cell surface heparan sulfate proteoglycans from … surface heparan sulfate proteoglycans from human...
TRANSCRIPT

THE JOURNAL 0 1992 by The American Society for Biochemistry and
OF BIOLOGICAL CHEMISTRY Molecular Biology, Inc.
Vol . 267, No. 28, Issue of October 5, PP 20435-20443,1992 Printed in U. S A.
Cell Surface Heparan Sulfate Proteoglycans from Human Vascular Endothelial Cells CORE PROTEIN CHARACTERIZATION AND ANTITHROMBIN I11 BINDING PROPERTIES*
(Received for publication, March 23, 1992)
Griet MertensS, Jean-Jacques Cassiman, Herman Van den Berghe, Jos Vermylent, and Guido Davidll From the Center for Human Genetics and the WCenter for Thrombosis and Vascular Research, University of Leuuen, B-3000 Leuuen, Belgium
Human aortic endothelial cells (HAEC) and human umbilical vein endothelial cells (HUVEC) were labeled with “SO:- for 48 h. The membrane-associated proteo- glycans were solubilized from these monolayers with detergent and purified by ion-exchange chromatogra- phy on Mono Q , incorporation in liposomes, and gel filtration. The liposome-intercalated proteoglycans were ‘251-iodinated and treated with heparitinase be- fore SDS-polyacrylamide gel electrophoresis. Radio- labeled proteins with apparent molecular masses of 130, 60, 46, 35, and 30 kDa (HAEC) and 180, 130, 62, 43, and 35 kDa (HUVEC) were detected by auto- radiography. Further characterization by affinity chromatography on immobilized monoclonal antibod- ies and by Northern blot analysis provided evidence for the expression of syndecan, glypican, and fibrogly- can in human endothelial cells. Most of the heparan sulfate which accumulated in the subendothelial ma- trix was implanted on a 400-kDa core protein. This protein was immunologically related to perlecan and bound to fibronectin. Binding studies on immobilized antithrombin I11 suggested that all membrane-associ- ated heparan sulfate proteoglycan forms had the ca- pacity to bind to antithrombin I11 but that high affinity binding was more typical for glypican. Most of the proteoglycans isolated from the extracellular matrix also bound only with low affinity to antithrombin 111. These results imply that glypican may specifically con- tribute to the antithrombotic properties of the vascular wall.
Heparan sulfate proteoglycans are found almost ubiqui- tously on the cell surface and in the extracellular matrix of mammalian cells. A wide range of biological functions, includ- ing growth control, cell adhesion, and anticoagulant activity,
*These investigations were supported by Grants 3.0066.87 and 3.0073.91 from the “Nationaal Fonds voor Wetenschappelijk Onder- zoek” of Belgium, by an award from the Bekales Foundation, and by the Inter-university Network for Fundamental Research sponsored by the Belgian Government (1987-1991). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Recipient of a fellowship from the “Instituut tot aanmoediging van het Wetenschappelijk Onderzoek in Nijverheid en Landbouw.”
7 Research Director of the “Nationaal Fonds voor Wetenschappe- lijk Onderzoek” of Belgium. To whom correspondence should be addressed: Center for Human Genetics, University of Leuven, Cam- pus Gasthuisberg, 0 & N6, Herestraat 49, B-3000 Leuven, Belgium. Tel.: 32-16-215863; Fax: 32-16-215997.
have been ascribed to the heparan sulfate proteoglycans (for review see Kjell6n and Lindahl, 1991). The heparan sulfate chains play a crucial role in most of the biological properties of these proteoglycans. Heparan sulfate can bind to extracel- lular matrix components such as laminin, fibronectin, colla- gen, and thrombospondin (for review see Jackson et al., 1991), is essential for the binding and signal transduction of growth factors (Rapraeger et al., 1991; Yayon et al., 1991), and anchors several enzymes, including lipoprotein lipase to the cell layer (Saxena et al., 1990). The most extensively studied interaction of heparin and heparan sulfate, however, is that with anti- thrombin 111. High affinity binding to this proteinase inhibitor which results in an anticoagulant effect depends on the pres- ence of a specific pentasaccharide sequence in these glycosa- minoglycans (Thunberg et al., 1982). The structural require- ments for the interactions of heparan sulfate with other proteins are not known but seem less stringent (Bengtsson et al., 1980).
The core proteins, on the other hand, target the glycosa- minoglycans to their strategic positions on the cell surface and in the extracellular matrix and determine their accumu- lation and turnover at these sites. Several core proteins, which constitute different gene products, have been described. Lung fibroblasts for example synthesize a 400-kDa heparan sulfate proteoglycan core protein which binds to fibronectin and accumulates in the extracellular matrix (Heremans et al., 1990). This proteoglycan is related to perlecan, the proteogly- can which accumulates in basement membranes, but is not related to the cell surface-associated heparan sulfate proteo- glycans of these cells which are characterized by core proteins of 125 kDa, 80 kDa (syndecan), 64 kDa (glypican), 48 kDa (fibroglycan) and 35 kDa (Lories et al., 1989). Glypican and its associated heparan sulfate, which are membrane-linked through a GP1’-anchor structure, are cleared from the cell surface by shedding, whereas all other cell surface proteogly- cans appear subject to endocytosis (David et al., 1990).
Taking into account the structural heterogeneity of the core proteins and the multiplicity of functions ascribed to the heparan sulfate chains, the question arises whether function- ally distinct heparan sulfate chains may occur on different core proteins. In this respect the endothelial cell system is of particular interest, since endothelial proteoglycans appear involved in specific biological functions which include the
The abbreviations used are: GPI, glycosyl phosphatidylinositol; HAEC, human aortic endothelial cells; HUVEC, human umbilical vein endothelial cells; GdnHC1, guanidinium chloride; mAb, mono- clonal antibody; PBS, phosphate-buffered saline; HS, heparan sul- fate; CS, chondroitin sulfate; PG, proteoglycan; SDS, sodium dodecyl sulfate; Mops, 3-(N-morpholino)propanesulfonic acid kb, kilo- base(s).
20435

20436 Endothelial Heparan Sulfate Proteoglycans
anchorage and transcytosis of lipoprotein lipase (Shimada et al., 1981; Saxena et al., 1991), the binding of basic fibroblast growth factor, possibly as a “supply” for the high affinity fibroblast growth factor receptor (Saksela et al., 1988), and an antithrombotic activity (Marcum et al., 1986). Recently, two distinct membrane-associated heparan sulfate proteogly- cans that bind antithrombin I11 have been isolated from cloned rat microvascular endothelial cells (Kojima et al., 1992a, 1992b), whereas earlier studies on venous and aortic vascular endothelial cells had identified a heparan sulfate proteoglycan with a core protein of 350 kDa which accumu- lates in the extracellular matrix (Kinsella and Wight, 1988; Saku and Furthmayr, 1989; Lindblom et al., 1989). We have extended these studies and report the isolation and charac- terization of several distinct heparan sulfate proteoglycans from human aortic and umbilical vein endothelial cells. Be- sides the previously described matrix-associated heparan sul- fate proteoglycan, we identified an important heparan sulfate fraction which could associate with artificial liposomes. These heparan sulfate chains were carried by several different hy- drophobic core proteins that were related to the membrane- associated core proteins previously isolated from human lung fibroblasts. Binding studies on immobilized antithrombin I11 suggested that all membrane-associated heparan sulfate pro- teoglycans, to some extent, can bind antithrombin I11 with high affinity but that glypican may bind preferentially. Like most of the cell surface proteoglycans, the heparan sulfate proteoglycan that accumulated in the subendothelial matrix displayed a low binding affinity for antithrombin 111.
MATERIALS AND METHODS
Cell Culture-Human aortic endothelial cells HAEC (AG09979, Institute for Medical Research, Camden, NJ) were grown on gelatin (0.1%)-coated dishes in Medium 199 (Flow Laboratories) supple- mented with 20% (v/v) fetal calf serum, 50 pg/ml of heparin, and 50 pg/ml of ECGF (Sigma).
Human umbilical vein endothelial cells (HUVEC) were prepared according to the procedure of Jaffe et al. (1973). The cells were grown on tissue-culture flasks coated with gelatin (0.1%) in Medium 199 containing 2 mM L-glutamine, 100 units/ml of penicillin, 100 pg/ml of streptomycin, 20 mM Hepes, 10% fetal calf serum, and 10% human serum. As a control for the endothelial origin of the cultured cells, a preparation was assayed for von Willebrand Factor-related antigen by using an immunofluorescence assay as described by Hoyer et al. (1973). The cells were cultivated for a maximum of two passages.
