catalase, bskat-1 an antioxidant enzyme as a model for...
TRANSCRIPT

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 129
5.1. Introduction
5.1.1. Radiation induced enzyme alternation in astrobiological important
microorganisms
Evidence for the possible interplanetary transfer of biological materials began
with experiments testing the resistance of microbes to the space environmental
conditions (Horneck, 1993; Wickramasinghe, 2004). In space, microbes would be
subjected to different stresses, including extreme vacuum, desiccation, solar and cosmic
radiation, microgravity, and both extreme hot and cold temperatures (Nicholson et al.,
2000). Of these factors, solar UV is the most harmful (Rampelotto et al., 2009; Horneck
et al., 1994; Nicholson et al., 2000; Roya et al., 2002). However, spaceflight
experiments demonstrate that with minimal UV shielding, several types of microbes can
survive for years at exposures to the harsh environment of space (Roya et al., 2002;
Rettberg et al., 2002). Furthermore, it was estimated that, if shielded by two meters of
meteorite, a substantial number of spores would survive after 25 million years in space
(Horneck et al., 2002). Because of their high resistance to different extreme conditions,
spores of Bacillus subtilis are the most widely used model microorganism for these
studies (Nicholson et al., 2000; Horneck, 1993; Horneck et al., 1994). However, various
other microbes have been used, including vegetative cells of the soil bacteria
Deinococcus spp. and Rhodococcus erythropolis, some halophilic Archaea Halorubrum
spp, Halobacterium spp, Chroococcidiopsis and others (Pabulo, 2010).
It was argued that, these upper atmospheric microorganisms were of cometary
origin and thereby cometary panspermia stood vindicated. Although, it was claimed that
bacteria and fungi can be found over the altitude range 18-39 km, such results were

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 130
generally dismissed on the basis of contamination and were particularly interest in
astrobiology (Shivaji et al., 2006, Shivaji et al., 2009). In atmosphere, evidence for the
occurrence of microorganisms at altitudes of 17-85 samples have been collected in the
higher atmosphere. These samples were obtained using a meteorological rockets, a
specially designed direct-flow sampler and cryosamplers sent up on a balloon
(Wainwright et al., 2003; Shivaji et al., 2006, Shivaji et al., 2009). The dynamic
chemical and biological interactions were very complex, and these organisms that
survive in this environment must tolerate higher level of UV and ionizing irradiation. At
higher altitude, the ionizing radiation (X-ray, γ-ray), UV light and redox cycling causes
the generation of reactive oxygen species during aerobic respiration of microorganism.
These include, superoxide (O2-1
), peroxide (O2-2
), hydrogen peroxide (H2O2), hydroxyl
(OH)- and hydroperoxyl (HOO)
- radicals were generated (Halliwell and Gutteridge,
1999, Hassan and Fridovich, 1979; Vile and Tyrrell, 1993; Jurkiewicz and Buettner,
1994). These oxidants cause chemical modification of the cellular components, nucleic
acids, proteins, and lipids, resulting in a number of metabolic malfunctioning, aging
process, mutagenesis, and cell death (Halliwell and Gutteridge, 1999). Consequently, it
has been proposed that, these microorganisms operate several defense mechanisms
against reactive oxygen intermediates by the action of scavenging enzymes such as
catalase, peroxidase, superoxide dismutase (SOD) and by a number of antioxidants
occurring in cells (Cha and Kim, 1996 & 1999; Halliwell and Gutteridge, 1999).
5.1.2. Catalase
Bacteria have specific enzymes to detoxify potentially lethal reactive oxygen
species (ROS), including superoxide anion radical (O2), hydrogen peroxide (H2O2) and

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 131
hydroxyl radical (HO-). The effects of ROS result in the oxidation of various cellular
components such as DNA, RNA, proteins and lipids. Thus, bacterial defense
mechanisms against oxidative stress play a very important role in their survival.
Catalase [hydrogen peroxide: hydrogen-peroxide oxidoreductase (EC1.11.1.6)] was one
of the central defense enzymes against oxidative stress, which catalyzes the conversion
of H2O2 to water and molecular oxygen (Reaction 1).
2H2O2 2H2O + O2 (Reaction 1)
Many bacteria possess two types of catalases, typical catalase which catalyzes
H2O2 only and catalase-peroxidase which also has an associated peroxidase activity.
They are distinct from each other in many properties. Typical catalase generally
contains heamin prosthetic groups and shows a broad optimum pH in the range of 5-10.
It was resistant to treatment with ethanol/chloroform (Margoliash et al., 1960).
Catalase-peroxidase, which has an optimum pH at 6-6.5, was relatively more sensitive
to temperature and ethanol/ chloroform than the typical catalases (Goldberg and
Hochman, 1989; Hochman et al., 1992).
5.1.3. Classification and catalytic properties of catalase
Three classes of proteins unrelated based on sequence and structure exhibit
significant catalase activity. The classes that were classified as monofunctional, bi
functional and non-heme or Mn-containing catalases. A typical monofunctional catalase
was subdivided based on large or small subunits. Phylogenetic analyses have
demonstrated the existence of two distinct clades or sub groupings of small subunit
enzymes and one clade of large subunit enzymes among the monofunctional catalases
(Klotz et al., 1997). The second, less widespread class was composed of bifunctional,

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 132
heme containing catalase-peroxidases that were closely related by sequence and
structure to plant peroxidases. The third class of catalase includes the nonheme or Mn-
containing catalases.
5.1.4. Radiation induced catalase regulation in bacteria
The ROS hypothesis for UV damage has been studied using catalases, which
destroy peroxide and other ROS (Eisenstark, 1989). Because wild type E. coli strains
were able to resist complete inactivation by two known catalases in E. coli are
suspected to provide protection (Eisenstark, 1989). The 84 kDa catalase,
hydroperoxidase-I (HPI), was membrane bound, encoded by katG and has catalase and
peroxidase activity (Triggs-Raine et al., 1990). HPI was expressed only during
anaerobic growth was predicted to repair after UVA damage by increasing cell
permeability to antioxidants (Leven et al., 1990). The 93 kDa catalase,
hydroperoxidase-II (HPII), was cytoplasmic, encoded by katE, and has only catalase
activity (Van Ossowski et al., 1991). HPI was expressed during aerobic exponential
growth (Mulvey et al., 1990) but its role in UVA damage remains unclear due to
contradicting evidence. The katG, katE double knockout E. coli strain, UM2, has
previously shown increased sensitivity to UVA consistent with a role in UVA resistance
(Amiri et al., 2007). However, the katG knockout strain, UM197, did not suffer after
UVA exposure compared to its isogenic parent, suggesting that only katE was required
for protection (Amiri et al., 2007). However, the UM2 strain complemented with katE
also failed to protect against UVA (Cheng et al., 2009) speculatively because of over-
expression of HPII by the pBAD24 construct (Banerjee et al., 2009). Lack of protection
was thought to be because of the photosensitizing properties of HPII at high

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 133
concentrations (Cheng et al., 2009). Another study successfully created an alternative
catalase double knockout of the UM197 strain, CKLW09W2, using antisense katE
DNA (akatE) in the pBAD24 vector (Chan et al., 2010); however, the authors did not
test the strain’s ability to resist UVA damage. To test for HPII-mediated UVA
protection at different HPII concentrations, this study controlled expression of the akatE
message in CKLW09W2 using the inducible araC promoter on the pBAD24 construct.
The aim of this study was to control expression levels of HPII in HPI knockout cells to
elucidate whether HPII alone has protective properties against UVA damage at lower
concentrations and determine the expression level above which photosensitizing
properties of HPII take over.
5.1.5. Stability of catalase
The unusual resistance to denaturation and proteolysis supports the catalases
with large subunits. Resistance to pH and thermal denaturation was noted early in the
study of catalases from E. coli and other enteric bacteria used as a diagnostic test to
differentiate among catalases in crude extracts (Meir and Yagil, 1984; Goldberg and
Hochman, 1989). Recent research on thermal stability of catalase revealed that, there
was actually a small increase in activity above 60°C and that activity began to drop only
above 80°C, with a Tm of 83°C (Switala et al., 1989). Boiling in buffer, or alternatively,
heating to 65°C in 5.6 M urea was required to dissociate the dimer. While not as stable
as HPII, small subunit catalases such as bovine liver catalase (BLC) still exhibited
enhanced stability for an enzyme from an organism normally growing at 37°C, with a
Tm for loss of activity of 56°C. The thermal stability of HPII has also been utilized as a
purification step (Switala and Loewen, 2002). Catalases, generally, were also resistant

