by and (from the department of chemistry, ames) · pdf file · 2003-02-26importance...
TRANSCRIPT

NEOPYRITHIAMINE AND THE THIAMINASE OF FISH TISSIJF,S*
BY ROBERT RIDGELY SEALOCK AND HILDA S. WHITE
(From the Department of Chemistry, Iowa State Colleye, Ames)
(Received for publication, June 20, 1949)
The enzyme of the visceral tissues of carp, mullet, and other fishes which destroys thiamine by cleaving the pyrimidylmethylene group from the thiazole portion may be either inhibited or activated by appropriate analogues of the vitamin. Sealock and Goodland (I) have shown that 3-(o-aminobenzyl)-4-methylthiazolium chloride specifically and competi- tively inhibits the enzymatic destruction of thiamine. On the other hand, when the amino group is present in the meta instead of the ortho position, the corresponding compound, 3-(m-aminobenzyl)-4-methylthiazolium chlo- ride fails to inhibit the enzyme. Instead activation is observed (2). Incubation of the enzyme and thiamine with this analogue results in very significantly increased destruction of the thiamine. As a result of these findings it has been suggested (2) that the position of the amino group relative to the methylene attachment to the thiazole ring is of utmost importance to the process of inhibition. In fact it would appear that both the inhibitor and the substrate combine with the enzyme by means of their respective amino groups (1, 2). This suggestion became even more valid when it was found that the activation by the m-amino compound or a substitute for it, m-nitroaniline (or m-aminobenzoic acid), resulted from an entirely different mechanism (3). These latter compounds produce their activation by serving as “reactors” during the enzymatic destruction of thiamine, being converted into secondary amino derivatives with the pyrimidylmethylene residue.
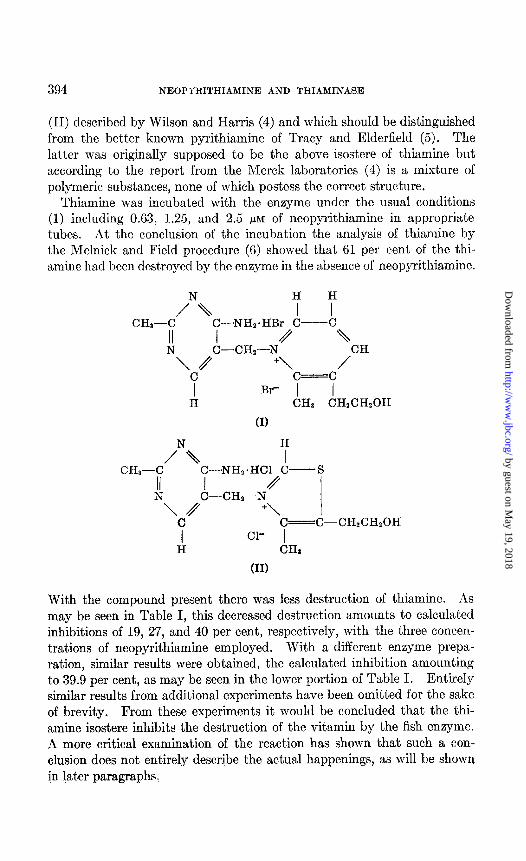
That the amino group is not the only requisite to an effective inhibitor may also be concluded, since o-aminobenzyl alcohol does not inhibit the enzyme and the aminopyrimidylmethylenesulfonic acid derived from thi- amine is a very poor inhibitor (1). It is quite conceivable that the latter compound, if present in the form of an aminopyrimidylmethylene quater- nary salt rather than a sulfonic acid, would then become a more effective inhibitor. Such a compound is available in neopyrithiamine or 1-((4- amino - 2 - methyl) - 5 - pyrimidylmethyl) - 2 - methyl - 3 - (p - hydroxy- ethyl)-pyridinium bromide hydrobromide (I), the isostere of thiamine
* The authors arc indebted to Dr. Karl Folkers, Director of Organic and Bio- chemical Research of the Research and Development Division of Merck and Com- pany, Inc., for making available a supply of neopyrithiamine and to the Williams- Waterman Fund of the Research Corporation for a grant-in-aid in support of this investigation.