Confluent cultures (-4 X lo4 cells/cm2) were labeled for 48 h with 5 pCi (1.8 X lo5 Bq) of carrier-free H,35S04 (Du Pont-New England Nuclear)/ml of culture medium (25 ml of medium/175-cm2 culture flask).
Isolation of the Cell Surface Proteoglycans-Confluent cell layers (-3 X IO7 cells/experiment) were rinsed three times with cold phos- phate-buffered saline (PBS). Rinsed endothelial cells were extracted with Triton X-100 buffer (0.5% Triton X-100, 10 mM Tris/HCl, pH 8.0, 150 mM NaC1) containing 50 mM 6-aminohexanoic acid, 10 mM EDTA, 5 mM N-ethylmaleimide, 5 mM benzamidine, 1 mM phenyl- methanesulfonyl fluoride, and 1 pg/ml of pepstatin A as proteinase inhibitors (Lories et al., 1986). The detergent extract was cleared by centrifugation (10,000 X g; 60 min), concentrated by adsorption on DEAE-Trisacryl M, and eluted in urea buffer (6 M urea, 0.5% (v/v) Triton X-100,50 mM Tris/HCl, pH 8) containing 750 mM NaC1. This eluate was submitted to ion-exchange chromatography on a Mono Q HR 5/5 column (Pharmacia LKB Biotechnology Inc.). After eluting the bulk of the bound proteins with a 0-0.6 M linear NaCl gradient (18 mM/ml) in urea buffer (Lories et al., 1987), the retained proteo- glycans were rinsed with urea buffer containing 50 mM octyl glucoside instead of Triton X-100, eluted in 4 M GdnHCl buffer (see below) containing 75 mM octyl glucoside and proteinase inhibitors, and incorporated into lipid vesicles as described before (Lories et al., 1987). The proteoglycan/liposome mixture was fractionated on a (1 X 100 cm) Sepharose CL4B column in 4 M GdnHCl buffer (4 M GdnHC1, 100 mM 6-aminohexanoic acid, 10 mM EDTA, 10 mM N- ethylmaleimide, 5 mM benzamidine, 50 pg/ml of bovine serum albu-
min, 10 bg/ml of heparin, 10 pg/ml of chondroitin sulfate, and 50 mM sodium acetate, pH 5.8) at a flow rate of 3 ml/h, to separate hydro- phobic liposome-associated proteoglycans from nonhydrophobic pro- teoglycans and free glycosaminoglycan chains. Excluded liposome- proteoglycan complexes were dissociated by addition of Triton X-100 (0.5% (v/v) final concentration) and rechromatographed at a flow rate of 3 ml/h on a (1 X 100 cm) Sepharose CL4B column in 4 M GdnHCl buffer containing 0.5% (v/v) Triton X-100 (Lories et al., 1987).
Isolation of the Matrix-associated HSPG-The heparan sulfate proteoglycans from the extracellular matrix that remained attached to the culture flasks after extraction of the cell layers with detergent were solubilized in 6 M GdnHCl and purified by CsCl density gradient centrifugation in 4 M GdnHCl, ion-exchange chromatography on Mono Q in urea buffer, and gel filtration over Sepharose C1-4B in 4 M GdnHCl buffer, as described by Heremans et al. (1988).
Glycosaminoglycan Zdentifi~ation-~~SOi- incorporated in heparan sulfate and chondroitin sulfate was measured by cetylpyridinium chloride precipitation of samples degraded by thermolysin, chondroi- tinase ABC, and nitrous acid, pH 1.5, and control samples, as de- scribed by Lories et al. (1986).
Radiolabeling and Zmmunopurification of the Proteoglycans-Puri- fied cell surface and matrix proteoglycan samples (from -3 x IO7 cells) in 4 M GdnHCl buffer were dialyzed against urea buffer, adsorbed on 200 pl of DEAE-Trisacryl M beads, and radioiodinated using NaIz5I and chloramine T as described by Lories et al. (1987). The further immunopurification of these proteoglycans on immobi- lized monoclonal antibodies was performed as described before (Lor- ies et al., 1989).
Enzyme Digestions-Digestion of the ’*’I-labeled proteoglycans with heparitinase, with chondroitinase ABC (Seikagaku Kogyo Co.) or with both enzymes, was done as before (Lories et al., 1987). For treatment with phosphatidylinositol-specific phospholipase C (Boeh- ringer Mannheim), the proteoglycan samples were dissolved in 50 mM octyl glucoside, 10 mM EDTA, 50 mM Tris/HCl, pH 7.4, and incubated with 50 milliunits of enzyme for 3 h a t 37 “C.
Gel Electrophoresis and Western Blotting-Gel electrophoresis in 4-10% SDS-polyacrylamide gradient gels was done as described by Lories et al. (1987). Electrophoresis in SDS-agarose gels (4% Nusieve agarose, Pharmacia Fine Chemicals, Uppsala, Sweden) was performed in buffer containing 90 mM Tris, 90 mM borate, 2 mM EDTA, 0.1% SDS, pH 8.0, for 3 h at 50 V.
In blotting experiments, the separated materials were electrotrans- ferred from the gel to a Zeta-probe membrane (Bio-Rad) (15 h, 30 V). After transfer and inactivation for 1 h in PBS with 0.5% casein and 600 mM NaC1, the membrane was incubated with the appropriate monoclonal antibodies in PBS with 0.5% casein. The membrane was then rinsed three times in PBS containing 0.5% casein and 600 mM NaCl and further incubated for 1 h with alkaline phosphatase- conjugated rabbit anti-mouse immunoglobulins (Promega), diluted 1/ 5000 in PBS which contained 0.5% casein. After three washes in PBS which contained 0.3% Tween 20, and two washes in 1 mM MgClz, 50 mM bicarbonate, pH 9.5, the membrane was incubated with the chemiluminescent substrate 3-(2’-spiroadamantane)-4-methoxy-4- (3”-phosphoryloxy)phenyl-1,2-dioxethane disodium salt (AMPPD) (Tropix, Bedford, MA) (10 p1 of AMPPD in 1 ml of bicarbonate buffer) for 5 min and submitted to autoradiography.
Monoclonal Antibodies-The monoclonal antibodies (mAb) used for these experiments were all raised against proteoglycans isolated from normal human fetal lung fibroblasts.
mAbs 2E9, 6G12, and S1 are directed against the core proteins of different cell surface HSPGs of human lung fibroblasts (Lories et al., 1989). The mAbs 4B1, 9C9, 3H8, and 7H9, in contrast, all recognize the same matrix-associated HSPG (Heremans et al., 1989).
Fibronectin Binding Assay-The fibronectin binding assay was performed as described by Heremans et al. (1990). A 10-pg aliquot of purified human plasma fibronectin (GIBCO-BRL) was submitted to SDS-electrophoresis to separate the fibronectin from possible con- taminants in the sample and electroblotted to a nitrocellulose mem- brane. The membrane was inactivated in PBS, 0.5% casein and incubated overnight at 4 “C with heparitinase-digested ’261-labeled heparan sulfate proteoglycan from the subendothelial matrix. After rinsing, the membrane was submitted to autoradiography to localize any bound proteoglycan core and stained with polyclonal rabbit anti- fibronectin and peroxidase-conjugated swine anti-rabbit antibodies (Dakopatts, Glostrup, Denmark) to detect the position of fibronectin.
Affinity Chromatography on Antithrombin III-Antithrombin I11 was purified from human plasma as described by Miller-Anderson et
Endothelial Heparan
al. (1974) and covalently linked to CNBr-activated Sepharose C1-4B (3 mg/ml) in the presence of acetylated heparin. Free heparan sulfate chains, prepared by p-elimination of proteoglycans isolated from the culture medium of mouse mammary epithelial cells and known to contain the J-O-s,ulfated glucosamine residues that mark the anti- thrombin-binding region (Pejler and David, 1987), were used to test the binding properties of the immobilized antithrombin. The 35S- labeled glycosaminoglycans were applied to the antithrombin column (8 ml of gel) in 50 mM NaCl in assay buffer (10 mM Tris/HCl, pH 7.4, containing 10 pg/ml of acetylated bovine serum albumin and 0.1% Triton X-100). After an overnight incubation at 4 "C, the column was washed with 50 mM NaCl in assay buffer (10 column volumes) and eluted at a flow rate of 9 ml/h with a 0.05-2 M NaCl gradient (15 mM/ml) in assay buffer. Materials eluting above 0.4 M NaCl were concentrated, reapplied (after dialysis) to the column, and eluted as before. Nearly 95% of this "high affinity" glycosaminoglycan fraction reeluted above 0.4 M NaCl, indicating that the binding and elution properties of these materials were specific.
