changes in peripheral blood lymphocyte subsets in elderly subjects are associated with an impaired...
TRANSCRIPT

Changes in peripheral blood lymphocyte subsets in elderlysubjects are associated with an impaired function of the
hypothalamic�/pituitary�/adrenal axis
Victor Martınez-Taboada a,b,*, Marıa Jose Bartolome b, Jose Antonio Amado c,Ricardo Blanco a, Maria Teresa Garcıa-Unzueta c,
Vicente Rodrıguez-Valverde a, Marcos Lopez-Hoyos b
a Services of Rheumatology, Immunology, and Endocrinology, Santander, Spainb Hospital Universitario Marques de Valdecilla, 39008 Santander, Spain
c Facultad de Medicina, Universidad de Cantabria, Santander, Spain
Received 10 May 2002; received in revised form 17 July 2002; accepted 9 September 2002
Abstract
A growing body of evidence indicates that ageing brings a progressive disruption in the immune and endocrine
systems. However, very few reports have correlated the changes in the immune system with the endocrine function in
the elderly. The aim of the present study was to investigate the changes occurring in the peripheral blood lymphocyte
subpopulations with age and correlate them with the hypothalamic�/pituitary�/adrenal (HPA) function. We determined
the peripheral blood lymphocyte phenotype and the T cell receptor usage by flow cytometry analysis. The HPA
function was evaluated by the basal serum levels of adrenal steroids and the response to stimulation with a low-dose
ACTH. In the elderly, we observed a decrease of major T subsets together with an increase of NK cells and activated T
cells. With regard to the HPA function, the most significant decline was found in dehydroepiandrosterone (DHEA) and
dehydroepiandrosterone-sulphate (DHEAS). A close correlation between immune changes with ageing and DHEA
response to ACTH stimulation was found. The present study showed an inverse correlation of lymphocyte changes with
the plasma levels of steroids, especially DHEA and its metabolite, DHEAS. This association was not found for other
steroids and points for the possibility of using DHEA to correct the immunological decline associated with ageing.
# 2002 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Ageing; Lymphocyte subsets; HPA axis; DHEA; T cell expansions
1. Introduction
Ageing is accompanied by a number of quanti-
tative and qualitative changes in the immune
response which are grouped by the term of
immunosenescence (Pawelec and Solana, 1997).* Corresponding author. Tel./fax: �/34-942-20-3453
E-mail address: [email protected] (V. Martınez-Taboada).
Mechanisms of Ageing and Development 123 (2002) 1477�/1486
www.elsevier.com/locate/mechagedev
0047-6374/02/$ - see front matter # 2002 Elsevier Science Ireland Ltd. All rights reserved.
PII: S 0 0 4 7 - 6 3 7 4 ( 0 2 ) 0 0 1 1 6 - 1

As a consequence, elderly individuals show anincreased susceptibility to neoplasias, infectious
diseases and autoimmune processes (Castle, 2000).
Most of the immune changes during the ageing
process have been extensively studied in mice
(Mountz et al., 2001). Recently, many reports
have addressed this issue in humans by different
approaches. Some of them have described immu-
nophenotypical changes in peripheral blood lym-phocyte subsets detected by flow cytometry, such
as an increase in CD3�/CD7�/ cells, a decrease of
naive and an increase of memory T cells, an
expansion of CD28�/ T cells or an increase of
NK cells (Malinowski and Rapaport, 1995; Fag-
noni et al., 1996; Ginaldi et al., 2001). Other
studies have reported a functional impairment of
lymphocytes in ageing: thymic involution, im-paired lymphocyte proliferative responses, in-
creased susceptibility of T cells to apoptosis,
impairment in type 2 cytokine production and
abnormal NK function (Mackall and Gress, 1997;
Aggarwal and Gupta, 1998; Sakata-Kaneko et al.,
2000; Bruunsgaard et al., 2000; Solana and Mar-
iani, 2000).
The immune system is not the only one thatsuffers from senescence. The endocrine system also
undergoes important changes during ageing. En-
docrinosenescence carries a substantial decline in
the hormone levels of at least three endocrine axes:
hypothalamic�/pituitary�/gonadal, hypothalamic�/
pituitary�/adrenal (HPA) and growth hormone-
insulin-like growth factor I (Roshan et al., 1999).
Among them, the HPA axis is the one that bestintegrates the neuroendocrine and immune system.
The HPA axis undergoes a number of age-related
changes that can play a possible role in the
occurrence of several age-associated pathological
conditions (Ferrari et al., 2001).
Despite the evidence indicating a close commu-
nication between the neuroendocrine and immune
systems, there have been quite few reports in thisfield (Straub et al., 1998). Specifically, it has not
been investigated if the changes in peripheral
blood lymphocyte subsets are related to the
impaired HPA axis. Thus, the aim of the present
study was to investigate the changes occurring in
the peripheral blood lymphocyte subpopulations
in the elderly and to correlate them with the serum
hormone levels of the HPA axis and with theadrenal reserve measured through the response of
those hormones to low-dose ACTH.
2. Materials and methods
2.1. Subjects and samples
Following informed consent, 34 healthy young
donors (14 male and 20 female; mean age: 24.89/
3.1 years; range: 20.1�/30.4) and 23 healthy aged
volunteers (nine male and 14 female; mean age:
73.09/8.0 years, range: 60.4�/94.3) were included in
the study. All the subjects were in healthy condi-
tion, according to accurate clinical investigations
and to hematological and biochemical parameters.None of the donors was taking drugs that could
affect the immune system or had a prior history of
neoplasia, infectious or autoimmune disease. Sam-
ples obtained from young healthy women were
taken during the early follicular phase. The study
received the approval of the ethical committee of
our hospital.
