changes of organelles associated with the differentiation...
TRANSCRIPT

/ . Embryol. exp. Morph. Vol. 43, pp. 107-121, 197S ]Ç)J Printed in Great Britain © Company of Biologists Limited 1978
Changes of organelles associated with the differentiation of epidermal
melanocytes in the mouse
By TOMOHISA HIROBE 1 AND T A K U J I TAKEUCHI 2
From the Biological Institute, Tohoku University, Japan
SUMMARY
Electron microscopic observations on normally differentiating and a-MSH (melanocyte-stimulating hormone)-treated epidermal melanocytes of newborn mouse skin were carried out. The process of melanocyte differentiation from premelanosome-containing melanoblasts was investigated in detail with respect to melanosomes as markers.
Melanoblasts containing unmelanized premelanosomes gradually decreased in number after birth, while the number of melanocytes rapidly increased. The epidermis of a-MSH-treated 3-day-old mice and normal 6-day-old mice contained melanocytes with numerous fully melanized melanosomes, and with no or only a few melanoblasts.
Changes in other organelles in differentiating melanocytes were also noticeable. Golgi apparatus and RER (rough endoplasmic reticulum) decreased in number during the normal or a-MSH-induced differentiation of the epidermal melanocytes, though the number of mitochondria showed no notable change. The number of SER (smooth endoplasmic reticulum) per cell did not change in the cells of newborn mice, while in a-MSH-treated cells the number increased significantly. These results led us to an assumption that Golgi apparatus or RER transforms into other forms of organelles including melanosomes and SER during the differentiation of melanocytes.
INTRODUCTION
We have reported that in the mouse differentiated melanocytes (the cells positive to the dopa reaction) appeared in the epidermis after birth and increased in number until 4 days of age (about 100 cells/0-1 mm2), then gradually decreased in number and disappeared by 30 days of age. On the other hand, the number of melanoblasts plus melanocytes (the cells positive to the combined dopa-premelanin reaction) remained constant (about 140 cells/0-1 mm2) until 4 days of age and then decreased (Hirobe & Takeuchi, 1977). The number of epidermal melanocytes was also shown to increase dramatically when newborn mice were injected with a-MSH (melanocyte-stimulating hormone). No change was observed in the number of melanoblasts plus melanocytes after the treat-
1 Author's address: Department of Biology, Faculty of Education, Iwate University, Ueda, Morioka, 020 Japan.
2 Author's address: Biological Institute, Tohoku University, Aoba-yama, Sendai, 980 Japan.

108 T. HIROBE AND T. TAKEUCHI
ment with a-MSH. Therefore, MSH seemed to induce differentiation in terms of initiation of melanogenesis in pre-existing melanoblasts.
The object of this study was to detect the effect of MSH on melanocytes at the level of organelle. Morphological change and changes in the number of melanosomes, Golgi apparatus including Golgi cisterna and Golgi vesicles, RER (rough endoplasmic reticulum), SER (smooth endoplasmic reticulum) and mitochondria in the normally differentiating and a-MSH-treated melanocytes were investigated in detail.
MATERIALS AND METHODS
Newborn C57BL/10J mice were injected subcutaneously at the dorsal side with a-MSH (a gift from Ciba-Geigy; 1 figjg BW) or Hanks' BSS (balanced salt solution) for control. After the treatment, pieces of skin were excised from the dorsal side of the animals. For electron microscopy, skins from 1-day-old, 3-day-old (Hanks' BSS-treated), 3-day-old (a-MSH-treated) and 6-day-old mice were fixed with 4 % glutaraldehyde in 0-1 M phosphate buffer (pH 7-4) for 2 h and washed with phosphate buffer and postfixed with 1 % osmium tetroxide in 0-1 M phosphate buffer (pH 7-4) for 2 h at 2-4 °C. After the fixation, they were dehydrated through a series of graded ethanols and embedded in Epon 812 (Luft, 1961). The ultra-thin sections were cut in an LKB Ultrotome 4802 A, stained with uranyl acetate and lead citrate (Reynolds, 1963), and examined with a Hitachi HS-9 electron microscope. All groups were triplicated and numerous blocks were examined with the electron microscope.
