brusatol- as potent chemotherapeutic regimen and …...brusatol- as potent chemotherapeutic regimen...
TRANSCRIPT

Brusatol- as potent chemotherapeutic regimen and its role on reversing EMT transition
Aarthi.R, Dept of Pharmacology, PSG College of Pharmacy, Coimbatore
Abstract Cancer is highly resistant to chemotherapeutic agents and their prognosis after resistance is found to be poor. The development of new therapeutic entities is badly needed for this deadly malignancy. In this review article, we elaborately given information about brusatol, a natural quassinoid isolated from a Chinese herbal medicine named Bruceae Fructus, that it possess potent cytotoxic effect against different types of cell lines. Its anti-cancer effects has been demonstrated through different cell lines and its cytoxicity studies has been discussed briefly. In this review we also summarise possible mechanism for brusatol and its action on EMT reversal process. This article is expected to widen the knowledge of brusatol as potent chemotherapeutic agent and also indicating that brusatol is a promising adjunct to the current chemotherapeutic regimen for any mostly any type of cancer.
Keywords -Brusatol, adjuvant chemotherapeutic agent, EMT
INTRODUCTION Cancer is one of the leading causes of human death. Despite significant advances in cancer research throughout the decades, treatment of cancer is still facing severe challenges. Chemotherapy is one of the common types of cancer treatment and has been used as monotherapy or in combination with surgery or radiotherapy to treat cancer patients. However, chemotherapy drugs, both classical cytotoxic drugs and molecular targeted drugs, have been challenged by drug resistance, a major cause of cancer treatment failure and cancer-related mortality. In the last decade, tremendous effort has been paid to develop targeted cancer therapies. A number of monoclonal antibody drugs and small molecules, especially kinase inhibitors, have been developed and entered clinic in the hope of improving anticancer efficacy. While many of the targeted therapy drugs showed promising outcomes with improved overall survival, a large number of the patients receiving targeted therapy developed drug resistance after long-term drug administration [1]. Currently, more than 100 targeted cancer therapy drugs have been approved for cancer patients and much more are in clinical investigations. Therefore, cancer drug resistance will be a key factor to determine the success of the upcoming targeted therapy drugs. Drug resistance (or chemoresistance) can be divided into two groups: intrinsic (or de novo) drug resistance and acquired drug resistance [2,3]. Intrinsic drug resistance refers that the resistance factors have existed in the bulk of tumor cells before the drug treatment, whereas acquired drug resistance comes from that the resistance factors are developed during the drug treatment. Drug resistance arises from a broad range of mechanisms, such as drug efflux, drug metabolism, drug target mutations, etc. Recently, epithelial–mesenchymal transition (EMT) has received increasing attention for its role in cancer drug resistance [4].Over the last decade, substantial efforts have been devoted to determine the genes and pathways that are involved in tumor invasion and metastasis [5]. It has been suggested that increase in cell movement, scattering and epithelial-to-mesenchymal transition (EMT) are among the main properties of advanced tumors [6].
In recent years, herbal medicines or natural compounds, either used alone or combined with conventional chemotherapeutic agents, have been shown to have beneficial effects on diverse cancers [7]. Brusatol (BR), a natural quassinoid diterpenoid isolated from Bruceae Fructus, exhibited the most potent in vitro anti-tumor action among all the isolated quassinoids [8]. Furthermore, it was reported that brusatol acted as a unique inhibitor of the Nrf2 pathway that sensitized various cancer cells and A549 xenografts to chemotherapeutic drugs, suggesting brusatol might be a promising candidate for combating chemo-resistance and has the potential to be developed into an adjuvant chemotherapeutic agent. Brusatol can promote anti cancer property and inhibit EMT/metastasis via different mechanisms and pathways in human tumors[9]. In this review, we summarize the mechanisms by which Brusatol affects cancer cells, its activity, possible toxic effects and EMT action as well as experimental and (pre)clinical data supporting its potential application as therapeutic agent.
Brusatol: Quassinoids are a gathering of mixes separated from plants of the Simaroubaceae family, which have been utilized for a long time in people prescription. These atoms picked up notice after the underlying revelation of the counter leukemic action of one part, bruceantin, in 1975. As of now more than 150 quassinoids have been detached and arranged dependent on their substance structures and natural properties researched in vitro and in vivo [10]. Brusatol (BR), a characteristic quassinoid diterpenoid detached from Bruceae Fructus, showed the most powerful in vitro enemy of tumor activity among all the secluded quassinoids [11]. As of now, home grown drugs or common mixes, either utilized as a monotherapy or joined with traditional chemotherapeutic specialists, have been accounted for to apply gainful impacts on the treatment of different kinds of disease [12]. Bruceae Fructus alludes to the product of Brucea javanica (L.) Merr. ('Ya‑Dan‑Zi' in Chinese), and was at first recorded in Supplementations to the Compendium of Chinese Materia Medica. Bruceae Fructus has been connected to treat different infirmities, including malignant growth, amoebic looseness of the bowels and intestinal sickness, since the Ming Dynasty
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1753

(1364‑1644 AD) [13,14]. The antitumor movement of Bruceae Fructus is viewed as a standout amongst the most significant natural exercises of this plant, and it has been ordinarily endorsed to treat different sorts of disease in China. In earlier years, rising proof has been furnished concerning the antitumor activity of Bruceae Fructus [13]. B. javanica is rich in quassinoids, which are considered the dominating fixings in charge of its checked antitumor movement [15]. Brusatol (BR; C26H32O11), the synthetic structure of which is exhibited in Fig. 1, is one of the major quassinoids detached from B. javanica. This compound has been accounted for to apply stamped anti‑inflammatory [16], antimalarial [17] and antitumor exercises [18-21]. Likewise, BR has been shown to extraordinarily obstruct the atomic factor erythroid 2‑related factor 2 pathway, in this way sharpening different malignancy cells in vitro and A549 mouse xenografts to chemotherapeutic operators. These discoveries recommended that BR might be viewed as a promising possibility for battling chemoresistance and for further improvement into a viable adjuvant for chemotherapy drugs [22].
