chapter 16: cytoskeleton - queen's universitypost.queensu.ca/~biol330/16 cytoskeleton.pdf ·...
TRANSCRIPT

Chapter 16: Cytoskeleton
Know the terminology:Actin, microfilament, thin filament, tubulin, microtubule, intermediate filament, microtubule-associated protein, flagella, cilia, pseudopodia, lamellapodia, fillipodia, MTOC, linker proteins, accessory proteins, motor proteins, myosin, dynein, kinesin, capping, cross-linkers, MAPs, focal contacts, cadherins
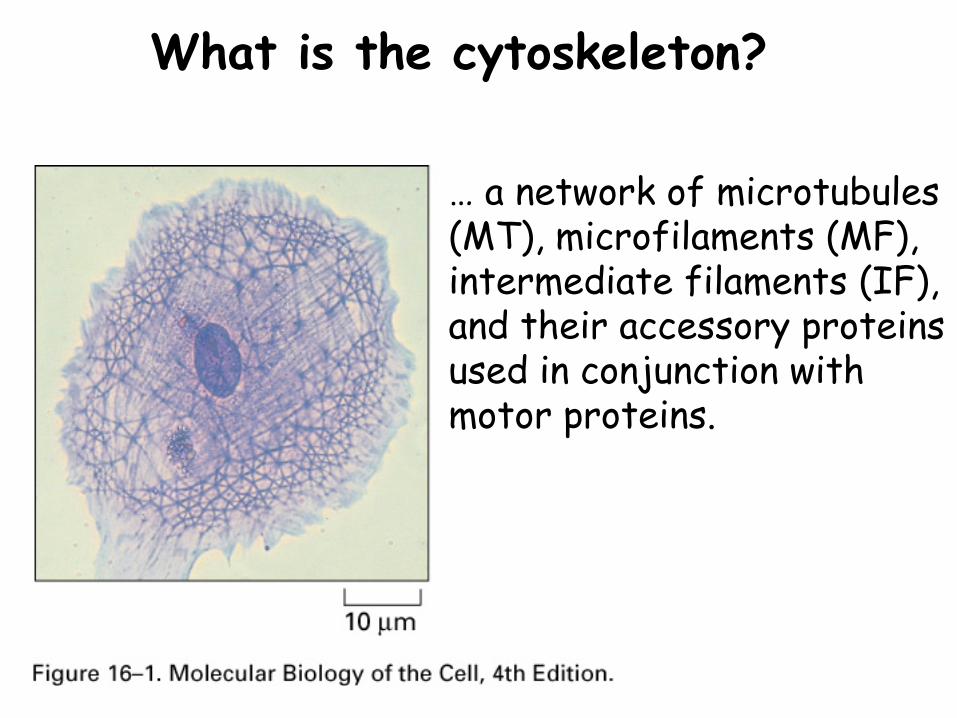
What is the cytoskeleton?
… a network of microtubules (MT), microfilaments (MF), intermediate filaments (IF), and their accessory proteins used in conjunction with motor proteins.

Chapter 16: Cytoskeleton Outline:I. Cytoskeletal elements and their roles
II. Cytoskeleton structure and assembly
III. Regulation of assembly
IV. Molecular motors
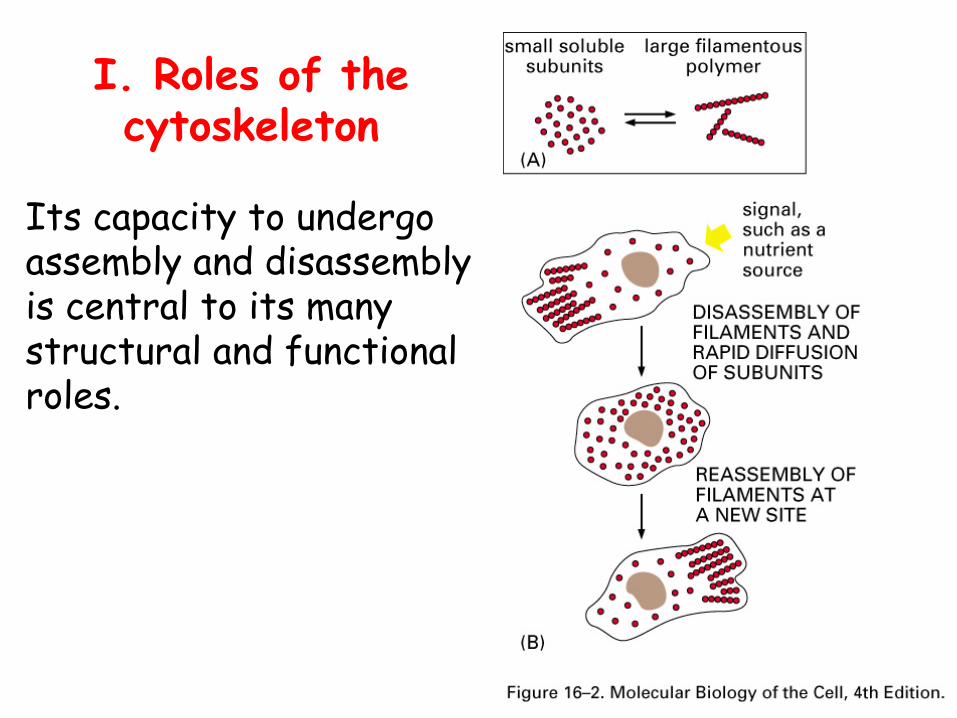
I. Roles of the cytoskeleton
Its capacity to undergo assembly and disassembly is central to its many structural and functional roles.

I. Roles of the cytoskeleton
Cell structure•Cell shape (microvilli)•Internal organization (membrane networks)•Physical robustness (erythrocyte deformability)
Cell function•Cell movement (lamellapodia, flagella, cilia)•Organismal movement (muscles)•Intracellular traffic (vesicle and organelle traffic)•Signal transduction

II. Cytoskeletal structure
Microfilaments (~5nm diameter)
Intermediate filaments (~10nm)
Microtubules (~25nm)
Increasing size
Cytoskeleton is composed primarily of 3 types of polymers

Polymer structure
Polymers are constructed from repeating monomers
Microfilaments (mainly polymers of actin)
Intermediate filaments (diverse subunits)
Microtubules (mainly polymers of tubulin)
How are these analogous to muscles, ligaments, and bones of a musculoskeletal system?

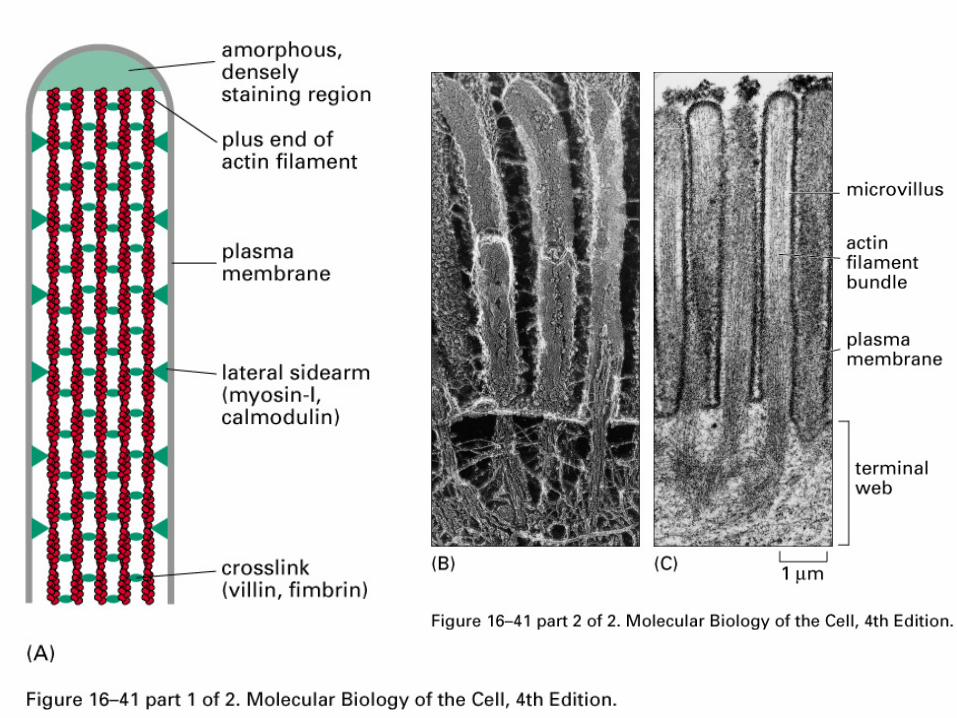
Microfilaments
Most organisms have multiple isoforms of actin-most cells have β, γ-muscles have α
Very conserved structure between isoforms and across species, but subtle variations are important
Actin polymers form microfilaments that are used to build fibers and networks
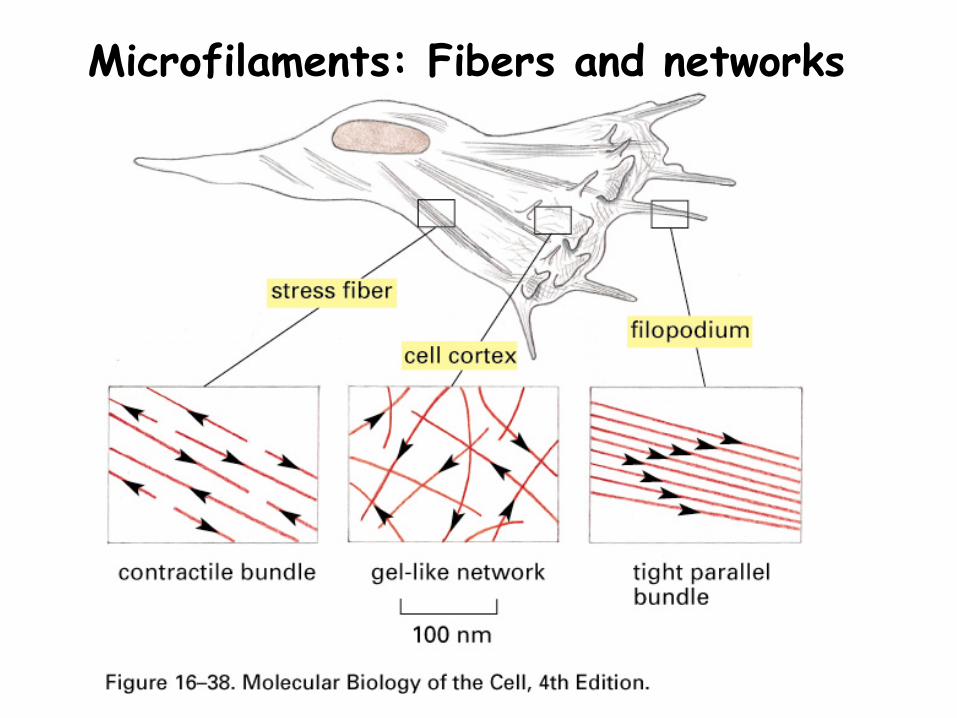
Microfilaments: Fibers and networks
Microfilaments

MicrotubulesMost organisms have multiple isoforms of both α and β tubulin (Other tubulins (γ) are involved in MT assembly)
Very conserved structure between isoforms and across species but subtle variations are important
Microtubules are organized to form an intracellular network, radiating from the MT organizing center (MTOC)
Also the basis of cilia and flagella
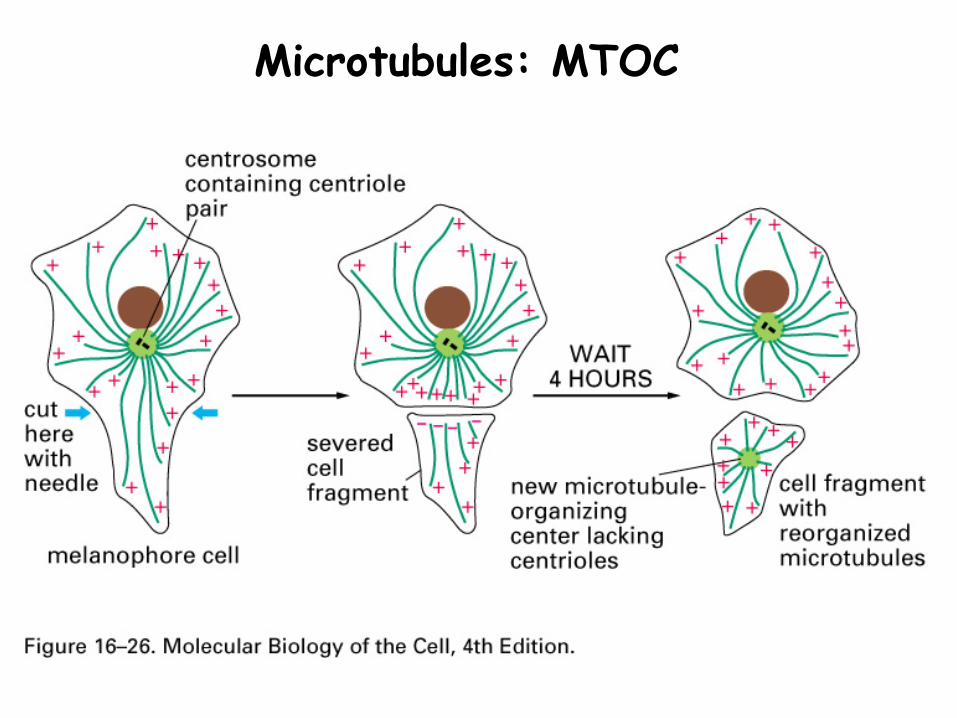
Microtubules: MTOC
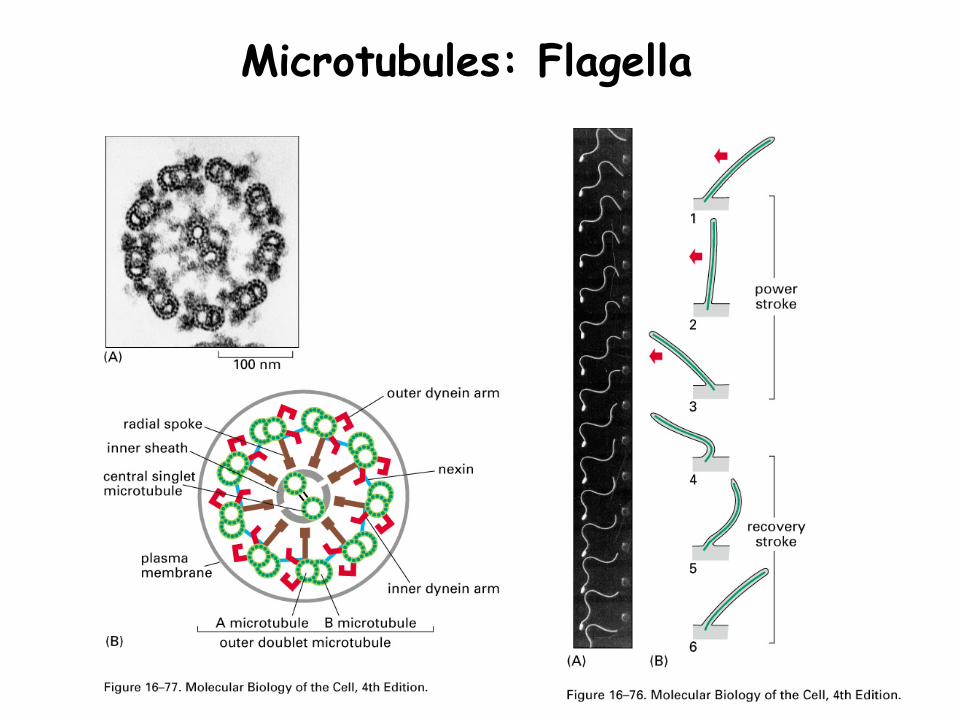
Microtubules: Flagella