2.2. Flow cytometry
Blood samples for cytometric studies were
collected in tubes containing potassium-ethylene-
diamine tetraacetic acid (EDTA, Becton Dickin-
son, Plymouth, UK). Absolute counts of
lymphocytes were always calculated. Lymphocyte
counts and T subsets were calculated by two-
colour fluorescence flow cytometry using a panelof specific monoclonal antibodies (mAbs) for the
following cell surface proteins: CD3, CD4, CD8,
CD16, CD56 (Becton Dickinson, San Jose, CA),
CD45RA, CD45RO, HLA-DR (Pharmingen, San
Diego, CA), CD25, CD28, CD57 (Labgen, Lab-
clinics, Barcelona, Spain). The mAbs were con-
jugated to fluorescein isothiocyanate (FITC) or
phycoerythrin (PE). The panel of mAbs usedallowed us to study the major T lymphocyte
subsets (Table 1).
Percentages of CD4�/ and CD8�/ T lympho-
cytes bearing a specific variable b chain of the T
cell receptor (TCRBV) were obtained by staining
with specific mAbs against the following TCRBV
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/14861478

which represent more than 40% of total repertoire:
TCRBV2, TCRBV3, TCRBV5S2/S3, TCRBV5S1,
TCRBV6.7, TCRBV8, TCRBV12, TCRBV13,
TCRBV17 (T Cell Diagnostics, Woburu, MA).TCRBV-specific expansion was defined as
TCRBV specificities expressed at a frequency
greater than the mean plus 3 standard deviation
(S.D.) of the young healthy donors or as a value of
more than 20%.
Whole blood (100 ml) was incubated with a
combination of these mAbs for 30 min at room
temperature. Following incubation, the sampleswere lysed and fixed with 2 ml of FACS Lysing
Solution (Becton Dickinson). At least 20 000
lymphocytes per sample, gated by the FSC/SSC
parameters, were analysed using the CellQuest
software (Becton Dickinson). The cut-off for
positive fluorescence was set to includeB/1% of
negative control mouse IgG conjugates (Becton
Dickinson). Compensation levels were adjustedwith two paired mAbs and checked daily.
2.3. Hypothalamic�/pituitary�/adrenal function
To assess the HPA function, the low dose
Synacthen test (Ciba Laboratories, Horsham,
UK) was performed between 7.30 and 9.30 h in
25 young (nine males/16 females) and 16 elderly
healthy subjects (7/9). The test consisted of an
intravenous bolus of 1 mg of (1�/24) ACTH in thevolunteers, who rested for 15 min before starting
the test (Bridges et al., 1998). A baseline blood
sample was taken before the (1�/24) ACTH was
administered to measure blood levels of the
hormones described below. Additionally, blood
samples at 30 and 60 min after the bolus were
drawn for measuring the increase in the serumconcentrations of cortisol, androstenedione (ASD)
and dehydroepiandrosterone (DHEA). The incre-
ment in the steroids was calculated as the differ-
ence between peak and baseline levels (Hurel et al.,
1996).
2.4. Measurement of serum hormonal levels
Serum hormonal levels were quantified by
established specific direct RIAs: cortisol (Diagnos-
tic Products Corporation, Los Angeles, CA,
USA), 11-deoxycortisol (11DOC, ICN Pharma-
ceuticals, Costa Mesa, CA, USA), 17 a-hydro-
xyprogesterone (17OHP, Diagnostic Products
Corporation), ASD (Diasorin, Stillwater, MN,
USA), DHEA (Diagnostic Systems Laboratories,Webster, TX, USA) and its metabolite, dehydroe-
piandrosterone sulphate (DHEAS, Diagnostic
Products Corporation). Intra and interassay coef-
ficients of variation were below 11 and 13% in each
test, respectively. According to the manufacturer’s
data, the cross-reactivity among the different
hormones measured was always less than 2%.
2.5. Statistical analysis
Statistical analysis was performed with STATIS-
TICA software package (Macintosh, Apple Com-
puter Inc., Cupertino, CA, USA). Comparison of
variables between groups was done using Mann�/
Whitney test. Correlations were analysed by the
Spearman test with the Bonferroni correction.
3. Results
3.1. Peripheral blood lymphocyte subpopulations
Peripheral blood T lymphocytes (CD3�/) levels
were lower in aged compared with young subjects.
This decrease could be attributed to both majortypes of CD4�/ and CD8�/ T lymphocytes (Table
2). Elderly donors showed a significant increase in
the frequencies of NK cells (CD3�/CD16�/
CD56�/) and also T cells with a killer phenotype
(CD3�/CD16�/CD56�/). In this regard, although
T cells expressing the CD57 NK cell marker
Table 1
Definition of major lymphocyte subsets analysed in the present
studied according to their expression of clusters of differentia-
tion (CD)
T cell subset Characteristic CD
Naıve CD45RA�/CD45RO�/
Memory CD45RA�/CD45RO�/
Early activation CD25�/
Late activation HLADR�/
Cytolytic CD3�/CD16�/CD56�/
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/1486 1479

represent minor subpopulations of peripheral
blood T cells, they were also increased within
both CD4�/ and, especially, CD8�/cells in elderly
controls (Table 2).Although some other molecules have been
recently defined for naıve and memory T cells,
we employed CD45RA and CD45RO, respec-
tively, as cellular surface markers (Table 2; Vitetta
et al., 1991). Naıve CD4�/ T cells showed a
significant decrease in the elderly with respect to
young subjects (P B/0.05). Likewise, naıve CD8�/
T cells were also importantly decreased in the agedindividuals. On the other hand, blood counts of
memory CD4�/ T cells showed a remarkable
increase as compared with young healthy donors
(P B/0.05). However, absolute numbers of mem-
ory CD8�/ T cells did not show any relevant
difference between both groups (Table 2).