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 134
to treatment with an ethanol: chloroform mixture, another property that could be
exploited as a tool for catalase purification (Switala and Loewen, 2002). Both thermal
stability and resistance of the majority of the protein to proteolysis can be explained in
terms of a very rigid, stable structure that resists unfolding, thereby preventing access to
the protease active site. Resistance to proteolysis was an advantage. Because, it was
expressed in stationary phase, a period of rapid protein turnover and elevated protease
levels. There would have been a strong selective pressure to retain this property, but it
was not clear whether resistance to proteolysis evolved in E. coli and thermal stability
was an inadvertent outcome. Alternatively, the catalase could have originated in a
thermophilic organism and been horizontally transferred to E. coli, where protease
resistance has been the selective pressure for retention of thermal resistance.
5.2. Materials and Methods
5.2.1. Chemicals
All the chemicals used for the experiments were of analytical grade and
chemicals were of the highest possible purity. For the preparation of culture media, the
chemicals used such as, Bacto peptone, Beef extract, Tryptone, Starch, yeast extract and
agar were purchased from Himedia laboratory (Mumbai, India). Other Inorganic salts
such as, NaCl, MgSO4, ferrous sulphate, magnesium chlorides, KH2PO4, NaHCO3,
Na2CO3 were purchased from Merck chemicals (Mumbai, India). Certain trace element
such as, Na2EDTA; ZnSO4 MnCl2, H3BO3, CaCl2, CuCl2, NiCl2, Na2MoO4, Na2SO3, and
FeSO4 purchased from Sigma-Aldrich (Mumbai, India) and Himedia laboratory
(Mumbai, India). (NH4)2SO4 was purchased from Himedia laboratory (Mumbai, India).
Phenyl Sepharose High Performance column was purchased from Pharmacia (Uppsala,

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 135
Sweden) Gel filtration chromatographic resin such as Superdex 200 HR was purchased
from Pharmacia (Uppsala, Sweden)
5.2.2. Bacterial strains
Bacillus stratosphericus was adopted or purchased from Microbial Type Culture
Collection (MTCC) Chandigarh, India and Center for Cellular and Molecular Biology
(CCMB) Hyderabad, India. This strain was deposited previously in MTCC under
accession number 7305 and is publically available. The strain was previously isolated
from the stratosphere (approximately 41 km high altitude) using cryogenic tubes for air
sampling experiments from high altitudes. The physicochemical properties of the strain
are found to be grow in between 8 and 37ºC, but not at 40ºC. Growth occurs between
pH 6-10, but not at pH 5 or pH 11. The strain has extreme resistant to UV radiation and
other ionizing radiations (Shivaji et al., 2006).
5.2.3. Reference strains
Dienococcus radiodurans R1 and E.Coli cells were used as reference strains.
Because Dienococcus radiodurans R1 is currently known as extreme radiation resistant
microorganism on earth
5.3.3. Culture conditions for Bacillus stratosphericus
The lyophilized Bacillus stratosphericus cells were initially cultivated
aerobically on nutrient agar medium of pH 7.0 at 40ºC for two days. Further,
B.stratosphericus was sub cultured in a medium containing 8 g/l–1
yeast extract, 8 gl-
1extract from meat and 1 g/l
–1 KH2PO4 at 40ºC and pH 7.0, buffered with 50 mM
NaHCO3/Na2CO3. Then, 1% (w/w) of a trace element solution (2.5 gl–1
Na2EDTA; 0.1
g/l–1
ZnSO4·7H2O; 0.030 g/l–1
MnCl2·4H2O; 0.3g/l–1
H3BO3; 0.2 g/l–1
CaCl2·6H2O;

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 136
0.01g/l–1
CuCl2·2H2O; 0.02g/l–1
NiCl2·6H2O; 0.9g/l–1
Na2MoO4·2H2O; 0.03 g/l–
1Na2SO3·5H2O, and 1g/l
–1FeSO4·7H2O) was added to the incubation mixture. For
reference strains Dienococcus radiodurans R1 was cultured in a TYGM medium (1%
tryptone 0.5% yeast extract, 0.2% glucose, and 0.2% L-methionine) at 30ºC with
continuous aeration at 150 rpm (Soung and Lee, 2000; Yun and Lee, 2001). E.Coli cells
were grown in nutrient broth medium of pH 7.0 at 37ºC
5.2.4. Radiation experiments
B. stratosphericus cells were cultured in the media as described earlier and
treated with different doses of UVA and γ- radiations (1 kGy to 10 kGy).
Ultraviolet-A exposure
Late log phase cells of B. stratosphericus were serially diluted with normal
saline and plated. Cells were exposed to different doses of UVA radiation using Xenon
lamp (15 W; Philips UV lamp, India) and 300 nm cut off band pass filter. The plates
were wrapped with aluminum foil and incubated at 32ºC for 48h and the number of
colonies appeared on plates were recorded. For control, D. radiodurans and E.coli cells
were cultured and exposed different dose of UVA light and the percent survivility was
calculated.
γ-radiation exposure
The cells of B. stratosphericus and reference strains were suspended in sterile
phosphate buffered saline (PBS) and exposed to different doses (1 kGy to 10 kGy) of γ-
radiation (radiation dose 4.87 kGy/h) using Cobalt 60
irradiator. Appropriate dilution of
treated cells was placed on media and number of colony forming units was counted
after 48h of incubation at 37ºC and the percent survivility was calculated.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 137
5.2.5. Catalase induction by UV and γ -irradiation on bacterial cells
For induction experiment, the bacterial cells was cultured in TYGM (1%
tryptone, 0.5% yeast extract, 0.2% glucose, and 0.2% methionine) with continuous
shaking condition (150 rpm) at 40ºC of pH 7.0 for 48 hours. Catalase was induced by
pre-incubating the bacterial suspension with continuous UV (UVA) irradiation (200-
400 nm) (15 W; Philips UV lamp, India) for an hour. The UV dose was measured using
VLX-3W radiometer (Radiometer, France). For γ-irradiation, cell suspensions were
exposed to different doses (1 kGy to 10 kGy) of γ-radiation (radiation dose 4.87 kGy/h)
using Co60
irradiator of strain were irradiated at room temperature for 1 h. Then, the cell
suspension was cultured on TYGM liquid media for 48 hours. The growth of bacterial
strain in both cases was monitored at 600 nm (Schmazdu UV-Visible
spectrophotometer, Japan). The cells were harvested and suspended in 50 mM
potassium phosphate buffer, pH 7.0 were used for preparation of cell free extracts.
5.2.6. Preparation of cell extraction
UV and γ-irradiated culture broth of B stratosphericus was centrifuged at 10,000
X g for 20 min at 4ºC to separate the cells from the broth. The pellet containing cells
were suspended in minimum amount of 50 mM potassium phosphate buffer (pH 7.0)
and kept at 4ºC for an hour. After incubation with appropriate buffer, the protein was
extracted by sonication for 10 min in ice-bath and can be used as a crude extract. The
protein was concentrated to several folds by Ultrafiltration (PM-10 membrane, Amicon,
Beverly, MA, USA).

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 138
5.2.7. Activity profile of catalase in crude extract
The ultra filtration extract were resolved on 8.5% gel by PAGE in Tris-glycine
buffer (Hedrick and Smith, 1968; Wayne and Diaz, 1986). Electrophoresis was carried
out at 75 volts through the stacking gel, and at 100-150 volts through the resolving gel,
using vertical mini slab gel unit (BIORAD, USA) filled with running buffer.
Visualization of catalase bands resolved by the treatment of gels with horseradish
peroxidase prior to incubating in 5 mM H2O2 solution followed by diaminobenzidine. In
the latter staining method, gels were incubated initially with 5 mM hydrogen peroxide
and followed by a freshly prepared mixture of 2% ferric chloride and 2% potassium
ferric cyanide.
5.2.8. Catalase activity assay
Catalase activity in sonic extract of cells was measured by
spectrophotometrically by monitoring the decrease in absorbance at 240 nm due to
decomposition of hydrogen peroxide in 50 mM potassium phosphate buffer, pH 7.0 at
25ºC. One unit of Catalase was defined as the disappearance of one µmol of hydrogen
peroxide per minute (ε = 43.6 M-1
cm-1
) (Nak and Young, 2000).
5.2.9. Estimation of protein concentration
Protein concentration was measured by the standard procedure of Lowry et al.,
(1953). A standard plot was developed using Bovine Serum Albumin (BSA) of different
concentration and the protein concentration of test samples was determined for crude
preparation and each steps of the purification.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 139
5.2.10. Purification of Catalase-BsKat-1
The cell lysate was subjected to fractionation by ammonium sulfate precipitation
at 40% saturation to remove impurities, followed by 70% saturation in a second step to
precipitate the Bskat. The precipitated proteins were collected by centrifugation, and the
pellet was dissolved in phosphate buffer (50 mM, pH 7.0) with 1.0 M (NH4)2SO4. The
solution was applied to a Phenyl Sepharose High Performance column (Pharmacia,
Uppsala, Sweden) equilibrated with 1.0 M (NH4)2SO4 in phosphate buffer. Fractions
containing protein were eluted with a linear decreasing gradient of 1.0-0 M (NH4)2SO4.
Catalase active fractions were pooled and concentrated in an ultrafiltration cell
(Sartorius, Epsom, UK) using a membrane with a 100 kDa cutoff. The concentrated
enzyme solution was subjected to a Superdex 200 HR 10/30 column (Pharmacia,
Uppsala, Sweden) previously equilibrated with phosphate buffer (50 mM, pH 7.0)
containing 0.15 M NaCl. The column fractions were eluted with the same buffer. The
active fractions were collected and stored -20ºC until further use.
5.2.11. Molecular mass determination
The molecular weight of the enzyme analyzed using 12.5% non-denaturating
polyacrylamide gels (BIORAD, USA) essentially as described by Laemmli, (1970).
After electrophoresis, gels were stained for 1 h or until clear visualization of protein
bands with the Colloidal protein strainer (Aristogene Biosciences, Bangalore, India).
The apparent molecular weight of the purified enzyme was determined by comparing
with Rf values of standard proteins (Bio-Rad Laboratories, Hercules, CA). The native
molecular mass of catalase was estimated by gel filtration using Sephacryl 200 HR
using Myosin (205 kDa), Phosphorylase B (97 kDa), Ovalbumin (45 kDa), Trypsinogen