393
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
394 NEOPYRITHIAMINE AND THIAMINASE
(II) described by Wilson and Harris (4) and which should be distinguished from the better known pyrithiamine of Tracy and Elderfield (5). The latter was originally supposed to be the above isostere of thiamine but according to the report from the Merck laboratories (4) is a mixture of polymeric substances, none of which postess the correct structure.
Thiamine was incubated with the enzyme under the usual conditions (1) including 0.63, 1.25, and 2.5 pM of neopyrithiamine in appropriate tubes. At the conclusion of the incubation the analysis of thiamine by the Melnick and Field procedure (6) showed that 61 per cent of the thi- amine had been destroyed by the enzyme in the absence of neopyrithiamine.
CH*mc/N\ C-NH$*HBr C---C
II I
N\ // C-CH%--N
/ XCH
+\ /
;:
- Br-
H H, H,,,OH a a 2
(I)
/“\ Ii I
CHa-c C-NH% .HCI C- 8 II I //
N\ // C-CHs-N
$: “\C =C-CH#HzOH
Cl- 1 H CHa
(II)
With the compound present there was less destruction of thiamine. As may be seen in Table I, this decreased destruction amounts to calculated inhibitions of 19, 27, and 40 per cent, respectively, with the three concen- trations of neopyrithiamine employed. With a different enzyme prepa- ration, similar results were obtained, the calculated inhibition amounting to 39.9 per cent, as may be seen in the lower portion of Table I. Entirely similar results from additional experiments have been omitted for the sake of brevity. From these experiments it would be concluded that the thi- amine isostere inhibits the destruction of the vitamin by the fish enzyme. A more critical examination of the reaction has shown that such a con- clusion does not entirely describe the actual happenings, as will be shown in later paragraphs,
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

R. R. SEALOCK AND II. S. WHITE 395
In order to use the Melnick and Field method of analysis in the above experiment, it was of course necessary to show that neopyrithiamine neither interfered nor reacted to give xylene-extractable chromogenic material. Under the same conditions employed for thiamine analysis, neopyrithi- amine gave no color value even when the reagent was allowed to react for 0.5, 6, and 24 hours before adding the xylene. At the same time, the pres- ence of neopyrithiamine had no effect on the thiamine color value.
It is of interest to point out that this failure of neopyrithiamine to react in the Melnick and Field method furnishes strong evidence relative to the nature of the reaction obtained with diazotized p-aminoacetophenone and thiamine. The presence of the alkali in the reagent causes the opening of the thiazole ring and formation of the sulfhydryl group, as is well known
Apparent Inhibition of ‘I’h.ia?ninase with L%Teop?jrithiamine
The incubations were carried out in 5 ml. of 0.08 AI phosphate buffer ant1 4 per cent sodium chloride at pEI 7.4 and 37.5’ for 2 hours. The enzyme was the soluble portion from the amount and preparation indicated. 2.5 PM of thiamine (5 X 1OP mole per liter) were present in each tube.
Enzyme Dl-215,40 mg. equiva- lents per tube
Enzyme S3-110, 60 mg. cquiva- lents per tube
-.-
Neopyrithiamine Thiamine destroyed
moles per 1. x 10‘ NM
0 1.58 1.25 1.22 2.50 1.11 5.00 0.92 0 0.98 5.0 0.59
-
-
Inhibition
pm ccnl
10.1 26.5 39.6
39.9
(7). If we assume that the Melnick and Field reaction involves coupling of the diazo reagent with the sulfhydryl group in order to form the colored (xylene-soluble) complex, it is obvious that neopyrithiamine, which neither contains sulfur nor can undergo analogous pyridine ring opening, should not react in this method. While considerable evidence has accumulated in this laboratory that this is the mechanism of the reaction, a more complete description will be presented later.