RESULTS
Changes in the maturation of melanosomes in the epidermal melanoblasts and melanocytes
In epidermis of 1-day-old, 3-day-old, 3-day-old (a-MSH-treated) and 6-day-old mice, the number of melanosomes of different stages and the percentage of
FIGURES 1 AND 2
Fig. 1. Electron micrograph of the epidermal melanoblast of 1-day-old mouse. Golgi apparatus (G) is well developed. Some stage-I melanosomes (small arrow) are seen. Insertion shows the Golgi area of another melanoblast of 1-day-old mouse. Stage-II melanosome (large arrow) is seen. RER, rough endoplasmic reticulum; SER, smooth endoplasmic reticulum; M, mitochondrion; C, centriole. x 23000 (insertion, x 53400).
Fig. 2. Electron micrograph of the epidermal melanocyte of 1-day-old mouse. All stages of melanosomes are seen. Insertion shows melanosomes of stages I-1V, ;;?l5 stage-I melanosome; m2, stage-II melanosome; m3, stage-Ill melanosome; w4, stage-IV melanosome; G, Golgi apparatus; RER, rough endoplasmic reticulum; M, mitochondrion; K, basal keratinocyte; BM, basement membrane, x23400 (insertion, x 50000).

Organelles in differentiating melanocytes 109
fi ,><gf
E MB 43

110 T. H I R O B E A N D T. T A K E U C H 1
100 -
80 -
60
40 -
2 0 -
<-> 0
80
(a) 1 -day-old .v = 21-79 N = 200
(h) 3-day-old (Hanks) -v = 54-82 N = 200
6 0 -
4 0 -
2 0 -
(c) 3-day-old (a-MSH) .v = 82-52 yV=200
"T" 20
- r -40
" T -
60
(ci) 6-day-old x = 80-87
TV=200
) 100 20
I + IV/l + ll + III + lV(%)
Fig. 3. Changes in the maturation of melanosome in the epidermal melanoblasts and melanocytes. The percentages of stage-Ill, -IV melanosomes against total melanosomes (I, II, III, IV) are shown, (a) 1-day-old mice; (b) Hanks' BSS-treated 3-day-old mice; (c) a-MSH-treated 3-day-old mice; (d) 6-day-old mice. The numbers of melanosomes were counted for 200 figures of nucleate cells in all groups, and the percentages per cell of stage-Ill, -IV melanosomes against total melanosomes were calculated.
FIGURES 4 AND 5
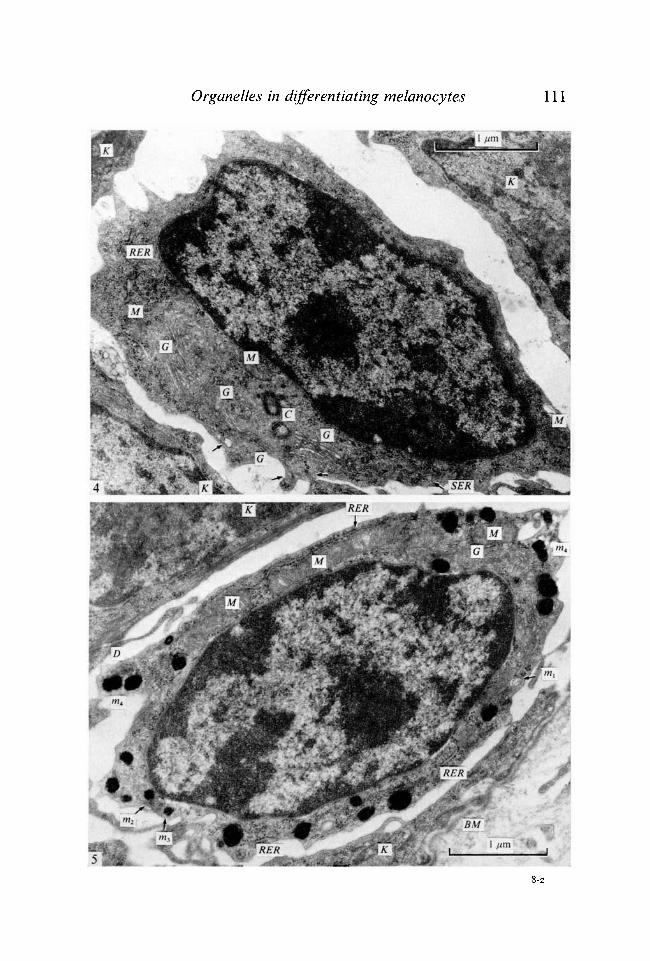
Fig. 4. Electron micrograph of the epidermal melanoblast of 3-day-old mice. Golgi apparatus (G) and RER are well developed. Some stage-I melanosomes (arrow) are seen. SER, smooth endoplasmic reticulum; M, mitochondrion; C, centrioles ; K, basal keratinocyte. x 28 000. Fig. 5. Electron micrograph of the epidermal melanocyte of 3-day-old mouse. Numerous melanized melanosomes are seen, m^ stage-I melanosome; m2, stage-11 melanosome; m3, stage-Ill melanosome; w4, stage-IV melanosome; G, Golgi apparatus; RER, rough endoplasmic reticulum; M, mitochondrion; K, basal keratinocyte; BM, basement membrane; D, desmosome. x 26800.