'"1-Labeled endothelial proteoglycan samples (representing the isolate from -3 X 10' cells, and containing -1.8 pg of HS or the equivalent of -45 pmol of HS chains) were fractionated on the antithrombin column as described above. Alternatively, the radioio- dinated cell surface proteoglycans (from -3 X lo7 cells) or the 35S04- labeled proteoglycans from 100 ml of conditioned culture medium were immunopurified, dissolved in 300 p1 of 50 mM NaCl in assay buffer, and mixed with 300 pg of the immobilized antithrombin. After an overnight incubation at 4 "C, the beads were washed and stepwise eluted using 4 column volumes of, successively, 0.05, 0.2, 0.4, and 2 M NaCl in assay buffer. When tested in this assay, more than 90% of the free HS chains (from mouse mammary epithelial cells) with high affinity for antithrombin I11 were eluted in the 2 M step. In both binding assays the immobilized antithrombin was present in 5-10 X 103-fold molar excess over the amount of HS added.
Isolation of RNA and Northern Blot Analysis-The poly(A)+ RNA from -3 X lo7 cultured human umbilical vein endothelial cells was extracted in 5 M guanidinium isothiocyanate and isolated by phenol/ chloroform extraction in the presence of sodium acetate (0.1 M), two isopropanol precipitations, and oligo(dT)-cellulose chromatography as described before (Marynen et al., 1989). Aliquots of 3 pg of denatured poly(A)+ RNA/lane were submitted to electrophoresis in 1.2% agarose gels containing 0.2 M Mops, 0.05 M sodium acetate, pH 7.0, 10 mM EDTA, and 6% formaldehyde and further analyzed as described by David et al. (1990).
RESULTS
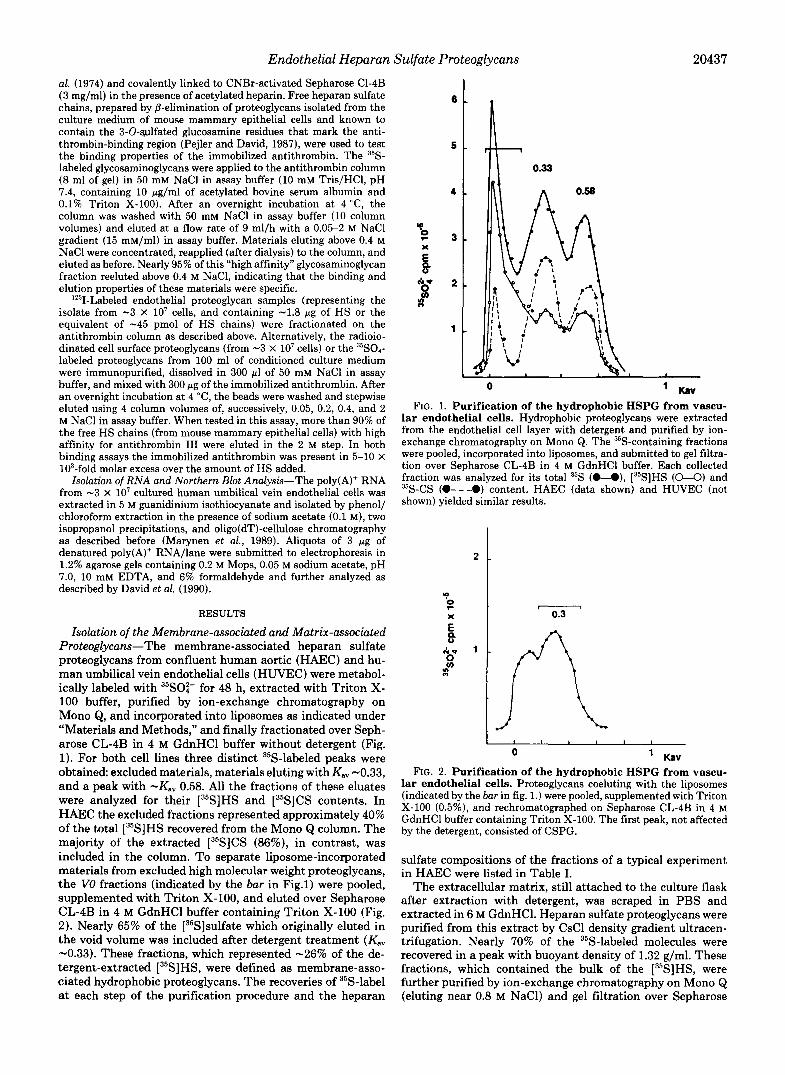
Isolation of the Membrane-associated and Matrix-associated Proteoglycans-The membrane-associated heparan sulfate proteoglycans from confluent human aortic (HAEC) and hu- man umbilical vein endothelial cells (HUVEC) were metabol- ically labeled with ""SO:- for 48 h, extracted with Triton X- 100 buffer, purified by ion-exchange chromatography on Mono Q, and incorporated into liposomes as indicated under "Materials and Methods," and finally fractionated over Seph- arose CL-4B in 4 M GdnHCl buffer without detergent (Fig. 1). For both cell lines three distinct 35S-labeled peaks were obtained excluded materials, materials eluting with K,, -0.33, and a peak with -Kav 0.58. All the fractions of these eluates were analyzed for their [35S]HS and [35S]CS contents. In HAEC the excluded fractions represented approximately 40% of the total [35S]HS recovered from the Mono Q column. The majority of the extracted [35S]CS (86%), in contrast, was included in the column. To separate liposome-incorporated materials from excluded high molecular weight proteoglycans, the VO fractions (indicated by the bar in Fig.1) were pooled, supplemented with Triton X-100, and eluted over Sepharose CL-4B in 4 M GdnHCl buffer containing Triton X-100 (Fig. 2). Nearly 65% of the [35S]sulfate which originally eluted in the void volume was included after detergent treatment (KaV -0.33). These fractions, which represented -26% of the de- tergent-extracted [35S]HS, were defined as membrane-asso- ciated hydrophobic proteoglycans. The recoveries of 35S-label at each step of the purification procedure and the heparan
Sulfate Proteoglycans 20437
4
..
E 8
$j- 2
f 1
0 ' Kav FIG. 1. Purification of the hydrophobic HSPG from vascu-
lar endothelial cells. Hydrophobic proteoglycans were extracted from the endothelial cell layer with detergent and purified by ion- exchange chromatography on Mono Q. The 35S-containing fractions were pooled, incorporated into liposomes, and submitted to gel filtra- tion over Sepharose CL-4B in 4 M GdnHCl buffer. Each collected fraction was analyzed for its total 35S (O-O), [35S]HS (U) and 35S-CS (O---O) content. HAEC (data shown) and HUVEC (not shown) yielded similar results.
2 .
In - 0.3
E 8
R-J 1 In s n
I 1 I a
0 ' Kav FIG. 2. Purification of the hydrophobic HSPG from vascu-
lar endothelial cells. Proteoglycans coeluting with the liposomes (indicated by the bar in fig. 1.) were pooled, supplemented with Triton X-100 (0.5%), and rechromatographed on Sepharose CL-4B in 4 M GdnHCl buffer containing Triton X-100. The first peak, not affected by the detergent, consisted of CSPG.
sulfate compositions of the fractions of a typical experiment in HAEC were listed in Table I.
The extracellular matrix, still attached to the culture flask after extraction with detergent, was scraped in PBS and extracted in 6 M GdnHCl. Heparan sulfate proteoglycans were purified from this extract by CsCl density gradient ultracen- trifugation. Nearly 70% of the 35S-labeled molecules were recovered in a peak with buoyant density of 1.32 g/ml. These fractions, which contained the bulk of the [%]HS, were further purified by ion-exchange chromatography on Mono Q (eluting near 0.8 M NaCl) and gel filtration over Sepharose

20438 Endothelial Heparan Sulfate Proteoglycans TABLE I
Isolation of the endothelial membrane-associated proteoglycans Recoveries of "S-label and [3sS]HS composition of the fractions
during the purification of the hydrophobic proteoglycans from HAEC. The detergent extract of the cells was concentrated by absorption on DEAE-Trisacryl M. The bound label was eluted and further purified by ion-exchange chromatography over Mono Q. Labeled materials requiring salt concentrations >0.6 M for elution from Mono Q were eluted with 4 M GdnHCl and octyl glucoside, mixed with phosphati- dylcholine, and dialyzed. The label-liposome mixture recovered at the end of the dialysis was fractionated by gel filtration chromatography over Sepharose C1-4B in 4 M GdnHCl in the absence of Triton X- 100. The excluded label which coeluted with the liposomes was mixed with Triton X-100 and rechromatographed over Sepharose CL-4B in the presence of Triton X-100. The label which became included as a result of the detergent treatment was defined as hydrophobic proteo- glycan. The heparan sulfate composition of the fractions is expressed as a percentage of the total 3sS-glycosaminoglycan in that particular fraction.
