calcium modulates fatty acid dynamics in rat liver plasma membranes
TRANSCRIPT

Eur. J. Biochem. 132, 517-524 (1983) $j FEBS 1983
Calcium Modulates Fatty Acid Dynamics in Rat Liver Plasma Membranes
Friedhelm SCHROEDER and Carlos SOLER-ARGILAGA
Department of Pharmacology, School of Medicine, University of Missouri - Columbia, Columbia, MO
(Received January 5 , 1983) - EJB 830004
Modulation of free fatty acid binding in isolated rat liver plasma membranes was evaluated using the fluores- cent fatty acids trans-parinaric and cis-parinaric acid as analogues for saturated and unsaturated fatty acids, respectively. Binding of trans-parinarate but not cis-parinarate was inhibited by physiological levels of Ca2 +. The effect was reversed by addition of excess EGTA. Calcium decreased the aqueous to lipid partition coefficient, K,, of truns-parinaric acid for liver plasma membranes while increasing the K,, for cis-parinaric acid. In addition, Ca2 + also altered the fluorescence lifetime, the quantum yield, and the relative partitioning of trans-parinaric and cis-parinaric acid into fluid and solid phases. Calcium and EGTA did not affect the binding of 1,6-diphenyl- 1,3,5-hexatriene. The effect of Ca2+ on the liver plasma membrane structure was to increase the rigidity of the membrane, primarily the solid domain. The fluorescence polarization of Irons-parinarate, cis-parinarate, and 1,6-diphenyl-l,3,5-hexatriene at 24 "C in liver plasma membranes in the absence of Ca" was 0.295 f 0.008, 0.253 0.007, and 0.284 f 0.005, respectively. Calcium (2.4 mM) increased the polarization of these probe molecules in liver plasma membranes by 8- 10%. EGTA (3.4 mM) reversed or abolished the increase in polari- zation. Thus, the fluorescent fatty acids fruns-parinarate and cis-parinarate may be used to monitor fatty acid binding by isolated membranes, to evaluate factors such as Ca2+ which modulate fatty acid binding, and to investigate the microenvironment in which the fatty acids reside. The data suggest that Ca2+ may be an important regulator of Fatty acid uptake by the liver plasma membrane, and thereby interact with intermediary metabolism of lipids at a step not involving lipolytic or synthetic enzymes.
Fatty acids play a major role both in cellular energy metabolism and in cellular membrane structure and function. Despite extensive investigations into cellular uptake of Fatty acids, there is still no clear answer as to how fatty acids enter the cell. Early data of Goodman [I] and Spector et al. [2] indi- cated that the fatty acids were first adsorbed to erythrocyte and Ehrlich ascites cellular binding sites from which they were internalized by unknown mechanisms. Recently, Weisiger et al. [3], using a perfused liver preparation and Abumrad et al. [4], using adipocytes, determined that fatty acid transfer across the cell membrane was dependent on binding of an albumin-fdtty acid complex or free fatty acid to a receptor on the cell surface. The existence of such a saturable fatty acid protein carrier in the plasma membrane of hepatocytes, embryonic chick cardiac cells, and of rat adipocytes was also postulated by others [5 - 71. In contrast, a simple diffusion mechanism for fatty acid transfer across plasma membranes was also demonstrated by several investigators [4,8,9]. In either case, most of these studies have been hampered in their measurement of fatty acid uptake because of complex inter- relationships and exchange equilibria between free fatty acids and albumin binding sites, cell membrane binding sites, intra- cellular binding sites, and intracellular metabolism among other factors [4]. In addition, nothing is known of dynamic events such as binding and partitioning of fatty acids within cell surface membranes or the role that modulating factors such as fatty acid structure, Ca2+, and hormones play in determining intramembrane fatty acid dynamics. Whole cell data with Ehrlich ascites cells indicated that the permeation
Symbols. Fa, absorbance-corrected fluorescence; Fre, relative fluo- rescence efficiency; F,,, corrected fluorescence emission; t, fluorescence lifetime.
of Fatty acids across the surface membrane may indeed depend on fatty acid unsaturation. Uptake appeared to in- crease linearly with decreasing number of double bonds [lo]. Another modulator, calcium, altered lipid dynamics in rat hepatocyte plasma membranes [l I ] and elicited gross struc- tural changes in liver plasma membranes upon binding of insulin [I 2,131. The binding affinity of Ca2+ itself to rat liver cell membranes was regulated by hormones such as insulin, glucagon, epinephrine, and cyclic AMP [14,15]. Roles for Ca2+ in lipid metabolism were also evident. Interference with calcium fluxes blocked the effects of glucagon on lipid mctab- olism indicating that calcium distribution and metabolic effects were functionally related [16]. Alterations in the distri- bution of intracellular calcium changed the incorporation of fatty acids into glycerides of leukocytes [17]. Hormonc- induced redistribution of calcium may regulate the rate of fatty acid oxidation during different metabolic states and thereby alter the partitioning of fatty acids between the path- ways of oxidation and esterification [I 81. Calcium stimulated fatty acid synthesis in isolated rat hepatocytes [19]. Despite these advances little is known about molecular events or f x - tors regulating fatty acid binding, uptake, or transport across the liver plasma membrane. Calcium levels in thc plasma membrane could be involved in fine adjustment of such an uptake mcchanism. Certainly the ion could provide a tre- mendous driving force for fatty acid uptake across the liver plasma membrane due to its 103-fold greater abundance in the extracellular fluid.
Therefore, in the present work the effect of Ca2+ on the dynamics and binding of fatty acids by isolated rat liver plasma membranes was investigated using fluorescent fatty acids. The advantages of using the fluorescent parinaroyl fatty acids are several. Trans and cis forms of the 18-carbon fatty

51x
acid, parinaric acid, are available. The trans form resembles a saturated fatty acid in structure, while the cis form resembles an unsaturated fatty acid. Neither trans-parinarate nor cis-parinarate fluoresce in aqueous solution [I 3,20 - 271 ; they fluoresce only in hydrophobic environments (e.g. within a membrane). An additional advantage is that fatty acid bind- ing by the membranes can be measured as an increase in fluo- rescence without separation of the liver plasma membrane from unbound free fatty acid. The fluorescent fatty acids, in in addition to providing quantitative information about fatty acid dynamics within the plasma membranes, provide quali- tative and quantitative information on the membrane micro- environment within which they reside.
MATERIALS AND METHODS
trans-Parinaric and cis-parinaric acid were obtained from Molecular Probes Inc. (Plano, TX). 1,6-Diphenyl-l,3,5-hexa- triene was purchased from Aldrich Chemical Co. (Milwaukee, Wl).
Preparation of' Liver Plasma Membranes. Male Sprague- Dawley rats [strain CRL : CD(SD)BR, Charles River Breeding Labs, Wilmington, MA] weighing 200- 300 g were fed a standard laboratory diet (Purina Laboratory Chow) ad libi- rum prior to sacrifice. The liver plasma membranes were iso- lated by the method of Pohl [28] as described in the previous paper of this series [29]. Protein was determined by the method of Lowry et al. [30].