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 140
(24 kDa) and Aprotinin (6.5 kDa) as a molecular weight standards. pI of the native
enzyme was determined by the method described by Thompson et al., (2003), except
for perioxidase staining. The native pI was determined from the standards range from
4.45 to 9.6 (Biorad, Hercules, CA).
5.2.12. Effect of temperature, pH and stability
To determine the influence of temperature on enzymatic activity, enzyme
samples were incubated at temperatures range from 15 to 60⁰C. Thermostability was
investigated by pre incubating the samples at temperatures range of 45-75⁰C. After
various time intervals (0.5 to 3 hours), samples were withdrawn and residual activity
was determined at the standard assay conditions. The pH optimum of catalase activity
was determined in various buffers from pH 4.0 to pH 12.0. The buffers used were
Formic acid buffer (pH 3.3 to 4.3); Trimethylamine/acetic acid buffer (pH 4.3 to 5.3);
Trimethylamine/carbonate buffer (pH 5.9 to 6.9) and Phosphate buffer (pH 7 to 12) at
optimum temperature.
5.2.13. Effects of inhibitors, metal ions and spectral studies
Effects of inhibitors on catalase activity were tested using 3-amino-1, 2, 4-
triazol, sodium azide, hydroxylamine hydrochloride and sodium cyanide at a
concentration of 0.1 to 1.0 mM. The enzyme (1 mg/ml) was assayed by pre incubated
with the inhibitor for five min prior to assay at room temperature. The absorption
spectra of native and reduced enzyme were measured on Schmidzu UV-Visible
Spectrophotometer (Schmidzu, Japan). For this, the native enzyme was reduced with 1
mM sodium dithionite, and enzyme treated with 10 mM KCN were pre incubated for 5
min at 45⁰C in phosphate buffer (50 mM, pH 10.0). The reaction was monitored

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 141
spectrophotometrically as described previously (Hochman and Goldberg, 1991). The
protoheme type and content of the enzyme was determined through the formation of a
pyridine hemochrome in absorption spectra. The molar absorption coefficient for the
pyridine hemochrome was assumed as 191.5 mM-1
cm-1
(Falk, 1964). Effect of some
metal ions on enzyme activity was determined by pre incubating at a final concentration
of 1 mM Zn2+
,Fe2+
,Ca2+
,Mg2+
,Cu2+
(Chloride form) 10 mM EDTA and histidine at 45
°C at pH 10. The enzyme activity was determined after incubation with the substrate for
30 min.
5.3. Results
5.3.1. B. stratosphericus showed differential response to UV-A and γ radiation
compared to D radiodurans and E.coli
The late log phase cells of B. stratosphericus were exposed to different doses of
γ and UVA radiation and cell survival was monitored on TYGM agar (described
earlier). Survival of all three strains (including reference strains) were determined
following γ-irradiation at doses up to 12 kGy. The results of γ-radiation exposure
showed the differential surveillance of B. stratosphericus cells/spores. When compared
to the reference strains, B. stratosphericus had shown nearly two log cycles loss in γ-
radiation beyond 4 kGy (34%). In contrast to D.radiodurans, which is currently known
as highest radiation resistance organism on earth, shown resistant beyond 8 kGy
exposure of γ-radiation (68%). However, the other mesophilic control strain E.coli cells
did not appeared much resistant for γ-radiation above 2 kGy (6%) (Fig.5.1)
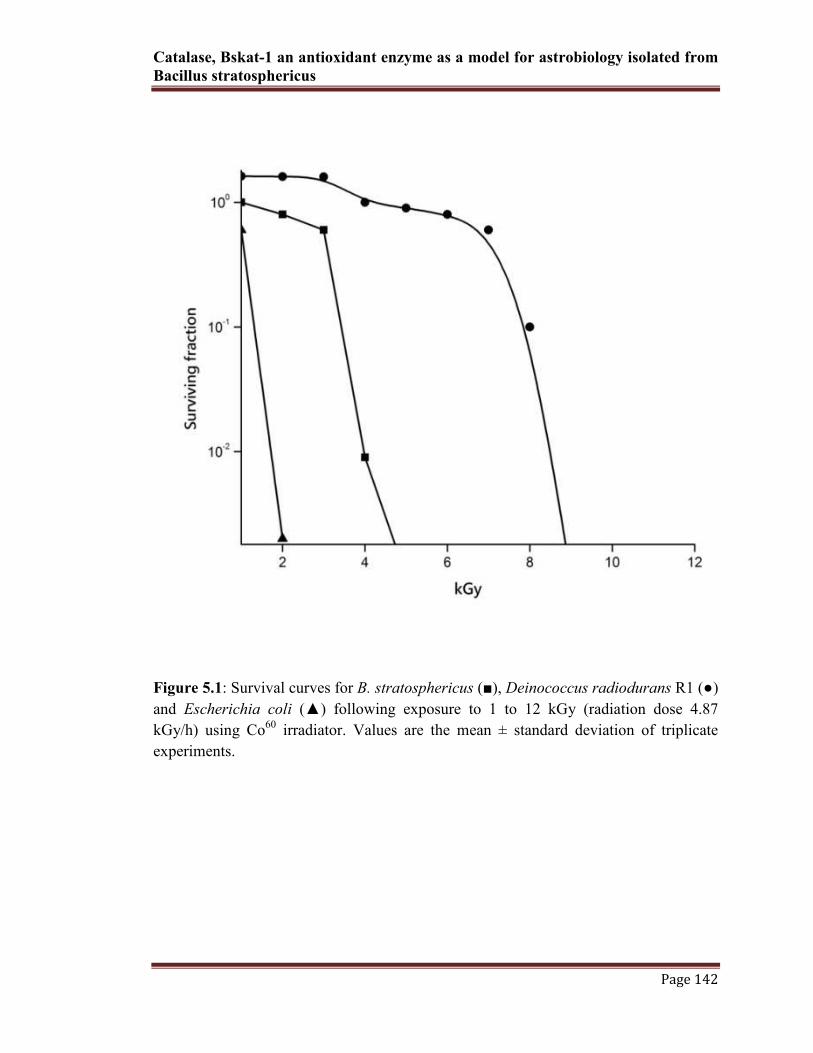
Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 142
Figure 5.1: Survival curves for B. stratosphericus (■), Deinococcus radiodurans R1 (●)
and Escherichia coli (▲) following exposure to 1 to 12 kGy (radiation dose 4.87
kGy/h) using Co60
irradiator. Values are the mean ± standard deviation of triplicate
experiments.
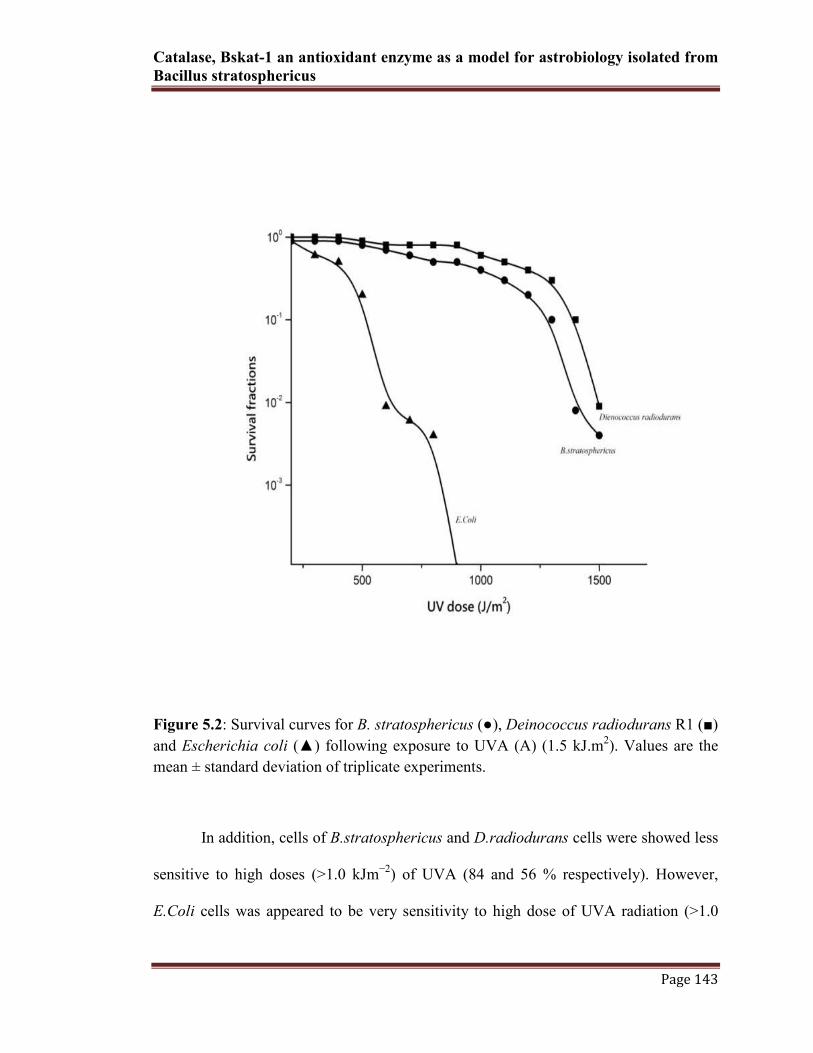
Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 143
Figure 5.2: Survival curves for B. stratosphericus (●), Deinococcus radiodurans R1 (■)
and Escherichia coli (▲) following exposure to UVA (A) (1.5 kJ.m2). Values are the
mean ± standard deviation of triplicate experiments.
In addition, cells of B.stratosphericus and D.radiodurans cells were showed less
sensitive to high doses (>1.0 kJm−2
) of UVA (84 and 56 % respectively). However,
E.Coli cells was appeared to be very sensitivity to high dose of UVA radiation (>1.0