Step "S recovery Composition
CPm % HS Triton X 100-extraction 4,708,000 DEAE concentration
Fall-through 3,419,000 Eluted 1,134,000
Eluted <0.6 M 141,000 Eluted >0.6 M 785,000 52
Liposome incorporation 760,000 Gel filtration 1 (-Triton
Mono Q
x-100) Included 283,000 Excluded 108,000 76
Gel filtration 2 (+Triton x-100)
Excluded 22,900 Included 42,500 >95
CL-4B in 4 M GdnHCl buffer (eluting with K,, < 0.1) (not shown). From the recovery at this stage it was calculated that up to 30% of the heparan sulfate proteoglycans could be extracted from the cell layer with detergent, whereas nearly 70% were associated with the extracellular matrix.
Similar results were obtained for HUVEC (not shown). Based on the yield of [3sS]HS at the final step and on the specific activity of the SO:- in the culture medium (1.1 x lo7 dpm/pmol), we calculated that -1.8 pg of HS were recovered as membrane-associated proteoglycan from -3 x lo7 HUVEC.
Characterization of the Matrix-associated Heparan Sulfate Proteoglycans-The heparan sulfate proteoglycans isolated from the HAEC or HUVEC matrix were labeled with "'1, and heparitinase-treated and nontreated iodinated samples were submitted to SDS-agarose electrophoresis, followed by trans- fer to a Zeta-probe membrane and autoradiography. Nonre- duced digested samples showed a band of M, -400,000. Treat- ing the samples with 100 mM P-mercaptoethanol (10 min at 100 "C in the presence of 1% SDS) decreased the mobility of this band (Fig. 3).
Immunostaining of a Western blot of unreduced matrix heparan sulfate proteoglycans, using a mixture of monoclonal antibodies raised against the perlecan-related matrix HSPG of human lung fibroblasts (Heremans et al., 1989), visualized the same high molecular weight band as detected by autora- diography (Fig. 4).
Further evidence for similarities between the matrix-asso- ciated HSPG from human fibroblasts and vascular endothelial cells was found by functional analysis. In prior studies we had identified a high affinity interaction between fibronectin and the core protein of the HSPG which is associated with the extracellular matrix of human lung fibroblasts (Heremans et
-200
- 97 -68
- + + - FIG. 3. Characterization of the matrix-associated HSPG
from vascular endothelial cells. Similar amounts of '*"I-labeled matrix-associated HSPG from HUVEC were incubated with (+) or without (-) heparitinase, submitted to SDS-polyacrylamide gradient gel electrophoresis under reducing ( R ) or nonreducing ( U R ) condi- tions (5 X lo3 cpm/lane), and detected by autoradiography. Similar results were obtained for HAEC (not shown).
- + Mr x 10-
-200
- 97 -68
FIG. 4. Immunological cross-reactivity of the matrix-asso- ciated HSPG from vascular endothelial cells. Nontreated (-1 and heparitinase digested (+) matrix-associated HSPG from HUVEC were analyzed by SDS-polyacrylamide gel electrophoresis, followed by electrotransfer to a Zeta-probe membrane and immunostaining with a mixture of mAbs raised against the large HSPG isolated from the extracellular matrix of human lung fibroblasts (Heremans et al., 1989). Similar results were obtained for HAEC (not shown).
al., 1990). To test for similar properties of the core of the matrix-associated HSPG in endothelial cell cultures, human plasma fibronectin was fractionated on a 4% Nusieve agarose gel and electrotransferred to a nitrocellulose membrane. After inactivation with casein, the membrane was incubated with heparitinase-digested 12'I-labeled HSPG from the extracellu- lar matrix of HUVEC. Autoradiography showed a binding of the radiolabeled core protein to the immobilized fibronectin (Fig. 5).
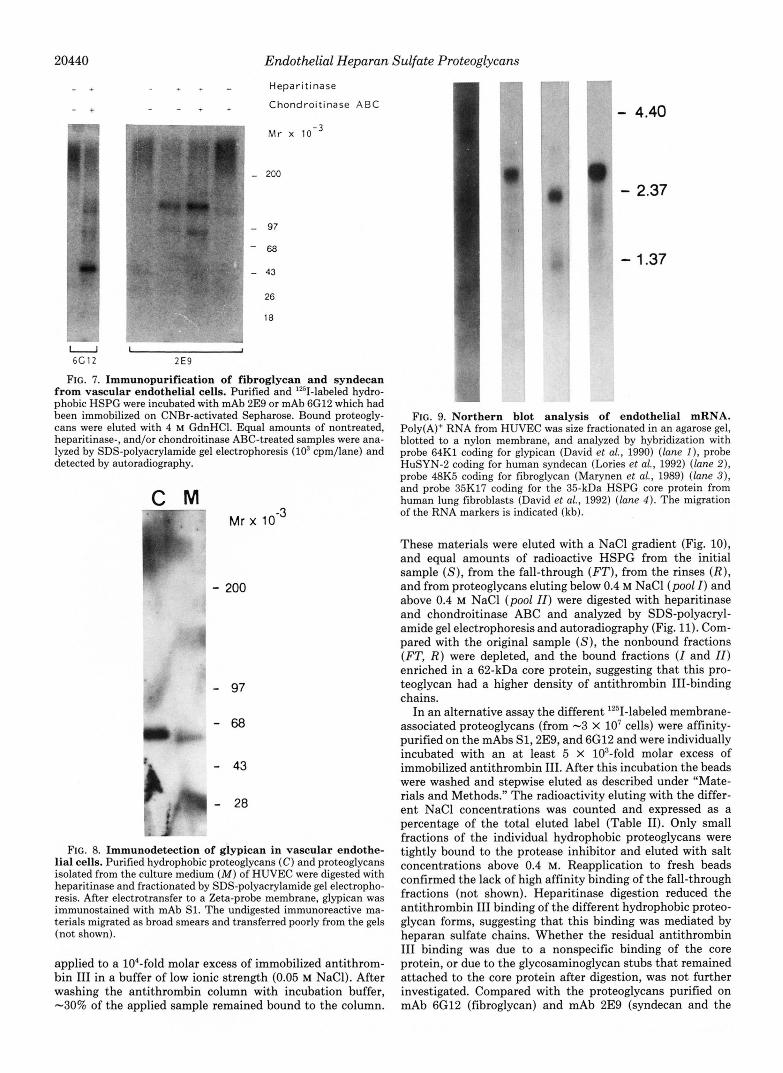
Characterization of the Membrane-associated Heparan Sul- fate Proteoglycans-To identify the core proteins of the hy- drophobic proteoglycans from vascular endothelial cells, "'1- labeled proteoglycan samples were subjected to SDS-poly- acrylamide gel electrophoresis and autoradiography. The in- tact proteoglycans migrated as a broad band in the high molecular weight region of the gel. In both HAEC (Fig. 6A) and HUVEC (Fig. 6 B ) multiple bands could be visualized after digestion with heparitinase. These bands represented

Endothelial Heparan Sulfate Proteoglycans 20439
aFN lcore n n
FIG. 5. Binding of the core protein of the endothelial ma- trix-associated HSPG to plasma fibronectin. Human plasma fibronectin (10 pg/lane) was electroblotted on a nitrocellulose mem- brane after SDS-gel electrophoresis in a 4% Nusieve agarose gel. An inactivated membrane strip was incubated with heparitinase-treated "'I-labeled HSPG (lo6 cpm) from the HUVEC extracellular matrix ( I core). A control strip was stained with an anti-fibronectin mAb (nFN).
core proteins with apparent molecular masses of -130,60,46, 35, and 30 kDa in HAEC and -180, 130,62,43, and 35 kDa in HUVEC. After chondroitinase ABC treatment some addi- tional proteins were identified a 100 kDa core protein in HUVEC and 100- and 80-kDa proteins in HAEC (Fig. 6, A and B ) . The 80-kDa core protein was recognized by mAb 7D8, an antibody which reacts with a CD44-related CSPG (David et al., 1989) (result not shown).