Extraction, fractionation, isolation and characterization: M. Zhao did extraction on plant Fructus Bruceae. Dried plant material of Fructus Bruceae (30 kg) was ground into little pieces with an electrical blender and refluxed in 80% watery EtOH for 1 h. The blend was sifted and the buildup re‐extracted twice. The pooled filtrates were dissipated to dryness in a revolving evaporator under diminished weight. The slurry buildup was suspended in boiling water and after that moved into a separatory channel. The arrangement was divided with hexane, ethyl acetic acid derivation and 1‐butanol progressively to get the hexane‐soluble‐fraction (HF, oil, 9.0% w/w yield), ethyl acetate‐soluble‐fraction (EAF, 0.45% w/w) and 1‐butanol‐soluble‐fraction (BF, 1.1% w/w). The EAF was connected to a macroporous gum segment (D 101) eluted with 80% fluid methanol and CH3)2CO successively to yield two eluates. The 80% watery methanol eluate was then isolated into 40 subfractions (Fr. 1– 40) on a Diaion HP‐20 segment eluted with blends of H2O and MeOH (10% MeOH→100% MeOH). Portions 1– 8 were then connected to a Diaion HP‐20ss segment eluted with an angle of H2O and MeOH (10% MeOH→45% MeOH) to get 30 subfractions (Fr. I1– I30). Parts I1– I10 were additionally isolated on a RP‐18 segment utilizing blends of H2O and MeOH (5% MeOH→20% MeOH) as the portable stage and pursued by re‐crystallization to yield bruceine H (12 mg) and bruceine D (600 mg). Divisions 20– 27 were isolated into six portions by Diaion HP‐20 chromatography. The third part was additionally isolated into 11 divisions (Fr. II1– II11) on a Sephadex LH‐20 section eluted with methanol. Portion II2 was then connected to a Diaion HP‐20ss section and eluted with H2O– MeOH (70:30→20:80) to acquire 25 parts (Fr. II2‐1– 25). Yadanzioside G (120 mg) and yadanzioside A (20 mg), javanicoside C (17 mg) and bruceantinoside A (25 mg) were filtered from Fr. II2‐2– 6, Fr. II2‐7– 9 and Fr. II2‐12– 15, separately, utilizing a semi‐preparative HPLC under the
accompanying conditions: versatile stage, water and acetonitrile (ACN); stream rate, 5 mL/min; dissolvable angle, 25% ACN– 30% ACN from 0 min to 20 min. Portions 33– 39 were additionally isolated on a Diaion HP‐20 segment and pursued by refinement on a Sephadex LH‐20 section (eluted with methanol) to yield brusatol (60 mg). The structures of bruceine D (Lee et al., 1979), bruceine H (Sakaki et al., 1984), yadanziosides A, G (Sakaki et al., 1985), javanicoside C (Kim et al., 2004), bruceantinoside An (Okano et al., 1981) and brusatol (Harigaya et al., 1989) were clarified by looking at their 1H, 13C NMR spectroscopic information with those distributed already. The structures were then additionally affirmed by the methods for APCI‐MS: bruceine D, C20H26O9, m/z 411 [M+ H]+; bruceineH,C20H26O10, m/z 427 [M + H]+; yadanzioside A, C32H44O16, m/z 685 [M + H]+; yadanzioside G, C36H48O18, m/z 769 [M + H]+; javanicoside C, C32H40O16, m/z 681 [M + H]+; bruceantinoside A, C34H46O16, m/z 711 [M + H]+; and brusatol, C26H32O11, m/z 477 [M + H]+[23].
Cytotoxicity assay: M. Zhao directed examinations utilizing the concentrates and the detached mixes. The cytotoxic impacts of these quassinoids were tried on two pancreatic adenocarcinoma cell lines including PANC‐1 and SW1990, and the outcomes are appeared in Fig. 1. What's more, their IC50 esteems on the development of these two pancreatic disease cell lines are appeared Table 1. Among them, brusatol showed the most strong cytotoxicity on both cell lines, with IC50 estimations of 0.36 μM and 0.10 μM, separately. It is worthy1968 (Sim et al., 1968), and has been accounted for to have antileukemic (Lee et al., 1984; Mata‐Greenwood et al., 2002; Hitotsuyanagi et al., 2006), calming (Hall et al., 1983), antitrypanosomal (Bawm et al., 2008) and antitobacco mosaic infection (Yan et al., 2010) exercises. Its in vitro antipancreatic adenocarcinoma action has not been accounted for previously. The promising trial discoveries render this concoction constituent a potential antipancreatic disease operator justifying further pharmaceutical improvement. Further robotic examinations on brusatol would give basic bits of knowledge into its enemy of malignant growth impact [23].
Pharmacokinetic, Metabolic profiling and Elimination of Brusatol: In the study conducted by Nan Guofter intravenous administration of brusatol (1mg/kg,) the plasma concentration in rats decreased rapidly, consistent with the pharmacokinetic characteristics observed in their other study. The average cumulative excretion rate of brusatol was found to be 5.82% in urine during 24 h, and 0.71% in bile during 12 h, which indicated majority of the drug were excreted as metabolites. Four metabolites were identified and rationalization, M438 is the hydrolysis product, M536 is the hydroxylation product, M618 is the glucuronidation product, and M424 is the demethylation or hydrolysis product of M438. The study reported 4 metabolites of brusatol for the first time, provide deep insight into brusatol metabolism and promote the development of effective drugs for anti-cancer[24].
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1754

Table 1. Effects of Fructus Bruceae extracts and isolates on the viability of human pancreatic cancer cells as
measured by SRB assay
Fig 1. Structure of brusatol
Mechanism: Recent studies with cultured cells and mouse xenograft models unequivocally bolster the thought that Nrf2 is an extraordinary focus to defeated chemoresistance. Overexpression of Nrf2 in malignant growth cells that have low basal dimensions of Nrf2, upgraded obstruction in an assortment of disease cells including neuroblastoma, bosom, ovarian, prostate, lung, and pancreatic disease cells [24– 29]. Consolidated utilization of Nrf2-siRNA and platins hindered the development of A549 xenografts in mice (30). Also, thinks about have exhibited that concealment of Nrf2 by Keap1 overexpression sharpened SPEC-2 cells, which are gotten from sort II endometrial disease, and SPEC-2 xenografts to cisplatin [27]. In help of a job for Nrf2 in chemoresistance, articulation of Nrf2 in malignancy cells expanded amid procurement of medication opposition [31, 32]. Altogether, these outcomes show that Nrf2 adds to chemoresistance saw in numerous kinds of malignancies starting from various organs. Further, this outlines the earnest need to recognize intensifies that stifle the Nrf2 pathway and form them into druggable mixes to improve the adequacy of malignant growth medications. Brusatol specifically restrained the Nrf2 Pathway. To battle Nrf2-interceded chemoresistance, they scanned for aggravates that smother the Nrf2 pathway by screening countless items for their capacity to repress ARE-luciferase movement utilizing a steady cell line, MDA-MB-231-ARE-
Luc [32]. A plant extricate from Brucea javanica (L) Merr. (Simaroubaceae), an evergreen bush developed in Southeast Asia and Northern Australia, was found to repress ARE-luciferase movement and the protein dimensions of Nrf2. In this manner, the plant separate was additionally fractionated and filtered mixes were tried for their capacity to repress the Nrf2 pathway, which brought about the ID of brusatol, a quassinoid. Brusatol hinders ARE-luciferase movement in a portion subordinate way in the MDA-MB-231-ARELuc stable cell line (Fig. 2A).