The lack of interference of neopyrithiamine in the analytical method has permitted the calculation of apparent inhibition in the experiment recorded in Table I. However, the similarity in structure of the com- pounds is such that the enzyme might be expected to produce the same cleavage of neopyrithiamine that it does of thiamine. In order to test this possibility it is necessary to have a method of analysis for the former. The possibility that it would undergo the alkaline oxidation and subse-
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

396 NEOPYRITHIAMINE AND THIAMINASE
quent production of fluorescence as does thiamine in the thiochrome procedure (8) was therefore tested. Solutions of neopyrithiamine, such as were used in the enzyme experiments, containing 2.5 PM in 10 ml. of 0.04 M phosphate buffer and 5 per cent trichloroacetic acid, were diluted with water. 5 ml. of a 1:250 dilution treated with 3 ml. of alkaline ferri- cyanide and read directly without butyl alcohol extraction gave gal- vanometer readings of 11 (corrected for the blank) which correspond to the readings obtained with approximately one-fifth the quantity of thi- amine. With dilutions of 1:50, scale readings of 75 were obtained and it became obvious that the new compound could be determined by means of
UJ 3L 3
100
2 4 6 8 I 2 3 TIME, MINS, TIME, HRS.
FIG. 1 FIG. 2
FIG. 1. The development of fluorescence with neopyrithiamine in the thiamine- thiochrome procedure. 0.025 PM of neopyrithiamine in 5 ml. of solution was treated with alkaline ferricyanide and the readings obtained directly with the reaction mix- ture.
FIG. 2. The destruction of neopyrithiamine by carp thiaminase measured at differ- ent time intervals. 2.5 pM of neopyrithiamine in 5 ml. of reaction mixture were in- cubated with the enzyme in the usual fashion, the soluble portion from 50 mg. of acetone-desiccated powder being used in each 5 ml.
the thiochrome procedure. At concentrations of 0 to 0.02 PM per 5 ml. of aliquot analyzed the galvanometer readings were found to be proportional to the concentration. With this method 5.70 moles of neopyrithiamine yield fluorescence equivalent to 1 mole of thiamine. All readings have been taken at 90 seconds after addition of the reagent, although more recently it has been found that maximum fluorescence is not developed in this length of time. Analysis of this factor has led to the results in Fig. 1 which shows that the maximum value is obtained at approximately 4 minutes. The curve likewise shows that the 90 second interval used in these experiments is beyond the time of rapid reaction, so that probably no great error was introduced. The fluorescence from neopyrithiamine does not appear in the absence of ferricyanide (analytical control) and
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
R. R. SEALOCK AND H. S. WHITE 397
may be extracted with isobutanol as in the usual thiochrome procedure. However, this latter step was not necessary in these experiments, because of the relatively high concentration of compound employed. Neopyrithi- amine may also be removed from solution by Decalso, as employed in the more complete thiochrome procedure, and elution may be accomplished in the usual fashion, although this likewise proved unnecessary in the experiments described here.
With the possibility of determining neopyrithiamine established, this compound was incubated with the enzyme under the same conditions employed with thiamine. In each tube the soluble material from 40 mg.
TABLE II Destruction of Neopyrithiamine by Thiaminase
‘Ihe incubations were carried out as described in Table I except for the modifica- tions indicated and for the substitution of neopgrithiamine in the amounts indicated for thiamine.