The core proteins of the hydrophobic HSPG from HUVEC were further characterized by affinity chromatography using a panel of monoclonal antibodies raised against the different membrane-associated HSPG of human lung fibroblasts (Lor- ies et al., 1989). mAb 6G12, which reacts with fibroglycan (Marynen et al., 1989), retained the HUVEC-HSPG with the 43-kDa core (Fig. 7). HSPG purified on mAb 2E9, which in fibroblasts reacts with syndecan and one other HS proteogly- can (Lories et al., 1992), yielded a core protein of 130 kDa after heparitinase digestion. An additional band of 100 kDa was obtained when the sample was digested with both hepar- itinase and chondroitinase ABC, suggesting that the latter core protein carried both HS and CS chains and represented syndecan (Fig. 7). Finally, SI, an anti-glypican mAb (David et al., 1990), reacted with the HSPG that yielded the 62-kDa core protein after heparitinase digestion (Fig. 8). Similar results were obtained for HAEC (not shown).
A - + - + Heparitinase . " - Heparitinase
FIG. 6. Heterogeneity of the hy- - - + + Chondroitinase ABC - + * - Chondroilinase ABC drophobic proteoglycans in vascular endothelial cells. Hydrophobic proteo- Mr x alvcans. isolated from HAEC ( A ) and
.. . I ._ -
HUVEC ( R ) were labeled with '251.'Non- 1 treated, heparitinase-, and/or chondroi- I - 200 tinase ABC-treated samples were ana- !
lyzed by electrophoresis in SDS-poly- acrylamide gradient gels (1.5 X lo3 cpm/ lane for HAEC; 5 X lo3 cpm/lane for HUVEC) and detected by autoradiogra- phy.
I
1 - 97 ! - 68
43 I -
w " ~ - 2 8 ,
- 18
In fibroblasts, glypican is anchored to the plasma mem- brane via a GPI, whereas fibroglycan and syndecan are com- ponents with a transmembrane orientation. To investigate the possibility of membrane insertion through a GPI anchor in endothelial cells, the '251-labeled samples from HUVEC were fractionated on immobilized mAb S1 and on mAb 2E9, and aliquots of the immunopurified proteoglycans were treated with phosphatidylinositol-specific phospholipase C. Phospholipase C-treated and nontreated samples were inter- calated into liposomes, whereupon liposome-incorporated and nonincorporated materials were separated by ultracentrifu- gation on a sucrose gradient (Andres et al., 1989). As a result of phospholipase C treatment, the ratio of liposome-incorpo- rated to nonincorporated mAb SI-purified proteoglycans was reduced from 1 to 0.3, while this proportion remained constant for proteoglycans purified on mAb 2E9. This indicated that at least a fraction of the mAb SI-bound membrane HSPG had a GPI-anchor.
Mab S1 reacted also with a HSPG from the culture medium conditioned by HUVEC (Fig. 8). The core protein of this nonhydrophobic medium proteoglycan was slightly smaller than the core protein of the hydrophobic cell surface-associ- ated proteoglycan which reacted with the same antibody (Fig. 8). Similar results had been obtained previously for membrane and medium-associated glypican in human lung fibroblasts (David et al., 1990), and similar experiments with proteogly- cans isolated from HAEC were in accordance with these findings (results not shown).
Finally, the expression of cell surface proteoglycans in human vascular endothelial cells was also studied at the transcriptional level. Northern blots of poly(A)+ RNA from HUVEC were hybridized with 32P-labeled probes coding for fibroglycan, glypican, syndecan, and the 35-kDa HSPG core protein of human lung fibroblasts and submitted to autora- diography. The poly(A)+ RNA contained a glypican RNA of -3.7 kb, whereas the human syndecan probe detected a major -2.5-kb mRNA. The fibroglycan probe and the 35-kDa core protein probe revealed mRNA bands of respectively -2.3 and -2.6 kb (Fig. 9). These results confirmed the expression of syndecan, glypican, and fibroglycan in vascular endothelial cells and suggested that the endothelial and fibroblastic pro- teoglycans which yielded a 35-kDa core were also identical or related to each other.
Antithrombin ZZZ Binding Properties of the Endothelial HSPG-Since we were not able to isolate sufficient amounts of 35S-labeled materials from the different hydrophobic HSPG forms to test for antithrombin I11 binding of the free HS chains, unfractionated preparations of hydrophobic proteo- glycans (from -3 X lo7 HUVEC) were labeled with lZ5I and
B
! 1
1 - 200
I
97
6 8
43
j 26
Endothelial Heparan Sulfate Proteoglycans
- " - Heparitinase " Chondroitinase A B C
5 -
Mr x
- 200
- 97
-68
- 43
26
18
" W
- 4.40
0 - 2.37
- 1.37
U 6C12
~, L I I
2E9
FIG. I. Immunopurification of fibroglycan and syndecan from vascular endothelial cells. Purified and '251-labeled hydro- phobic HSPG were incubated with mAb 2E9 or mAb 6G12 which had been immobilized on CNBr-activated Sepharose. Bound proteogly- cans were eluted with 4 M GdnHC1. Equal amounts of nontreated, heparitinase-, and/or chondroitinase ABC-treated samples were ana- lyzed by SDS-polyacrylamide gel electrophoresis (lo3 cpm/lane) and detected by autoradiography.
C M "
Mr x l o 3
- 200
- 97 -68
- 43
- 28
FIG. 8. Immunodetection of glypican in vascular endothe- lial cells. Purified hydrophobic proteoglycans (C) and proteoglycans isolated from the culture medium (M) of HUVEC were digested with heparitinase and fractionated by SDS-polyacrylamide gel electropho- resis. After electrotransfer to a Zeta-probe membrane, glypican was immunostained with mAb S1. The undigested immunoreactive ma- terials migrated as broad smears and transferred poorly from the gels (not shown).
applied to a 104-fold molar excess of immobilized antithrom- bin I11 in a buffer of low ionic strength (0.05 M NaC1). After washing the antithrombin column with incubation buffer, -30% of the applied sample remained bound to the column.
FIG. 9. Northern blot analysis of endothelial mRNA. Poly(A)+ RNA from HUVEC was size fractionated in an agarose gel, blotted to a nylon membrane, and analyzed by hybridization with probe 64K1 coding for glypican (David et al., 1990) (lane I ) , probe HuSYN-2 coding for human syndecan (Lories et al., 1992) ( l a m 2), probe 48K5 coding for fibroglycan (Marynen et al., 1989) (lane 3), and probe 35K17 coding for the 35-kDa HSPG core protein from human lung fibroblasts (David et al., 1992) (lane 4 ) . The migration of the RNA markers is indicated (kb).