3.2. Frequencies of peripheral blood T cells
expressing activation molecules
The expression of CD25 and HLA-DR onCD4�/ and CD8�/ T cells was evaluated as an
indicator of the activation status (Rea et al., 1999;
Zola, 2000). No remarkable difference in the
marker of early activation was found in either
CD4�/ or CD8�/ T cells. Furthermore, the early
activated cells were rather low (Table 3). Regard-
ing late activation, significant differences between
young and aged subjects were observed. Thus,
proportions, but not absolute numbers, of CD4�/
HLA-DR�/ T cells increased in the elderly ascompared with the young population (P B/0.05).
Furthermore, within the CD8�/ T cell subset aged
individuals showed a higher percentage and abso-
lute number of cells expressing HLA-DR (Table
3).
As another marker of T cell function, frequen-
cies and absolute numbers of CD4�/ and CD8�/ T
cells expressing the costimulatory molecule CD28were determined. CD28 is a key molecule in the
successful activation of a T cell since its ligation
transduces the second signal (Chambers and
Allison, 1999). Both CD4�/CD28�/ and CD8�/
CD28�/ T cells were significantly decreased (P B/
0.05) in the elderly in comparison with healthy
young individuals (Table 3).
3.3. TCRBV expression on peripheral blood T cells
The TCRBV usage within the CD4�/ subset wasvery similar between elderly and young donors
(Fig. 1). However, a significant increase of CD8�/
T cells expressing TCRBV2 and BV8 was observed
in young individuals.
We defined a TCRBV expansion as greater than
the mean�/3 S.D. of the young data or as a
Table 2
Percentages and absolute numbers of the main peripheral blood lymphocyte subsets in healthy elderly subjects as compared with young
controls
Percentage (%) Absolute number (cells per mm3)
Elderly Young Elderly Young
CD3�/ 68.49/10.4* 74.19/6.56 11549/313* 14189/362
CD4�/ 409/8.7 429/6.6 6709/186* 8039/231
CD8�/ 28.49/8.9 32.19/6.4 4859/218* 6159/192
CD3�/CD16�/CD56�/ 14.39/7.2* 8.99/4.7
CD3�/CD16�/CD56�/ 3.89/4* 2.39/1.8
CD4�/CD57�/ 10.79/11* 2.39/2.5 669/64* 209/27
CD8�/CD57�/ 499/13.1* 23.89/12.9 2439/144* 1529/112
CD4�/CD45RA�/ 309/11.7* 48.69/8.7 2099/113* 3959/159
CD8�/CD45RA�/ 64.39/7.9* 70.79/9.3 3159/163* 4309/133
CD4�/CD45RO�/ 72.49/13.1* 50.99/10.1 4729/122* 4029/123
CD8�/CD45RO�/ 39.69/8.6* 31.99/10.3 1919/86 2009/94
*, P B/0.05 elderly compared with young controls.
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/14861480

frequency �/20%. Remarkably, elderly controls
had a higher frequency of expanded TCRBV
populations than young controls in peripheral
blood CD4�/ (52.2 vs. 5.9%, P B/0.001) and
CD8�/ (47.8 vs. 17.6%, P B/0.05) T cells.
The distribution of TCRBV-specific expansions
in each group of healthy donors is shown in Table
4. Within the young volunteers, two individuals
had TCRBV expansion in the CD4�/ subset. Each
of them displayed only one TCRBV expansion. In
contrast, twelve out of the 23 elderly donors
showed 13 expansions within the CD4�/ subset
(1, 1 expansion/individual), being TCRBV13.1 and
BV17 clearly overexpressed in elderly subjects
(Table 4). In young healthy controls, TCRBV-
specific expansions were more frequent in CD8�/
T cells than in the CD4�/ compartment. Six out of
the 34 individuals showed six TCRBV-specific
expansions (one expansion/individual). On theother hand, CD8�/ TCRBV-specific expansion
was a common finding, with almost half of the
elderly individuals showing one TCRBV expan-
sion.
3.4. Plasma levels of steroids in healthy elderly
subjects
As shown in Fig. 2, the lower basal concentra-
tions and responses to the ACTH test were found
Table 3
Percentages and absolute numbers of peripheral blood lymphocytes expressing activation molecules in healthy elderly subjects as
compared with young controls
Percentage (%) Absolute number (cells per mm3)
Elderly Young Elderly Young
CD4�/CD25�/ 4.99/2.4 4.19/2.7 349/20 349/26
CD8�/CD25�/ 1.39/0.8 1.39/1.3 69/4 79/7
CD4�/HLA DR�/ 3.39/1.9* 1.99/1.4 219/15 159/12
CD8�/HLA DR�/ 5.39/3.4* 2.29/1.5 249/18* 139/9
CD4�/CD28�/ 87.79/13.9* 97.89/2.7 5989/218* 7649/260
CD8�/CD28�/ 35.89/12.2* 62.19/13.8 1729/89* 3629/135
*, P B/0.05 elderly compared with young controls.