Intermediate filamentsComposed of many different types of subunits
Each subunit is a tetramer
Filaments can be bundled together or cross-linked via accessory proteins
Types include keratins (hair, scales, nails, claws) and neurofilaments
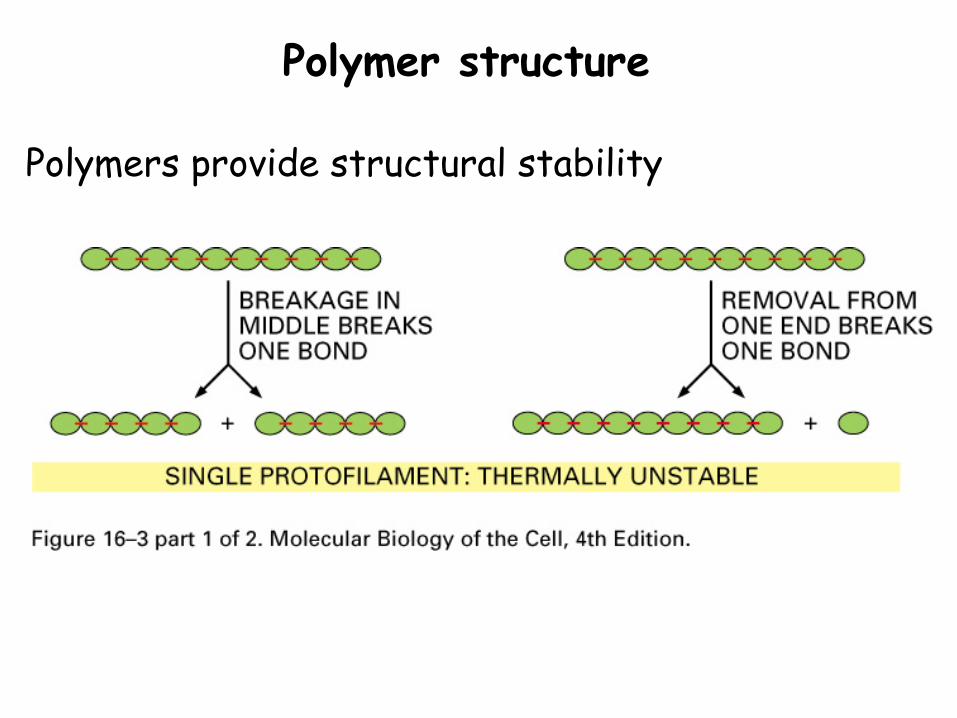
Polymer structure
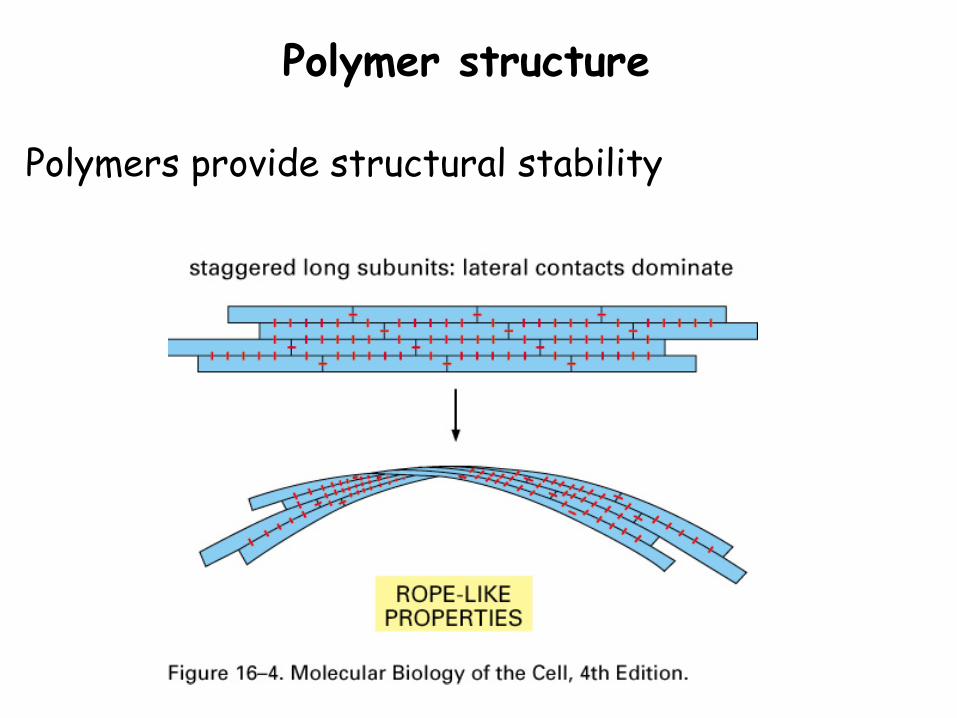
Polymers provide structural stability
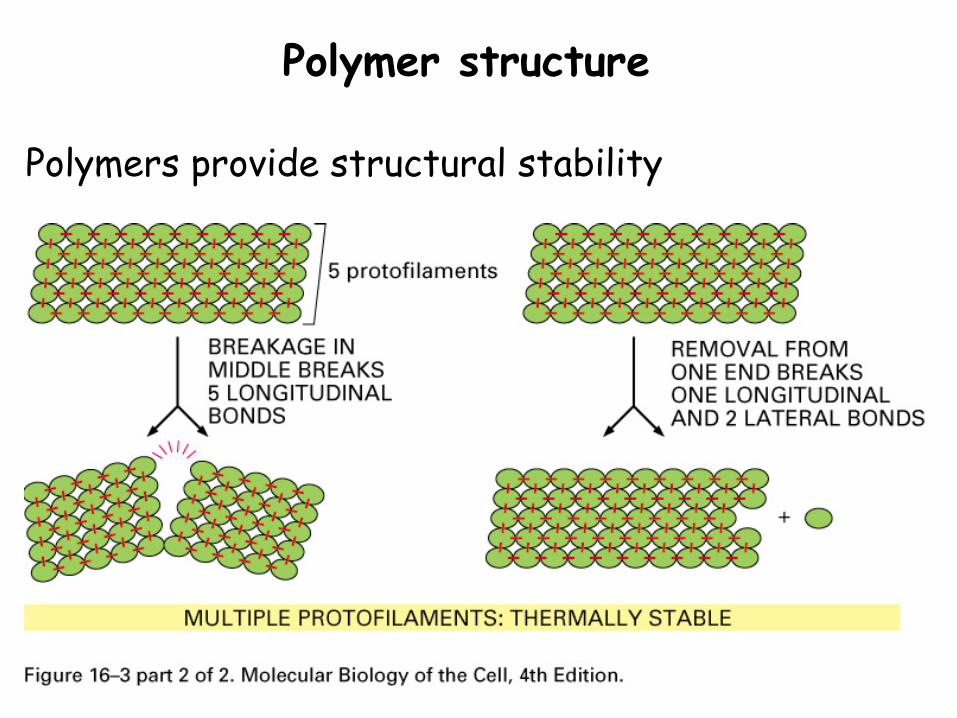
Polymer structure
Polymers provide structural stability
Polymer structure
Polymers provide structural stability

Tubulin polymerization and microtubulesTubulin is a dimer composed of:
α−tubulin & β−tubulin
Both proteins are GTP-binding proteins -the GTP in α−tubulin is part of its structure-the GTP in β−tubulin can be hydrolyzed
Tubulin monomers join end to end to form protofilaments
Protofilaments join side by side to form microtubules
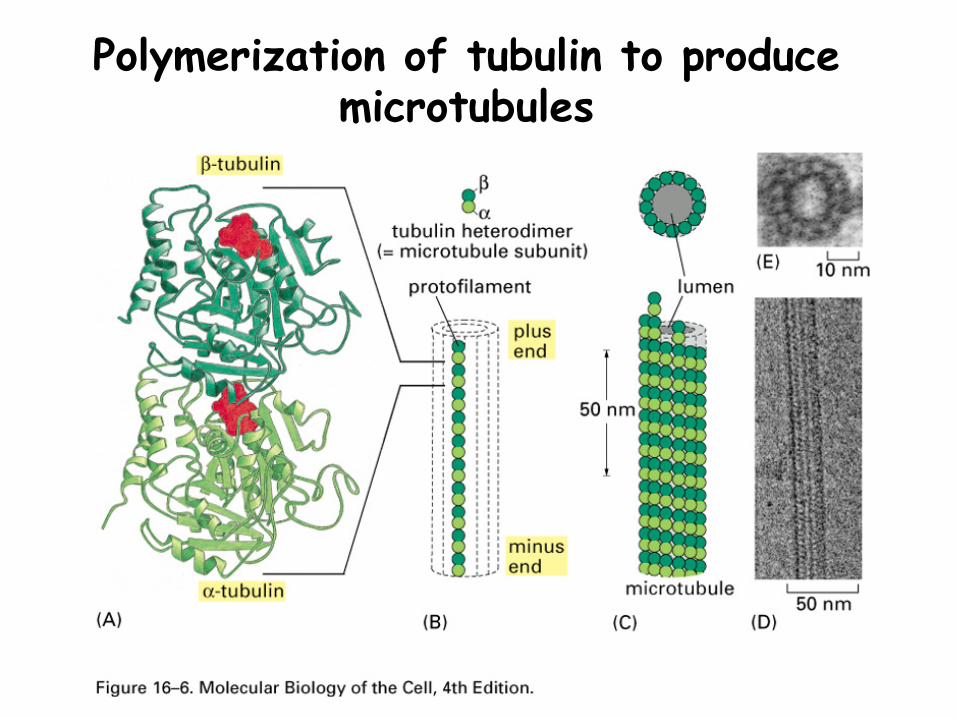
Polymerization of tubulin to produce microtubules
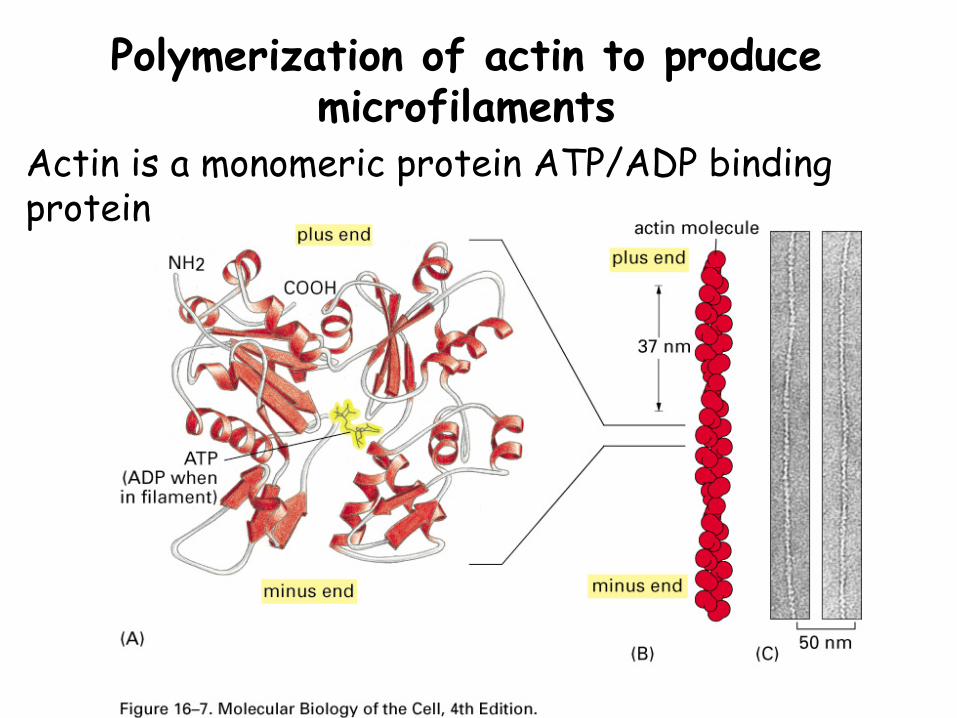
Polymerization of actin to produce microfilaments
Actin is a monomeric protein ATP/ADP binding protein