A
B
C
Figure 2. (A) Brusatol inhibited ARE-dependent luciferase activity. MDA-MB-231-ARE-Luc cells were treated with several doses of brusatol for 16 h. (B). Brusatol-mediated reduction of Nrf2 was reversible. (Upper) A549 cells were
treated with 40 nM of brusatol for the indicated time points. (Lower) A549 cells were pretreated with 40 nM of brusatol
for 4 h and then brusatol was removed and cells were further cultured for the indicated periods.(C) Brusatol reduced the protein levels of Nrf2 and its downstream
genes. A549 cells were treated with the indicated doses of brusatol for 16 h.
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1755

Besides, 40 nM of brusatol essentially diminished the protein dimension of Nrf2 after just 2– 4 h of treatment and had the capacity to keep up decreased Nrf2 protein levels contrasted with control for up with 72 h (Fig. 2B, Upper). Curiously, brusatol yet not brucein C, another quassinoid with a comparative synthetic structure, diminished Nrf2 in a portion subordinate way (Fig. 2C, Nrf2,). To determine if the impacts of brusatol are reversible, we tried the capacity of Nrf2 to recuperate after expulsion of brusatol. Once more, brusatol diminished Nrf2 protein levels when contrasted and untreated cells (Fig. 1B, Lower). Besides, following the evacuation of brusatol, Nrf2 protein level immediately recouped inside 1 h and outperformed those of basal dimensions at 2– 4 h. Along these lines, Nrf2 protein levels achieved harmony 8– 24 h after the expulsion of brusatol (Fig. 2B, Lower). Notwithstanding Nrf2, the protein dimension of Nrf2-target qualities, including γGCS, MRP1, and MRP2, was likewise decreased in a portion subordinate way, while just a slight decrease in NQO1 was observed(Fig. 2C). Brusatol on Radiosensitivity: Xiaohui Sun, Qin Wang performed experiment for brusatol on radiosensitivity. Nrf2 is an atomic translation factor that secures cells by coordinating cell stress signals, coordinating different transcriptional programs [33], just as inclusion in different cell forms, for example, expansion, separation, movement, apoptosis, and angiogenesis [34,35]. Raised Nrf2 protein levels are seen in malignant growths, for example, lung, head and neck, nerve bladder, and pancreatic disease [36,37], and an addition of Nrf2 work improves cell multiplication and presents radioresistance and chemoresistance in these disease types [38]. To improve the adequacy of disease medications, specialists
have built up a progression of systems directing the dimension of Nrf2 protein. the Nrf2 inhibitor, brusatol, as a novel radiosensitizer, which could conquer the radioresistance of lung malignant growth cells by advancing ROS creation and expanding DNA harm. In accordance with their investigation, double treatment with brusatol and radiotherapy could viably repress the expansion of malignancy cells. Bruastol on oxidative Stress: In the investigation directed by Tongde Wu, it was affirmed the raised dimension of Nrf2 and its downstream reaction which keeps up the low dimensions of ROS in the CSC improved mammosphere subpopulation. Past work from the lab has shown that brusatol hindered the Nrf2 pathway through improved ubiquitination and debasement of Nrf2 [39]. Brusatol treatment prompted diminished articulation of Nrf2 at the protein level and a smothered cancer prevention agent reaction (Figure 3) which results in stamped height of intracellular ROS particularly in mammospheres (Figures 4A and B). We showed that mammospheres with higher Nrf2 articulation are progressively helpless to brusatol-intervened refinement to taxol treatment (Figures 4E and F). The system basic the upregulation of the Nrf2 pathway in mammospheres is as yet indistinct. In a lung malignant growth model, Pan et al. [40] revealed that 26S proteasome movement is down-directed in lung malignancy stem-like cells spread in vitro. Since Nrf2 is additionally exposed to 26S proteasomal debasement, it is very conceivable that the diminished Nrf2 corruption may offer ascent to abnormal state of Nrf2 giving mammospheres a development favorable position and protection from chemo drugs[41].
Fig3. Brusatol suppresses Nrf2-mediated response in both mammospheres and adherent cells. (A and B) Immunoblot analysis of the indicated proteins
from cell lysates from tertiary mamospheres (Mammo.) and adherent (Ad.) MCF-7 (A) and MDA-MB-231 (B) Cells, treated with PBS or brusatol. (C and D) qPCR analysis of the mRNA levels of indicated genes with cell lysates from tertiary mammospheres and adherent MCF-7 (C) and MDA-MB-231 (D) Cells with indicated treatments. For qRT-PCR, results are expressed as mean_SD. (n¼3 independent experiments). NQO1 activity (E) and intracellular glutathione levels (F) Were measured in single cell suspension from tertiary mammospheres and adherent MCF-7 and MDA-MB-231 cells. Results are
expressed as mean_SD. (n¼3 independent experiments).