Fig. 1. Expression of TCRBV proteins on peripheral blood
CD4�/ (upper) and CD8�/ (lower) T cells of young (grey) and
elderly (white) healthy subjects. Bars represent mean percen-
tages and error bars represent the S.D. Asterisks denote
differences between the different groups with a P B/0.05
calculated by the Mann�/Whitney analysis.
Table 4
Specific TCRBV families expanded in peripheral blood CD4�/
and CD8�/ cells in healthy elderly subjects as compared with
young controls
CD4�/ cells CD8�/ cells
Elderly Young Elderly Young
BV2 1 �/ 1 1
BV3 �/ �/ 2 1
BV5S2/S3 2 1 2 �/
BV5S1 1 �/ 2 �/
BV6.7 �/ �/ 2 1
BV8 1 �/ �/ 1
BV12 1 �/ �/ �/
BV13.1 4 1 1 1
BV17 3 �/ 2 1
Total 13 2 12 6
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/1486 1481
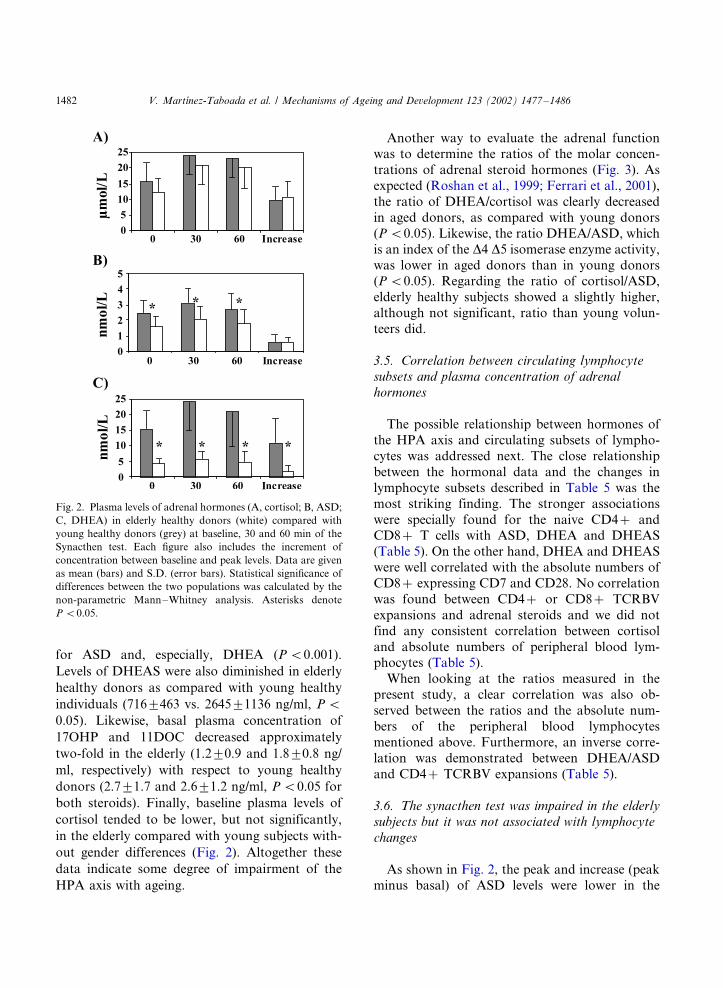
for ASD and, especially, DHEA (P B/0.001).
Levels of DHEAS were also diminished in elderly
healthy donors as compared with young healthy
individuals (7169/463 vs. 26459/1136 ng/ml, P B/
0.05). Likewise, basal plasma concentration of
17OHP and 11DOC decreased approximately
two-fold in the elderly (1.29/0.9 and 1.89/0.8 ng/
ml, respectively) with respect to young healthy
donors (2.79/1.7 and 2.69/1.2 ng/ml, P B/0.05 for
both steroids). Finally, baseline plasma levels of
cortisol tended to be lower, but not significantly,
in the elderly compared with young subjects with-
out gender differences (Fig. 2). Altogether these
data indicate some degree of impairment of the
HPA axis with ageing.
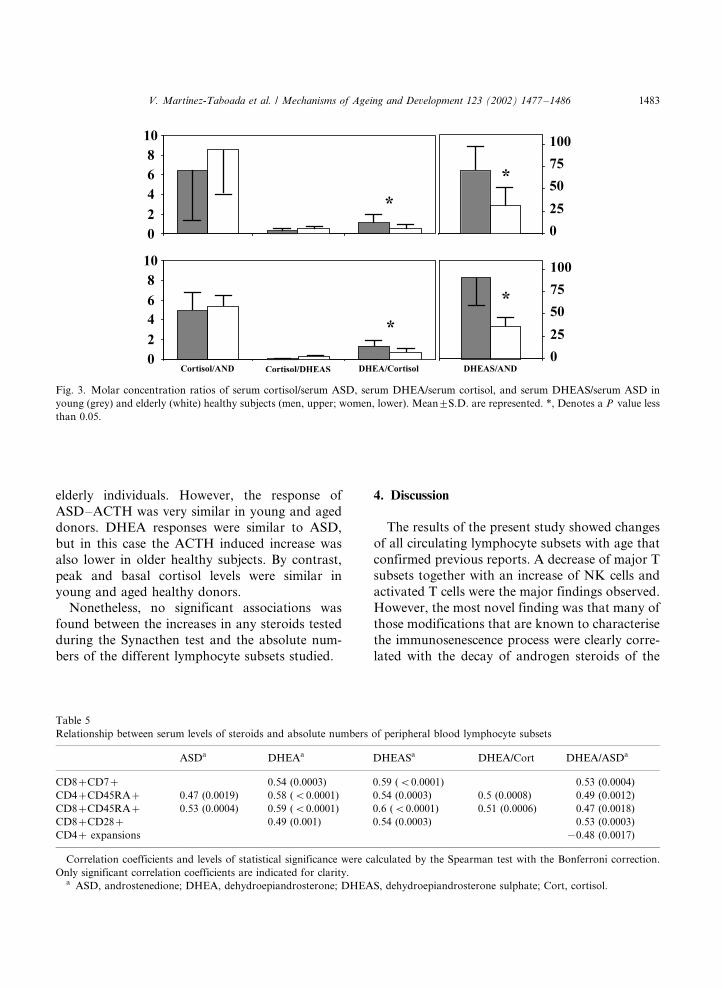
Another way to evaluate the adrenal functionwas to determine the ratios of the molar concen-
trations of adrenal steroid hormones (Fig. 3). As
expected (Roshan et al., 1999; Ferrari et al., 2001),
the ratio of DHEA/cortisol was clearly decreased
in aged donors, as compared with young donors
(P B/0.05). Likewise, the ratio DHEA/ASD, which
is an index of the D4 D5 isomerase enzyme activity,
was lower in aged donors than in young donors(P B/0.05). Regarding the ratio of cortisol/ASD,
elderly healthy subjects showed a slightly higher,
although not significant, ratio than young volun-
teers did.