Organelles in differentiating melanocytes 111
w
RER
[SËR]
m Wà
V* %'k+. .**•*>
,'., - £-' ^ '*'
r;'t- W f r *:•«* *'
8-2

112 T. HIROBE AND T. TAKEUCHI
mature melanosomes were recorded for 200 electron micrographs of nucleate cells.
In the epidermis of 1-day-old mice, loose cells which possessed no desmosomes and clear cytoplasm distinct from other epidermal cells were observed. They contained well-developed Golgi apparatus, RER and some stage-I melanosomes (Fig. 1) and a few stage-II melanosomes (Fig. 1, insert). They seemed to be melanoblasts devoid of melanin formation. The stage of melanosome maturation was categorized according to Fitzpatrick, Hori, Toda & Seiji (1969): stage I and stage II include unmelanized immature premelanosomes, while melanized melanosomes are classified as stage III and IV. Melanocytes which possessed some stage-Ill, -IV melanosomes were also observed (Fig. 2), though cells with high melanosome maturation grade (III + IV/I + II + III + IV %) were few (Fig. 3).
In the epidermis of 3-day-old mice (Hanks' BSS-treated control) some undifferentiated melanoblasts which contained some stage-I melanosomes were observed (Fig. 4). Many melanocytes with melanized melanosomes were also found (Fig. 5). They contained many more stage-Ill, -IV melanosomes than the cells from 1-day-old mice. On the other hand, melanocytes from the a-MSH-treated mice (Fig. 6) contained many more stage-Ill, -IV melanosomes than the control. They contained no melanoblasts that had only stage-I, -II melanosomes (Fig. 3).
In the epidermis of 6-day-old mice, a few melanoblasts and mature melanocytes (Fig. 7) containing many more stage-Ill, -IV melanosomes were observed (Fig. 3). In the epidermis of 6-day-old mice, melanocytes were located in the granular layer (30 % of total melanocytes) and in the basal layer (70 % of total melanocytes) of the epidermis. This indicates that some differentiated melanocytes migrated to the granular layer, though most of the differentiated basal layer melanocytes migrated into hair follicles.
The percentages of melanoblasts of 1-day-old, 3-day-old (Hanks), 3-day-old (a-MSH) and 6-day-old mice were 52-5, 28, 0 and 4%, respectively. Thus, the proportion of melanoblasts in the melanoblast-melanocyte population in the epidermis decreased from 52-5 to 4% in the first 5 days after birth, and decreased from 52-5 to 0% in the first 2 days after the injection of a-MSH.
FIGURES 6 AND 7
Fig. 6. Electron micrograph of the epidermal melanocyte of a-MSH-treated 3-day-old mouse. Numerous mature melanosomes are seen. No Golgi apparatus are seen. mu stage-I melanosome; m2, stage-II melanosome; w3, stage-Ill melanosome; ra4, stage-lV melanosome; RER, rough endoplasmic reticulum; SER, smooth endoplasmic reticulum; M, mitochondrion, x 21000. Fig. 7. Electron micrograph of the epidermal melanocyte of 6-day-old mice. Numerous mature melanosomes are seen, but no Golgi apparatus. m4, stage-IV melanosome; RER, rough endoplasmic reticulum; SER, smooth endoplasmic reticulum; M, mitochondrion, x 12800.