Nucleotide hydrolysis
The NTPs in the monomers can be hydrolyzed to NDPs (Tubulin GTP to GDP, Actin ATP to ADP)
Monomers with NDP are more likely to dissociate than are monomers with NTP.
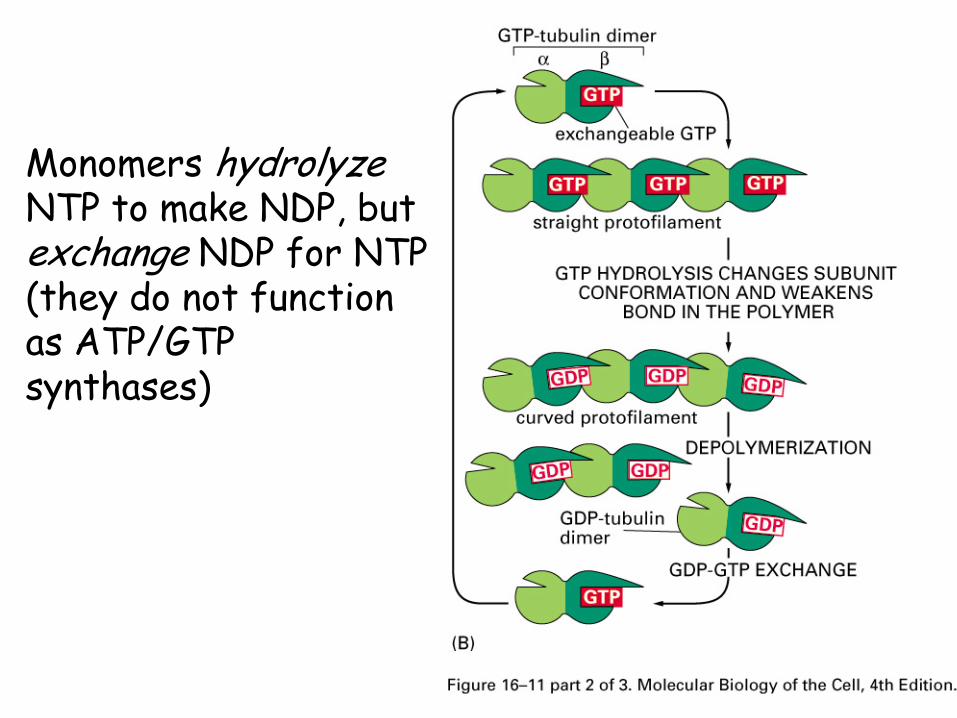
Monomers hydrolyzeNTP to make NDP, but exchange NDP for NTP (they do not function as ATP/GTP synthases)
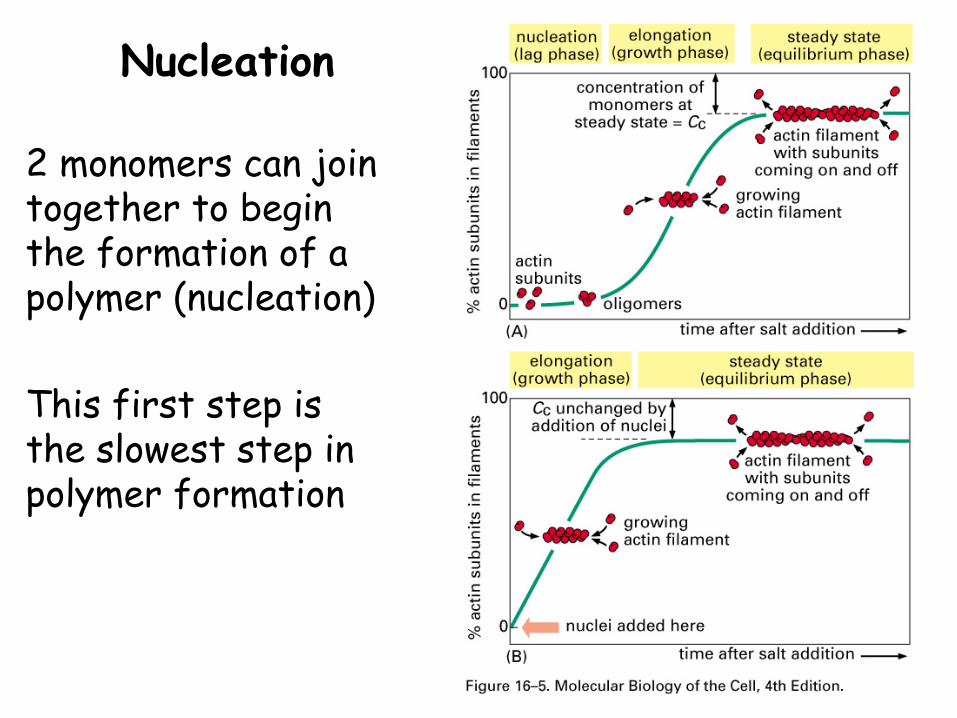
Nucleation
2 monomers can join together to begin the formation of a polymer (nucleation)
This first step is the slowest step in polymer formation

Polarity
The growing polymer has a plus (+) end.
While monomers can bind to either end, they are much more likely to:
-bind to the + end-leave the minus (-) end

Assembly and disassembly
Assembly and disassembly occur simultaneously
The direction of growth and rate of growth are controlled
At a Critical concentration (Cc) the rates of assembly and disassembly are equal.
The plus and minus ends have different Cc values
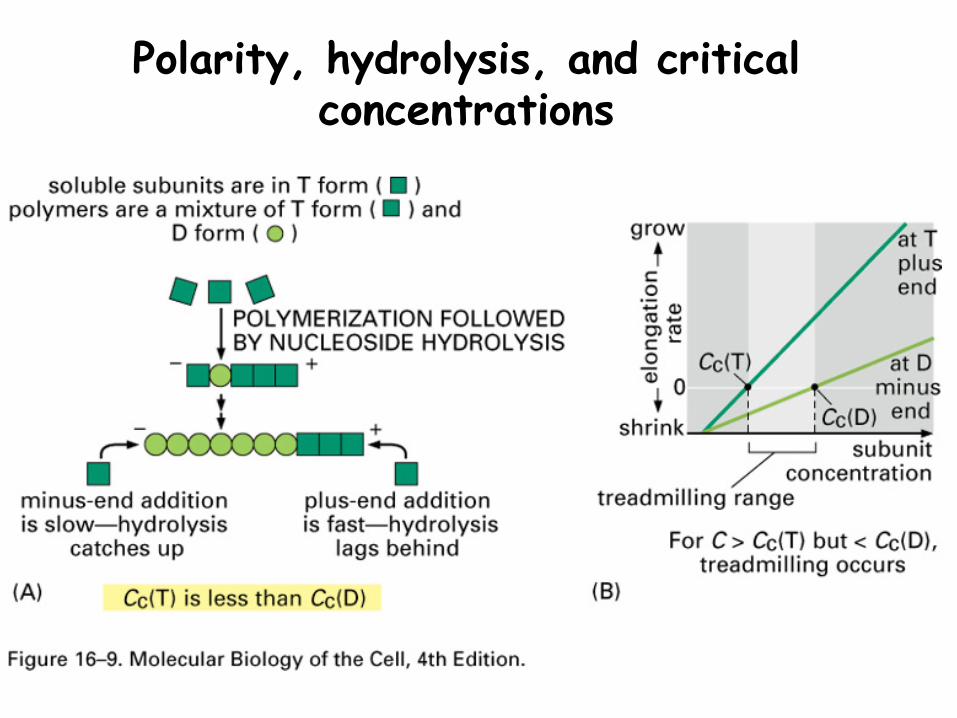
Polarity, hydrolysis, and critical concentrations

Capping effects on polymerization
If the plus end of the polymer has a series of monomers with NTPs, this is referred to as a “cap”
A capped polymer is more likely to grow.
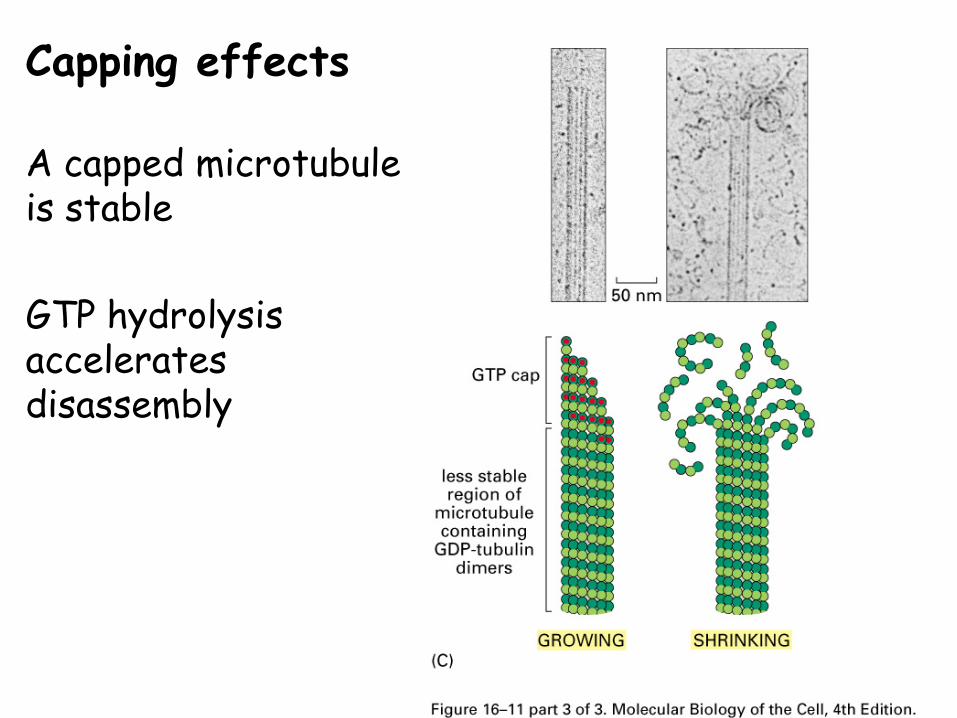
Capping effects
A capped microtubule is stable
GTP hydrolysis accelerates disassembly

Summary
Cytoskeletal proteins are polymers of repeating subunits.
Many structural and functional roles within cells
Assembly and disassembly important to their actions
They have intrinsic polarity, tending to grow from the + end.
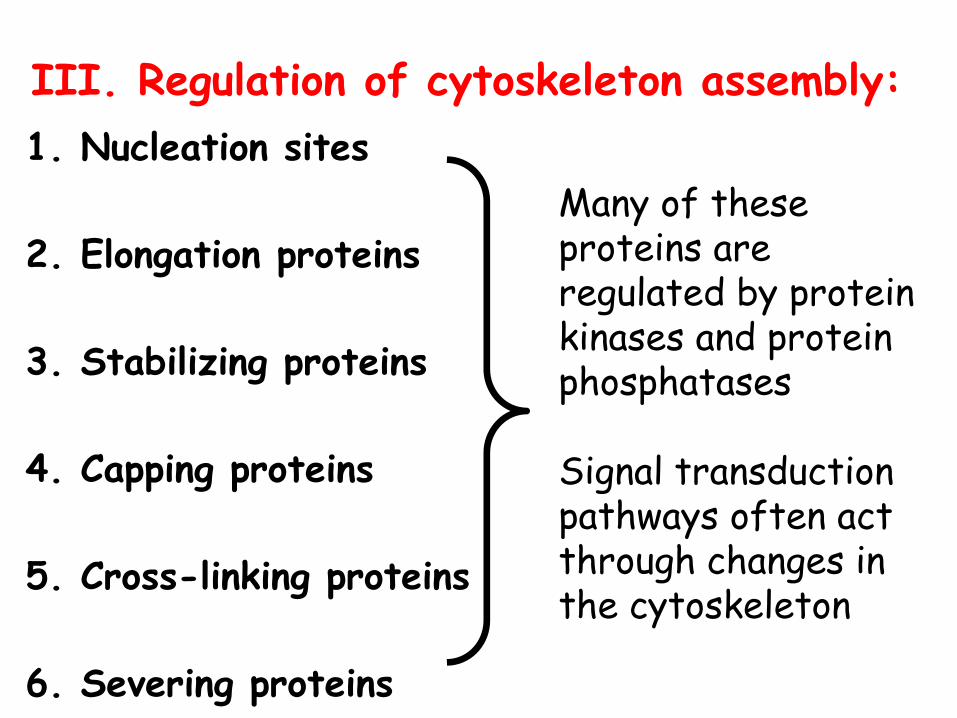
III. Regulation of cytoskeleton assembly:1. Nucleation sites
2. Elongation proteins
3. Stabilizing proteins
4. Capping proteins
5. Cross-linking proteins
6. Severing proteins
Many of these proteins are regulated by protein kinases and protein phosphatases
Signal transduction pathways often act through changes in the cytoskeleton

Regulation of cytoskeleton assembly: 1. Nucleation sites
1. Microtubule organizing center (MTOC)
A structure around paired centrioles, near the nucleus
Acts as nucleating site for MT growth
Binds the minus ends of outwardly directing MT