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 144
kJm−2
) (less than 10%). The percent surveillance of all three strains under radiation
exposure was determined by mean SD values of colony forming units (Fig. 5.2).
5.3.2. Enhanced production of catalase (Bskat-1) activity of Bacillus stratosphericus
after exposure to UV and γ- radiation
The exposure of UV (Fig.5.3) and γ-radiation (Fig 5.4) on cells yielded
significant changes in activities of the antioxidant enzyme (catalase). γ-radiation
induces gradual increase in catalase activity in B. stratosphericus and D. radiodurans at
an exposure of 2 to 6 kGy respectively. The maximum induction of catalase was
determined at an exposure of 6 kGy (about 98%), however its activity was lowered to
80% beyond 6 kGy in control strain D radiodurans. In B. stratosphericus, while
absorbed 96% increase in activity at 4 kGy. However, dose beyond 4 kGy resulted in
drastic fall in activity (10%) from initial. On the contrary, pattern of change in catalase
activity was different in case of gamma irradiated E.coli. This strain, when irradiated at
2 kGy experienced less than one fold decrease in catalase.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 145
Figure 5.3: Catalase activity of B. stratosphericus, D. radiodurans R1 and E. coli at 1
to 12 kGy (radiation dose 4.87 kGy/h). The highest residual activity of B.
stratosphericus was observed at 4 kGy. Values were the mean ± standard deviation of
triplicate experiments.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 146
Figure 5.4: Catalase activity curves for B. stratosphericus (circle), Deinococcus
radiodurans R1 (box) and Escherichia coli (triangle) after following exposure to UVA
for one hour. Values were the mean ± standard deviation of triplicate experiments.
At an exposure of UV (>1.0 kJm−2) was also experienced the drastic changes in
enzyme activity in both B. stratosphericus and control group. It was noted that,
irradiation at >1.0 kJm−2 significant increased in the activity was observed (63.1% fold)
in B. stratosphericus and 60.23% in D radiodurans. Whereas, in E.coli, the effect of
UV does not affect much on the enzyme activity (8.23%). These results attributed that,
UV and γ-radiation induce catalase activity in B. stratosphericus and this may be
involve in protection from the oxidative stress generated from radiations.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 147
5.3.3. Purification and PAGE pattern of radiation induced catalase Bskat-1
Two chromatographic purification steps were employed for catalase Bskat-1.
The purification by initial HIC column, using decreasing gradient of 1.0-0 M
(NH4)2SO4, resulted in two peaks eluted at 800 and 420-440 mM (NH4)2SO4
respectively (Fig 5.5). At this stage, the fractions were pooled and enzyme activity was
determined to be 20,900 U/mg with a 6.76 fold, which was higher than the crude
extract. Further, the purification fold was enhanced to 9.77 fold with specific activity of
30,200 U/mg by concentrated on ultra filtration. The purification strategy for Bskat-1
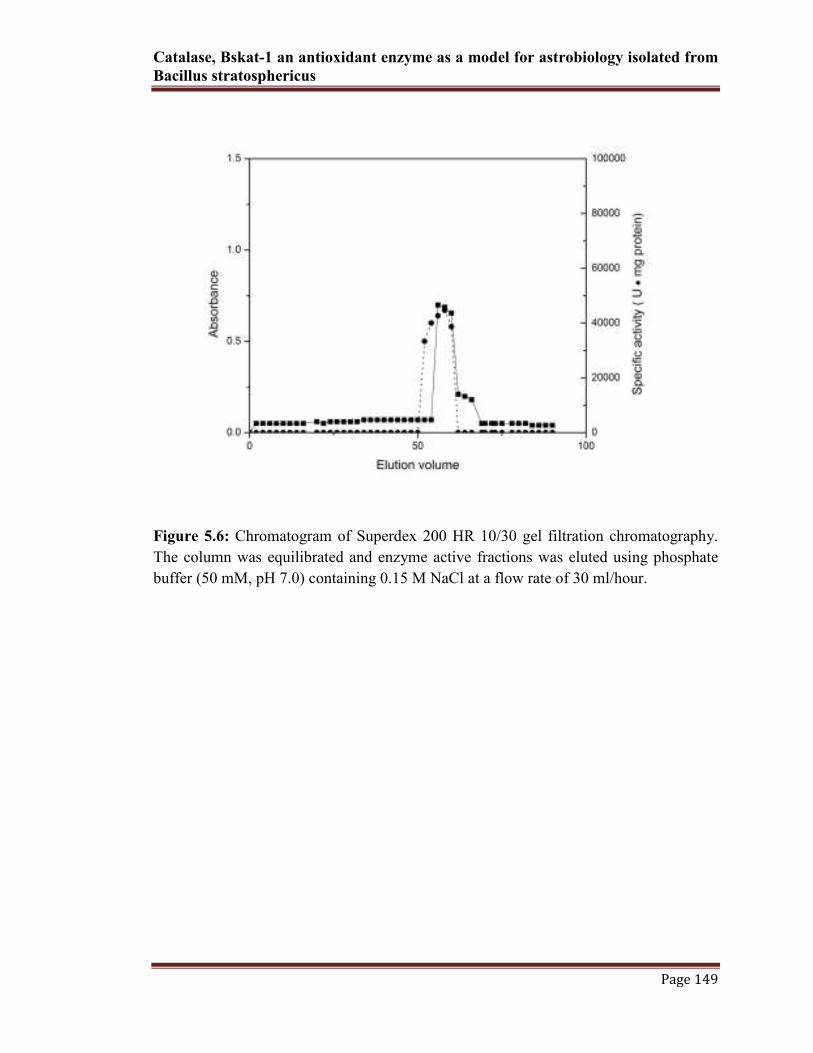
was determined to highest in superdex 200 HR gel filtration chromatographic column.
This step was resulted in single enzyme active peak (Fig 5.6). At this final purification
steps, the activity of the purified enzyme was determined to be 45,250 U/mg and 14.67
fold. The detailed series of purification steps and corresponding enzyme activity were
summarized in Table 5.1. Upon chromatographic elution from gel filtration, the
molecular mass of the purified enzyme was determined by 12.5% denaturating PAGE.
The electrophoresis pattern of the purified enzyme showed a single dark massive band
corresponding to 28 kDa (Fig 5.7). However, exact molecular weight of the enzyme was
determined by Sephacryl 200 HR gel filtration chromatography using standard markers
(described in material and methods). The exact molecular weight of the enzyme was
corresponds to 120 kDa (Fig. 5.8), which was indication of an enzyme with four
identical subunits. Since, catalase was a homo tetramer, containing four identical
subunits, which corresponds to the molecular weight of 120 kDa (characteristic of
catalase). The activity staining for catalase was determined by 8% native PAGE