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1756
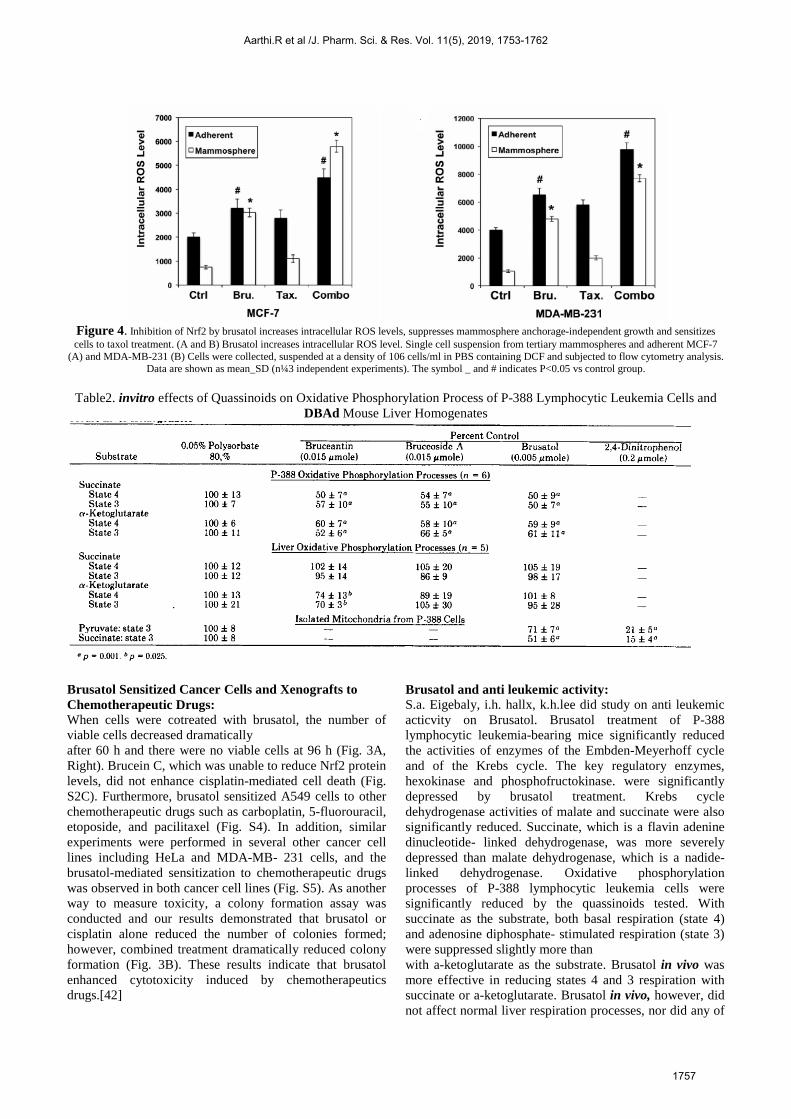
Figure 4. Inhibition of Nrf2 by brusatol increases intracellular ROS levels, suppresses mammosphere anchorage-independent growth and sensitizes cells to taxol treatment. (A and B) Brusatol increases intracellular ROS level. Single cell suspension from tertiary mammospheres and adherent MCF-7
(A) and MDA-MB-231 (B) Cells were collected, suspended at a density of 106 cells/ml in PBS containing DCF and subjected to flow cytometry analysis. Data are shown as mean_SD (n¼3 independent experiments). The symbol _ and # indicates P<0.05 vs control group.
Table2. invitro effects of Quassinoids on Oxidative Phosphorylation Process of P-388 Lymphocytic Leukemia Cells and
DBAd Mouse Liver Homogenates
Brusatol Sensitized Cancer Cells and Xenografts to Chemotherapeutic Drugs: When cells were cotreated with brusatol, the number of viable cells decreased dramatically after 60 h and there were no viable cells at 96 h (Fig. 3A, Right). Brucein C, which was unable to reduce Nrf2 protein levels, did not enhance cisplatin-mediated cell death (Fig. S2C). Furthermore, brusatol sensitized A549 cells to other chemotherapeutic drugs such as carboplatin, 5-fluorouracil, etoposide, and pacilitaxel (Fig. S4). In addition, similar experiments were performed in several other cancer cell lines including HeLa and MDA-MB- 231 cells, and the brusatol-mediated sensitization to chemotherapeutic drugs was observed in both cancer cell lines (Fig. S5). As another way to measure toxicity, a colony formation assay was conducted and our results demonstrated that brusatol or cisplatin alone reduced the number of colonies formed; however, combined treatment dramatically reduced colony formation (Fig. 3B). These results indicate that brusatol enhanced cytotoxicity induced by chemotherapeutics drugs.[42]
Brusatol and anti leukemic activity: S.a. Eigebaly, i.h. hallx, k.h.lee did study on anti leukemic acticvity on Brusatol. Brusatol treatment of P-388 lymphocytic leukemia-bearing mice significantly reduced the activities of enzymes of the Embden-Meyerhoff cycle and of the Krebs cycle. The key regulatory enzymes, hexokinase and phosphofructokinase. were significantly depressed by brusatol treatment. Krebs cycle dehydrogenase activities of malate and succinate were also significantly reduced. Succinate, which is a flavin adenine dinucleotide- linked dehydrogenase, was more severely depressed than malate dehydrogenase, which is a nadide-linked dehydrogenase. Oxidative phosphorylation processes of P-388 lymphocytic leukemia cells were significantly reduced by the quassinoids tested. With succinate as the substrate, both basal respiration (state 4) and adenosine diphosphate- stimulated respiration (state 3) were suppressed slightly more than with a-ketoglutarate as the substrate. Brusatol in vivo was more effective in reducing states 4 and 3 respiration with succinate or a-ketoglutarate. Brusatol in vivo, however, did not affect normal liver respiration processes, nor did any of
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1757

the quassinoids affect in vitro liver respiration at 0.015 pmole. The mitochondria studies showed that brusatol did not uncouple oxidative phosphorylation like 2,4-dinitropheriol does. Nor did hrusatol treatment stimulate mitochondrial adenosine triphosphatase activity, which would facilitate the uncoupling of mitchondrial oxidative phosphorylation. Rather, brusatol appeared to act on the mitochondria1electron-transport chain. In vivo studies showed drastic increases in the reduced forms of chain cofactors with either inalate or succinate as the substrate. In vivo hrusatol effects were more striking than in vitro effects on reducing cofactors, which may explain the observed increased effects of hrusatol on in vivo states 4 and 3 respiration after 3 days of treatment as compared to in uitro effects. UV in vitro studies showed that brusatol chemically interacted with nadide increasing the reduced form ahsorbance at 340 nm. Cytochrome c, a heme, also was reduced in the presence of hrusatol to the ferrous form with an increase in absorbance at 550nm. Brusatol on HCV and hepatoma cells: Brusatol has been appeared to have an enemy of multiplication impact on disease cells including chemoresistant cells. In spite of the fact that the exact system by which brusatol restrains Nrf2 isn't completely comprehended, it was demonstrated that brusatol and related mixes hinders protein synthesis[44]. Besides, brusatol specifically hinders the Nrf2 pathway, and the decrease of Nrf2 is through improvement of ubiquitination and debasement of Nrf2 [45]. Thus, the change of mRNA articulation saw in the present examination could be an optional wonder after decrease of Nrf2 protein brought about by brusatol. An ongoing report exhibited that brusatol diminished the Nrf2 protein level in a post-translational way, since this decrease seemed in all respects ahead of schedule (from 30 min to 12 h) after its organization, with maximal hindrance at around 2 [46]. The present investigation utilizing the HPI cells also demonstrated that decrease of Nrf2 was maximal at 2 h after the organization of brusatol supporting the post-translational instrument for the decrease in the Nrf2 protein level by brusatol. The present examination led byYuko Murakami showed enlargement of the double enemy of HCV and anticancer impacts of sorafenib by blend with brusatol, in any event in vitro. Their mix accomplished practically complete concealment of HCV contamination at 72 h after their organization. For clinical application, mix treatment utilizing drugs with an alternate pharmaceutical instrument prompts the decrease of unfriendly impacts. We subsequently imagine that brusatol could be clinically appropriate in blend with sorafenib for the treatment of HCC, particularly when associative with HCV contamination. In any case, there are worries that hindrance of Nrf2 may result in askew consequences for non-malignancy cells, accordingly causing surprising antagonistic impacts. Such antagonistic occasions may happen not just on the grounds that Nrf2 is a transcriptional controller that controls a variety of qualities incorporating qualities associated with host safeguard and digestion, yet