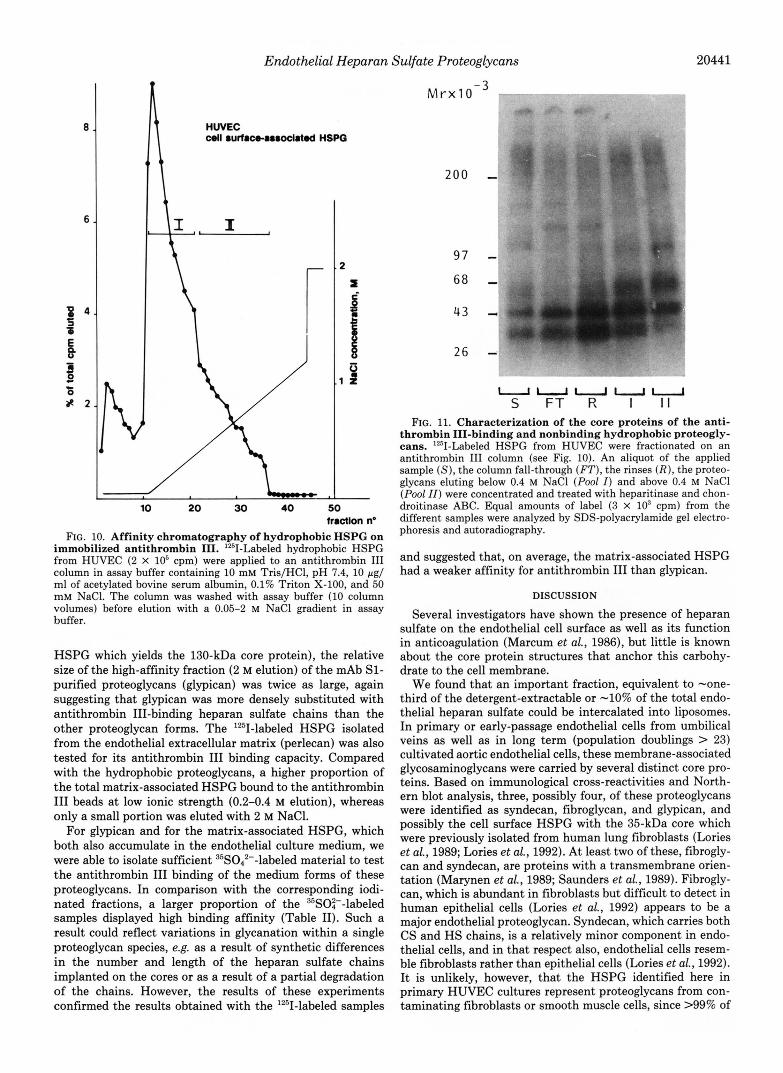
These materials were eluted with a NaCl gradient (Fig. lo), and equal amounts of radioactive HSPG from the initial sample (S), from the fall-through (FT), from the rinses ( R ) , and from proteoglycans eluting below 0.4 M NaCl (pool I ) and above 0.4 M NaCl (pool 11) were digested with heparitinase and chondroitinase ABC and analyzed by SDS-polyacryl- amide gel electrophoresis and autoradiography (Fig. 11). Com- pared with the original sample (S), the nonbound fractions (FT, R ) were depleted, and the bound fractions ( I and 11) enriched in a 62-kDa core protein, suggesting that this pro- teoglycan had a higher density of antithrombin III-binding chains.
In an alternative assay the different '251-labeled membrane- associated proteoglycans (from -3 X lo7 cells) were affinity- purified on the mAbs S1,2E9, and 6G12 and were individually incubated with an at least 5 X 103-fold molar excess of immobilized antithrombin 111. After this incubation the beads were washed and stepwise eluted as described under "Mate- rials and Methods." The radioactivity eluting with the differ- ent NaCl concentrations was counted and expressed as a percentage of the total eluted label (Table 11). Only small fractions of the individual hydrophobic proteoglycans were tightly bound to the protease inhibitor and eluted with salt concentrations above 0.4 M. Reapplication to fresh beads confirmed the lack of high affinity binding of the fall-through fractions (not shown). Heparitinase digestion reduced the antithrombin I11 binding of the different hydrophobic proteo- glycan forms, suggesting that this binding was mediated by heparan sulfate chains. Whether the residual antithrombin I11 binding was due to a nonspecific binding of the core protein, or due to the glycosaminoglycan stubs that remained attached to the core protein after digestion, was not further investigated. Compared with the proteoglycans purified on mAb 6G12 (fibroglycan) and mAb 2E9 (syndecan and the
Endothelial Heparan Sulfate Proteoglycans 20441
8
6
z 4 i6
E B 3 r L
0 % Z
2
9
P 1 1 2
10 20 30 40 50 hactlon no
FIG. 10. Affinity chromatography of hydrophobic HSPG on immobilized antithrombin 111. '*'I-Labeled hydrophobic HSPG from HUVEC (2 X 10' cpm) were applied to an antithrombin I11 column in assay buffer containing 10 mM Tris/HCl, pH 7.4, 10 rg/ ml of acetylated bovine serum albumin, 0.1% Triton X-100, and 50 m M NaCI. The column was washed with assay buffer (10 column volumes) before elution with a 0.05-2 M NaCl gradient in assay buffer.
HSPG which yields the 130-kDa core protein), the relative size of the high-affinity fraction (2 M elution) of the mAb S1- purified proteoglycans (glypican) was twice as large, again suggesting that glypican was more densely substituted with antithrombin 111-binding heparan sulfate chains than the other proteoglycan forms. The lZ5I-labeled HSPG isolated from the endothelial extracellular matrix (perlecan) was also tested for its antithrombin I11 binding capacity. Compared with the hydrophobic proteoglycans, a higher proportion of the total matrix-associated HSPG bound to the antithrombin I11 beads at low ionic strength (0.2-0.4 M elution), whereas only a small portion was eluted with 2 M NaCl.
For glypican and for the matrix-associated HSPG, which both also accumulate in the endothelial culture medium, we were able to isolate sufficient 3sS042--labeled material to test the antithrombin I11 binding of the medium forms of these proteoglycans. In comparison with the corresponding iodi- nated fractions, a larger proportion of the 3sSO:--labeled samples displayed high binding affinity (Table 11). Such a result could reflect variations in glycanation within a single proteoglycan species, e.g. as a result of synthetic differences in the number and length of the heparan sulfate chains implanted on the cores or as a result of a partial degradation of the chains. However, the results of these experiments confirmed the results obtained with the '2sI-labeled samples
200 -
97 - 6 8 -
26 -.
uuuuu S FT R I I t
FIG. 11. Characterization of the core proteins of the anti- thrombin 111-binding and nonbinding hydrophobic proteogly- cans. '2'II-Labeled HSPG from HUVEC were fractionated on an antithrombin I11 column (see Fig. 10). An aliquot of the applied sample (S), the column fall-through (FT), the rinses ( R ) , the proteo- glycans eluting below 0.4 M NaCl (Pool I) and above 0.4 M NaCl (Pool ZZ) were concentrated and treated with heparitinase and chon- droitinase ABC. Equal amounts of label (3 X lo3 cpm) from the different samples were analyzed by SDS-polyacrylamide gel electro- phoresis and autoradiography.
and suggested that, on average, the matrix-associated HSPG had a weaker affinity for antithrombin I11 than glypican.
DISCUSSION
Several investigators have shown the presence of heparan sulfate on the endothelial cell surface as well as its function in anticoagulation (Marcum et al., 1986), but little is known about the core protein structures that anchor this carbohy- drate to the cell membrane.
We found that an important fraction, equivalent to -one- third of the detergent-extractable or -10% of the total endo- thelial heparan sulfate could be intercalated into liposomes. In primary or early-passage endothelial cells from umbilical veins as well as in long term (population doublings > 23) cultivated aortic endothelial cells, these membrane-associated glycosaminoglycans were carried by several distinct core pro- teins. Based on immunological cross-reactivities and North- ern blot analysis, three, possibly four, of these proteoglycans were identified as syndecan, fibroglycan, and glypican, and possibly the cell surface HSPG with the 35-kDa core which were previously isolated from human lung fibroblasts (Lories et al., 1989; Lories et al., 1992). At least two of these, fibrogly- can and syndecan, are proteins with a transmembrane orien- tation (Marynen et dl., 1989; Saunders et al., 1989). Fibrogly- can, which is abundant in fibroblasts but difficult to detect in human epithelial cells (Lories et al., 1992) appears to be a major endothelial proteoglycan. Syndecan, which carries both CS and HS chains, is a relatively minor component in endo- thelial cells, and in that respect also, endothelial cells resem- ble fibroblasts rather than epithelial cells (Lories et al., 1992). It is unlikely, however, that the HSPG identified here in primary HUVEC cultures represent proteoglycans from con- taminating fibroblasts or smooth muscle cells, since >99% of
2 0442 Endothelial Heparan Sulfate Proteoglycans
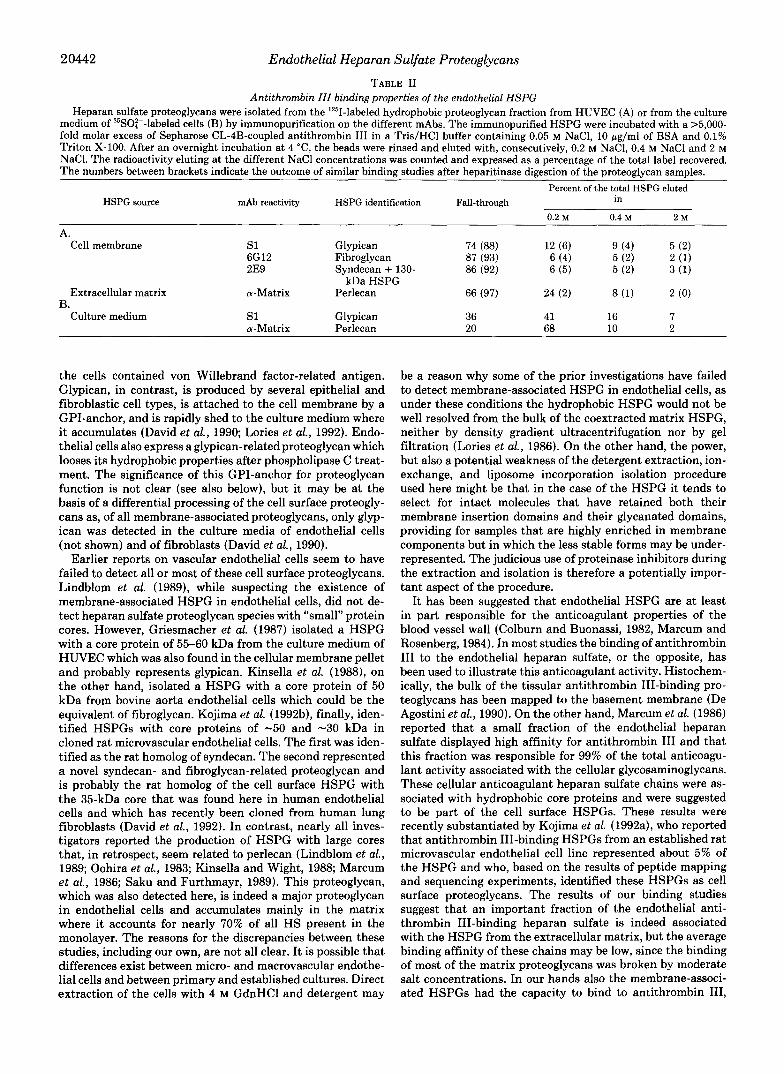
TABLE I1 Antithrombin III binding properties of the endothelial HSPG
Heparan sulfate proteoglycans were isolated from the 'Z51-labeled hydrophobic proteoglycan fraction from HUVEC (A) or from the culture medium of 35SO:--labeled cells (B) by immunopurification on the different mAbs. The immunopurified HSPG were incubated with a >5,000- fold molar excess of Sepharose CL-4B-coupled antithrombin I11 in a Tris/HCl buffer containing 0.05 M NaCl, 10 pg/ml of BSA and 0.1% Triton X-100. After an overnight incubation at 4 "C, the beads were rinsed and eluted with, consecutively, 0.2 M NaCI, 0.4 M NaCl and 2 M NaCI. The radioactivity eluting at the different NaCl concentrations was counted and expressed as a percentage of the total label recovered. The numbers between brackets indicate the outcome of similar binding studies after heparitinase digestion of the proteoglycan samples.