Incorporation of Fluorrscmce Probe Molecules in Liver Plasma Membranes. Unless otherwise specified, trans-pari- narate, cis-parinarate, and 1,6-diphenyl-l,3,5-hexatriene were incorporated into liver plasma membranes at probe to phos- pholipid molar ratios of 1 : 100 or less as described elsewhere [13,20-231, and in the previous paper of this series [29]. The buffers used were 0.15 M Tris, pH 7.2, to which (A) 2.4 mM Ca2+, (B) 1 mM EGTA, or (C) 1 mM EGTA + 3.4 mM Ca2' were added. All samples were stirred continuously with a rcsident stirring bar in the cuvette using a Mark I1 micro- stirrer (Lawrence Instr. Ltd, London, Ontario, Canada).
Absorption Spectroscopy. Spectra were recorded with a Cary 14 UV-Vis scanning spectrophotometer and by the computer-centered spectrofluorimeter [31- 331. The refrac- tive index, n, and the polarizability, a, were calculated as described in the preceding paper [29].
Fluorescence Spectroscopy. Fluorescence measurements were made with two instruments. Absorbance, absorbance- corrected fluorescence, relative fluorescence efficiency, and corrected fluorescence emission were determined simultane- ously with the computer-centered spectrofluorimeter of Hol- land et al. 131,331, while fluorescence lifetime, 5, and steady- state fluorescence polarization, P, were determined using a T format SLM 4500 Subnanosecond Speclrofluorimeter (SLM Instruments Inc., Champaign-Urbana, IL). Details of the procedures for measuring these quantities are given in the previous paper in this series [29]. Fluorescence quantum yields, Qtotal, were determined experimentally as described earlier [29], from fluorescence and absorbance measurements as well as from fluorescence lifetime data.
Purtitioning of Probe Molecules between Aqueous and Liver Plusma Membrane Phases. A direct binding assay [29] was used to determine the mole fraction partition coefficient.
C'llculu tion of D issoc ia t ion Cons tun ts for trans- Par inur ic and cis-Parinaric Acid in Liver Plasma Membranes. Dissocia- tion constants, Kd? were determined as described in the pre- ceding paper [29] from double-reciprocal plots of [absor-
bance-corrected fluorescence]-' vs [parinaric acid] ~ in which either the membrane concentration was maintained constant at 50 yg protein/ml buffer while the probe concentration was varied from 0- 73 yM or the probe concentration was varied.
Determination of the Partition of' Parinuric Acid between Fluid and Solid Liuid Phases in Rut Liver Plasma Membranes. The partitioning of probe molecules between coexisting solid and fluid phases was determined as described in the preceding paper [29].
Fatty Acid Binding. The binding of fluorescent fatty acids, trans-parinarate and cis-parinarate, by liver plasma mem- branes was determined by measurement of the increase in absorbance-corrected fluorescence as a function of time when the fatty acids were incubated with the liver plasma mem- branes as stated above [29].
RESULTS
Effect of Ca2+ and EGTA on trans-Purinaric and &-Parinuric Acid Binding to Rat Liver Plasma Membranes
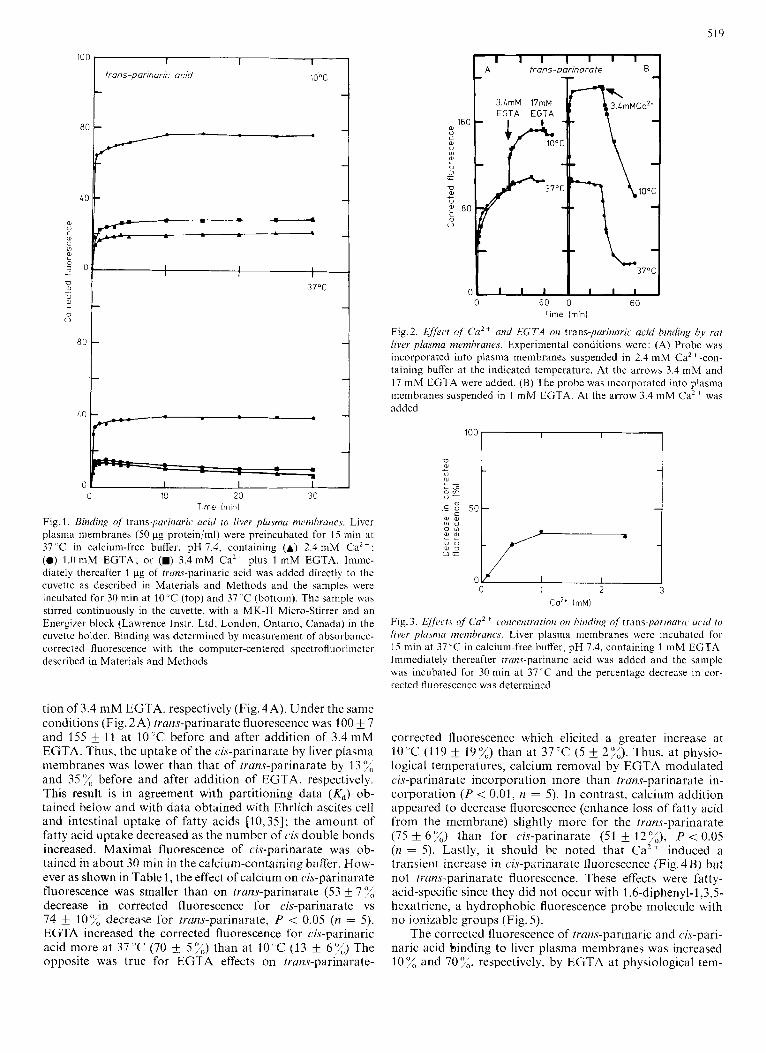
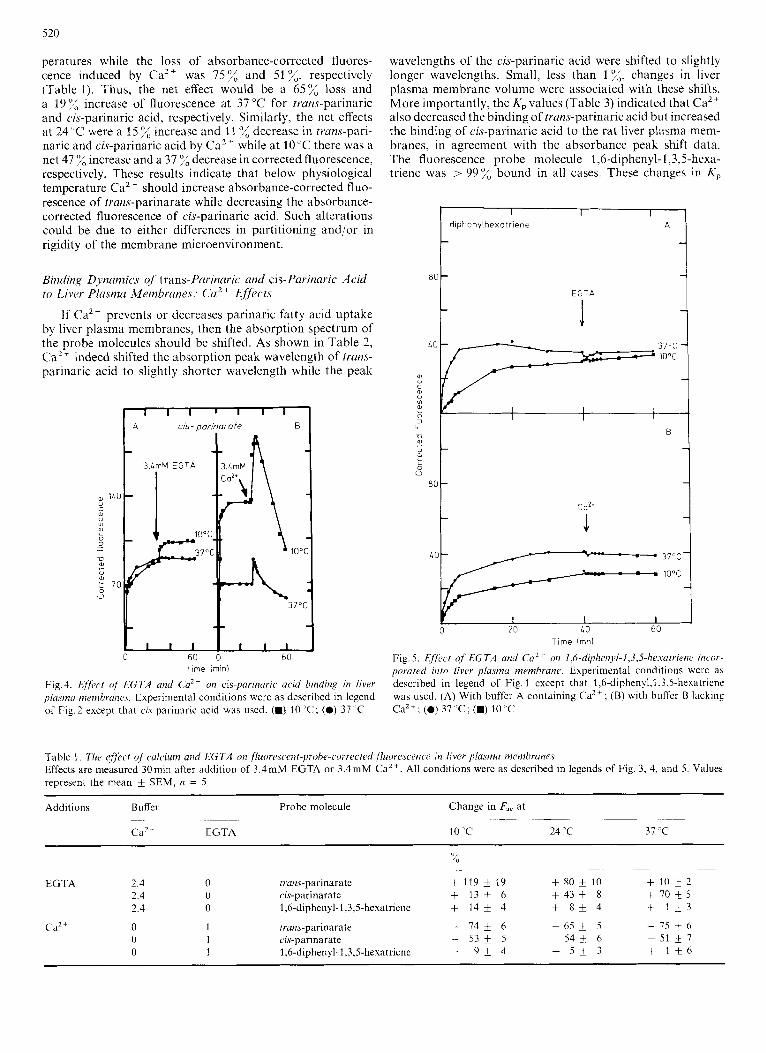
The binding of trans-parinaric acid by liver plasma mem- branes occurred very rapidly (Fig. 1). The corrected fluores- cence due to trans-parinaric acid increased to maximal values within 1-3 min both at 10 "C and 3 7 T . Similar data were obtained with cis-parinarate (data not shown). Two types of binding experiments were performed. First, the fluores- cent fatty acid, calcium and/or EGTA were added at the same time to liver plasma membranes in order to determine if Ca2+ could prevent fatty acid binding. Second, the liver plasma membranes were preincubated with the fluorescent fatty acid followed by addition of Ca2+ and/or EGTA to determine if the Ca2+ could elicit release of the fluorescent fatty acids from the membranes. Fatty acid binding at IO'C measured by absorbance-corrected fluorescence intensity of trans-parinarate in liver plasma membranes was approxi- mately 0.5-times lower in the calcium-containing buffer (2.4 m M Ca2+) than in the calcium-deficient buffer (1 mM EGTA) ; while at 37 ̂ C binding was approximately equivalent (compare Fig.2A vs 2B). The 2.4 mM Ca2+ concentration was chosen because this concentration elicited a maximal decrease of trans-parinaric acid absorbance-corrected fluores- cence (Fig. 3), and because it was the same as the physiological calcium concentration found in plasma [34]. When 3.4 mM EGTA was added to the liver membranes in 2.4 mM calcium buffer (Fig. 5 A) the corrected fluorescence increased 119 f 1 0 X a t IO"Cand5 2%at37"C(seeTable l).Incontrast, addition of 3.4 mM calcium to liver plasma membranes in buffer containing 1 mM EGTA decreased the corrected fluo- rescence by 74 f 10 % and 75 fi 6 % at 10 "C and 37 'C, respec- tively (Fig. 2 B and Table 1). The increase in trans-parinarate, corrected fluorescence, in 2.4 mM Ca2+ buffer after addition of 3.4 mW EGTA (Fig. 2) was slow and required longer than 30 min to reach a maximum. In the 1 mM EGTA buffer, the corrected fluorescence reached a maximal value in 1 - 3 min. Thus, calcium appeared to both prcvent the incorporation of trans-parinarate into liver plasma membranes and to induce loss of trans-parinarate from the membrane.