Organelles in differentiating melanocytes 113
gg *isa
•t HP Ï€ -^W* » ^ •-». P E B I ' - M *
%.jEsa f t
• S ^ / i ^ -
114 T. HIROBE AND T. TAKEUCH1
2 0 -
15 -
10 -
5 -
1 1 1 1 3 6
Days after birth
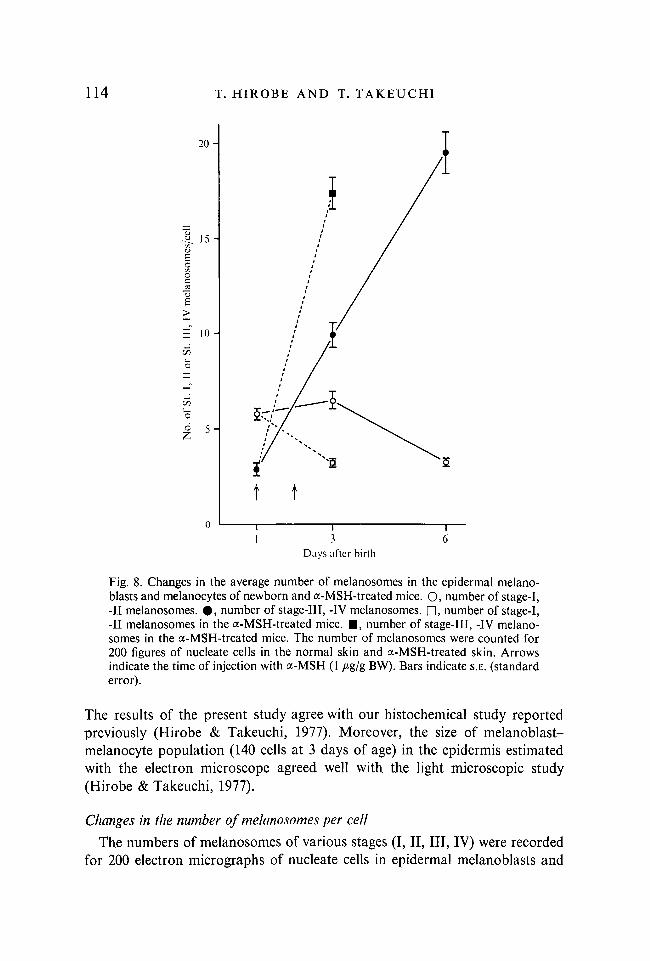
Fig. 8. Changes in the average number of melanosomes in the epidermal melanoblasts and melanocytes of newborn and a-MSH-treated mice. O, number of stage-I, -II melanosomes. • , number of stage-Ill, -IV melanosomes. D, number of stage-1, -II melanosomes in the a-MSH-treated mice. • , number of stage-Ill, -IV melanosomes in the a-MSH-treated mice. The number of melanosomes were counted for 200 figures of nucleate cells in the normal skin and a-MSH-treated skin. Arrows indicate the time of injection with a-MSH (1 /*g/g BW). Bars indicate S.E. (standard error).
The results of the present study agree with our histochemical study reported previously (Hirobe & Takeuchi, 1977). Moreover, the size of melanoblast-melanocyte population (140 cells at 3 days of age) in the epidermis estimated with the electron microscope agreed well with the light microscopic study (Hirobe & Takeuchi, 1977).
Changes in the number of melanosomes per cell
The numbers of melanosomes of various stages (I, II, III, IV) were recorded for 200 electron micrographs of nucleate cells in epidermal melanoblasts and

Organelles in differentiating melanocytes 115
1-4-
1-2 -
1-0-
£ 0-8 ^
a 0 - 6 -
0 -4 -
0 - 2 -
1 3
Days after birth Fig. 9. Changes in the average number of Golgi apparatus in the epidermal melano-blasts and melanocytes of newborn and a-MSH-treated mice. O, number of Golgi apparatus per melanoblast. • , number of Golgi apparatus per melanocyte. • , number of Golgi apparatus per melanocyte in the a-MSH-treated mice. The numbers of Golgi apparatus were counted for 200 figures of nucleate cells in the normal skin and the a-MSH-treated skin. Arrows indicate the time of injection with a-MSH (1 /tg/g BW). Bars indicate S.E.
melanocytes at 1 day, 3 days (Hanks' BSS control), 3 days (a-MSH) and 6 days of age (Fig. 8).