MTOC
MTs anchored by γ-tubulin and accessory proteins that form ring structures
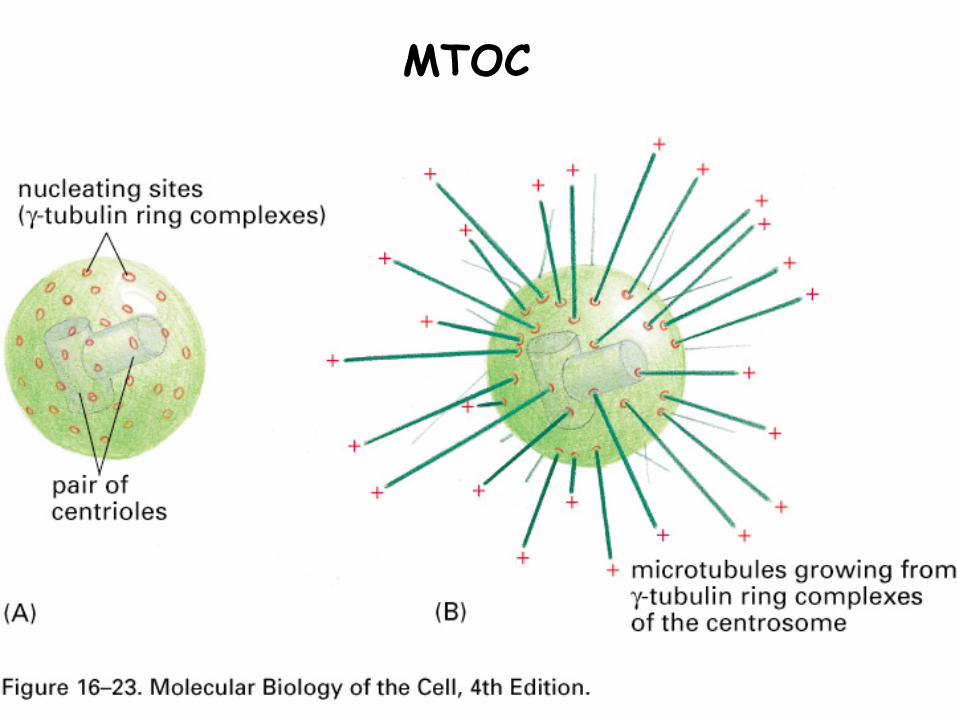
MTOC

Regulation of cytoskeleton assembly: 1. Nucleation sites
2. Actin related proteins (ARP)
Structurally similar to actin
Binds minus end to stabilize it, allowing growth only at plus end
Can also bind other actin filaments to create network
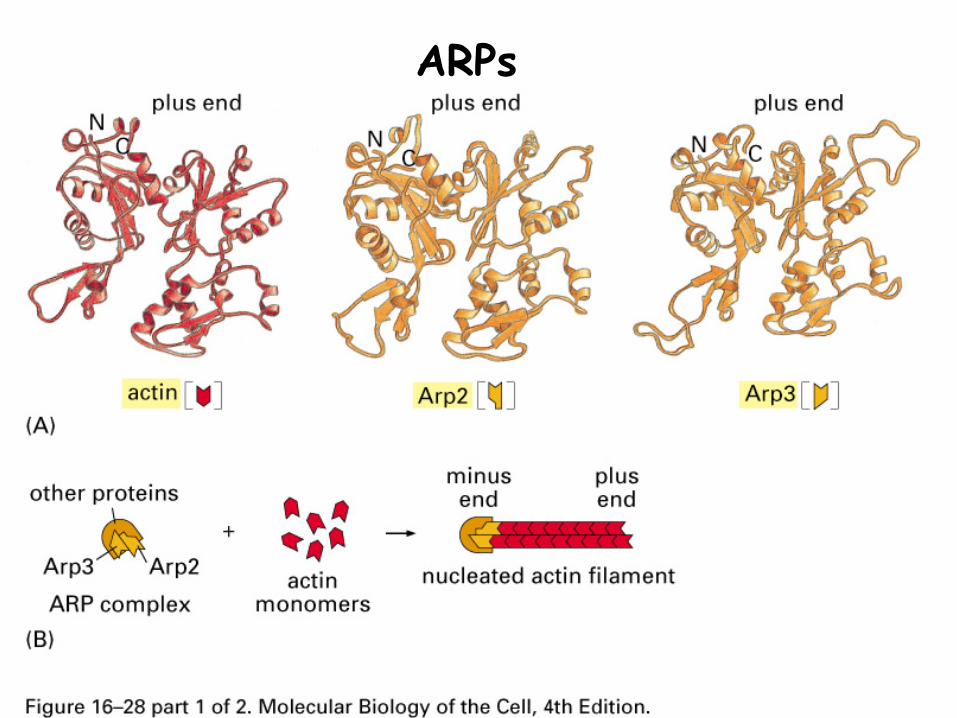
ARPs
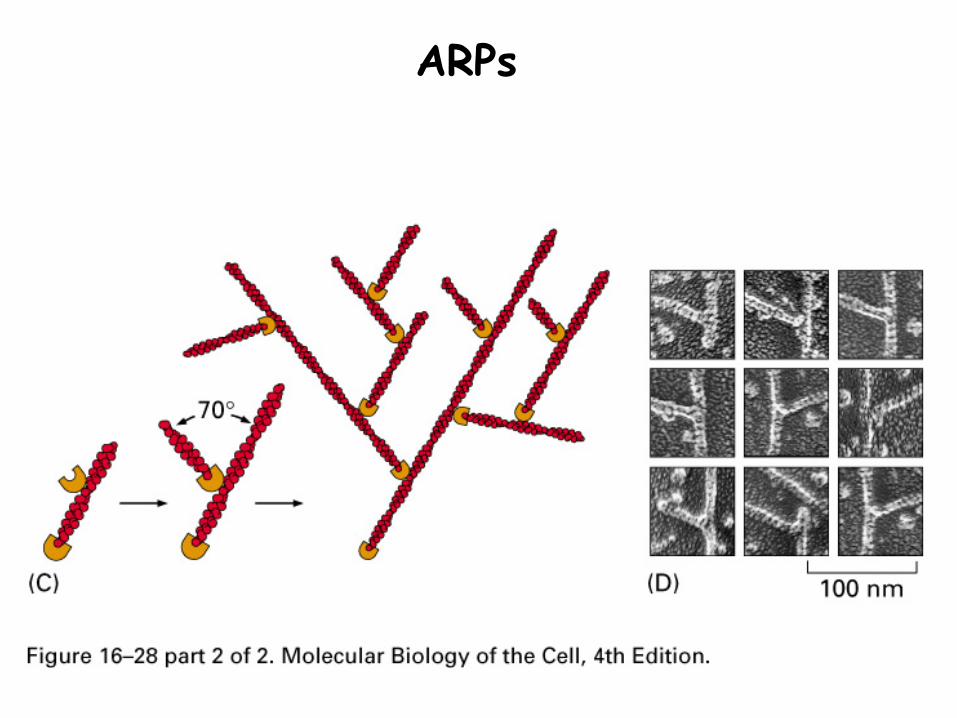
ARPs

1. Both microtubules and microfilaments can bind proteins that alter the ability of monomers to incorporate into the polymer
Actin-profilin assembles faster than actin aloneActin-thymosin assembles slower than actin alone
Stathmin sequesters tubulin to reduce the concentration of free monomers
Regulation of cytoskeleton assembly: 2. Elongation control
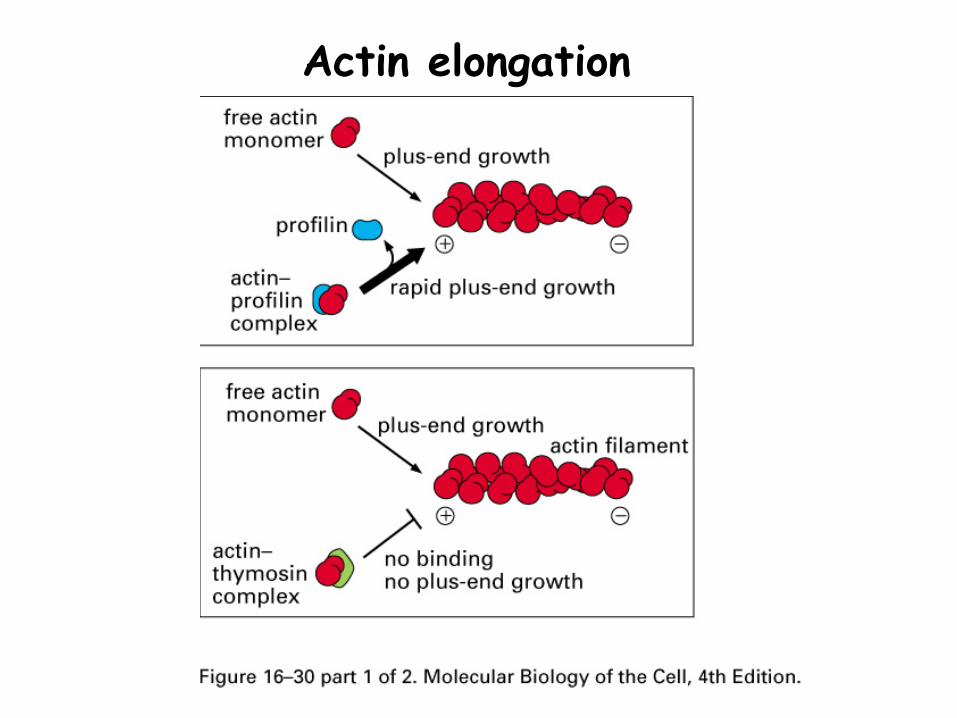
Actin elongation
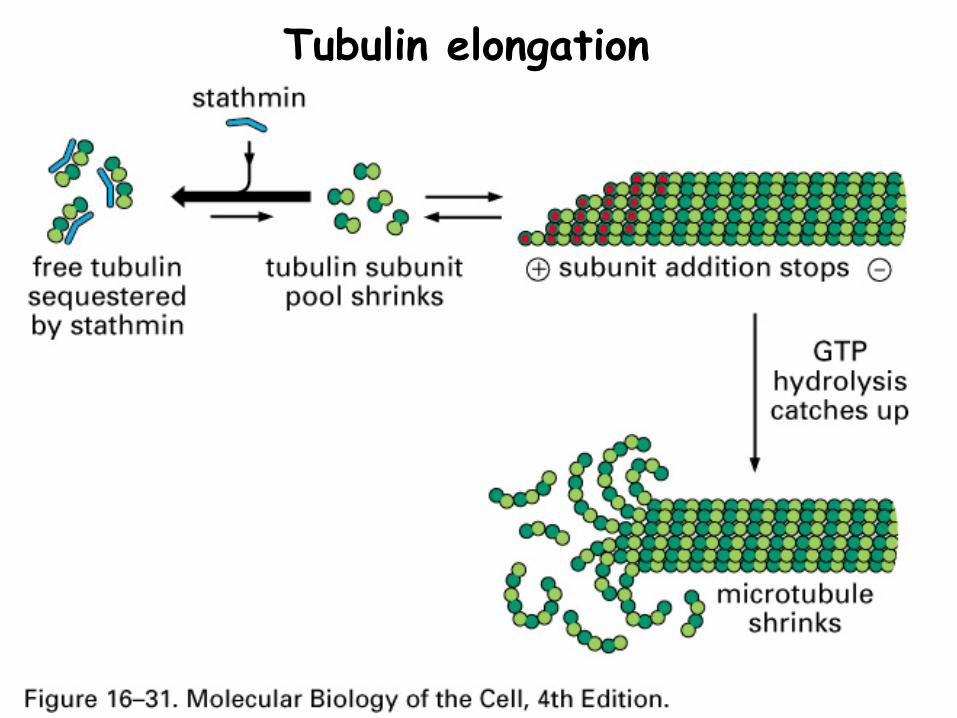
Tubulin elongation

Many microtubule associated proteins (MAPs) bind to the sides of microtubules to stabilize or destabilize (example: tau protein)
Many proteins bind to microfilaments to stabilize or destabilize the filament (example: cofilin)
Regulation of cytoskeleton assembly: 3. Stabilization control
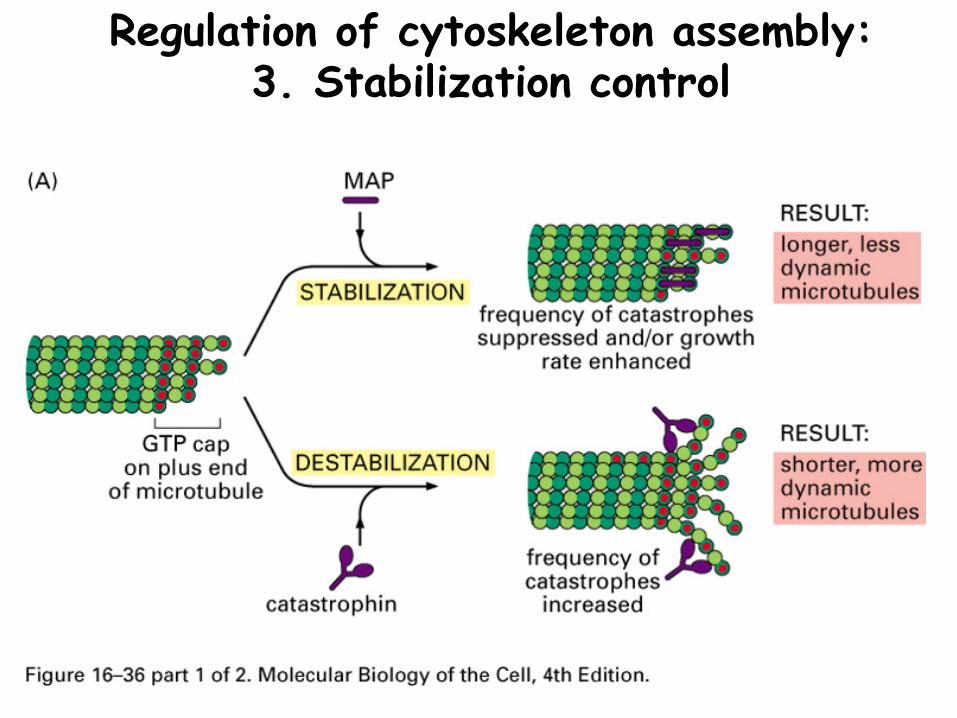
Regulation of cytoskeleton assembly: 3. Stabilization control

Both microfilaments and microtubules have proteins that can bind the ends to alter the ability to assemble or disassemble
Actin: tropomodulin, CapZMicrotubules: γ-tubulin
Regulation of cytoskeleton assembly: 4. Capping proteins