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 148
(materials and methods), resulted in appearance of single white fluorescent colored
band in dark background (Fig. 5.7).
.
Figure 5.5: Bskat-1 was purified from phenyl sepharose high performance HIC
chromatography. The column was previously equilibrated with 1.0M (NH4)2SO4 in
phosphate buffer and fractions were eluted with a linear decreasing gradient of 1.0–0 M
(NH4)2SO4 at a flow rate of 30 ml/hour. The enzyme active fraction was eluted at 420
mM concentration (NH4)2SO4 (dotted lines).
Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 149
Figure 5.6: Chromatogram of Superdex 200 HR 10/30 gel filtration chromatography.
The column was equilibrated and enzyme active fractions was eluted using phosphate
buffer (50 mM, pH 7.0) containing 0.15 M NaCl at a flow rate of 30 ml/hour.

Cata
lase
, B
skat-
1 a
n a
nti
oxid
an
t en
zym
e as
a m
od
el f
or
ast
rob
iolo
gy i
sola
ted
fro
m B
aci
llu
s st
rato
sph
eric
us
P
ag
e 1
50
T
ota
l
act
ivit
y
(U)
Tota
l
pro
tein
(mg)
Sp
ecif
ic
act
ivit
y
(U/m
g)
Yie
ld
(%)
Pu
rifi
cati
on
(fold
)
Cel
l ex
tra
ct
Ph
enyl
Sep
haro
se
(HIC
)
Ult
ra f
iltr
ati
on
Su
per
dex
200
220
X10
-4
234
X10
-4
160
X10
-4
54.3
X10
-4
712
112
53
10
3090
20,9
00
30,2
00
45,2
50
100
106
72.7
24.6
8
1.0
6.7
6
9.7
7
14.6
4
On
e un
it (
U)
of
acti
vit
y i
s d
efin
ed, as
act
ivit
y r
equ
ired
des
troyin
g o
ne
µm
ol
H2
O2
per
min
Tab
le 5
.1:
Puri
fica
tion s
tage
pat
tern
of
cata
lase
(BsK
at-1)
from
the B. Stratosp
hericus

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 151
Figure 5.7: Non-reducing 12.5% PAGE pattern of purified Bskat-1 from B.
stratosphericus. Lane A, standard protein molecular weight markers. Lane B, untreated
native Bskat showing single band corresponding to 28 kDa. Lane C, activity profile of
catalase band was resolved on gel by incubating the gels in 5 mM H2O2, 2% ferric
chloride and 2% potassium ferric cyanide. The catlase band showing colored band in
dark background.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 152
Figure 5.8: Native molecular weight of Bskat-1 was determined using Sephacryl 200
HR gel filtration column chromatography. The column was previously equilibrated with
50 mM phosphate buffer, pH 7.0. The standards used were Myosin (205 kDa),
Phosphorylase B (97 kDa), Ovalbumin (45 kDa), Trypsinogen (24 kDa) and Aprotinin
(6.5 kDa) as a molecular weight standards. The molecular weight of Bskat-1 was 120
kDa (indicated on marked lines)
5.3.4. Effect of temperature and pH of Bskat-1
The activity of Bskat-1 as a function of temperature and pH was shown in
Figures 5.8 and 5.9. The enzyme had virtually no activity at below 15°C, and activity
increased with increasing temperature up to the maximum activity at 55°C. In contrast,
it had activity over the range of 20-55°C with an optimum temperature for activity at
45°C (Fig. 5.8a). This was slightly thermostable when compared to other mesophilic
catalase.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 153
(a)
(b)
Figure 5.8: (a) & (b) Effect of temperature and thermostability on Bskat-1 from B.
stratosphericus. The optimum temperature of enzymes activity was determined by
incubated for 30 min at 15 to 60⁰C prior to initiation of the reaction. The thermostability
was assayed at 45 to 75⁰C at different interval of time. Data represents the mean of at
least three independent experiments.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 154
The stability of the Bskat-1 was observed approximately 13% increase in
activity observed at 45°C for 1 h of incubation. At 55°C, the residual activity was 80%,
which was higher than that of initial 30 min of incubation time (60%). Post hours of
incubation, the residual activity was drastically lowered. When the enzyme was
incubated at more elevated temperatures (65-75°C), the enzyme activity appears to
cease more rapidly (Fig. 5.8b). It was determined that, Bskat-1 had activity over a broad
pH range of 4-12, with the optimal activity at pH 10. This pH range of activity was
comparable to typical monofunctional catalases (Fig. 5.9). The typical pI of the native
enzyme was determined to be 7.8.
Figure 5.9: Bskat-1 activity as a function of pH. Various buffers from pH 4.0 to pH
12.0 [Formic acid buffer (pH 3.3 to 4.3); Trimethylamine / acetic acid buffer (pH 4.3 to
5.3); Trimethylamine / carbonate buffer (pH 5.9 to 6.9) and Phosphate buffer (pH 7 to
12)] were used at optimum temperature. Data represents the mean of three independent
experiments

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 155
5.3.5. Effect of metal ions, inhibitors and spectral studies
Metal ions (1 mM) such as Na+, Mg
2+, K
+, and Mn
2+ did not shown effect on
catalase activity. However, Co2+
and Ba2+
showed a partial inhibition on the enzyme
(Table 5.2). Histidine and EDTA at a concentration of 10 mM reduced activity of 19%
and 27%, respectively. Catalase activity was strongly inhibited by hydroxylamine
hydrochloride, sodium cyanide and sodium azide at a concentration of (0.1 to 1 mM).
Catalase specific inhibitor like, 3-Amino-1, 2, 4-triazole had minimum inhibitory action
on Bskat-1 (Table 5.3). Inhibitory values of these chemical were corresponds to the
residual activity of the enzyme.
Table 5.2: Effect of inhibitors on BsKat-1 activity
Inhibitor Final concentration(mM) % inhibition
Sodium azide 0.1 89
1.0 97
Sodium cyanide 0.1 75
1.0 99
3-Amino-1,2,4-triazole 0.1 23
1.0 29
Hydroxylamine hydrochloride 0.1 82
1.0 99

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 156
Table 5.3: Effect of metal ions on BsKat-1 activity
Like a typical heme containing catalase, Bskat-1 showed a strong Soret peaks at
410 nm (Fig. 5.10a). This provides evidence that the Bskat-1 was a heme catalase rather
than a Mn-catalase. The peak was shifted from 410 to 422 nm on following the addition
of 10 mM sodium cyanide (Fig. 5.10a). This indicates that, binding of cyanide to the
heme moiety of the Bskat-1. However, spectra remained unchanged after the treatment
of the enzyme with 1 mM sodium dithionite. This characteristic was consistent with the
above observation that Bskat-1 lacks peroxidase activity. Treatment of the enzyme with
pyridine/NaOH and sodium dithionite produced pyridine hemochrome whose major
absorption peaks shifted from 410 to 420, 522 and 560 nm (Fig. 5.10b). This indicates
that Bskat-1 contains protoheme IX as its heme group. The protoheme content of this
enzyme was determined from the absorption of its pyridine hemochromogen at 420 nm.
There are 3.1 molecules of protoheme IX per tetrameric molecule of Bskat-1. The heme
content as well as the RZ (Reinheitszahl) A410/A280 ratio of 0.54 again indicates that
Bskat-1 shares properties of typical monofunctional catalases.
Control None 100
NaCl 1mM 89
KCl - 86
MnCl2 - 99
CoCl2 - 54
MgCl2 - 91
BaCl2 - 51
Histidine 10 mM 19
EDTA - 27
Effectors Concentration * Residual Activity (%)

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 157
(a)
(b)
Figure 5.10: (a) Absorption spectrum of native Bskat-1 and changes in spectra after the
treatment of 10 mM cyanide and 1 mM dithionite.
(b) Absorption spectrum of pyridine hemochrome of Bskat-1