ExpsPt
I
II
-
--
Enzyme present
None ‘I
Dl-215, 40 mg. equivalents Dl-215, 40 “ “ Dl-215, 40 “ “ Dl-215, 40 “ equivalents Dl-215, 40 “ “ Same, boiled
I‘ “
-
1
Neopyrithiamine
Concentra- tion
nozes per 1. x IO’ 2.50 5.00 1.25 2.50 5.00 2.5 5.0 2.5 5.0
-- Destruction
Qcr ccn:
0.02 1.6 0.03 1.2 0.60 96.2 1.20 96.0 1.44 58.0 1.25 100.0 1.52 60.8 0.04 3.2 0.14 5.7
of acetone-desiccated powder was used as the source of the enzyme. Neo- pyrithiamine at the levels of 0.63, 1.25, and 2.5 PM was added to appropri- ate tubes. At the end of the 2 hour incubation period, the neopyrithiamine remaining was determined. At the two lower levels of compound almost complete destruction occurred, as may be seen in Table II. At the higher level only 57.6 per cent was destroyed. Thus the reaction in this respect is similar to the thiamine reaction. As also may be seen in Table II incu- bation of the compound in the buffer solution alone caused no appreciable destruction. In the second experiment shown in Table II incubation of 1.25 and 2.5 PM of neopyrithiamine with the same amount of enzyme prepa- ration resulted in 100 and 60.8 per cent destruction respectively. In contrast, incubation with an equivalent amount of previously boiled enzyme extract produced essentially no destruction of the added neo-
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

398 SEOPYRITHIAMINE AND THIAMISBSE
pyrithiamine. Additional experiments have yielded similar results; how- ever, the enzymatic nature of the reaction is even more evident in the graph of neopyrithiamine destruction with time, shown in Fig. 2. The curve obtained is typical of enzyme reactions and in fact resembles closely the analogous curve obtained with thiamine (9).
With these results it may then be concluded that neopyrithiamine suffi- ciently resembles thiamine in structure so that it too can undergo destruc- tive reaction by the fish enzyme. It also may be concluded that the ap- parent inhibition recorded in Table I is not inhibition but instead is a case of enzymatic action in the presence of competitive substrates. Each of the substrates when present in the same enzyme solution is competing with the other for the enzyme. Confirmation of this conclusion has been obtained in experiments in which the enzymatic destruction of each substrate alone has been compared with that obtained when the two are present in the same solution. In such an experiment thiamine destruction is deter- mined by the Melnick and Field procedure, whereas the total neopyrithi- amine and thiamine destruction is measured by the thiochrome method. By deducting the former value from the latter the amount of neopyrithi- amine destroyed was then calculated. When 2.5 PM of each compound were employed with the particular enzyme preparation used, analytical difficulties were encountered due to the unfavorable ratio in chromogenic value in the fluorescence method. Consequently a precise evaluation of the neopyrithiamine destruction was difficult. By employing 4 times as much neopyrithiamine as thiamine a more favorable ratio was obtained for analytical purposes. With 5.0 pM of the former and 1.25 pM of the latter it was found that simultaneous destruction of each compound occurred, as may be seen in Table III.
It should be pointed out also that neopyrithiamine is only the second substrate, even though an artificial one, to be discovered for this enzyme. We, of course, may discount cocarboxylase, even though it is also destroyed, since it is merely the pyrophosphate ester of the original substrate. None of the other thiamine analogues which have been studied have been found to undergo the enzymatic reaction, although some of them are excellent competitive inhibitors.
The results obtained in this study permit at least tentative conclusions relative to one aspect of the mechanism of the thiaminase action. Since it is almost certain that neopyrithiamine is incapable of undergoing the ring opening characteristic of the thiamine-thiazole ring in alkaline solution, it may be argued that the enzymatic destruction of neopyrithiamine occurred with the molecule in the form indicated on a previous page. If the enzyme is able to produce its characteristic reaction with neopyrithi- amine, which is incapable of ring opening, then one may tentatively con-
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

R. R. SEALOCR AKD H. S. WHITE 399
elude that the enzyme reaction in the case of thiamine is entirely analogous. Or in other words, we would suggest that the thiaminase action involves a cleavage of the linkage between the two ring units of thiamine when the thiazole moiety is present in the intact and closed ring form. This con- clusion is also supported by the fact that the intact thiazole unit can be recovered from the enzymatic reaction as a product of the reaction.