Assembly of MT and MF into 3 dimensional networks requires proteins that interconnect strands
Regulation of cytoskeleton assembly: 5. Cross-linking proteins
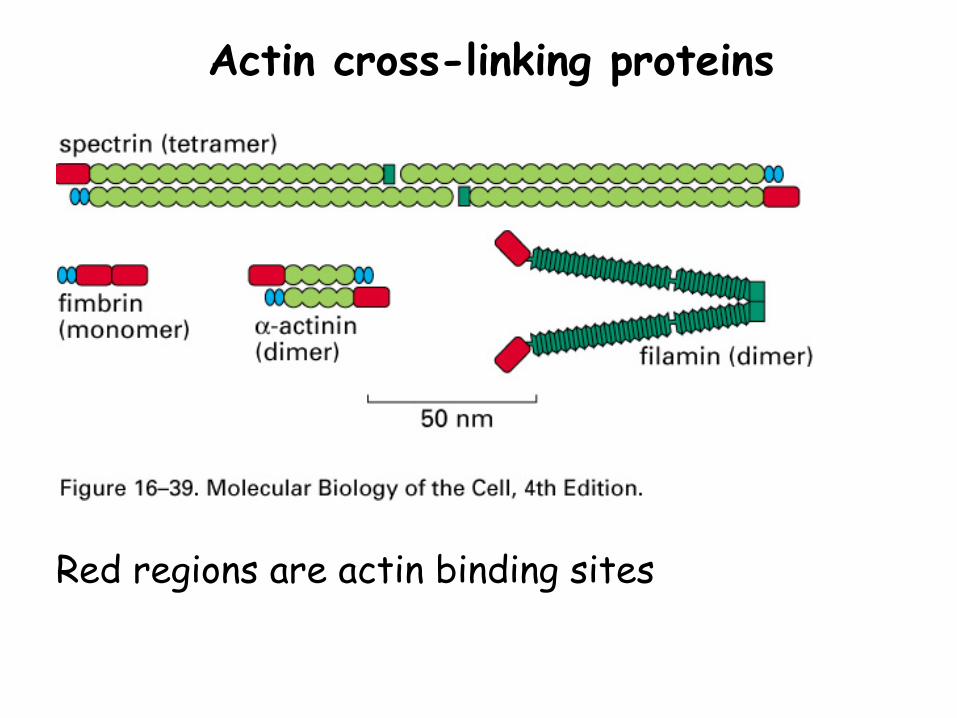
Actin cross-linking proteins
Red regions are actin binding sites
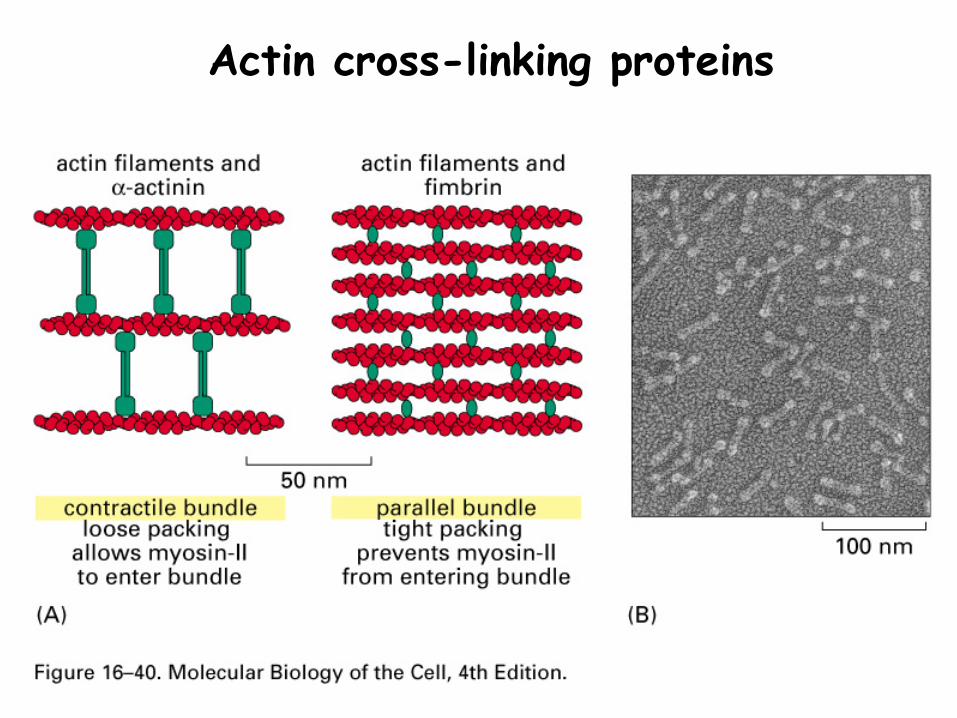
Actin cross-linking proteins
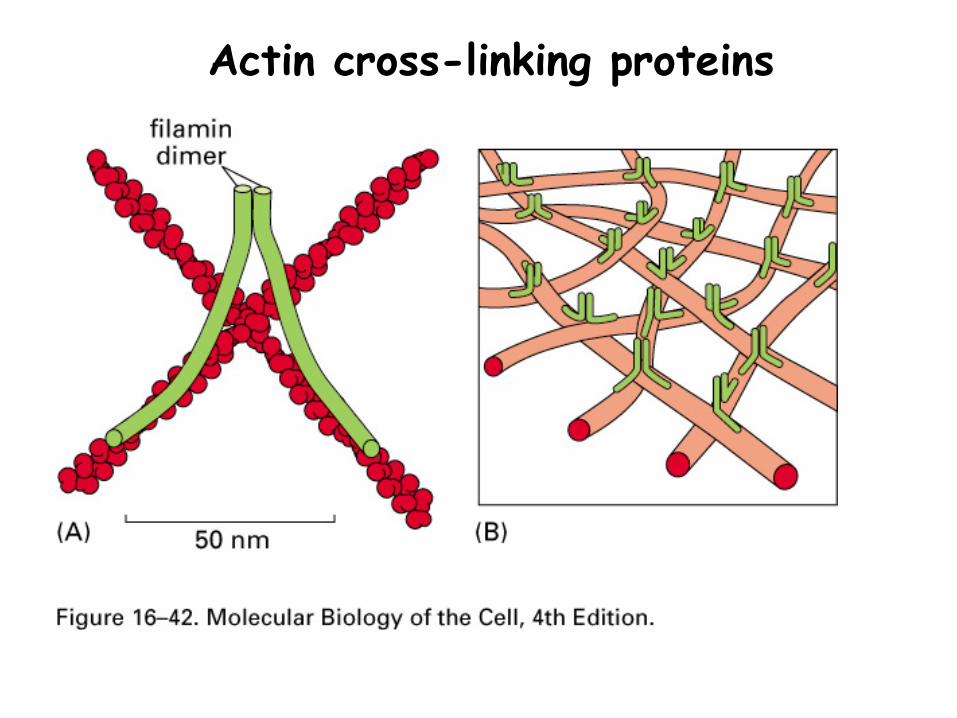
Actin cross-linking proteins

Rapid disassembly of the cytoskeleton is triggered by proteins that cut MT and MF, allowing de-polymerization
Katanin cuts MT, using the energy of ATP hydrolysis
Gelsolin cuts MF in response to high Ca2+
Regulation of cytoskeleton assembly: 6. Severing proteins
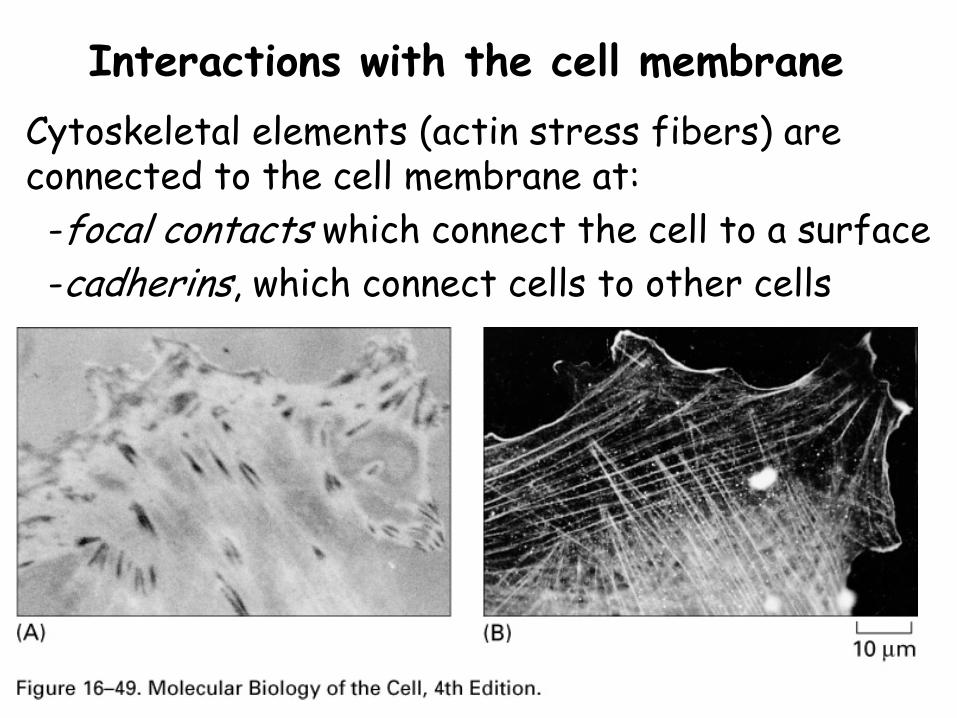
Interactions with the cell membraneCytoskeletal elements (actin stress fibers) are connected to the cell membrane at:-focal contacts which connect the cell to a surface-cadherins, which connect cells to other cells
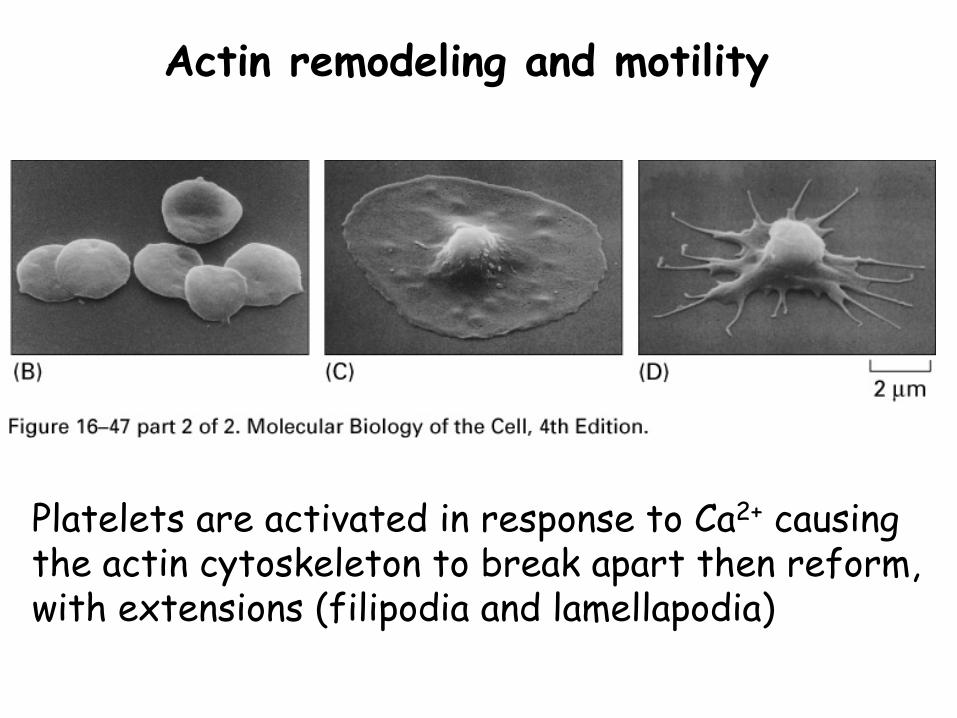
Actin remodeling and motility
Platelets are activated in response to Ca2+ causing the actin cytoskeleton to break apart then reform, with extensions (filipodia and lamellapodia)
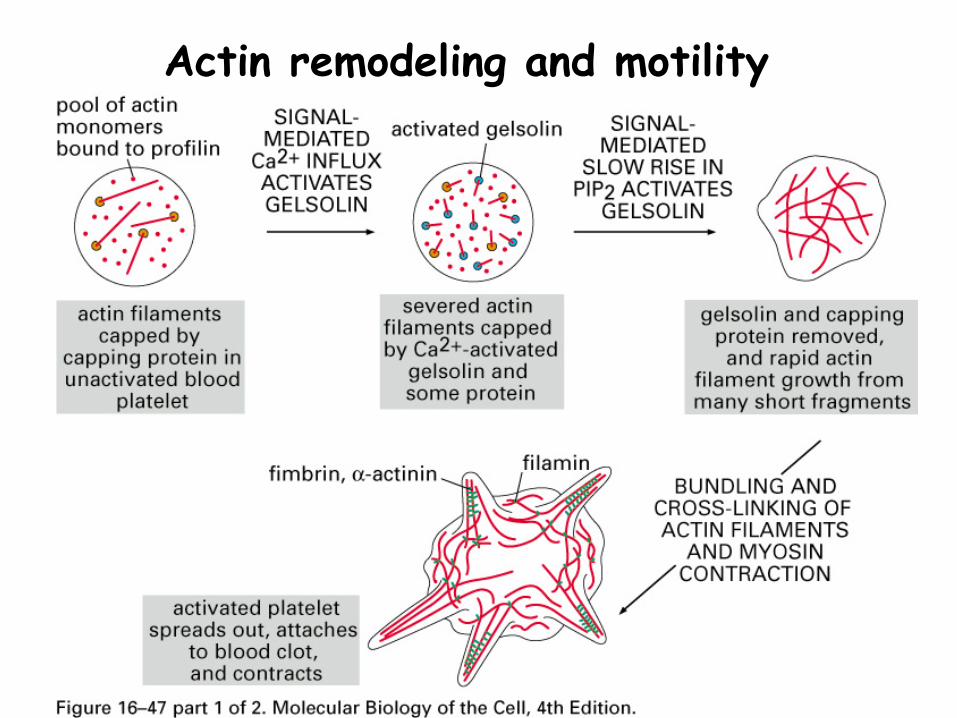
Actin remodeling and motility
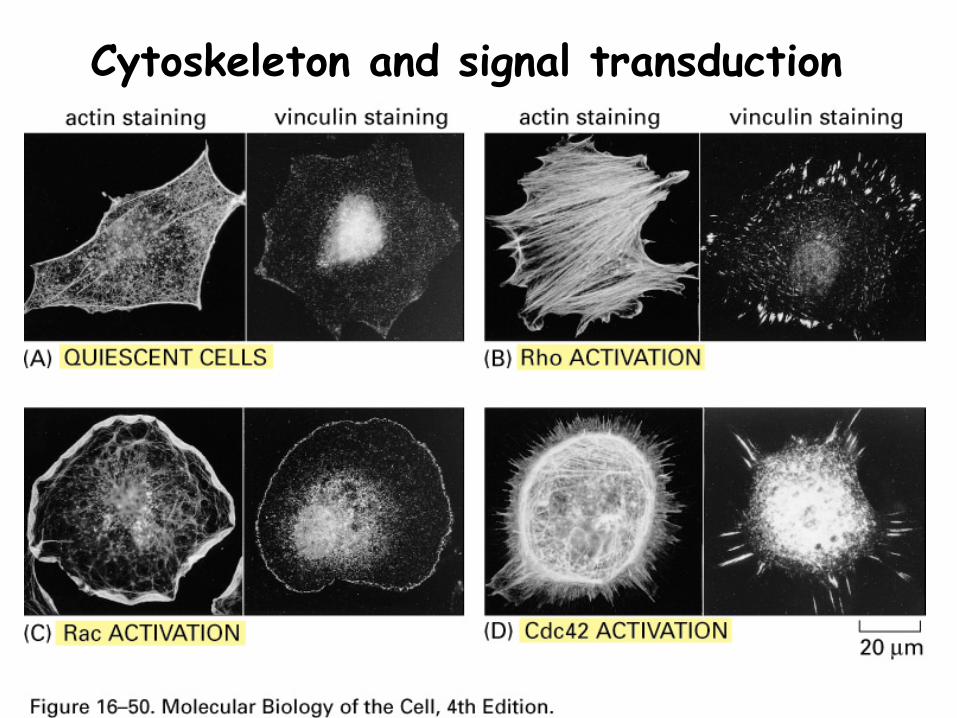
Cytoskeleton and signal transduction