Since naturally occurring unsaturated fatty acids have cis double bonds, cis-parinarate should behave similarly to cis- unsaturated fatty acids while truns-parinate should resemble saturated fatty acids. The uptake of cis-parinarate is shown in Fig. 4. The absorbance-corrected fluorescence reached maxima of 87 f 6 and 100 & 4 at 10 "C before and after addi-
519
100 1 I I
irons-parinuric acid 10°C
U q 0 ti
37oc
8ot
0 10 20 30 Time lrnin)
Fig. 1. Binding of' trans-parinuric acid to liver pla.sma nzemhrancs. Liver plasma membranes (50 pg protein/ml) were preincubated for 15 min at 37 ' C in calcium-free buffer, pH 7.4, containing (A) 2.4 mM CaZt ; (0) 1.0 mM EGTA; or (W) 3.4 mM Ca2+ plus 1 mM EGTA. Imme- diately thereafter 1 pg of trans-parinaric acid was added directly to the cuvette as described in Materials and Methods and the samples were incubated for 30 min at 10 'C (top) and 37 "C (bottom). The sample was stirred continuously in the cuvette, with a MK-I1 Micro-Stirrer and an Energizer block (Lawrence Instr. Ltd. London, Ontario, Canada) in the cuvette holder. Binding was determined by measurement of absorbance- corrected fluorescence with the computer-centered spectrofluorimeter described in Materials and Methods
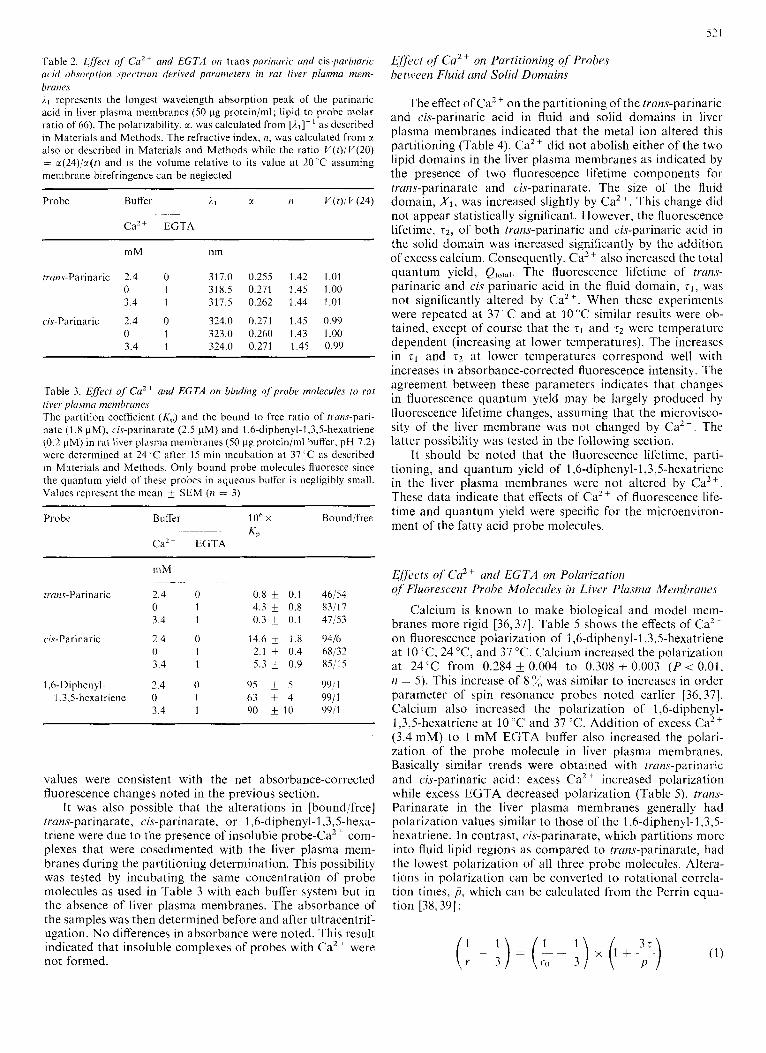
tion of 3.4 mM EGTA, respectively (Fig. 4A). Under the same conditions (Fig. 2A) trans-parinarate fluorescence was 100 & 7 and 155 f 11 at 10 "C before and after addition of 3.4 mM EGTA. Thus, the uptake of the cis-parinarate by liver plasma membranes was lower than that of trans-parinarate by 13% and 35%, before and after addition of EGTA, respectively. This result is in agreement with partitioning data ( K d ) ob- tained below and with data obtained with Ehrlich ascites cell and intestinal uptake of fatty acids [10,35]; the amount of fatty acid uptake decreased as the number of cis double bonds increased. Maximal fluorescence of cis-parinarate was ob- tained in about 30 min in the calcium-containing buffer. How- ever as shown in Table l , the effect of calcium on cis-parinarate fluorescence was smaller than on trans-parinarate (53 f 7 % decrease in corrected fluorescence for cis-parinarate vs 74 k 10'x decrease for trans-parinarate, P < 0.05 (n = 5). EGTA increased the corrected fluorescence for cis-parinaric acid more at 37 'C (70 & 5 %) than at 10°C (13 6 7 3 The opposite was true for EGTA effects on tram-parinarate-
I I I l I I I A trans-parinarate B
160 U C a
Ul
2 2
a U
U
- - 1
: 80 L 0 0
0 0 60 0 60
Time (mini
Fig.2. &#kt of Ca2+ and EGTA on trans-pavinaric acid binding by rut liver plasma membranes. Experimental conditions were : (A) Probe was incorporated into plasma membranes suspended in 2.4 mM Cazt-con- taining buffer at the indicated temperature. At the arrows 3.4 m M and 17 m M EGTA were added. (B) The probe was incorporated into plasma membranes suspended in 1 mM EGTA. At the arrow 3.4 niM CaZt was added
La2* (mM)
Fig. 3. Ejfi~crs of Ca2 ' c.onreritration ow hinding of trans-parinaric w i d to livcr plasnia membranes. Liver plasma membranes were incubated for 15 min at 37°C in calcium-free buffer, pH 7.4, containing 1 mM EGTA. Immediately thereafter rrans-parinaric acid was added and the sample was incubated for 30 min at 37°C and the percentage decrease in cor- rected fluorescence was determined
corrected fluorescence which elicited a greater increase at 10°C (119 f 19%) than at 3 7 T ( 5 -t 2%). Thus, at physio- logical temperatures, calcium removal by EGTA modulated cis-parinarate incorporation more than trans-parinarate in- corporation ( P < 0.01, n = 5). In contrast, calcium addition appeared to decrease fluorescence (enhance loss of fatty acid from the membrane) slightly more for the trans-parinarate (75 f 6%) than for cis-parinarate (51 f 1279, P < 0.05 (n = 5). Lastly, it should be noted that Ca2+ induced a transient increase in cis-parinarate fluorescence (Fig. 4 B) but not trans-parinarate fluorescence. These effects were fatty- acid-specific since they did not occur with 1,6-diphenyl-l,3,5- hexatriene, a hydrophobic fluorescence probe molecule with 110 ionizable groups (Fig. 5).
The corrected fluorescence of trans-parinaric and cis-pari- na rk acid binding to liver plasma membranes was increased 10% and 70%, respectively, by EGTA at physiological tem-
520