If the conclusions at this point are correct, then it would be predicted that the compounds which specifically inhibit the destruction of thiamine by the fish thiaminase would also inhibit the destruction of neopyrithiamine. Likewise, compounds producing activation of thiamine destruction would have a similar effect upon neopyrithiamine destruction. Preliminary ex-
TABLE III
Simultaneous Destruction of Thiamine and Neopyrithiamine
The incubations were carried out as described in Table I except for the modifica- tions indicated. The enzyme preparation in each tube represented the soluble por- tion from 33 mg. of Enzyme 53-138. Thiamine was determined by the Melnick-Field procedure and neopyrithiamine by subtracting the Melnick-Field value from t,he total fluorophotometric values.
Compound present
Single substrates Thiamine. Neopyrithiamine.
Combined substrates Thiamine................. Neopyrithiamine.
- Concentration
2.5 10.0
2.5 10.0
Compound destroyed
P.M per Ccnl
0.792 63.3 0.860 17.2
0.190 15.2 0.870 17.1
periments in this direction entirely confirmed both predictions. ,&Amino- ethyl-4-methylthiazolium chloride, previously shown to be an excellent inhibitor of this enzyme (I), was found to inhibit neopyrithiamine destruc- tion, and m-nitroaniline (3) caused an increased enzymatic destruction of neopyrithiamine. Thus additional evidence for identity of the enzyme producing destruction in the case of the two substrates is obtained.
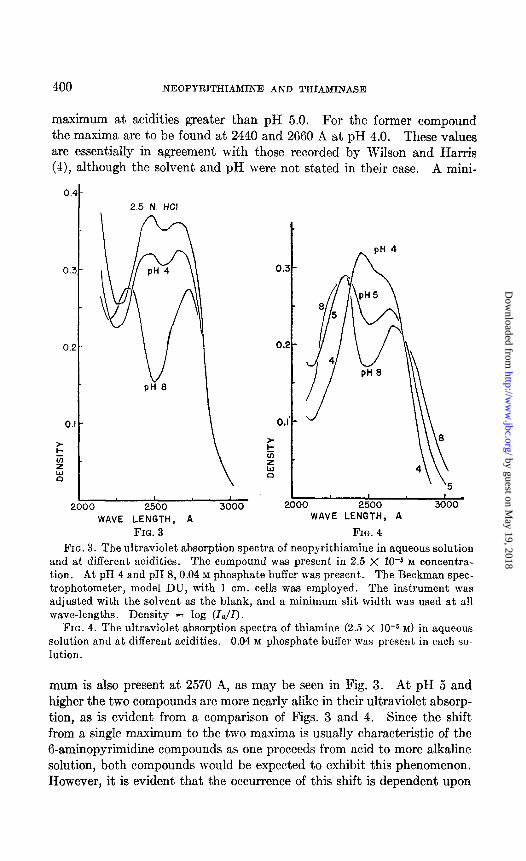
In conjunction with the analyses made in the enzymatic experiments, the ultraviolet absorption spectra of neopyrithiamine were studied. Even though it is a 6-aminopyrimidine derivative and accordingly absorbs ultraviolet light as thiamine, there are sufficient differences so that each of the two compounds may be quantitatively determined in the same solu- tion. However, it is necessary to maintain control of the PH. For ex- ample, in 2.5 N hydrochloric acid and at pH 0.45 and 1.25, neopyrithi- amine exhibits two maxima, whereas thiamine exhibits only a single
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
400 NEOPYRITHIAMINE AND THIAMINASE
maximum at acidities greater than pH 5.0. For the former compound the maxima are to be found at 2440 and 2660 A at pH 4.0. These values are essentially in agreement with those recorded by Wilson and Harris (4), although the solvent and pH were not stated in their case. A mini-
I , I I
)O 2500 3000 IO 2500 3000
WAVE LENGTH, A WAVE LENGTH, A
FIG. 3 FIG. 4
FIG. 3. The ultraviolet absorption spectra of neopyrithiamine in aqueous solution and at different acidities. The compound was present in 2.5 X 10-5 M concentra- tion. At pH 4 and pH 8,0.04 M phosphate buffer was present. The Beckman spec- trophotometer, model DU, with 1 cm. cells was employed. The instrument was adjusted with the solvent as the blank, and a minimum slit width was used at all wave-lengths. Density = log (lo/Z).