3.5. Correlation between circulating lymphocyte
subsets and plasma concentration of adrenal
hormones
The possible relationship between hormones of
the HPA axis and circulating subsets of lympho-
cytes was addressed next. The close relationship
between the hormonal data and the changes in
lymphocyte subsets described in Table 5 was the
most striking finding. The stronger associations
were specially found for the naive CD4�/ andCD8�/ T cells with ASD, DHEA and DHEAS
(Table 5). On the other hand, DHEA and DHEAS
were well correlated with the absolute numbers of
CD8�/ expressing CD7 and CD28. No correlation
was found between CD4�/ or CD8�/ TCRBV
expansions and adrenal steroids and we did not
find any consistent correlation between cortisol
and absolute numbers of peripheral blood lym-phocytes (Table 5).
When looking at the ratios measured in the
present study, a clear correlation was also ob-
served between the ratios and the absolute num-
bers of the peripheral blood lymphocytes
mentioned above. Furthermore, an inverse corre-
lation was demonstrated between DHEA/ASD
and CD4�/ TCRBV expansions (Table 5).
3.6. The synacthen test was impaired in the elderly
subjects but it was not associated with lymphocyte
changes
As shown in Fig. 2, the peak and increase (peak
minus basal) of ASD levels were lower in the
Fig. 2. Plasma levels of adrenal hormones (A, cortisol; B, ASD;
C, DHEA) in elderly healthy donors (white) compared with
young healthy donors (grey) at baseline, 30 and 60 min of the
Synacthen test. Each figure also includes the increment of
concentration between baseline and peak levels. Data are given
as mean (bars) and S.D. (error bars). Statistical significance of
differences between the two populations was calculated by the
non-parametric Mann�/Whitney analysis. Asterisks denote
P B/0.05.
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/14861482
elderly individuals. However, the response of
ASD�/ACTH was very similar in young and aged
donors. DHEA responses were similar to ASD,
but in this case the ACTH induced increase was
also lower in older healthy subjects. By contrast,
peak and basal cortisol levels were similar in
young and aged healthy donors.
Nonetheless, no significant associations was
found between the increases in any steroids tested
during the Synacthen test and the absolute num-
bers of the different lymphocyte subsets studied.
4. Discussion
The results of the present study showed changes
of all circulating lymphocyte subsets with age that
confirmed previous reports. A decrease of major T
subsets together with an increase of NK cells and
activated T cells were the major findings observed.
However, the most novel finding was that many of
those modifications that are known to characterise
the immunosenescence process were clearly corre-
lated with the decay of androgen steroids of the
Fig. 3. Molar concentration ratios of serum cortisol/serum ASD, serum DHEA/serum cortisol, and serum DHEAS/serum ASD in
young (grey) and elderly (white) healthy subjects (men, upper; women, lower). Mean9/S.D. are represented. *, Denotes a P value less
than 0.05.
Table 5
Relationship between serum levels of steroids and absolute numbers of peripheral blood lymphocyte subsets
ASDa DHEAa DHEASa DHEA/Cort DHEA/ASDa
CD8�/CD7�/ 0.54 (0.0003) 0.59 (B/0.0001) 0.53 (0.0004)
CD4�/CD45RA�/ 0.47 (0.0019) 0.58 (B/0.0001) 0.54 (0.0003) 0.5 (0.0008) 0.49 (0.0012)
CD8�/CD45RA�/ 0.53 (0.0004) 0.59 (B/0.0001) 0.6 (B/0.0001) 0.51 (0.0006) 0.47 (0.0018)
CD8�/CD28�/ 0.49 (0.001) 0.54 (0.0003) 0.53 (0.0003)
CD4�/ expansions �/0.48 (0.0017)
Correlation coefficients and levels of statistical significance were calculated by the Spearman test with the Bonferroni correction.
Only significant correlation coefficients are indicated for clarity.a ASD, androstenedione; DHEA, dehydroepiandrosterone; DHEAS, dehydroepiandrosterone sulphate; Cort, cortisol.