peratures while the loss of absorbance-corrected fluores- cence induced by Ca2+ was 75% and 51%, respectively (Table 1). Thus, the net effect would be a 65% loss and a 19% increase of fluorescence at 37°C for trans-parinaric and cix-parinaric acid, respectively. Similarly, the net effects at 24 "C were a 15 increase and 1 1 % decrease in trans-pari- naric and cis-parinaric acid by Ca2+ while at 1O"C there was a net 47 o/, increase and a 37 "/, decrease in corrected fluorescence, respectively. These results indicate that below physiological temperature Ca2 + should increase absorbance-corrected fluo- rescence of trans-parinarate while decreasing the absorbance- corrected fluorescence of cis-parinaric acid. Such alterations could be due to either differences in partitioning and/or in rigidity of the membrane microenvironment.
Biiiding Dynamics of trans-Parinaric and cis-Parinaric Acid to Liver P l a ~ m a Membranes: Ca2' E f f c t s
If CaZ+ prevents or dccreases parinaric fatty acid uptake by liver plasma membranes, then the absorption spectrum of the probe molecules should be shifted. As shown in Table 2, Ca2+ indeed shifted the absorption peak wavelength of frans- parinaric acid to slightly shorter wavelength while the peak
I I I ' I I I A cis- Dorinorote B
Time (mini
Fig.4. Effect of' LGTA unrl Ca" OII cis-parinuric acid binding in liver plusinn mernhra17e.r. Experimental conditions were as dcscribed in legend of Fig.2 except that cis-parinaric acid was used. (M) 10 ' C ; (0) 37 'C
wavelengths of the cis-parinaric acid were shifted to slightly longer wavelengths. Small, less than 1 x, changes in liver plasma membrane volume were associated with these shifts. More importantly, the K p values (Table 3) indicated that Ca" also decreased the binding of trans-parinaric acid but increased the binding of cis-parinaric acid to the rat liver plasma mem- branes, in agreement with the absorbance peak shift data. The fluorescence probe molecule 1,6-diphenyl-I ,3,5-hexa- triene was > 99% bound in all cases. These changes in K ,
0 20 10 60 Time (mn)
Fig. 5 . Effect of EGTA a i d Ca2+ on 1,6-diph~~i1~~l-1,3,5-hL.sarriene incor- poraretl into fivcr pfasmu membrane. Experimental conditions were as described in legend of Fig. 1 except that 1,6-diphenyl,l,3,5-hexatriene was used. (A) With buffer A containing CaZ+; (B) with buffcr B lacking CaZ+; (0) 37 " C ; (.) 10 'C
Table 1. The Cffeecf uf' culciurn and EGTA on , f luorr~st.ent-~?robe-co~~Kc~~(i fluorescence in fiver plasma membranes Effects are measured 30niin after addition of 3.4mM EGTA or 3.4mM CaZ+. All conditions were as described in legends of Fig. 3, 4, and 5 . Values represent the mean k SEM, it = 5
Additions Buffer Probe molecule Change in F,, at ~~ ~~ ~ -~
CaZ + EGTA IO'C 24 ' C 37 'C ~
~~ ~~
x - ~-
EGTA 2 4 0 tian$-parindrdte + 119 * I 9 + 80 f 10 + 1 0 * 2
3 4 0 1,6-diphenyl-1,3,5-hexatriene + 1 4 + 4 + 8 + 4 + 1 + 3
0 1 c zr-parniarate - 5 3 * 5 - 5 4 t 6 - 5 1 + 7
2 4 0 ru-parinarate + 1 3 A 6 + 4 3 * 8 + 7 0 + 5
Cd2 + 0 1 trans-parinarate - 7 4 i 6 - 6 5 + 5 - 7 5 + 6
0 1 1,6-d1phenyl-l,3,5-hexdtr1enc - 9 + 4 + 5 + 3 + 1 + 6
521
Table 2. Effect of' Cu2+ and EGTA on trans-parinuric und cis-parinuric acid ah.sorption spectrum derived parameters in rat liver plasma mem- branes il represents the longest wavelength absorption peak of the parinaric acid in liver plasma membranes (50 pg proteinjml; lipid to probe molar ratio of 66). The polarizability, a, was calculated from [?.,I-' as dcscribed in Materials and Methods. The refractive index, n , was calculated from a also or described in Materials and Methods while the ratio V ( t ) / V ( 2 0 ) = x(24)/x(t) and is the volume relative to its value at 20 'C assuming membrane birefringence can be neglected
CaZ+ EGTA
mM nm
trwwParinaric 2.4 0 317.0 0.255 1.42 1.01 0 1 318.5 0.271 1.45 1.00 3.4 1 317.5 0.262 1.44 1.01
c.ir-Parinaric 2.4 0 324.0 0.271 1.45 0.99 0 1 323.0 0.260 1.43 1.00 3.4 1 324.0 0.271 1.45 0.99
Table 3. Effect of Ca2+ and EGTA on bindinx oJprobr molecules to raf liver plusma menibrunes The partition coefficient (K,) and the bound to free ratio of tra~w-pari- nate (1.8 pM), cis-parinarate (2.5 pM) and 1,6-diphenyl-l,3,S-hexatriene (0.2 pM) in rat liver plasma membranes (50 pg proteinjml buffer, pH 7.2) were determined at 24 -C after IS min incubation at 37 ' C as described In Materials and Methods. Only bound probe molecules fluoresce since the quantum yield of these probes in aqueous bulf'er is negligibly small. Values represent the mean SEM (n = 3)
___ ~
Probe BuHer l o h x Bound'free KP ~~~
Cd2+ EGTA
n~ M -
tran.s-Parinaric 2.4 0 0.8 f 0.1 46/54 0 1 4.3 f 0.8 83/17 3.4 1 0.3 f 0.1 47/53
0 1 2.1 * 0.4 68/32 3.4 1 5.3 k 0.9 85/15
ch-Parinaric 2.4 0 14.6 k 1.8 9416
1 .G-Diphenyl- 2.4 0 9S k 5 99il 1,3,5-hexatriene 0 1 63 k 4 9911
3.4 1 90 f 10 9Y/l
values were consistent with the net absorbance-corrected fluorescence changes noted in the previous section.