Percent of the total HSPG eluted HSPG source mAb reactivity HSPG identification Fall-through in
0.2 M 0.4 M 2 M
A. Cell membrane s1 Glypican 74 (88) 12 (6) 9 (4) 5 (2)
6G12 Fibroglycan 87 (93) 6 (4) 5 (2) 2 (1) 2E9 Syndecan + 130- 86 (92) 6 ( 5 ) 5 (2) 3 (1)
Extracellular matrix a-Matrix Perlecan 66 (97) 24 (2) 8 (1) 2 (0)
Culture medium s 1 Glypican 36 41 16 7 a-Matrix Perlecan 20 68 10 2
kDa HSPG
B.
the cells contained von Willebrand factor-related antigen. Glypican, in contrast, is produced by several epithelial and fibroblastic cell types, is attached to the cell membrane by a GPI-anchor, and is rapidly shed to the culture medium where it accumulates (David et al., 1990; Lories et al., 1992). Endo- thelial cells also express a glypican-relatedproteoglycan which looses its hydrophobic properties after phospholipase C treat- ment. The significance of this GPI-anchor for proteoglycan function is not clear (see also below), but it may be at the basis of a differential processing of the cell surface proteogly- cans as, of all membrane-associated proteoglycans, only glyp- ican was detected in the culture media of endothelial cells (not shown) and of fibroblasts (David et al., 1990).
Earlier reports on vascular endothelial cells seem to have failed to detect all or most of these cell surface proteoglycans. Lindblom et al. (1989), while suspecting the existence of membrane-associated HSPG in endothelial cells, did not de- tect heparan sulfate proteoglycan species with "small" protein cores. However, Griesmacher et al. (1987) isolated a HSPG with a core protein of 55-60 kDa from the culture medium of HUVEC which was also found in the cellular membrane pellet and probably represents glypican. Kinsella et al. (19881, on the other hand, isolated a HSPG with a core protein of 50 kDa from bovine aorta endothelial cells which could be the equivalent of fibroglycan. Kojima et al. (1992b), finally, iden- tified HSPGs with core proteins of -50 and -30 kDa in cloned rat microvascular endothelial cells. The first was iden- tified as the rat homolog of syndecan. The second represented a novel syndecan- and fibroglycan-related proteoglycan and is probably the rat homolog of the cell surface HSPG with the 35-kDa core that was found here in human endothelial cells and which has recently been cloned from human lung fibroblasts (David et al., 1992). In contrast, nearly all inves- tigators reported the production of HSPG with large cores that, in retrospect, seem related to perlecan (Lindblom et al., 1989; Oohira et al., 1983; Kinsella and Wight, 1988; Marcum et al., 1986; Saku and Furthmayr, 1989). This proteoglycan, which was also detected here, is indeed a major proteoglycan in endothelial cells and accumulates mainly in the matrix where it accounts for nearly 70% of all HS present in the monolayer. The reasons for the discrepancies between these studies, including our own, are not all clear. It is possible that differences exist between micro- and macrovascular endothe- lial cells and between primary and established cultures. Direct extraction of the cells with 4 M GdnHCl and detergent may
be a reason why some of the prior investigations have failed to detect membrane-associated HSPG in endothelial cells, as under these conditions the hydrophobic HSPG would not be well resolved from the bulk of the coextracted matrix HSPG, neither by density gradient ultracentrifugation nor by gel filtration (Lories et al., 1986). On the other hand, the power, but also a potential weakness of the detergent extraction, ion- exchange, and liposome incorporation isolation procedure used here might be that in the case of the HSPG it tends to select for intact molecules that have retained both their membrane insertion domains and their glycanated domains, providing for samples that are highly enriched in membrane components but in which the less stable forms may be under- represented. The judicious use of proteinase inhibitors during the extraction and isolation is therefore a potentially impor- tant aspect of the procedure.
It has been suggested that endothelial HSPG are at least in part responsible for the anticoagulant properties of the blood vessel wall (Colburn and Buonassi, 1982, Marcum and Rosenberg, 1984). In most studies the binding of antithrombin I11 to the endothelial heparan sulfate, or the opposite, has been used to illustrate this anticoagulant activity. Histochem- ically, the bulk of the tissular antithrombin 111-binding pro- teoglycans has been mapped to the basement membrane (De Agostini et al., 1990). On the other hand, Marcum et al. (1986) reported that a small fraction of the endothelial heparan sulfate displayed high affinity for antithrombin I11 and that this fraction was responsible for 99% of the total anticoagu- lant activity associated with the cellular glycosaminoglycans. These cellular anticoagulant heparan sulfate chains were as- sociated with hydrophobic core proteins and were suggested to be part of the cell surface HSPGs. These results were recently substantiated by Kojima et al. (1992a), who reported that antithrombin 111-binding HSPGs from an established rat microvascular endothelial cell line represented about 5% of the HSPG and who, based on the results of peptide mapping and sequencing experiments, identified these HSPGs as cell surface proteoglycans. The results of our binding studies suggest that an important fraction of the endothelial anti- thrombin 111-binding heparan sulfate is indeed associated with the HSPG from the extracellular matrix, but the average binding affinity of these chains may be low, since the binding of most of the matrix proteoglycans was broken by moderate salt concentrations. In our hands also the membrane-associ- ated HSPGs had the capacity to bind to antithrombin 111,

Endothelial Heparan Sulfate Proteoglycans 20443
and all the identified forms bound antithrombin to a certain extent. As a whole our results are consistent with the data of Kojima et al. (1992a) in microvascular endothelial cells, who established that active and inactive HSPGs (in terms of antithrombin I11 binding) yielded similar peptide patterns. However, the HSPG which in HUVEC more typically dis- played high affinity binding for antithrombin I11 was glypican, a proteoglycan which was not recovered or formally identified among the antithrombin-binding proteoglycans from micro- vascular endothelial cells. This suggests that glypican, some- what more often than other proteoglycans, contains the spe- cific pentasaccharide sequence required for high affinity bind- ing to antithrombin 111. If these antithrombin I11 binding sequences are randomly distributed among the heparan sul- fate chains of the different endothelial proteoglycans, these results would suggest that the glypican core protein on average carries somewhat more and/or longer heparan sulfate chains than the other core proteins. On the other hand, the indication for more antithrombin I11 binding sequences in glypican com- pared with other proteoglycans raises the possibility that the core protein structure may have some influence with regard to HS modifications. The existence of a GPI anchor which may direct the proteoglycan to different membrane compart- ments, the more complex tertiary structure as a result of multiple intrachain disulfide bridges, and the location of glycan attachment sites close to the carboxyl terminus of the glypican core protein are features that single out glypican from fibroglycan and syndecan (which are transmembrane proteins that lack cysteines and have their chain attachment sites clustered at the amino terminus) and may influence the efficiency of the heparosan modification enzymes. Final ex- planations of these results will have to await antithrombin I11 binding studies with the corresponding free heparan sulfate chains and the analysis of the prevalence of these specific pentasaccharide sequences in the different fractions.