The number of melanosomes per cell increased markedly after birth; a-MSH accelerated the increase in the number of melanosomes. In melanoblasts from 1-day-old mice, stage-I and -II melanosomes were prominent. Then stage-Ill and -IV melanosomes increased drastically in number, while those of stages I and II decreased.
Changes in the numbers of Golgi apparatus, RER, SER and mitochondria
The numbers of Golgi apparatus, RER, SER and mitochondria were also recorded for 200 electron micrographs of nucleate cells in the epidermal melanoblasts and melanocytes at 1 day, 3 days (Hanks' BSS control), 3 days (a-MSH) and 6 days of age. A continuous and distinctive ER (endoplasmic reticulum)

116 T. HIROBE A N D T. T A K E U C H I
100 -
80 -
60 -
40 -
20 -
0 100 -
80 -
60 -
40 -
20 -
0 100
80
60
40
20
0 100
80
60
40
20
0
Melanocyte
(«) 1-day-old .Y = 0-8! N=95
(h) 3-day-old (Hanks) .7=0-45 JV=144
(c) 3-day-old (a-MSH) .7=0-19 7V=200
(cl) 6-day-old .7=0-28 JV=192
Melanoblast
.7=0-95 JV=105
.7=1-16 JV=56
No melanoblasl
.7=0-75 TV=8
I~I i~i 1 2 3 4 5 6 7 0 1 2 3 4
No. of Golgi apparatus/cell 5 6
Fig. 10. Decrease in the number of Golgi apparatus in the epidermal melanoblasts and melanocytes after birth and a-MSH treatment. The numbers of Golgi apparatus per cell are shown, (a) 1-day-old mice, left: melanocyte; right: melanoblast; (b) Hanks' BSS-treated 3-day-old mice, left: melanocyte; right: melanoblast; (c) a-MSH-treated 3-day-old mice, left: melanocyte; (d) 6-day-old mice, left; melanocyte; right: melanoblast. The numbers of Golgi apparatus were counted for 200 figures of nucleate cells in the normal skin and the a-MSH-treated skin.
in a cell was counted as one unit. An ER unit in which RER and SER are connected was counted as one RER.
As shown in Fig. 9, a remarkable change was detected in the number of Golgi apparatuses. They were numerous in the melanoblasts and melanocytes of 1-day-old mice. However, their number decreased rapidly in the melanocytes of 3-day-old (Hanks' BSS control) and 6-day-old mice, while the melanoblasts from 3-day-old mice (Hanks' BSS control) still contained numerous Golgi apparatus (Fig. 4). In the epidermal melanocytes of a-MSH-treated skin, a rapid decrease in the number of Golgi apparatus was also demonstrated (Fig. 6).

Organelles in differentiating melanocytes 117
8 -
6 -
4 -
t t
1 3
Days after birth
Fig. 11. Changes in the average number of RER in the epidermal melanoblasts and melanocytes of newborn and a-MSH-treated mice. O, number of RER per melanoblast. # , number of RER per melanocyte. • , number of RER per melanocyte in the a-MSH-treated mice. The numbers of RER were counted for 200 figures of nucleate cells in the normal skin and the a-MSH-treated skin. Arrows indicate the time of injection with a-MSH (1 /tg/g BW). Bars indicate S.E.
This suggests that Golgi apparatus tends to decrease in number as melanoblast differentiates and that MSH stimulates the decrease in the number of Golgi apparatus. Fig. 10 shows the histogram of the number of Golgi apparatus. No changes in the morphological features of Golgi apparatus, including the Golgi cisterna and Golgi vesicles, were observed.
Changes in the number of RER per cell were also recognized. As shown in Fig. 11, the average number of RER per cell decreased after birth. A similar result was obtained in the melanocytes of a-MSH-treated mouse skin. There seemed to be no change in morphological features of RER (Figs. 1, 2, 4-7).