It was also possible that the alterations in [bound/free] trans-parinarate, cis-parinarate, or 1,6-diphenyl-1,3,5-hexa- triene were due to the presence of insoluble probe-Ca2' com- plexes that were coscdimented with the liver plasma inem- branes during the partitioning determination. This possibility was tested by incubating the same concentration of probe molecules as used in Table 3 with each buffer system but in thc absence of liver plasma membranes. The absorbance of the samples was then determined before and after ultracentrif- ugation. No differences in absorbance were noted. This result indicated that insoluble complexes of probes with Ca2+ were not formed.
Efject of Ca2+ on Partitioning of Probes between Fluid and Solid Domains
The effect of CaZ + on the partitioning of the trans-parinaric and cis-parinaric acid in fluid and solid domains in liver plasma membranes indicated that the metal ion altered this partitioning (Table 4). Ca2+ did not abolish either of the two lipid domains in the liver plasma membranes as indicated by the presence of two fluorescence lifetime cotnponents for trans-parinarate and cis-parinarate. The size of the fluid domain, X I , was increased slightly by Ca". This change did not appear statistically significant. However, the fluorescence lifetime, t 2 , of both trans-parinaric and cis-parinaric acid in the solid domain was increased significantly by the addition of excess calcium. Consequently, Ca2+ also increased the total quantum yield, Qtotai. The fluorescence lifetime of trans- parinaric and ci.7-parinaric acid in the fluid domain, zl, was not significantly altered by Ca2 '. When these experiments were repeated at 37°C and at 10°C similar results were ob- tained, except of course that the zl and r2 were temperature dependent (increasing at lower temperatures). The increases in rI and 52 at lower temperatures correspond well with increases in absorbance-corrected fluorescence intensity. The agreement between these parameters indicates that changes in fluorescence quantum yield may be largely produced by fluorescence lifetime changes, assuming that the microvisco- sity of the liver membrane was not changed by Ca2+ . The latter possibility was tested in the following section.
It should be noted that the fluorescence lifetime, parti- tioning, and quantum yield of 1,6-diphenyl-l,3,5-hexatriene in the liver plasma membranes were not altered by Ca2+. These data indicate that effects of Ca2+ of fluorescence life- time and quantum yield were specific for the microenviron- ment of the fatty acid probe molecules.
Effects of' Cu2' untl EGTA on Polarization oj Fluorescent Probe Molecules in Livm Plusmi Men~brutzr.c
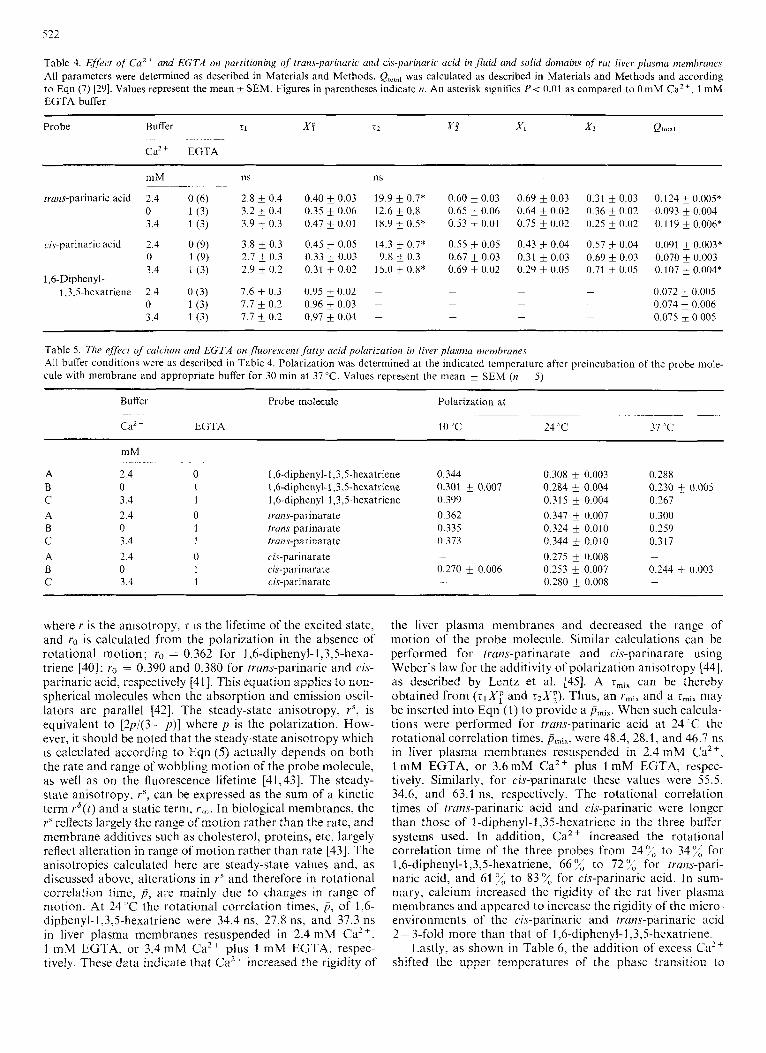
Calcium is known to make biological and model mcm- branes more rigid [36,37]. Table 5 shows the effects of Ca2+ on fluorescence polarization of I ,6-diphenyl-l,3,5-hexatriene at 10 "C, 24 "C, and 37 T. Calcium increased the polarization at 24' C from 0.284 k 0.004 to 0.308 k 0.003 ( P < 0.01, n = 5). This increase of 8 "/, was similar to increases in order parameter of spin resonance probes noted earlier [36,37]. Calcium also increased the polarization of 1,6-diphenyl- 1,3,5-hexatriene at 10°C and 37°C. Addition of excess Ca2+ (3.4 mM) to 1 mM EGTA buffer also increased the polari- zation of the probe molecule in liver plasma membranes. Basically similar trends were obtained with <vans-parinaric and cis-parinaric acid: excess Ca2 + increased polarization while excess EGTA decreased polarization (Table 5). trans- Parinarate in the liver plasma membranes generally had polarization values similar to those of the 1,6-diphenyl-l,3,5- hexatriene. In contrast, cis-parinarate, which partitions more into fluid lipid regions as compared to trans-parinarate, had the lowest polarization of all three probe molecules. Altera- tions in polarization can be converted to rotational correla- tion times, p , which can be calculated from the Perrin equa- tion [38,39] :
522
Table 4. Effect of Caz+ and EGTA on partitioning of trans-parinuric and cis-parinuric acid in fluid and solid domains of rat liver plasma memhranes All parameters were determined as described in Materials and Methods. Q,,,, was calculated as described in Materials and Methods and according to Eqn (7) [29]. Values represent the mean f SEM. Figures in parcnthcscs indicate n. An asterisk signifies P < 0.01 as compared to 0mM Ca2+, 1 mM EGTA buffer
Probe Buffer TI XP T 2 XP XI xz Q m a ~ . ~~ ~~ ~
Ca2 + EGTA
mM ns ns -~