Summary
Extracellular signals trigger changes in the cytoskeleton by altering the activity of proteins that bind MT and MF

Molecular motors
1. What are the molecular motors?
2. How do they catalyze different types of movement?

Molecular motors
Enzymes that use the hydrolysis of ATP to provide the energy to move along cytoskeletal “tracks”
Each specific type of motor protein moves:-on a specific type of track-in a characteristic direction (toward + or - end)
Myosin moves along actin (toward + end)Movement along microtubules uses kinesin (mostly toward +) and dynein (mostly -ve)
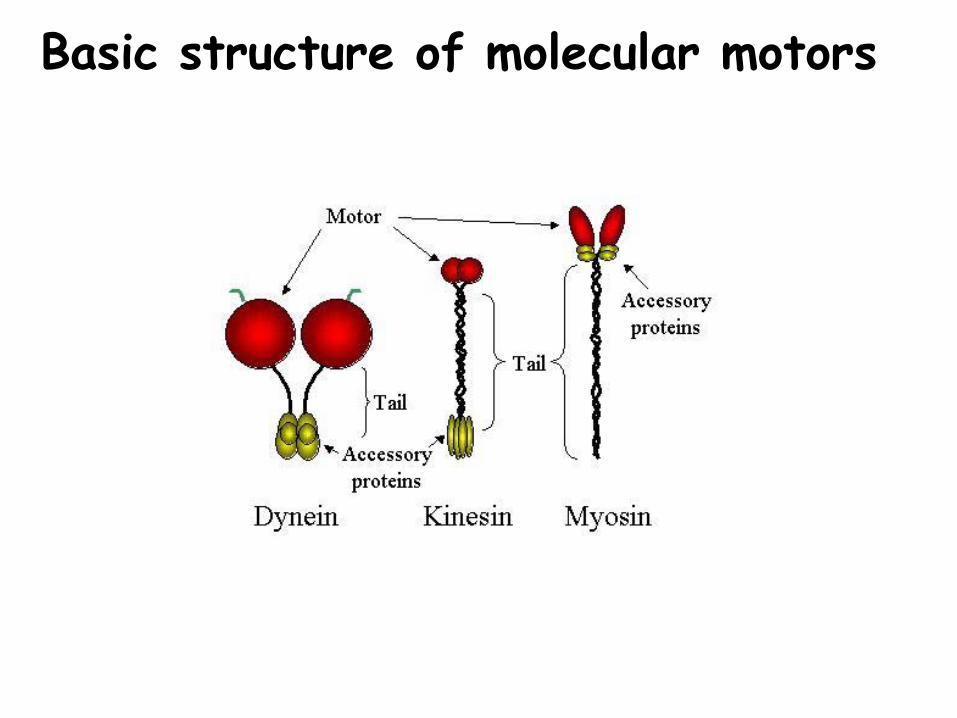
Basic structure of molecular motors

Basic structure of molecular motors
Catalytic (ATPase) head hydrolyses ATP causing a change in three dimensional structure.
A tail that interacts with other proteins such as:-other motors (e.g. muscle thick filament)-vesicle membrane proteins-organelles proteins-cell membrane proteins
Neck, connects head to tail, and may possess accessory proteins that modify the properties of the motor protein

Molecular motors:Myosin
Myosin (II) is composed of:•2 myosin heavy chains•4 myosin light chains
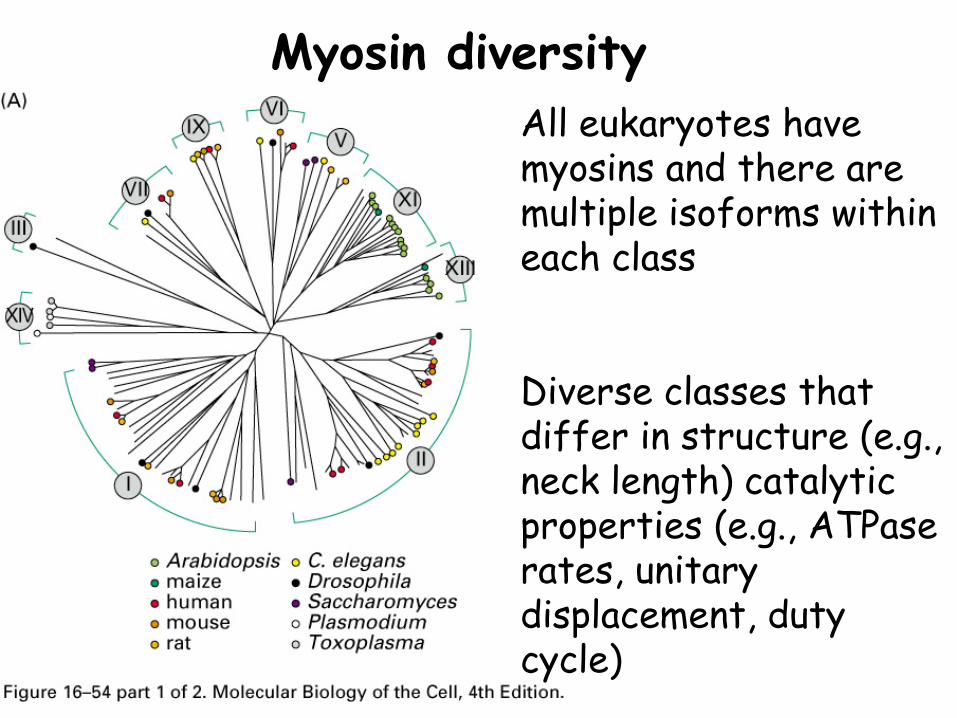
Myosin diversityAll eukaryotes have myosins and there are multiple isoforms within each class
Diverse classes that differ in structure (e.g., neck length) catalytic properties (e.g., ATPase rates, unitary displacement, duty cycle)
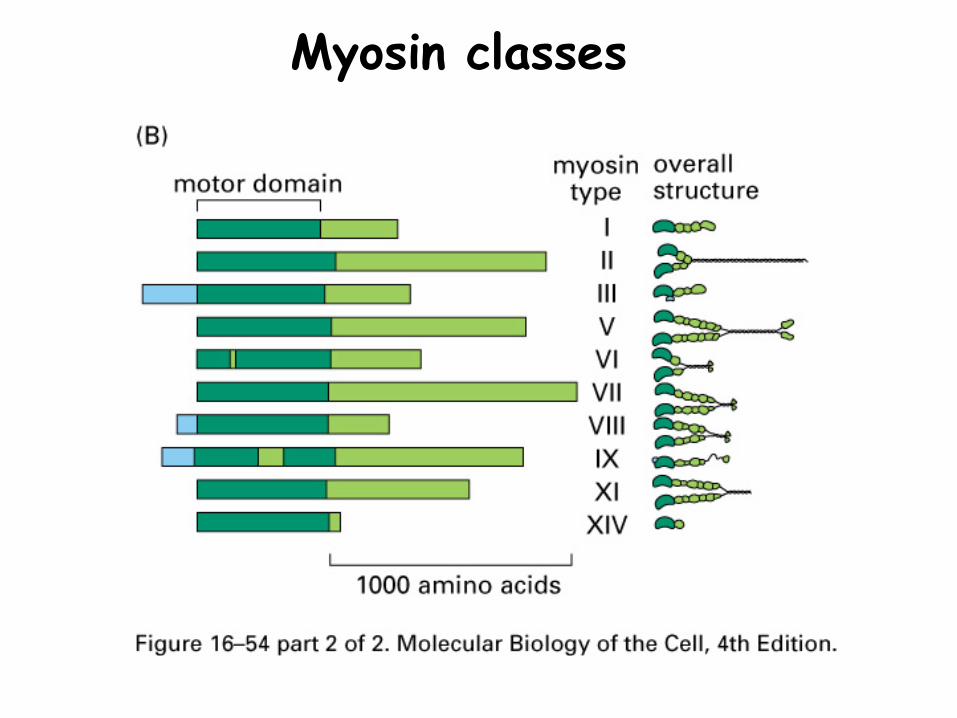
Myosin classes
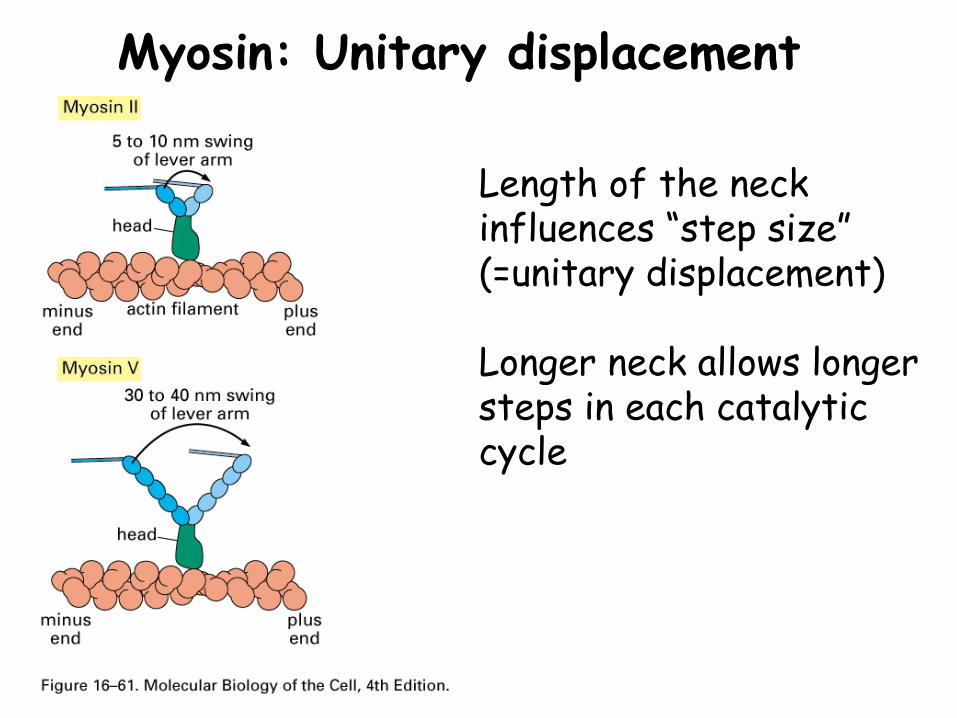
Myosin: Unitary displacement
Length of the neck influences “step size” (=unitary displacement)
Longer neck allows longer steps in each catalytic cycle
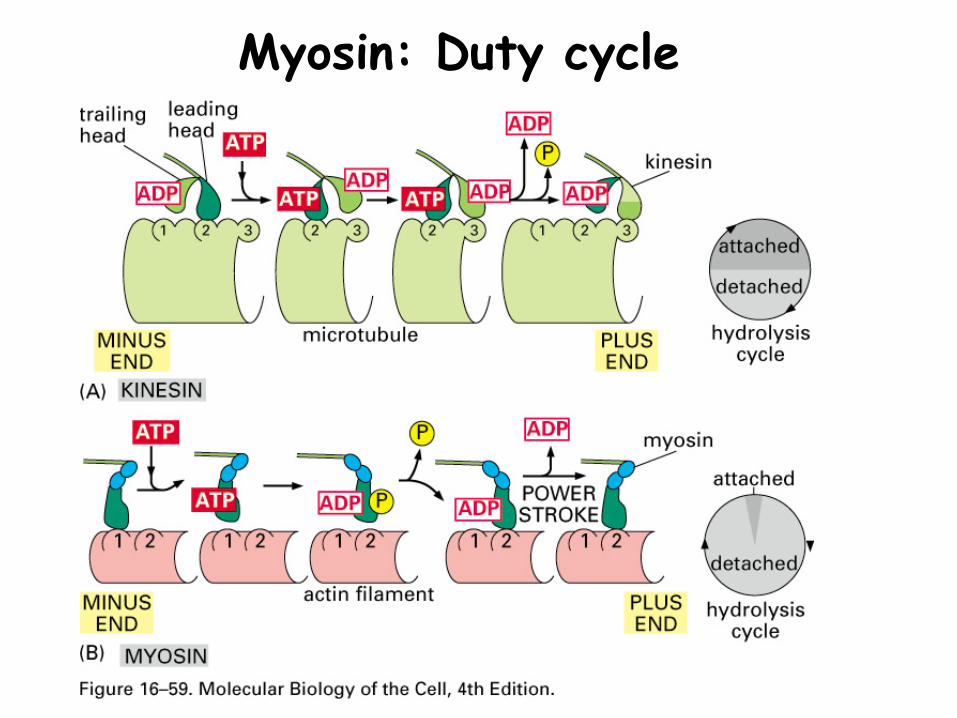
Myosin: Duty cycle