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 158
5.4. Discussion
It was generally accepted that, the bacteria and fungi are present in the
stratosphere unlikely to survive the high exposure to ultraviolet and other types of
ionizing radiation (Yinjie et al., 2008). It is not surprising that, the stratospheric
microbes exhibit potentially UV and ionizing resistant physiology. However, study on
differential mechanism of radio-protective nature of microorganism was explained. B.
simplex cells, which were relatively resistant to such radiation, whose vegetative cells
tend to clump together; essentially because the outer cells provide a UV barrier
(Wainwright et al., 2003). Another studies on Bacillus spores revealed that, the survival
of nearly 6 years in space when shielded from UV radiation. Whereas, when exposed to
the full range of space conditions, only a small proportion of the spores from the
innermost part of the samples were able to survive (Roya et al., 2002). Another
mechanism of Sarcina lutea, which forms UV protective packets of cells, is 100 times
more resistant to UV than E coli, while B. subtilis and Staphylococcus aureus were
respectively three and eight times more resistant (Whisler, 1940). It was known that,
genus Deinococcus were extraordinarily resistant to UV and ionizing radiation and the
mechanism has been studied on repairing the damaged DNA. There have been,
however, very few published studies on radiation surveillance in terrestrial bacterial
species Deinococcus spp. and Bacillus spores have been used as the model for space
radiation experiment (Wayne et al., 2000). Some evidence suggests that, radiation
resistance in D. radiodurans involves the use of RecBC and scavenging enzyme
systems against oxidative DNA damage (Nivedita et al., 2008). Basal levels of catalase
and SOD activities in cell extracts of D. radiodurans cultures pretreated with 10 mM

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 159
H2O2 were more resistant to the lethal effect of γ-irradiation. Changes in the
concentration and activities of scavenging enzymes, such as catalase (kat) and
superoxide dismutase (SOD) were valid as biomarkers in stratospheric bacterial species.
The DNA damage was due to radiation induced reactive oxygen intermediates, which
leads to the impairment of SOD and catalase, the two important components of the
antioxidants defense system (Imam et al., 2010).
In the present research work, the cells of B. stratosphericus showed the
differential surveillance in both UVA and γ-radiations. It had shown nearly two log
cycles loss in γ-radiation beyond 4 kGy. However, the data from the present and
previous studies, the resistance of D. radiodurans and E. coli was 8 kGy and 2 kGy of
γ-radiation respectively (Issei et al., 2006). Moreover, Roya et al., (2002) exposed
thirteen isolates to 5.5 kGy of γ-radiation with D37 (dose for 37% survival) and all
isolates were shown resistant up to 3.5 kGy. It was also determined that, only 7%
survival at the very high dose of 5.5 kGy. In comparison, the D37 for E. coli was lower
than 100 Gy (Roya et al., 2002). Although, B. stratosphericus and D.radiodurans cells
were showed less sensitive to high doses (>1.0 kJm−2
) of UVA radiation. Whereas,
E.coli cells was appeared to be very sensitivity to high dose (Issei et al., 2006).
Catalase Bskat-1 from B. stratosphericus was purified using two
chromatographic steps. However, it was reported that, the requirement of various
chromatographic steps were varied in different bacterial catalases. After, the complete
purification by Superdex 200 HR gel filtration chromatography, the specific activity of
purified Bskat-1 was determined to be 48,000 U/mg (Table 5.1). However, this was not
particularly interesting when compared with the catalase activity of other radioresistant

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 160
and other bacterial species. It was reported that, after four chromatographic purification
steps, the specific activity of catalase (KatA) from D. radiodurans was determined to be
68,800 U/mg (Issei et al., 2006). The highest catalase activity was observed in D.
radiophilus, which is 3-fold higher than that of D. radiopugnans, which showed the
lowest activity. Although, Thompson et al., (2003) purified catalase from thermophilic
Thermus brockianus using the three chromatographic steps, that resulted in 65-fold
purification to a specific activity of 5300 U/mg. However, Xunlong et al., (1998) over
expressed and purified the recombinant secretary catalase from B. subtils requires only
two chromatographic steps. In his study, the simple purification steps yielded nearly
homogeneous catalase, with ~70% recovery. The purified recombinant catalase has a
specific activity of 34,600 U/mg under optimal conditions. The purified Bskat-1 was
shown at single band at 28 kDa in SDS–PAGE analysis and the gel filtration results
showed an approximate native protein molecular mass of 120 kDa, indicative of an
enzyme with four identical subunits. This was slightly lower than those reported for
other tetrameric catalase enzymes. i.e., Bacillus sp. with 70.5 and 282 kDa, E. coli with
84.3 and 337 kDa, Rhodobacter capsulatus with 59 and 236 kDa, Neurospora crassa
with 80 and 320 kDa and T. brockianus with 42.5 and 178 kDa subunit and native
molecular mass, respectively (Yumoto et al., 1990; Thompson et al., 2003). With
respect to Bskat-1, comparable size of the catalases has been observed in Streptomyces
venezuelae (Knoch et al., 1989), Mycobacterium tuberculosis (Wayne and Diaz, 1986),
and K. pneumoniae (Goldberg and Hochman, 1989). However, all of these catalases
were homodimers. A manganese-containing catalase from the obligate thermophile
Thermoleophilum album (Allgood and Perry, 1986) consists of four identical subunits

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 161
of 34 kDa. The presence of two different subunits was known only in the case of the
catalase-peroxidase of Vitreoscilla (Abrams and Webster, 1990). Loewen and Switala,
(1986) have explained the appearance of the E. coli HPII catalase in the form of two
subunits of 90 and 92 kDa with proteolytic degradation. In comparision, the molecular
weight of the catalases from radiation resistant Deinococcus spp. was varied from 650
kDa in D. proteolyticus to 155 kDa in D.radiophilus. In comparision, catalase KatA
from D. radiodurans 65 kDa by SDS-PAGE analysis and 240 kDa by gel filtration,
suggesting that KatA forms a same homotetramer in solution. The isoelectric point of
Bskat-1 was 7.8. However, based on the available literature, pI of Bskat-1 was slightly
above when compared to those reported for catalases and catalase-peroxides from
Halobacterium halobium (4.0), Thermoascus aurantiacus (4.5), Vitreoscilla sp. of (5.0
and 5.2), and Anacystis nidulans of (4.7). (Obinger et al., 1997; Brown et al., 1995;
Abrams and Webster, 1990; Wang et al., 1998).
The temperature optimum of the Bskat-1 was slightly moderate thermostable. It
was shown activity between 25-55ºC, with temperature optimum of 45ºC. It was slight
high when compared to the radiation resistant Deinococcus spp. and other mesophilic
bacterial catalases. At 55-60ºC, the residual activity of Bskat-1 was reduced to 70%.
However, catalase (KatA) from D. radiodurans was active over a temperature range
from 20°C to 70°C, with optimum activity occurring at 30°C. Also, other commercially
available catalase enzyme from bovine liver and A. niger, the optimum assay
temperature demonstrating the enzymatic activity of catalases was 40°C and 50°C to
60°C, respectively. At 70°C, the activity of A. niger, bovine liver and D. radiodurans
catalases was reduced to 82%, 45% and 20%, respectively, of each activity at the

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 162
optimum assay temperature. It was reported that, psycrophilic bacteria V. rumoiensis S-
1T secrete catalase, which was shown optimum at 40°C and completely suppressed by
incubation at 60°C. While, the M. luteus catalase activity was suppressed at 70°C (Isao
et al., 2000). Like strain S-1T
, at specific time of incubation, Bskat-1 loses its activity
above 60°C by 70% residual activity. At 90°C, the Bskat-1 residual activity was
completely suppressed to 10% for 15 min. These results suggested that the
thermostability of Bskat-1 is similar to the strain S-1T
catalase and lower than those of
the A. niger, M. luteus and bovine liver catalases (Isao et al., 2000). The pH stability of
the Bskat-1 was active over broad pH range of 4-12 with the optimal activity at pH 10,
which was closely related catalase-peroxidase from thermophilic B. stearothermophilus.
These results indicate the activity of Bskat-1 was more likely to alkaline environment.
However, catalases from B. bacillus and KN25 was shown lower pH stability than
Bskat-1 (Bol and Yasbin, 1991). In comparison, KN25 and bovine liver catalases was
shown highest activity between pH 5.0 and 8.0, with an optimal pH was determined at
6.0 (Xunlong et al., 1998). Similar to Bskat-1, strain S-1T
catalase exhibit same pH range
for the activity (pH 6-10) (Isao et al., 2000). Indicative of results from thermal stability
and pH, it can confine that the Bskat-1 was similar to the strain S-1T
catalase.
Like other catalase and catalase-perioxidase from Bacillus SF (Marinka et al.,
2001); D. radiodurans (Issei et al., 2006); V. rumoiensis (Isao et al., 2000);
hydroxylamine, azide, cyanide and 3-amino-1, 2, 4-triazole (Marcinkeviciene et al.,,
1995) had inhibitory effects on Bskat-1. It has been reported that, inhibitory action of
hydroxylamine on nonsulfur phototrophic bacterium Rhodobacter sphaeroides ATH
2.4.1 catalase. It had cyanide, azide, and hydroxylamine decreased its activity by 72.5,