in addition on the grounds that brusatol influences a more extensive scope of qualities than siRNA against Nrf2. In this way, for clinical use of brusatol, it will be important to widely illuminate its poisonous quality preclinically in vitro and in vivo investigations. Brusatol on neurotoxicity There is plentiful proof that Aβ has a key job in activating neurotoxicity in neurons.[47] Although the precise components that intercede Aβ danger have not been generally comprehended, they might be associated with oxidative stress‐dependent apoptosis.[48] Based on these outcomes, thinks about directed by Xin Liu et al., inferred that hindering the neurotoxicity of glioma cells instigated by Aβ may control the advancement of glioma and in this way, treat glioma. In this investigation, brusatol was distinguished to ensure human glioma U‐251 cells against Aβ‐induced neurotoxicity. Also, brustaol directed the Nrf2/HO‐1 pathway in U‐251 cells to hinder neurotoxicity. In particular, they found that when treated with Aβ, cell viabilities were fundamentally diminished in U‐251 cells. A similar outcome was additionally distinguished in PC12 cells.[49] Moreover, Aβ was found to initiate cell apoptosis in U‐251 cells and direct the outflows of cleaved‐caspase3, Bax, and Bcl‐2 related with cell apoptosis. The Western smear investigation proposed that Aβ expanded the protein dimensions of Bax and cleaved‐caspase3, while decreased Bcl‐2 protein level in U‐251 cells. Besides, Aβ diminished the proportion of Bcl‐2/Bax articulation. Wang et al11 likewise detailed comparable outcomes in SH‐SY5Y cells. Furthermore, ROS development was accounted for to be engaged with the neurotoxicity prompted by Aβ.[50] Here, we distinguished that Aβ advanced the ROS levels in U‐251 cells to incite cell passing which was steady with a past study.[51] Moreover, Aβ was found to lessen the MMP associated with the early procedure of apoptosis in the their present investigation. Liu et al[52] exhibited that the MMP level in N2a cells treated with Aβ was fundamentally decreased. These results suggest that Aβ induced the neurotoxicity of U‐251 cells through affecting ROS generation, mitochondrial dysfunction, cell viability, and death. In summary, BR inhibited Aβ‐induced neurotoxicity in U‐251 cells and can effectively reverse a series of cellular changes caused by Aβ‐induced neurotoxicity. Further, BR regulates the PI3K/AKT/mTOR and Nrf2/ HO‐1 pathways to inhibit neurotoxicity in U‐251 cells. Therefore, they concluded that brusatol has a neuroprotective effect that may be used to treat gliomas. Brusatol on colorectal cancer Studies led by Eun-Taex Oh et al., demonstrated that c-Myc overexpression likewise diminishes collection of intracellular ferrous iron in brusatol-treated malignant growth cells under hypoxia (Figure 5A and 5B) by expanding mitochondrial ROS, in this manner lessening brusatol-prompted colorectal disease cell demise under hypoxia (Figure 6A and 6B). Also, brusatol treatment instigated malignant growth cell passing by advancing PHD-interceded debasement of HIF-1α, which thus
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1758

fundamentally smothered tumor development in both RKO and HCT116 xenografts. These outcomes firmly propose that brusatol expands PHD-initiated debasement of HIF-1α by repressing c-Myc articulation under hypoxia, in this manner diminishing mitochondrial ROS creation and causing malignant growth cell passing. Despite the fact that the the detailed mechanism by which brusatol diminishes c-Myc articulation stays misty, their discoveries all things considered exhibit that brusatol actuates cell demise in colorectal disease cells under hypoxia by advancing PHD-intervened corruption of HIF-1α[53].
A
B
Fig 5. Effect of c-Myc expression on intracellular ferrous iron concentrations in RKO (G) and HCT116 (H) cells
incubated with or without 100 nM brusatol under hypoxia. Data are presented as means ± SD (**P < 0.01, ***P <
0.001, ****P < 0.0001; ANOVA)
A
B
Fig6. Effects of c-Myc on brusatol-induced clonogenic cell death in RKO (G) and HCT116 (H) cells under hypoxia.
Data are presented as means ± SD (*P < 0.05, ***P < 0.001, ****P < 0.0001; ANOVA).
Toxic effects of brusatol on reproductive system To evaluate the impact of brusatol on female regenerative limit, the examination directed by Rujun Ma etal., intended to explore the danger and primer systems of Brusatol on mouse oocyte meiotic development. In the examination, they demonstrated that Brusatol treatment prompted the articulated loss of Nrf2 in mouse oocytes, which lead to oocyte development disappointment and shaft/chromosomal imperfections. In addition, their information likewise shown that the dimension of Cyclin B1 was controlled by Nrf2 in oocytes, which is basic for chromosome buildup and microtubule polymerization. In conclusion, oocyte quality is a basic component directing the fruitfulness of a female. In this way the investigation directed uncovered that the mouse oocytes development was disturbed after Brusatol treatment through axle morphology and chromosome buildup, which gave the proof to the harmful impacts of Brusatol on reproductive systems[54].