FIG. 4. The ultraviolet absorption spectra of thiamine (2.5 X IO-6 M) in aqueous solution and at different acidities. 0.04 M phosphate buffer was present in each so- lution.
mum is also present at 2570 A, as may be seen in Fig. 3. At pH 5 and higher the two compounds are more nearly alike in their ultraviolet absorp- tion, as is evident from a comparison of Figs. 3 and 4. Since the shift from a single maximum to the two maxima is usually characteristic of the 6-aminopyrimidine compounds as one proceeds from acid to more alkaline solution, both compounds would be expected to exhibit this phenomenon. However, it is evident that the occurrence of this shift is dependent upon
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

R. R. SEALOCK AND I-1. S. WHITE 401
the substituent attached to the 5-methylene group of the pyrimidine ring. When this shift does occur with typical 6-aminopyrimidine compounds, it has been observed that the pH at which the two maxima appear is a characteristic of the individual compound, and it even has been possible to distinguish and identify some of the different 6-aminopyrimidines by the pH value and other aspects of the shift.* This in turn emphasizes the necessity of control of pH in determining the absorption spectra of such compounds.
In the face of the findings with neopyrithiamine described above it is of interest to summarize briefly and illustrate the present status of thiamine with regard to its analogues. Thiamine is somewhat unique among bio- logically important compounds. On the one hand, there is the r81e of this substance as an important vitamin and accessory factor producing physi- ological effects, primarily through the enzymatic functioning of its pyro- phosphate ester, cocarboxylase. This role may be designated as that of physiological function. On the other hand, there is the destruction of thiamine by an enzyme present in a number of unrelated species, and at the moment, it is hardly conceivable that the latter phenomenon could be regarded as a component of the former. For each category there are one or more specific analogues that may be employed for the purpose of further analysis of mechanisms involved or useful inhibition. Neopyrithiamine is an antagonist to thiamine function (4), as is pyrithiamine (lo), and is also a substrate for the fish enzyme. In contrast, the o-aminobenzylthia- zolium compounds inhibit the fish enzyme (l), but show no antagonism toward thiamine function.2 The corresponding aminoethyl- and aminopro- pylthiazolium analogues exhibit similar behavior, being relatively strong in- hibitors of the enzymatic destruction of thiamine. With the m-amino- benzylthiazolium compounds neither antagonism to thiamine function nor inhibition of destruction is observed. Instead, activation of the destructive process is obtained. All of these compounds represent exten- sive modification of the molecular composition of thiamine and in no instance do we find them able to substitute for the vitamin in its physi- ological function. However, with only slight modification active substi- tutes are obtained. For example, the 2-ethylpyrimidine analogue of thi- amine possesses physiological activity (11). Without referring to numerous other interesting analogues which have been studied in more or less detail, it is obvious that a number of structural modifications are available for use in determining the biochemical properties and behavior of thiamine. In particular, they should make possible a most detailed analysis of the fundamental aspects of enzyme relationships of thiamine. The fact that
1 Sealock, R. R., and White, H. S., unpublished data. 2 Sealock, R. R., and Picken, J. C., unpublished data.
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

402 NEOPYRITHIAMINE AND THIAMINASE
they may also have practical uses yet to be discovered only emphasizes the interest in their further study.