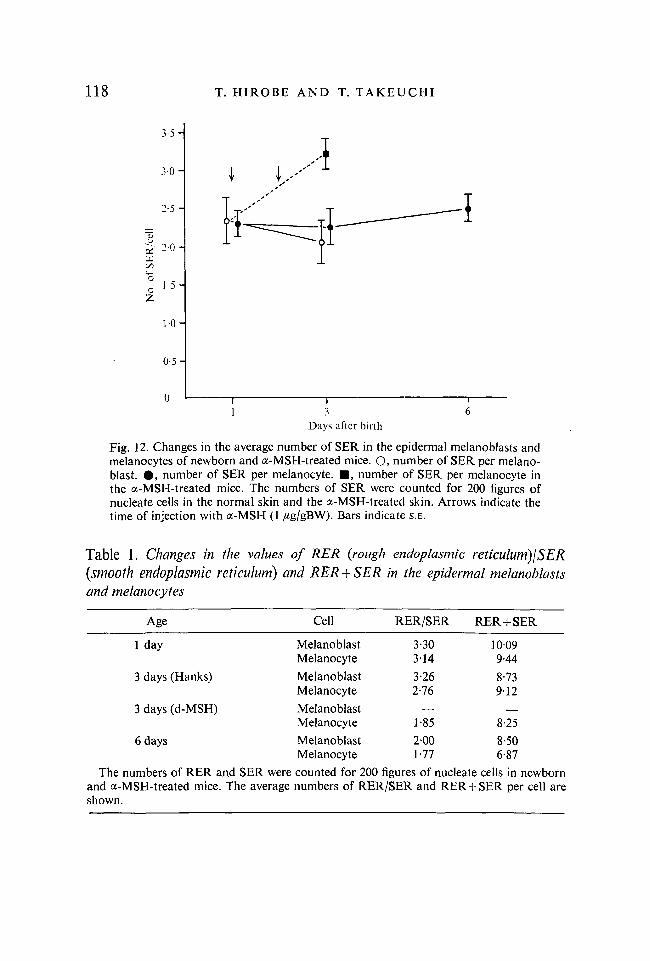
On the other hand, the number of SER per cell did not change in the epidermal melanoblasts and melanocytes after birth. The number was about 2*2-2-5/cell. In the a-MSH-treated melanocyte, however, a significant increase in the number of SER was revealed. The number was about 3-2 (Fig. 12). Values of RER/SER and of RER + SER are shown in Table 1. Both values decreased
118 T. HIROBE AND T. TAKEUCHI
3 - 5 -
3 0 -
2 0 -
1 0 -
0 - 5 -
1 1 1 1 3 6
Days after birth
Fig. 12. Changes in the average number of SER in the epidermal melanoblasts and melanocytes of newborn and a-MSH-treated mice. O, number of SER per melanoblast. • , number of SER per melanocyte. • , number of SER per melanocyte in the a-MSH-treated mice. The numbers of SER were counted for 200 figures of nucleate cells in the normal skin and the a-MSH-treated skin. Arrows indicate the time of injection with a-MSH (1 /ig/gBW). Bars indicate S.E.
Table 1. Changes in the values of RER (rough endoplasmic reticulum)!SER (smooth endoplasmic reticulum) and RER + SER in the epidermal melanoblasts and melanocytes
Age Cell RER/SER RER+SER
1 day Melanoblast Melanocyte
3-30 3-14
1009 9-44
3 days (Hanks) Melanoblast Melanocyte
3-26 2-76
8-73 912
3 days (d-MSH) Melanoblast Melanocyte 1-85 8-25
6 days Melanoblast Melanocyte
2-00 1-77
8-50 6-87
The numbers of RER and SER were and a-MSH-treated mice. The average shown.
counted for 200 figures of nucleate cells in newborn numbers of RER/SER and RER + SER per cell are

Organelles in differentiating melanocytes 119
in the 6 days after birth and after a-MSH treatment. This result indicates that RER decreases in the course of differentiation of melanocytes without any change in SER.
There was no marked difference in the numbers of mitochondria per melano-blast or melanocyte in 1-day-old, 3-day-old (Hanks' BSS control), 3-day-old (a-MSH) and 6-day-old mice. The number was about 3-5-4-5 per cell. No change in morphological features of mitochondria was detected (Figs. 1, 2, 4-7).