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1759
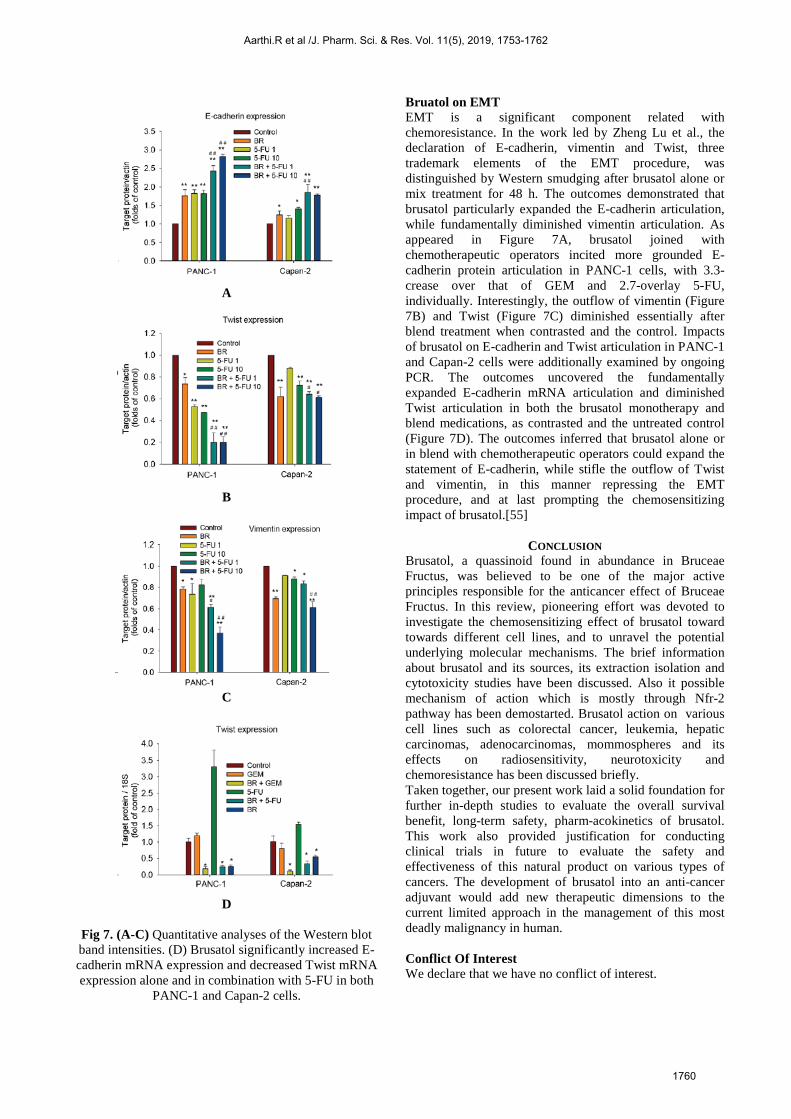
A
B
C
D
Fig 7. (A-C) Quantitative analyses of the Western blot band intensities. (D) Brusatol significantly increased E-cadherin mRNA expression and decreased Twist mRNA expression alone and in combination with 5-FU in both
PANC-1 and Capan-2 cells.
Bruatol on EMT EMT is a significant component related with chemoresistance. In the work led by Zheng Lu et al., the declaration of E-cadherin, vimentin and Twist, three trademark elements of the EMT procedure, was distinguished by Western smudging after brusatol alone or mix treatment for 48 h. The outcomes demonstrated that brusatol particularly expanded the E-cadherin articulation, while fundamentally diminished vimentin articulation. As appeared in Figure 7A, brusatol joined with chemotherapeutic operators incited more grounded E-cadherin protein articulation in PANC-1 cells, with 3.3-crease over that of GEM and 2.7-overlay 5-FU, individually. Interestingly, the outflow of vimentin (Figure 7B) and Twist (Figure 7C) diminished essentially after blend treatment when contrasted and the control. Impacts of brusatol on E-cadherin and Twist articulation in PANC-1 and Capan-2 cells were additionally examined by ongoing PCR. The outcomes uncovered the fundamentally expanded E-cadherin mRNA articulation and diminished Twist articulation in both the brusatol monotherapy and blend medications, as contrasted and the untreated control (Figure 7D). The outcomes inferred that brusatol alone or in blend with chemotherapeutic operators could expand the statement of E-cadherin, while stifle the outflow of Twist and vimentin, in this manner repressing the EMT procedure, and at last prompting the chemosensitizing impact of brusatol.[55]
CONCLUSION
Brusatol, a quassinoid found in abundance in Bruceae Fructus, was believed to be one of the major active principles responsible for the anticancer effect of Bruceae Fructus. In this review, pioneering effort was devoted to investigate the chemosensitizing effect of brusatol toward towards different cell lines, and to unravel the potential underlying molecular mechanisms. The brief information about brusatol and its sources, its extraction isolation and cytotoxicity studies have been discussed. Also it possible mechanism of action which is mostly through Nfr-2 pathway has been demostarted. Brusatol action on various cell lines such as colorectal cancer, leukemia, hepatic carcinomas, adenocarcinomas, mommospheres and its effects on radiosensitivity, neurotoxicity and chemoresistance has been discussed briefly. Taken together, our present work laid a solid foundation for further in-depth studies to evaluate the overall survival benefit, long-term safety, pharm-acokinetics of brusatol. This work also provided justification for conducting clinical trials in future to evaluate the safety and effectiveness of this natural product on various types of cancers. The development of brusatol into an anti-cancer adjuvant would add new therapeutic dimensions to the current limited approach in the management of this most deadly malignancy in human. Conflict Of Interest We declare that we have no conflict of interest.
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1760

REFERENCES 1. Juchum, M.; Gunther, M.; Laufer, S.A. Fighting cancer drug
resistance: Opportunities and challenges formutation-specific EGFR inhibitors. Drug Resist. Update 2015, 20, 12–28.