Myosin: Duty cycleDuty cycle: Proportion of time in a contractile cycle where myosin is attached to actin microfilament
Vesicle myosins have long duty cycles (~50%)-allows myosin to remain in contact with the actin microfilament-why doesn’t the vesicle fall off the microfilament when the myosin detaches?
Muscle myosins have short duty cycles (attached to the actin filament only ~5% of the time)
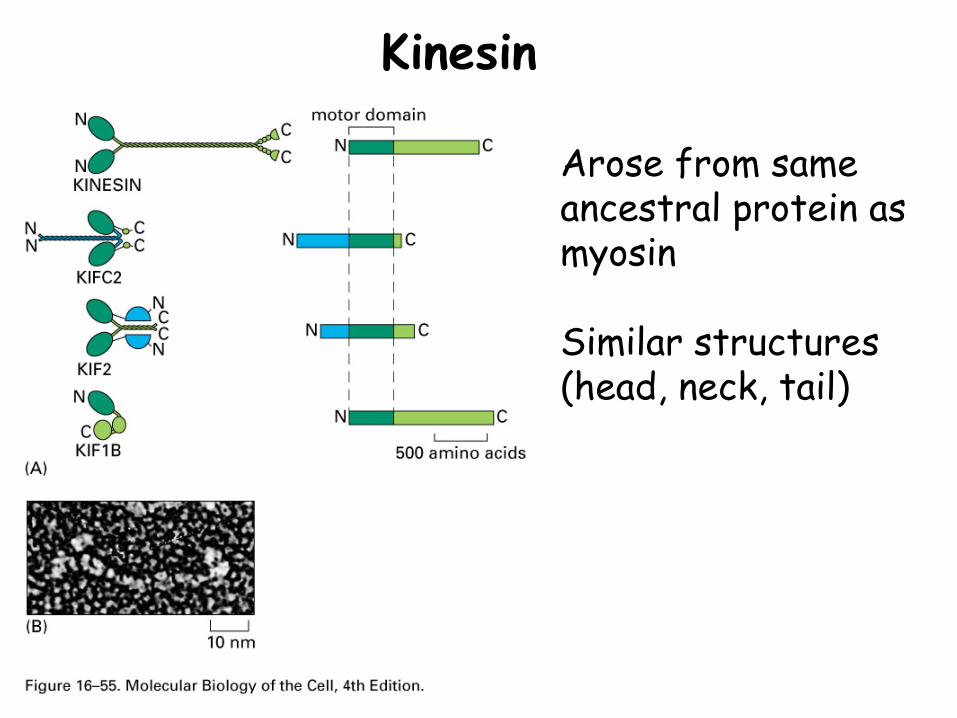
Kinesin
Arose from same ancestral protein as myosin
Similar structures (head, neck, tail)
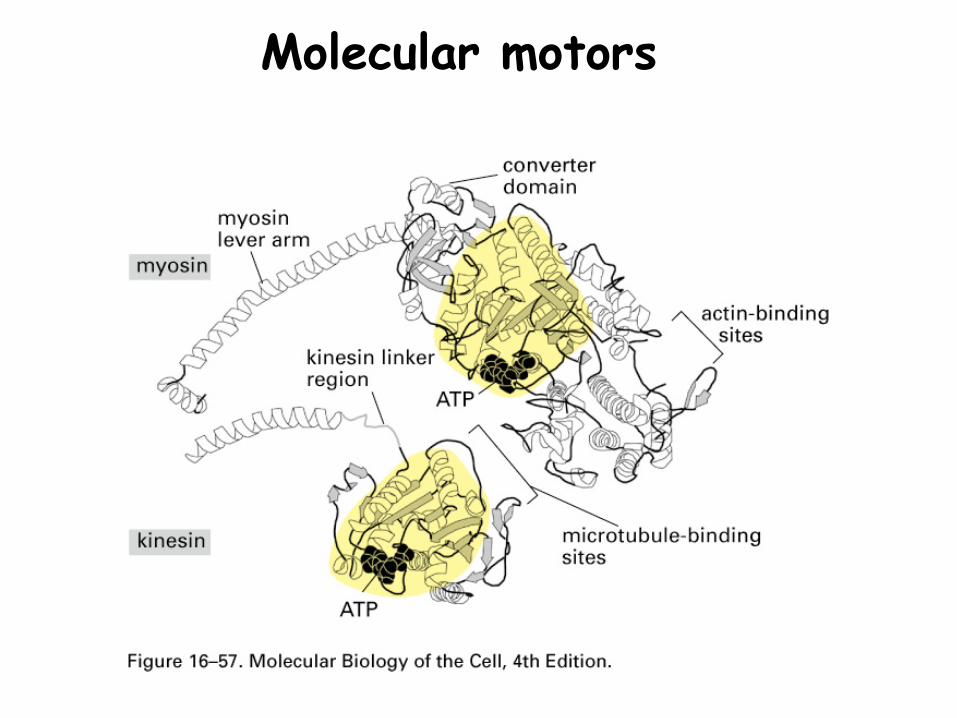
Molecular motors
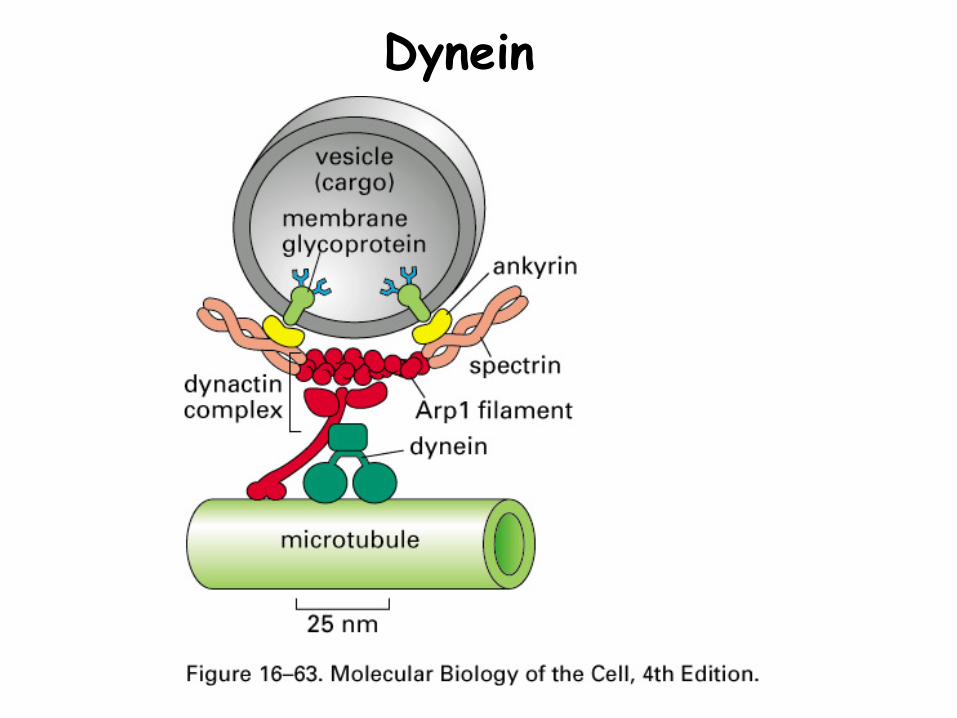
Dynein

Intracellular movementMotor proteins carry diverse intracellular particles and vesicles along cytoskeletal tracks
•Organelles•Secretory vesicles•Transport vesicles•Pigment granules
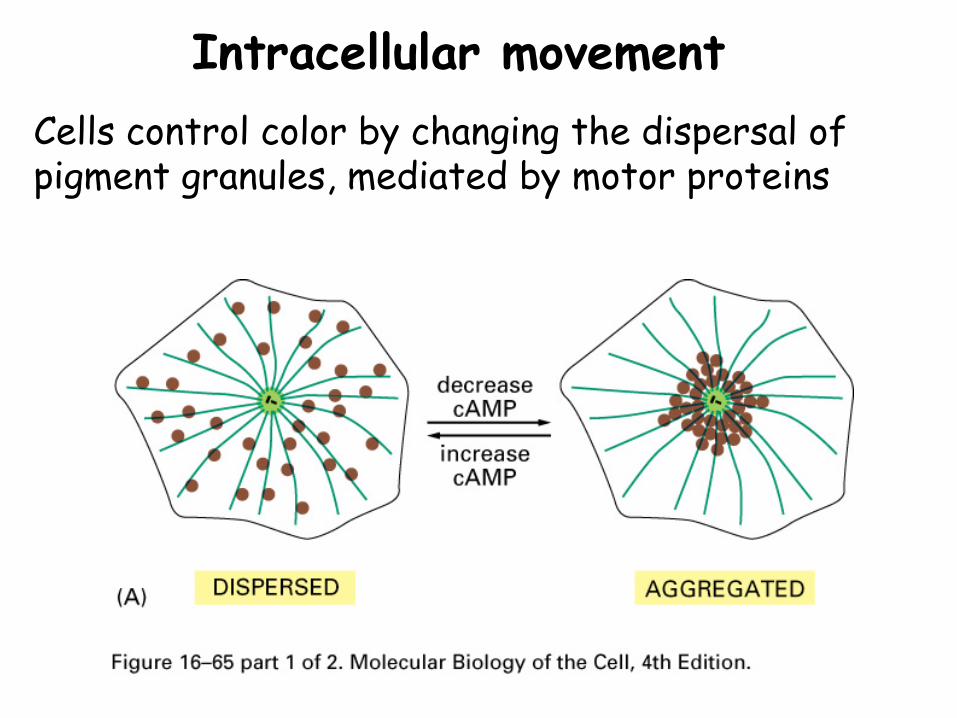
Intracellular movementCells control color by changing the dispersal of pigment granules, mediated by motor proteins
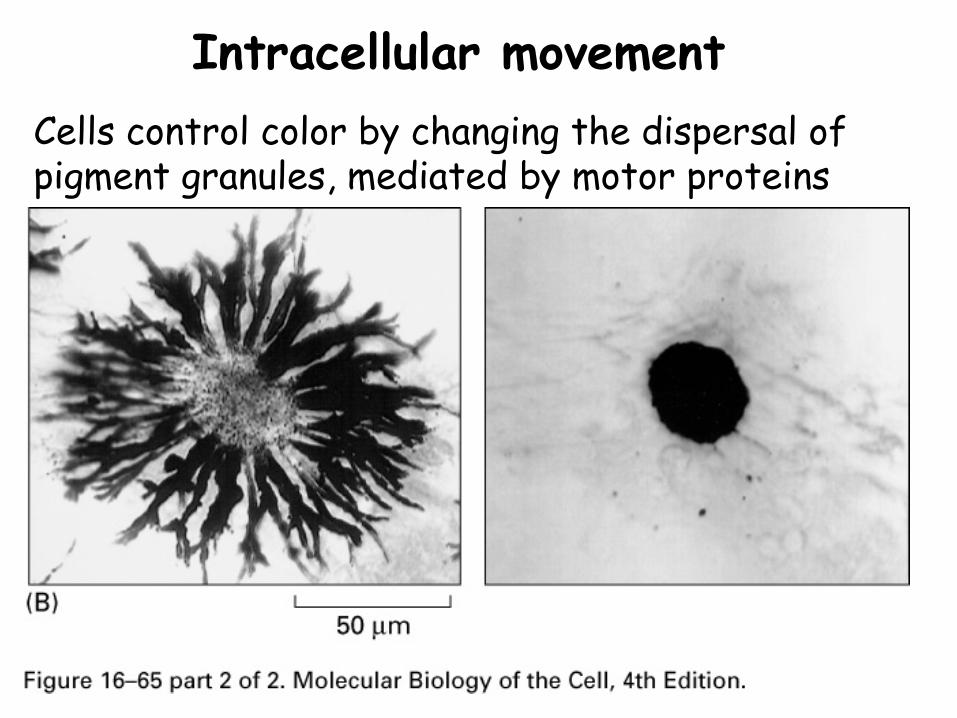
Intracellular movementCells control color by changing the dispersal of pigment granules, mediated by motor proteins
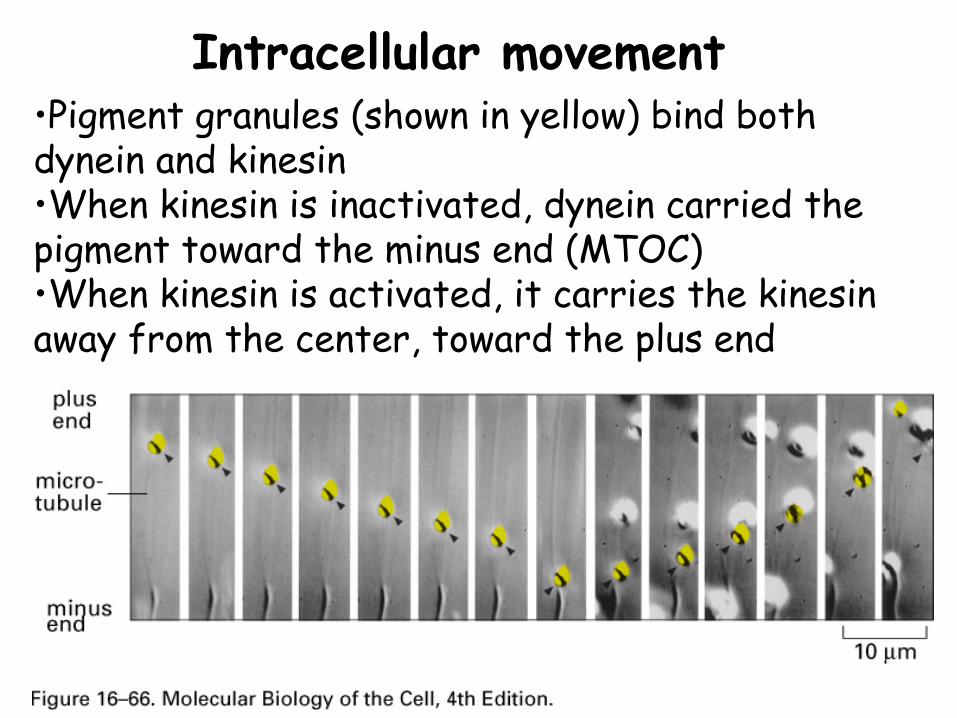
Intracellular movement•Pigment granules (shown in yellow) bind both dynein and kinesin•When kinesin is inactivated, dynein carried the pigment toward the minus end (MTOC)•When kinesin is activated, it carries the kinesin away from the center, toward the plus end
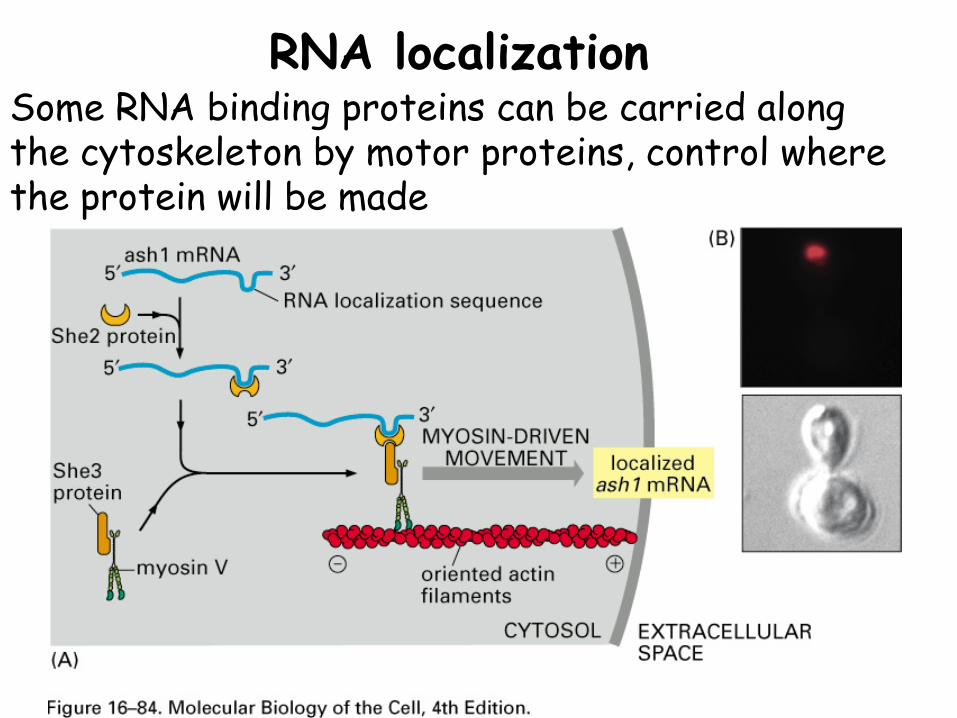
RNA localizationSome RNA binding proteins can be carried along the cytoskeleton by motor proteins, control where the protein will be made