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/1486 1483

HPA axis with age. Although correlation does notprove causal relationship, it suggests that this line
of evidence should merit further investigation.
The results of the blood immune parameters of
the present report are mostly in agreement with
previous works (Malinowski and Rapaport, 1995;
Fagnoni et al., 1996; Ginaldi et al., 2001). As
expected, both CD4�/ and CD8�/ blood T cells
significantly decreased in elderly donors (Ginaldiet al., 2001). We also found a significant increase
of blood lymphocytes expressing CD57 in elderly
subjects, especially in the CD8�/ subset. In this
regard, the loss of naive T cells associated with
ageing is particularly observed in the CD8�/ T cell
compartment (Fagnoni et al., 2000). However, we
did not find a deeper reduction of blood naive
CD8�/ T cells in aged subjects, probably becausethe use of CD45RA as the only marker of naive
cells underestimates the cells with such phenotype
(Rabin et al., 1995). In agreement with other
authors (Kukel et al., 1994; Fagnoni et al., 2000;
Ginaldi et al., 2000), we observed a decrease of
CD45RA�/ T cells with ageing associated with a
marked increase of T cells expressing CD45RO,
HLA-DR and CD57, which suggest a memoryphenotype (Kukel et al., 1994).
Our elderly population showed a marked reduc-
tion in both percentages and absolute numbers of
circulating CD28�/ T cells, especially within the
CD8�/ subset (Fagnoni et al., 1996; Boucher et al.,
1998; Nociari et al., 1999). This finding suggests
that ageing may affect the T cell activation path-
way, since CD28 is an essential costimulatorymolecule for a successful T cell activation (Cham-
bers and Allison, 1999). Thus, the particular
reduction of CD28 on the CD8 compartment
could explain the higher susceptibility to suffer
from viral infections and neoplasias, where CD8�/
T cells constitute important effector elements
(Zinkernagel, 1996). In disagreement with this
hypothesis, polyclonal activation experimentshave proved a functional integrity of the CD28
pathway in the elderly (Vidan et al., 1999).
Additional evidence in the present study for the
age-related disturbance in T cell homeostasis was
the higher frequency of TCRBV-specific expan-
sions in the CD8�/ subset (LeMaoult et al., 2000).
In a similar way to the altered expression of
surface molecules especially in the CD8�/ com-partment, it is possible that the decreased periph-
eral T cell repertoire diversity in the elderly makes
the immune system to recruit a very limited set of
T cell clones against an antigenic challenge. As
commented above, this restricted repertoire of
CD8�/ cells helps to understand the higher
susceptibility to disease in the elderly. In contrast,
young individuals with a conserved thymic outputrespond to an antigen by recruiting many T cell
specificities (LeMaoult et al., 2000). On the other
hand, despite previous studies have found quite a
few expansions within the CD4�/ compartment
(Posnett et al., 1994), the elderly population in the
present work showed an increased frequency of
TCRRBV-specific expansions in the CD4�/ T
cells. A possible explanation for this finding canbe found in the negative correlation observed
between CD4�/ TCRBV expansions and the
ration DHEA/ASD. It is possible that the aged
population studied in the present study would be
especially deficient for those steroids and could not
regulate the TCRBV expansions after repeated
stimuli.
Although there was some trend to decrease inthe secretion of cortisol, the most significant
decline was found for the androgen steroids of
the HPA axis, mainly DHEA and DHEAS (Be-
langer et al., 1994; Parker et al., 2000). Such
deficiency has been shown to be due to an
alteration in the zona reticularis of the adrenal
cortex (Parker et al., 2000; Giordano et al., 2001).
Plasma ASD concentration was lower in theelderly than in young subjects but this diminished
sensitivity of ASD synthetizer enzymes with age
was normalised by low-dose exogenous ACTH.
However, DHEA response to exogenous ACTH
was lower in aged subjects. This finding indicates
that low-dose exogenous ACTH stimulation is not
able to normalise DHEA synthesis, and confirms
that the D5 steroid pathway, which transforms 17-hydroxypregnenolone to DHEA, is the most
seriously affected by age (Parker et al., 2000).
Interestingly enough, the steroids under investi-
gation have been found to have a modulating
effect on the immune system (Khorram et al.,
1997; Solerte et al., 1999). It has been suggested
that the reduced levels of cortisol may be respon-
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/14861484

sible in part of the T cell expansions observed in
elderly, since clonal deletion of peripheral T cells
in vivo is induced by glucocorticoids (Gonzalo et
al., 1993). On the other hand, DHEA administra-
tion is able to increase in the elderly the number of
blood T cells expressing activation and NK
markers, such as CD25 or CD56 (Khorram et
al., 1997). Besides, DHEAS also enhances the NK
cell cytotoxicity in aged individuals (Solerte et al.,
1999). However, in our aged population the
increased expression of activation markers on
circulating T cells and the augmented numbers of
peripheral blood NK cells were inversely corre-
lated with the plasma concentration of both
DHEA and its metabolite, DHEAS (data not
shown). Since DHEA and glucocorticoids seem
to have opposite actions on the immune system
(Daynes and Araneo, 1990), it is possible that
those discrepancies could be due to a counter-
action of cortisol, which was almost kept in our
aged population as compared with young donors.
While in men DHEA and DHEAS can be pro-
duced by both the adrenals and the testes, post-
menopausal origin of androgens in women is
exclusively adrenal (Couzinet et al., 2001). There-
fore, these women may become critically depen-
dent on adrenal androgens to regulate peripheral
blood lymphocyte subsets.