trans-parinaric acid 2.4 0 (6) 2.8 f 0.4 0.40 * 0.03 19.9 * 0.7* 0.60 f 0.03 0.69 f 0.03 0.31 f 0.03 0.124 0.005* 0 l ( 3 j 3.2 0.4 0.35 k 0.06 12.6 f 0.8 0.65 0.06 0.64 k 0.02 0.36 f 0.02 0.093 f 0.004 3.4 l ( 3 ) 3.9 f 0.3 0.47 k 0.01 18.9 * 0.5* 0.53 f 0.01 0.75 k 0.02 0.25 f 0.02 0.119 f 0.006*
cis-parinaric acid 2.4 0 (9j 3.8 * 0.3 0.45 k 0.05 14.3 f 0.7* 0.55 f 0.05 0.43 k 0.04 0.57 k 0.04 0.091 & 0.003* 0 1 (9) 2.7 k 0.3 0.33 k 0.03 9.8 * 0.3 0.67 f 0.03 0.31 f 0.03 0.69 f 0.03 0.070 f 0.003 3.4 1 (3) 2.9 0.2 0.31 f 0.02 15.0 0.8* 0.69 k 0.02 0.29 f 0.05 0.71 f 0.05 0.107 & 0.004*
1,h-Diphenyl- 1,3,5-hexatriene 2.4 0 (3) 7.6 f 0.3 0.95 f 0.02 - - - - 0.072 & 0.005
0 l ( 3 ) 7.7 f 0.2 0.96 f 0.03 - ~ - - 0.074 * 0.006 3.4 1(3 j 7.7 0.2 0.97 & 0.04 - ~ - - 0.075 t_ 0.005
Table 5 . The eflect of calcium and EGTA on ,fluorescent fatty acid polarization in liver plasma membranes All buffer conditions were as described in Table 4. Polarization was determined at the indicated temperature after preincubation of the probe mole- cule with membrane and appropriate buffer for 30 min at 37°C. Values represent the mean * SEM (n = 5 )
Buffer Probe molecule Polarization at ~
~ ~
CaZ + EGTA 10 'C 24 "C 37 "C
mM
2 4 0 3 4 2 4 0 3 4 2 4 0 1 4
1 ,G-diphenyl- 1,3,5-hexatr-iene 1,6-diphenyl-l,3,5-hexatriene 1,6-diphenyl-l,3,5-hexatriene trans-parinarate trans-parinarate irans-pa rinarate cis-parinarate cis-parinarate cis-parinarate
0.344 0.301 f 0.007 0.399 0.362 0.335 0.373
0.270 f 0.006 -
__
0.308 f 0.003 0.284 i 0.004 0.315 k 0.004 0.347 f 0.007 0.324 f 0.010 0.344 * 0.010 0.275 f 0.008 0.253 f 0.007 0.280 k 0.008
0.288 0.230 & 0.005 0.267 0.300 0.259 0.317
~
0.244 i 0.003 -
where I' is the anisotropy, z is the lifetime of the excited state, and ro is calculated from the polarization in the absence of rotational motion ; r~ = 0.362 for 1,6-diphenyl-l,3,5-hexa- triene [40]; YO = 0.390 and 0.380 for trans-parinaric and cis- parinaric acid, respectively [42]. This equation applies to non- spherical molecules when the absorption and emission oscil- lators are parallel [42]. The steady-state anisotropy, r s , is equivalent to [2p/(3 -p)] where p is the polarization. How- ever, it should be noted that the steady-state anisotropy which is calculated according to Eqn ( 5 ) actually depends on both the rate and range of wobbling motion of the probe molecule, as well as on the fluorescence lifetime [41,43]. The steady- state anisotropy, r s , can be expressed as the sum of a kinetic term r b ( t ) and a static term, r W . In biological membranes, the rs reflects largely the range of motion rather than the rate, and membrane additives such as cholesterol, proteins, etc, largely reflect alteration in range of motion rather than rate [43]. The anisotropies calculated here are steady-state values and, as discussed above, alterations in r s and therefore in rotational correlation time, p, are mainly due to changes in range of motion. At 24'C the rotational correlation times, p, of 1,6- diphenyl-I ,3,5-hexatriene were 34.4 ns, 27.8 ns, and 37.3 ns in liver plasma membranes resuspended in 2.4 mM Ca2+, 1 mM EGTA, or 3,4 mM Ca2+ plus 1 mM EGTA, respec- tively. These data indicate that Ca2+ increased the rigidity of
the liver plasma membranes and decreased the 1-ange of motion of the probe molecule. Similar calculations can be performed for tram-parinarate and cis-parinarate using Weber's law for the additivity of polarization anisotropy [44], as described by Lentz et al. [45]. A zmix can be thereby obtained from (zl Xf' and rzX;) . Thus, an rmix and a smiX may be inserted into Eqn (1) to provide a Pmir. When such calcula- tions were performed for trans-parinaric acid at 24 'C the rotational correlation times, Pmix, were 48.4, 28.1, and 46.7 ns in liver plasma membranes resuspended in 2.4 mM Ca2+, 1 mM EGTA, or 3.6 mM Ca2+ plus I mM EGTA, respec- tively. Similarly, for cis-parinarate these values were 55.5, 34.6, and 63.1 ns, respectively. The rotational correlation times of trans-parinaric acid and cis-parinaric were longer than those of l-diphenyl-1,35-hexatriene in the three buffer systems used. In addition, CaZ + increased the rotational correlation time of the three probes from 241,; to 347; for 1,6-diphenyl-l,3,5hexatriene, 66 o/, to 72 o/, for trans-pari- naric acid, and 61 for cis-parinaric acid. In sum- mary, calcium increased the rigidity of the rat liver plasma membranes and appeared to increase the rigidity of the micro- environments of the cis-parinaric and trans-parinaric acid 2- 3-fold more than that of 1,6-diphenyl-I ,3,5-hexatriene.