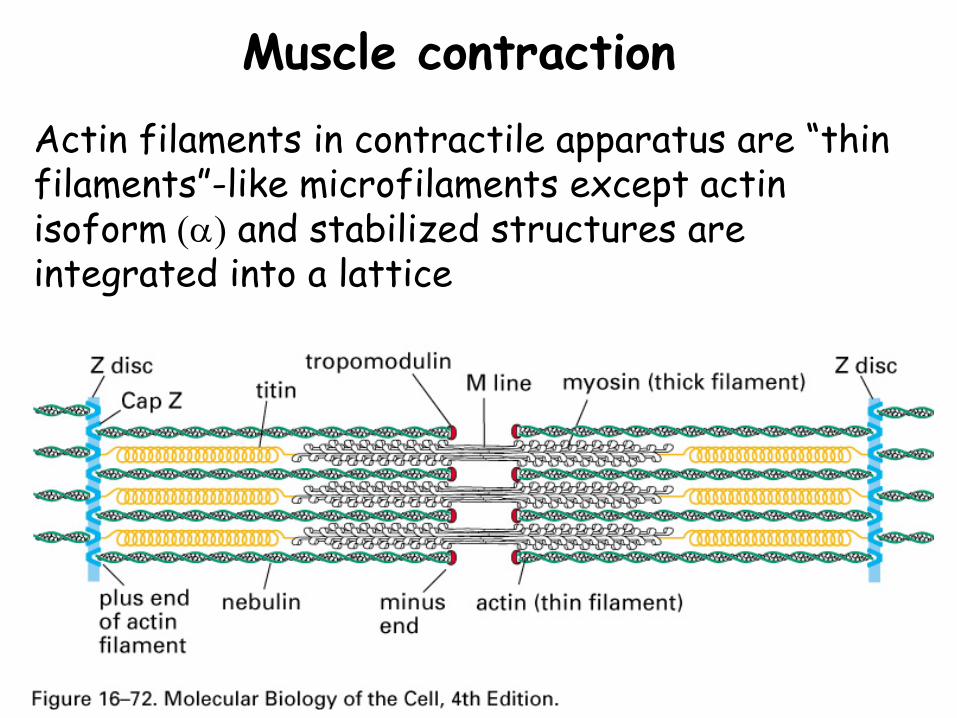
Muscle contractionActin filaments in contractile apparatus are “thin filaments”-like microfilaments except actin isoform (α) and stabilized structures are integrated into a lattice
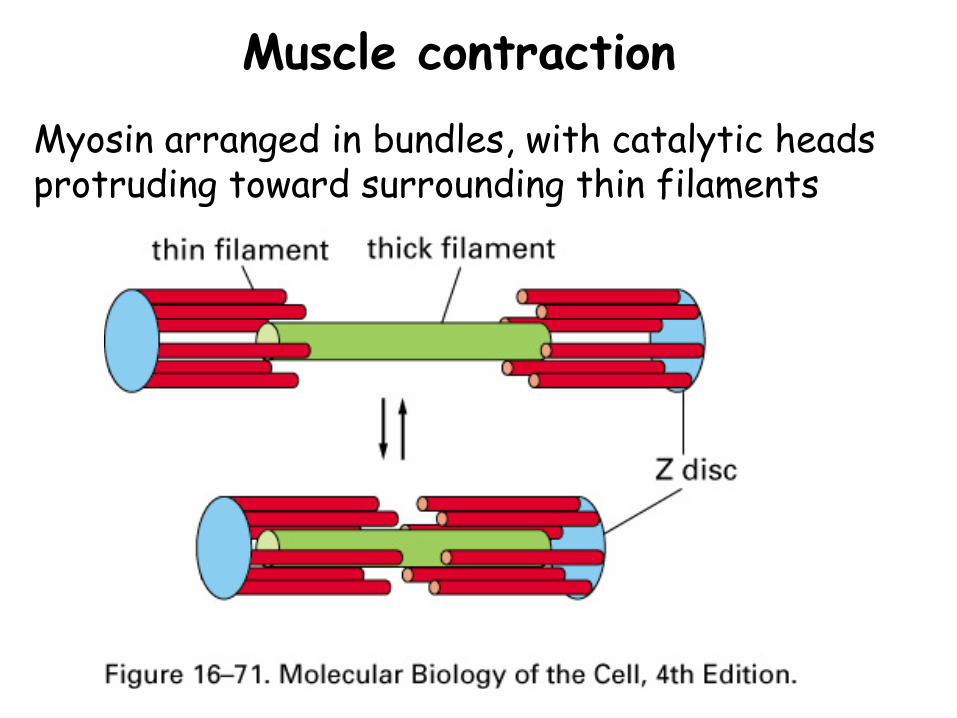
Muscle contractionMyosin arranged in bundles, with catalytic heads protruding toward surrounding thin filaments
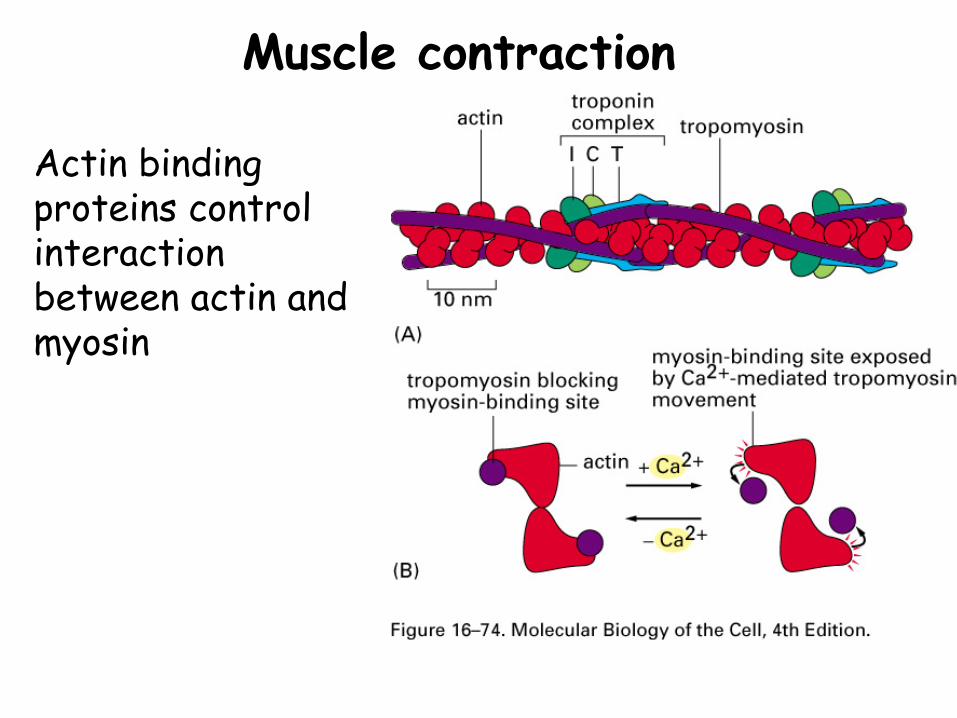
Muscle contraction
Actin binding proteins control interaction between actin and myosin

Cilia and flagella
Similar structures of 9 microtubule doublets and 2 microtubule singlets
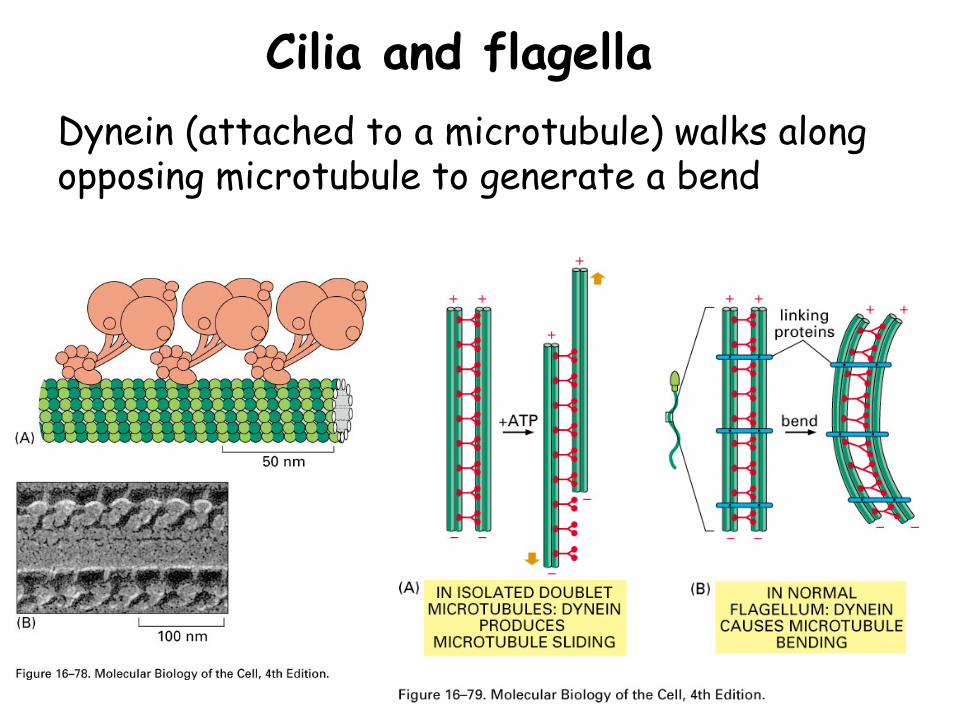
Cilia and flagellaDynein (attached to a microtubule) walks along opposing microtubule to generate a bend

Summary•Motor protein genes are ancient, with large gene families for most motor proteins
•Molecular motors use ATP hydrolysis to induce conformational changes that allow the motor to walk along a cytoskeletal filament
•Tail regions of motor proteins bind diverse cargo (organelles, vesicles, particles)
•Diversity in motor activity can be obtained by altering the nature of the cytoskeleton and the activity of the motor protein