In summary, the present study shows that
ageing is accompanied by changes in the distribu-
tion of peripheral blood lymphocytes and that
many of these changes keep a good correlation
with the plasma levels of steroids, especially
DHEA and its metabolite, DHEAS. Furthermore,
present data points at the possibility that admin-
istration of DHEA would be used to correct the
immunophenotypical alterations induced by age.
Acknowledgements
This work was partially supported by grants
from the ‘Fundacion Marques de Valdecilla’ (1998
and 2000) and FIS (FIS98/0846), Spain. We are
very grateful to Barbara Garcıa for English
revision of the manuscript.
References
Aggarwal, S., Gupta, S., 1998. Increased apoptosis of T cell
subsets in aging humans: altered expression of Fas (CD95),
Fas Ligand, Bcl-2 and Bax. J. Immunol. 160, 1627�/1637.
Belanger, A., Candas, B., Dupont, A., Cusan, L., Diamond, P.,
Gomez, J.L., Labrie, F., 1994. Changes in serum concentra-
tions of conjugated and unconjugated steroids in 40�/80
year old men. J. Clin. Endocrinol. Metab. 79, 1086�/1090.
Boucher, N., Dufeu-Duchesne, T., Vicaut, E., Farge, D.,
Effros, R.B., Schachter, F., 1998. CD28 expression in T
cell aging and human longevity. Exp. Gerontol. 33, 267�/
282.
Bridges, N.A., Hindmarsh, P.C., Pringle, P.J., Honour, J.W.,
Brook, C.G.D., 1998. Cortisol, androstenedione (A4),
dehydroepiandrosterone sulphate (DHEAS) and 17 hydro-
xyprogesterone (17OHP) responses to low doses of (1-24)
ACTH. J. Clin. Endocrinol. Metab. 83, 3750�/3753.
Bruunsgaard, H., Pedersen, A.N., Schroll, M., Skinhoj, P.,
Pedersen, B.K., 2000. Proliferative responses of blood
mononuclear cells (BMNC) in a cohort of elderly humans:
role of lymphocyte phenotype and cytokine production.
Clin. Exp. Immunol. 119, 433�/440.
Castle, S.C., 2000. Clinical relevance of age-related immune
dysfunction. Clin. Infect. Dis. 31, 578�/585.
Couzinet, B., Meduri, G., Lecce, M.G., Young, J., Brailly, S.,
Loosfelt, H., Milgrom, E., Schaison, G., 2001. The post-
menopausal ovary is not a major androgen-producing
gland. J. Clin. Endocrinol. Metab. 86, 5060�/5066.
Chambers, C.A., Allison, J.P., 1999. Costimulatory regulation
of T cell function. Curr. Opin. Cell Biol. 11, 203�/210.
Daynes, R.A., Araneo, B.A., 1990. Contrasting effects of
glucocorticoids on the capacity of T cells to produce the
growth factor interleukin 2 and interleukin 4. Eur. J.
Immunol. 19, 2319�/2325.
Fagnoni, F.F., Vescovini, R., Mazzola, M., Bologna, G.,
Nigro, E., Lavagetto, G., Franceschi, C., Passeri, M.,
Sansoni, P., 1996. Expansion of cytotoxic CD8�/CD28�/
T cells in healthy ageing people, including centenarians.
Immunology 88, 501�/507.
Fagnoni, F.F., Vescovini, R., Passeri, G., Bologna, G.,
Pedrazzoni, M., Lavagetto, G., Casti, A., Franceschi, C.,
Passeri, M., Sansoni, P., 2000. Shortage of circulating naive
CD8�/ T cells provides new insights on immunodeficiency
in aging. Blood 95, 2860�/2868.
Ferrari, E., Cravello, L., Muzzoni, B., Casarotti, D., Paltro, M.,
Solerte, S.B., Fioravanti, M., Cuzzoni, G., Pontiggia, B.,
Magri, F., 2001. Age-related changes of the hypothalamic�/
pituitary�/adrenal axis: pathophysiological correlates. Eur.
J. Endocrinol. 144, 319�/329.
Ginaldi, L., De Martinis, M., Modesti, M., Loreto, F., Corsi,
M.P., Quaglino, D., 2000. Immunophenotypical changes of
T lymphocytes in the elderly. Gerontology 46, 242�/248.
Ginaldi, L., De Martinis, M., D’Ostilio, A., Marini, L., Loreto,
F., Modesti, M., Quaglino, D., 2001. Changes in the
expression of surface receptors on lymphocyte subsets in
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/1486 1485

the elderly: quantitative flow cytometric analysis. Am. J.
Hematol. 67, 63�/72.
Giordano, R., Di Vito, L., Lanfranco, F., Broglio, F., Benso,
A., Gianotti, L., Grottoli, S., Ghigo, E., Arvat, E., 2001.
Elderly subjects show severe impairment of dehydroepian-
drosterone sulphate and reduced sensitivity of cortisol and
aldosterone response to the stimulatory effect of ACTH (1-
24). Clin. Endocrinol. 55, 259�/265.
Gonzalo, J.A., Gonzalez-Garcia, A., Martinez, C., Kroemer,
G., 1993. Glucocorticoid-mediated control of the activation
and clonal deletion of peripheral T cells in vivo. J. Exp.
Med. 177, 1239�/1246.
Hurel, S.J., Thompson, C.J., Watson, M.J., Harris, M.M.,
Baylis, P.H., Kendall-Taylor, P., 1996. The short Synacthen
and insulin stress tests in the assessment of the
hypothalamic�/pituitary�/adrenal axis. Clin. Endocrinol.