DISCUSSION
Our results show that many melanocytes with melanized melanosomes appeared 6 days after birth in the epidermis of C57BL/10J mice, while the number of melanoblasts with unmelanized stage-I, -II melanosomes decreased. The increase in the number of melanocytes was accelerated by treating with a-MSH. The increase in the number of melanocytes in the melanoblast-melanocyte population in the epidermis after birth and in the a-MSH-treated epidermis seems to be the result of induction of tyrosinase activity and rapid formation of melanosomes in the melanoblasts previously located in the epidermis. Therefore, our previous report (Hirobe & Takeuchi, 1977) of an increase in the number of dopa-positive melanocytes after birth and a drastic increase in the number of dopa-positive melanocytes after treatment with a-MSH agrees well with the present electron microscopic study. Melanin synthesis appears shortly after the formation of stage-I, -II melanosomes, and the number of melanosomes then rapidly increases, so that the formation of melanosomes and the synthesis of melanin occur together. The number of melanosomes per cell in the a-MSH-treated mice is the same as that of melanosomes in 6-day-old mice. The epidermal melanocytes in 6-day-old mice transfer many melanosomes into surrounding keratinocytes, so epidermal melanocytes probably possess a limited number of melanosomes per cell.
Weiss & Zelickson (1975) reported that immature melanoblasts which possessed premelanosomes were first identified in 15-day foetal C57BL/6J mice, and that epidermal melanocytes with both unmelanized and melanized melanosomes appeared from the 16th and 17th day of gestation, and then increased with increasing number of melanosomes. On the 4th day after birth, fewer melanocytes were present and epidermal melanocytes could not be identified at 18 days after birth. Their qualitative electron microscopic observation agrees with our present study.
Our present report also demonstrated that the numbers of Golgi apparatus, RER and SER seemed to relate to the formation of melanosomes during the differentiation of melanocytes.
The number of Golgi apparatus in the epidermal melanocytes decreased dramatically during the normal differentiation after birth and after a-MSH-treatment. The origin of Golgi apparatus in a cell is not established yet, though

120 T. HIROBE AND T. TAKEUCHI
it is supposed that the Golgi apparatus originates from SER (Claude, 1970; Whaley, 1975). On the other hand, Golgi apparatus has been considered to play the most important role in the formation of melanosomes (Seiji, Shimao, Birbeck & Fitzpatrick, 1963; Toda & Fitzpatrick, 1971; Eppig & Dumont, 1972). Although numerous Golgi vesicles were observed in the melanoblasts containing few premelanosomes, the number of vesicles decreased as melanosomes dramatically increased in number during normal and a-MSH-induced differentiation. Stage-I, -II melanosomes were observed near Golgi vesicles, while stage-Ill, -IV melanosomes were found far from Golgi apparatus. These observations seem to indicate that the Golgi vesicles are taking part in the formation of melanosomes as suggested by Novikoff, Albala & Biempica (1968).
We assume that in the process of melanocyte differentiation the formation of nascent Golgi cisterna of premelanosome-containing melanoblast is overwhelmed by the transformation process into vesicles, which in turn are incorporated in the nascent melanosomes. However, it should be emphasized that Golgi apparatus did not disappear but was merely reduced in number.
The number of RER per cell also decreased in the newborn melanocyte and in the a-MSH-treated melanocyte. In the epidermal melanoblasts and melanocytes, some stage-I and stage-II melanosomes were observed to connect with RER. We suppose that RER decreases in. number, producing many premelanosomes by budding in the process of melanocyte differentiation in the normal and the a-MSH-induced development. Indeed, Moyer (1963) and Stanka (1971) have proposed that melanosomes originates from RER. In addition, it is probable that a part of RER transforms itself into SER and then into melanosomes (Maul, 1969). The above-mentioned indication that the decline of Golgi apparatus and of RER associate the enhancement of melanosomes during melanocyte differentiation leads us to an assumption that melanosomes are formed of at least two components derived from Golgi vesicles and RER. This idea agrees well with the observation reported by one of us for phaeomelanosome formation (Sakurai, Ochiai & Takeuchi, 1975). Contrary to the changes in the numbers of Golgi apparatus and RER, SER did not change in number in 6 days after birth. The number of SER per cell increases only in the a-MSH-treated mice. It is probable that a part of Golgi cisterna remains as SER which does not yet take part in melanogenesis because of the limited processing capacity in the period observed. Therefore, the number of SER increases after the a-MSH treatment. We suppose that in the normal condition SER retains its number in the melanoblasts and melanocytes in the balance of loss and reproduction.