Lastly, as shown in Table 6, the addition of excess Ca2+ shifted the upper temperatures of the phase transition to
to 83

Table 6. Effect of Cn2 on churucteristic temperuture of l,6-diphenyl- 1 . 3 , 5 - 1 1 e x a ~ ~ i ~ ~ n c ~ , f l ~ ~ ~ ~ ~ ~ . ~ ~ e ~ i ~ e in rut liver plrrsmcr memhrunes Characteristic temperatures were determined from Arrhenius plots as desctibed in the legend to Fig.2 [ZO]
Buffer Characteristic temperature
CaZ + EGTA . .-
mM "C
2.4 0 21 19 0 1 22 18 3 4 1 30 17
. ~~ -
higher temperature. These data are also consistent with CaZ +
rigidifying the rat liver plasma membrane.
DISCUSSION
Mechanism of trans-Parinarate and cis-Parinarate Uptake
Fatty acid uptake by cells may occur by a receptor- mediated [3 - 71 or a passive diffusion mediated [4,8,9] pro- cess. Abumrad [4] suggested that at low unbound fatty acid concentrations (less than 0.06pM) the receptor-mediated pathway of fatty acid uptake dominated and was half- saturated, while at 2.5 pM free fatty acid both receptor- mediated and diffusion-mediated uptake occurred to an equal extent. In the present investigation, the free fatty acid concen- tration was varied from 0.01 - 10 pM and binding was evaluated at equilibrium conditions. In contrast, Abumrad et al. [4] measured only initial rates of binding (the first 15 s) of radioactive fatty acid. In general, however, the concen- trations of fluorescent fatty acids used herein were high enough such that at equilibrium the receptor-mediated mech- anism was saturated and the diffusion-mediated mechanism accounted for the remainder of the uptake. In addition, others have shown that unsaturated fatty acids are taken up at slower rates than saturated fatty acid by Ehrlich ascites cells [lo]. Similarly, we have obtained K,, Kd, and bound/free values for truns-parinaric and clr-parinaric acid in rat liver plasma membranes consistent with this relationship. Thus, the all-trans-parinaric acid resembles a saturated fatty acid in its binding while the cis-parinaric acid (cis, truns, trans-, cis-) resembled cis unsaturated fatty acid in its uptake.
Culcium Alters Binding of trans-Parinaric and cis-Parinaric Acid to Rut Liver Plasma Mernhrunt?s
Consistent with the possibility that Ca2+ alters fatty acid binding by making the solid domains of the rat liver plasma membrane more rigid is phospholipid compositional data indicating that significant quantities of negatively charged phospholipid which Ca2+ can bind are present in liver plasma membranes : 8.5 phosphatidylinositol, 4.0 "/, phosphatidyl- serine and 0.4% phosphatidic acid [46]. Addition of Ca2+ to phosphatidylserine-containing micelles or membranes in- creased the rigidity of the membranes [36, 37, 471 altered the conformation of the lipid polar head groups [47], and shifted the location of fluorescent probe molecules within the mem- branes 1471. Although modulation of fatty acid binding could also be achicved by interaction of Ca2+ with membrane-bound proteins, the majority of binding sites would be expected to be negatively charged lipids.
The results presented herein and elsewhere [48,,49] indi- cate that calcium does not modulate fatty acid uptake by forming insoluble complexes with the free acid. Hexane extracts calcium-fatly acid complexes in two phase partition systcins of hexane and aqueous buffer [48,49]. Ca2+ may also interact with cis-parinaric acid in membranes [24,27]. These rcsults would imply that Ca2+ cannot extract the fatty acid from a hydrophobic environment (hexane or a membrane). Other investigations have shown that 4sCa2+ binding to fatty acid monolayers or fatty acid in single or mixed monolayers of dipalmitoylglycerophosphocholine or dioleoylglycerophos- phocholine is insignificant at pH 7.0 due to a dramatic shift in pK of the fatty acid carboxyl group (from 4 or 5 to 10) [SO]. Thus, it seems more likely as shown herein that calcium will not extract fatty acid from membranes as a precipitable com- plex but may reduce the binding of the fatty acid by some other means, possibly by increasing rigidity of the membrane and thereby altering the partitioning of the fatty acid between the membrane and aqueous interiors, and/or also by altering the partitioning between fluid and solid domains of the liver plasma membrane. In the latter cases the parinarate fatty acids would become less fluorescent.
The increase in fluorescence of cis-parinarate fluorescence a short time after 3.4 mM Ca2+ addition noted herein is con- sistent with the view that the calcium rigidifies the liver plasma membrane thcreby giving rise to a higher quantum yield of cis-parinarate fluorescence. Shortly thereafter, the cis-pari- narate may be removed from the membrane resulting in an overall fluorescence decrease after 30 min. Livingston and Schachter 151 ] demonstrated that Ca2+ modulated lipid dynamics in the rat hepatocyte membrane by direct and indirect mechanisms. The direct mechanism was rapid with a half-time of 10- 15 niin at 3 7 T , was proportional to Ca2+ in the range of 0- 4 mM, and was readily reversible by addi- tion of excess EGTA. This mechanism accounted for the increased rigidity of the hepatocyte membrane and appears very similar to the mechanism described herein for Ca2+ effects on rat liver plasma membrane structure. The second mechanism was indirect, required Ca2+, was slow with a half-time of 75 min, plateaued, and could not be reversed upon chelation of Ca2+ by EDTA. The latter mechanism was due to Ca2+ activation of phospholipase A activity and re- sulted in altered membrane fluidity that was not reversible by EDTA. The latter mechanism was not investigated in the present work.
Significance
Free fatty acids are rapidly taken up by cells and either incorporated into glycerolipids or oxidized. However, a small amount remains in unesterified form in a pool which has a high turnover rate [S2- 541. Most of this free fatty acid (about 70%) is easily removed by albumin and appears to be asso- ciatcd with the plasma membrane 12,531. Similar results have been obtained with free trans-parinaric acid 1481. The pres- ence of significant amounts of free fatty acids in membranes affects membrane fluidity 1101 as well as alters biological effects including aggregation of platelets [55], activity of adenylate cyclase [56], cheniotaxis [57], and phagocytosis [lo, 571. Thus, free fatty acids have an important role in bio- logical processes as well as cellular metabolism. Ca2+ and hormones that regulate calcium binding to membrane [I I , 12, 14,16,51] influence fatty acid metabolism [16-191. These effects may be mediated through the intracellular redistri- bution of Ca2+. As shown herein, such a redistribution may

alter the fatty acid binding to the plasma membranes of liver cells. chemistry, 16, 813-819.
Association, Missouri Affiliate, and the USPHS (GM 31651 and CA 24339). The authors wishes to especially thank Dr Murray Heimberg for critical reading of the manuscript and stimulating discussions.