2. Gottesman, M.M. Mechanisms of cancer drug resistance. Ann. Rev. Med. 2002, 53, 615–627.
3. Kartalou, M.; Essigmann, J.M. Mechanisms of resistance to cisplatin. Mutat. Res. 2001, 478, 23–43.
4. Du B, Shim J. Targeting epithelial–mesenchymal transition (EMT) to overcome drug resistance in cancer. Molecules. 2016 Jul 22;21(7):965.
5. A. Bahrami, A. Hesari, M. Khazaei, S.M. Hassanian, G.A. Ferns, A. Avan, The therapeutic potential of targeting the BRAF mutation in patients with colorectal cancer. J Cell Physiol. 233, 2162–2169 (2018)
6. F. Bocci, H. Levine, J.N. Onuchic andM.K. Jolly Deciphering the dynamics of epithelial-mesenchymal transition and cancer stem cells in tumor progression, arXiv preprint arXiv:1808.09113, (2018)
7. Yin SY, Wei WC, Jian FY, Yang NS. Therapeutic applications of herbal medicines for cancer patients. Evid Based Complement Alternat Med. 2013; 2013:302426.
8. Zhao M, Lau ST, Leung PS, Che CT, Lin ZX. Seven quassinoids from Fructus Bruceae with cytotoxic effects on pancreatic adenocarcinoma cell lines. Phytother Res. 2011; 25:1796–800.
9. Ren D, Villeneuve NF, Jiang T, Wu T, Lau A, Toppin HA, Zhang DD. Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proc Natl Acad Sci USA. 2011; 108:1433–38.
10. Fiaschetti G, A Grotzer M, Shalaby T, Castelletti D, Arcaro A. Quassinoids: From traditional drugs to new cancer therapeutics. Current medicinal chemistry. 2011 Jan 1;18(3):316-28.
11. Zhao M, Lau ST, Leung PS, Che CT, Lin ZX. Seven quassinoids from Fructus Bruceae with cytotoxic effects on pancreatic adenocarcinoma cell lines. Phytother Res. 2011; 25:1796–800.
12. Beutler, J. A.; Kang, M. I.; Robert, F.; Clement, J. A.; Pelletier, J.; Colburn, N. H.; McKee, T. C.; Goncharova, E.; McMahon, J. B.; Henrich, C. J. Quassinoid inhibition of AP-1 function does not correlate with cytotoxicity or protein synthesis inhibition. J. Nat. Prod., 2009, 72(3), 503-6.
13. Fukamiya, N.; Lee, K. H.; Muhammad, I.; Murakami, C.; Okano, M.; Harvey, I.; Pelletier, J. Structure-activity relationships of quassinoids for eukaryotic protein synthesis. Cancer Lett., 2005, 220(1), 37-48.
14. Jin, X.; Jin, H. R.; Lee, D.; Lee, J. H.; Kim, S. K.; Lee, J. J. A quassinoid 6alpha-tigloyloxychaparrinone inhibits hypoxiainducible factor-1 pathway by inhibition of eukaryotic translation initiation factor 4E phosphorylation. Eur. J. Pharmacol., 2008, 592(1-3), 41-7.
15. Cuendet, M.; Pezzuto, J. M. Antitumor activity of bruceantin: an old drug with new promise. J. Nat. Prod., 2004, 67(2), 269-72.
16. Cuendet, M.; Christov, K.; Lantvit, D. D.; Deng, Y.; Hedayat, S.; Helson, L.; McChesney, J. D.; Pezzuto, J. M. Multiple myeloma regression mediated by bruceantin. Clin. Cancer Res., 2004, 10(3), 1170-9.
17. Kupchan, S. M.; Britton, R. W.; Lacadie, J. A.; Ziegler, M. F.; Sigel, C. W. The isolation and structural elucidation of bruceantin and bruceantinol, new potent antileukemic quassinoids from Brucea antidysenterica. J. Org. Chem., 1975, 40(5), 648-54.
18. Grieco, P. A.; Speake, J. D. Synthetic Studies on Quassinoids: Total Synthesis and Biological Evaluation of (+)-Des-Dchaparrinone. J. Org. Chem., 1998, 63(17), 5929-5936.
19. Polonsky, J. Quassinoid Bitter Principles. Fortschr. Chem. Org. Naturst., 1973, 30, 101-50.
20. Gurel, G.; Blaha, G.; Moore, P. B.; Steitz, T. A. U2504 determines the species specificity of the A-site cleft antibiotics: the structures of tiamulin, homoharringtonine, and bruceantin bound to the ribosome. J. Mol. Biol., 2009, 389(1), 146-56.
21. Wiseman, C. L.; Yap, H. Y.; Bedikian, A. Y.; Bodey, G. P.; Blumenschein, G. R. Phase II trial of bruceantin in metastatic breast carcinoma. Am. J. Clin. Oncol., 1982, 5(4), 389-91.
22. Arseneau, J. C.; Wolter, J. M.; Kuperminc, M.; Ruckdeschel, J. C. A Phase II study of Bruceantin (NSC-165, 563) in advanced malignant melanoma. Invest. New Drugs, 1983, 1(3), 239-42.
23. Zhao M, Lau ST, Leung PS, Che CT, Lin ZX. Seven quassinoids from Fructus Bruceae with cytotoxic effects on pancreatic
adenocarcinoma cell lines. Phytotherapy Research. 2011 Dec;25(12):1796-800.
24. Guo N, Xu X, Yuan G, Chen X, Wen Q, Guo R. Pharmacokinetic, metabolic profiling and elimination of brusatol in rats. Biomedical Chromatography. 2018 Dec;32(12):e4358.
25. Wang XJ, et al. (2008) Nrf2 enhances resistance of cancer cells to chemotherapeutic drugs, the dark side of Nrf2. Carcinogenesis 29:1235–1243.
26. Zhang P, et al. (2010) Loss of Kelch-like ECH-associated protein 1 function in prostate cancer cells causes chemoresistance and radioresistance and promotes tumor growth. Mol Cancer Ther 9:336–346.
27. Homma S, et al. (2009) Nrf2 enhances cell proliferation and resistance to anticancer drugs in human lung cancer. Clin Cancer Res 15:3423–3432.
28. Cho JM, Manandhar S, Lee HR, Park HM, Kwak MK (2008) Role of the Nrf2- antioxidant system in cytotoxicity mediated by anticancer cisplatin: Implication to cancer cell resistance. Cancer Lett 260:96–108.