Ultimately, the finding that in polarized cell types all gly- cosyl phosphatidylinositol-anchored proteins appear to be restricted to the apical surface (Rodriguez-Boulan and Nel- son, 1989) may prove more relevant and significant for indi- vidual proteoglycan functions than small to moderate differ- ences in antithrombin-binding activities. If the apical target- ting rule is also true for glypican, this could mean that glypican becomes concentrated at the luminal side of the endothelium where it could act as a "first line" physiological anticoagulant, even if chains with anticoagulant activity were not restricted to glypican. Then, the shedding of glypican, if it also occurs in vivo, may result in the release of a soluble anticoagulant from the endothelial cell surface into the blood stream. As proposed by de Agostini et al. (1990) the small amounts of luminal anticoagulant HSPG may be critically placed to bind plasma antithrombin, accelerate the action of the protease inhibitor, and thereby regulate the ability of hemostatic mechanisms at the blood vessel wall interface. The possibly larger quantities of abluminal anticoagulant HSPG could serve as a reservoir or backup system that may be brought into play when extensive damage occurs to the overlying endothelium. This may extend to other proteoglycan functions as well. Lipoprotein lipase is bound to the vascular endothelial cell surface through heparin-like glycosaminogly-
cans (Shimada et al., 1981) and depends on a saturable trans- port system which requires HSPG for passage across the endothelial cells (Saxena et al. 1991). Since lipoprotein lipase bound to heart cell cultures can be released by the action of phosphatidylinositol-specific phospholipase C (Chajek-Shaul, 1989), an apically exposed glypican would also be a candidate for binding lipoprotein lipase to the endothelial luminal cell surface.
Acknowledgments-We thank Helga Ceulemans, Christien Coo- mans, An Ray6, and Trees Vancoetsem for their expert technical assistance.
REFERENCES Andres, J. L., Stanley, K., Cheifetz, S., and Massagu6, J. (1989) J. Cell Biol.
109,3137-3145 Bengtsson, O., Olivecrona, T., Hook, M., Riesenfeld, J., and Lindahl, U. (1980)
Biochem. J. 189,625-633 Chajek-Shaul, T., Halimi, O., Ben-Naim, M., Stein, 0.. and Stein, Y. (1989)
Biochim. Biophys. Acta 1014,178-183 Colburn, P., and Buonassisi, V. (1982) Biochem. Biophys. Res. Commun. 104 ,
220-227 David, G., Lories, V., Decock, B., Marynen, P., Cassiman, J. J., and Van den
Berghe, H. (1990) J. Cell Biol. 111 , 3165-3176 David, G., Van der Schueren, B., Marynen, P., Cassiman, J.-J., and Van den
Berghe, H. (1992) J. Cell Biol. 118,961-969 De Agostini, A. I., Watkins, S. C., Slayter, H. S., Youssoufian, H., and Rosen-
berg, R. D. (1990) J. Cell Biol. 3 , 1293-1304 Griesmacher, A,, Hennes, R., Keller, R., and Greiling, H. (1987) Eur. J .
Biochem. 168.95-101 Heremans, A,, De Cock, B., Cassiman, J. J., Van den Berghe, H., and David,
G. (1990) J. Biol. Chem. 265,8716-8724 Heremans, A,, Cassiman, J. J., Van den Berghe, H., and David, G. (1988) J.
Biol. Chem. 263,4731-4739 Heremans, A., Van der Schueren, B., De Cock, B., Paulsson, M., Cassiman, J.
J., Van den Berghe, H., and David, G. (1989) J. Cell Biol. 109,3199-3211 Hoyer, L. W., de 10s Santos, R. P., and Hoyer, J. R. (1973) J. Clin. Inuest. 5 2 ,
2737-2744 Jackson, R. L., Busch, S. J., and Cardin, A. D. (1991) Physiol. Reu. 7 1 , 481-
Jaffe. E. A,. Nachman. R. L.. Becker. C. G.. and Minick. C. R. (1973) J. Clin.
-~ ~~~
539
Invest. 5 2 , 2745-2756 Kinsella, M. G., and Wight, T. N. (1988) Biochemistry 2 7 , 2136-2144 Kjellh, L., and Lindahl, U. (1991) Annu. Reu. Biochem. 60,443-475 Kojiama, T., Leone, C. W., Marchildon, G. A., Marcum, J. A,, and Rosenberg,
KoJlama, T., Shworak, N. W., and Rosenberg, R. D. (1992b) J . Biol. Chem. R : D. (1992a) J . Biol. Chem. 267,4859-4869
9 R 7 AQ7LAQ77 Lindblom, A. I., Carlstedt, I., and Fransson, L.-A (1989) Biochem. J. 261 , 145- "., "." " , ,
167 Lories, V., David, G., Cassiman, J. J., and Van den Berghe, H. (1986) Eur. J.
Lories, V., De B, Biochem. 1 5 8
Lones, V., Cassl (1987) J. Biol,.
*""
oeck, H., David, G., Cassiman, J. J., and Van den Berghe, H. ,351-360
Chem. 262,854-859 man, J. J., Van den Berghe, H., and David, G. (1989) J . Biol.
Lories, V., Cassiman, J. J., Van den Berghe, H., and David, G. (1992) J . Biol.
Marcum, J. A,, and Rosenberg, R. D. (1984) Biochemistry 23,1730-1737 Marcum, J. A,, Atha, D. H., Fritze, L. M. S., Nawroth, P., Stern, D., and
Marynen, P., Zhang, J., Cassiman, J. J., Van den Berghe, H., and David, G.
Miller-Andersson, M., Borg, H., and Anderson, L.-0. (1974) Thromb. Res. 5 ,
Chem. 2 6 4 , 7009-7016
Chem. 2 6 7 , 1116-1122
Rosenberg, R. D. (1986) J. Biol. Chem. 2 6 1 , 7507-7517
(1989) J. Biol. Chem. 2 6 4 , 7017-7024
439-452 Oobira, A., Wight, T. N., and Bornstein, P., (1983) J. Biol. Chem. 2 5 8 , 2014-
Pejler, G., and David, G. (1987) Biochem. J. 248,69-77 Rapraeger, A. C., Krufka, A., and Olwin, B. B. (1991) Science 2 5 2 , 1705-1708 Rodriguez-Boulan, E., and Nelson, W. J. (1989) Science 2 4 5 , 718-725 Saksela, O., Moscatelli, D., Sommer, A., and Rifkin, D. B. (1988) J. Cell Biol.
Saku, T., and Furthmayr, H. (1989) J . Biol. Chem. 264,3514-3523 Saunders, S., Jalkanen, M., O'Farrell, S., and Bernfield, M. (1989) J. Cell Biol.
Saxena, U., Klein, M. G., and Goldberg, I. J. (1990) J. Biol. Chem. 2 6 5 , 12880-
Saxena, U., Klein, M. G., and Goldberg, I . J. (1991) Proc. Natl. Acad. Sci.
Shimada, K., Gill, P. J., Silbert, J. E., Douglas, W. H. J., and Fanburg, B. L.
Thunberg, L., Backstrom, G., and Lindahl, U. (1982) Carbohydr. Res. 100 ,
~~~ ~~-
2021
107 , 743-751
108 , 1547-1556
12886
U. S. A. 88,2254-2258
(1981) J. Clin. Inuest. 6 8 , 995-1002
2 ~ 7 ~ 4 1 n Yayon, A,, Klagsbrun, M., Esko, J. D., Leder, P., and Ornitz, D. M. (1991) Cell
VU" TI"
64,841-848