26. Sklar, L. A., Hudson, B. S., Petersen, M. & Diamond, J . (1977) Bio-
This work was supported in part, by from the ~~~~i~~~ H~~~~ 27. Sklar, L. A., Miljanich, G. P. & Drdtz, E. A. (1979) Biocl~emisrr.~, 18, 1707 - 1716.
28. Pohl, s. L. (1976) in Methods in Recepptor Research (Blecher, M., ed.) pp. 159-174, Marcel Dekker Inc., New York.
29. Schroeder. F. (1983) Eur. J . Biochem. 132. 509 - 516.
REFERENCES 1. 2.
3.
4.
5.
6 .
7.
8.
9.
10.
1 1 .
12. 13. 14.
15. 16.
17 18 19
20 21
22
23
Goodman, D. D. (1958) J . Clin. Invest. 37, 1729-1735. Spector, A. A., Stcinberg, D. & Tanaka, A. (1967) J . Biol. Chem.
Weisiger, R., Gollan, J. & Ockner, R. (1981) Science (Wask. DC)
Abumrad, N. A,, Perkins. R. C., Park, J. H. & Park, C. R. (1981)
Mahadevan, S. & Sauer, F. (1974) Arch. Biochem. Biophys. 164,
Samuel, D., Paris, S. & Ailhaud, G. (1976) Eur. J . Biochem. 64,
Paris, S., Samuel, D., Romey, G. & Ailhaud, G. (1979). Biochin~ie (Paris) 61, 361 -361.
DeGrella, R. F. & Light, R. J . (1980) J . B i d . Chem. 255, 9731- 9748.
DeGrella, R. F. & Light, R. J. (1980) J . Biol. Chem. 255, 9739- 9745.
Spector, A. A. & Fletcher, J . E. (1978) in Disturbances in Lipid and Lipoprotein Metabolism (Dietschy, J. M . , Gotto, A. M. & Ontko, J . A., eds) pp. 229-249, Williams and Wilkins Co., Baltimore, MD.
Livingtone, C. J. & Schater, D. (1980) J . Biol. C'hem. 255, 10902- 10908.
Luly, P. & Shinitzky, M. (1979) Biochemistry, 18, 445-450. Schroeder, F. (1982) J . Membr. Bid . 68, 1 - 10. Shlatz, L. & Marinetti, G. V. (1972) Science (Wash. DC) 176,
Kiss, Z. (1979) Eur. J . Biochem. 95, 607-611. Andia-Waltenbaugh, A. M., Kimura, S., Wood, J., Divakaran, P.
Tou, J . S. (1979) Biochim. Biophys. Acta, 572, 307-313. Otto, D. A. & Ontko, J. A. (1978) J . Biol. Chem. 253, 789-799. Cook, G. A., Sullivan, A. C. & Ontko, J. A. (1977) Arch. Biochem.
Schroeder, F. & Goh, E. H. (1980) Chem. Phys. Lipids, 26,207-224. Schroeder, F., Holland, J. F. & Vagelos, P. R. (1976) J . B id . Chem.
Schroeder, F., Holland, J. F. & Vagelos, P. R. (1976) J . Bid. C'hem.
240, 3 032 - 1041.
211,1048-1050.
J . Biol. Chem. 256, 9183-9191.
185 - 193.
583 - 595.
175 - 177.
& Friedman, N. (1978) Life Sci. 23, 2437-2444.
BiophJJs. 179,310-321.
251, 6739.
251. 6747-6756.
j i
30. Lowry,O. H., Rosebrough, N. J., Farr,A. L. & Randall, R. J.(1971)
31. Holland, J. F., Teets, R. E. & Timmick, A. (1973) Anal. Chem. 45,
32. Holland, J . F., Teets, R. E., Sindmack, G. & Timmick, A. (1977)
33. Christman, D. R., Crouch, S. R., Holland, J . F. & Timmick, A.
34. Reference deleted. 35. Reference deleted. 36. Sauerheber, R. D., Zimmerinan, T. S., Esgate, J. A., VanderLoan,
37. Cullis, P. R. & Verkleij, A. .I. (1979) Biochim. Biophys. Acta, 552,
38. Weber, G. (1973) Adv. Protein Chem. 8, 415-4460, 39. Shinitzky, M. & Inbar, M. (1974) J . Mol. Biol. 85, 603-615. 40. Shinitzky, M., Dianous, A.-C. Gitler, C. & Weber, G. (1971) Bio-
41. Wolber, P. K . & Hudson, B. S. (1981) Biochemistry, 20, 2800-
42. Gilmore, R., Cohn, N. & Glaser, M. (1979) Biochemistry, 18, 1042-
43. Kinositd, K., Kataoka, R., Kimura, Y., Gotoh, 0. & Ikegami, A.
44. Weber, G. (1952) Biochem. J . 51, 145-167. 45. Lentz, B. R., Barenholz, Y. & Thompson, T. E. (1976) Biochmzistry,
46. Ray, T. K., Skipski, V. P., Barclay, M., Essner, E. & Archibald, F.
47. Harris, W. E. (1977) Chem. Phys. Lipids, 19, 243-254 48. Moskal, J . R., Emaus, R. K. & Holland, J. F. (1977) Fed. Proc. 37,
49. Hauser, H., Guyer, W. & Howell, K. (1979) Biochemisrr.v, 18, 3285-
50. Blaustein, M. (1974) Rev. Phys. Biochem. Pharmacol. 70, 33 - 82. 51. Livingstone, C. J. & Schdchter, D. (1980) Biochemistry, 19, 4823-
52. Spector, A. A. & Steinberg, D. (1965) J . Biol. Chem. 240, 3747-
53. Spector, A. A. (1971) frog. Biochem. Pharmacol. 6 , 130-176. 54. Hoak, J. C., Spector, A. A.. Fry, G. L. & Warner, E. D. (1970)
J . Bid . Chem. 193, 267-275.
145-153.
Anal. Cl7em. 49, 706-717.
(1 980) Anal. Chem. 52, 29 1 - 295.
W. P. & Gordon, L. M. (1980) J . Membr. B i d . 52, 201-219.
546 - 551.
chemistry, 10, 2106-21 13.
2810.
1071.
(1981) Biochemistry, 20, 4270- 4277.
15, 4529 - 4537.
M. (1969) J . Biol. Chem. 244, 5528-5536.
1597, a1805.
3291.
4827.
3753.
Nature (Lond.) 228, 1330-1332. Schroeder. F. (1978) Biochim. Biouhvs. Ac fa , 511, 356- 376. 55. Fain, J. N. & Sheppherd, R. E. (1975) J . B i d . Chem. 250, 6586-
~I ._ 24. Sklar, L. A,, Hudson, B. S. & Simoni, R. D. (1977) J . Biol. Chem. 6592.
3433- 3437. 16, 5100-5108. 56. Orly, J. & Schramm, M. (1975) Proc. Nut1 Acad. Sci. USA, 72,
819- 82X. 25. Sklar, L. A., Hudson, B. & Simoni, K. D. (1977) Biochemistry, 16,
57. Hawley, H. P. & Gordon, G. B. (1976) Lab. Invest. 34, 216-222.
F. Schroeder*, Department of Pharmacology, School of Medicine, University of Missouri, Columbia, Missouri, USA 65201
* To whom requests for reprints should be made.