It has been reported that in the process of differentiation of liver cells (Parsa, 1974) and pancreatic ascinar cells (Parsa, Marsh & Fitzgerald, 1969), Golgi apparatus, RER and SER quantitatively change. These observations together with our results are examples of functional differentiation reflected in the changes of organelles.

Organelles in differentiating melanocytes 121
No change in the number of mitochondria was observed either in normal or in a-MSH-induced differentiation. The observed changes of the Golgi apparatus, RER and SER are therefore not due to artifacts.
This work was supported by Grant 244004 from the Ministry of Education.
REFERENCES
CLAUDE, A. (1970). Growth and differentiation of cytoplasmic membranes in the course of lipoprotein granule synthesis in the hepatic cell. I. Elaboration of elements of the Golgi complex. J. Cell Biol. 47, 745-766.
EPPIG, J. J. & DUMONT, J. N. (1972). Cytochemical localization of tyrosinase activity in pigmented epithelial cells of Rana pipiens and Xenopus laevis larvae. J. Ultrast. Res. 39, 397-410.
FITZPATRICK, T. B., HORI, Y., TODA, K. & SEIJI, M. (1969). Melanin 1969: some definitions and problems. Jap. J. Derm. Ser. B. 79, 278-282.
HIROBE, T. & TAKEUCHI, T. (1977). Induction of melanogenesis in the epidermal melano-blasts of newborn mouse skin by MSH. / . Embryol. exp. Morph. 37, 79-90.
LUFT, J. H. (1961). Improvement in epoxy resin embedding methods. / . biophys. biochem. Cytol. 9, 409-414.
MAUL, G. G. (1969). Golgi-melanosome relationship in human melanoma in vitro. J. Ultra-struct. Res. 26, 163-176.
MOYER, F. H. (1963). Genetic effects on melanosome fine structure and ontogeny in normal and malignant cells. Ann. N. Y. Acad. Sei. 100, 584-606.
NOVIKOFF, A. B., ALBALA, A. & BIEMPICA, L. (1968). Ultrastructural and cytochemical observation on B-16 and Harding-Passey mouse melanomas. The origin of premelano-somes and compound melanosomes. / . Histochem. Cytochem. 16, 299-319.
PARSA, I. (1974). Liver cell differentiation in a chemically defined medium. Morphologic and enzymatic comparisons of in utero rat liver and liver rudiment grown in organ culture. Am. J. Pathol. 76, 197-122.
PARSA, I., MARSH, W. H. & FITZGERALD, P. J. (1969). Pancreas acinar cell differentiation. I. Morphologic and enzymatic comparisons of embryonic rat pancreas and pancreatic anläge grown in organ culture. Am. J. Pathol. 57, 457-487.
REYNOLDS, E. S. (1963). The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. / . Cell Biol. 17, 208-212.
SAKURAI, T., OCHIAI, H. & TAKEUCHI, T. (1975). Ultrastructural change of melanosomes associated with agouti pattern formation in mouse hair. Devi Biol. 47, 466-471.
SEIJI, M., SHIMAO, K., BIRBECK, M. S. C. & FITZPATRICK, T. B. (1963). Subcellular localization of melanin biosynthesis. Ann. N.Y. Acad. Sei. 100, 497-533.
STANKA, P. (1971). Electronenmikroskopische Untersuchung über die Prämelanosomenent-stehung im retinalen Pigmentepithel von Hühnerembryonen. Z. Zellforsch. Mikrosk. Anat. 112, 120-128.
TODA, K. & FITZPATRICK, T. B. (1971). The origin of melanosomes. Biology of Normal and Abnormal Melanocytes, (ed. T. Kawamura, T. B. Fitzpatrick & M. Seiji), pp. 265-278. Tokyo: Univ. Tokyo Press.
WEISS, L. W. & ZELICKSON, A. S. (1975). Embryology of the epidermis: ultrastructural aspects. III. Maturation and primary appearance of dendritic cells in the mouse with mammalian comparisons. Acta Dermatovener. 55, 431-442.
WHALEY, W. G. (1975). The Golgi apparatus. Cell Biol. Monogr. 2, 1-178.
(Received 16 March 1977, revised 7 July 1977)