29. Hong YB, et al. (2010) Nuclear factor (erythroid-derived 2)-like 2 regulates drug resistance in pancreatic cancer cells. Pancreas 39:463–472.
30. Singh A, et al. (2008) RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res 68:7975–7984.
31. Shim GS, Manandhar S, Shin DH, Kim TH, Kwak MK (2009) Acquisition of doxorubicin resistance in ovarian carcinoma cells accompanies activation of the NRF2 pathway. Free Radic Biol Med 47:1619–1631.
32. Kim SK, et al. (2008) Increased expression of Nrf2/ARE-dependent anti-oxidant proteins in tamoxifen-resistant breast cancer cells. Free Radic Biol Med 45:537–546.
33. Sporn, M.B.; Liby, K.T. Nrf2 and cancer: The good, the bad and the importance of context. Nat. Rev. Cancer 2012, 12, 564–571.
34. Na, H.K.; Surh, Y.J. Oncogenic potential of Nrf2 and its principal target protein heme oxygenase-1. Free Radic.Biol. Med. 2014, 67, 353–365.
35. Ganan-Gomez, I.;Wei, Y.; Yang, H.; Boyano-Adanez, M.C.; Garcia-Manero, G. Oncogenic functions of the transcription factor Nrf2. Free Radic. Biol. Med. 2013, 65, 750–764.
36. Homma, S.; Ishii, Y.; Morishima, Y.; Yamadori, T.; Matsuno, Y.; Haraguchi, N.; Kikuchi, N.; Satoh, H.; Sakamoto, T.; Hizawa, N.; et al. Nrf2 enhances cell proliferation and resistance to anticancer drugs in human lung cancer. Clin. Cancer Res. 2009, 15, 3423–3432.
37. Singh, A.; Bodas, M.; Wakabayashi, N.; Bunz, F.; Biswal, S. Gain of Nrf2 function in non-small-cell lung cancer cells confers radioresistance. Antioxid. Redox Signal. 2010, 13, 1627–1637. [CrossRef] [PubMed]
38. Ren D, Villeneuve NF, Jiang T, et al. Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proc Nat Acad Sci U S A 2011;108:1433–1438.
39. Pan J, Zhang Q, Wang Y, You M. 26S proteasome activity is down-regulated in lung cancer stem-like cells propagated in vitro. PloS one 2010;5:e13298.
40. Diehn M, Cho RW, Lobo NA, et al. Association of reactive oxygen species levels and radioresistance in cancer stem cells. Nature 2009;458:780–783.
41. Leonard GD, Fojo T, Bates SE. The role of ABC transporters in clinical practice. Oncologist 2003;8:411–424.
42. Ren D, Villeneuve NF, Jiang T, Wu T, Lau A, Toppin HA, Zhang DD. Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proceedings of the National Academy of Sciences. 2011 Jan 25;108(4):1433-8.
43. ElGebaly SA, Hall IH, Lee KH, Sumida Y, Imakura Y, Wu RY. Antitumor agents XXXV: Effects of brusatol, bruceoside A, and bruceantin on P‐388 lymphocytic leukemia cell respiration. Journal of pharmaceutical sciences. 1979 Jul;68(7):887-90.
44. Willingham W 3rd, Considine RT, Chaney SG, Lee KH, Hall IH. Reversibility of protein synthesis inhibition by quassinoid antineoplastic agents in a rabbit reticulocyte system. Biochem Pharmacol. 1984;33(2):330–3.
45. Ren D, Villeneuve NF, Jiang T, Wu T, Lau A, Toppin HA, Zhang DD. Brusatol enhances the efficacy of chemotherapy by inhibiting
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1761

the Nrf2-mediated defense mechanism. Proc Natl Acad Sci U S A. 2011;108(4):1433–8.
46. Olayanju A, Copple IM, Bryan HK, Edge GT, Sison RL, WongMW, Lai ZQ, Lin Z-X, Dunn K, Sanderson CM, et al. Brusatolprovokes a rapid and transient inhibition of Nrf2 signaling andsensitizes mammalian cells to chemical toxicity—implications fortherapeutic targeting of Nrf2. Free Radic Biol Med. 2015;78:202–12.
47. Selkoe DJ. Alzheimer's disease results from the cerebralaccumulation and cytotoxicity of amyloid beta‐protein. J AlzheimersDis. 2001;3:75‐80.
48. Loo DT, Copani A, Pike CJ, Whittemore ER, Walencewicz AJ,Cotman CW. Apoptosis is induced by beta‐amyloid in cultured central nervous system neurons. Proc Natl Acad Sci USA.1993;90:7951‐7955.
49. Liu JY, Chen XX, Chen HY, et al. Downregulation of aquaporin 9exacerbates beta‐amyloid‐induced neurotoxicity in Alzheimer'sdisease models in vitro and in vivo. Neuroscience. 2018;394:72‐82.
50. Behl C, Davis JB, Lesley R, Schubert D. Hydrogen peroxide mediates amyloid beta protein toxicity. Cell. 1994;77:817‐827.
51. Telerman A, Ofir R, Kashman Y, Elmann A. 3,5, 4′‐trihydroxy‐6,7,3′‐trimethoxyflavone protects against beta amyloid‐induced neurotoxicity through antioxidative activity and interference with cell signaling. BMC Complement Altern Med.2017;17:332.
52. Liu J, Wang M. Carvedilol protection against endogenous Abeta‐induced neurotoxicity in N2a cells. Cell Stress Chaperones. 2018;23:695‐702.
53. Oh ET, Kim CW, Kim HG, Lee JS, Park HJ. Brusatol-mediated inhibition of c-Myc increases HIF-1α degradation and causes cell death in colorectal cancer under hypoxia. Theranostics. 2017;7(14):3415.
54. Ma R, Li H, Zhang Y, Lin Y, Qiu X, Xie M, Yao B. The toxiceffects and possible mechanisms of Brusatol on mouse oocytes. PloSone. 2017 May 18;12(5):e0177844.
55. Lu Z, Lai ZQ, Leung AW, Leung PS, Li ZS, Lin ZX. Exploring brusatol as a new anti-pancreatic cancer adjuvant: biologicalevaluation and mechanistic studies. Oncotarget. 2017 Oct17;8(49):84974.
Aarthi.R et al /J. Pharm. Sci. & Res. Vol. 11(5), 2019, 1753-1762
1762