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 163
76.4, and 85.1%, respectively (Detlef and Michael, 1998). It was also confirmed that, 2-
Mercaptoethanol and sodium dithionite at a concentration of was a less-potent inhibitor.
Other various metal ions like Na+, Mg
2+, K
+, and Mn
2+ had a stimulatory effect on
catalase activity. However, in case of Bskat-1, there was a partial inhibition was
observed by Co2+
and Ba2+
metal ions. Nevertheless, its inhibitory effect could not be
quantitated by the catalase assay because of interference with the oxygen electrode
(Detlef and Michael, 1998). In relation to catalase from Thermus brockianu, Bskat-1 is a
typical heme containing catalase (Thompson et al., 2003). Bskat-1 had shown a strong
Soret peaks at 410 nm. This provides evidence that the Bskat-1 was a heme catalase
rather than a Mn-catalase as have been described in Thermus species (Kagawa et al.,
1999). It was described that, typical Mn-catalases lacks the Soret peak (Kagawa et al.,
1999; Yumoto et al., 1990; Brown et al., 1995; Abrams and Webster, 1990). Although,
KatG from Mycobacterium tuberculosis had a Soret peak at 408 nm (Johnsson et al.,
1997). The Soret band was shifted to 422 nm upon the addition of CN- . Similar shifts
have also been described for other catalases but varying degree (Hochman and
Shemesh, 1987; Goldberg and Hochman, 1989; Hyoung-pyo et al., 1994; Isao et al.,
2000). However, sodium dithionite did not alter any spectrum of the native enzyme
indicating the negative effect on Bskat-1. Major absorption at 522 and 560 nm for
Bskat-1(with respect to range), which was typical for protoheme IX for other catalases
(Detlef and Michael, 1998; Hyoung-pyo et al., 1994; Isao et al., 2000). The Bskat-1 has
similar absorption for typical heme containing catlase and share common spectral
attribute among currently known catalases.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 164
Results from this investigation revealed that, a typical mono functional, moderate
temperature and alkali stable catalase (Bskat-1) was a potent antioxidant enzyme system
for radiation induced oxidative stress in B. stratosphericus. These extreme properties of
the novel enzyme could be a potential candidate for studying the mechanism of
protecting radiation-induced stress in space radiation experiments.

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 165
5.5. References
1. Abrams JJ, Webster DA (1990) Purification, partial characterization and
possible role of catalase in the bacterium Vitreoscilla. Arch. Biochem. Biophys.
279 (1): 54-59.
2. Allgood GS, Perry JJ (1986) Characterization of a manganese-containing
catalase from the obligate thermophileThermoleophilum album. J. Bacteriol.
168: 563-567
3. Amiri N, Finkbeiner M, Hamilton S, Kibenge P (2007) The role of reactive
oxygen species in UVA-mediated killing of Escherichia coli. J. Exp. Microbiol.
Immunol. 11: 42-46.
4. Banerjee S, Salunkhe S, Apte-Deshpande A, Mandi N, Mandal G, Padmanabhan
S (2009) Over-expression of proteins using a modified pBAD24 vector in E. coli
expression system. Biotechnol. Lett. 31: 1031-1036
5. Bol DK, Yasbin RE (1991) The isolation, cloning and identification of a
vegetative catalase gene from Bacillus subtilis. Gene. 109: 31–37
6. Brown-Peterson NJ, Salin ML (1995) Purification and characterization of a
mesohalic catalase from the halophilic bacterium Halobacterium halobium. J.
Bacteriol. 177 (2): 378-384.
7. Cha MK, Kim IH (1996) Purification and characterization of thiol specific
antioxidant protein from human liver: A mer5-like human isozyme. J. Biochem.
Mol. Biol. 29: 236-240.
8. Cha MK, Kim IH (1999) Thioredoxin in the periplasmic space of Escherichia
coli as a physiological electron donor to periplasmic thiol peroxidase, p20. J.
Biochem. Mol. Biol. 32:168-172
9. Chan E, Kuang L, Lau A, Wang J (2010) Antisense mRNA method as an
alternative to generate a catalase double knockout phenotype in a Escherichia
coli katG mutant. J. Exp. Microbiol. Immunol. 14: 127134
10. Cheng M, Chow A, Ho J, Luk B (2009) katE complementation fails to protect
against UV-A-mediated killing in catalase-deficient Escherichia coli. J. Exp.
Microbiol. Immunol. 13: 63-66.
11. Detlef PT, Michael B (1998) Purification and characterization of a catalase from
the nonsulfur phototrophic bacterium Rhodobacter sphaeroides ATH 2.4.1 and
its role in the oxidative stress response. Arch. Microbiol. 169: 503-508
12. Eisenstark A (1989) Bacterial genes involved in response to near-ultraviolet
radiation. Adv. Genetics. 26:100-129
13. Falk JE (1964) Porphyrins and Metalloporphyrins; Elsevier: Amsterdam,
14. Goldberg I, Hochman A (1989) Purification and characterization of a novel type
of catalase from the bacterium Klebsiella pneumoniae. Biochim Biophys Acta.
991: 330-336
15. Goldberg I, Hochman A (1989) Three different types of catalases in Klebsiella
pneumoniae. Arch. Biochem. Biophys. 268: 124-128
16. Halliwell B (1990) How to characterize a biological antioxidant. Free Radical
Res. Commun. 9: 1-32.
17. Halliwell B, Gutteridge JMC (1999) Free Radicals in Biology and Medicine. 3rd
edition. pp: 105-350. Oxford Univ. Press, Oxford, UK

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 166
18. Hassan HM, Fridovich I (1979) Intracellular production of superoxide radical
and of hydrogen peroxide by redox active compounds. Arch. Biochem. Biophys.
196: 385-395
19. Hedrick JL, Smith AJ (1968) Size and charge isomer separation and estimation
of molecular weights of proteins by disc gel electrophoresis. Arch. Biochem.
Biophys. 126: 155-164.
20. Hochman A, Figueredo A, Wall JD (1992) Physiological function of
hydroperoxidase in Rhodobacter capsulatus. J Bacteriol. 174: 3386-3391
21. Hochman A, Goldberg I (1991) Purification and characterization of a catalase-
peroxidase from the photosynthetic bacterium Rhodopseudomonas capsulata. J.
Biol. Chem. 262: 6871-6876.
22. Horneck G (1993) Responses of Bacillus subtilis spores to space environment:
Results from experiments in space. Origin. Life Evol. Biosph. 23: 37-52.
23. Horneck G, Buecker H, Reitz G (1994) Long-term survival of bacterial spores in
space. Adv. Space Res. 10: 41-45
24. Horneck G, Miliekowsky C, Melosh HJ, Wilson JW, Cucinotta FA, Gladman B
(2002) Viable transfer of microorganisms in the solar system and beyond. In
Astrobiology: The Quest for the Conditions of Life. (Horneck G, Baumstark-
Khan C. eds.), Springer. Berlin, Germany.
25. Hyoung-pyo K, Jong-Soo L, Yung CH, Jung-Hye R (1994) Characterization of
the major catalase from Streptomyces coelicolor ATCC 10147. Microbiology.
140: 123391-3397
26. Imam AAM, Usama MM, Alaa GO, Alaa El-Din HS (2010) Effects of
ultraviolet A on the activity of two metabolic enzymes, DNA damage and lipid
peroxidation during early developmental stages of the African catfish, Clarias
gariepinus (Burchell, 1822) Fish. Physiol. Biochem. (2010) 36: 605–626
27. Isao Y, Daisen I, Hideaki I, Anita I, Nobutoshi I, Hidetoshi M, Hidetoshi O,
Kosei K (2003) Purification and characterization of a catalase from the
facultative psychrophilic bacterium Vibrio rumoiensis S-1T
exhibiting high
catalase activity. J. Bacteriol. 182 (7): 1903-1909
28. Issei K, Takashi T, Haitham S, Issay N, Shotaro Y, Koichi U, Kenji I (2006)
Characterization of monofunctional catalase KatA from radioresistant Bacterium
Deinococcus radiodurans. J. Biosci. Bioeng. 101(4): 315-321
29. Johnsson K, Froland WA, Schultz PG (1997) Overexpression, purification, and
characterization of the catalase-peroxidase KatG from Mycobacterium
tuberculosis. J. Biol.Chem. 272(5): 2834-2840.
30. Jurkiewicz BA, Buettner GR (1994) Ultraviolet light induced free radical
formation in skin: An electron paramagnetic resonance study. Photochem.
Photobiol. 59: 1-4
31. Kagawa M, Murakoshi N, Nishikawa Y, Matsumoto G, Kurata Y, Mizobata T,
Kawata Y, Nagai J (1999) Purification and cloning of a thermostable manganese
catalase from a thermophilic bacterium. Arch. Biochem. Biophys. 363(2): 346-
355.
32. Klotz MG, Klassen GR, Loewen PC (1997) Phylogenetic relationships among
prokaryotic and eukaryotic catalases. Mol. Biol. Evol. 14: 951-958