EXPERIMENTAL
The thiaminase used in these experiments was prepared from acetone- desiccated powders of carp viscera (9) by extracting with 10 per cent sodium chloride in 0.2 M phosphate buffer at pH 7.4. The combined extracts were diluted with the solvent so that 2 ml. contained the soluble portion of the indicated amounts of the powder and were adjusted to pH 7.4 before being used. 2 ml. of this extract plus the desired volumes of substrates and inhibitors or activators were placed in test-tubes and when necessary the volume made to 5 ml. with water. The tubes were incubated for 2 hours at 37.5”, and unless otherwise indicated, the reaction was stopped by the addition of 5 ml. of 10 per cent trichloroacetic acid. After at least 30 minutes the precipitated protein was removed by filtration. For thiamine determination 2 ml. aliquots of the filtrates were analyzed by the Melnick-Field method. For determination of neopyrithiamine 5 ml. aliquots of a solution made by diluting the filtrates 1: 50 were treated with 3 ml. of an alkaline ferricyanide solution (3 ml. of 1 per cent potassium ferricyanide diluted to 100 ml. with 15 per cent sodium hydroxide), or, in order to determine the blank galvanometer reading, with 3 ml. of 15 per cent sodium hydroxide. In determining the total fluorescence in the presence of both substrates, 2 ml. aliquots of the filtrates were diluted to 250 ml., and 5 ml. aliquots analyzed as above. The fluorophotometer was adjusted with a quinine solution to allow maximum use of the gal- vanometer scale. In all analyses comparison was made with unincubated controls to which trichloroacetic acid was added immediately after the addition of the substrate.
SUMMARY
In preliminary experiments neopyrithiamine was found to inhibit the destruction of thiamine by the thiaminase of carp tissue.
The calculated inhibition produced by neopyrithiamine proved not to be inhibition in the usual sense, for this analogue in turn undergoes the same destructive reaction with thiaminase as does thiamine. Neopyrithi- amine therefore possesses in its isosteric structure the necessary compo- nents for complete action of the enzyme. It is therefore an additional substrate for this enzyme. When the two compounds are present in the same enzyme solution, they may be regarded as competing substrates.
Neopyrithiamine neither reacts nor interferes in the Melnick and Field diazotizkd p-aminoacetophenone method for the determination of thiamine.
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

It. R. SEALOCK AND H. S. WHITE 403
It does, however, react under the conditions of the thiochrome procedure and may be quantitatively determined by this method.
With appropriate control of pH, thiamine and neopyrithiamine may be readily distinguished by means of their ultraviolet absorption spectra, for the analogue exhibits two maxima in acid solution in contrast to thiamine which exhibits only one maximum at acidities less than pH 5.
BIBLIOGRAPHY
1. Scalock, R. R., and Goodland, R. L., J. Am. Chem. Sot., 66,507 (1944). 2. Sealock, R. R., and Livermore, A. H., J. Biol. Chem., 177, 553 (1949). 3. Sealock, R. R., and Davis, N. C., J. Riol. Chem., 17’7, 987 (1949). 4. Wilson, A. N., and Harris, S. A., Abstracts, Division of Biological Chemistry,
American Chemical Society, Washington, 18C (1948). 5. Tracy, A. H., and Elderfield, R. C., J. Org. Chem., 6,54 (1941). 6. Melnick, D., and Field, H., Jr., J. Biol. Chem., 127, 515 (1939). 7. Zima, O., and Williams, R. R., Ber. them. Ges., 73, 941 (1940). 8. Hennessy, D. J., Incl. and Eng. Chem., Anal. Ed., 13,216 (1941). 9. Sealock, R. R., Livermore, A. I-I., and Evans, C. A., J. Am. Chem. Sot., 66, 935
(1943). 10. Woolley, D. W., and White, A. G., J. Ezp. Med., 78,489 (1943). 11. Stein, G. A., Sampson, W. L., Cline, J. K., and Stevens, J. R., J. Am. Ch,em.
B a oc., 63,2059 (1941).
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Robert Ridgely Sealock and Hilda S. WhiteTHIAMINASE OF FISH TISSUESNEOPYRITHIAMINE AND THE
1949, 181:393-403.J. Biol. Chem.
http://www.jbc.org/content/181/1/393.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/181/1/393.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on May 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from