44, 141�/146.
Khorram, O., Vu, L., Yen, S.S., 1997. Activation of immune
function by dehydroepiandrosterone (DHEA) in age-ad-
vanced men. J. Gerontol. A Biol. Sci. Med. Sci. 52, M1�/
M7.
Kukel, S., Reinhold, U., Oltermann, I., Kreysel, H.W., 1994.
Progressive increase of CD7�/ T cells in human blood
lymphocytes with ageing. Clin. Exp. Immunol. 98, 163�/168.
LeMaoult, J., Messaoudi, I., Manavalan, J.S., Potvin, H.,
Nikolich-Zugich, D., Dyall, R., Szabo, P., Weksler, M.E.,
Nikolich-Zugich, J., 2000. Age-related dysregulation in
CD8 T cell homeostasis: kinetics of a diversity loss. J.
Immunol. 165, 2367�/2373.
Mackall, C.L., Gress, R.E., 1997. Thymic aging and T-cell
regeneration. Immunol. Rev. 160, 91�/102.
Malinowski, K., Rapaport, F.T., 1995. Effects of aging upon
the expression of differentiation and class II MHC antigens
on the surface of T lymphocytes from normal human
subjects. Cell. Immunol. 162, 68�/73.
Mountz, J.D., Van Zant, G.E., Zhang, H.G., Grizzle, W.E.,
Ahmed, R., Williams, R.W., Hsu, H.C., 2001. Genetic
dissection of age-related changes of immune function in
mice. Scand. J. Immunol. 54, 10�/20.
Nociari, M.M., Telford, W., Russo, C., 1999. Postthymic
development of CD28�/CD8�/ T cell subset: age-associated
expansion and shift from memory to naive phenotype. J.
Immunol. 162, 3327�/3335.
Parker, C.R., Soctt, M.S., Azziz, R., Crabbe, S.L., Hines, G.A.,
Boots, L.R., Bae, S., 2000. Effects of aging on adrenal
function in the human: responsiveness and sensitivity of
adrenal androgens and cortisol to adrenocorticotropin in
premenopausal and postmenopausal women. J. Clin. En-
docrinol. Metab. 85, 48�/54.
Pawelec, G., Solana, R., 1997. Immunosenescence. Immunol.
Today 18, 514�/516.
Posnett, D.N., Sinha, R., Kabak, S., Russo, C., 1994. Clonal
populations of T cells in normal elderly humans: the T cell
equivalent to ‘benign monoclonal gammapathy’. J. Exp.
Med. 179, 609�/618.
Rabin, R.L., Roederer, M., Maldonado, Y., Petru, A., Herzen-
berg, L.A., Herzenberg, L.A., 1995. Altered representation
of naive and memory CD8 T cell subset in HIV-infected
children. J. Clin. Invest. 95, 2054�/2060.
Rea, I.M., McNerlan, S.E., Alexander, H.D., 1999. CD69,
CD25, and HLA-DR activation antigen expression on
CD3�/ lymphocytes and relationship to serum TNF-alpha,
IFN-gamma, and sIL-2R levels in aging. Exp. Gerontol. 34,
79�/93.
Roshan, S., Nader, S., Orlander, P., 1999. Review: ageing and
hormones. Eur. J. Clin. Invest. 29, 210�/213.
Sakata-Kaneko, S., Wakatsuki, Y., Matsunaga, Y., Usui, T.,
Kita, T., 2000. Altered Th1/Th2 commitment in human
CD4�/ T cells with ageing. Clin. Exp. Immunol. 120, 267�/
273.
Solana, R., Mariani, E., 2000. NK and NK/T cells in human
senescence. Vaccine 18, 1613�/1620.
Solerte, S.B., Fioravanti, M., Vignati, G., Giustina, A.,
Cravello, L., Ferrari, E., 1999. Dehydroepiandrosterone
sulfate enhances natural killer cell cytotoxicity in humans
via locally generated immunoreactive insulin-like growth
factor I. J. Clin. Endocrinol. Metab. 84, 3260�/3267.
Straub, R.H., Konecna, L., Hrach, S., Rothe, G., Kreutz, M.,
Scholmerich, J., Falk, W., Lang, B., 1998. Serum dehy-
droepiandrosterone (DHEA) and DHEA sulfate are nega-
tively correlated with serum interleukin-6 (IL-6), and
DHEA inhibits IL-6 secretion from mononuclear cells in
man in vitro: possible link between endocrinosenescence and
immunosenescence. J. Clin. Endocrinol. Metab. 83, 2012�/
2017.
Vidan, M.T., Fernandez-Gutierrez, B., Hernandez-Garcia, C.,
Serra, J.A., Ribera, J.M., Perez-Blas, M., Regueiro, J.R.,
Banares, A., Jover, J.A., 1999. Functional integrity of the
CD28 co-stimulatory pathway in T lymphocytes from
elderly subjects. Age Ageing 28, 221�/227.
Vitetta, E.S., Berton, M.T., Burger, C., Kepron, M., Lee, W.T.,
Yin, X.M., 1991. Memory B and T cells. Ann. Rev.
Immunol. 9, 487�/498.
Zinkernagel, R.M., 1996. Immunology taught by viruses.
Science 271, 173�/178.
Zola, H., 2000. Markers of cell lineage, differentiation and
activation. J. Biol. Regul. Homeostatic Agents 14, 218�/219.
V. Martınez-Taboada et al. / Mechanisms of Ageing and Development 123 (2002) 1477�/14861486