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 167
33. Knoch M, Van Pee KH, Vining LC, Lingens F (1989) Purification, properties
and immunological detection of a bromoperoxidase catalase from Streptomyces
venezuelae and from a chloramphenicol-nonproducing strain. J. Gen. Microbiol.
135: 2493-2502
34. Laemmli UK (1970) Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature. 277: 680-685.
35. Leven S, Heimberger A, Eisenstark A (1990) Catalase HP1 influence membrane
permeability in Escherichia coli following near-UV stress. Biochem. Biophys.
Res. Commun. 171: 1224-1228.
36. Loewen PC, Switala J (1986) Purification and characterization of catalase HPII
from Escherichia coli K12. Biochem. Cell. Biol. 64: 638-646
37. Lowry OH, Rosebrough, NJ, Farr AL, Randall RJ (1951) Protein measurement
with the Folin phenol reagent. J. Biol. Chem. 193: 265–275.
38. Marcinkeviciene JA, Magliozzo RS, Blanchard JS (1995) Purification and
characterization of the Mycobacterium smegmatis catalase peroxidase involved
in isoniazid activation. J. Biol. Chem. 270: 22290-22295
39. Margoliash E, Novogrodsky A, Schekter A (1960) Irreversible reaction of 3-
amino-1:2:4-triazole and related inhibitors with the protein of catalase. Biochem
J. 74: 339-348
40. Marinka G, Gilbert OF, Andreas P, Fritz L, Karl-Heinz R, Artut CP, Georg MG
(2001) A catalase-peroxidase from a newly isolated thermoalkaliphilic Bacillus
sp. with potential for the treatment of textile bleaching effluents Extremophiles.
5: 423-429
41. Meir E, Yagil E (1984) Catalase-negative mutants of Escherichia coli. Curr.
Microbiol. 11: 13-18
42. Mulvey M, Switala J, Borys A, Loewen P (1990) Regulation of transcription of
katE and katF in Escherichia coli. J. Bacteriol. 172: 6713-6720.
43. Nak-Kuyn S, Young NL (2000) Iso-catalase Profiles of Deinococcus spp. J.
Biochem. Mol. Biol. 33(5): 412-416
44. Nicholson WL, Munakata N, Horneck G, Melosh HJ, Setlow P (2000)
Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial
environments. Microbiol. Mol. Biol. Rev. 64: 548-572
45. Nivedita PK, Vidya AK, Hari SM (2008) RecBC enzyme overproduction affects
UV and gamma radiation survival of Deinococcus radiodurans. DNA. Repair. 7:
40-47
46. Obinger C, Regelsberger G, Strasser G, Burner U, Peschek GA (1997)
Purification and characterization of a homodimeric catalase-peroxidase from the
cyanobacterium Anacystisnidulans. Biochem. Biophys. Res. Commun. 235: 545-
552.
47. Pabulo HR (2010) Resistance of microorganisms to extreme environmental
conditions and its contribution to Astrobiology. Sustainability. 2: 1602-1623
48. Rampelotto PH, Rosa MB, Schuch NJ, Munakata N (2009) Exobiological
application of spore dosimeter in studies involving solar UV radiation. Orig.
Life. Evol. Biosph. 39: 373-374
49. Rettberg P, Eschweiler U, Strauch K, Reitz G, Horneck G, Wänke H, Brack A,
Barbier B (2002) Survival of microorganisms in space protected by meteorite

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 168
material: Results of the experiment-Exobiologie of the perseus mission. Adv.
Space Res. 30: 1539-1545
50. Roya S, Renu N, Dennis S, Frank TR, Joseph MD, Marvin S, Leon O, Roger JT,
Jocelyne D (2002) Microbial survival of space vacuum and extreme ultraviolet
irradiation: strain isolation and analysis during a rocket fight. FEMS. Microbiol.
Lett. 215: 163-168
51. Shivaji S, Chaturvedi P, Begum Z, Pindi PK, Manorama R, Padmanaban DA,
Shouche YS, Pawar S, Vaishampayan P, Dutt CBS, Datta GN, Manchanda RK,
Rao UR, Bhargava PM, Narlikar JV (2009) Janibacter hoylei sp. nov., Bacillus
isronensis sp. nov. and Bacillus aryabhattai sp. nov., isolated from cryotubes
used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol.
59: 2977-2986
52. Shivaji S, Chaturvedi P, Suresh K, Reddy GSN, Dutt CBS, Wainwright MJ,
Narlikar V, Bhargava PM (2006) Bacillus aerius sp. nov., Bacillus aerophilus
sp. nov., Bacillus stratosphericus sp. nov. and Bacillus altitudinis sp. nov.,
isolated from cryogenic tubes used for collecting air samples from high
altitudes. Int. J. Sys. Evol. Microbiol. 56: 1465-1473
53. Soung NK, Lee YN (2000) Iso-catalase profiles of Deinococcus spp. J.
Biochem. Mol. Biol. 33: 412-416
54. Switala J, Loewen PC (2002) Diversity of properties among catalases. Arch.
Biochem. Biophys. 401: 145-154
55. Switala J, O’Neil JO, Loewen PC (1999) Catalase HPII from Escherichia coli
exhibits enhanced resistance to denaturation. Biochemistry. 38: 3895-3901
56. Thompson VS, Schaller KD, Apel WA (2003) Purification and characterization
of a novel thermo-alkali stable catalase from Thermus brockianus. Biotechnol.
Prog. 19(4): 1292-1299
57. Triggs-Raine B, Doble B, Mulvey M, Sorby P, Loewen P (1988) Nucleotide
sequence of katG, encoding catalase HPI of Escherichia coli. J. Bacteriol. 170:
4415-4419
58. Van Ossowski I, Mulvey M, Leco O, Borys A, Loewen P (1991) Nucleotide
sequence of Escherichia coli KatE, which encodes catalase HPII. J. Bacteriol.
173: 514-520.
59. Vile GF, Tyrrell RM (1993) Oxidative stress resulting from ultraviolet A
irradiation of human skin fibroblast leads to heme oxygenase-dependent
increase in ferritin. J. Bol. Chem. 268: 14678-14681
60. Wainwright M, Wickramasinghe NC, Narlikar JV, Rajaratnam P (2003)
Microorganisms cultured from stratospheric air samples obtained at 41 km.
FEMS. Microbiol. Lett. 218: 161-165.
61. Wang H, Tokusige Y, Shinoyama H, Fujii T, Urakami T (1998) Purification and
characterization of a thermostable catalase from culture broth of Thermoascus
aurantiacus. J. Ferment. Bioeng. 85(2): 169-173
62. Wayne LG, Diaz GA (1986) A double staining method for differentiating
between two classes of mycobacterial catalase in polyacrylamide electrophoresis
gels. Anal. Biochem. 157: 89–92

Catalase, Bskat-1 an antioxidant enzyme as a model for astrobiology isolated from
Bacillus stratosphericus
Page 169
63. Wayne LN, Nobuo M, Gerda H, Henry JM, Peter S (2000) Resistance of
Bacillus endospores to extreme terrestrial and extraterrestrial environments.
Microbiol. Mol. Biol. Rev. 64(3): 548-572
64. Whisler BA (1940) The efficacy of ultra-violet light sources in killing bacteria
suspended in air. Iowa State Coll. J. Sci. 14: 215-231
65. Wickramasinghe C (2004) The universe: A cryogenic habitat for microbial life.
Cryobiology. 48: 113–125
66. Xunlong SMF, Yujie Z, Xin GPZ (2008) Overexpression, purification and
characterization of a recombinant secretary catalase from Bacillus subtilis.
Biotechnol Lett. 30: 181-186
67. Yinjie Y, Shiho I, Shin-ichi Y, Akihiko Y (2008) UV-resistant bacteria isolated
from upper troposphere and lower stratosphere. Biol. Sci. Space. 22(1): 18-25
68. Yumoto I, Fukumori Y, Yamanaka T (1990) Purification and characterization of
catalase from a facultative alkalophilic Bacillus. J. Biochem. 108: 583-587.
69. Yun YS, Lee YN (2001) Superoxide dismutase profiles in the mesophilic
Deinococcus species. J. Microbiol. 39: 232-235.