×106 bp K.C.
2×10-6 8×10-3 3×10-1 9 Cot 1/2

3-81
• 复性速度常数 K2 与 DNA 的复杂性成反比。• Cot(1/2) 越高, DNA 越复杂。• 利用 C/Co 值和 Cot 作图就可以求出 Cot(1/2) 值。
• 复性程度的检测方法① 羟基磷灰石柱层析② 测定 S1 核酸梅水解 DNA 的量③ 测定增色效应

DNA Supercoiling
Closed circular molecule
Supercoiling & energy
Topoisomer & topoisomerase

1. Almost all DNA molecules in cells can be considered as circular, and are on average negatively supercoiled.
Counter helical turn

2. Negative supercoiled DNA has a higher torsional ( 扭转的 ) energy than relaxed DNA, which facilitates the unwinding of the helix, such as during transcription initiation or replication
3. Topoisomer: A circular dsDNA molecule with a specific linking number which may not be changed without first breaking one or both strands.

Supercoiled structure of circular DNA molecules with low writhe. Note that the helical nature of the DNA duplex is omitted for clarity.
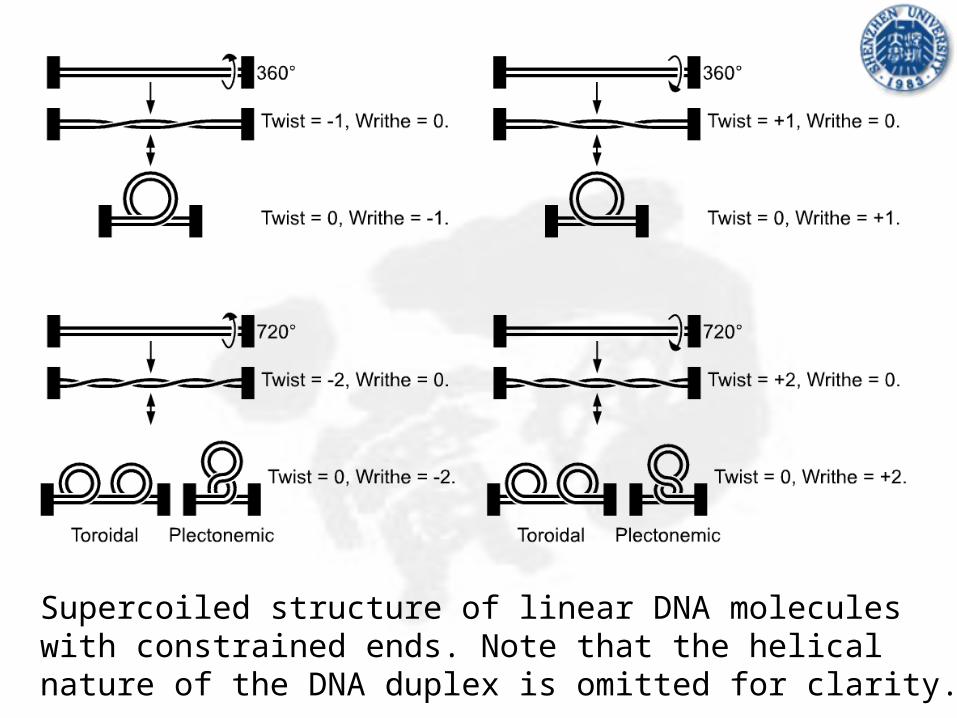
Supercoiled structure of linear DNA molecules with constrained ends. Note that the helical nature of the DNA duplex is omitted for clarity.

Topoisomerases exist in cell to regulate the level of supercoiling of DNA molecules.
Type I topoisomerase: breaks one strand and change the linking number in steps of ±1.
TypeII topoisomerase: breaks both strands and change the linking number in steps of ±2.
Gyrase (促旋酶) : introduce the negative supercoiling (resolving the positive one and using the energy from ATP hydrolysis.

Ethidium bromide (intercalator): locally unwinding of bound DNA, resulting in a reduction in twist and increase in writhe.
Topoisomerases Type I: break one strand of the DNA , and change the linking number in steps of ±1.Type II: break both strands of the DNA , and change the linking number in steps of ±2.